

Abstract
Discontinuation of the contingency between a response and its reinforcer sometimes produces a temporary increase in the response before its rate decreases, a phenomenon called the extinction burst. Prior clinical and basic studies on the prevalence of the extinction burst provide highly disparate estimates. Existing theories on the extinction burst fail to account for the dynamic nature of this phenomenon, and the basic behavioral processes that control response bursting remain poorly understood. In this paper, we first review the basic and applied literature on the extinction burst. We then describe a recent refinement of the concatenated matching law called the temporally weighted matching law that appears to resolve the above‐mentioned issues regarding the extinction burst. We present illustrative translational data based conceptually on the model. Finally, we discuss specific recommendations derived from the temporally weighted matching law regarding procedures clinicians could implement to potentially mitigate or prevent extinction bursts.
Keywords: extinction burst, problem behavior, temporally weighted matching law, translational research
When the contingency between a response and its reinforcing consequence is terminated, the resulting effect on the rate of the target response is a characteristic reduction to low or zero levels, a process called extinction (EXT; Skinner, 1938). However, under certain arrangements (e.g., multiple fixed‐ratio [FR] 1 EXT; Azrin et al., 1966; Skinner, 1938), the target response may temporarily increase prior to showing the characteristic decelerating extinction curve. Research has shown that this temporary potentiating effect of extinction also occurs during respondent or Pavlovian extinction (Rohrbaugh et al., 1972). Behavior analysts refer to such transitory increases in target responding as a burst (Catania, 2013; Skinner, 1938) or bursting (Lerman et al., 1999), and as an extinction burst (Lerman & Iwata, 1995, 1996; Lerman et al., 1999) when the increase in target responding coincides with the introduction of extinction.
Children with autism spectrum disorder or intellectual disability often display severe destructive behavior (e.g., self‐injurious behavior [SIB], aggression) that poses great risks to self, others, or the environment and represent immense barriers to community integration (Borthwick‐Duffy, 1994; Crocker et al., 2006). Such problem behavior is often treated with interventions derived from a functional analysis that involve extinction of the response. Functional analyses identify the environmental antecedents and consequences that occasion and reinforce the behavior. For example, functional communication training (FCT; Carr & Durand, 1985; Fisher et al., 2021) involves extinction of problem behavior and reinforcement of an alternative communication response with the consequence that previously reinforced problem behavior. In other treatment arrangements, the consequence that previously reinforced problem behavior is delivered on a response‐independent, time‐based schedule (Hagopian et al., 1994; Vollmer et al., 1993). Results from epidemiological studies and meta‐analyses indicate that treatments based on functional analyses, like FCT, typically reduce problem behavior by 90% or more and are much more effective than other treatments (e.g., Greer et al., 2016; Iwata et al., 1994; Richman et al., 2015). In some cases, function‐based treatments can reduce problem behavior to clinically acceptable levels without extinction (e.g., Lalli et al., 1999; Lomas et al., 2010); however, treatment effectiveness often decreases when the schedule of reinforcement is thinned to make the treatment more practical for caregivers (e.g., Briggs et al., 2018; see Briggs et al., 2019 and Pace et al., 1994, for notable exceptions).
Despite the impressive findings regarding the effectiveness of function‐based treatments like FCT, these interventions can induce untoward effects when individuals experience periods of nonreinforcement at the start of treatment, during reinforcement schedule thinning, or when the alternative communication response stops producing reinforcement (Briggs et al., 2018; Brown et al., 2020; DeRosa et al., 2015; Fisher, Greer, Fuhrman et al., 2018; Fisher, Greer, Mitteer et al., 2018; Fisher et al., 2019, Greer et al., 2020; Haney et al., 2022; Mitteer et al., 2022). That is, when periods of extinction are introduced, three untoward byproducts may appear: (a) the predominant target response (e.g., head banging) may temporarily increase in frequency, duration, or intensity (i.e., an extinction burst); (b) other topographies of responding within the same response class may increase or emerge (e.g., face slapping partially replacing head banging); or (c) topographies of responding outside the response class (i.e., nontarget responses) may emerge (e.g., extinction‐induced aggression replacing SIB). However, these untoward effects of extinction are not ubiquitous (Katz & Lattal, 2021; Lerman & Iwata, 1995); they occur in some cases, but not others. Thus, understanding the variables that give rise to extinction‐related increases in problem behavior is critical to further improving the effectiveness of function‐based treatments like FCT.
In this paper, we describe and compare basic and applied studies that have examined the prevalence of extinction bursts among human/clinical participants (e.g., Lerman & Iwata, 1995; Lerman et al., 1999) and among nonhuman/laboratory subjects (e.g., Lattal et al., 2020). The human studies have found lower than expected prevalence rates (e.g., Lerman et al., 1999), and the nonhuman studies have found near‐zero prevalence rates, when applying the same ascertainment criteria used in the human/clinical studies.
We hypothesize that these disparate prevalence estimates occur because (a) applied research participants often experience greater decreases in reinforcer‐consumption time when extinction begins than do basic research participants; and (b) applied researchers typically include reinforcer‐consumption time when calculating baseline response rates, but basic researchers often do not. Further, we suggest that including and analyzing the effects of reinforcer‐consumption time is important because it may help to explain the conditions under which extinction bursts are likely and unlikely to occur. Further, we describe a recent account hypothesizing that extinction bursts can be explained using the general principles of the matching law. That is, during baseline, individuals allocate their time between the target response (e.g., problem behavior) and reinforcer consumption (which the individual does consistently when reinforcement is available). At the start of extinction, individuals temporarily allocate more time to problem behavior because it still has value due to its reinforcement history and because reinforcer consumption is no longer available to compete with it. Problem behavior decreases shortly thereafter because its value decreases as time in extinction increases.
We describe prior theories of extinction‐induced problem behavior, which we argue do not explain the temporal patterns of extinction bursts. We then present a refinement of the concatenated matching law developed by Shahan (2022), called the temporally weighted matching law (TWML). We argue that the TWML addresses the limitations of prior theoretical accounts of extinction bursts by incorporating reinforcer‐consumption time into the model and estimating the current value of each response option based on its reinforcement history. These aspects of the TWML render it applicable to dynamically changing reinforcement conditions, like the introduction of extinction. We then describe some clinical implications of the TWML and provide illustrative translational data based conceptually on the TWML. Finally, we discuss potential strengths and limitations of the TWML that should be tested and addressed through future research.
Estimating the Prevalence of Extinction Bursts
Although extinction bursts have been discussed in the applied‐ and basic‐research literatures for decades, no well‐integrated, quantitative theories or experimental analyses have identified and evaluated the variables that control the phenomenon (Katz & Lattal, 2020; Shahan, 2022). Extinction‐induced bursting and other problem behavior have been described as prototypical features of the extinction process in both applied and basic research (e.g., Azrin et al., 1966; Keller & Schoenfeld, 1950; Lerman et al., 1999; North, 1950; Skinner, 1938; Terrace, 1966; Thompson & Bloom, 1966); however, recent basic investigations and conceptualizations of extinction‐induced behavior have raised doubts about the ubiquity and generality of at least one of these byproducts, the extinction burst (Katz & Lattal, 2020, 2021; Lattal et al., 2013; Lattal et al., 2020).
Katz and Lattal (2020) introduced extinction after they exposed pigeons' key pecking to reinforcement on FR or variable‐ratio (VR) schedules of reinforcement in baseline. Similarly, Lattal et al. (2020) exposed pigeons' key pecking (Experiment 1) and rats' bar pressing (Experiments 2 and 3) to extinction following baselines in which responding produced reinforcement on multiple schedules (VR schedules alternated with yoked variable‐interval [VI] schedules). Katz and Lattal removed reinforcement time from the baseline response‐rate calculation and observed increases in response rates for a minority of applications and primarily during the first minute of the first extinction session that followed a block of VR sessions, but the increase never lasted for an entire extinction session (i.e., 0 of 106 applications showed bursting during the first whole sessions of extinction [0%]; 21 of 106 applications showed bursting in the first minute of extinction [19.8%]). Lattal et al. removed reinforcement time from the baseline response‐rate calculations in Experiment 1 (pigeons), but not in Experiments 2 and 3 (rats), and the experimenters rarely observed increased response rates when examining responding on a minute‐by‐minute or whole‐session basis (12% of applications showed increased responding during extinction in the minute‐by‐minute analysis but none met the Lerman & Iwata, 1995, criteria for an extinction burst). As mentioned above, we hypothesize that experimenters are more likely to observe extinction bursts following large drops in reinforcement time from baseline to treatment and when they include reinforcement time in the baseline response‐rate calculations. Katz and Lattal programmed large drops in reinforcement time in some cases but did not include reinforcement time in the baseline response‐rate calculations. Lattal et al. included reinforcement time in the baseline response‐rate calculations but programmed considerably smaller decrements in reinforcement time during the transition from baseline to extinction.
In an applied study aimed at determining the prevalence of extinction‐induced bursting and aggression in a clinical population, Lerman et al. (1999) examined whole‐session data for evidence of extinction bursts among 41 individuals with moderate to profound intellectual disability referred for the treatment of SIB. Of those, 21 received treatment with extinction alone, and 13 (62%) of this subgroup showed an extinction burst, defined as an increase in responding during any of the first three sessions of treatment above the response rates measured during the last five baseline sessions (or all sessions for shorter baselines). Thus, whereas Katz and Lattal (2020) and Lattal et al. (2020) never observed an extinction burst that met the Lerman & Iwata (1995) criteria among their pigeons or rats exposed to extinction alone, most of the participants exposed to extinction alone in the Lerman et al. study did. We believe that this difference is because Lerman et al. delivered reinforcement on dense (i.e., FR 1) schedules, thus producing large decrements in reinforcer‐consumption time. In addition, they included reinforcement time in the baseline response‐rate calculations when transitioning from baseline to extinction alone. On the other hand, Katz and Lattal and Lattal et al. sometimes included one of these factors, but not both.
Differences in Reinforcement Schedules and Response‐Rate Calculations
The discrepancies between applied and basic research on extinction‐induced behavior, as exemplified by the Lattal et al. (2020) and Lerman et al. (1999) studies, are probably due to two primary factors mentioned above and explained more fully here. One is that applied research participants typically experience greater decreases in reinforcer‐consumption time when extinction begins than basic research participants. This is because applied researchers often use denser reinforcement schedules than basic researchers (e.g., FR 1 vs. VI 1 min, respectively), and applied researchers often deliver reinforcers for longer periods than basic researchers (e.g., 20‐s access to an iPad vs. 3‐s access to food, respectively). Thus, a human clinical participant accessing an iPad for 20 s after each problem response could spend 80% of a baseline session consuming reinforcement (see example below), whereas a pigeon accessing food for 3 s about once every min would spend about 5% of session time consuming reinforcement. Although these differences between how basic and applied researchers program reinforcement schedules and calculate response rates are not universal, we believe that this is an important distinction because we hypothesize that larger drops in reinforcement at the start of extinction‐based treatments increase the probability of an extinction burst relative to smaller drops in reinforcement.
A second factor that probably contributes to the higher prevalence of extinction bursts often observed by applied researchers is that applied researchers typically include reinforcer‐consumption time when calculating baseline response rates (though again, this is not universal; cf. Ibañez et al., 2019). For example, in a 5‐min baseline session in which problem behavior produces access to its functional reinforcer for 20 s on an FR 1 schedule, an efficient human participant might emit 12 responses and produce 240 s of reinforcement access. Thus, in this example, the participant spends 4 min of the 5‐min baseline session with access to the functional reinforcer and 1 min of the 5‐min baseline session without access to the functional reinforcer. When calculating the baseline response rate as basic researchers often do, the baseline rate equals 12 responses per minute (12 responses / 1 min, when one excludes reinforcement time). However, when calculating the baseline response rate as applied researchers typically do, the baseline rate equals 2.4 responses per minute(12 responses / 5 min, when one includes the 4 min of reinforcement time).
These different methods of calculating baseline response rates can greatly influence whether an extinction burst is identified because during periods of extinction, no reinforcement intervals occur, and all session time is included in the calculation of the response rate. So, if 24 responses occur during the first 5 min of extinction, it would produce a rate of 4.8 responses per minute(24 responses / 5 min). This response rate would not constitute an extinction burst if one excluded reinforcement time when calculating baseline response rates (i.e., 4.8 < 12 responses per minute), but would constitute an extinction burst if one included reinforcement time (i.e., 4.8 > 2.4 responses per minute).
Indeed, in recent research with rats, Nist and Shahan (2021) directly showed that inclusion of all session time (including reinforcer‐consumption time) when calculating baseline response rates increased the prevalence of the extinction burst, especially following reinforcement on small FR schedules like those commonly used in applied studies. Baselines that arrange small FR schedules are likely to produce more reinforcer deliveries per session and thus larger proportions of session time with reinforcement access. Therefore, whether reinforcement time is or is not included in the calculation of baseline response rates is particularly important when assessing extinction bursts following baselines that arrange dense schedules of reinforcement like small FR schedules. Although the findings of Nist and Shahan need to be replicated, they are consistent with our hypothesis that investigators are more likely to observe extinction bursts at a higher prevalence rate if they (a) program large drops in reinforcement at the start of extinction (e.g., shifting from an FR 1 to extinction; programming long reinforcement intervals) and (b) include reinforcer‐consumption time when calculating baseline response rates. When Nist and Shahan did both, they identified extinction bursts at rates like Lerman et al. (1999) and much higher than those observed by Katz and Lattal (2020) and Lattal et al. (2020).
It is worth noting that the early basic researchers who first identified what is now called the extinction burst appear to have included reinforcement time when calculating baseline response rates. For example, Skinner (1938) presented typical cumulative records of lever pressing by rats trained on an FR 1 schedule in his Figure 4, and once the rats learned the schedule, they pressed the lever between 2 and 3 times per minute. 1 In Skinner's Figure 7, he showed typical cumulative records of lever pressing during extinction following training on an FR 1 schedule. During the first 1 to 3 minutes of extinction, all four rats displayed what might be called an extinction burst by pressing the lever between 6 and 17 times per minute. 2 However, it should be noted that Skinner used a modified kymograph (to record responding) that did not remove reinforcement time during the FR 1 training. This led Skinner to the following interpretation of the observed responding at the start of extinction: “When the first response to the lever fails to supply the stimulus for the next member of the usual chain, the response is elicited again immediately, and a high rate of elicitation is maintained for a short time” (Skinner, 1938, p. 74).
Which Calculation Method is Better?
One might ask, then, does including reinforcement time in the calculation of response rates during baseline falsely promulgate a spurious and meaningless extinction burst, or does excluding reinforcement time conceal a legitimate and meaniful one? The answer probably depends on the purpose of the investigation. If one is interested in studying response patterns during transitions to determine whether the local rate of responding temporarily accelerates before decreasing when extinction is introduced, then removing reinforcement time when calculating baseline response rates makes perfect sense. Without removing baseline reinforcement time, the experimenter might observe an apparent, but specious, increase in response rate during extinction due to the decrease in time spent by the participant accessing and consuming the reinforcer when transitioning from baseline to extinction (Keller & Schoenfeld, 1950). However, if one is interested in whether a participant shows more problematic responses in actual clock time during an extinction‐based treatment relative to baseline, then including reinforcement time when calculating baseline response rates seems more appropriate. That is, if an individual displays head banging 12 times per 5‐min session in baseline and 24 times per 5‐min session in the first extinction‐based treatment session, that increase is socially meaningful, even if the increase is primarily due to the absence of time spent accessing and consuming the reinforcer. Such an increase in response rate might increase the risk of injury produced by the behavior (e.g., detached retinas from SIB; Hyman et al., 1990), especially if the individual also displays an increase in the intensity of SIB. In addition, a parent might be unwilling or unable to continue to ignore attention‐reinforced SIB during the initiation of an extinction‐based treatment if their child is emitting twice as many instances of SIB during the first 5 min of treatment than the child did immediately prior to treatment (i.e., an apparent worsening in behavior resulting from what seems to be an ineffective treatment). When this occurs, the parent may resume delivering the functional reinforcer following SIB because the individual stops engaging in SIB and consumes the reinforcer while it is available, thereby lowering the overall rate of SIB (i.e., a negative reinforcement contingency maintaining caregiver behavior; e.g., Mitteer et al., 2018).
Narrower and Broader Definitions of Extinction Bursts
Researchers also may differ in terms of how they define an extinction burst. Researchers studying nonhuman animals (e.g., Lattal et al., 2020) typically define an extinction burst more narrowly than researchers studying human clinical populations. For example, nonhuman animal studies generally (but not always) focus on the rate of a narrowly defined response topography as the target response (e.g., a pigeon pecking a circular key with sufficient force to activate an attached microswitch). Other potentially relevant topographies may occur but are typically not counted or differentiated from the target response. For example, pecks to the wall of the chamber near the response key (e.g., Blough, 1959) and emotional responses (e.g., Terrace, 1966) may go uncounted, whereas vigorous aggressive responses toward the response apparatus (e.g., Keller & Schoenfeld, 1950) are typically counted the same as any other key peck.
By contrast, when studying human clinical populations, researchers often describe extinction bursts as including a variety of topograhies of problem behavior, which may or may not have been a part of the originally defined response class. For example, Thackeray and Richdale (2002), in a study on pediatric sleep disorders, targeted settling, co‐sleeping (with parents), and night‐waking for treatment with extinction for three children with intellectual disability. They described the extinction burst that occurred for one child when treatment commenced as follows: “C1's parents put him back to bed 259 times in 2½ hours on the first night, and he tried behaviours such as screaming, crying, kicking, hitting the walls, pulling faces, throwing up, and spitting” (Thackeray & Richdale, 2002, p. 221). Obviously, these additional topographies of problem behavior are clinically important events that would have been missed if the researchers monitored only the originally defined target responses of settling, co‐sleeping, and night‐waking. These discrepancies between how different researchers define target responses and extinction bursts can lead to markedly different interpretations of the observed results.
Defining and Characterizing Extinction Bursts
Katz and Lattal (2021) reviewed nine operational definitions of an extinction burst from the literature. Eight definitions were from basic studies with nonhuman animals and one by Lerman and colleagues described above that was first applied in clinical studies on extinction bursts (Lerman & Iwata, 1995; Lerman et al., 1999) and was later adapted for use with other behavioral phenomena (e.g., relapse of problem behavior; Briggs et al., 2018; Haney et al., 2022; Mitteer et al., 2022; Muething et al., 2020). All the definitions listed by Katz and Lattal characterized an extinction burst in terms of an increase in the rate of responding at the start of extinction relative to target response rates in baseline, but the criteria used to determine whether an increase had occurred varied considerably across studies. As examples, Schramm‐Sapyta et al. (2006) required a 15% increase in responding during the first session of extinction relative to the mean of the last 2 days of reinforcement; Niyuhire et al. (2007) required statistically significant differences in group means between the last baseline session and the first 15 min of extinction; and, as previously mentioned, Lerman and Iwata (1995) required a higher rate of responding in any of the first three extinction sessions relative to highest rate observed in the last five baseline sessions (or all sessions for shorter baselines).
Katz and Lattal (2021) concluded that attempts to develop a single definition of the extinction burst are misguided. They suggested that researchers consider the environmental conditions in effect during baseline (e.g., FR 1, VI 60) and extinction (e.g., within or between‐session cessation of reinforcement), as well as dimensions of the target response (e.g., rate, force, duration) when defining an extinction burst. We agree, and also suggest that it is important for researchers to consider the purpose of their research carefully when determining whether to (a) include or exclude reinforcement time in the calculation of baseline response rates, (b) define the target response narrowly or broadly, and (c) measure and consider reporting other potentially relevant response topographies (e.g., emotional responses, novel responses) that may not have been a part of the reinforced response class during baseline.
Finally, although the definitions described by Katz and Lattal (2021) show considerable variation, two characteristics of the extinction burst that appear to be common across definitions, at least implicitly, are that extinction bursts (a) occur close in time to the onset of extinction and (b) disappear with continued exposure to extinction. Thus, a convincing theoretical explanation of the extinction burst should, at a minimum, account for both characteristics.
Prior Theories of Extinction‐Induced Problem Behavior
Three prior theories of extinction‐induced problem behavior are that these side effects of extinction result from frustration, adventitious reinforcement, or schedule‐induction. In what follows, we describe each of these theoretical accounts, and we argue that each one fails to account for the dynamic characteristics of the extinction burst.
Frustration‐Induced Problem Behavior
Dollard et al. (1939) hypothesized that blocking a goal or omitting a scheduled reinforcer results in frustration, which in turn produces aggression. Later versions of the frustration hypothesis stated that aggression represented one of several potential responses produced by frustration (Berkowitz, 1969; Miller, 1941). Similarly, Amsel (1958, 1992) observed that rats ran faster during the second runway of a two‐runway task on trials where they did not receive reinforcement at the completion of the first runway relative to trials where they did. Based on this observation, Amsel posited that the loss of an anticipated reward (e.g., upon completing the first runway) produced an emotional reaction that he labeled primary frustration. He hypothesized that primary frustration, in turn, invigorated a variety of responses, including faster running in the second component of the two‐runway task (i.e., the frustration effect) or a burst of responding at the start of extinction (e.g., Thomas & Papini, 2001).
The major limitations of this theory are that frustration is treated as an independent (or intervening) variable, but it is not well defined and is difficult to scale and manipulate directly. In addition, the presence of this intervening variable is typically inferred from the observed change in the dependent variable, which results in circular reasoning (e.g., faster running indicates that frustration is present, but the inferred frustration is then used to explain the faster running). Finally, as mentioned above, a complete theory of extinction bursts should account for both the occurrence of a response burst at the start of extinction and its rapid remission shortly thereafter. That is, when extinction is implemented with high procedural integrity, extinction bursts tend to be short lived (Katz & Lattal, 2020; Nist & Shahan, 2021), which seems inconsistent with the frustration hypothesis. That is, if extinction causes frustration, and frustration causes a burst of responding, why does bursting not persist longer than it typically does? Surely, extended exposures to extinction are more frustrating than temporary exposures, and more frustration should lead to more bursting, but this assumption is not consistent with the research on extinction bursts. Additionally, Coe et al. (1983) found that cortisol levels in rats, which represent a biological correlate of frustration and stress, continued to increase during extinction long past the time when extinction bursts are typically observed in rats.
Adventitious Reinforcement
Contiguous pairings between a response and a reinforcer can maintain, or partially maintain, the target response, especially after a transition from contingent to time‐based reinforcer deliveries with similar reinforcement rates (Lattal, 1972; Ringdahl et al., 2001; Vollmer et al., 1997). As such, one potential explanation of extinction‐induced problem behavior is that the introduction of extinction results in response variability, and these various responses (e.g., induced aggression) come into contiguous contact with reinforcement (Frederiksen & Peterson, 1977). This phenomenon has been reported in applied studies involving both contingent and response‐independent delivery of reinforcement during function‐based treatment of problem behavior (e.g., Fisher et al., 1993; Vollmer et al., 1997; Wacker et al., 1990). For example, researchers evaluating the effects of FCT during the early 1990s reported that problem behavior and the functional communication response (FCR) can form an adventitious response chain that maintains problem behavior even though the latter response is correlated with extinction (e.g., Fisher et al., 1993; Wacker et al., 1990). That is, the individual displays problem behavior, which is on extinction, and then immediately thereafter displays the FCR, which produces the reinforcer, and the delivery of the reinforcer comes to maintain both problem behavior and the FCR. As such, more recent evaluations of FCT have typically included a changeover delay to prevent reinforcer deliveries from occurring in close temporal proximity to problem behavior (e.g., Borrero et al., 2010; Fisher, Greer, Fuhrman et al., 2018; Fisher, Greer, Mitteer et al., 2018; Greer et al., 2016). Similarly, basic researchers evaluating schedule‐induced aggression routinely include a changeover delay to prevent adventitious reinforcement of aggression (Frederiksen & Peterson, 1977). However, the inclusion of such changeover delays has not appreciably reduced extinction‐induced behavior (cf. Fisher, Greer, Mitteer et al., 2018). Thus, it is unlikely that adventitious reinforcement is responsible for extinction bursts or other extinction‐induced behavior (e.g., aggression). In addition, like the frustration hypothesis, the adventitious‐reinforcement hypothesis fails to account for the observation that bursts typically occur at the start of extinction and resolve shortly thereafter. This is because adventitious pairings of the target response and the delivery of alternative reinforcement could potentially occur at any point during treatment and not just at its onset.
Schedule‐Induced Problem Behavior
Electric shock (e.g., O'Kelly & Steckle, 1939; Ulrich & Azrin, 1962) and other forms of primary aversive stimulation (Ulrich & Azrin, 1962) can induce aggressive behavior when a conspecific is available, and aversive stimuli also can maintain escape or avoidance responding (Dinsmoor et al., 1958; Sidman, 1962). Similarly, Azrin and colleagues (1961, 1966) observed that schedules of positive reinforcement that contained discriminable periods of nonreinforcement (or extinction) can similarly maintain escape behavior or induce aggression. In addition, brief bursts of target responding have been observed immediately after shock delivery during avoidance schedules (e.g., De Villiers, 1974) and at the onset of extinction (e.g., Azrin et al., 1966). Moreover, during both primary aversive stimulation and extinction, these bursts have been described qualitatively as the subject attacking the response apparatus (Keller & Schoenfeld, 1950; Pear et al., 1972). Though electric shock and extinction are clearly not equivalent, the above‐described similarities between the behavioral effects of primary aversive stimulation and periods of extinction during or following schedules of positive reinforcement have led to the conclusion that such periods of extinction represent a form of aversive stimulation. It has also led to the hypothesis that this form of aversive stimulation induces bursts of the target response or aggression and maintains avoidance responding through the same behavioral process as primary aversive stimulation (Frederiksen & Peterson, 1977). Although this hypothesis certainly seems plausible, given the above‐mentioned similarities between the effects of aversive stimulation and extinction, it does not account for the dynamic nature of extinction bursts. That is, extinction bursts, when observed, typically occur early on during extinction (e.g., during the first few sessions—Lerman et al., 1999; the first minute—Katz & Lattal, 2020; Nist & Shahan, 2021; or even the first few seconds, when data are graphed and presented on a fine‐grained, second‐by‐second basis [see the bottom temporal stream in Figure 1 of this paper and the top panel of Figure 2 in Azrin et al., 1966]). Thereafter, the rate of the target response decreases steadily or even rapidly as extinction continues (see the top panel of Figure 2 in Azrin et al., 1966). It is difficult to envision how extinction could become aversive and induce more responding almost immediately after the onset of extinction and then shortly thereafter reverse its effects on the target response and produce progressively less responding.
Figure 1.
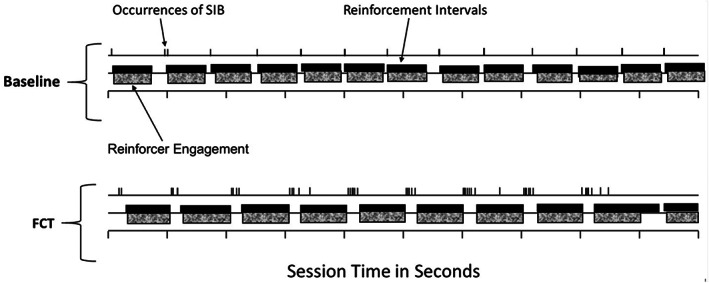
Temporal Patterns of SIB and Reinforcer Engagement Note. The top temporal stream shows occurrences of self‐injurious behavior (SIB; tick marks), periods of reinforcer delivery (black bars), and periods of reinforcer engagement (patterned bars) during baseline when responding was on a fixed‐ratio 1 schedule of reinforcement. The bottom temporal stream shows this same information during a representative session of functional communication training (FCT) when therapists guided Carson (from Fisher, Greer, Mitteer et al., 2018) to emit the functional communication response (not shown) according to a 5‐s prompt delay. Extinction bursts are visible during FCT when reinforcer consumption was unavailable to compete with SIB during the 5‐s prompt delays.
It seems reasonable to posit that experiencing longer periods of extinction should be more aversive than experiencing shorter periods of extinction. Consistent with this supposition, Thomas (1965) compared the effects of different durations of time‐out from reinforcement and found that 5‐min periods of time‐out produced more avoidance responses than briefer durations of time‐out. Similarly, Kaufman and Baron (1968) found that 120‐s periods of time‐out generally produced longer interresponse times than shorter durations of time‐out.
If the schedule‐induced hypothesis is correct, then extinction bursts should begin after at least some exposure to extinction (e.g., after 10 s) so that the individual experiences and discriminates the aversiveness of a period of nonreinforcement, and extinction bursts should be most probable at the point in time when extinction is most aversive (e.g., perhaps after about 5 min of nonreinforcement, based on the Thomas, 1965 study). Contrary to the schedule‐induced hypothesis, our data in the bottom panel of Figure 1 and the data in the top panel of Figure 2 in Azrin et al. (1966) show that extinction bursts can occur almost immediately after the onset of extinction, and Figure 2 from Azrin et al. shows that responding can rapidly decrease below baseline levels well before the time at which a period of nonreinforcement should reach its peak level of aversiveness.
Extinction Bursts as Temporally Weighted Choice Responding
As previously indicated, a satisfactory explanation of extinction bursts should account for why the bursts occur shortly after the start of extinction and disappear soon thereafter. Shahan (2022) suggested that a refinement of the concatenated matching law, called the TWML, accounts for both characteristics of the extinction burst. In the remainder of this paper, we discuss the TWML, its implications for clinical intervention, and its potential strengths and limitations.
The matching law (Herrnstein, 1961) is a quantitative theory of choice postulating that individuals allocate their behavior to response options (i.e., B 1 and B 2 ) according to the relative reinforcement rates obtained from those options (i.e., R 1 and R 2 ), such that,
| (1) |
Recently, researchers have begun to extend the matching law, both conceptually and quantitatively, to conditions in which one or more responses are correlated with extinction, such as resurgence (e.g., Greer & Shahan, 2019; Shahan et al., 2020; Shahan & Craig, 2017). One potential advantage of this approach is that it provides a way to analyze and predict how responding reacts to changing reinforcement contingencies, like the introduction of extinction for problem behavior with or without differential reinforcement of an appropriate alternative response (DRA). However, it is important to note that the original version of the matching law described by Equation 1 is difficult to apply quantitatively when one or both responses are associated with extinction, as typically occurs at the start of a function‐based treatment like FCT. This is because Equation 1 is based only on current reinforcement rates for the two responses; thus, the equation does not quantify how a history of reinforcement affects responding once extinction is implemented. As a result, the equation suggests that when extinction is implemented for one response (e.g., problem behavior), both the reinforcement rate and the predicted response allocation toward that option drop immediately to zero. Unfortunately, extinction typically does not have such immediate and dramatic effects on behavior.
Shahan and Craig (2017) described one way to address this limitation of the matching law by providing a means to calculate how extinction affects the value of response options over time. Their approach is based on the concatenated matching law (Baum & Rachlin, 1969), which suggests that individuals allocate their behavior (i.e., B 1 and B 2 ) in relation to the values (i.e., V 1 and V 2 ) of those options such that,
| (2) |
where value depends not only on the obtained rate of reinforcement but also on the magnitude, quality, and immediacy of the reinforcers associated with each response. To determine how extinction affects the current value of the response options, Shahan and Craig calculated the current value of each response option by weighting all known previously experienced reinforcement rates (i.e., the reinforcement history) according to their relative recencies. That is, based on the postulate that more recently experienced reinforcement rates influence current responding more than those experienced further in the past, they weighted reinforcement rates according to the temporal weighting rule (Devenport & Devenport, 1994). Mathematically, the weightings (i.e., w x ) for a series of past experiences are calculated as the relative recency of those experiences such that,
| (3) |
where t represents time from the previous experience to the present, and n represents the number of previous experiences under consideration. Then, the value of an option is obtained by multiplying each experienced reinforcement rate (i.e., R x ) by the appropriate weighting (i.e., w x ) for when it occurred in the past and summing across all the experiences such that,
| (4) |
When reinforcement rates remain constant across time, Equation 4 returns the programmed reinforcement rate as the value of the option. When extinction is in effect, R x = 0 for that specific time in the series of experiences. As time in extinction continues, V decreases because the weightings for previous reinforcement experiences generated by Equation 3 decrease hyperbolically as they drift into the past. According to this rule, when problem behavior first contacts extinction, it still has value due to its recent history of reinforcement. For example, after five baseline sessions in which problem behavior produced reinforcement on a VI 30‐s schedule (producing 120 reinforcers per hour), its value would decrease from 120 in baseline to 51.2 in the first session of extinction (introduced in Session 6). However, the value of problem behavior decreases rapidly during the initial sessions of extinction, as older reinforcement experiences are replaced by experiences with extinction (e.g., from 51.2 in the first session of extinction to 33.3 in the second), and the value of problem behavior decreases more gradually over subsequent periods of time (e.g., from 20.2 in the fourth session of extinction to 17.1 in the fifth session of extinction).
Applying the TWML to Resurgence
Shahan and Craig (2017) first applied the temporal weighting rule within the concatenated matching law to explain resurgence, and they called this approach resurgence as choice (RaC) theory. In this paper, we use the more general term TWML (Shahan, 2022) rather than RaC because we are applying the model beyond just resurgence. Resurgence is an increase in the rate of a target response (e.g., problem behavior) previously reduced by differential reinforcement when reinforcement conditions worsen (e.g., when an alternative response [e.g., the FCR] contacts a disrupter [e.g., extinction]). As applied to FCT and resurgence, the TWML suggests that allocation of behavior to a problem behavior and an appropriate alternative behavior are governed by the relative values associated with those behaviors such that,
| (5) |
where B T and B Alt are the rates of problem behavior and the FCR, respectively, and V T and V Alt are the reinforcement values associated with those options. In general terms, the equation suggests that FCT reduces problem behavior relative to baseline because extinction reduces the value of this response while the value of the FCR increases simultaneously, and thus the allocation of responding to problem behavior decreases. When problem behavior first contacts extinction during FCT, the TWML predicts that it still has considerable value due to its recent history of reinforcement during baseline. But, as treatment with FCT proceeds, the value of problem behavior decreases rapidly because the more recent experiences with problem behavior not producing reinforcement become more influential (or carry more weight) than reinforcers delivered in prior baseline sessions. Over time, however, the rate at which the value of problem behavior decreases slows (according to a hyperbolic decay function). Later, when the FCR stops producing reinforcement (e.g., because a parent is tending to an infant sibling), the value of the FCR drops rapidly because it has just contacted extinction. By contrast, the current value of problem behavior decreases more slowly because the current extinction sessions would be much less influential than those that occurred when extinction was first introduced for problem behavior at the start of FCT (see Greer & Shahan, 2019, and Shahan & Greer, 2021, for more applied examples).
During this initial period in which both responses are now on extinction and the value of the FCR is decreasing more rapidly than the value of problem behavior, the relative value of problem behavior increases temporarily and produces resurgence. For example, if the current values of problem behavior (i.e., V T ) and the FCR (i.e., V Alt ) are 50 and 100, respectively, the relative value of problem behavior would be 50 / (50 + 100) or 0.33. If, during a subsequent resurgence challenge, the value of problem behavior then drops by 10 and the value of the FCR by 50, because that latter response contacted extinction more recently, the relative value of problem behavior would increase to 0.44 [40 / (40 + 50)]. Under such a condition, the matching law, adjusted using the temporal weighting rule, predicts that the relative rate of problem behavior should increase because its relative value increases, and we should observe resurgence of problem behavior.
Applying the TWML to Extinction Bursts
Employing this general approach, Shahan (2022) suggested that the same basic processes may account for the extinction burst. With the extinction burst, the two response options during baseline are engaging in problem behavior (i.e., B T ) or behavior related to consuming or otherwise engaging with the reinforcer when it is present (i.e., B r ) such that,
| (6) |
where V T and V r are the values associated with those two options. When the reinforcer is present, allocation of behavior in clock time (i.e., including reinforcement time) is likely to strongly favor B r over B T , and the relative rate of problem behavior (i.e., B T ) is likely to be low. However, when extinction is implemented and access to the reinforcer (and thus reinforcement‐related behavior) is eliminated, there is a temporary increase in the relative value of the target behavior, thus producing a temporary increase in allocation to the target behavior. Such a reallocation could be the source of the extinction burst (see Shahan, 2022, for full quantitative details).
For example, during baseline, an individual may typically emit problem behavior (e.g., head banging), which produces its functional reinforcer (e.g., on an FR 1 schedule). While the reinforcer is available (e.g., 20‐s access to an iPad), the individual typically allocates their responding to consuming that reinforcer, and few or no instances of problem behavior occur during the 20‐s reinforcement interval. That is, when given a choice between emitting head banging and consuming the reinforcer, the individual almost always chooses the latter response option. Thus, as described in the earlier example, an efficient participant might head bang 12 times in a 5‐min session and produce a response rate of 2.4 responses per minute if we include reinforcement time in the calculation. By contrast, when treatment with extinction alone is initiated, the individual no longer has the option to consume the reinforcer. During this period at the start of treatment, the TWML predicts an increase in problem behavior (i.e., an extinction burst) because its relative value increases temporarily. This temporary increase in relative value occurs because (a) reinforcer consumption is no longer available as a competing response option, and (b) the value of problem behavior remains high due to its recent history of reinforcement during baseline. That is, the TWML predicts a temporary increase in the target response at the start of extinction because, in baseline, reinforcer consumption competes with the target response. However, at the start of extinction (when extinction is implemented alone), reinforcer consumption is no longer available as an alternative response option and, therefore, the relative value of the target response increases.
The TWML also provides a reasonable explanation for why extinction bursts tend to be short lived. That is, according to the TWML, extinction bursts occur at the start of extinction because that is when the effects of prior reinforcer deliveries for problem behavior have the most influence or value. So, at the start of extinction, when the value of problem behavior is still high and reinforcer consumption is no longer a competing response option, the individual allocates nearly all their time to problem behavior, and an extinction burst is observed. As time in extinction increases, the influence or value of the reinforcers delivered for problem behavior during baseline decreases. Thus, as the individual has increased experience with extinction of problem behavior, the value of problem behavior decreases rapidly, and the rate of problem behavior decreases to match its value, thereby ending the extinction burst. The TWML appears to be the only theory that explains why bursts occur when extinction is initiated and why they resolve shortly thereafter.
Applying the TWML to DRA Treatments
When treatment with DRA (e.g., FCT) is initiated, rather than extinction alone, the individual has the option to access the functional reinforcer via the alternative response, so the relative value of problem behavior would be lower when DRA is initiated relative to extinction alone. Thus, the TWML predicts lower levels of problem behavior at the start of DRA than at the start of extinction alone, a prediction that is consistent with the empirical findings of Lerman and Iwata (1995) and Lerman et al. (1999).
Nevertheless, extinction bursts, as defined in the clinical literature, sometimes occur when problem behavior is treated with DRA. The TWML similarly suggests that these bursts during DRA should occur during times when reinforcement is unavailable (i.e., when both problem behavior and the alternative response contact a period of extinction). For example, Fisher, Greer, Mitteer et al. (2018; see also DeRosa et al., 2015) compared the effects of briefer (e.g., 1‐ to 2‐s) and longer (e.g., 5‐ to 40‐s) periods in which reinforcement was unavailable during FCT. Such periods of nonreinforcement can occur when initiating FCT due to (a) prompt delays designed to promote independent communication responses (DeRosa et al., 2015; Fisher, Greer, Mitt et al., 2018) or (b) changeover delays designed to prevent adventitious reinforcement of problem behavior (e.g., Borrero et al., 2010; Greer et al., 2016). Fisher et al. observed extinction bursts, as defined by Lerman et al. (1999), in five of six applications of the conditions with 5‐ to 40‐s periods of nonreinforcement and in none of the six applications with 1‐ to 2‐s periods of nonreinforcement.
Fisher, Greer, Mitteer et al. (2018) attributed these bursts of problem behavior to the presence of the establishing operation during periods of nonreinforcement. That is, they hypothesized that during periods of nonreinforcement, the establishing operation evokes responses that have produced the reinforcer in the past, problem behavior in this case. However, like the frustration, adventitious‐reinforcement, and schedule‐induced hypotheses described above, this account of extinction bursts does not address the dynamic nature of extinction bursts. That is, extinction bursts typically occur early on during extinction (e.g., during the first few sessions—Lerman et al., 1999, or even just the first minute—Katz & Lattal, 2020). Thereafter, rates of the target response decrease, even though the establishing operation remains in effect. Thus, the establishing‐operation account offered by Fisher et al. provides a potential explanation of why bursts of problem behavior may occur at the start of extinction (with or without DRA), but it does not explain why rapid reductions in problem behavior typically follow an extinction burst. That is, if the establishing operation remains in effect, why does problem behavior decrease rapidly following an initial burst? By contrast, the TWML provides an explanation of the dynamic nature of extinction bursts, as described above.
According to the TWML, extinction bursts should occur during DRA treatments when reinforcer consumption is not available as a competing response and the current value of problem behavior is considerably higher than the value of the alternative response (see Shahan, 2022, for equations). That is, during a DRA treatment like FCT, individuals are likely to allocate their responding among three options: (a) problem behavior, (b) the alternative response (e.g., the FCR), and (c) reinforcer consumption. The TWML predicts that reinforcer consumption should occur whenever it is available; the alternative response should predominate when reinforcer consumption is unavailable and the alternative response reliably produces the reinforcer; and problem behavior should increase (i.e., an extinction burst) when its value well exceeds the value of the alternative response. During FCT, extinction bursts are most likely to occur during periods of nonreinforcement for the FCR shortly after treatment commences because that is when the influence or value of prior reinforcer deliveries for problem behavior is still relatively high, the value of the FCR is relatively low (because it has not had an extended history of reinforcement), and reinforcer consumption is temporarily unavailable as a response option.
Figure 1 shows the within‐session patterns of SIB and reinforcer engagement in relation to session time and reinforcement intervals during a typical baseline session (top timeline) and a typical FCT session from the extended establishing‐operation condition (bottom timeline) for the participant named Carson in Fisher, Greer, Mitteer et al. (2018). Consistent with the TWML, during this typical baseline session: (a) Carson almost always displayed a single SIB shortly after a reinforcement interval ended, which then produced the next reinforcement interval, and (b) during nearly all reinforcement intervals, Carson engaged with the reinforcer throughout most of the interval and emitted no SIB.
During the FCT session, Carson typically displayed SIB shortly after a reinforcement interval ended, just as in baseline. However, the first occurrence of SIB did not produce the next reinforcement interval and more occurrences of SIB typically followed, thus producing the observed extinction burst. That is, consistent with the TWML, the extinction burst occurred at the start of treatment during a period of extinction when the relative value of SIB was high and the FCR was not yet a viable option because Carson had not learned to produce reinforcement via this alternative response. The next reinforcement interval only came after the therapist physically prompted Carson to emit the FCR, which occurred after a 5‐s prompt delay or after a 3‐s changeover delay with no SIB, whichever came later. These data are consistent with the hypothesis that individuals choose to engage with the functional reinforcer when it is available and to emit the response that historically produced that reinforcer primarily when the reinforcer is unavailable. The data are also consistent with the hypothesis that bursts of problem behavior occur shortly after FCT commences when neither problem behavior (because it is on extinction) nor the alternative response (because the individual has not yet mastered the new contingency) efficiently produces the next reinforcement interval.
Prior applied studies on the effects of time‐based schedules of alternative reinforcement on problem behavior, often called noncontingent reinforcement (Vollmer et al., 1993), have also shown similar patterns of responding that are consistent with the TWML (Fisher et al., 1999; Hagopian et al., 2000; Marcus & Vollmer, 1996). That is, participants allocate their responding to consuming the free, alternative reinforcer that is delivered via time‐based schedules whenever it is available and allocate their responding to problem behavior when free reinforcement is unavailable. For example, Fisher et al. (1999) compared the effectiveness of denser and leaner schedules of alternative reinforcement on problem behavior when the investigators continued to deliver contingent reinforcement for problem behavior on an FR 1 schedule. Consistent with the TWML, participants emitted more problem behavior under the leaner time‐based schedule relative to the denser one. Moreover, within‐session response patterns showed that they emitted problem behavior and accessed contingent reinforcement primarily when time‐based reinforcement was unavailable (i.e., during the interreinforcement intervals for the time‐based schedules). Thus, the participants preferred consuming the time‐based reinforcer over emitting problem behavior (and then obtaining contingent reinforcement), and this accounted for the greater reductions in problem behavior during the denser versus leaner time‐based schedules. That is, time‐based reinforcement was available for more of the session time with the denser schedules, and problem behavior primarily occurred when the time‐based reinforcement was unavailable.
Implications of the TWML for Clinical Intervention
Because the TWML is a refined form of the concatenated matching law, in which extinction bursts are hypothesized to be a function of the value of the target response relative to the value of all concurrently available responses, the theory provides specific predictions about variables that can be manipulated to increase or decrease the probability of extinction bursts when extinction‐based treatments are initiated. That is, the concatenated matching law states that increasing the rate, magnitude, or quality of reinforcement relative to the alternative response options (e.g., the FCR, reinforcer engagement) can increase the relative value of the target response (e.g., problem behavior). For example, delivering a higher rate of reinforcement for problem behavior during baseline (e.g., FR 1) and a lower rate of reinforcement for the alternative response during treatment (e.g., FR 5) should increase the likelihood of an extinction burst. According to the TWML, the larger the decrease in the rate of reinforcement that occurs with the switch from baseline to treatment, the larger the observed extinction burst. Conversely, according to the TWML, preventing a decrease in the rate of reinforcement when treatment is initiated should prevent an extinction burst and lead to a more rapid reduction in problem behavior.
More generally, the TWML predicts that manipulating any variable that affects choice responding (e.g., rate, magnitude, immediacy, and quality of reinforcement) can increase or decrease the probability of an extinction burst. That is, individuals are more likely to display a burst of problem behavior if they experience a decrease in the rate, magnitude, immediacy, or quality of reinforcement when treatment is initiated, and less likely to show bursting if the rate, magnitude, immediacy, or quality of reinforcement remains the same or increases when treatment starts.
One advantage of quantitative theories of behavior like the TWML is that they allow behavior analysts to model and make predictions about the potential outcome of a clinical or experimental manipulation before finalizing and conducting the intervention or study. For example, Figure 2 shows the formal quantitative predictions of the TWML for a hypothetical experiment comparing the effects of extinction implemented alone or in combination with relatively denser and leaner DRA schedules. For these mathematical simulations, we programmed a VI 1.5‐s schedule for problem behavior during each baseline. In the dense DRA condition, we programmed the same VI 1.5‐s schedule for the alternative response and placed problem behavior on extinction. In the lean DRA condition, we programmed a VI 90‐s schedule for the alternative response and placed problem behavior on extinction.
Figure 2.
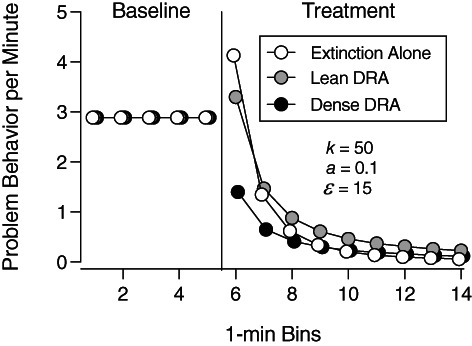
Predictions of the TWML When Reinforcement Rates Drop or Stay the Same During Treatment Note. The temporally weighted matching law (TWML) predicts that extinction bursts are correlated with reductions in the rate of reinforcement when treatment commences, with larger decreases in the rate of reinforcement producing larger initial increases in responding. DRA = differential reinforcement of alternative behavior. See Shahan (2022) for TWML parameter definitions and full quantitative details.
As Figure 2 shows, these quantitative simulations predict that a relatively large extinction burst will occur when extinction is implemented alone, because implementing extinction alone produces the largest drop in the rate of reinforcement (i.e., from about 2.8 reinforcers per minute to 0 reinforcers per minute). The TWML predicts a smaller extinction burst in the lean DRA condition, because it produces a smaller drop in the rate of reinforcement (i.e., from about 2.8 reinforcers per minute to about 0.6 reinforcers per minute). Finally, the TWML predicts a rapid drop in the rate of problem behavior without an extinction burst in the dense DRA condition, because it does not produce a drop in the rate of reinforcement (i.e., 2.8 reinforcers per minute in baseline and in treatment).
We have conducted parallel simulations in which we manipulated other reinforcement parameters (e.g., reinforcer quality remaining the same or reduced at the start of treatment), and those simulations produced results consistent with the results shown in Figure 2. These quantitative predictions of the TWML need to be tested empirically. If validated, they suggest that clinicians should work to ensure that the rate, magnitude, immediacy, and quality of reinforcement remain the same or increase relative to baseline when they initiate differential reinforcement treatments like FCT.
The TWML also predicts that a decrease in one reinforcement parameter that might ordinarily produce an extinction burst (e.g., reduced rate of reinforcement at the start of treatment) could be counteracted by manipulating another reinforcement parameter (e.g., increasing the quality of reinforcement). For example, individuals with no preexisting communication repertoire are likely to experience a reduction in reinforcement rate at the start of FCT until they learn and master the new FCR. According to the TWML, we may be able to mitigate extinction bursts for such individuals by increasing the quality of reinforcement delivered for the FCR to counteract the negative effects of the decrease in reinforcement rate. Although this technique for mitigating extinction bursts is hypothetical and untested at this point, it represents a plausible extension of principles of choice responding and behavioral economics that have considerable empirical support (DeLeon et al., 2021; Fisher & Mazur, 1997; Podlesnik et al., 2021; Weinsztok & DeLeon, 2021).
Illustrative Translational Data
As previously mentioned, the TWML predicts that large drops in reinforcer‐consumption time at the start of treatment should increase the likelihood of an extinction burst; thus, preventing drops in reinforcer‐consumption time should prevent extinction bursts. To illustrate these predicted effects, we conducted a baseline in which the target response (i.e., hitting a pad made of foam covered with vinyl) produced reinforcement, followed by a phase in which we discontinued reinforcement for pad hits (i.e., extinction) and delivered reinforcement on either a dense or lean time‐based schedule. We randomly assigned one child to receive the dense schedule and the other child to receive the lean schedule during treatment. Based on the TWML, we hypothesized that (a) the child exposed to the dense schedule during treatment would show a rapid reduction in pad hits without an extinction burst and (b) the child exposed to the lean schedule during treatment would show a burst of pad hits before the response decreased to low levels.
Participants
Allie was a 5‐year‐old female and Sam was a 4‐year‐old male at the time of the study. Both individuals showed typical development; they spoke using full sentences, carried on conversations with the experimenters, and followed the experimenter's instruction (e.g., “Sit here”; “If you hit the pad, like this, I will give you the iPad”). Neither participant had a previous history of receiving reinforcement for engaging in the target response (i.e., pad hit).
Procedure
Pretraining
At the start of the session, the experimenter allowed the participant 2 s to hit the pad independently (neither child did). After the 2 s elapsed, the experimenter physically guided the child to hit the pad and then immediately delivered the reinforcer (i.e., an edible every 10 s along with access to attention and an iPad for the entire 20‐s reinforcement interval for Allie; access to attention and an iPad for the entire 20‐s reinforcement interval for Sam). After every two trials, the experimenter increased the duration of the prompt delay according to the following progression: 2 s, 5 s, 10 s, 20 s. After the child hit the pad independently three consecutive times, pretraining ended.
Baseline
During baseline, the experimenter terminated the prompts and delivered reinforcement for pad hits as described above on a VR 2 schedule. Baseline ended after the participant experienced 15 reinforcement intervals.
Dense Time‐Based Reinforcement (Allie Only)
During this phase, the experimenter discontinued reinforcement for pad hits (i.e., extinction), delivered one edible every 10 s, and provided continuous access to attention and the iPad throughout the 5‐min phase.
Lean Time‐Based Reinforcement (Sam Only)
During this phase, the experimenter discontinued reinforcement for pad hits (i.e., extinction) and provided 20‐s access to attention and the iPad once every 60 s during the 5‐min phase (e.g., access to attention and the iPad during the first 20 s of the first minute, the first 20 s of the second minute, etc.).
Results and Discussion
Figure 3 shows the results for Allie in the top panel and the results for Sam in the bottom panel. As can be seen, Allie showed relatively high but variable rates of pad hits during baseline. During dense, time‐based reinforcement, pad hits immediately decreased to zero and remained at zero throughout the 5‐min phase, presumably because the reinforcer was available continuously and reinforcer consumption effectively competed with pad hits. Sam showed relatively high and somewhat less variable rates of pad hits during baseline. However, during lean, time‐based reinforcement, Sam displayed an extinction burst during the first minute of this phase, when pad hits increased to a rate higher than all the rates observed during the 1‐min intervals in baseline. Thereafter, pad hits decreased rapidly to zero.
Figure 3.
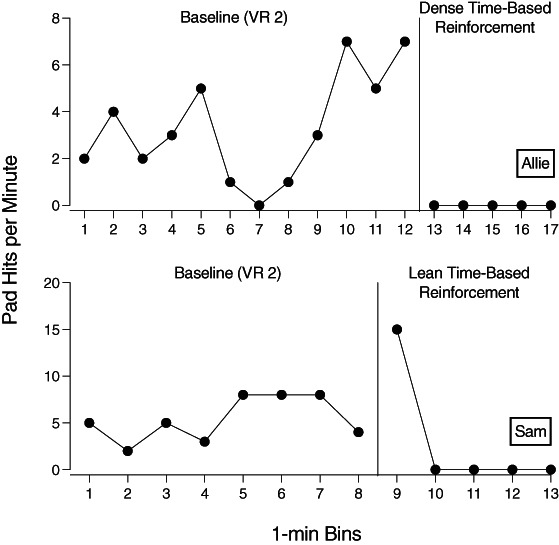
Rates of Pad Hits During Baseline and Dense and Lean Time‐Based Schedules
Note. VR = variable ratio.
Thus, the child exposed to the dense schedule of reinforcement during the treatment phase showed a rapid (i.e., immediate) decrease in pad hits and did not show an extinction burst, whereas the child exposed to the lean schedule showed an extinction burst before pad hits decreased to low levels. In addition, no pad hits occurred during reinforcer‐consumption time during treatment for either participant. For Sam, pad hits commenced about 5 s after the first reinforcer‐consumption time ended during treatment. He emitted 15 pad hits in the next 23 s (i.e., the extinction burst), and then pad hits ceased for the remainder of the phase. These results comport to the conceptual predictions of the TWML. Unfortunately, we could not effectively test the quantitative predictions of the TWML with these data. We collected all the baseline and treatment data in one 15‐ to 20‐min session, and the limited amount of data and high degree of variability did not allow for reliable estimates of fitted model parameters.
Potential Strengths and Limitations of the TWML
Because the TWML is a new refinement of the concatenated matching law, it has not received sufficient empirical testing to fully describe its strengths and limitations. Clearly, the TWML does not account for all the behavioral phenomena that occur during extinction in its current form. However, a potential strength of the model is that it is based on the concatenated matching law, and it should be applicable to those variables that traditionally have been addressed by the matching law (e.g., reinforcer rate, delay, and quality; response effort), but under dynamically changing reinforcement conditions. That is, the TWML generally predicts that the variables that affect relative response rates during steady‐state, baseline conditions similarly affect relative response rates when reinforcement is reduced or eliminated, with the caveat that those effects are mediated by the timing (or recency) of past reinforcer deliveries. Another potential strength of the TWML is that adding terms to the equation to account for discrimination learning could potentially allow the model to address variables like prior exposure(s) to extinction or the introduction of extinction in one context (e.g., the clinic) but not in another (e.g., the home; see Shahan et al., 2020, for a recent example).
A limitation of the current form of the TWML is that it does not account for several potentially important variables involved in the extinction process. For example, the current TWML does not account for the effects of habituation during escape extinction (McSweeney & Swindell, 2002; in part, because the TWML has only been applied to appetitive reinforcement schedules). In addition, the TWML does not account for the emergence of new responses during extinction (e.g., extinction‐induced aggression). Thompson and Bloom (1966) described a temporal pattern of extinction bursts of the target response, which typically occurred at its highest levels during the first minute of extinction, followed by extinction‐induced aggression, which typically occurred at its highest levels during the second and third minutes of extinction. This temporal covariance between extinction bursts and extinction‐induced aggression suggests that aggression, and perhaps other forms of extinction‐induced variability, may be most likely to occur after the relative value of the target response decreases substantially. If this is true, then the TWML could potentially predict the onset of response variability during extinction. However, even if the TWML can predict when new responses are likely to emerge during extinction, the theory does not provide a conceptual explanation or basis for the generation of new responses during extinction, and we should look elsewhere for such explanations (Machado, 1993; Nergaard & Holth, 2020; Neuringer et al., 2001).
Another potential limitation is that in the clinical literature on function‐based treatments, like FCT and the time‐based delivery of alternative reinforcement, we sometimes observe that problem behavior decreases to zero in the first treatment session (see participants Gordon and Henry in Figure 2 of Betz et al., 2013), and such immediate and total decreases in responding are not predicted by the TWML. Immediate decreases in problem behavior to near‐zero levels likely occur when participants (a) receive their initial FCT training outside of the experimental sessions and prior to the formal introduction of FCT (as occurred with Gordon and Henry in Betz et al., 2013) or (b) have well‐established rule‐governed behavior and the experimenters provide the participants with contingency‐specifying rules (e.g., “I will give you the iPad back if you say ‘iPad please’, but not if you hit me.”; cf. Tiger & Hanley, 2004). In the first scenario, the TWML is not applicable because the change in reinforcement conditions occurred outside of the experimental preparation. In the second scenario, the TWML is not applicable because the observed change in responding resulted from instructional (or stimulus) control, and matching‐based quantitative models like the TWML are most relevant to concurrently available response options primarily under the control of consequent stimuli.
Finally, the TWML has difficulty dealing with large temporal gaps in the data being analyzed, which can sometimes occur in clinical settings. For example, an individual with severe problem behavior may attend clinic 2 days in a row and go through most of the baseline sessions, then be away from the clinic for a week due to illness, and then return to clinic for 3 days to finish the baseline and initiate the treatment sessions. The week that the individual spent away from the clinic due to illness can greatly alter the ability of the TWML to fit and explain the individual's observed results.
Summary
In this paper, we discussed the strengths and limitations of prior conceptualizations of the extinction burst and described a reconceptualization of the extinction burst based on a revised version of the concatenated matching law called the TWML. The TWML calculates the current value of the target response by temporally weighting all past reinforcer deliveries, with more recent deliveries weighted more heavily than preceding ones, based on the postulate that recent experiences influence responding more so than prior experiences. In addition, when applied to the extinction burst, the TWML conceptualizes reinforcer engagement or consumption as a response option that reliably competes with the target response during baseline. However, reinforcer consumption is unavailable when extinction is implemented alone, or it may be available less reliably when extinction of the target response is combined with DRA or the time‐based delivery of alternative reinforcement. When reinforcement time is eliminated during extinction alone or substantially reduced during DRA, the relative value of the target response increases, which in turn can increase its rate, thus producing an extinction burst. The TWML also explains why extinction bursts tend to be short lived. That is, extinction bursts are temporary because the relative value of problem behavior decreases as time in extinction increases and its history of reinforcement moves further into the past.
Finally, because the TWML is a revised form of the concatenated matching law, it posits that any reinforcement parameter that influences choice responding (e.g., reinforcer magnitude) can increase or decrease the probability of an extinction burst. For example, large decreases in the magnitude of reinforcement when FCT is initiated should increase the probability of an extinction burst and preventing large decreases in the magnitude of reinforcement should decrease the probability of an extinction burst. Clinicians who wish to prevent or mitigate extinction bursts of problem behavior should consider the predictions of the TWML when developing and implementing extinction‐based treatments. In addition, applied researchers should test the predictions of the TWML with individuals with autism or intellectual disability referred for the treatment of problem behavior under typical clinical conditions. Finally, basic researchers should test the predictions of the TWML under a wide range of controlled conditions to provide a formal assessment of the quantitative predictions of the theory, evaluate its boundary conditions, and identify potential clinical refinements.
Grants 2R01HD079113, 5R01HD083214, and 5R01HD093734 from The National Institute of Child Health and Human Development provided partial support for this work.
Determined by extracting the two most extreme response patterns in Figure 4 from Skinner (1938) using the freely available program, WebPlotDigitizer (Rohatgi, 2020). The lowest and highest values were 2.2 and 3.1 responses per minute across 14.9 and 13.6 min of steady‐state responding, respectively.
Associate Editor, Dorothea Lerman
REFERENCES
- Amsel, A. (1958). The role of frustrative nonreward in noncontinuous reward situations. Psychological Bulletin, 55(2), 102–119. 10.1037/h0043125 [DOI] [PubMed] [Google Scholar]
- Amsel, A. (1992). Frustration theory: Many years later. Psychological Bulletin, 112(3), 396–399. 10.1037/0033-2909.112.3.396 [DOI] [PubMed] [Google Scholar]
- Azrin, N. H. (1961). Time‐out from positive reinforcement. Science, 133(3450), 382–383. 10.1126/science.133.3450.382 [DOI] [PubMed] [Google Scholar]
- Azrin, N. H. , Hutchinson, R. R. , & Hake, D. F. (1966). Extinction‐induced aggression. Journal of the Experimental Analysis of Behavior, 9(3), 191–204. 10.1901/jeab.1966.9-191 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baum, W. M. , & Rachlin, H. C. (1969). Choice as time allocation. Journal of the Experimental Analysis of Behavior, 12(6), 861–874. 10.1901/jeab.1969.12-861 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berkowitz, L. (1969). The frustration‐aggression hypothesis revisited. In Berkowitz L. (Ed.), Roots of aggression. Atherton Press. [Google Scholar]
- Betz, A. M. , Fisher, W. W. , Roane, H. S. , Mintz, J. C. , & Owen, T. M. (2013). A component analysis of schedule thinning during functional communication training. Journal of Applied Behavior Analysis, 46(1), 219–241. 10.1002/jaba.23 [DOI] [PubMed] [Google Scholar]
- Blough, D. S. (1959). Delayed matching in the pigeon. Journal of the Experimental Analysis of Behavior, 2(2), 151–160. 10.1901/jeab.1959.2-151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borrero, C. S. , Vollmer, T. R. , Borrero, J. C. , Bourret, J. C. , Sloman, K. N. , Samaha, A. L. , & Dallery, J. (2010). Concurrent reinforcement schedules for problem behavior and appropriate behavior: Experimental applications of the matching law. Journal of the Experimental Analysis of Behavior, 93(3), 455–469. 10.1901/jeab.2010.93-455 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borthwick‐Duffy, S. A. (1994). Epidemiology and prevalence of psychopathology in people with mental retardation. Journal of Consulting and Clinical Psychology, 62(1), 17–27. 10.1037/0022-006X.62.1.17 [DOI] [PubMed] [Google Scholar]
- Briggs, A. M. , Dozier, C. L. , Lessor, A. N. , Kamana, B. U. , & Jess, R. L. (2019). Further investigation of differential reinforcement of alternative behavior without extinction for escape‐maintained destructive behavior. Journal of Applied Behavior Analysis, 52(4), 956–973. 10.1002/jaba.648 [DOI] [PubMed] [Google Scholar]
- Briggs, A. M. , Fisher, W. W. , Greer, B. D. , & Kimball, R. T. (2018). Prevalence of resurgence of destructive behavior when thinning reinforcement schedules during functional communication training. Journal of Applied Behavior Analysis, 51(3), 620–633. 10.1002/jaba.472 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown, K. R. , Greer, B. D. , Craig, A. R. , Sullivan, W. E. , Fisher, W. W. , & Roane, H. S. (2020). Resurgence following differential reinforcement of alternative behavior implemented with and without extinction. Journal of the Experimental Analysis of Behavior, 113(2), 449–467. 10.1002/jeab.588 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carr, E. G. , & Durand, V. M. (1985). Reducing behavior problems through functional communication training. Journal of Applied Behavior Analysis, 18(2), 111–126. 10.1901/jaba.1985.18-111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Catania, A. C. (2013). Learning (5th ed.). Sloan Publishing. [Google Scholar]
- Coe, C. L. , Stanton, M. E. , & Levine, S. (1983). Adrenal responses to reinforcement and extinction: Role of expectancy versus instrumental responding. Behavioral Neuroscience, 97(4), 654–657. 10.1037/0735-7044.97.4.654 [DOI] [PubMed] [Google Scholar]
- Crocker, A. G. , Mercier, C. , Lachapelle, Y. , Brunet, A. , Morin, D. , & Roy, M.‐E. (2006). Prevalence and types of aggressive behaviour among adults with intellectual disabilities. Journal of Intellectual Disability Research, 50(9), 652–661. 10.1111/j.1365-2788.2006.00815.x [DOI] [PubMed] [Google Scholar]
- DeLeon, I. G. , Fernandez, N. , Goldman, K. J. , Schieber, E. , Greer, B. D. , & Reed, D. D. (2021). Behavioral economics. In Fisher W. W., Piazza C. C., & Roane H. S. (Eds.), Handbook of applied behavior analysis, 2 nd Edition (pp. 115–132). Guilford Publishing. [Google Scholar]
- DeRosa, N. M. , Fisher, W. W. , & Steege, M. W. (2015). An evaluation of time in establishing operation on the effectiveness of functional communication training. Journal of Applied Behavior Analysis, 48(1), 115–130. 10.1002/jaba.180 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devenport, L. D. , & Devenport, J. A. (1994). Time‐dependent averaging of foraging information in least chipmunks and golden‐mantled ground squirrels. Animal Behaviour, 47(4), 787–802. 10.1006/anbe.1994.1111 [DOI] [Google Scholar]
- De Villiers, P. A. (1974). The law of effect and avoidance: A quantitative relationship between response rate and shock‐frequency reduction. Journal of the Experimental Analysis of Behavior, 21(2), 223–235. 10.1901/jeab.1974.21-223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dinsmoor, J. A. , Hughes, L. H. , & Matsuoka, Y. (1958). Escape‐from‐shock training in a free‐response situation. The American Journal of Psychology, 71(2), 325–337. 10.2307/1420077 [DOI] [PubMed] [Google Scholar]
- Dollard, J. , Miller, N. E. , Doob, L. W. , Mowrer, O. H. , & Sears, R. R. (1939). Frustration and aggression. Yale University Press. 10.1037/10022-000 [DOI]
- Fisher, W. W. , Greer, B. D. , & Bouxsein, K. J. (2021). Developing function‐based reinforcement procedures for problem behavior. In Fisher W. W., Piazza C. C., & Roane H. S. (Eds.), Handbook of applied behavior analysis, 2 nd Edition (pp. 337–351). Guilford Publishing. [Google Scholar]
- Fisher, W. W. , Greer, B. D. , Fuhrman, A. M. , Saini, V. , & Simmons, C. A. (2018). Minimizing resurgence of destructive behavior using behavioral momentum theory. Journal of Applied Behavior Analysis, 51(4), 831–853. 10.1002/jaba.499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher, W. W. , Greer, B. D. , Mitteer, D. R. , Fuhrman, A. M. , Romani, P. W. , & Zangrillo, A. N. (2018). Further evaluation of differential exposure to establishing operations during functional communication training. Journal of Applied Behavior Analysis, 51(2), 360–373. 10.1002/jaba.451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher, W. W. , & Mazur, J. E. (1997). Basic and applied research on choice responding. Journal of Applied Behavior Analysis, 30(3), 387–410. 10.1901/jaba.1997.30-387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher, W. , Piazza, C. , Cataldo, M. , Harrell, R. , Jefferson, G. , & Conner, R. (1993). Functional communication training with and without extinction and punishment. Journal of Applied Behavior Analysis, 26(1), 23–36. 10.1901/jaba.1993.26-23 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher, W. W. , Saini, V. , Greer, B. D. , Sullivan, W. E. , Roane, H. S. , Fuhrman, A. M. , Craig, A. R. , & Kimball, R. T. (2019). Baseline reinforcement rate and resurgence of destructive behavior. Journal of the Experimental Analysis of Behavior, 111(1), 75–93. 10.1002/jeab.488 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher, W. W. , Thompson, R. H. , DeLeon, I. G. , Piazza, C. C. , Kuhn, D. E. , Rodriguez‐Catter, V. , & Adelinis, J. D. (1999). Noncontingent reinforcement: Effects of satiation versus choice responding. Research in Developmental Disabilities, 20(6), 411–427. 10.1016/S0891-4222(99)00022-0 [DOI] [PubMed] [Google Scholar]
- Frederiksen, L. W. , & Peterson, G. L. (1977). Schedule‐induced aggression in humans and animals: A comparative parametric review. Aggressive Behavior, 3(1), 57–75. 10.1002/1098-2337(1977)3:1<57::AID-AB2480030106>3.0.CO;2-D [DOI] [Google Scholar]
- Greer, B. D. , Fisher, W. W. , Retzlaff, B. J. , & Fuhrman, A. M. (2020). A preliminary evaluation of treatment duration on the resurgence of destructive behavior. Journal of the Experimental Analysis of Behavior, 113(1), 251–262. 10.1002/jeab.567 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer, B. D. , Fisher, W. W. , Saini, V. , Owen, T. M. , & Jones, J. K. (2016). Functional communication training during reinforcement schedule thinning: An analysis of 25 applications. Journal of Applied Behavior Analysis, 49(1), 105–121. 10.1002/jaba.265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer, B. D. , & Shahan, T. A. (2019). Resurgence as Choice: Implications for promoting durable behavior change. Journal of Applied Behavior Analysis, 52(3), 816–846. 10.1002/jaba.573 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagopian, L. P. , Crockett, J. L. , Van Stone, M. , DeLeon, I. G. , & Bowman, L. G. (2000). Effects of noncontingent reinforcement on problem behavior and stimulus engagement: The role of satiation, extinction, and alternative reinforcement. Journal of Applied Behavior Analysis, 33(4), 433–449. 10.1901/jaba.2000.33-433 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagopian, L. P. , Fisher, W. W. , & Legacy, S. M. (1994). Schedule effects of noncontingent reinforcement on attention‐maintained destructive behavior in identical quadruplets. Journal of Applied Behavior Analysis, 27(2), 317–325. 10.1901/jaba.1994.27-317 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haney, S. D. , Greer, B. D. , Randall, K. R. , & Mitteer, D. R. (2022). Relapse during the treatment of pediatric feeding disorders. Journal of Applied Behavior Analysis, 55(3), 704–726. 10.1002/jaba.913 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrnstein, R. J. (1961). Relative and absolute strength of response as a function of frequency of reinforcement. Journal of the Experimental Analysis of Behavior, 4(3), 267–272. 10.1901/jeab.1961.4-267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hyman, S. L. , Fisher, W. , Mercugliano, M. , & Cataldo, M. F. (1990). Children with self‐injurious behavior. Pediatrics, 85(3), 437–441. 10.1542/peds.85.3.437 [DOI] [PubMed] [Google Scholar]
- Ibañez, V. F. , Piazza, C. C. , & Peterson, K. M. (2019). A translational evaluation of renewal of inappropriate mealtime behavior. Journal of Applied Behavior Analysis, 52(4), 1005–1020. 10.1002/jaba.647 [DOI] [PubMed] [Google Scholar]
- Iwata, B. A. , Pace, G. M. , Dorsey, M. F. , Zarcone, J. R. , Vollmer, T. R. , Smith, R. G. , Rodgers, T. A. , Lerman, D. C. , Shore, B. A. , Mazaleski, J. L. , Goh, H. L. , Cowdery, G. E. , Kalsher, M. J. , McCosh, K. C. , & Willis, K. D. (1994). The functions of self‐injurious behavior: An experimental‐epidemiological analysis. Journal of Applied Behavior Analysis, 27(2), 215–240. 10.1901/jaba.1994.27-215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katz, B. , & Lattal, K. A. (2020). An experimental analysis of the extinction‐induced response burst. Journal of the Experimental Analysis of Behavior, 114(1), 24–46. 10.1002/jeab.611 [DOI] [PubMed] [Google Scholar]
- Katz, B. R. , & Lattal, K. A. (2021). What is an extinction burst? A case study in the analysis of transitional behavior. Journal of the Experimental Analysis of Behavior, 115(1), 129–140. 10.1002/jeab.642 [DOI] [PubMed] [Google Scholar]
- Kaufman, A. , & Baron, A. (1968). Suppression of behavior by timeout punishment when suppression results in loss of positive reinforcement. Journal of the Experimental Analysis of Behavior, 11(5), 595–607. 10.1901/jeab.1968.11-595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keller, F. S. , & Schoenfeld, W. N. (1950). Principles of psychology: A systematic text in the science of behavior. Appleton‐Century‐Crofts. 10.1037/11293-000 [DOI] [Google Scholar]
- Lalli, J. S. , Vollmer, T. R. , Progar, P. R. , Wright, C. , Borrero, J. , Daniel, D. , Hoffner Barthold, C. , Tocco, K. , & May, W. (1999). Competition between positive and negative reinforcement in the treatment of escape behavior. Journal of Applied Behavior Analysis, 32(3), 285–296. 10.1901/jaba.1999.32-285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lattal, K. A. (1972). Response–reinforcer independence and conventional extinction after fixed‐interval and variable‐interval schedules. Journal of the Experimental Analysis of Behavior, 18(1), 133–140. 10.1901/jeab.1972.18-133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lattal, K. A. , Kuroda, T. , & Cook, J. E. (2020). Early extinction effects following intermittent reinforcement: Little evidence of extinction bursts. Journal of the Experimental Analysis of Behavior, 114(1), 47–59. 10.1002/jeab.616 [DOI] [PubMed] [Google Scholar]
- Lattal, K. A. , St. Peter, C. , & Escobar, R. (2013). Operant extinction: Elimination and generation of behavior. In Madden G. J., Dube W. V., Hackenberg T. D., Hanley G. P., & Lattal K. A. (Eds.), APA handbook of behavior analysis, Vol. 2. Translating principles into practice (pp. 77–107). American Psychological Association. 10.1037/13938-004 [DOI] [Google Scholar]
- Lerman, D. C. , & Iwata, B. A. (1995). Prevalence of the extinction burst and its attenuation during treatment. Journal of Applied Behavior Analysis, 28(1), 93–94. 10.1901/jaba.1995.28-93 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerman, D. C. , & Iwata, B. A. (1996). Developing a technology for the use of operant extinction in clinical settings: An examination of basic and applied research. Journal of Applied Behavior Analysis, 29(3), 345–382. 10.1901/jaba.1996.29-345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerman, D. C. , Iwata, B. A. , & Wallace, M. D. (1999). Side effects of extinction: Prevalence of bursting and aggression during the treatment of self‐injurious behavior. Journal of Applied Behavior Analysis, 32(1), 1–8. 10.1901/jaba.1999.32-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lomas, J. E. , Fisher, W. W. , & Kelley, M. E. (2010). The effects of variable‐time delivery of food items and praise on problem behavior reinforced by escape. Journal of Applied Behavior Analysis, 43(3), 425–435. 10.1901/jaba.2010.43-425 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Machado, A. (1993). Learning variable and stereotypical sequences of responses: Some data and a new model. Behavioural Processes, 30(2), 103–129. 10.1016/0376-6357(93)90002-9 [DOI] [PubMed] [Google Scholar]
- Marcus, B. A. , & Vollmer, T. R. (1996). Combining noncontingent reinforcement and differential reinforcement schedules as treatment for aberrant behavior. Journal of Applied Behavior Analysis, 29(1), 43–51. 10.1901/jaba.1996.29-43 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McSweeney, F. K. , & Swindell, S. (2002). Common processes may contribute to extinction and habituation. The Journal of General Psychology, 129(4), 364–400. 10.1080/00221300209602103 [DOI] [PubMed] [Google Scholar]
- Miller, N. E. (1941). I. The frustration‐aggression hypothesis. Psychological Review, 48(4), 337–342. 10.1037/h0055861 [DOI] [Google Scholar]
- Mitteer, D. R. , Greer, B. D. , Fisher, W. W. , Briggs, A. M. , & Wacker, D. P. (2018). A laboratory model for evaluating relapse of undesirable caregiver behavior. Journal of the Experimental Analysis of Behavior, 110(2), 252–266. 10.1002/jeab.462 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitteer, D. R. , Greer, B. D. , Randall, K. R. , & Haney, S. D. (2022). On the scope and characteristics of relapse when treating severe destructive behavior. Journal of Applied Behavior Analysis, 55(3), 688–703. 10.1002/jaba.912 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muething, C. , Call, N. , Pavlov, A. , Ringdahl, J. , Gillespie, S. , Clark, S. , & Mevers, J. L. (2020). Prevalence of renewal of problem behavior during context changes. Journal of Applied Behavior Analysis, 53(3), 1485–1493. 10.1002/jaba.672 [DOI] [PubMed] [Google Scholar]
- Nergaard, S. K. , & Holth, P. (2020). A critical review of the support for variability as an operant dimension. Perspectives on Behavior Science, 43(3), 579–603. 10.1007/s40614-020-00262-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neuringer, A. , Kornell, N. , & Olufs, M. (2001). Stability and variability in extinction. Journal of Experimental Psychology: Animal Behavior Processes, 27(1), 79. 10.1037/0097-7403.27.1.79 [DOI] [PubMed] [Google Scholar]
- Nist, A. N. , & Shahan, T. A. (2021). The extinction burst: Impact of reinforcement time and level of analysis on measured prevalence. Journal of the Experimental Analysis of Behavior, 116(2), 131–148. 10.1002/jeab.714 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niyuhire, F. , Varvel, S. A. , Thorpe, A. J. , Stokes, R. J. , Wiley, J. L. , & Lichtman, A. H. (2007). The disruptive effects of the CB1 receptor antagonist rimonabant on extinction learning in mice are task‐specific. Psychopharmacology, 191(2), 223–231. 10.1007/s00213-006-0650-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- North, A. J. (1950). Improvement in successive discrimination reversals. Journal of Comparative and Physiological Psychology, 43(6), 442–460. 10.1037/h0061372 [DOI] [PubMed] [Google Scholar]
- O'Kelly, L. I. , & Steckle, L. C. (1939). A note on long enduring emotional responses in the rat. The Journal of Psychology, 8(1), 125–131. 10.1080/00223980.1939.9917655 [DOI] [Google Scholar]
- Pace, G. M. , Ivancic, M. T. , & Jefferson, G. (1994). Stimulus fading as treatment for obscenity in a brain‐injured adult. Journal of Applied Behavior Analysis, 27(2), 301–305. 10.1901/jaba.1994.27-301 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pear, J. J. , Moody, J. E. , & Persinger, M. A. (1972). Lever attacking by rats during free‐operant avoidance. Journal of the Experimental Analysis of Behavior, 18(3), 517–523. 10.1901/jeab.1972.18-517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podlesnik, C. A. , Jimenez‐Gomez, C. , & Kelley, M. E. (2021). Matching and behavioral momentum: Quantifying choice and persistence. In Fisher W. W., Piazza C. C., & Roane H. S. (Eds.), Handbook of applied behavior analysis, 2 nd Edition (pp. 94–114). Guilford Publishing. [Google Scholar]
- Richman, D. M. , Barnard‐Brak, L. , Grubb, L. , Bosch, A. , & Abby, L. (2015). Meta‐analysis of noncontingent reinforcement effects on problem behavior. Journal of Applied Behavior Analysis, 48(1), 131–152. 10.1002/jaba.189 [DOI] [PubMed] [Google Scholar]
- Ringdahl, J. E. , Vollmer, T. R. , Borrero, J. C. , & Connell, J. E. (2001). Fixed‐time schedule effects as a function of baseline reinforcement rate. Journal of Applied Behavior Analysis, 34(1), 1–15. 10.1901/jaba.2001.34-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rohatgi, A. (2020). WebPlotDigitizer (Version 4.4) [Computer software]. https://automeris.io/WebPlotDigitizer/index.html
- Rohrbaugh, M. , Riccio, D. C. , & Arthur, A. (1972). Paradoxical enhancement of conditioned suppression. Behaviour Research and Therapy, 10(2), 125–130. 10.1016/s0005-7967(72)80005-6 [DOI] [PubMed] [Google Scholar]
- Schramm‐Sapyta, N. L. , Olsen, C. M. , & Winder, D. G. (2006). Cocaine self‐administration reduces excitatory responses in the mouse nucleus accumbens shell. Neuropsychopharmacology, 31(17), 1444–1451. 10.1038/sj.npp.1300918 [DOI] [PubMed] [Google Scholar]
- Shahan, T. A. (2022). A theory of the extinction burst. Perspectives in Behavior Science, 45, 495‐519. 10.1007/s40614-022-00340-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahan, T. A. , Browning, K. O. , & Nall, R. W. (2020). Resurgence as Choice in Context: Treatment duration and on/off alternative reinforcement. Journal of the Experimental Analysis of Behavior, 113(1), 57–76. 10.1002/jeab.563 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahan, T. A. , Browning, K.O. , Nist, A. N. , & Sutton, G. M. (2020), Resurgence and downshifts in alternative reinforcement rate. Journal of the Experimental Analysis of Behavior, 114(2), 163–178. 10.1002/jeab.625 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahan, T. A. , & Craig, A. R. (2017). Resurgence as Choice. Behavioural Processes, 141, 100–127. 10.1016/j.beproc.2016.10.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahan, T. A. , & Greer, B. D. (2021). Destructive behavior increases as a function of reductions in alternative reinforcement during schedule thinning: A retrospective quantitative analysis. Journal of the Experimental Analysis of Behavior, 116(2), 243–248. 10.1002/jeab.708 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidman, M. (1962). Reduction of shock frequency as reinforcement for avoidance behavior. Journal of the Experimental Analysis of Behavior, 5(2), 247–257. 10.1901/jeab.1962.5-247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skinner, B. F. (1938). The behavior of organisms: An experimental analysis. Appleton‐Century. [Google Scholar]
- Terrace, H. S. (1966). Stimulus control. In Honig W. K. (Ed.), Operant behavior: Areas of research and application. Appleton‐Century‐Crofts. [Google Scholar]
- Thackeray, E. J. , & Richdale, A. L. (2002). The behavioural treatment of sleep difficulties in children with an intellectual disability. Behavioral Interventions, 17(4), 211–231. 10.1002/bin.123 [DOI] [Google Scholar]
- Thomas, J. R. (1965). Discriminated time‐out avoidance in pigeons. Journal of the Experimental Analysis of Behavior, 8(5), 329–338. 10.1901/jeab.1965.8-329 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas, B. L. , & Papini, M. R. (2001). Adrenalectomy eliminates the extinction spike in autoshaping with rats. Physiology & Behavior, 72(4), 543–547. 10.1016/S0031-9384(00)00448-0 [DOI] [PubMed] [Google Scholar]
- Thompson, T. , & Bloom, W. (1966). Aggressive behavior and extinction‐induced response‐rate increase. Psychonomic Science, 5(9), 335–336. 10.3758/BF03328426 [DOI] [Google Scholar]
- Tiger, J. H. , & Hanley, G. P. (2004). Developing stimulus control of preschooler mands: An analysis of schedule‐correlated and contingency‐specifying stimuli. Journal of Applied Behavior Analysis, 37(4), 517–521. 10.1901/jaba.2004.37-517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulrich, R. E. , & Azrin, N. H. (1962). Reflexive fighting in response to aversive stimulation. Journal of the Experimental Analysis of Behavior, 5(4), 511–520. 10.1901/jeab.1962.5-511 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vollmer, T. R. , Iwata, B. A. , Zarcone, J. R. , Smith, R. G. , & Mazaleski, J. L. (1993). The role of attention in the treatment of attention‐maintained self‐injurious behavior: Noncontingent reinforcement and differential reinforcement of other behavior. Journal of Applied Behavior Analysis, 26(1), 9–21. 10.1901/jaba.1993.26-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vollmer, T. R. , Ringdahl, J. E. , Roane, H. S. , & Marcus, B. A. (1997). Negative side effects of noncontingent reinforcement. Journal of Applied Behavior Analysis, 30(1), 161–164. 10.1901/jaba.1997.30-161 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wacker, D. P. , Steege, M. W. , Northup, J. , Sasso, G. , Berg, W. , Reimers, T. , Cooper, L. , Cigrand, K. , & Donn, L. (1990). A component analysis of functional communication training across three topographies of severe behavior problems. Journal of Applied Behavior Analysis, 23(4), 417–429. 10.1901/jaba.1990.23-417 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weinsztok, S. C. , & DeLeon, I. G. (2021). The mitigating effects of enhanced reinforcer magnitude and quality on treatment degradation. Journal of Applied Behavior Analysis, 55(2), 547–571. 10.1002/jaba.910 [DOI] [PubMed] [Google Scholar]