

ABSTRACT
Acute respiratory distress syndrome (ARDS) is an acute inflammatory condition with a dramatic increase in incidence since the beginning of the coronavirus disease 19 (COVID‐19) pandemic. Neutrophils play a vital role in the immunopathology of severe acute respiratory syndrome coronavirus 2 (SARS‐CoV‐2) infection by triggering the formation of neutrophil extracellular traps (NETs), producing cytokines including interleukin‐8 (CXCL8), and mediating the recruitment of other immune cells to regulate processes such as acute and chronic inflammation, which can lead to ARDS. CXCL8 is involved in the recruitment, activation, and degranulation of neutrophils, and therefore contributes to inflammation amplification and severity of disease. Furthermore, activation of neutrophils also supports a prothrombotic phenotype, which may explain the development of immunothrombosis observed in COVID‐19 ARDS. This review aims to describe hyperinflammatory ARDS due to SARS‐CoV‐2 infection. In addition, we address the critical role of polymorphonuclear neutrophils, inflammatory cytokines, and the potential targeting of CXCL8 in treating the hyperinflammatory ARDS population.
Keywords: ARDS, COVID‐19, CXCL8, immunomodulators
Neutrophils play a role in the immunopathology SARS‐CoV‐2 infection by triggering the formation of NETs, producing cytokines including interleukin‐8 (CXCL8), and mediating the recruitment of other immune cells to regulate inflammation, which can lead to ARDS. CXCL8 is involved in the recruitment, activation, and degranulation of neutrophils, and therefore contributes to inflammation amplification and severity of disease. Furthermore, activation of neutrophils also supports a prothrombotic phenotype, which may explain the development of immunothrombosis observed in COVID‐19 ARDS.

Introduction
Acute respiratory distress syndrome (ARDS) is an acute inflammatory and life‐threatening respiratory condition with a mortality rate that may exceed 40% [1]. Before the emergence of the coronavirus disease 2019 (COVID‐19) pandemic, caused by severe acute respiratory syndrome coronavirus‐2 (SARS‐CoV‐2), there were an estimated 190,000 cases of ARDS annually in the United States, associated with 74,500 deaths per year [2]. However, in the context of COVID‐19, 29–42% of patients with COVID‐19 have developed ARDS, with a mortality of 15–52% [3]. Common causes of ARDS include pneumonia and nonpulmonary sepsis. In addition, risk factors such as older age may place patients at higher risk for developing ARDS [4, 5]. ARDS is characterized by acute onset of severe hypoxemia, diffuse bilateral pulmonary infiltrates, and extensive pulmonary edema induced by increased vascular permeability, reduced compliance, and protein‐rich fluid in the alveolar space combined with accumulation of activated immune cells such as neutrophils [6].
Neutrophil extracellular trap (NET) formation may lead to compromised gas exchange and profound hypoxemic respiratory failure [7]. NETs are extracellular structures composed of granule, nuclear, and mitochondrial constituents assembled on a scaffold of decondensed chromatin with antimicrobial proteins and peptides [8]. NETs play a crucial role in host defense and pathogens clearance during infection; however, dysregulation of NETs can lead to autoimmune and inflammatory disorders [9]. The formation of NETs occur through various molecular mechanisms related to neutrophil death. This process is mainly stimulated by the induction of mitogen phorbol 12‐myristate 13‐acetate (PMA), which results in the activation of PKC, leading to reactive oxygen species (ROS) production through the activation of different enzymes such as NAD phosphate (NADPH), oxidase, myeloperoxidase (MPO), and through the release of neutrophil elastase (NE) from granules into the cytoplasm. NE then migrates to the nucleus of neutrophils where it leads to nuclear membrane disintegration. Moreover, various proteins have also been found to prime neutrophils leading to the release of NETs [10, 11, 12].
Recent attempts to define precision management strategies for the heterogeneous population of patients with ARDS have emphasized defining hyper‐inflammatory versus hypo‐inflammatory phenotypes of ARDS [13]. The hyperinflammatory phenotype is characterized by higher plasma levels of inflammatory biomarkers and is more common in patients with sepsis or pneumonia and on vasopressors. Because of the high concentrations of cytokines and chemokines present in patients with severe COVID‐19, they are likely to be classified as having a hyper‐inflammatory phenotype [14]. This review focuses on hyperinflammatory ARDS, particularly due to SARS‐CoV‐2 infection, and addresses the critical role of the polymorphonuclear neutrophils and the inflammatory cytokine storm caused by an accumulation of high plasma levels of inflammatory cytokines [15].
Role of neutrophils in cytokine storm in COVID‐19 ARDS
Patients with COVID‐19 ARDS tend to have higher levels of neutrophils in the plasma and BALF, which have been correlated with poor outcomes and illness severity [16, 17]. Neutrophil infiltration of the lungs is a hallmark of ARDS [18]. Activated neutrophils trigger oxidative stress, release proteases, and form NETs, resulting in lung damage [19]. Infiltration of neutrophils at the site of infection, and their degranulation and release of NETs in response to microbial stimuli, produces an increase of cytokines and chemokines that might result in cytokine storm and contribute to ARDS [20, 21]. The unregulated excess of cytokines further causes lung inflammation resulting in diffuse alveolar damage, septic shock, and multiple organ dysfunction [22]. Neutrophils are pivotal effector cells in the human innate immune defense against infections that migrate to infected tissues in multiple ways including rolling, adhesion, crawling, and transmigration [23]. They are known to have protective roles against intracellular pathogens such as viruses and mycobacteria. But, they are also involved in the immunopathology of SARS‐CoV‐2 infection by triggering NETosis, the process of NET formation, to result in severe tissue damage and immunological and inflammatory processes (Figure 1) [24]. Based on these findings, targeting neutrophils could represent a new therapeutic strategy in acute lung injury and inflammation.
Figure 1.
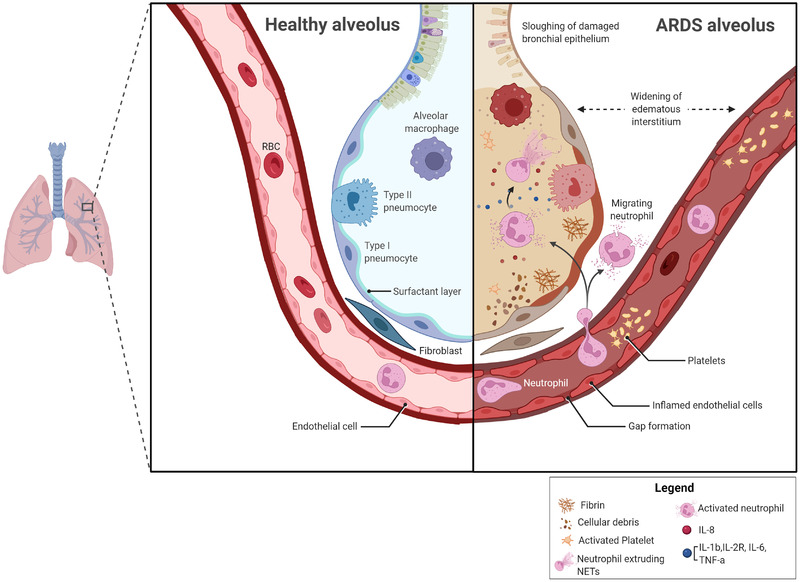
Infiltration of neutrophil in the lung contribute to ARDS. In ARDS, activated alveolar macrophages recruit neutrophils, and induce the secretion of IL‐8, into the lungs; this process leads to alveolar pathology associated with the development and severity of the condition. Excessive accumulation of neutrophils in the alveolar space may trigger release of reactive oxygen species, and neutrophil extracellular traps (NETs) which can result in epithelial injury. An excess of inflammatory cytokines further causes immune cell infiltration into inflamed lungs resulting in the pathologic hallmark of diffuse alveolar damage. ARDS, Acute respiratory distress syndrome; NETs, neutrophil extracellular traps; CXCL8, Interleukin 8; IL‐1b, interleukin 1 beta; IL‐2R, Interleukin 2 receptor; IL‐6, interleukin 6; TNF‐α, tumor necrosis factor alfa.
Effectors of neutrophil activation in COVID‐19 ARDS
Inappropriate activation of neutrophils can lead to tissue or organ injury, thrombus formation, vascular leakage or necrosis that may lead to the development of a wide range of diseases, including ARDS and COVID‐19 ARDS [25, 26]. Specifically, this inappropriate hyperactivation is caused by a cascade of cellular processes (oxidation, degranulation, NETosis and/or cytokine overproduction) and it is characterized by the production of various neutrophils effectors, ultimately leading to host tissue/organ damage. The most commonly reported effectors are ROS, granular enzymes such as MPO, NE, cathepsin G (CatG), NETs and various pro‐inflammatory cytokines (Figure 2) [9, 27, 28]. Additionally, increasing evidence shows that oxidative stress plays an essential role in endothelial dysfunction and can lead to coagulation, thrombosis, and atherosclerosis [29, 30]. Patients with COVID‐19 ARDS have damage to pulmonary microvascular endothelial cells due to higher levels of ROS that contribute to pulmonary vascular injury, permeability defects, and thrombotic complications [30]. In support of this hypothesis, ROS produced by the NADPH oxidase complex (Nox2) has been associated with thrombotic events in COVID‐19 patients [31].
Figure 2.
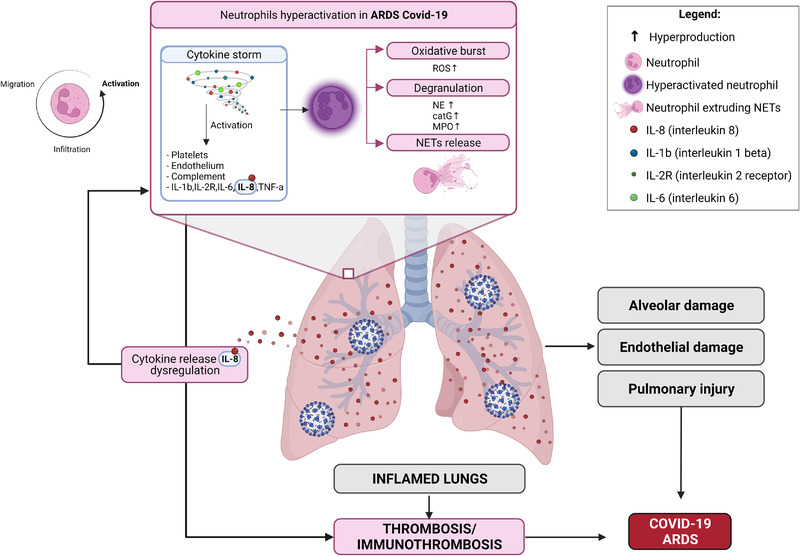
Neutrophil hyperactivation in ARDS COVID‐19. Neutrophil hyperactivation causes an enhancement of cellular processes including oxidation, degranulation, NETosis and increases in the production of various neutrophils effectors, ultimately leading to host tissue/organ damage. The most commonly reported effectors are ROS, MPO, NE, CatG, NETs and various pro‐inflammatory cytokines. IL‐1b, interleukin 1beta; IL‐2R, Interleukin 2 receptor; IL‐6, interleukin 6; CXCL8, Interleukin 8; TNF‐a, tumor necrosis factor alfa; ROS, reactive oxygen species; NE, neutrophil elastase; CatG, cathepsin G; MPO, myeloperoxidase; NETs, neutrophil extracellular traps; ARDS, Acute respiratory distress syndrome
Increased serum levels of NE, a proteolytic enzyme, in patients with COVID‐19 and fatal ARDS have been reported, suggesting a key role of neutrophil serine proteinases (NSPs) in COVID‐19 pneumonia‐driven ARDS [32]. This observation deserves further investigation to define the role of NSPs, and to evaluate if NSPs’ elimination, by using cathepsin C inhibitors which block NSP maturation, could be a potential therapeutic strategy to prevent the pulmonary damage in patients with COVID‐19 [33]. Myeloperoxidase, the principal enzyme of peroxidases that catalyzes H2O2 to produce toxic ROS, was demonstrated to be a local mediator of alveolar damage [34]. COVID‐19 disease severity is correlated with MPO‐DNA complexes as a measure of NETs in plasma and tracheal aspirates of patients with COVID‐19 [35]. Exaggerated NET formation and high concentrations of NET markers have been observed in hospitalized patients, including circulating free DNA (cfDNA), DNA‐MPO and DNA‐NE complexes [36, 37].
Several neutrophil subpopulations exert different biological functions [38]. Among them, high density neutrophils (HDNs) are abundantly represented both in physiological and pathological settings. Whereas, low‐density neutrophils (LDNs), an immature neutrophil population, and low‐density granulocytes (LDGs), a distinct set of pro‐inflammatory granulocytes, differ from LDNs in surface marker expression and are mainly associated with pathological conditions [39, 40]. LDN subpopulations that expresses intermediate levels of CD16 (CD16Int) have been identified in blood samples of patients with severe COVID‐19, and exhibit a pro‐inflammatory phenotype with phagocytic capacity, NET formation, and interactions with platelets leading to a hypercoagulable state [41]. Additionally, a significant increase of the LDG subpopulation, the generally less characterized neutrophil subpopulation in viral infections, has been observed in the blood of patients with COVID‐19, thus demonstrating an increase in neutrophil recruitment and activation of this disease [42]. These findings support the hypothesis that hyperactivation of specific neutrophilic subpopulations is an essential feature of inflammatory progression and pathogenesis of COVID‐19 ARDS, and that neutrophil effectors produced by the over‐activated process can consequently lead to tissue/organ damage and disease progression.
Role of NETs in COVID‐19 ARDS
The neutrophilic infiltration into lung tissue in COVID‐19 ARDS is associated with NETs release and surrogate markers of NETs, such as DNA‐MPO and DNA‐citrullinated histone‐3 complexes, linked with disease severity [36]. Further evidence suggests that SARS‐CoV‐2 can directly stimulate human neutrophils to release NETs, through the stimulation of angiotensin‐converting enzyme 2 (ACE2) and transmembrane serine protease 2 (TRPMSS2) axis [43]. ACE2 is a key mediator of viral entry into the host cells in SARS‐CoV‐2 infection due to its interaction with the glycoprotein Spike (S) and its expression in several host cells, including lung pneumocytes, epithelial cells, and endothelial cells, and it is crucial in release of NETs by neutrophils [43]. ACE2‐mediated SARS‐CoV‐2 cell entry causes a reduction of ACE2 levels on the cell surface, leading to worse disease severity from neutrophil infiltration, vascular permeability, and lung edema [44].
NETs cause endothelial damage and necroinflammation via complement activation linked to the release of thrombogenic NETs combined with Tissue Factor (TF) which may lead to multiorgan failure and death [45, 46]. In parallel, NETs have been proposed to promote thrombus formation during COVID‐19 that leads to multiple organ system dysfunction [47]. Severe disease is associated with markers of increased coagulopathy, thus suggesting a potential interplay between the coagulation system and NET formation [48]. Occlusion of small pulmonary vessels, by aggregated NETs, have been observed in lungs of patients who died from COVID‐19 [35]. Interestingly, the recent use of a combination of high‐dimensional single‐cell analysis and ex vivo functional assays of neutrophils from patients with COVID‐19 ARDS, compared with patients with non‐COVID ARDS, was used to identify a distinct landscape of neutrophil activation in COVID‐19 ARDS that was intrinsically programmed during SARS‐CoV‐2 infection [49]. Thus, demonstrating that neutrophils in COVID‐19 ARDS are functionally primed to produce high amounts of NETs. This molecular mechanism of neutrophil priming escapes conventional therapy with corticosteroids, like dexamethasone, paving the way to consider neutrophil priming in COVID‐19 as a promising new target for adjunctive treatments against severe COVID‐19.
The concept of activated neutrophils that produce various cytotoxic products, NETS, and pro‐inflammatory cytokines (IL‐1β, IL‐6, IL‐8, TNF‐α, MCP‐1, and GM‐CSF) leading to enhanced inflammation and tissue damage and lung dysfunction is established in ARDS and COVID‐19 ARDS [50, 51, 52]. A prospective study investigated the presence of NETs in blood and lung samples of patients with critical COVID‐19, and demonstrated that plasma NET levels peaked early after intensive care unit (ICU) admission and correlated with plasma levels of chemokines and inflammatory markers [53]. NET quantity correlated with disease severity and infection duration, with continued detection of NETs in bronchial and alveolar spaces [37]. All these findings support the hypothesis of the crucial role of NETs in severe COVID‐19 pneumonia and ARDS, and their potential contribution to the pathologic changes observed in severe cases.
Neutrophil role in immunothrombosis in COVID‐19 ARDS
COVID‐19 acutely causes lung injury, primarily affecting the vascular endothelium, as evident by the finding of endothelitis and capillaritis in patients with diffuse severe lung alveolar damage and ARDS [54]. Venous thromboembolism (VTE) has been reported to occur in up to one‐third of COVID‐19 cases and is associated with severe disease, worse clinical outcomes, and an increase in mortality [55, 56]. Higher VTE rates in patients with severe COVID‐19, compared to matched cohorts with other forms of ARDS, suggest that additional mechanisms beyond typical risk factors of hospitalized critically ill patients (e.g. immobility and severe illness) exist [57]. Microvascular coagulopathy and immunothrombosis observed in patients with severe COVID‐19 are partially responsible for the hypercoagulation consequences from severe COVID‐19 (Figure 3) [58]. This cascade can be activated by SARS‐CoV‐2 itself, but also by hypoxemia, activated complement and pro‐inflammatory cytokines (such as IL‐1β and IL‐6) released as part of the cytokine storm that can trigger endothelial cells injury [59, 60, 61, 62].
Figure 3.

Microvascular coagulopathy and immunothrombosis processes in patients with severe COVID‐19. A dysregulated inflammatory condition contributes to a prothrombotic phenotype characterized by activation of platelets, endothelial dysfunction and complement activation. This inflammatory induced coagulopathy can result in excessive formation of immunologically mediated thrombi primarily in the microvasculature, a process known as immunothrombosis. The microvascular coagulopathy and immunothrombosis observed in patients with severe COVID‐19 may be responsible for severe multiple organ dysfunction like ARDS. IL‐1b, interleukin 1β; IL‐2, Interleukin 2; IL‐6, interleukin 6; CXCL8, Interleukin 8; TNF‐a, tumor necrosis factor α; INF – γ, interferon gamma; NETs, neutrophil extracellular traps; ARDS, Acute respiratory distress syndrome.
Neutrophil activation induced by thrombin and factor Xa, through fibrinogen, fibrin, and C5a, can lead to increased production of IL‐6 and CXCL8, to result in a strong procoagulant response and neutrophil‐derived TF and NETs formation [63, 64]. As previously reported, in severe COVID‐19, high‐dimensional flow cytometric analysis of circulating neutrophils revealed a highly activated phenotype, LDN, that is more prone to spontaneously forming NETs and is associated with microthrombosis and organ damage [35, 37, 65]. NET‐driven thrombosis is largely platelet‐dependent, and have been identified as a major contributor to neutrophil‐related thrombo‐inflammation. Von Willebrand Factor (VWF), released by endothelial cells and platelets, can result in increased platelet adhesion and fibrin formation, and can contribute to immunothrombosis. Histone proteins, present in the DNA fragments of NETs, are potent Damage‐Associated Molecular Patterns (DAMP) molecules also capable of initiating a positive inflammatory feedback loop [66, 67, 68].
Several cell types of the immune system can contribute to cytokine storm, including polymorphonuclear neutrophils (PMN), through NET formation [69]. Cytokine storm, through the release of proinflammatory cytokines and chemokines can exaggerate the immunothrombotic and coagulation response to create immune thrombi formation [70, 71]. Both IL‐6 and IFN‐γ increase platelet production and activity, while TF expression on endothelial cells and monocytes increases endothelial dysfunction augmenting coagulopathy. In addition, IL‐2 can further promote coagulation by decreasing fibrinolysis as a consequence of plasminogen activator inhibitor‐1 (PAI‐1) release [72]. CXCL8 has a key role in the production of the prothrombotic phenotype by attracting neutrophils to the site of infection and causing NETs formation [63]. Dysregulated signaling in the CXCL8/CXCR1/2 axis may initiate and perpetuate a self‐sustaining loop leading to an activated, prothrombotic neutrophil phenotype, characterized by intense degranulation and NET formation [73, 74]. Serum and plasma from patients with COVID‐19 demonstrated NET release and increased adhesion to activated platelets from healthy neutrophils. [65, 75]
Petito et al. found thrombotic complications in 22.2% of 36 patients with COVID‐19 and suggested a crucial role of neutrophil activation as compared to platelet activation, because NET formation was the biomarker most associated with increased thrombosis and overall disease severity. Thrombotic complications were observed more often in severe disease, including patients admitted to the ICU and requiring mechanical ventilation, as compared to patients with mild COVID‐19 [76]. Thus, this suggests that NET formation is a possible biomarker to predict disease severity and the potential for thrombotic complications [76].
Neutrophils and NETs have been previously implicated in the pathophysiology of thrombosis in VTE, as well as ARDS and sepsis [77, 78]. In this context, ARDS is defined as a “NETopathy” in which higher levels of NETs in the plasma and BALF are a characteristic feature of transfusion‐associated and pneumonia‐related ARDS [79, 80, 81]. There remains an unmet need in the context of immunothrombosis in COVID‐19 and ARDS‐related syndrome. Future and ongoing clinical trials may help to identify agents with “immunothrombosis targeted interventions” that modulate the immune response and the cytokine cascade.
Targeting neutrophils and NETosis in COVID‐19
There are several investigational anti‐inflammatory strategies under evaluation as potential immunological therapies for hyperinflammatory COVID‐19 ARDS (Table 1). Corticosteroids are currently recommended in severe COVID‐19 patients due to their ability to inhibit cytokine release by blocking the NF‐κB pathway. Other commonly utilized agents include anti‐IL‐6 monoclonal antibodies, tocilizumab or sarilumab, and agents targeting the JAK‐STAT pathway, such as baricitinib. A recent prospective meta‐analysis of 10,930 patients hospitalized for COVID‐19, demonstrated that IL‐6 antagonists were associated with lower 28‐day all‐cause mortality [82]. Still, many current immunomodulators have unknown benefits in early cytokine release syndrome.
Table 1.
Targeted Immunological Therapies for COVID‐19 ARDS
| Medication | Mechanism | PHASE | Patient Population | NCT |
|---|---|---|---|---|
| Lenzilumab | Inhibitory GMCSF mAb | III | Severe and Critical COVID‐19 Pneumonia | NCT04351152 |
| Mavrilimumab | Inhibitory GMCSF mAb | II/III | Severe COVID‐19 Pneumonia | NCT04447469 |
| Otilimab | Inhibitory GMCSF mAb | II | Severe COVID‐19 Pneumonia | NCT04376684 |
| Anakinra | IL‐1 receptor antagonist | III | Severe COVID‐19 Pneumonia | NCT04680949 |
| Canakinumab | Inhibitory IL‐1 mAb | III | Severe COVID‐19 Pneumonia | NCT04362813 |
| Sarilumab | Inhibitory IL‐6 receptor mAb | II | Moderate and Severe COVID‐19 Pneumonia | NCT04357808 |
| Tociluzumab | Inhibitory IL‐6 receptor mAb | III | Severe COVID‐19 Pneumonia | NCT04320615 |
| Siltuximab | Inhibitory IL‐6 mAb | III | Acute Respiratory Failure by COVID‐19 Pneumonia | NCT04330638 |
| Reparixin | IL‐8–CXCR1/2 pathway inhibitors | III | Severe COVID‐19 Pneumonia | NCT04878055 |
| Dornase alfa | Aerosolized DNAse | III | Acute Respiratory Failure by COVID‐19 Pneumonia | NCT04402970 |
| DNase I | II | Acute Respiratory Failure by COVID‐19 Pneumonia | NCT04541979 | |
| rh‐DNase | II | Acute Respiratory Failure by COVID‐19 Pneumonia | NCT04445285 | |
| Mesenchymal stem cell therapy | I/IIa | Acute Respiratory Failure by COVID‐19 Pneumonia | NCT04524962 |
https://clinicaltrials.gov/ct2/home last access 03.03.2022
Recognizing the important role of neutrophils in severe COVID‐19 has led to the investigation of novel potential therapeutic strategies targeting neutrophil recruitment and/or activation. CXCL8, a key neutrophil chemotactic factor and its binding to the cognate CXC receptor types 1 and 2 (CXCR1/2), mediates neutrophil recruitment, activation, and NETs release to potentiate inflammation and multiple organ system dysfunction in severe COVID‐19 [83, 84]. Elevated CXCL8 levels have been found in patients at risk for ARDS, and found useful as a prognostic biomarker in patients with severe COVID‐19 and ARDS [85]. In the setting of COVID‐19, CXCL8 has been significantly associated with time until death and time from hospital admission until death [86]. In a study of 24 critically ill adults, CXCL8 was shown to be significantly correlated with 30‐day mortality (r = 0.50), respiratory failure (r = ‐0.68), acute kidney injury (r = 0.73), and elevated CRP levels [87]. Li et al. demonstrated a correlation of CXCL8 levels and survival after 14 days from their ICU admission [88]. It was hypothesized that IL‐6 levels spike first and promote the recruitment of CXCL8 later, resulting in the continuous cytokine cascade [88]. Elevated admission CXCL8 plasma levels (>30 pg/mL) have been shown to have a sensitivity of 54.90% and specificity of 90.26% in predicting mortality in patients with COVID‐19, or can be utilized to predict the length of hospital stay until death [89]. This data lends further support that CXCL8 may be a prognostic biomarker for hospital mortality along with other poor outcomes, and supports new clinical trials targeting CXCL8 as strategy to treat severe COVID‐19.
Several CXCL8 inhibitors have now entered clinical evaluation in patients with severe COVID‐19 pneumonia (Table 1). Reparixin, a CXCR1/2 allosteric inhibitor has completed a multicenter, randomized, adaptive phase 2/3 clinical study (NCT04794803). The phase 2 portion demonstrated positive results in patients with severe COVID‐19 pneumonia who received oral reparixin and had a lower incidence of the primary composite clinical outcome relative to those receiving standard of care [90]. A larger phase 3 clinical study is ongoing to confirm the role of reparixin in limiting disease progression in patients with severe COVID‐19 (NCT05254990). Another pro‐inflammatory cytokine, granulocyte‐macrophage colony stimulating factor (GM‐CSF), plays a major role in neutrophil differentiation and activation leading to downstream inflammation caused by cytokines, and activation of myeloid cells. LIVE‐AIR, a phase 3 randomized, double‐blind, placebo‐controlled trial, demonstrated a survival benefit due to lenzilumab, an anti‐GM‐CSF monoclonal antibody, in patients without invasive mechanical ventilation and with early‐stage COVID‐19 disease [91]. IL‐1β is a major inducer of CXCL8 expression and release, as well as other neutrophil recruiting chemotactic factors. The SAVE‐MORE trial investigated anakinra, a recombinant human IL‐1 receptor antagonist, in a Phase 3 double‐blind, randomized, controlled trial in 594 patients with moderate to severe COVID‐19 and demonstrated a significant improvement in clinical status (World Health Organization Clinical Progression Scale) and mortality when compared to placebo [92]. However, other conflicting studies have demonstrated that blockade of IL‐1 has no effect in clinical improvement in patients with COVID‐19 [93].
The role of NETs in the development of ARDS complications has prompted the investigation of the therapeutic potential of agents designed to reduce NETs formation or accumulation. Protein arginine deiminase 4 (PAD‐4) inhibitors, such as CL‐Amidine, are being evaluated in COVID‐19 due to their potential role in thrombotic complications and ability to attenuate NET formation [94]. ROS may lead to activation of PAD‐4, an enzyme involved in histone citrullination, leading to decondensed chromatin, “net‐like” occlusions, and thrombus formation [94]. Further modulation of NETosis is being investigated by administration of DNase agents to suppress cytokine levels in patients with COVID‐19 by targeting cell‐free DNA from NETs [95]. Recombinant human DNase, is being investigated in patients presenting with COVID‐19 to reduce sputum NETs and improve oxygenation [96, 97]. Sivelestat, a NE inhibitor, has been shown to improve pulmonary function and ventilator free days in patients with acute lung injury by inhibiting neutrophil chemotaxis and downregulating the NF‐kB pathway [98, 99]. Sivelestat may also be beneficial in preventing the development of disseminated intravascular coagulation (DIC), a coagulopathic complication contributed to excess NE, as it has shown to reduce DIC scores [100].
Conclusions
The COVID‐19 pandemic has resulted in a dramatically greater number of patients with ARDS over the past several years. The complex relationship of neutrophils, the coagulation system, NETosis, and the hyperimmune response has provided clinical challenges for its management. This composite pattern supports the evaluation of potential therapeutic agents targeting the dysregulated neutrophil response in SARS‐CoV‐2 infection to mitigate disease complications. The observation that elevated levels of CXCL8 are associated with poor prognosis in severe COVID‐19 suggests a potential role in targeting CXCL8 in the future to impact hyperinflammation and improve patient outcomes.
Conflicts of interest
R.B. and A.K. are consultants for Dompé U.S. Inc. E.M.G., M.C.C., M.Z., C.M., F.M., and M.A. are full‐time employees of Dompé farmaceutici S.p.A.
Author contributions
All authors contributed equally in editing and proofreading of the manuscript. M.C.C., C.M., E.G., and M.Z. wrote the manuscript and G.C. prepared the figures. R.B., A.K., M.A., and F.M. provided critical analysis and overall guidance of the science.
Acknowledgments
This work was supported by Dompé farmaceutici S.p.A. All figures have been created with BioRender.com.
Data availability statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
References
- 1. Pham, T. and Rubenfeld, G.D. , Fifty years of research in ARDS. The epidemiology of acute respiratory distress syndrome. A 50th birthday review. Am J Respir Crit Care Med. 2017. 195(7): 860‐70. [DOI] [PubMed] [Google Scholar]
- 2. Hendrickson, K.W. , Peltan, I.D. and Brown, S.M. , The Epidemiology of acute respiratory distress syndrome before and after coronavirus disease 2019. Crit Care Clin. 2021. 37(4): 703‐16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Peltan, I.D. , Caldwell, E. , Admon, A.J. , Attia, E.F. , Gundel, S.J. , Mathews, K.S. , et al., Characteristics and outcomes of US patients hospitalized With COVID‐19. Am J Crit Care. 2022. 31(2): 146‐57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Meyer, N.J. , Gattinoni, L. and Calfee, C.S. , Acute respiratory distress syndrome. Lancet. 2021. 398(10300): 622‐37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Johnston, C.J. , Rubenfeld, G.D. and Hudson, L.D. , Effect of age on the development of ARDS in trauma patients. Chest. 2003. 124(2): 653‐9. [DOI] [PubMed] [Google Scholar]
- 6. Force, A.D.T. , Ranieri, V.M. , Rubenfeld, G.D. , Thompson, B.T. , Ferguson, N.D. , Caldwell, E. , et al., Acute respiratory distress syndrome: the Berlin Definition. JAMA. 2012. 307(23): 2526‐33. [DOI] [PubMed] [Google Scholar]
- 7. Bourenne, J. , Carvelli, J. and Papazian, L. , Evolving definition of acute respiratory distress syndrome. J Thorac Dis. 2019. 11(Suppl 3): S390‐S3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Brinkmann, V. , Reichard, U. , Goosmann, C. , Fauler, B. , Uhlemann, Y. , Weiss, D.S. , et al., Neutrophil extracellular traps kill bacteria. Science. 2004. 303(5663): 1532‐5. [DOI] [PubMed] [Google Scholar]
- 9. Kaplan, M.J. and Radic, M. , Neutrophil extracellular traps: double‐edged swords of innate immunity. J Immunol. 2012. 189(6): 2689‐95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Pruchniak, M.P. and Demkow, U. , Potent NETosis inducers do not show synergistic effects in vitro. Cent Eur J Immunol. 2019. 44(1): 51‐8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Kirchner, T. , Moller, S. , Klinger, M. , Solbach, W. , Laskay, T. and Behnen, M. , The impact of various reactive oxygen species on the formation of neutrophil extracellular traps. Mediators Inflamm. 2012. 2012:849136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Papayannopoulos, V. , Metzler, K.D. , Hakkim, A. and Zychlinsky, A. , Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J Cell Biol. 2010. 191(3): 677‐91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Calfee, C.S. , Delucchi, K. , Parsons, P.E. , Thompson, B.T. , Ware, L.B. , Matthay, M.A. , et al., Subphenotypes in acute respiratory distress syndrome: latent class analysis of data from two randomised controlled trials. Lancet Respir Med. 2014. 2(8): 611‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Bos, L.D.J. , Artigas, A. , Constantin, J.M. , Hagens, L.A. , Heijnen, N. , Laffey, J.G. , et al., Precision medicine in acute respiratory distress syndrome: workshop report and recommendations for future research. Eur Respir Rev. 2021. 30(159). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Ruan, Q. , Yang, K. , Wang, W. , Jiang, L. and Song, J. , Clinical predictors of mortality due to COVID‐19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Med. 2020. 46(5): 846‐8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Voiriot, G. , Dorgham, K. , Bachelot, G. , Fajac, A. , Morand‐Joubert, L. , Parizot, C. , et al., Identification of bronchoalveolar and blood immune‐inflammatory biomarker signature associated with poor 28‐day outcome in critically ill COVID‐19 patients. Scientific reports. 2022. 12(1): 9502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Dentone, C. , Vena, A. , Loconte, M. , Grillo, F. , Brunetti, I. , Barisione, E. , et al., Bronchoalveolar lavage fluid characteristics and outcomes of invasively mechanically ventilated patients with COVID‐19 pneumonia in Genoa, Italy. BMC Infect Dis. 2021. 21(1): 353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Zemans, R.L. and Matthay, M.A. , What drives neutrophils to the alveoli in ARDS? Thorax. 2017. 72(1): 1‐3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Zhu, Y. , Chen, X. and Liu, X. , NETosis and Neutrophil Extracellular Traps in COVID‐19: Immunothrombosis and Beyond. Frontiers in Immunology. 2022. 13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Shah, R.D. and Wunderink, R.G. , Viral Pneumonia and Acute Respiratory Distress Syndrome. Clin Chest Med. 2017. 38(1): 113‐25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Eworuke, E. , Major, J.M. and Gilbert McClain, L.I. National incidence rates for Acute Respiratory Distress Syndrome (ARDS) and ARDS cause‐specific factors in the United States (2006‐2014). J Crit Care. 2018. 47:192‐7. [DOI] [PubMed] [Google Scholar]
- 22. McElvaney, O.J. , McEvoy, N.L. , McElvaney, O.F. , Carroll, T.P. , Murphy, M.P. , Dunlea, D.M. , et al., Characterization of the Inflammatory Response to Severe COVID‐19 Illness. Am J Respir Crit Care Med. 2020. 202(6): 812‐21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Niedzwiedzka‐Rystwej, P. , Grywalska, E. , Hrynkiewicz, R. , Bebnowska, D. , Wolacewicz, M. , Majchrzak, A. , et al., Interplay between Neutrophils, NETs and T‐Cells in SARS‐CoV‐2 Infection‐A Missing Piece of the Puzzle in the COVID‐19 Pathogenesis? Cells. 2021. 10(7). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Muraro, S.P. , De Souza, G.F. , Gallo, S.W. , Da Silva, B.K. , De Oliveira, S.D. , Vinolo, M.A.R. , et al., Respiratory Syncytial Virus induces the classical ROS‐dependent NETosis through PAD‐4 and necroptosis pathways activation. Sci Rep. 2018. 8(1): 14166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Kruger, P. , Saffarzadeh, M. , Weber, A.N. , Rieber, N. , Radsak, M. , von Bernuth, H. , et al., Neutrophils: Between host defence, immune modulation, and tissue injury. PLoS Pathog. 2015. 11(3): e1004651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Reyes, L. , AS‐G, M. , Morrison, T. , Howden, A.J.M. , Watts, E.R. , Arienti, S. , et al., A type I IFN, prothrombotic hyperinflammatory neutrophil signature is distinct for COVID‐19 ARDS. Wellcome Open Res. 2021. 6:38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Goud, P.T. , Bai, D. and Abu‐Soud, H.M. , A Multiple‐Hit Hypothesis Involving Reactive Oxygen Species and Myeloperoxidase Explains Clinical Deterioration and Fatality in COVID‐19. Int J Biol Sci. 2021. 17(1): 62‐72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Tamassia, N. , Bianchetto‐Aguilera, F. , Arruda‐Silva, F. , Gardiman, E. , Gasperini, S. , Calzetti, F. , et al., Cytokine production by human neutrophils: Revisiting the “dark side of the moon”. Eur J Clin Invest. 2018. 48 Suppl 2:e12952. [DOI] [PubMed] [Google Scholar]
- 29. Jin, Y. , Ji, W. , Yang, H. , Chen, S. , Zhang, W. and Duan, G. , Endothelial activation and dysfunction in COVID‐19: from basic mechanisms to potential therapeutic approaches. Signal Transduct Target Ther. 2020. 5(1): 293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Cekerevac, I. , Turnic, T.N. , Draginic, N. , Andjic, M. , Zivkovic, V. , Simovic, S. , et al., Predicting Severity and Intrahospital Mortality in COVID‐19: The Place and Role of Oxidative Stress. Oxid Med Cell Longev. 2021. 2021:6615787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. DiNicolantonio, J.J. and McCarty, M. , Thrombotic complications of COVID‐19 may reflect an upregulation of endothelial tissue factor expression that is contingent on activation of endosomal NADPH oxidase. Open Heart. 2020. 7(1). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Zerimech, F. , Jourdain, M. , Onraed, B. , Bouchecareilh, M. , Sendid, B. , Duhamel, A. , et al., Protease‐antiprotease imbalance in patients with severe COVID‐19. Clin Chem Lab Med. 2021. 59(8): e330‐e4. [DOI] [PubMed] [Google Scholar]
- 33. Korkmaz, B. , Lesner, A. , Marchand‐Adam, S. , Moss, C. and Jenne, D.E. , Lung Protection by Cathepsin C Inhibition: A New Hope for COVID‐19 and ARDS? J Med Chem. 2020. 63(22): 13258‐65. [DOI] [PubMed] [Google Scholar]
- 34. Aratani, Y. , Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch Biochem Biophys. 2018. 640:47‐52. [DOI] [PubMed] [Google Scholar]
- 35. Middleton, E.A. , He, X.Y. , Denorme, F. , Campbell, R.A. , Ng, D. , Salvatore, S.P. , et al., Neutrophil extracellular traps contribute to immunothrombosis in COVID‐19 acute respiratory distress syndrome. Blood. 2020. 136(10): 1169‐79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Zuo, Y. , Yalavarthi, S. , Shi, H. , Gockman, K. , Zuo, M. , Madison, J.A. , et al., Neutrophil extracellular traps in COVID‐19. JCI Insight. 2020. 5(11). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Leppkes, M. , Knopf, J. , Naschberger, E. , Lindemann, A. , Singh, J. , Herrmann, I. , et al., Vascular occlusion by neutrophil extracellular traps in COVID‐19. EBioMedicine. 2020. 58:102925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Silvestre‐Roig, C. , Fridlender, Z.G. , Glogauer, M. and Scapini, P. , Neutrophil Diversity in Health and Disease. Trends Immunol. 2019. 40(7): 565‐83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Villanueva, E. , Yalavarthi, S. , Berthier, C.C. , Hodgin, J.B. , Khandpur, R. , Lin, A.M. , et al., Netting neutrophils induce endothelial damage, infiltrate tissues, and expose immunostimulatory molecules in systemic lupus erythematosus. J Immunol. 2011. 187(1): 538‐52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Denny, M.F. , Yalavarthi, S. , Zhao, W. , Thacker, S.G. , Anderson, M. , Sandy, A.R. , et al., A distinct subset of proinflammatory neutrophils isolated from patients with systemic lupus erythematosus induces vascular damage and synthesizes type I IFNs. J Immunol. 2010. 184(6): 3284‐97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Morrissey, S.M. , Geller, A.E. , Hu, X. , Tieri, D. , Ding, C. , Klaes, C.K. , et al., A specific low‐density neutrophil population correlates with hypercoagulation and disease severity in hospitalized COVID‐19 patients. JCI Insight. 2021. 6(9). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Cabrera, L.E. , Pekkarinen, P.T. , Alander, M. , Nowlan, K.H.A. , Nguyen, N.A. , Jokiranta, S. , et al., Characterization of low‐density granulocytes in COVID‐19. PLoS Pathog. 2021. 17(7): e1009721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Veras, F.P. , Pontelli, M.C. , Silva, C.M. , Toller‐Kawahisa, J.E. , de Lima, M. , Nascimento, D.C. , et al., SARS‐CoV‐2‐triggered neutrophil extracellular traps mediate COVID‐19 pathology. J Exp Med. 2020. 217(12). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Eguchi, S. , Kawai, T. , Scalia, R. and Rizzo, V. , Understanding Angiotensin II Type 1 Receptor Signaling in Vascular Pathophysiology. Hypertension. 2018. 71(5): 804‐10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Skendros, P. , Mitsios, A. , Chrysanthopoulou, A. , Mastellos, D.C. , Metallidis, S. , Rafailidis, P. , et al., Complement and tissue factor‐enriched neutrophil extracellular traps are key drivers in COVID‐19 immunothrombosis. J Clin Invest. 2020. 130(11): 6151‐7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Czaikoski, P.G. , Mota, J.M. , Nascimento, D.C. , Sonego, F. , Castanheira, F.V. , Melo, P.H. , et al., Neutrophil Extracellular Traps Induce Organ Damage during Experimental and Clinical Sepsis. PLoS One. 2016. 11(2): e0148142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Llitjos, J.F. , Leclerc, M. , Chochois, C. , Monsallier, J.M. , Ramakers, M. , Auvray, M. , et al., High incidence of venous thromboembolic events in anticoagulated severe COVID‐19 patients. J Thromb Haemost. 2020. 18(7): 1743‐6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Connors, J.M. and Levy, J.H. , COVID‐19 and its implications for thrombosis and anticoagulation. Blood. 2020. 135(23): 2033‐40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Panda, R. , Castanheira, F.V. , Schlechte, J.M. , Surewaard, B.G. , Shim, H.B. , Zucoloto, A.Z. , et al., A functionally distinct neutrophil landscape in severe COVID‐19 reveals opportunities for adjunctive therapies. JCI Insight. 2022. 7(2). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Liu, Y. , Du, X. , Chen, J. , Jin, Y. , Peng, L. , Wang, H.H.X. , et al., Neutrophil‐to‐lymphocyte ratio as an independent risk factor for mortality in hospitalized patients with COVID‐19. J Infect. 2020. 81(1): e6‐e12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Bhaskar, S. , Sinha, A. , Banach, M. , Mittoo, S. , Weissert, R. , Kass, J.S. , et al., Cytokine Storm in COVID‐19‐Immunopathological Mechanisms, Clinical Considerations, and Therapeutic Approaches: The REPROGRAM Consortium Position Paper. Frontiers in immunology. 2020. 11: 1648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Hussman, J.P. , Cellular and Molecular Pathways of COVID‐19 and Potential Points of Therapeutic Intervention. Frontiers in pharmacology. 2020. 11: 1169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Teluguakula, N. , Neutrophils Set Extracellular Traps to Injure Lungs in Coronavirus Disease 2019. J Infect Dis. 2021. 223(9): 1503‐5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Goshua, G. , Pine, A.B. , Meizlish, M.L. , Chang, C.H. , Zhang, H. , Bahel, P. , et al., Endotheliopathy in COVID‐19‐associated coagulopathy: evidence from a single‐centre, cross‐sectional study. Lancet Haematol. 2020. 7(8): e575‐e82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Klok, F.A. , Kruip, M. , van der Meer, N.J.M. , Arbous, M.S. , Gommers, D. , Kant, K.M. , et al., Incidence of thrombotic complications in critically ill ICU patients with COVID‐19. Thromb Res. 2020. 191: 145‐7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Middeldorp, S. , Coppens, M. , van Haaps, T.F. , Foppen, M. , Vlaar, A.P. , Muller, M.C.A. , et al., Incidence of venous thromboembolism in hospitalized patients with COVID‐19. J Thromb Haemost. 2020. 18(8): 1995‐2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Helms, J. , Tacquard, C. , Severac, F. , Leonard‐Lorant, I. , Ohana, M. , Delabranche, X. , et al., High risk of thrombosis in patients with severe SARS‐CoV‐2 infection: a multicenter prospective cohort study. Intensive Care Med. 2020. 46(6): 1089‐98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Bradley, B.T. , Maioli, H. , Johnston, R. , Chaudhry, I. , Fink, S.L. , Xu, H. , et al., Histopathology and ultrastructural findings of fatal COVID‐19 infections in Washington State: a case series. Lancet. 2020. 396(10247): 320‐32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Zhang, S. , Liu, Y. , Wang, X. , Yang, L. , Li, H. , Wang, Y. , et al., SARS‐CoV‐2 binds platelet ACE2 to enhance thrombosis in COVID‐19. J Hematol Oncol. 2020. 13(1): 120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Price, L.C. , McCabe, C. , Garfield, B. and Wort, S.J. , Thrombosis and COVID‐19 pneumonia: the clot thickens! Eur Respir J. 2020. 56(1). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Gupta, A. , Madhavan, M.V. , Sehgal, K. , Nair, N. , Mahajan, S. , Sehrawat, T.S. , et al., Extrapulmonary manifestations of COVID‐19. Nat Med. 2020. 26(7): 1017‐32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Magro, C. , Mulvey, J.J. , Berlin, D. , Nuovo, G. , Salvatore, S. , Harp, J. , et al., Complement associated microvascular injury and thrombosis in the pathogenesis of severe COVID‐19 infection: A report of five cases. Transl Res. 2020. 220:1‐13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Iliadi, V. , Konstantinidou, I. , Aftzoglou, K. , Iliadis, S. , Konstantinidis, T.G. and Tsigalou, C. , The Emerging Role of Neutrophils in the Pathogenesis of Thrombosis in COVID‐19. International journal of molecular sciences. 2021. 22(10). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Fletcher‐Sandersjoo, A. and Bellander, B.M. , Is COVID‐19 associated thrombosis caused by overactivation of the complement cascade? A literature review. Thromb Res. 2020. 194:36‐41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Nicolai, L. , Leunig, A. , Brambs, S. , Kaiser, R. , Weinberger, T. , Weigand, M. , et al., Immunothrombotic Dysregulation in COVID‐19 Pneumonia Is Associated With Respiratory Failure and Coagulopathy. Circulation. 2020. 142(12): 1176‐89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. de Bont, C.M. , Boelens, W.C. and Pruijn, G J M. , NETosis, complement, and coagulation: a triangular relationship. Cell Mol Immunol. 2019. 16(1): 19‐27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Zucoloto, A.Z. and Jenne, C.N. , Platelet‐Neutrophil Interplay: Insights Into Neutrophil Extracellular Trap (NET)‐Driven Coagulation in Infection. Front Cardiovasc Med. 2019. 6:85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Fuchs, T.A. , Brill, A. , Duerschmied, D. , Schatzberg, D. , Monestier, M. , Myers D.D., Jr. , et al., Extracellular DNA traps promote thrombosis. Proceedings of the National Academy of Sciences of the United States of America. 2010. 107(36): 15880‐5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Keshari, R.S. , Jyoti, A. , Dubey, M. , Kothari, N. , Kohli, M. , Bogra, J. , et al., Cytokines induced neutrophil extracellular traps formation: implication for the inflammatory disease condition. PLoS One. 2012. 7(10): e48111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Gorham, J. , Moreau, A. , Corazza, F. , Peluso, L. , Ponthieux, F. , Talamonti, M. , et al., Interleukine‐6 in critically ill COVID‐19 patients: A retrospective analysis. PLoS One. 2020. 15(12): e0244628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Kim, J.S. , Lee, J.Y. , Yang, J.W. , Lee, K.H. , Effenberger, M. , Szpirt, W. , et al., Immunopathogenesis and treatment of cytokine storm in COVID‐19. Theranostics. 2021. 11(1): 316‐29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Du, F. , Liu, B. and Zhang, S. , COVID‐19: the role of excessive cytokine release and potential ACE2 down‐regulation in promoting hypercoagulable state associated with severe illness. J Thromb Thrombolysis. 2021. 51(2): 313‐29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Wang, J. , Jiang, M. , Chen, X. and Montaner, L.J. , Cytokine storm and leukocyte changes in mild versus severe SARS‐CoV‐2 infection: Review of 3939 COVID‐19 patients in China and emerging pathogenesis and therapy concepts. J Leukoc Biol. 2020. 108(1): 17‐41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Kaiser, R. , Leunig, A. , Pekayvaz, K. , Popp, O. , Joppich, M. , Polewka, V. , et al., Self‐sustaining IL‐8 loops drive a prothrombotic neutrophil phenotype in severe COVID‐19. JCI Insight. 2021. 6(18). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Bonaventura, A. , Vecchie, A. , Dagna, L. , Martinod, K. , Dixon, D.L. , Van Tassell, B.W. , et al., Endothelial dysfunction and immunothrombosis as key pathogenic mechanisms in COVID‐19. Nature reviews Immunology. 2021. 21(5): 319‐29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Petito, E. , Falcinelli, E. , Paliani, U. , Cesari, E. , Vaudo, G. , Sebastiano, M. , et al., Association of Neutrophil Activation, More Than Platelet Activation, With Thrombotic Complications in Coronavirus Disease 2019. J Infect Dis. 2021. 223(6): 933‐44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Lefrancais, E. , Mallavia, B. , Zhuo, H. , Calfee, C.S. and Looney, M.R. , Maladaptive role of neutrophil extracellular traps in pathogen‐induced lung injury. JCI Insight. 2018. 3(3). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Denning, N.L. , Aziz, M. , Gurien, S.D. and Wang, P. , DAMPs and NETs in Sepsis. Frontiers in immunology. 2019. 10:2536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Mitsios, A. , Arampatzioglou, A. , Arelaki, S. , Mitroulis, I. and NETopathies?, R.K. , Unraveling the Dark Side of Old Diseases through Neutrophils. Frontiers in immunology. 2016. 7:678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Rebetz, J. , Semple, J.W. and Kapur, R. , The Pathogenic Involvement of Neutrophils in Acute Respiratory Distress Syndrome and Transfusion‐Related Acute Lung Injury. Transfus Med Hemother. 2018. 45(5): 290‐8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Bendib, I. , de Chaisemartin, L. , Granger, V. , Schlemmer, F. , Maitre, B. , Hue, S. , et al., Neutrophil Extracellular Traps Are Elevated in Patients with Pneumonia‐related Acute Respiratory Distress Syndrome. Anesthesiology. 2019. 130(4): 581‐91. [DOI] [PubMed] [Google Scholar]
- 82. Group WHOREAfC‐TW , Shankar‐Hari, M. , Vale, C.L. , Godolphin, P.J. , Fisher, D. , Higgins, J.P.T. , et al., Association Between Administration of IL‐6 Antagonists and Mortality Among Patients Hospitalized for COVID‐19: A Meta‐analysis. Jama. 2021. 326(6): 499‐518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Narasaraju, T. , Tang, B.M. , Herrmann, M. , Muller, S. , Chow, V.T.K. and Radic, M. , Neutrophilia and NETopathy as Key Pathologic Drivers of Progressive Lung Impairment in Patients With COVID‐19. Frontiers in pharmacology. 2020. 11:870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Group, R.C. , Horby, P. , Lim, W.S. , Emberson, J.R. , Mafham, M. , Bell, J.L. , et al., Dexamethasone in Hospitalized Patients with Covid‐19. N Engl J Med. 2021. 384(8): 693‐704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Wang, Y. , Wang, H. , Zhang, C. , Zhang, C. , Yang, H. , Gao, R. , et al., Lung fluid biomarkers for acute respiratory distress syndrome: a systematic review and meta‐analysis. Crit Care. 2019. 23(1): 43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Li, H. , Zhang, J. , Fang, C. , Zhao, X. , Qian, B. , Sun, Y. , et al., The prognostic value of IL‐8 for the death of severe or critical patients with COVID‐19. Medicine (Baltimore). 2021. 100(11): e23656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Bulow Anderberg, S. , Luther, T. , Berglund, M. , Larsson, R. , Rubertsson, S. , Lipcsey, M. , et al., Increased levels of plasma cytokines and correlations to organ failure and 30‐day mortality in critically ill Covid‐19 patients. Cytokine. 2021. 138:155389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Li, J. , Rong, L. , Cui, R. , Feng, J. , Jin, Y. , Chen, X. , et al., Dynamic changes in serum IL‐6, IL‐8, and IL‐10 predict the outcome of ICU patients with severe COVID‐19. Ann Palliat Med. 2021. 10(4): 3706‐14. [DOI] [PubMed] [Google Scholar]
- 89. Luo, Y. , Mao, L. , Yuan, X. , Xue, Y. , Lin, Q. , Tang, G. , et al., Prediction Model Based on the Combination of Cytokines and Lymphocyte Subsets for Prognosis of SARS‐CoV‐2 Infection. J Clin Immunol. 2020. 40(7): 960‐9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Landoni, G. , Piemonti, L. , Monforte, A.D. , Grossi, P. , Zangrillo, A. , Bucci, E. , et al., A Multicenter Phase 2 Randomized Controlled Study on the Efficacy and Safety of Reparixin in the Treatment of Hospitalized Patients with COVID‐19 Pneumonia. Infect Dis Ther. 2022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Temesgen, Z. , Burger, C.D. , Baker, J. , Polk, C. , Libertin, C.R. , Kelley, C.F. , et al., Lenzilumab in hospitalised patients with COVID‐19 pneumonia (LIVE‐AIR): a phase 3, randomised, placebo‐controlled trial. Lancet Respir Med. 2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Kyriazopoulou, E. , Poulakou, G. , Milionis, H. , Metallidis, S. , Adamis, G. , Tsiakos, K. , et al., Early treatment of COVID‐19 with anakinra guided by soluble urokinase plasminogen receptor plasma levels: a double‐blind, randomized controlled phase 3 trial. Nat Med. 2021. 27(10): 1752‐60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Declercq, J. , Van Damme, K.F.A. , De Leeuw, E. , Maes, B. , Bosteels, C. , Tavernier, S.J. , et al., Effect of anti‐interleukin drugs in patients with COVID‐19 and signs of cytokine release syndrome (COV‐AID): a factorial, randomised, controlled trial. Lancet Respir Med. 2021. 9(12): 1427‐38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Elliott, W., Jr. , Guda, M.R. , Asuthkar, S. , Teluguakula, N. , Prasad, D.V.R. , Tsung, A.J. , et al., PAD Inhibitors as a Potential Treatment for SARS‐CoV‐2 Immunothrombosis. Biomedicines. 2021. 9(12). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Lee, Y.Y. , Park, H.H. , Park, W. , Kim, H. , Jang, J.G. , Hong, K.S. , et al., Long‐acting nanoparticulate DNase‐1 for effective suppression of SARS‐CoV‐2‐mediated neutrophil activities and cytokine storm. Biomaterials. 2021. 267:120389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Fisher, J. , Mohanty, T. , Karlsson, C.A.Q. , Khademi, S.M.H. , Malmstrom, E. , Frigyesi, A. , et al., Proteome Profiling of Recombinant DNase Therapy in Reducing NETs and Aiding Recovery in COVID‐19 Patients. Mol Cell Proteomics. 2021. 20:100113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Holliday, Z.M. , Earhart, A.P. , Alnijoumi, M.M. , Krvavac, A. , Allen, L.H. and Schrum, A.G. , Non‐Randomized Trial of Dornase Alfa for Acute Respiratory Distress Syndrome Secondary to Covid‐19. Frontiers in immunology. 2021. 12:714833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Tamakuma, S. , Ogawa, M. , Aikawa, N. , Kubota, T. , Hirasawa, H. , Ishizaka, A. , et al., Relationship between neutrophil elastase and acute lung injury in humans. Pulm Pharmacol Ther. 2004. 17(5): 271‐9. [DOI] [PubMed] [Google Scholar]
- 99. Aikawa, N. , Ishizaka, A. , Hirasawa, H. , Shimazaki, S. , Yamamoto, Y. , Sugimoto, H. , et al., Reevaluation of the efficacy and safety of the neutrophil elastase inhibitor, Sivelestat, for the treatment of acute lung injury associated with systemic inflammatory response syndrome; a phase IV study. Pulm Pharmacol Ther. 2011. 24(5): 549‐54. [DOI] [PubMed] [Google Scholar]
- 100. Hayakawa, M. , Katabami, K. , Wada, T. , Sugano, M. , Hoshino, H. , Sawamura, A. , et al., Sivelestat (selective neutrophil elastase inhibitor) improves the mortality rate of sepsis associated with both acute respiratory distress syndrome and disseminated intravascular coagulation patients. Shock. 2010. 33(1): 14‐8. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.