

Abstract
Extensive research has examined arsenic (As) bioavailability in contaminated soils and is routinely assessed using in vitro bioaccessibility (IVBA) assays. Analysis of differences in bioaccessibility measurements across IVBA assays and phases is expected to provide valuable insights into geochemical mechanisms controlling soil As bioaccessibility and bioavailability. Soil iron (Fe) content and As speciation are expected to significantly influence IVBA gastric and intestinal phases due to fluctuations in precipitation-dissolution chemistry and sorption reactivity as pH and assay chemical complexity changes. The aim of this review was to examine these relationships by 1) conducting a meta-analysis (n = 47 soils) determining the influence of total Fe on As bioaccessibility measurements and 5 IVBA assays and 2) investigating the effect of As speciation on gastric/intestinal phase IVBA and in vitro-in vivo correlations. Our findings indicate that soil Fe content and As speciation heterogeneity are important in elucidating variability of bioaccessibility measurements across IVBA assays and gastrointestinal phases. Greater focus on coupled As speciation and Fe precipitation chemistry may (1) improve our understanding of soil geochemical factors and assay constituents that influence As in vitro-in vivo correlations and (2) resolve variability in the precision of oral relative bioavailability (RBA) estimated using IVBA assays for soils possessing heterogenous As speciation and Fe composition.
Keywords: Arsenic, bioaccessibility, bioavailability, coprecipitation, iron, meta-analysis, sorption, speciation
Arsenic as a soil contaminant
Arsenic (As) is a geogenic and anthropogenic-released contaminant that has been the focus of extensive global research efforts (Monteiro de Oliveira et al. 2021). Arsenic is a metalloid group 1 carcinogen and the most frequently occurring contaminant on the priority list of hazardous substances (Bradham et al. 2018; WHO 2019). Arsenic exposure is known to be associated with multiple diseases such as diabetes, cardiovascular disease, reproductive dysfunctions, anemia, and cancer (Argos et al. 2010; Cohen et al. 2013; da Silva et al. 2017; Golub, Macintosh, and Baumrind 1998; Kim et al. 2013; Medina et al. 2017; Schuhmacher–Wolz et al. 2009; Vahter 2008). Aqueous As exposure may facilitate the highest degree of increased disease burden as As is commonly sequestered in soils; however, As sequestration mechanisms may be disrupted during changing biochemical conditions upon ingestion (Eggleton and Thomas 2004; Nelson et al. 2018; Sparks 2003), potentially facilitating increased exposure and subsequent toxicological effects. Due to the potential risks associated with exposure to As, investigating As sources and biogeochemical As controls such as pH, redox potential, and soil mineralogy dictating environmental and biological mobility is critical (Fendorf et al. 2010; Griggs et al. 2021; Mandal and Suzuki 2002; Smedley and Kinniburgh 2002).
Understanding As bioavailability is paramount to estimating health risks associated with soil As exposure. Oral bioavailability (absolute bioavailability, ABA) refers to the portion of the oral As dose that is absorbed from the gastrointestinal tract (GIT) into the systemic circulation. Arsenic ABA can be described as the product of two processes: bioaccessibility and absorptive transport (Bradham et al. 2011; NRC 2003). Soil As becomes bioaccessible when released from soil particles in a form that can be transported across the intestinal barrier (Ruby et al. 1999). Current literature of physiological transport of As in the GIT identified water-soluble arsenate (anionic AsVO4) and arsenite (neutral AsIIIO3) as the primary transportable species in mammals (Bradham et al. 2018). Bioaccessibility is measured via in vitro bioaccessibility (IVBA) assays that have been designed to simulate the dissolution of soil As in the GIT of animals. IVBA assays offer an inexpensive, expedient method for quantifying the bioaccessible fraction of the soil As dose (Griggs et al. 2021; Nelson et al. 2018). Relative bioavailability (RBA) of As in soil refers to the ratio of the ABA of As in soil to the ABA of a highly soluble reference form of As (sodium arsenate; Bradham et al. 2011; Brattin and Casteel 2013). RBA is directly measured through in vivo animal bioassays, typically via mouse or swine models, and are used as a biological surrogate for humans (Bradham et al. 2011; Brattin and Casteel 2013; Li et al. 2021). Animal models of soil As RBA provided the empirical basis for regression models relating soil As RBA and IVBA (Brattin et al. 2013; Diamond et al. 2016). The strong correlation observed between IVBA and RBA (in vitro-in vivo correlations, IVIVC) demonstrates the importance of bioaccessibility in determining the oral bioavailability of soil As (Bradham et al. 2018). For a relatively large data set of 83 soils, IVBA explained approximately 90% of the variance in RBA; however, absolute residuals for some soils were as high as 34% (Diamond et al. 2016). Thus, while the IVIVC performed superbly for predicting the relationship between IVBA and RBA for the dataset, uncertainty in predicting RBA for any single future soil from its corresponding IVBA would be larger (prediction interval ± 19%). It is likely that many factors contribute to the prediction interval of the As IVIVC, which has prompted extensive research on elucidating the geochemical and physiological determinants of bioaccessibility, with IVBA assays as the primary research model (Bradham et al. 2018; Griggs et al. 2021). Of these factors, soil Fe is expected to be a major variable as Fe phases ubiquitously sequester As; however, the effects of this sequestration process on bioaccessibility and bioavailability are poorly resolved.
Many unknowns remain regarding biogeochemical drivers of bioaccessibility variability observed across soils and IVBA assays. Therefore, the aim of this review was to 1) assess the influence of total soil Fe and/or As-Fe sorption characteristics on As bioaccessibility across 5 IVBA extractant chemistries, 2) discuss the effect of As speciation on gastric and intestinal phase IVBA and bioaccessibility correlations with bioavailability measurements, and 3) provide a framework for future research investigating soil chemical drivers of As bioaccessibility and influences on IVIVC. A meta-analysis was conducted of previously published As bioaccessibility data for 47 soils across 5 IVBA methods to investigate connections between soil Fe content, assay chemistry, and As speciation that affect bioaccessibility measurements and IVIVC. Although datasets containing paired bioaccessibility or bioavailability data and As speciation are limited, this review presents the current state of science while identifying avenues of research warranting future investigations.
Arsenic chemistry and common stabilization reactions with soils
Arsenic chemistry and the surrounding soil biogeochemical conditions not only mediate As cycling in soil but may also influence bioavailability of soil As upon ingestion. To better understand variability in As IVBA and RBA measurements across soils (Figure 1), relationships between geochemical conditions including pH, redox, and soil mineralogy need to be understood in the context of As speciation. Here and throughout, bioaccessibility refers to in vitro measurements collected via IVBA extraction whereas bioavailability refers to in vivo measurements that are representative of the mouse or swine model, typically reported as RBA.
Figure 1.
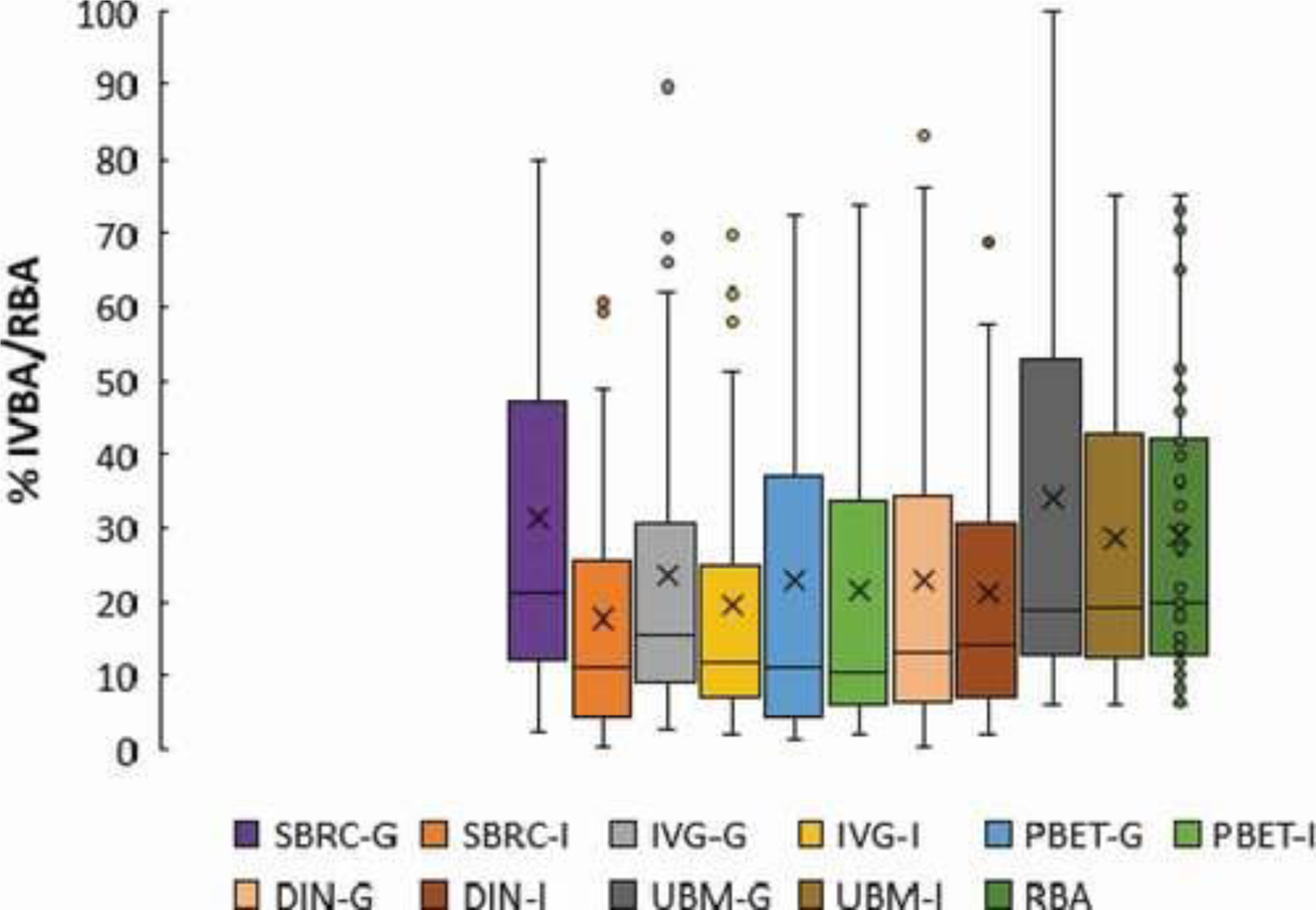
Box and whisker plots showing variability in mean (x), median (horizontal line), lower and upper quartile (end of boxes) and extreme (whiskers) % in vitro bioaccessibility (IVBA) values (by assay and phase), and % relative bioavailability (RBA) measurements for the 47 soils included in the meta-analysis. All IVBA assays shown include data for the gastric (-G) and intestinal (-I) assay phases
Chemistry of inorganic arsenic species
Pentavalent arsenate (As(V)) and trivalent arsenite (As(III)) are the predominant inorganic As species commonly found in soils (Fendorf et al. 2010; Sparks 2003). Redox interactions mediate the distribution of each species, with As(V) typically being predominant in oxygenated, well-drained conditions (Khalid et al. 2017; Kuivenhoven and Mason 2019; Lim, Shukor, and Wasoh 2014; Marlborough and Wilson 2015; Thouin et al. 2016). Arsenate is negatively charged across most environmentally relevant pH ranges (pH >2) (Akter et al. 2005; Fendorf et al. 2010), making As(V) preferentially sequestered by positively charged surfaces in soils (Chappell, Chiswell, and Olszowy 1995; Marlborough and Wilson 2015). For example, Fe(III) (oxyhydr)oxides below the point of zero charge, the pH at which mineral variable surface charge is neutral, have positive surface charge with high affinity for oxyanions such as arsenate and phosphate (Sparks 2003). Conversely, negatively charged surfaces such as clay minerals with permanently negative charge via isomorphic substitution, organic matter, and other soil constituents may drive mobility of As(V) introduced into the soil system (Strawn, Bohn, and O’Connor 2019), limiting soil As sequestration. However, As(V) typically remains sequestered in soils due to strong sorption interactions with positively charged metal oxide minerals and ligand exchange processes (Fendorf et al. 2010; Strawn, Bohn, and O’Connor 2019). Arsenic associated with amorphous metal oxide minerals was found to exhibit higher bioaccessibility, whereas crystalline phases generally possess lower bioaccessibility (Kim, Yoo, and Baek 2014; Meunier et al. 2010; Palumbo-Roe and Klinck 2007) (Figure 2). However, the effect of metal oxide interactions with specific As species on soil As bioaccessibility is poorly understood.
Figure 2.

Conceptual model showcasing expected bioaccessibility of As phases ranging from least to greatest based on current literature (left, middle; adapted from Meunier et al. 2010). We then compare this to a range of processes that may facilitate changes in bioaccessibility measurements (right) but require further investigation
Reducing environmental conditions (~100 mV at circumneutral pH) facilitate conversion of As(V) to As(III) (Akter et al. 2005; Sowers et al. 2017), with reduction commonly facilitated by microbial activity, redox-active minerals such as manganese oxides, and/or organic matter in suboxic to anoxic environments (inundated soils) (Fendorf and Kocar 2009; Gillispie et al. 2015). Unlike As(V), As(III) is neutral at common soil pH conditions (negatively charged ~ pH ≥9) which drives increased mobility in soil systems and exposure risks for biota (Akter et al. 2005). Speciation of As mediates environmental mobility and stability when exposed to changes in surrounding chemical conditions (Abbas et al. 2018; Gillispie et al. 2015; Smedley and Kinniburgh 2002; Sparks 2003), making As(V) and As(III) sequestration critical to mitigating toxicological effects.
Mineral phases
Precipitation of As mineral phases readily form with Fe as pH and/or redox conditions change; however, precipitation pathways may be As species dependent. Ions precipitating with As(V) are typically dictated by pH conditions. At basic conditions, calcium and magnesium, in addition to other basic cations, may precipitate with As(V), whereas polyvalent Al and Fe(III) cations may be favored at acidic pH conditions (Fendorf et al. 2010). Substitution of As(V) in minerals containing oxyanions of similar size such as phosphate is another significant fraction of As(V) containing minerals. Sulfides are a major driver of As(III) solubility and are highly influenced by sulfidic precipitates (Fendorf et al. 2010). Sulfur rich, reducing conditions tend to favor As(III) mineral precipitation and may be preceded by metal sulfide sorption of As(III).
Numerous investigators demonstrated that mineral solubility is positively correlated with As bioaccessibility and bioavailability, with current knowledge of As species and bioaccessibility illustrated in Figure 2 (Meunier et al. 2010; Ollson et al. 2016; Smith and Steinmaus 2009; Stevens et al. 2018). Arsenic minerals that are known to exhibit low bioaccessibility include As-sulfides (realgar, orpiment, and arsenopyrite) and Fe arsenates (scorodite and kankite) (Karna et al. 2017; Kim, Yoo, and Baek 2014; Meunier, Koch, and Reimer 2011; Meunier et al. 2010; Toujaguez et al. 2013; Whitacre et al. 2017). Conversely, calcium-arsenate is an As mineral that has been shown as highly soluble under gastric-like conditions (Meunier et al. 2010; Whitacre et al. 2017). The bioaccessibilities of the aforementioned As mineral phases for both species are sensible given that As minerals that typically form at acidic pH, such as arsenopyrite and scorodite, are resistant to destabilization following exposure to acidic gastric conditions; whereas those typically formed at basic conditions (calcium arsenate) are preferentially solubilized. Therefore, recognizing typical precipitation conditions of specific As minerals is important for assessing expected bioaccessibility. Mineralogy of Fe may also be critically important for bioaccessibility estimates (Ollson et al. 2016). Soils from a gold mine tailings system were found to possess higher As bioaccessibility when associated with goethite and amorphous Fe compared to lower As bioaccessibility for arsenopyrite and scorodite (Toujaguez et al. 2013). In addition, Karna et al. (2017 found that Pb substituted jarosite (plumbojarosite; Pb2+0.5Fe3+3(OH)6(SO4)2)) resulted in low Pb bioaccessibility (1–10% IVBA) and bioavailability (RBA <10%). Arsenate is also capable of substitution into jarosite (Paktunc and Dutrizac 2003), suggesting that As-substituted jarosite may promote low bioaccessibility and/or bioavailability (Figure 2). Both examples showcase the need to understand Fe mineralogy, as Fe mineralogy is expected to mediate As mineral stability and surface sorption processes in soils.
Sorbed phases
Metal oxide sorption and coprecipitation processes are major sources of As immobilization (Fendorf et al. 2010; Gillispie et al. 2015; Sparks 2003). As(V) and As(III) may bind to a variety of soil minerals with Fe(III) metal oxides typically exhibiting the highest adsorption maximum for both species (Dixit and Hering 2003) (Figure 3). Sorption mechanism(s) dictate the stability of sequestered As species, representing the fraction of total As that is either resilient to desorbing conditions or readily mobilized in unfavorable conditions. Both As(V) and As(III) may sorb to metal oxides via high strength inner sphere complexes involving ligand exchange; whereas As(III) may also participate in relatively weaker outer sphere complexation that relies on electrostatic differences (Fendorf et al. 2010; Gillispie et al. 2015; Goldberg and Johnston 2001; Sparks 2003; Waychunas et al. 1993). Inner sphere complexation is defined as a sorption process involving ionic or covalent bonding to a sorbent such as a metal oxide mineral surface, whereas outer sphere complexation is an electrostatic-driven process where a chemical does not form a direct bond with the sorbent (Strawn, Bohn, and O’Connor 2019). Therefore, inner sphere complexation is markedly stronger and more resistant to destabilization compared to outer sphere complexation processes. Surface-sorbed As(V) on Fe and Al oxides are dominated by bidentate, binuclear (double-corner sharing) inner sphere complexes (Fendorf et al. 2010). Goldberg and Johnston (2001) confirmed that As(V) forms inner-sphere complexes on amorphous Al and Fe oxides, whereas As(III) forms both inner- and outer-sphere complexes on amorphous Fe oxide, and solely outer-sphere complexes on amorphous Al oxides (Goldberg and Johnston 2001). Partial outer sphere complexation of As(V) to hematite was reported by Catalano et al. (2008), but As(V) is still expected to predominantly participate in inner sphere complex reactions with most metal oxide minerals. Sequestration of As oxyanions may be interrupted and/or destabilized in the presence of competing ions (Strawn, Bohn, and O’Connor 2019). Fendorf et al. (2010) showed that phosphate oxyanions are atomically similar to arsenate and bind via similar mechanisms; therefore, the presence of phosphate is often expected to facilitate decreased As sorption.
Figure 3.

Magnitude and pH-dependency of As(V) (arsenate – ∆) and as(III) (arsenite – ○) adsorption to ferrihydrite (green) and goethite (yellow) at an initial concentration of 100 mM (7.5 g As/L) as (data taken from Dixit and Hering 2003). Sorbent concentration was 0.03 g L−1 hydrous ferric oxide and 0.5 g L−1 goethite compared to the IVBA pH ranges of the gastric and intestinal phases
Coprecipitation reactions of Fe with As commonly occur in soil systems with fluctuating redox and/or pH conditions and are expected to affect As availability along the GIT. Coprecipitation is the process of an aqueous metal precipitating in the presence of another aqueous constituent such as dissolved organic C and/or As (Sparks 2003). Coprecipitation of ferrihydrite with natural organic matter (NOM) is considered as a key factor influencing As cycling in soils (Kim, Hwang, and Baek 2015). NOM may influence adsorption, oxidation, and speciation of As at Fe (oxyhydr)oxide surfaces (Xue et al. 2019). NOM may inhibit the adsorption of As(V), especially at pH values greater than 4.5, which interferes with maximum sorption potential, ultimately resulting in an abundance of easily solubilized As in the soil (Kim, Hwang, and Baek 2015; Sparks 2003). These results suggest that As bioaccessibility may increase in the presence of NOM. However, effects of NOM on As(V) and As(III) bioaccessibility and bioavailability are currently under-investigated, warranting future IVBA studies. Further, the effect of Ca and/or CaCO3-associated As phases on bioaccessibility and bioavailability measurements may be more complicated than previously thought. Although As phases associated with Ca and/or CaCO3 are typically expected to be highly bioaccessible and bioavailable due to dissolution in the acidic conditions of the gastric phase, preliminary investigations by our research group found that bioaccessibility did not resolve RBA for high pH soils containing As-CaCO3 coprecipitates and/or As sorbed to Ca and/or carbonate phases, potentially sorbed via Ca-bridging complexes (Personal communication). Antelo, Arce, and Fiol (2015) reported that calcium bridging influenced As sorption to ferrihydrite (Fe(III) oxyhydroxide), emphasizing the importance of probing the stability of these complexes. This study is preliminary but suggests that the role of these phases may be overly simplified in the current state of science (Figure 2).
Background on As bioaccessibility and bioavailability assays
Over the past 25 years, various IVBA methods have been developed to estimate RBA through in vivo-in vitro correlations (IVIVCs). Validation and regulatory acceptance criteria developed by the U.S. Environmental Protection Agency (USEPA 2007), as adapted from the Interagency Coordinating Committee on the Validation of Alternative Methods (ICCVAM), have been applied to an in vitro bioaccessibility (IVBA) assay as a reliable predictor of RBA. This IVBA assay, Solubility Bioaccessibility Research Consortium (SBRC) assay, is now part of EPA’s Test Methods for Evaluating Solid Waste: Physical/Chemical Methods SW-846 as EPA Method 1340 (EPA 2017).
Other common IVBA assays include, but are not limited to, the following: Physiologically Based Extraction Test (PBET) (Ruby et al. 1996), In Vitro Gastrointestinal extraction method (IVG) (Rodriguez et al. 1999), Deutsches Institut für Normunge.V. (DIN) (Hack and Selenka 1996; Rotard et al. 1995), Unified Bioaccessibility Research Group of Europe Method (UBM) (DIN 2000; Oomen et al. 2002; Wragg et al. 2011), and Simulator of the Human Intestinal Microbial Ecosystem (SHIME) (Van de Wiele et al. 2007, 2015). These assays differ in many ways: (1) chemical composition of digestive fluids, (2) number of digestive components used (i.e. inclusion of a saliva phase, gastric phase (GP), intestinal phase (IP), and/or colon phase), (3) biogeochemistry (i.e. pH, Eh, chemical amendments, solids-to-solution ratio, food component), and (4) incubation time (Table 1). In addition, GP and IP IVBA extractions may be run sequentially. However, detailed understanding of complex relationships between As chemistry, assay constituents, and soil geochemical properties, and how these relationships influence assay-specific IVIVCs, is limited. Methodological differences between assays have been known to result in variability in contaminant bioaccessibility measurements (Bradham et al. 2018; Griggs et al. 2021; Juhasz et al. 2014, 2015; Li et al. 2015; Oomen et al. 2002; Rodriguez et al. 1999; Smith et al. 2014). For example, differences in soil-to-solution ratio between assays have been attributed to variations in bioaccessibility %. As the ratio decreases, bioaccessibility increases or remains unchanged (Yang et al. 2003) except in the case of glycine-based gastric solutions (SBRC) in which increases in liquid-to-soil ratios have been shown to result in gastric phase As bioaccessibility being greater than that of glycine-free GI fluids due to Fe-glycine interactions (Meunier et al. 2010). Variations also exist within phases of the same assay such as gastric vs. intestinal phases (Denys et al. 2012; Juhasz et al. 2014, 2007; Wragg and Cave 2003; Wragg et al. 2011).
Table 1.
Five prominent and/or recently developed IVBA assays. Digestive phases, sample mass required, digestive constitutes used, pH of digestive phases, food component inclusion, sample solid to digestive solution ratio, phase time, and experiment temperature for each assay are showcased
| Assay | Phase | Sample Mass (g) | Digestive constituent(s) | pH | Food* | Solid/Solution ratio | Incubation time (h) | Temperature (°C) |
|---|---|---|---|---|---|---|---|---|
| SBRC/ EPA Method 1340 |
Gastric | 1 | 30.03 g glycine | 1.5 | No | 1:100 | 1 | 37 |
| Intestinal | 1.75 g bile, 0.5 g pancreatin | 7 | 4 | |||||
| PBET | Gastric | 0.4 | 0.5 g malate, 0.5 g citrate, 420 µL lactic acid, 500 µL acetic acid, 1.25 g pepsin | 1.3, 2.5, & 4** | No | 1:160 | 1 | 37 |
| Intestinal | 70 mg porcine bile salts, 20 mg porcine pancreatin | 7 | 4 | |||||
| IVG | Gastric | 4 | 8.77 g 0.15 M NaCl, 10 g 1% porcine pepsin | 1.8 | Yes; 200 g Dough | 1:150 | 1 | 37 |
| Intestinal | 1.75 g porcine 0.035% bile and 0.5 g 0.35% pancreatin | 5.5 | 1 | |||||
| DIN | Gastric | 2 | 1 g pepsin, 3 g mucin, 2.9 g NaCl, 0.7 g KCl, 0.27 g KH2PO4 | 2 | Yes; 50 g Dried Whole Milk Powder | 1:50 | 2 | 37 |
| Intestinal | 9 g bile, 9 g pancreatin, 0.3 g, 0.15 M trypsin | 7.5 | 1:100 | 6 | ||||
| UBM | Saliva | 0.6 | 9 mL synthetic saliva fluid | 6.5 (± 0.5) | No | 1:37.5 | 5 min*** | 37 |
| Gastric | 13.5 mL synthetic gastric fluid | 1.2 (± 0.2) | 1:100 | 1 | ||||
| Intestinal | 27 mL synthetic duodenal, 9 mL synthetic bile* | 6.3 (± 0.2) | 4 |
Food component is often optional.
pH 1.3 (fasting), 2.5 (food ingestion), 4 (emptying).
Saliva added to the GP after 5 minutes.
Past studies evaluated bioaccessibility measurements in the same set of soils across multiple IVBA assays. While emphasis of these studies has generally been the assessment of IVIVCs across assays, data presented also enable evaluation of soil and assay geochemical mechanisms that may influence bioaccessibility and/or bioavailability. Individually, past studies were limited in the number of soils examined to explore such mechanisms. Therefore, a meta-analysis was conducted to compile a larger set of soils to explore possible geochemical mechanisms influencing bioaccessibility measurements across assays and assay phases. Collectively, 47 soils from 4 previously published studies (Juhasz et al. 2014, 2015, 2009; Li et al. 2015) were included in the meta-analysis, with the analysis focusing on assessing variability in bioaccessibility measurements across assays and assay phases, and how select soil geochemical properties reported in these studies (i.e., total Fe and As speciation) influenced IVBA and IVIVC variability. These studies were selected because As IVBA was assessed across the same five assays (SBRC, IVG, PBET, DIN, and UBM), with the exception of Juhasz et al. (2009), which did not include the UBM assay. Total Fe was reported in all four studies; however, only one study (Juhasz et al. 2014) provided As speciation data. Variability in IVBA and RBA measurements by assay and phase across the 47 soils included in the 4 studies is shown in Figure 1. Additional analyses conducted as part of the meta-analysis included evaluating change in % IVBA from gastric to intestinal phase within an assay. Assay-specific relationships between total soil Fe and 1) change in IVBA from gastric to intestinal phase, and 2) variability in IVBA measurement in the same soil across IVBA assays were also explored. Finally, variability in RBA prediction based upon soil As speciation was evaluated for the 10 soils where As speciation data were provided.
Influence of biogeochemical factors on Asenvironmental mobility and GIT solubility
Soil chemical and physical properties previously discussed are expected to significantly mediate As bioaccessibility and bioavailability. Primary drivers dictating soil As mobility include As speciation, Eh-pH conditions, As mineral phases, and metal oxides present (Borch et al. 2010; Bowell 1994; Nelson et al. 2018; Raven, Jain, and Loeppert 1998; Sparks 2003; Sposito 2008; Wilkie and Hering 1996). Of these properties, the connection of As(V) and As(III) to Fe(III) (oxyhydr)oxides is especially important when assessing biogeochemical constraints of As bioaccessibility and bioavailability. As discussed earlier, Fe(III) (oxyhydr)oxides are high-affinity sorbents of As(V) and As(III) ubiquitously found in soils (Gamble, Givens, and Sparks 2018); however, fluctuating biogeochemical conditions such as acidic pH, reducing conditions, and competing ions may promote enhanced As availability in soils (Borch et al. 2010; Deng et al. 2020; Gillispie et al. 2015; Sowers et al. 2017; Voegelin, Weber, and Kretzschmar 2007) and may impact bioaccessibility and bioavailability measurements. Identifying the relationship of soil Fe and As speciation under changing IVBA conditions is crucial to discovering mediators of bioaccessibility and bioavailability.
pH as a key variable governing bioaccessibility measurements
The complex relationship of pH, As, and Fe (oxyhydr)oxides has been extensively studied in soil systems; however, this process is also of critical importance when estimating As release along the GI tract. Stability of As minerals and As-bearing Fe (oxyhydr)oxides are associated with pH conditions. Ingested As-bearing soils are exposed to highly acidic conditions along the GI tract (gastric phase pH = 1.2–4; Table 1) (Bradham et al. 2018), potentially facilitating destabilization of previously sequestered As and absorption by the body. This process may be well modeled via bioaccessibility assays, with assay gastric phases showing increased As solubilization as As-bearing Fe (oxyhydr)oxide phases undergo dissolution (Juhasz et al. 2007; Ollson et al. 2016; Rodriguez et al. 1999; Ruby et al. 1996). Dissolved Fe may reprecipitate in the subsequent intestinal phase as pH elevates to the circumneutral range (Cornell and Schwertmann 2003; Sparks 2003), potentially promoting As–Fe (co-)precipitation phenomenon that may result in As bioaccessibility decrease when comparing GP to IP (Juhasz et al. 2014, 2015). Therefore, pH is commonly cited as a driving force in determining As bioaccessibility for contaminated soils (Brattin et al. 2013; Juhasz et al. 2015; Yang et al. 2002).
Results of the meta-analysis with respect to exploring changes in IVBA when comparing assay gastric and intestinal phases are illustrated in Figure 4. For the SBRC assay, all 47 soils demonstrated reduction in bioaccessibility from gastric to intestinal phase. This significant reduction in SBRC-estimated As bioaccessibility may be driven, in part, by the larger difference in pH of the GP (pH = 1.5) and IP (pH = 7) in the SBRC assay relative to other IVBA assays (Table 1). Increasing pH from the gastric to the intestinal phase promotes precipitation of aqueous Fe, potentially sequestering previously aqueous As. Furthermore, Juhasz et al. (2015) reported that Fe concentration in the SBRC assays decreased 23-fold from the gastric to the intestinal phase, supporting the conclusion that As-Fe coprecipitation was likely. Conversely, in PBET, DIN, and UBM assays, approximately half the soils demonstrated a rise in IVBA in the intestinal phase and half exhibited a fall, with the median change near zero. Data for the IVG assay generally favored an IVBA reduction in the intestinal phase; however, soils still ranged from positive to negative % change when comparing gastric to intestinal phase IVBA. The presence of Fe-chelating organic acids and increased chemical complexity of these four assays compared to more chemically “simple” SBRC method might resolve observed differences in alterations in IVBA from gastric to intestinal phases by assay (Figure 4). Chelation of Fe in the presence of diverse organic moieties may inhibit coprecipitation of As and Fe (Juhasz et al. 2015) in the intestinal phase. Less Fe available for precipitation and sorption reactions with As is expected to have resulted in elevated As in solution, increasing estimated bioaccessibility for these assays. This trend is supported by our meta-analysis, where IP bioaccessibility measured in the SBRC bioaccessibility is less likely to be affected by chelation effects than in the other four assays analyzed.
Figure 4.

Change in % IVBA from gastric to intestinal phase across the 5 IVBA assays evaluated (n = 47 soil for SBRC, IVG, PBET, and DIN; n = 35 soils for UBM). Positive values represent an increase in IVBA from gastric to intestinal phase within an assay, whereas negative values represent a decrease
Differences in gastric phase pH have been cited as a contributor to variability in bioaccessibility measured in different IVBA assays applied to the same soils (Juhasz et al. 2015; Oomen et al. 2002; Smith et al. 2014). This assertion spawned several investigations that tested the effect of variations in pH on bioaccessibility. Brattin et al. (2013) examined the influence of pH (1.5 to 7.5) in the SBRC IVBA assay on As bioaccessibility in 18 soils. The trend in all soils was a reduction in IVBA with increasing pH, with the highest values noted for all soils at pH 1.5. Ruby et al. (1999) found that As bioaccessibility in two residential soils and house dust collected near a historical copper smelter was threefold lower in the PBET assay when GP pH was 2.5 than compared to pH 1.3 (Ruby et al. 1996, 1999). Similarly, Smith et al. (2014) altered pH in SBRC and PBET assays while investigating 5 As-contaminated brownfield soils and noted SBRC to exhibit the highest IVBA compared to PBET at pH 1.5 and 2.5, although elevated PBET acidity at pH 1.5 did increase observed As bioaccessibility compared to pH 2.5. In summary, pH plays a major role in governing As IVBA measurement, predominantly via pH-driven effects on the dissolution-precipitation chemistry of metal oxide minerals.
Total soil iron (Fe) explains bioaccessibility variability across soils and IVBA assays
In addition to pH, total soil Fe is expected to be an influential soil property driving bioaccessibility estimates. To further explore the role(s) of Fe on gastric and intestinal phase IVBA, the relationship was investigated between total soil Fe and relative % difference (RPD) in within-assay gastric versus intestinal phase IVBA values across each of the 5 IVBA assays examined (Figure 5) (Juhasz et al. 2014, 2015, 2009; Li et al. 2015). RPD between gastric and intestinal phase IVBA, in this context, was calculated as:
where
Figure 5.

Relationship between total soil Fe (x-axis) and relative percent difference (RPD) in As IVBA from gastric to intestinal phase (y-axis) by assay
Increasing soil Fe concentration was associated with more negative RPD values for all 5 IVBA assays evaluated, indicating soils with higher Fe concentration generally displayed a stronger relative decrease in % IVBA from gastric to intestinal phase. The strength of this relationship, however, varied by assay, and was only statistically significant (α = 0.05) for the SBRC, IVG, and PBET assays. SBRC showed the strongest correlation with soil Fe explaining approximately 39% of the variance in RPD. The data suggest that co-precipitation of As with Fe may be a particularly dominant mechanism driving differences between gastric and intestinal-phase %IVBA in the SBRC assay. Weaker correlations between gastric and intestinal phase RPD with increasing Fe were observed for the other assays, which may be attributed to Fe-chelation effects associated with assay-specific constituents limiting coprecipitation in the intestinal phase in these assays.
The manner in which Fe may influence variability in IVBA measurements observed across IVBA assay was also determined. Further, phase by means of Spearman Rank Correlation, in which soil Fe concentration and % IVBA values were first ranked, and then linear regression models fit based upon ranking was also investigated. Spearman Rank Correlation was used because total soil Fe and IVBA values were not normally distributed across the meta-data set. The Spearman correlation coefficients for a given assay and phase ranged from 0.51 to 0.84 (Table 2). Correlation coefficients were generally higher in the intestinal phase for the five assays (mean = 0.7, range = 0.57 to 0.84) than gastric-phase assays (mean = 0.6, range = 0.51 to 0.73). Higher correlation coefficients in IP assays may be explained by the dual roles Fe plays in controlling IP IVBA, where increasing soil Fe content may first mitigate the solubilization of As at GP pH, and then promote enhanced As-Fe coprecipitation at intestinal IP pH. This may also be explained by variations in the reactivity of Fe phases. Soil Fe mineral phases may be more readily solubilized in GP depending upon crystallinity, whereas Fe precipitation in the IP is expected to occur uniformly among all solubilized Fe present. Of particular interest is identifying to what extent Fe modifies bioavailability in vivo. The Spearman’s correlation coefficient between soil Fe and % RBA for the 47 soils evaluated in the meta-analysis was 0.58, more closely matching those detected in gastric-phase systems. The data suggest that co-precipitation of As with Fe noted in intestinal-phase assays may not be as important a factor in controlling As RBA in vivo; however, more research is needed.
Table 2.
Spearman’s rank correlation between total soil Fe and a) % IVBA by IVBA assay and phase and b) RBA
| Gastric | Intestinal | |
|---|---|---|
| SBRC | 0.60 | 0.77 |
| IVG | 0.59 | 0.66 |
| PBET | 0.60 | 0.64 |
| DIN | 0.51 | 0.57 |
| UBM | 0.73 | 0.84 |
| RBA | 0.58 | |
Increasing soil Fe concentration was also positively associated with elevated variability in % IVBA measurements in a given soil across assays and phases, as measured by coefficient of variation (CoV) (Figure 6(a)). Specifically, for the 47 soils evaluated, soil Fe concentration accounted for 44% of variability in COV in measured IVBA values across both IVBA assay and phase (R2 = 0.44). When within-soil IVBA variability was assessed for GP or IP separately, total Fe accounted for 13% and 36% of variability in IVBA, respectively (Figures 6(b,c), respectively). All three relationships (GP and IP combined, gastric-only, and intestinal-only) were significant at α = 0.05. One possible explanation for this observed behavior is differences in dissolution rates across heterogenous Fe mineral phases in the gastric phase, where the relationship between soil Fe and As IVBA may vary across Fe-As minerals. Enhanced mineral crystallinity is linked to diminished surface area; therefore, dissolution rate is expected to decrease with increasing crystallinity. However, the intestinal phase is more dependent upon available aqueous Fe that does not consist of heterogenous phases; therefore, it is conceivable that total Fe better resolves intestinal-phase variability. Characterization of Fe mineralogy, therefore, is expected to be important for understanding the stability of As-bearing Fe (oxyhydr)oxides phases, especially in the gastric phase, and may better resolve IVBA variability compared to total soil Fe concentration.
Figure 6.

Relationship between total soil Fe (x-axis) and coefficient of variation (CoV) in measured soil as IVBA (y-axis) across 10 IVBA assays/phases (5 assays x 2 phases per assay) for a) gastric and intestinal phases (GP and IP, respectively) combined, b) gastric only assays, and c) intestinal only assays)
Arsenic complexation interactions
Sorption interactions of As with reactive minerals ubiquitously found in soil systems such as Fe (oxyhydr)oxides) are essential to As sequestration as surrounding chemical conditions fluctuate (Dixit and Hering 2003; Redman, Macalady, and Ahmann 2002; Reeder, Schoonen, and Lanzirotti 2006; Sparks 2003). Resilience to diverse chemical conditions is especially important when evaluating As bioaccessibility in soils. Arsenic adsorption onto mineral surfaces is strongly affected by pH and exhibits speciation-specific behaviors (Dixit and Hering 2003; Goldberg and Johnston 2001; Sowers et al. 2017). As(V) typically sorbs to metal oxide minerals to a higher extent than As(III) across most soil pH values with As(III) typically sorbing to a greater extent only in highly alkaline soils (pH >8) (Dixit and Hering 2003; Fendorf et al. 2010; Goldberg and Johnston 2001). At pH below the point of zero charge (PZC) of Fe (oxyhydr)oxide (between pH of 7–9) minerals, As(V) commonly sorbs to a higher extent and via more potent bonding mechanisms, such as inner sphere complexation, compared to As(III) (Fendorf et al. 2010; Sowers et al. 2017; Sparks 2003; Waychunas et al. 1993). This strength of sorption and persistence of As(V) was clearly observed in Smith et al., where five Fe-rich soils were analyzed via As XANES after being exposed to SBRC gastric phase (pH 1.5) or modified PBET gastric phase (pH 1.5). For both the original soils and IVBA assay treated soils, As(V) speciation remained consistent for all soils tested (≥88% As(V)) with As minerals also found to be stable (Smith et al. 2014). However, soils with detectable As(III) were not evaluated, exemplifying the need for future investigation of As(III)-containing soils. Smith et al. (2014) stated that PBET organic acids may affect sorption to Fe oxides; however, it remains unknown how organic acids may selectively passivate As mineral sorbents and if these interactions favor reduced sorption of As(V) versus As(III). Similarly, Yang et al. (2002); (2005) reported a significant relationship between As(V) adsorption and Fe content in soils, which generally reduced As(V) bioaccessibility with increasing Fe content. Previously (Juhasz et al. 2014, 2015, 2007; Juhasz, Weber, and Smith 2011; Juhasz et al. 2009) examined this relationship further by determining potential sorption and precipitation mechanisms affecting As bioaccessibility in gastric and intestinal-phase assays. Sorbent stability when exposed to gastric and intestinal-phase assays also needs to be considered, as pH fluctuations and competing ions may drive increases in soluble As.
Although As(III) is frequently more mobile in the environment, information on effects of As speciation on bioaccessibility and bioavailability are lacking in the current literature base, because bioaccessibility investigations have nearly exclusively examined total soil As or been conducted on highly oxidized, As(V)-rich soil systems. Due to the lack of charge of As(III) and decreased strength of sorption at acidic pH (Dixit and Hering 2003; Fendorf et al. 2010; Yang et al. 2012), As(III) may be more readily mobilized from soil particles in the GIT, facilitating enhanced bioaccessibility and bioavailability. In the current literature, limited studies reported soil As speciation and correlated speciation to RBA and/or IVBA%. Investigation of mice and swine models found inorganic As(V) and As(III) to display similar bioavailability, suggesting that once released from the soil matrix in the GIT, these two species may exhibit similar bioavailability (Juhasz et al. 2007). Yang et al. (2005) compared bioaccessibility in aerobic soils spiked with inorganic As(III) and As(V) over a 6-month time period. Initially, bioaccessibility of As(III) spiked soils was significantly higher than that of As(V) spiked soils, likely attributed to greater mobility of As(III) and different interspecies adsorption behaviors in soils. After aging the soils, a significant amount of As(III) was oxidized to As(V) thus reducing As bioaccessibility (Yang et al. 2005). These results are in agreement with current understandings of environmental mobility of As(III) and As(V) and the likelihood that speciation influences on mobility may also persist in IVBA and RBA %. However, this concept needs to be examined in further studies. Yang et al. (2005) postulated these conclusions using solely the PBET assay, and it is not known if results would be consistent across assays due to biogeochemical interactions with differing IVBA constituents. Soil properties such as pH, Fe, Mn, cation exchange capacity and silt content may increase As(III) adsorption (Meunier et al. 2010; Yang et al. 2005); therefore, further investigation of the soils varying in chemical and physical characteristics is essential to discovering the manner in which As(V) and As(III) bioaccessibility may be affected.
Connecting pH, As speciation, and As–Fe interactions to resolve bioaccessibility and bioavailability
The effect of As speciation and IVBA assay chemistry on As sequestration provides a unique window into mechanistic processes underlying As bioaccessibility measurements (Figures 2 and 3). Arsenate and arsenite sorption envelopes (pH 3.5–10.5) for ferrihydrite and goethite from Dixit and Hering (2003) seminal Fe(III) (oxyhydr)oxide As reactivity manuscript are presented in Figure 3, modified to include overlaid IVBA assay and phase pH ranges. As shown in Figure 3, significant differences in As(V) and As(III) sorption with pH may be observed. Arsenate sorption was lowered for both ferrihydrite and goethite with increasingly basic pH, whereas arsenite sorption was elevated with rising pH until approximately pH 7.5. Ferrihydrite As(V) and As(III) sorption capacities are approximately 20- and 10-fold greater, respectively, than goethite; however, both Fe(III) (oxyhydr)oxides possess similar sorption patterns with changing pH (Dixit and Hering 2003; Fendorf et al. 2010). Sorption envelopes for both inorganic As species (Figure 3) visually indicate the critical relationship of As species with assay-dependent pH conditions. Gastric pH conditions may facilitate Fe dissolution conditions (pH <2) dependent upon the Fe phase present and soil characteristic including organic matter content, surface area, and particle size (Juhasz et al. 2014; Sparks 2003). However, As(III) present may be favorably released in this phase compared to As(V) as indicated by twice the sorption maximum of As(V) to ferrihydrite/goethite compared to As(III). Interestingly, the circumneutral approximate 6–7.5 pH conditions of the intestinal phase may promote equally favorable retention of As(V) and As(III). These differences are related to changing surface charge of the Fe(III) (oxyhydr)oxide with fluctuating pH. The PZC for ferrihydrite and goethite typically exists at approximately 6.5 to 7.5 (Cornell and Schwertmann 2003; Juhasz et al. 2014; Sowers et al. 2017; Sparks 2003), indicating that pH below this value results in the sorbent, ferrihydrite, or goethite being positively charged, increasing preference for negatively charged As(V) oxyanions (Sparks 2003; Sposito 2008). Arsenite is neutrally charged at most environmental pH conditions resulting in similarities in sorption of As(V) and As(III) near the PZC (Fendorf et al. 2010; Sparks 2003; Sposito 2008); therefore, As(V) and As(III) may behave similarly in the IP. Understanding the relationship between soil, IVBA, and/or stomach pH compared to PZC may be crucial when interpreting IVBA and RBA results as mineral-specific PZC may affect As sorption, especially for As(V), further exemplifying the need to elucidate soil Fe phases and speciation changes throughout the GIT.
Coprecipitation, as previously discussed, for both As species is still expected when shifting from highly acidic to more basic conditions promoting Fe(III) precipitation in the presence of aqueous As (Cornell and Schwertmann 2003; LeMonte et al. 2017; Sposito 2008). Similar sorption of As(III) indicates that As species-dependent interactions may be negligible in the IP, but might be significantly different in the acidic gastric phase. Speciation driven interactions might dictate significant sorption phenomena during IVBA assay extractions and are currently understudied in the bioaccessibility literature base. In addition, the ionic strength and competing ions of IVBA assays are expected to impact sorption (Fendorf et al. 2010; Sparks 2003), subsequently affecting bioaccessibility measurements. Indeed, future research examining the relationship between contaminant sorption to assay pH conditions may resolve mechanisms mediating bioaccessibility measurements for soils.
Due to pH-dependency of As speciation-specific interactions with metal oxide minerals, especially Fe(III) minerals, As bioaccessibility and bioavailability in soils may be significantly impacted by IVBA assay or mouse model, respectively, and soil pH. Dixit and Hering (2003) found As(V) sorption extent to ferrihydrite and goethite exhibited a significantly increased rate of change with pH compared to As(III), suggesting pH is a more important variable influencing As(V) sorption compared to As(III). Evidence indicates that there may be elevated variability in As IVBA estimates across methodologies and/or IVBA phase (gastric vs. intestinal) for soils dominated by As(V) due to pH-sensitive sorption reactions, whereas As(III) variability may alter significantly less. Furthermore, the gastric phase IVBA pH (SBRC approximately pH 1.5) is lower than typical mouse stomach pH (approximately 3) (Bradham et al. 2018, 2011). Release of As(V) at pH 3 in the mouse stomach may occur to a higher extent due to diminished sorption maximum at a relatively less acidic pH condition. This in theory might contribute to lower %IVBA measured at pH 1.5 compared to %RBA; however, solution-dissolution chemistry of the specific Fe phase present is expected to play an integral role at gastric phase IVBA pH. Therefore, pH-mediated differences in As gastric phase bioaccessibility versus mouse model bioavailability may be challenging to discern due to joint sorption and precipitation-dissolution chemical processes.
To provide initial insight into the influence of As speciation on IVIVC, a IVIVC residual analysis was conducted across 5 assays for 10 soils where speciation data was provided (Figure 7) (Juhasz et al. 2014). IVIVC residuals, in this context, represent the difference between directly measured % IVBA value for a given soil and predicted value based upon fitted linear regression line, where positive residuals are associated with directly measured IVBA values that are greater than regression model predicted values, and negative residuals are associated with the opposite scenario. Soils labeled 1 through 5 were reported to contain solely As(V) phases, whereas soils labeled 6 through 10 were reported to contain a mixture of As(V) and As(III). Generally, the closer to zero the residual, the more accurate the prediction of RBA for that particular soil and assay. For the gastric-phase residuals (Figure 7(a)), As(V) only soils (left side) generally displayed lower residuals than soils containing both As(V) and As(III) species (right side). The data suggest that soils with more complex As speciation may be associated with less accurate RBA prediction across gastric-phase assays. However, speciation complexity alone cannot predict IVIVC prediction error, as evidenced by soils 6 and 9 having residuals similar to soils containing As(V), only. Furthermore, it remains unclear whether speciation complexity in terms of As valence state, mineral vs. sorbed, or a combination of factors are most important. Conversely, no clear pattern was noted from residual analysis for the intestinal phase (Figure 7(b)). This may be due to total soil Fe being a more important mechanism controlling IVBA in intestinal-phase assays independent of As speciation than in gastric phases. It was also found that IVIVC residuals for a specific soil to be consistently positive or negative across assay for both gastric and intestinal phases. Although the scope of this dataset is limited, this preliminary investigation indicates that increasing As speciation complexity is expected to influence prediction of RBA from IVBA assays, warranting future investigations combining As bioaccessibility, bioavailability, and speciation analyses.
Figure 7.

In vitro – in vivo correlation (IVIVC) residuals in (a) gastric and (b) intestinal phase IVBA assays reported by Juhasz et al. (2014) (n = 10 soils) for each of the five assays. Soils labeled 1 through 5 contain As(V) only, whereas soils 6 through 10 contain a mixture of As(V) and As(III). Residuals, in this context, represent the difference between the directly measured % IVBA value for a given soil and the predicted value based on the fitted linear regression line. Generally, the closer to zero the residual, the more accurate the prediction of RBA for that particular soil and assay
Techniques for investigating arsenic speciation and binding mechanisms in soil systems
Improved understanding of the roles As speciation and Fe precipitation chemistry play in RBA and IVBA measurements, will be dependent, in part, on access to tools and techniques that can reliably and precisely measure soil As and Fe valence states, sorption/mineral phases, and binding mechanisms. Fortunately, direct spectroscopic approaches, coupled with indirect batch experiment techniques, are continually improving our ability to elucidate biogeochemical controls of As environmental mobility, bioaccessibility, and bioavailability. The need for this crucial insight has led to advances in indirect batch experiments and direct spectroscopic approaches over approximately the past 25 years. Benchtop experiments may include performing paired-adsorption and desorption experiments in the presence of high ionic strength solutions and/or matrices containing competing ions such as phosphate (Balint et al. 2020; Deng et al. 2020). Decreased sorption and/or increased desorption in the presence of a high ionic strength solution may be indicative of disruption of outer-sphere sorption (Sparks 2003). Increases in the desorbed As fraction in the presence of a competing ion compared to the high ionic strength desorption results are often indirectly employed to indicate the fraction of As that may be sorbing via inner-sphere complexes (Sparks 2003). Raman and Fourier-transform infrared (FTIR) vibrational spectroscopies have been successfully utilized to probe As species specific binding mechanisms (Goldberg and Johnston 2001). However, X-ray absorption spectroscopy (XAS) is the predominant technique utilized to investigate As chemistry in soils (Fendorf et al. 2010).
Arsenic K-edge XAS spectra may be collected for soils using synchrotron facilities such as Stanford Synchrotron Radiation Lightsource or Advanced Photon Source. Spectra are collected either using X-ray transmission and/or fluorescence technique typically by placing a solid sample such as soil or mouse model components in a beamline that runs tangential to the synchrotron ring. Arsenic K-edge spectra collected may be divided into two regions that provide different insights into As chemistry. X-ray absorption near-edge structure (XANES) spectroscopy is the region typically existing in an energy range of −50 to 100 eV relative to the As absorption edge (Kelly, Hesterberg, and Ravel 2008). Valence state of As in the soil may be determined based upon location of the As absorption edge in the XANES region, whereas the As(V) absorption edge is located at approximately 11875 eV and the As(III) edge is at approximately 11871 eV. Further, linear combination fitting (LCF) using spectra of known As standards may also provide structural insight into As sorbed or mineral phases (Sowers et al. 2017); however, the LCF of the extended X-ray absorption fine-structure (EXAFS) region – ranging from the absorption edge to approximately 1,000 eV above the edge – may provide the most robust structural information present (Kelly, Hesterberg, and Ravel 2008). In addition, shell-fitting analysis may be performed to probe As bonding distances with coordinated oxygen, Fe, and/or other associated elements including metals, metalloids, and/or nonmetals (Kelly, Hesterberg, and Ravel 2008; Waychunas et al. 1993). Estimations of bond distance may be used to elucidate bonding mechanisms that include the presence of inner- or outer-sphere complexes in addition to showcasing the type of inner-sphere complexation present such as monodentate mononuclear, bidentate mononuclear or bidentate binuclear (Fendorf et al. 2010; Sowers et al. 2017; Sparks 2003). Atomic-level sorption behavior markedly mediates the observed As sorption stability. Paired As XANES and EXAFS analyses of soils provide a powerful window into As valence state, phase, and binding mechanism that is unmatched by other techniques. The primary limitation of this technique is that accessibility might be an issue. It can be challenging to acquire beamtime to conduct these analyses, and it is difficult to obtain appropriate instructions for performing XANES, especially EXAFS data processing. Unwanted photooxidation of As(III) is a common issue when performing XAS but is typically prevented using a cryostat. Further, performing XAS on samples post-IVBA extraction may be restricting due to sample mass constraints and analysis of remaining solution phases typically requires challenging beamline setups. However, XAS-based techniques remain at the forefront of investigations examining the chemistry of As in soils and are often paired with ancillary batch, spectrometric, and spectroscopic techniques.
Persisting knowledge gaps and concluding remarks
Elucidation of chemical factors controlling bioaccessibility is critical to understanding how soil heterogeneity affects As bioavailability. Although, on theoretical grounds, As speciation is expected to significantly influence bioaccessibility and bioavailability, few studies examined the relationship between As speciation and bioaccessibility or bioavailability. As(V) and As(III) interact with Fe phases uniquely and are driven by pH-dependent sorption (Figure 3) and co-precipitation reactions (Borch et al. 2010; Fendorf and Kocar 2009; Sparks 2003). Recently, Stevens et al. (2018) showed the promise and challenges of connecting As speciation with RBA measurements using the RBA of As standards to estimate RBA in soils. There is a significant opportunity to build upon this work via continued speciation analyses and correlation of results to IVBA assays, which is also presented in our IVIVC residual analysis findings (Figure 7). However, As speciation should not be solely considered to drive bioaccessibility and bioavailability measurements and should be assessed in conjunction with other soil chemical properties.
Soil Fe content was identified as a significant explanatory variable dictating expected heterogeneity of bioaccessibility results across all assays and assay phases (Figures 6 and 7) (Juhasz et al. 2014, 2015, 2009; Li et al. 2015). Due to the association between As(V/III) and Fe, the biochemical composition including pH, competing ions, and microbiology of the gastric and intestinal IVBA phases make resolving the impact of specific As species critical to predicting drivers of bioaccessible As in soils. Similarly, further investigations correlating As speciation and Fe content when measuring bioaccessibility assays might prove pivotal to the accurate determination of bioaccessibility for soil ranging in mineralogy and composition.
Other soil factors that may affect bioaccessibility include soil chemical and physical properties that have been well explored for soil systems but not in relation to speciation-specific bioaccessibility and bioavailability. Manganese oxides are prevalent in soil systems and were extensively investigated with respect to relationship with As species (Fischel et al. 2015; Ying, Kocar, and Fendorf 2012); however, the effect of manganese oxides on As bioaccessibility is poorly understood. Manganese oxides are highly redox active and were found to promote oxidation of As(III) to As(V) resulting in As sorbing and/or precipitating on the surface of a manganese oxide mineral (Fischel et al. 2015; Yang et al. 2005; Ying, Kocar, and Fendorf 2012). Oxidation to As(V) may promote increased sorption to other minerals such as Fe (oxyhydr)oxides and aluminosilicates compared to As(III) (Fischel et al. 2015; Sparks 2003). This process may decrease solubilized As, and oxidation of environmentally mobile As(III) to more favorably sequestered As(V) by soil manganese oxides and may exert a considerable, yet unknown, effect on bioaccessibility and bioavailability estimations specific to soils where As(III) is or was previously present. However, there is currently a lack of research regarding the correlation between As bioaccessibility and bioavailability in the presence of soil Mn oxides. A combination of model and applied investigations using As(III)-containing systems/soils may shed light on the relationship between manganese oxides and As bioaccessibility and bioavailability.
The influence of soil organic matter (OM) interactions on bioaccessibility and bioavailability is under-investigated. OM contains heterogeneous distributions of reactive organic moieties that may surface passivate Fe sorbents. Limited research shows that As bioaccessibility is expected to rise with increasing OM content (Grafe, Eick, and Grossl 2001; Meunier et al. 2010). However, soil OM consisting of diverse carbon compound classes may have variable reactivity for As and may be As species dependent. Exploring how OM heterogeneity affects As(V) and As(III) may lead to important advances in how soil OM needs to be viewed in the context of bioaccessibility and bioavailability.
Future investigations examining the relationship between As speciation and soil composition to assay chemistry have the potential to provide an informative window into explaining bioaccessibility and bioavailability results across heterogenous soil systems, potentially revealing critical soil chemical indicators that exert the greatest effect on bioaccessibility measurements and bioavailability predictions. Increased understanding of chemical mediators of bioaccessibility has the potential to inform future bioaccessibility investigations and remediation strategy/technology development in As-contaminated soil systems.
Acknowledgments
Portions of this work were funded by U.S. Environmental Protection Agency Office of Superfund Remediation and Technology Innovation (OSRTI) under contract 68HERH19D022 (Task Order 68HERH19F0313). We thank Kasey Kovalcik and Wilson Salls for their internal reviews of this manuscript. This document is being subjected to review by the Center of Environmental Measurement and Modeling (CEMM) for publication. Approval does not signify that the contents reflect the views of the Agency, nor does mention of trade names or commercial products constitute endorsement or recommendation for use.
Funding
The author(s) reported there is no funding associated with the work featured in this article.
Footnotes
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Abbas G, Murtaza B, Bibi I, Shahid M, Niazi NK, Khan MI, Amjad M, and Hussain M. 2018. Arsenic uptake, toxicity, detoxification, and speciation in plants: Physiological, biochemical, and molecular aspects. Int J Environ Res Public Health 15 (1):59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akter KF, Owens G, Davey DE, and Naidu R. 2005. Arsenic speciation and toxicity in biological systems. Rev Environ Contam Toxicol 184:97–149. [DOI] [PubMed] [Google Scholar]
- Antelo J, Arce F, and Fiol S. 2015. Arsenate and phosphate adsorption on ferrihydrite nanoparticles. Synergetic interaction with calcium ions. Chem. Geol 410:53–62. [Google Scholar]
- Argos M, Kalra T, Rathouz PJ, Chen Y, Pierce B, Parvez F, Islam T, Ahmed A, Rakibuz-Zaman M, Hasan R, et al. 2010. Arsenic exposure from drinking water, and all-cause and chronic-disease mortalities in Bangladesh (HEALS): A prospective cohort study. The Lancet 376 (9737):252–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balint R, Celi L, Barberis E, Prati M, and Martin M. 2020. Organic phosphorus affects the retention of arsenite and arsenate by goethite. Journal of Environmental Quality 49 (6):1655–66. [DOI] [PubMed] [Google Scholar]
- Borch T, Kretzschmar R, Kappler A, Cappellen PV, Ginder-Vogel M, Voegelin A, and Campbell K. 2010. Biogeochemical redox processes and their impact on contaminant dynamics. Environ. Sci. Technol 44 (1):15–23. [DOI] [PubMed] [Google Scholar]
- Bowell R 1994. Sorption of arsenic by iron oxides and oxyhydroxides in soils. Applied Geochemistry 9 (3):279–86. [Google Scholar]
- Bradham KD, Diamond GL, Burgess M, Juhasz A, Klotzbach JM, Maddaloni M, Nelson C, Scheckel K, Serda SM, Stifelman M, et al. 2018. In vivo and in vitro methods for evaluating soil arsenic bioavailability: Relevant to human health risk assessment. Journal of Toxicology and Environmental Health B 21 (2):83–114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradham KD, Scheckel KG, Nelson CM, Seales PE, Lee GE, Hughes MF, Miller BW, Yeow A, Gilmore T, Serda SM, et al. 2011. Relative bioavailability and bioaccessibility and speciation of arsenic in contaminated soils. Environ. Health Perspect 119 (11):1629–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brattin W, Drexler J, Lowney Y, Griffin S, Diamond G, and Woodbury L. 2013. An in vitro method for estimation of arsenic relative bioavailability in soil. Journal of Toxicology and Environmental Health, Part A 76 (7):458–78. [DOI] [PubMed] [Google Scholar]
- Brattin W, and Casteel S. 2013. Measurement of arsenic relative bioavailability in swine. Journal of Toxicology and Environmental Health, Part A 76 (7):449–57. [DOI] [PubMed] [Google Scholar]
- Catalano JG, Park C, Fenter P, and Zhang Z. 2008. Simultaneous inner-and outer-sphere arsenate adsorption on corundum and hematite. Geochim. Cosmochim. Acta 72 (8):1986–2004. [Google Scholar]
- Chappell J, Chiswell B, and Olszowy H. 1995. Speciation of arsenic in a contaminated soil by solvent extraction. Talanta 42 (3):323–29. [DOI] [PubMed] [Google Scholar]
- Cohen SM, Arnold LL, Beck BD, Lewis AS, and Eldan M. 2013. Evaluation of the carcinogenicity of inorganic arsenic. Crit. Rev. Toxicol 43 (9):711–52. [DOI] [PubMed] [Google Scholar]
- Cornell RM, and Schwertmann U. 2003. The iron oxides: Structure, properties, reactions, occurrences and uses John Wiley & Sons. [Google Scholar]
- da Silva RF, Borges CDS, de Almeida Lamas C, Cagnon VHA, and de Grava Kempinas W. 2017. Arsenic trioxide exposure impairs testicular morphology in adult male mice and consequent fetus viability. Journal of Toxicology and Environmental Health, Part A 80 (19–21):1166–79. [DOI] [PubMed] [Google Scholar]
- de Oliveira M, Caixeta EC, Santos ES, and Pereira BB. 2021. Arsenic exposure from groundwater: Environmental contamination, human health effects, and sustainable solutions. Journal of Toxicology and Environmental Health, Part B 24 (3):119–35. [DOI] [PubMed] [Google Scholar]
- Deng Y, Weng L, Li Y, Chen Y, and Ma J. 2020. Redox-dependent effects of phosphate on arsenic speciation in paddy soils. Environ. Pollut 264:114783. [DOI] [PubMed] [Google Scholar]
- Denys S, Caboche J, Tack K, Rychen G, Wragg J, Cave M, Jondreville C, and Feidt C. 2012. In vivo validation of the unified BARGE method to assess the bioaccessibility of arsenic, antimony, cadmium, and lead in soils. Environ. Sci. Technol 46 (11):6252–60. [DOI] [PubMed] [Google Scholar]
- Diamond GL, Bradham KD, Brattin WJ, Burgess M, Griffin S, Hawkins CA, Juhasz AL, Klotzbach JM, Nelson C, Lowney YW, et al. 2016. Predicting oral relative bioavailability of arsenic in soil from in vitro bioaccessibility. Journal of Toxicology and Environmental Health, Part A 79 (4):165–73. [DOI] [PubMed] [Google Scholar]
- DIN D (2000). Soil quality-absorption availability of organic and inorganic pollutants from contaminated soil material. DIN E, 19738. [Google Scholar]
- Dixit S, and Hering JG. 2003. Comparison of arsenic (V) and arsenic (III) sorption onto iron oxide minerals: Implications for arsenic mobility. Environ. Sci. Technol 37 (18):4182–89. [DOI] [PubMed] [Google Scholar]
- Eggleton J, and Thomas KV. 2004. A review of factors affecting the release and bioavailability of contaminants during sediment disturbance events. Environ Int 30 (7):973–80. [DOI] [PubMed] [Google Scholar]
- EPA. (2017). Method 1340: In vitro bioaccessibility assay for lead in soil: US EPA Washington, DC. [Google Scholar]
- Fendorf S, and Kocar BD. 2009. Biogeochemical processes controlling the fate and transport of arsenic: Implications for South and Southeast Asia. Advances in Agronomy 104:137–64. [Google Scholar]
- Fendorf S, Nico PS, Kocar BD, Masue Y, and Tufano KJ. 2010. Arsenic chemistry in soils and sediments. Developments in Soil Science 34:357–78. [Google Scholar]
- Fischel MHH, Fischel JS, Lafferty BJ, and Sparks DL. 2015. The influence of environmental conditions on kinetics of arsenite oxidation by manganese-oxides. Geochem. Trans 16 (1):15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gamble AV, Givens AK, and Sparks DL. 2018. Arsenic speciation and availability in orchard soils historically contaminated with lead arsenate. Journal of Environmental Quality 47 (1):121–28. [DOI] [PubMed] [Google Scholar]
- Gillispie EC, Sowers TD, Duckworth OW, and Polizzotto ML. 2015. Soil pollution due to irrigation with arsenic-contaminated groundwater: Current state of science. Current Pollution Reports 1 (1):1–12. [Google Scholar]
- Goldberg S, and Johnston CT. 2001. Mechanisms of arsenic adsorption on amorphous oxides evaluated using macroscopic measurements, vibrational spectroscopy, and surface complexation modeling. Journal of Colloid and Interface Science 234 (1):204–16. [DOI] [PubMed] [Google Scholar]
- Golub MS, Macintosh MS, and Baumrind N. 1998. Developmental and reproductive toxicity of inorganic arsenic: Animal studies and human concerns. Journal of Toxicology and Environmental Health, Part B 1 (3):199–237. [DOI] [PubMed] [Google Scholar]
- Grafe M, Eick M, and Grossl P. 2001. Adsorption of arsenate (V) and arsenite (III) on goethite in the presence and absence of dissolved organic carbon. Soil Science Society of America Journal 65 (6):1680–87. [Google Scholar]
- Griggs JL, Thomas DJ, Fry R, and Bradham KD. 2021. Improving the predictive value of bioaccessibility assays and their use to provide mechanistic insights into bioavailability for toxic metals/metalloids – A research prospectus. Journal of Toxicology and Environmental Health, Part B 24 (7):307–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hack A, and Selenka F. 1996. Mobilization of PAH and PCB from contaminated soil using a digestive tract model. Toxicol. Lett 88 (1–3):199–210. [DOI] [PubMed] [Google Scholar]
- Juhasz AL, Smith E, Weber J, Rees M, Rofe A, Kuchel T, Sansom L, and Naidu R. 2007. Comparison of in vivo and in vitro methodologies for the assessment of arsenic bioavailability in contaminated soils. Chemosphere 69 (6):961–66. [DOI] [PubMed] [Google Scholar]
- Juhasz AL, Weber J, and Smith E. 2011. Predicting arsenic relative bioavailability in contaminated soils using meta analysis and relative bioavailability–bioaccessibility regression models. Environ. Sci. Technol 45 (24):10676–83. [DOI] [PubMed] [Google Scholar]
- Juhasz AL, Weber J, Smith E, Naidu R, Rees M, Rofe A, Kuchel T, and Sansom L. 2009. Assessment of four commonly employed in vitro arsenic bioaccessibility assays for predicting in vivo relative arsenic bioavailability in contaminated soils. Environ. Sci. Technol 43 (24):9487–94. [DOI] [PubMed] [Google Scholar]
- Juhasz AL, Herde P, Herde C, Boland J, and Smith E. 2014. Validation of the predictive capabilities of the Sbrc-G in vitro assay for estimating arsenic relative bioavailability in contaminated soils. Environ. Sci. Technol 48 (21):12962–69. [DOI] [PubMed] [Google Scholar]
- Juhasz AL, Herde P, Herde C, Boland J, and Smith E. 2015. Predicting arsenic relative bioavailability using multiple in vitro assays: Validation of in vivo–in vitro correlations. Environ. Sci. Technol 49 (18):11167–75. [DOI] [PubMed] [Google Scholar]
- Karna RR, Noerpel M, Betts AR, and Scheckel KG. 2017. Lead and arsenic bioaccessibility and speciation as a function of soil particle size. Journal of Environmental Quality 46 (6):1225–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelly S, Hesterberg D, and Ravel B. 2008. Analysis of soils and minerals using X‐ray absorption spectroscopy. Methods of Soil Analysis Part 5—Mineralogical Methods 5:387–463. [Google Scholar]
- Khalid S, Shahid M, Niazi NK, Rafiq M, Bakhat HF, Imran M, Abbas T, Bibi I, and Dumat C. 2017. Arsenic behaviour in soil-plant system: Biogeochemical reactions and chemical speciation influences. In Enhancing cleanup of environmental pollutants, ed. Anjum N, Gill S, and Tuteja N, 97–140. Cham: Springer. [Google Scholar]
- Kim EJ, Hwang BR, and Baek K. 2015. Effects of natural organic matter on the coprecipitation of arsenic with iron. Environ Geochem Health 37 (6):1029–39. [DOI] [PubMed] [Google Scholar]
- Kim EJ, Yoo J-C, and Baek K. 2014. Arsenic speciation and bioaccessibility in arsenic-contaminated soils: Sequential extraction and mineralogical investigation. Environ. Pollut 186:29–35. [DOI] [PubMed] [Google Scholar]
- Kim NH, Mason CC, Nelson RG, Afton SE, Essader AS, Medlin JE, Levine KE, Hoppin JA, Lin C, Knowler WC. 2013. Arsenic exposure and incidence of type 2 diabetes in Southwestern American Indians. Am. J. Epidemiol 177(9):962–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuivenhoven M, and Mason K (2019). Arsenic (Arsine) toxicity StatPearls [Internet]: StatPearls Publishing. [PubMed] [Google Scholar]
- LeMonte JJ, Stuckey JW, Sanchez JZ, Tappero R, Rinklebe JR, and Sparks DL. 2017. Sea level rise induced arsenic release from historically contaminated coastal soils. Environ. Sci. Technol 51 (11):5913–22. [DOI] [PubMed] [Google Scholar]
- Li HB, Ning H, Li SW, Li J, Xue RY, Li MY, Wang MY, Liang JH, Juhasz AL, and Ma LQ. 2021. An interlaboratory evaluation of the variability in arsenic and lead relative bioavailability when assessed using a mouse bioassay. Journal of Toxicology and Environmental Health A 84 (14):593–607. [DOI] [PubMed] [Google Scholar]
- Li J, Li K, Cui X-Y, Basta NT, Li L-P, Li H-B, and Ma LQ, 2015. In vitro bioaccessibility and in vivo relative bioavailability in 12 contaminated soils: Method comparison and method development. Science of the Total Environment 532:812–20. [DOI] [PubMed] [Google Scholar]
- Lim K, Shukor M, and Wasoh H. 2014. Physical, chemical, and biological methods for the removal of arsenic compounds. Biomed. Res. Int 2014:503784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mandal BK, and Suzuki KT. 2002. Arsenic round the world: A review. Talanta 58 (1):201–35. [PubMed] [Google Scholar]
- Marlborough SJ, and Wilson VL. 2015. Arsenic speciation driving risk based corrective action. Science of the Total Environment 520:253–59. [DOI] [PubMed] [Google Scholar]
- Medina S, Xu H, Wang SC, Lauer FT, Liu KJ, and Burchiel SW. 2017. Low level arsenite exposures suppress the development of bone marrow erythroid progenitors and result in anemia in adult male mice. Toxicol. Lett 273:106–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meunier L, Koch I, and Reimer KJ. 2011. Effects of organic matter and ageing on the bioaccessibility of arsenic. Environ. Pollut 159 (10):2530–36. [DOI] [PubMed] [Google Scholar]
- Meunier L, Walker SR, Wragg J, Parsons MB, Koch I, Jamieson HE, Reimer KJ. 2010. Effects of soil composition and mineralogy on the bioaccessibility of arsenic from tailings and soil in gold mine districts of Nova Scotia. Environ. Sci. Technol 44(7):2667–74. [DOI] [PubMed] [Google Scholar]
- National Research Council. 2003. Bioavailability of Contaminants in Soils and Sediments: Processes, Tools, and Applications National Research Council National Academies Press. [Google Scholar]
- Nelson CM, Li K, Obenour DR, Miller J, Misenheimer JC, Scheckel K, Betts A, Juhasz A, Thomas DJ, and Bradham KD. 2018. Relating soil geochemical properties to arsenic bioaccessibility through hierarchical modeling. Journal of Toxicology and Environmental Health, Part A 81 (6):160–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ollson CJ, Smith E, Scheckel KG, Betts AR, and Juhasz AL. 2016. Assessment of arsenic speciation and bioaccessibility in mine-impacted materials. J. Hazard. Mater 313:130–37. [DOI] [PubMed] [Google Scholar]
- Oomen AG, Hack A, Minekus M, Zeijdner E, Cornelis C, Schoeters G, Verstraete W, Van de Wiele T, Wragg J, Rompelberg CJM. 2002. Comparison of five in vitro digestion models to study the bioaccessibility of soil contaminants. Environ. Sci. Technol 36(15):3326–34. [DOI] [PubMed] [Google Scholar]
- Paktunc D, and Dutrizac JE. 2003. Characterization of arsenate-for-sulfate substitution in synthetic jarosite using X-ray diffraction and X-ray absorption spectroscopy. The Canadian Mineralogist 41 (4):905–19. [Google Scholar]
- Palumbo-Roe B, and Klinck B. 2007. Bioaccessibility of arsenic in mine waste-contaminated soils: A case study from an abandoned arsenic mine in SW England (UK). Journal of Environmental Science and Health, Part A 42 (9):1251–61. [DOI] [PubMed] [Google Scholar]
- Raven KP, Jain A, and Loeppert RH. 1998. Arsenite and arsenate adsorption on ferrihydrite: Kinetics, equilibrium, and adsorption envelopes. Environ. Sci. Technol 32 (3):344–49. [Google Scholar]
- Redman AD, Macalady DL, and Ahmann D. 2002. Natural organic matter affects arsenic speciation and sorption onto hematite. Environ. Sci. Technol 36 (13):2889–96. [DOI] [PubMed] [Google Scholar]
- Reeder RJ, Schoonen MA, and Lanzirotti A. 2006. Metal speciation and its role in bioaccessibility and bioavailability. Reviews in Mineralogy and Geochemistry 64 (1):59–113. [Google Scholar]
- Rodriguez RR, Basta NT, Casteel SW, and Pace LW. 1999. An in vitro gastrointestinal method to estimate bioavailable arsenic in contaminated soils and solid media. Environ. Sci. Technol 33 (4):642–49. [Google Scholar]
- Rotard W, Christmann W, Knoth W, and Mailahn W. 1995. Bestimmung der resorptionsverfügbaren PCDD/PCDF aus Kieselrot. Umweltwissenschaften Und Schadstoff-Forschung 7 (1):3–9. [Google Scholar]
- Ruby MV, Davis A, Schoof R, Eberle S, and Sellstone CM. 1996. Estimation of lead and arsenic bioavailability using a physiologically based extraction test. Environ. Sci. Technol 30 (2):422–30. [Google Scholar]
- Ruby MV, Schoof R, Brattin W, Goldade M, Post G, Harnois M, Mosby DE, Casteel SW, Berti W, Carpenter M, et al. 1999. Advances in evaluating the oral bioavailability of inorganics in soil for use in human health risk assessment. Environ. Sci. Technol 33 (21):3697–705. [Google Scholar]
- Schuhmacher–Wolz U, Dieter HH, Klein D, and Schneider K. 2009. Oral exposure to inorganic arsenic: Evaluation of its carcinogenic and non-carcinogenic effects. Crit. Rev. Toxicol 39 (4):271–98. [DOI] [PubMed] [Google Scholar]
- Smedley PL, and Kinniburgh DG. 2002. A review of the source, behaviour and distribution of arsenic in natural waters. Applied Geochemistry 17 (5):517–68. [Google Scholar]
- Smith AH, and Steinmaus CM. 2009. Health effects of arsenic and chromium in drinking water: Recent human findings. Annu. Rev. Public Health 30 (1):107–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith E, Scheckel K, Miller BW, Weber J, and Juhasz AL. 2014. Influence of in vitro assay pH and extractant composition on As bioaccessibility in contaminated soils. Science of the Total Environment 473:171–77. [DOI] [PubMed] [Google Scholar]
- Sowers TD, Harrington JM, Polizzotto ML, and Duckworth OW. 2017. Sorption of arsenic to biogenic iron (oxyhydr) oxides produced in circumneutral environments. Geochim. Cosmochim. Acta 198:194–207. [Google Scholar]
- Sparks DL 2003. Environmental Soil Chemistry Elsevier. [Google Scholar]
- Sposito G 2008. The Chemistry of Soils Oxford university press. [Google Scholar]
- Stevens BN, Betts AR, Miller BW, Scheckel KG, Anderson RH, Bradham KD, Casteel SW, Thomas DJ, and Basta NT. 2018. Arsenic speciation of contaminated soils/solid wastes and relative oral bioavailability in swine and mice. Soil Systems 2 (2):27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strawn DG, Bohn HL, and O’Connor GA. 2019. Soil Chemistry John Wiley and Sons. [Google Scholar]
- Thouin H, Le Forestier L, Gautret P, Hube D, Laperche V, Dupraz S, and Battaglia-Brunet F. 2016. Characterization and mobility of arsenic and heavy metals in soils polluted by the destruction of arsenic-containing shells from the Great War. Science of the Total Environment 550:658–69. [DOI] [PubMed] [Google Scholar]
- Toujaguez R, Ono F, Martins V, Cabrera P, Blanco A, Bundschuh J, and Guilherme LRG. 2013. Arsenic bioaccessibility in gold mine tailings of Delita, Cuba. J. Hazard. Mater 262:1004–13. [DOI] [PubMed] [Google Scholar]
- USEPA. 2007. Guidance for evaluating the oral bioavailability of metals in soils for use in human health risk assessment. OSWER 9285: 7–80. [Google Scholar]
- Vahter M 2008. Health effects of early life exposure to arsenic. Basic Clin. Pharmacol. Toxicol 102 (2):204–11. [DOI] [PubMed] [Google Scholar]
- Van de Wiele TR, Oomen AG, Wragg J, Cave M, Minekus M, Hack A, Cornelis C, Rompelberg CJ, De Zwart LL, Klinck B, et al. 2007. Comparison of five in vitro digestion models to in vivo experimental results: Lead bioaccessibility in the human gastrointestinal tract. Journal of Environmental Science and Health, Part A 42 (9):1203–11. [DOI] [PubMed] [Google Scholar]
- Van de Wiele T, Van den Abbeele P, Ossieur W, Possemiers S, and Marzorati M. 2015. The simulator of the human intestinal microbial ecosystem (SHIME®). In The impact of food bioactives on health: in vitro and ex vivo models, ed. Verhoeckx K, Cotter P, López-Expósito I, Kleiveland C, Lea T, Mackie A, Requena T, Swiatecka D, and Wichers H, 305–17. Cham: Springer International Publishing. [PubMed] [Google Scholar]
- Voegelin A, Weber F-A, and Kretzschmar R. 2007. Distribution and speciation of arsenic around roots in a contaminated riparian floodplain soil: Micro-XRF element mapping and EXAFS spectroscopy. Geochim. Cosmochim. Acta 71 (23):5804–20. [Google Scholar]
- Waychunas G, Rea B, Fuller C, and Davis J. 1993. Surface chemistry of ferrihydrite: Part 1. EXAFS studies of the geometry of coprecipitated and adsorbed arsenate. Geochim. Cosmochim. Acta 57 (10):2251–69. [Google Scholar]
- Whitacre S, Basta N, Stevens B, Hanley V, Anderson R, and Scheckel K. 2017. Modification of an existing in vitro method to predict relative bioavailable arsenic in soils. Chemosphere 180:545–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WHO. 2019. Preventing disease through healthy environments: Exposure to arsenic: A major public health concern World Health Organization. [Google Scholar]
- Wilkie JA, and Hering JG. 1996. Adsorption of arsenic onto hydrous ferric oxide: Effects of adsorbate/adsorbent ratios and co-occurring solutes. Colloids Surf A Physicochem Eng Asp 107:97–110. [Google Scholar]
- Wragg J, and Cave M (2003). In-vitro methods for the measurement of the oral bioaccessibility of selected metals and metalloids in soils: A critical review: Environment agency Bristol [Google Scholar]
- Wragg J, Cave M, Basta N, Brandon E, Casteel S, Denys S, Gron C, Oomen A, Reimer K, Tack K, et al. 2011. An inter-laboratory trial of the unified BARGE bioaccessibility method for arsenic, cadmium and lead in soil. Science of the Total Environment 409:4016–30. [DOI] [PubMed] [Google Scholar]
- Xue Q, Ran Y, Tan Y, Peacock CL, and Du H. 2019. Arsenite and arsenate binding to ferrihydrite organo-mineral coprecipitate: Implications for arsenic mobility and fate in natural environments. Chemosphere 224:103–10. [DOI] [PubMed] [Google Scholar]
- Yang H-C, Fu H-L, Lin Y-F, and Rosen BP. 2012. Pathways of arsenic uptake and efflux. Curr Top Membr 69:325–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang JK, Barnett MO, Jardine PM, and Brooks SC. 2003. Factors controlling the bioaccessibility of arsenic (V) and lead (II) in soil. Soil and Sediment Contamination: An International Journal 12 (2):65–179. [Google Scholar]
- Yang J-K, Barnett MO, Zhuang J, Fendorf SE, and Jardine PM. 2005. Adsorption, oxidation, and bioaccessibility of As (III) in soils. Environ. Sci. Technol 39 (18):7102–10. [DOI] [PubMed] [Google Scholar]
- Yang J-K, Barnett MO, Jardine PM, Basta NT, and Casteel SW. 2002. Adsorption, sequestration, and bioaccessibility of As (V) in soils. Environ. Sci. Technol 36 (21):4562–69. [DOI] [PubMed] [Google Scholar]
- Ying SC, Kocar BD, and Fendorf S. 2012. Oxidation and competitive retention of arsenic between iron-and manganese oxides. Geochim. Cosmochim. Acta 96:294–303. [Google Scholar]