

Abstract
Inner Mongolia steppe is one of the suitable habitats for Didymodon species and a new species, Didymodonmanhanensis C. Feng & J. Kou from Manhan Mountain in semi-arid region in Inner Mongolia, China is described and illustrated. It is characterised by leaves incurved and slightly twisted when dry, spreading when moist, narrowly lanceolate from an ovate base; subulate and fragile leaf apices; distally bistratose leaf margins that are recurved in proximal 2/3–3/4; excurrent costa with guide cells in 2–3 layers and without ventral stereids; smooth laminal cells and red KOH laminal colour reaction. Our morphological analyses and molecular results, based on DNA sequences of ITS, rps4 and trnM-trnV, confirm that D.manhanensis belongs to a group that includes D.obtusus J. Kou, X.-M. Shao & C. Feng and D.daqingii J. Kou, R.H. Zander & C. Feng. This new species is compared with similar species and its phylogenetic position and ecology are discussed.
Keywords: Asia, Manhan Mountain, phylogenetic analysis, taxonomy
Introduction
Inner Mongolia, situated in Inner Eurasia, is located in the northern part of China and presents a strip distribution from the northeast to the west. The district habitats are temperate continental monsoon climate. The annual mean temperature is 8 °C, which increases from east to west and the annual precipitation is 35–530 mm, which decreases from southeast to northwest (Miao 2017). The area of grassland accounts for 60% of the whole Inner Mongolia and more than one-quarter of the total area of grassland in China. The grassland in Inner Mongolia is divided into three types: meadow steppe formed by, for example, Stipabaicalensis Roshev, Leymuschinensis (Trin.) Tzvelev; typical steppe formed by, for example, Stipagrandis P. Smirn., Stipakrylovii Roshev, Leymuschinensis (Trin.) Tzvelev and desert steppe formed by, for example, Stipaklemenzii Roshev, Stipaglareosa P.A. Smirn., Stipabreviflora Griseb (Hua et al. 2021). The main vegetation types of the steppes present distinct zonal features (Wu et al. 2005). The Inner Mongolia steppe is a suitable habitat for the Didymoodn Hedw. species and several new species were recently discovered (e.g. Kou et al. 2016a; Kou et al. 2019; Feng et al. 2022).
The taxonomy of genus Didymodon is complicated, involving the differentiation from related genera, such as Barbula Hedw. and the circumscriptions of its infrageneric sections (Zander 1993; Zander 2007; Zhang et al. in press). A recent important event associated with Didymodons. lat. was the split of the genus into seven smaller genera: Aithobryum R.H. Zander, Didymodons. str., Exobryum R.H. Zander, Fuscobryum R.H. Zander, Geheebia Schimp., Trichostomopsis Card. and Vinealobryum R.H. Zander, based on macro-evolutionary analysis and the dissilient genus concept applied (Zander 2013; Zander 2019). Although initially this revolutionary concept was considered unnecessary or unsupported (Blockeel and Kučera 2019), it has later been supported by molecular phylogenetic data, but with some alterations (Jiménez et al. 2022; Zhang et al. in press) and has gained acceptance by some other authors (e.g. Kou and Feng 2018; Osman et al. 2021; Feng et al. 2022). During our continuous investigations of xerophilic mosses, especially Pottiaceae Hampe, in China (e.g. Feng et al. 2016a, b; Kou et al. 2016b, 2018, 2019; Kou and Feng 2018), many specimens have been collected from different provinces. Amongst them, two samples collected from Manhan Mountain in Inner Mongolia of Didymodon s. lat. from stony habitats are different from species previously reported in the area (Li et al. 2001). They mostly resemble Didymodonobtusus J. Kou, X.-M. Shao & C. Feng. To clarify their taxonomic identity, we conducted phylogenetic analysis and confirm that these samples belong to the genus Didymodons. str. (Zander 2013), but do not match with any species known in the genus. Here, we describe this unknown moss as a new species.
Materials and methods
Morphological observations
Over 3000 specimens of the genus Didymodons. lat. were examined during our revision of Pottiaceae in China. More than 50 field investigations were conducted in past years and the specimens included in this study were housed in the Herbaria at IFP, KUN and NMAC. Microscopic examinations and measurements were taken with a ZEISS Primo Star light microscope and photomicrographs were obtained with a Canon EOS 70D camera, mounted on this microscope. Specimens were examined in 2% potassium hydroxide (KOH). Three plants were dissected from each collection and, for each shoot, every possible structure from the gametophyte had to be examined and a record kept of what was found for each individual species. Specific morphological and anatomical features of taxonomic importance were assessed mainly following Zander (1993). Leaves were always taken from the upper and middle parts of the stem and cross-sections were made in the middle part of the stem. Measurements of leaf width were taken at the base, mid-leaf and upper part. Cross-sections were made at mid-leaf.
Phylogenetic analyses
To test the phylogenetic position of the new species, two specimens collected from Manhan Mountain were sampled. Due to its great similarity with D.obtusus and Didymodondaqingii J. Kou, R.H. Zander & C. Feng, the isotypes of the two species were added to the dataset. We employed one nuclear (ITS) and two chloroplast markers (rps4 and trnM-trnV), which had been used successfully in previous phylogenetic studies in Didymodon s. lat. and enabled the re-use of earlier results and easier interpretation of new data (Werner et al. 2004, 2005, 2009; Kučera and Ignatov 2015; Kučera et al. 2018; Ronikier et al. 2018; Jiménez et al. 2022; Zhang et al. in press). Phylogenetic trees are created and shown separately. The complete list with sample names and GenBank accession numbers is presented in Tables 1 and 2. DNA extraction, PCR amplification and sequencing procedure followed the protocols described by Wang et al. (2010).
Table 1.
New sequences used in this study, including taxa vouchers information and GenBank accession numbers.
| Species | Voucher information | ITS | rps4 | trnM-trnV |
|---|---|---|---|---|
| Didymodonmanhanensis 4 | China, Inner Mongolia, Chao Feng 2016060162 | OL514237 | OL450506 | OL450515 |
| Didymodonmanhanensis a3 | China, Inner Mongolia, Chao Feng 2016060176 | OL514238 | OL450507 | OL450516 |
| Didymodonobtusus | China, Tibet, Xiao-Ming Shao & Jin Kou 20140815037 | OL514239 | OL450508 | OL450517 |
| Didymodondaqingii | China, Inner Mongolia, Chao Feng 20170605032 | OL514240 | OL450509 | OL450518 |
Table 2.
. Sequences from GenBank used in this study, including taxa and GenBank accession numbers.
| Species | ITS | rps4 | trnM-trnV |
|---|---|---|---|
| Acaulontriquetrum | MW398556 | ||
| Aloinarigida | MW398549 | ||
| Aloinellaandina | MW398550 | ||
| Andinellachurchilliana | MW398720 | ||
| Andinellacoquimbensis | MW398711 | ||
| Andinellaelata | MW398708 | ||
| Andinellagranulosa | MW398714 | ||
| Andinellalimensis | MW398710 | ||
| Andinellaoedocostata | MW398733 | ||
| Andinellapruinosa | MW398726 | ||
| Barbulaunguiculata | MW398553 | HM147777 | JQ890366 |
| Bryoerythrophyllumrecurvirostrum | MW398547 | JQ890468 | JQ890407 |
| Bryoerythrophyllumrubrum | MW398548 | ||
| Chenialeptophylla | MW398561 | ||
| Cinclidotusriparius | MW398554 | ||
| Crossidiumsquamiferum | MW398558 | ||
| Didymodonacutus | AY437111 | KP307551 | KP307667 |
| Didymodonalpinus | MW398606 | ||
| Didymodonandreaeoides | MW398768 | ||
| Didymodonanserinocapitatus | MW398649 | KP307545 | KP307640 |
| Didymodonasperifolius | MW398594 | JQ890472 | KP307600 |
| Didymodonaustralasiae (Trichostomumaustralasiae) | MW398737 | KP307571 | KP307651 |
| Didymodonbrachyphyllus (Vinealobryumbrachyphyllum) | MW398817 | ||
| Didymodonbuckii | MW398578 | ||
| Didymodoncaboverdeanus | MW398607 | ||
| Didymodoncalifornicus (Vinealobryumcalifornicum) | MW398819 | ||
| Didymodoncanoae | MW398584 | ||
| Didymodoncardotii | MW398729 | ||
| Didymodonchallaensis (Trichostomopsischallaensis) | MW398748 | ||
| Didymodonconstrictus | MW398613 | ||
| Didymodoncordatus | MW398664 | KP307564 | KP307668 |
| Didymodonditrichoides | MW398642 | ||
| Didymodoneckeliae (Vinealobryumeckeliae) | MW398826 | ||
| Didymodonedentulus | MW398685 | ||
| Didymodonepapillatus | MW398665 | ||
| Didymodonerosodenticulatus | MW398792 | MF536597 | MF536635 |
| Didymodonerosus | EU835148 | MF536609 | MF536646 |
| Didymodonfallax (Geheebiafallax) | MW398779 | KP307552 | KP307663 |
| Didymodonferrugineus (Geheebiaferruginea) | MW398796 | MF536588 | MF536625 |
| Didymodonfragilicuspis | KP307482 | ||
| Didymodonfuscus | MW398689 | KP307537 | KP307601 |
| Didymodonaff.fuscus | KP307546 | KP307615 | |
| Didymodongaochienii | KP307538 | KP307658 | |
| Didymodongelidus | MW398693 | ||
| Didymodongiganteus | MW398786 | KP307548 | KP307669 |
| Didymodonglaucus | MW398612 | ||
| Didymodonguangdongensis (Vinealobryumguangdongense) | MW398657 | ||
| Didymodonhedysariformis | MW398582 | KP307569 | KP307629 |
| Didymodonhengduanensis | MW398629 | ||
| Didymodonhegewaldiorum | MW398739 | ||
| Didymodonherzogii | MW398746 | ||
| Didymodonhumboldtii | MW398667 | ||
| Didymodonicmadophilus | MW398632 | KP307598 | KP307604 |
| Didymodonimbricatus | MW398646 | ||
| Didymodonincrassatolimbatus | MW398572 | ||
| Didymodonincurvus | MW398680 | ||
| Didymodoninsulanus (Vinealobryuminsulanum) | MW398811 | ||
| Didymodonjaponicus | MW398757 | ||
| Didymodonjimenezii | MW398622 | ||
| Didymodonjohansenii | MW398589 | KP307542 | KP307662 |
| Didymodonkunlunensis | MW398610 | ||
| Didymodonlaevigatus | MW398618 | ||
| Didymodonlainzii | MW398575 | ||
| Didymodonleskeoides (Geheebialeskeoides) | MW398777 | MF536604 | MF536642 |
| Didymodonluehmannii | MW398718 | ||
| Didymodonluridus | AY437098 | MF536587 | MF536624 |
| Didymodonmaschalogena | MW398615 | ||
| Didymodonmaximus (Geheebiamaxima) | MW398784 | MF536591 | MF536628 |
| Didymodonmesopapillosus | MW398758 | ||
| Didymodonmolendoides | MW398687 | ||
| Didymodonmongolicus | KU058175 | ||
| Didymodonmurrayae | KP307513 | KP307563 | KP307650 |
| Didymodonnevadensis | MW398730 | ||
| Didymodonnicholsonii (Vinealobryumnicholsonii) | MW398808 | ||
| Didymodonnigrescens | LC545516 | KP307543 | KP307611 |
| Didymodonnorrisii | MW398830 | KP307585 | KP307617 |
| Didymodonnovae-zelandiae | MW398769 | ||
| Didymodonobtusus | MW398666 | ||
| Didymodonoccidentalis | KP307533 | KP307599 | |
| Didymodonochyrarum | MW398763 | ||
| Didymodonparamicola (Trichostomopsisparamicola) | MW398740 | ||
| Didymodonpatagonicus | MW398675 | ||
| Didymodonperobtusus | KP307523 | KP307539 | KP307609 |
| Didymodonrevolutus (Husnotiellarevoluta) | MW398569 | JQ890471 | KP307646 |
| Didymodonrevolutusvar.africanus | MW398568 | ||
| Didymodonrigidulus | MW398602 | KP307589 | KP307647 |
| Didymodonrigidulusvar.subulatus | MW398672 | ||
| Didymodonrivicola | MW398599 | KP30756 | KP307607 |
| Didymodonsantessoni | MW398705 | ||
| Didymodonsicculus | MW398801 | MF536606 | MF536643 |
| Didymodonsinuosus | MW398567 | JQ890476 | JQ890410 |
| Didymodonspadiceus (Geheebiaspadicea) | MW398795 | MF536593 | MF536631 |
| Didymodonsubandreaeoides | AY437108 | KP307570 | KP307630 |
| Didymodontectorum | MW398659 | ||
| Didymodontibeticus | MW398638 | ||
| Didymodontomaculosus | AY437114 | ||
| Didymodontophaceus | MW398807 | MF536607 | MF536644 |
| Didymodontophaceusvar.anatinus | MF536589 | MF536626 | |
| Didymodontorquatus | MW398719 | ||
| Didymodonumbrosus (Trichostomopsisumbrosa) | MW398742 | ||
| Didymodonvalidus | MW398650 | ||
| Didymodonvinealis (Vinealobryumvineale) | MW398815 | JQ890475 | KP307606 |
| Didymodonvinealisvar.rubiginosus | MW398822 | ||
| Didymodonvulcanicus | MW398636 | ||
| Didymodonwaymouthii | MW398770 | ||
| Didymodonwisselii | MW398655 | ||
| Didymodonxanthocarpus | MW398696 | KP307534 | KP307638 |
| Didymodonzanderi | MW398585 | KP307535 | KP307621 |
| Dolotortulamniifolia | MW398555 | ||
| Erythrophyllopsisandina | MW398546 | ||
| Gertrudiellauncinicoma | MW398698 | ||
| Gertrudiellauncinicomavar.serratopungens | MW398701 | ||
| Guerramontesiamicrodonta | MW398543 | ||
| Hennediellaheimii | GQ339750 | ||
| Hennediellapolyseta | GQ339759 | ||
| Leptodontiumexcelsum | MW398545 | ||
| Microbryumcurvicolle | JX679986 | JX679936 | |
| Microbryumdavallianum | MW398557 | ||
| Pseudocrossidiumhornschuchianum | MW398551 | JQ890481 | JQ890420 |
| Pseudocrossidiumrevolutum | MW398552 | ||
| Pterygoneurumovatum | MW398560 | ||
| Sagenotortulaquitoensis | GQ339761 | ||
| Stegonialatifolia | MW398559 | ||
| Syntrichiaruralis | MW398564 | FJ546412 | FJ546412 |
| Tortulamuralis | MW398562 | JN581679 | JQ890421 |
| Tortulasubulata | MW398563 | ||
| Triquetrellaarapilensis | MW398544 | ||
| Tridontiumtasmanicum | MW398750 |
The sequences were aligned by using MAFFT 7.222 (Kazutaka and Daron 2013) and then edited in BioEdit 7.0.1 (Hall 1999). The concatenation of individual rps4 and trnM-trnV fragments was performed by our custom Perl script. Phylogenetic analyses were performed by using the Bayesian Inference (BI) and Maximum Likelihood (ML) methods. MrBayes 3.2.6 (Ronquist et al. 2012) was used for BI analyses under the GTR substitute model. The following was used: two Markov Chain Monte Carlo (MCMC) searches were run for 10 million generations each, with a sampling frequency of 1000. The first 25% of the trees were discarded as burn-in. The convergence between runs in all cases dropped below 0.01. ML analyses were executed in IQ-TREE 1.6.3 (Nguyen et al. 2014) under the TPM3u+F+R3 (for cpDNA) and TIM3e+I+G4 (for ITS) substitute models, respectively, selected by the ModelFinder programme (Kalyaanamoorthy et al. 2017), based on the Bayesian Information Criterion (BIC) and 1000 fast bootstrapping replicates were used. The final obtained trees were visualised and edited in FigTree v.1.4.0 (Rambaut 2014).
Results
The chloroplast (cp) and ITS alignments comprised 1313 and 1364 nucleotide sites, respectively. The BI and ML phylogenetic trees have a consistent topology, although there are different levels of support depending on the method. Hence, only the topologies with branch lengths from the BI trees are presented, with added support from the ML method on the respective trees (Figs 3–4). Although the inference from analysed chloroplast regions (Fig. 3) and the ITS (Fig. 4) agrees in most aspects, the position of the new species is different between the two above phylogenetic trees and, thus, both of them are reserved. The topology of the ITS dataset shows that D.manhanensis is nested within the monophyletic group comprising Didymodonepapillatus J. Kou, X.-M. Shao & C. Feng, Didymodonmongolicus D.-P. Zhao & T.-R. Zhang, Didymodonvalidus Limpr., Didymodonwisselii (Dixon) D.H. Norris & T.J. Kop. and Vinealobryumguangdongensis C. Feng & J. Kou and is sister to D.obtusus, but with weakly-supported values. In the combined plastid dataset, D.manhanensis is nested within the group including Didymodoncordatus Jur. and is sister to D.daqingii and Didymodonanserinocapitatus (X.J. Li) R.H. Zander, with well-supported values.
Figure 3.

Phylogenetic relationships (50% majority consensus tree) from the Bayesian Inference of the concatenated rps4 and trnM-trnV datasets. Numbers above branches indicate posterior probability from the BI analysis, followed by bootstrap values for the ML analysis.
Figure 4.
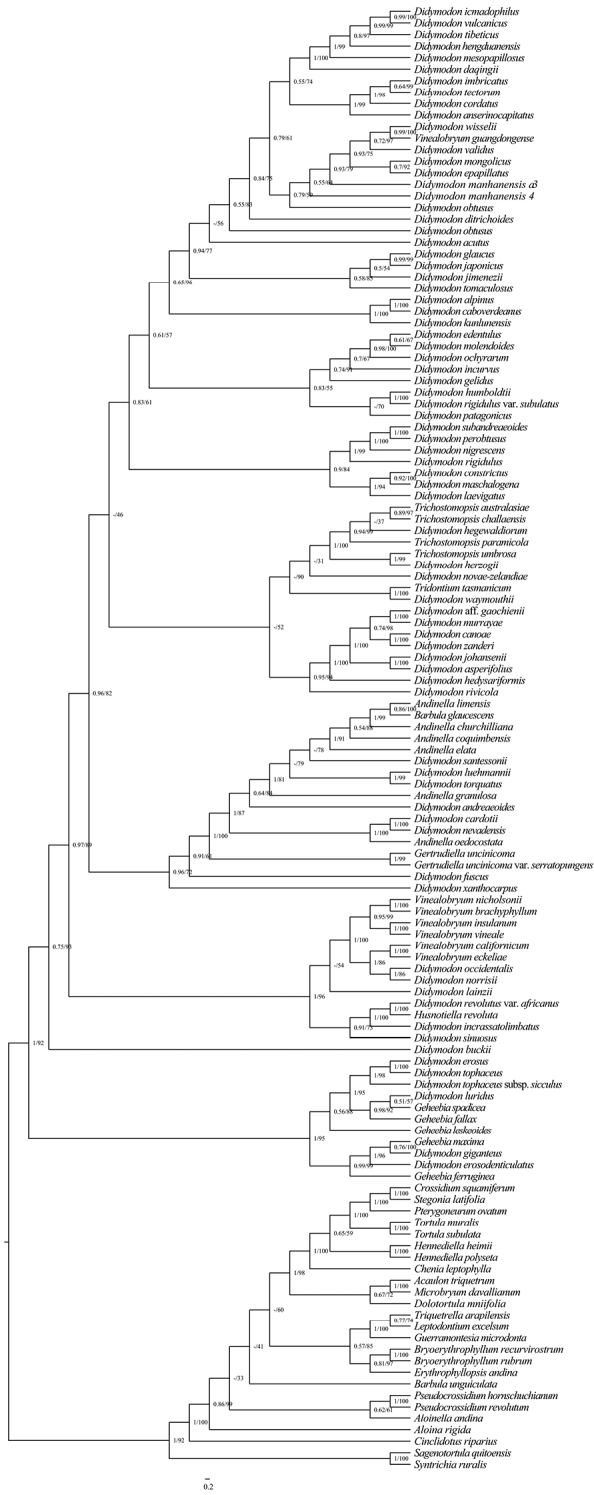
Phylogenetic relationships (50% majority consensus tree) from the Bayesian Inference on the ITS dataset. Numbers above branches indicate posterior probability from the BI analysis, followed by bootstrap values for the ML analysis.
Discussion
As indicated by Zander (1993), Didymodons. lat. is heterogeneous and could be profitably split. In our phylogenetic analyses, this genus is polyphyletic and its species can be classified within several well-supported monophyletic clades, which correspond to other phylogenetic studies of the genus (Feng et al. 2022; Jiménez et al. 2022; Zhang et al. in press). Our results reveal a close relationship between D.manhanensis and two recently-described species in China: D.daqingii and D.obtusus. Although the latter two species were considered identical by Sollman et al. (2020), they are not closely related in our phylogenetic analyses, based on both ITS and chloroplast data.
Didymodonmanhanensis is distinguished from all congeners by the following combination of diagnostic features: leaves incurved and slightly twisted when dry, spreading when moist, narrowly lanceolate from an ovate base; subulate and fragile leaf apices; distally bistratose leaf margins that are recurved in proximal 2/3–3/4; costal guide cells in 2–3 layers and without ventral stereids, smooth laminal cells and red KOH laminal colour reaction. This combination of characters suggests the placement of D.manhanensis in the sect.Didymodon (Zander 1978, 1993, 1998). Following the recent revolutionary work on the genus Didymodons. lat. by Zander (2013, 2019), morphologically, it belongs in the amended genus Didymodons. str. Its systematic position in Didymodons. str. was also confirmed by our phylogenetic analyses, based on both ITS and chloroplast data.
Chloroplast data support that D.manhanensis is closely related to D.cordatus and sister to both D.daqingii and D.anserinocapitatus. However, D.manhanensis differs morphologically from D.cordatus by the costa with guide cells in 2–3 layers and without ventral stereids and smooth laminal cells. It differs from D.daqingii by the leaves that are narrowly lanceolate from an ovate base, smooth laminal cells and red KOH laminal colour reaction; it differs from D.anserinocapitatus by the distally bistratose leaf margins and lack of swollen and deciduous leaf apex (Zander 2007). In the ITS analyses, there is successive branching of clades, including D.obtusus J. Kou, X.-M. Shao & C. Feng, D.manhanensis, D.epapillatus J. Kou, X.-M. Shao & C. Feng, D.mongolicus D.-P. Zhao & T.-R. Zhang, D.validus Limpr., Vinealobryumguangdongensis C. Feng & J. Kou and D.wisselii (Dixon) D.H. Norris & T.J. Kop. Amongst these species, D.manhanensis is most similar to D.obtusus, a species that was recently described from Tibet in China (Kou et al. 2018), but the former can be distinguished from the latter by its narrowly lanceolate leaves from an ovate base and spreading when moist, subulate and somewhat fragile leaf apex and unistratose distal lamina.
There are three species distributed in China that have excurrent costa and smooth laminal cells may be confused with the new species. Didymodonditrichoides (Broth.) X.-J. Li & S. He, a species known from North American, Asia (China) and the Atlantic Islands (Iceland) (Li et al. 2001; Zander 2007), differs from the new species by the unistratose leaf margins, costa with 1–2 layers of guide cells and with 0–1 layer of ventral stereids and yellowish KOH laminal colour reaction (Zander 2007). Didymodonvalidus Limpr. can be separated from D.manhanensis by the twisted and incurved leaves when dry, unistratose leaf margins, costa with 1 layer of guide cells and with 1–3 layers of ventral stereids and yellowish-green KOH laminal colour reaction (Shuayib et al. 2017).
The lanceolate to long-lanceolate leaves with a widely ovate base, distally bistratose leaf margins, excurrent costa and epapillose laminal cells are likewise found in Didymodonochyrarum J.A.Jiménez & M.J.Cano, a species described from tropical South America (Jiménez and Cano 2019), which may be confused with the new species. However, D.ochyrarum can be separated from D.manhanensis by its plane leaf margins, marginal basal cells running up the margin forming a distinctly differentiated area of transversely thick-walled cells and yellowish KOH laminal colour reaction.
Taxonomic treatment
. Didymodon manhanensis
C. Feng & J. Kou sp. nov.
70A98184-1F76-5333-8D1C-D128E33BD3A0
Figs 1 , 2 Chinese name: 蛮汉山对齿藓
Figure 1.

DidymodonmanhanensisA dry plants B moist plants C cross-section of stem D leaves E leaf apex F upper part of costa (dorsal) G upper part of costa (ventral) H axillary hairs. Photographed on 21 November 2021 by Chao Feng from the holotype (NMAC!).
Figure 2.

DidymodonmanhanensisA median leaf cells B basal juxtacostal cells C basal marginal cells; D–H cross-sections of leaves, sequentially from apex to base. Photographed on 21 November 2021 by Chao Feng from the holotype (NMAC!).
Type.
China. Inner Mongolia: Ulanqab City, Manhan Mountain, 40°39'19.2931"N, 112°19'36.3792"E, on soil under the grass, elevation 1417 m, 20 June 2016, Chao Feng 2016060162 (holotype: NMAC!; isotype: MO!).
Diagnosis.
It is distinguished from all congeners by the following combination of diagnostic features: leaves incurved and slightly twisted when dry, spreading when moist, narrowly lanceolate from an ovate base; subulate and fragile leaf apices; distally bistratose leaf margins that are recurved in proximal 2/3–3/4; costal guide cells in 2–3 layers and without ventral stereids, smooth laminal cells and red KOH laminal colour reaction.
Description.
Plants medium, growing in turfs, green-blackish distally, brown-blackish proximally. Stems very seldom branched, 0.8–1.6 cm in length, not papillose, transverse section rounded to rounded-pentagonal, central strand developed, sclerodermis present, hyalodermis absent; axillary hairs filiform, of 4–8 hyaline cells, the basal cell brown. Leaves crowded on stem, incurved and slightly twisted when dry, spreading when moist, narrowly lanceolate from an ovate base, constricted just above the base, 1.3–2.3 × 0.43–0.55 mm, distal lamina narrowly channelled ventrally; margins plane distally, recurved in proximal 2/3–3/4 of leaf, entire, distal margins bistratose; apex subulate, somewhat fragile; leaf base ovate, not sheathing, not decurrent; costa stout, tapering distally, 57.5–75 µm wide at base, excurrent as a long, thick subula, not spurred, ventral cells of costa in upper middle part of leaf quadrate or subquadrate, smooth, 4 rows of cells across costa ventrally at mid-leaf, dorsal cells of costa in upper middle part of leaf quadrate or subquadrate, smooth, transverse section semicircular to nearly rounded, epidermis present adaxially and abaxially, not or weakly bulging, ventral stereids absent, guide cells 10–16 in 2–3 layers, 2–4 layers of dorsal stereids, reniform or crescent-shaped, without hydroids; upper laminal cells quadrate to rhombic, usually with angular lumens, 7.5–10 × 5–10 µm, smooth, slightly thick-walled, weakly convex on both surfaces, distal lamina unistratose, basal cells weakly differentiated juxtacostally, rectangular, 12.5–37.5 × 5–7.5 µm, thin-walled, smooth; basal marginal cells subquadrate or quadrate, 5–8.75 × 6.25–7.5 µm, with weakly-thickened walls, smooth. Gemmae absent. Dioicous. Sporophytes unknown. KOH laminal colour reaction red.
Additional specimens examined.
China Inner Mongolia: Ulanqab City, Manhan Mountain, on soil under the grass, 20 June 2016, Chao Feng 2016060176 (NMAC).
Etymology.
The specific epithet refers to Manhan Mountain, the type locality.
Habitat and distribution.
Manhan Mountain is situated in Liangcheng County in the southern section of the Yinshan Mountains in the middle of Inner Mongolia, with an average altitude of approximately 1500 m (Huang et al. 2014). Its soil types are mainly leaching grey, cinnamonic soil (Lyu et al. 2012). The vegetation on Manhan Mountain is typical forest shrub vegetation, including natural forest that consists of Betulaplatyphylla Sukaczev and Populusdavidiana Dode, plantation that consists of Larixprincipis-rupprechtii Mayr and Pinussylvestrisvar.mongolica Litv., natural shrubs that consists of Ostryopsisdavidiana Decne., Spiraeasalicifolia L. and Rosadavurica Pall. and the herbaceous plants including Stipabungeana Trin., Cleistogenessquarrosa (Trin.) Keng, Lespedezabicolor Turcz., Carex spp. and Leymuschinensis (Trin.) Tzvelev (Zhang et al. 2017; Li et al. 2021). Didymodonmanhanensis is currently known only from the type locality at the foot of the Manhan Mountain, north-western Liangcheng County, Inner Mongolia, China, growing on soil under the grass.
Key to species morphologically similar to D.manhanensis
| 1 | Leaf apices apically swollen as a propagulum | D.anserinocapitatus |
| – | Leaf apices not swollen, usually evenly narrowing | 2 |
| 2 | Cells on the upper ventral surface of the costa elongate | D.wisselii |
| – | Cells on the upper ventral surface of the costa quadrate | 3 |
| 3 | Laminal cells smooth | 4 |
| – | Laminal cells papillose | 10 |
| 4 | Costa with 2–3 layers of guide cells and without ventral stereids | 5 |
| – | Costa with 1 layer of guide cells and with ventral stereids | 6 |
| 5 | Leaves patent to spreading when moist, leaf lamina bistratose | D.obtusus |
| – | Leaves spreading when moist, leaf lamina unistratose | D.manhanensis |
| 6 | Costa percurrent or ending before the apex | 7 |
| – | Costa long-excurrent | 8 |
| 7 | Leaf margins bistratose near apex | D.epapillatus |
| – | Leaf margins unistratose | D.mongolicus |
| 8 | Plants flagellate, leaves linear-lanceolate | D.ditrichoides |
| – | Plants thickly leaved, leaves short-lanceolate to long-lanceolate | 9 |
| 9 | Leaves appressed when dry | D.acutus |
| – | Leaves twisted or incurved when dry | D.validus |
| 10 | Leaf margins plane | D.tibeticus |
| – | Leaf margins recurved | 11 |
| 11 | Costa without ventral stereids | 12 |
| – | Costa with ventral stereids | 13 |
| 12 | Costa excurrent | D.daqingii |
| – | Costa ending below apex | D.imbricatus |
| 13 | Marginal basal cells forming a distinctly differentiated area of smooth and transversely thick-walled cells | D.hengduanensis |
| – | Marginal basal cells not forming a distinctly differentiated area | 14 |
| 14 | Distal laminal cell superficial walls thicker than the internal walls | D.mesopapillosus |
| – | Distal laminal cell superficial walls of same thickness as the internal walls | 15 |
| 15 | Laminal cells with low papillae over the transverse walls, which reach the two adjacent cells | 16 |
| – | Laminal cells with papillae situated over the lumina | 17 |
| 16 | Leaves spreading when moist | D.guangdongensis |
| – | Leaves erect to patent when moist | D.vulcanicus |
| 17 | Leaf margins recurved in proximal 1/4–3/4 | D.icmadophilus |
| – | Leaf margins strongly recurved or revolute to near apex | 18 |
| 18 | Leaf base squared in shape, costa slender | D.tectorum |
| – | Leaf base usually ovate in shape, costa stout | D.cordatus |
Supplementary Material
Acknowledgements
Sincerest thanks are given to Dr Richard H. Zander, Missouri Botanical Garden, for his consistent help during the authors’ study of the Pottiaceae in China and for his valuable comments on the manuscript. We really appreciate Dr Jan Kučera, University of south Bohemia, for providing many valuable suggestions on the molecular experiment and help to obtain the sequences of D.daqingii and D.obtusus. We are very grateful to Dr Matt von Konrat of the Field Museum, Dr Grzegorz J. Wolski of University of Lodz and one anonymous reviewer for their constructive criticisms. This work was supported by the Natural Science Foundation of China (grant no. 42001045, 32060051, 31660051), Shenzhen Key Laboratory of Southern Subtropical Plant Diversity (grant no. 99203030) and the Innovative team of China’s Ministry of Education-Research on the sustainable use of grassland resources (IRT_17R59).
Citation
Feng C, Zhang G-L, Wu T-T, Kou J (2022) Didymodon manhanensis (Pottiaceae, Bryophyta), a new species from Inner Mongolia steppe, China and its phylogenetic position, based on molecular data. PhytoKeys 197: 41–57. https://doi.org/10.3897/phytokeys.197.80531
Funding Statement
Natural Science Foundation of China (grant no. 42001045, 32060051, 31660051), Shenzhen Key Laboratory of Southern Subtropical Plant Diversity (grant no. 99203030) and the Innovative team of China's Ministry of Education-Research on the sustainable use of grassland resources (IRT_17R59)
References
- Blockeel T, Kučera J. (2019) Notes from the BBS Workshop on Didymodon. Field Bryology 121: 23–30. [Google Scholar]
- Feng C, Kou J, Yu C-Q, Shao X-M. (2016a) Encalyptagyangzeana C.Feng, X.-M.Shao & J.Kou (Encalyptaceae), a new species from Tibet, China. Journal of Bryology 38(3): 262–266. 10.1080/03736687.2015.1117170 [DOI] [Google Scholar]
- Feng C, Kou J, Yu C-Q, Shao X-M. (2016b) Bryoerythrophyllumzanderi (Bryophyta, Pottiaceae), a new species from Tibet, China. Nova Hedwigia 102(3–4): 339–345. 10.1127/nova_hedwigia/2015/0308 [DOI] [Google Scholar]
- Feng C, Kou J, Wu T-T, Wang J-L, Zhang G-L. (2022) Didymodonsinicus (Pottiaceae, Musci), a new species from China and its phylogenetic position based on molecular data. Nordic Journal of Botany 2022(3): e03424. 10.1111/njb.03424 [DOI]
- Hall T. (1999) BioEdit: A user-friendly biological sequence alignment program for Windows 95/98/NT. Nucleic Acids Symposium Series 41: 95–98. [Google Scholar]
- Hua Y-C, Sa R-L, Bao W-X. (2021) Temporal lag of grassland vegetation growth response to precipitation in Inner Mongolia. Journal of Northeast Forestry University 49(7): 47–55. [In Chinese] 10.13759/j.cnki.dlxb.2021.07.009 [DOI] [Google Scholar]
- Huang Y, Cha M-H, Wu X-D. (2014) Preliminary study on rodent control in different forest stands of Manhan Mountain Forest Farm in Inner Mongolia, China. Chinese journal of Vector Biology and Control 25(3): 273–274[+276]. [In Chinese] 10.11853/j.issn.1003.4692.2014.03.021 [DOI]
- Jiménez JA, Cano MJ. (2019) Didymodonochyrarum (Pottiaceae, Bryophyta), a new Andean species from tropical South America. Acta Musei Silesiae. Scientiae Naturales 68(1–2): 135–141. 10.2478/cszma-2019-0013 [DOI] [Google Scholar]
- Jiménez JA, Cano MJ, Guerra J. (2022) A multilocus phylogeny of the moss genus Didymodon and allied genera (Pottiaceae): Generic delimitations and their implications for systematics. Journal of Systematics and Evolution 60(2): 281–304. 10.1111/jse.12735 [DOI] [Google Scholar]
- Kalyaanamoorthy S, Minh BQ, Wong TKF, von Haeseler A, Jermiin LS. (2017) ModelFinder: Fast model selection for accurate phylogenetic estimates. Nature Methods 14(6): 587–589. 10.1038/nmeth.4285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kazutaka K, Daron MS. (2013) MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Molecular Biology and Evolution 30(4): 772–780. 10.1093/molbev/mst010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kou J, Feng C. (2018) Cheniazanderi C.Feng & J.Kou (Pottiaceae, Musci), a new species from Yunnan, China. Journal of Bryology 40(1): 39–44. 10.1080/03736687.2017.1408872 [DOI] [Google Scholar]
- Kou J, Feng L, Feng C. (2016a) Didymodoncanoae (Pottiaceae), a new moss species from Inner Mongolia, China. Annales Botanici Fennici 53(1–2): 27–30. 10.5735/085.053.0205 [DOI] [Google Scholar]
- Kou J, Feng C, Song S-S, Yu C-Q, Shao XM. (2016b) Hilpertiatibetica J.Kou, X.-M.Shao & C.Feng (Pottiaceae), a new species from Tibet, China. Journal of Bryology 38(1): 28–32. 10.1179/1743282015Y.0000000027 [DOI] [Google Scholar]
- Kou J, Feng C, Shao X-M. (2018) Didymodonobtusus (Bryophyta, Pottiaceae), a new species from Tibet, China. Phytotaxa 372(1): 97–103. 10.11646/phytotaxa.372.1.8 [DOI] [Google Scholar]
- Kou J, Zander RH, Feng C. (2019) Didymodondaqingii (Pottiaceae, Bryophyta), a new species from Inner Mongolia, China. Annales Botanici Fennici 56(1–3): 87–93. 10.5735/085.056.0113 [DOI] [Google Scholar]
- Kučera J, Ignatov MS. (2015) Revision of phylogenetic relationships of Didymodonsect.Rufiduli (Pottiaceae, Musci). Arctoa 24(1): 79–97. 10.15298/arctoa.24.11 [DOI] [Google Scholar]
- Kučera J, Blockeel TL, Erzberger P, Papp B, Soldan Z, Vellak K, Werner O, Ros RM. (2018) The Didymodontophaceus complex (Pottiaceae, Bryophyta) revisited: New data support the subspecific rank of currently recognized species. Cryptogamie. Bryologie 39(2): 241–257. 10.7872/cryb/v39.iss2.2018.241 [DOI] [Google Scholar]
- Li X-J, He S, Iwatsukiz Z. (2001) Pottiaceae. In: Li X-J, Crosby MR. (Eds) Moss flora of China.English Version (Vol. 2.) Science Press, Beijing and Missouri Botanical Garden Press, St. Louis, 114–249.
- Li Y-X, Ji M, Cao G-X, Xing Y-K, Yang Y-W, Wang Z-B, Zhao Z-J, Huo F-L, Zhao D-Y, Bai G-W. (2021) Study on soil physical and chemical properties of typical forest vegetation in Manhan Mountains. Journal of Inner Mongolia Forestry Science & Technology 47(2): 24–29. [In Chinese] [Google Scholar]
- Lyu J-B, Zhang Q-L, Yu N-N, Wang D-Z, Mo R-G, Zhang Z-H. (2012) The impacts of intermediate felling on the soil physicochemical characteristics and microorganism of Chinese Pine and Larix spp in Manhan Mountains. Forest Resources Management 4: 74–79[+85]. [In Chinese] 10.13466/j.cnki.lyzygl.2012.04.007 [DOI]
- Miao B-L. (2017) Vegetation dynamics and its response to climate change in arid and semi-arid areas – a case study of Inner Mongolia. Master’s thesis, Inner Mongolia University, China. [In Chinese]
- Nguyen L-T, Schmidt HA, von Haeseler A, Minh BQ. (2014) IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Molecular Biology and Evolution 32(1): 268–274. 10.1093/molbev/msu300 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osman IB, Hugonnot V, Daoud-Bouattour A, Muller SD. (2021) New bryophytes in Tunisia (North Africa). Part 1: Pottiaceae. Nova Hedwigia 113(1–2): 45–59. 10.1127/nova_hedwigia/2021/0645 [DOI] [Google Scholar]
- Rambaut A. (2014) FigTree, v1.4.2: Tree figure drawing tool. Molecular evolution, phylogenetics and epidemiology. https://github.com/rambaut/figtree/releases
- Ronikier M, Saługa M, Jiménez JA, Ochyra R, Stryjak‐Bogacka M. (2018) Multilocus DNA analysis supports Didymodongelidus (Musci, Pottiaceae) as a distinct endemic of the austral polar region. Acta Societatis Botanicorum Poloniae 87(4): e3609. 10.5586/asbp.3609 [DOI]
- Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Höhna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP. (2012) MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology 61(3): 539–542. 10.1093/sysbio/sys029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shuayib Y, Mamtimin S, Zhao D-P. (2017) Didymodonvalidus Limpr., new to moss flora of China. Xibei Zhiwu Xuebao 37(5): 1038–1041. 10.7606/j.issn.1000-4025.2017.05.1038 [DOI] [Google Scholar]
- Sollman P, Kučera J, Shevock JR, Ma W-Z, Long DG. (2020) Additions and taxonomic insights regarding several rarely or newly reported Pottiaceous mosses of China primarily from the Hengduan Mountains, Easternmost Himalayas. Chenia 14: 1–44. [Google Scholar]
- Wang QH, Jia Y, Liu Y, Chen ZD. (2010) The systematic position of Meteoriella S. Okamura (Musci) based on molecular and morphological data. Taxon 59(1): 93–100. 10.1002/tax.591010 [DOI] [Google Scholar]
- Werner O, Jiménez JA, Ros RM. (2004) The systematic position of the moss Kingiobryumparamicola (Pottiaceae) based on molecular and morphological data. The Bryologist 107(2): 215–221. 10.1639/0007-2745(2004)107[0215:TSPOTM]2.0.CO;2 [DOI]
- Werner O, Jiménez JA, Ros RM, Cano MJ, Guerra J. (2005) Preliminary investigation of the systematics of Didymodon (Pottiaceae, Musci) based on nrITS sequence data. Systematic Botany 30(3): 461–470. 10.1600/0363644054782198 [DOI] [Google Scholar]
- Werner O, Köckinger H, Jiménez JA, Ros RM. (2009) Molecular and morphological studies on the Didymodontophaceus complex. Plant Biosystems 143(Supp 1): S136–S145. 10.1080/11263500903226965 [DOI]
- Wu X-H, Cao Y-F, Chen S-H. (2005) The change of grassland ecological environment in Inner Mongolia and their dynamics response to climate change. Huabei Nongxuebao 20: 65–68. [In Chinese] [Google Scholar]
- Zander RH. (1978) New combinations in Didymodon (Musci) and a key to the taxa in North America north of Mexico. Phytologia 41(1): 11–32. 10.5962/bhl.part.20773 [DOI] [Google Scholar]
- Zander RH. (1993) Genera of the Pottiaceae: Mosses of harsh environments. Bulletin of the Buffalo Society of Natural Sciences 32: 1–378. [Google Scholar]
- Zander RH. (1998) A phylogrammatic evolutionary analysis of the moss genus Didymodon in North America north of Mexico. Bulletin of the Buffalo Society of Natural Sciences 36: 81–115. [Google Scholar]
- Zander RH. (2007) Didymodon Hedwig. In: Flora of North America Editorial Committee (Eds) Flora of North America north of Mexico (Vol 27).Oxford University Press, New York, 539–561.
- Zander RH. (2013) A Framework for Post-phylogenetic Systematics. Zetetic Publications, St. Louis.
- Zander RH. (2019) Macroevolutionary versus molecular analysis: Systematics of the Didymodon segregates Aithobryum, Exobryum and Fuscobryum (Pottiaceae). Hattoria 10: 1–38. 10.18968/hattoria.10.0_1 [DOI] [Google Scholar]
- Zhang X, Zhang Y, Gao L. (2017) A study on control of rodents in forest region of Manhanshan Mountain. Animal Husbandry and Feed Science 38(9): 63–67. [In Chinese]. 10.16003/j.cnki.issn1672-5190.2017.09.020 [DOI]
- Zhang G-L, Feng C, Kou J, Han Y, Zhang Y, Xiao H-X. (in press) Phylogeny and divergence time estimation of the genus Didymodon (Pottiaceae) based on nuclear and chloroplast markers. Journal of Systematics and Evolution. 10.1111/jse.12831 [DOI]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.