

Abstract
Background
Cadmium is a non-essential, third largest heavy metal contaminant with long retention time that poses environmental hazards. It emanating majorly from industrial processes and phosphate fertilizers. Cadmium is effortlessly assimilated by plants and leads to yield loss. Henceforth, identification of mechanisms to attenuate the heavy metal toxicity in crops is beneficial for enhanced yields.
Results
Beneficial soil bacteria have been known to combat both biotic and abiotic stress, thereby promoting plant growth. Amongst them, Pseudomonas fluorescens has been shown to enhance abiotic stress resistance in umpteen crops for instance maize and groundnut. Here, we investigated the role of P. fluorescens in conferring cadmium stress resistance in Arabidopsis thaliana. In silico analysis of PCR2 gene and promoter revealed the role, in cadmium stress resistance of A. thaliana. Real-time expression analysis employing qRT-PCR ratified the upregulation of AtPCR2 transcript under cadmium stress up to 6 folds. Total leaf (50%), biomass (23%), chlorophyll content (chlorophyll-a and b 40%, and 36 %) silique number (50%), and other growth parameters significantly improved on bacterial treatment of the 2mM Cd-stressed plants.
Conclusion
Moreover, generated 35s-promoter driven AtPCR2 over-expressing transgenic lines that exhibited resistance to cadmium and other heavy metal stress. Taken together, a crucial interplay of P. fluorscens mediated enhanced expression of AtPCR2 significantly induced cadmium stress resistance in Arabidopsis plants.
Supplementary Information
The online version contains supplementary material available at 10.1186/s43141-022-00457-7.
Keywords: Cadmium stress, PCR2, Pseudomonas, Transgenics, Arabidopsis
Background
Cadmium (Cd) is a non-essential, toxic heavy metal, enormously increasing in the environment due to industrialization. It is one of the global major pollutants, and it has highly toxic nature due to reactivity with sulfhydryl groups, which may inhibit the enzymatic activities [1]. Cd significantly inhibits the plant growth and biomass by interrupting water, mineral uptake, and photosynthesis [2]. It negatively impacts photosynthesis via disorganization of grana, thereby leading to reduction in chlorophyll biosynthesis [3]. Cd is toxic for plants even at low concentrations, while concentrations higher than 5–10 μg Cd g-1 leaf dry weight are lethal to plants [4]. In nature, plants exhibit an innate tolerance to Cd stress to a certain extent; however, at elevated Cd concentrations, improved growth and crop yields may only be garnered via employing novel strategies to withstand Cd or heavy metal toxicity for sustainable agriculture development.
Mutual interactions between beneficial mycorrhizae helper bacteria and plants have been reported to accelerate crop growth and yields. Recent studies revealed that bacterial interactions contributed to increased biotic and abiotic stress resistance in plants [5–10]. Pseudomonas fluorescens is a non-toxic, ecofriendly, and widely distributed rhizo-bacterium, which releases beneficial phyto-hormones, anti-microbial substances, metal chelators, and volatile organic compounds (VOCs) as well as excretes hydrolytic enzymes, such as proteases, cellulose, chitinase, and β-glucanase [11, 12]. This potent diversity of compounds directly or indirectly promotes growth, in addition to biotic and abiotic stress resistance in umpteen crops. Multiple reports exhibited the protective role of P. fluorescens in conferring stress resistance [12–14]. Conjointly, around 25 volatile bacterial odors elicited have been identified, which were correlated to numerous differentially expressing plant mRNAs related to stress responsiveness, hormonal regulation, and cell metabolism. Previously, Wang and coworkers [15] performed microarray analysis of Arabidopsis thaliana treated with P. fluorescens and revealed an upregulation in 95 genes at the transcript level. Amongst them, the plant cadmium resistance 2 gene (PCR2) exhibited increased (5.4-fold) transcript levels corresponding to heavy metal tolerance. This motivated us to elucidate the potential impact of Arabidopsis-Pseudomonas interaction on the degree of Cd resistance within the host plant.
The present investigation was undertaken in order to reveal the role of P. flourescens on Arabidopsis plant growth and heavy metal (Cd) tolerance. To validate the Arabidopsis thaliana PCR2 (AtPCR2) expression, we performed qRT-PCR in accordance to [15]. Invariably, AtPCR2 gene expression was monitored in P. fluorescens treated A. thaliana interaction studies during Cd stress. Further, to corroborate the findings from qRT-PCR of AtPCR2, in silico analysis of its promoter region was performed to highlight its role in Cd stress resistance. Moreover, we assayed the biomass, chlorophyll content, leaf, and silique numbers of A. thaliana Cd stressed lines that we primed with P. fluorescens in comparison to control lines (in the absence of bacterial treatment). To elucidate the underlying molecular mechanism, we have generated AtPCR2 transgenic lines employing floral dip method. Overexpression studies of PCR2 gene in Cd-stressed A. thaliana lines would mimic the P. fluorescens treatment and enable us to obtain insights into the mechanism of Cd-stress resistance. Insights into the influence of P. fluorescens on plant growth and heavy metal resistance in Arabidopsis may be extrapolated to other crops for modulating both productivity and stress resistance.
Methods
Bacterial suspension cultures
P. fluorescens KACC10327 was streaked onto Luria Bertani (LB) agar plates and grown at 28+1°C in dark for 24h. Single discrete colony was picked from LB agar plate using sterile tooth pick and transferred into liquid LB media followed by incubation at 28+1 °C (250 rpm) to yield 10-9ml−1colony-forming units (CFU), as calculated by optical density (OD) and serial dilution counts [6].
AtPCR2 transcript analysis in Arabidopsis thaliana treated with P. fluorescens
Plants were kept in growth chamber enabled with 16/8h light (with 120–150 μmol/m2 s) and dark photoperiod. Healthy growing 14 days old plants were subjected with P. fluorescens cells with 50 μl of (1×108 bacteria/ml) simultaneously control plant was treated with LB media. The total RNA was isolated from P. fluorescens treated A. thaliana ecotype Col-0 seedlings at various time points, i.e., 0, 3, 6, and 12 h post-inoculation. The total RNA was isolated by TRIZOL method (Invitrogen, San Diego, CA) according to manufacturer’s instructions. cDNA synthesis was performed using mRNA (1.0 μg) employing an ImProm-II first-strand cDNA synthesis system (Promega Corp., Madison, WI) with an oligo(dT)18 primer. Real-time qRT-PCR was performed using SYBR Green PCR Master Mix (Applied Biosystems, San Diego, CA) and an ABI Step One Plus thermocycler (Applied Biosystems). Cycling conditions were maintained as follows; 95°C for 10 min, followed by 40 cycles of 95°C for 30 s, 52°C for 25 s, and 72°C for 30 s. Each experiment was performed in triplicate with three independent replicates. The RT-PCR amplification and quantification of the expression of AtPCR2 gene (NM_001332138.1) transcript levels was normalized against Actin2 gene (NM_001338358.1) as an internal control. The list of primers used in this study is mentioned in the Table 1.
Table 1.
Primers used in this study listed in the table
| Gene | Primer | Product size (bp) |
|---|---|---|
| AtPCR2 F(Full length) | 5′-TACAAAACCTTACATTGCTTTC-3′ | 859 |
| AtPCR2 R(Full length) | 5′-TATTGTTTTGGTGTCACTTTCT-3′ | |
| AtPCR2 F(RT-PCR) | 5′-GGTCCACAGGCTTCTGTGAT-3′ | 99 |
| AtPCR2 R(RT-PCR) | 5′-CTACAATCTCGGCGACTTGG-3′ | |
| Actin-2 F | 5′-AGT GGT CGT ACA ACC GGT ATT GT-3′ | 92 |
| Actin-2 R | 5′-GAT GGC ATG AGG AAG AGA GAA AC-3′ |
In silico analysis of promoter
Upstream genomic sequences (∼1 kb from transcription start) of Arabidopsis thaliana PCR2 gene were retrieved from TAIR9. The cis-regulatory element analysis was performed using http://www.bioinformatics2.wsu.edu/cgi-bin/Athena/cgi/visualize.pl. Identified elements were highlighted with different colors in the Fig. 2
Fig. 2.

Putative cis-regulatory elements identified in the promoter regions of PCR2 gene from Arabidopsis thaliana. The approximate positions of putative cis-regulatory elements were predicted in the 1 kb upstream region of the PCR2 gene by PlantCARE database
Expression analysis of PCR2 gene using micro array data and co-expression
The expression data of Arabidopsis PCR2 genes were retrieved from the AtGenExpress (http://jsp.weigelworld.org/expviz/expviz.jsp) under various stresses as described previously [16]. The log2-transformed values were used to generate heat maps, and hierarchical clustering was performed using MeV software [17]. AtPCR2 gene regulatory network was identified by co-expression analysis tool http://genecat.mpg.de/cgi-bin/Ainitiator.py
Plant growth materials and treatments
A. thaliana (ecotype Col-0) seeds were initially surface sterilized with 70% ethanol for 5 min, followed by 2% sodium hypochlorite (NaOCl) for 10 min, and rinsed with sterile distilled or (milliQ) water about ten times. Seeds were germinated on MS media (half-strength) and 7-day-old seedlings of uniform growth were carefully transferred to pre-sterilized plastic pots (diameter 10cm, depth 8cm, one seedling per pot) containing 100g of heat-sterilized (95°C, 36 h) vermiculite and soil mix (1:1) supplied with Hoagland’s nutrient solution [25 mL of half strength; KNO3(5mM), (NH)4H2PO4 (1mM), Ca(NO3)2 (0.5mM), MgSO4 (0.5mM), Fe-Citrate (60μM), H3BO3(92μM), MnCl2·4H2O (18μM), ZnSO4·7H2O (1.6μM), CuSO4·5H2O (0.6μM), and (NH 4)6Mo7O24·4H2O (0.7 μM)] once a week. After 1 week, the soil was treated with bacterial suspension culture (10-9 cfu/ml in 1m LB) and soil inoculated with only liquid LB medium (1mL) was used as control. In case of administration of cadmium stress treatments, seedlings were supplied with nutrient solution supplemented with CdCl2 (1, 2 mM). The experiments were replicated in triplicate. Arabidopsis plants were maintained under controlled conditions at 23+1°C in the growth chamber.
Quantification of plant growth parameters
We employed Arabidopsis plants (30 days old) for determination of plant growth and physiological index measurements. Total plant leaf and silique numbers were counted. Plants were gently detached from the pots and fresh root and shoot weights, root length, and shoot height were measured. Leaf chlorophyll content was estimated according to [18]. Fresh leaf samples were ground thoroughly with acetone (80%) in the dark and centrifuged at 9000 g for 10 min at 4°C. Absorbance was measured at 645 and 663nm for collected supernatant on UV-2102C Spectrophotometer, Unico Instrument Co., Ltd, Shanghai, China.
Statistical analysis
Data generated by assaying of parameters for instance growth; physiological index, transgenic analysis, and so on were compiled in the form of mean standard deviations (n=6). Statistical analyses, one-way ANOVA, and Duncan’s multiple range tests were performed.
Cloning of AtPCR2 cDNA and plant transformation
The AtPCR2 cDNA expression cassette that comprised of 35S CaMV Promoter: AtPCR2 cDNA:NOS-terminator was cloned into pCB302ES plant transformation vector harbouring BASTA resistant gene “bar” as the selection marker and hemagglutinin (HA) epitope tag [19]. Further, the recombinant plant transformation vector harbouring the AtPCR2 expression construct was transformed into Arabidopsis thaliana employing the agrobacterium (GV3101)-mediated floral-dip method [20]. We generated putative T0, T1, T2, and T3 transgenic lines that were screened by PCR analyses employing genes specific primer for AtPCR2 and BAR. In order to perform stress experiments, T3 homozygous lines harboring AtPCR2 were used for further experiments.
Protein extraction and immunoblot analysis of AtPCR2 transgenic lines
Total solubilized rosette leaf proteins were extracted from transgenic and wild type lines using extraction buffer [SDS (2%), TRIS (pH 6.8; 60mM), β-mercaptoethanol (14.4mM), glycerol (10%), and bromophenol blue (0.1% (w/v)]. Total protein concentration was determined using Bio-Rad protein assay reagent (Bio-Rad Laboratories, Hercules, CA, USA) and BSA (bovine serum albumin; Kit II, Catalog No. 500-0002) as standard. Protein extracts (50 μg) were fractionated on a 10% SDS–PAGE and electro-blotted onto a nitrocellulose PVDF membrane (Hybond-C membrane; Amersham Biosciences, Piscataway, NJ, USA) according to [21]. AtPCR2-HA proteins were detected using anti-HA-HRP IgG antibodies (Cell Signaling, Hitchin, UK) and visualized using a Super Signal® West Femto maximum sensitivity substrate kit (Thermo Scientific, Waltham, USA).
Stress analysis of transgenic lines
Sterilized seeds of transgenic and WT lines were germinated on MS (half-strength) media. Seedlings (7 days old) were transferred to fresh MS (half-strength) media supplemented with various abiotic stressors for instance; salt (NaCl; 150 mM), heavy metal (CdCl2; 50 μM), cold (only MS media; 4°C), and (80μM CuSO4).
Results
P. fluorescens-induced AtPCR2 transcript expression confers Cd tolerance
The qRT-PCR was carried out to quantify the real-time expression profiles of AtPCR2 transcript in Arabidopsis lines that were treated with P. fluorescens for varying time periods in comparison to WT. The AtPCR2 transcript abundance was normalized against that of internal control actin. We found that AtPCR2 transcript levels increased in a time-dependent manner, i.e., AtPCR2 transcript levels enhanced by 1.75-, 4.8- and 6-folds after 3h, 6h, and 12h interaction with P. fluorscens, respectively (Fig. 1). Results strongly suggested that the enhanced expression ofAtPCR2 gene in Arabidopsis lines on interaction with P. fluorscens had a definite functional role in conferring cadmium tolerance to them.
Fig. 1.
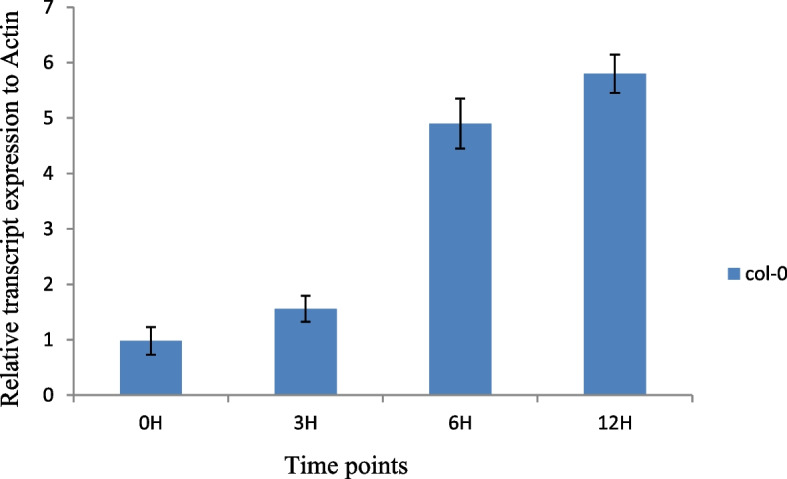
Relative AtPCR2 m-RNA expression level while Pseudomonas fluorescens and Arabidopsis thaliana interaction. Bars indicate standard deviation (n = 3). Statistically significant differences (Student’s t test) between transgenic lines and Col-0 were indicated (p < 0.05)
In silico analysis of promoter
In order to unveil the transcriptional regulation of AtPCR2 gene in Arabidopsis, we retrieved the 1 kb upstream regions or putative promoter sequences and analyzed them using PlantCARE. A number of important cis-regulatory elements and stress-responsive motifs were identified (Fig. 2). A few known stress-related cis-regulatory elements (CREs) involved in stress responsive expressions for instance-dehydration responsive MYB1AT and transcriptional activator of ABA (MYB2AT), cold-response element (Evening Element), Gibberellin responsive element (GARE), phytochrome-modulated (CCA1), and wound-responsive WRKY (W-Box) family elements were revealed in the putative promoter region of AtPCR2 gene. The presence of these stress-related motifs in the upstream promoter region of AtPCR2 gene may be directly correlated to the altered gene expression on interaction with P. fluorscens.
Microarray data and co-expression analysis
In response to stress, AtPCR2 gene expression was high in roots, whereas mild expression was detected in shoots. Interestingly, we noted that AtPCR2 expression was highly upregulated in response to cold, salt, oxidative stress, and UVB exposure in the roots across different time periods within 24 h (Supplementary Figures S1 and S2).
Pseudomonas fluorescens boosted Arabidopsis growth during Cd stress
P. fluorescens interaction led to enhanced growth of A. thaliana, even under cadmium stress (Fig. 3). Total plant biomass (fresh weight) increased by about 25%, with P. fluorescens treatment in comparison to untreated WT during normal conditions. However, during P. fluorescens treatment in combination with Cd stress (1mM, 2 mM), the plant biomass revealed further enhancement by 58 and 23 %, respectively, in comparison to stand alone Cd treatment (Fig. 4A). Leaf and silique number showed an increment of 69%, 50%, and 41.3%, 30.56%, during combinatorial treatment of cadmium stress (1mM and 2mM, respectively) along with the presence of P. fluorescens; in comparison to only Cd treatment (Fig. 4B, C). P. fluorescens significantly improved chlorophyll a and b content in Arabidopsis thaliana by 25% and 40% (P <0.01) and 45 % (P <0.05), 36 % (P <0.01) during cadmium treatments 1mM and 2mM, respectively (Fig. 5).
Fig. 3.

Col-0: Columbia-0 Arabidopsis line. a Control. b Pseudomonas fluorescens treated. Col-0 + Cd: Columbia-0 plants with cadmium stress. a 1mM. b 2mM. Col-0 + Cd + Pf: In the presence of Pseudomonas fluorescens cadmium treatment to Arabidopsis plants. a 1mM and b 2mM
Fig. 4.

Effects of Pseudomonas fluorescens on total biomass, leaf, and silique number of Arabidopsis under different CdCl2 concentrations. Values are means and bars indicate SDs (n=6) columns with a, b, and c letters indicate statistically significant difference at P< 0.05 (Duncan test)
Fig. 5.

Effects of P. fluorescens on chlorophyll a and b content of Arabidopsis under different CdCl2 concentrations. Values are means and bars indicate SDs (n=6) columns with a, b, and c letters indicate statistically significant difference at P< 0.05 (Duncan test)
Transgenic Arabidopsis thaliana lines expressing AtPCR2
To investigate the function of AtPCR2 in effectuating plant resistance responses against cadmium stress, we generated transgenic Arabidopsis lines over expressing AtPCR2 gene and selected two independent T3 homozygous lines; T3-12 and T3-18. Under normal growth condition, both line T3-12 and T3-18 exhibited similar growth and developmental patterns to WT (Figs. 6Aa, S3). Among selected transgenic lines, line T3-12 exhibited higher expression level of AtPCR2 than line T3-18 (Fig. 6Bb). Further, AtPCR2-HA proteins were detected in transgenic lines. Among transgenic lines, the accumulation of AtPCR2-HA was higher in T3-12 than T3-18. We detected the accumulation of AtPCR2-HA protein in the T3-12 and T3-18 transgenic Arabidopsis plants, whereas no AtPCR2-HA protein was accumulated in WT (Fig. 6b). The rubisco protein was used as a reference (54KD) (Fig. 6c) protein which is abundantly present in the universe).
Fig. 6.

A Plant transformation vector construction with restriction sites. B a Col-0 and transgenic lines (Col-0, T3-12, and T3-18) grown at normal conditions; b Lane B contains Western blot of Col-0, T3-12, and T3-18 (overexpressed PCR2 protein-detected 17kDa with HA-tagged antibodies); c Line C contains SDS-PAGE of the Col-0, T3-12, and T3-18 (Rubisco protein 54 kDa)
Overexpression of AtPCR2 confers resistance against stress
In a bid to investigate the role of AtPCR2 gene over expression in conferring stress resistance to A. thaliana, we exposed both transgenic and WT lines to various abiotic stresses. AtPCR2 over-expressing A. thaliana lines showed significantly enhanced resistance various stresses for instance; cadmium chloride (CdCl2), copper sulphate (CuSO4), sodium chloride (NaCl), and cold in comparison to WT. T3-12 line exhibited higher levels of stress resistance than T3-18 line (Figs. 7, S4).
Fig. 7.

Various abiotic stresses effect on AtPCR2 transgenic lines: A Heavy metal (50-μM cadmium chloride) treatment. B An 80-μM CuSO4 heavy metal treatment. C Cold stress treatment (4°C). D Salt stress (150-mM sodium chloride). Col-0, columbia-0 control; AtPCR2 line, transgenic Arabidopsis line
Discussion
Previously, it has been elucidated that beneficial soil bacteria promote growth of umpteen plant species [10, 22, 23]. Beneficent soil bacteria-mediated plant growth promotion induced during abiotic stress conditions has been observed in several cultivated and wild plant species including tomato (Solanum lycopersicum) [24], chickpea (Cicer arietinum) [25], and alfalfa (Medicago sativa) [26]. Numerous studies revealed that plant-growth promoting rhizobacteria (PGPR) improve plant growth and yield during adverse ambient conditions via production of a spectrum of siderophores, ACC-dehydrogenase, microbial-emitted volatiles (mVOCs), or chemicals and other hormone-modulated compounds. Hormone-modulated compounds trigger the differential expression of numerous transcripts involved in cell-wall modifications, primary and secondary metabolisms, stress responses, hormone regulation, and protein expression in Arabidopsis [16]. Similarly, Zhang and colleagues [6] have shown PGPR GB03-induced HKT1 gene expression augments salt resistance. A recent study has revealed that Azotobacter vinelandii strain SRI Az3 aids to combat chromium stress tolerance in rice plants via significantly boosting its antioxidant machinery [27].
The present study focuses on the underlying mechanisms of PCR2 upregulation and its role in imparting cadmium stress resistance to Arabidopsis. We elucidated the direct correlation between Pseudomonas triggered PCR2 gene induction and its contribution towards abiotic stress tolerance in Arabidopsis plants. Invariably, it provides direct evidence for high transcript induction of AtPCR2 that up-regulates by 5.4 folds within a 12-h span of interaction between Arabidopsis (host) and Pseudomonas (Fig. 1). In line with our results, Wang et al. data [16] demonstrate similar PCR2 transcript induction that peaks (six-fold) at 12h time-point, post interaction with Pseudomonas. In silico investigations of cis-regulatory elements in the upstream region of PCR2 gene reveals crucial stress regulatory elements for instances cold responsive MYB1, MYB2, gibberellin responsive GARE and salicylic acid responsive, and wound responsive WRKY family W-box (Fig. 2). PCR2 co-expression analysis suggests that it is closely related tothe PLAC8 family of proteins (At4g23470, At1g52200) and AtPCR1, which play imperative roles in determining fruit size and heavy metal transportation [1] (Fig. S1). Microarray expression analysis across multiple abiotic stress treatments reveals differential expression of PCR2 with abundance in roots as compared to aerial parts (Fig. S2). In silico analysis exhibits numerous resident cis-acting elements in the upstream promoter region of PCR2 that evidently displays its crucial involvement in combating abiotic stress.
During stress conditions, enhanced growth of Arabidopsis plants directly correlates to the presence of Pseudomonas in soil. Our results are consistent with previous reports demonstrating that Pseudomonas inoculation in soil significantly increases stress tolerance in plants [13, 14]. Previous studies suggest that P. fluorescens can tolerate high concentrations of Cd [28]. Moreover, a strain of P. aeruginosa performs as an efficient Cd accumulator [29]. Reports are consistent with our analysis of improved Cd tolerance levels in A. thaliana (up to ~0.75mM) on interaction with P. fluorescens. Further, an exposure of Arabidopsis plants to CdCl2 (1–2 mM) results in significantly high levels of tolerance to Cd along with growth enhancements in comparison to WT (Fig. 3).
Leaf development plays a crucial role in plant survival and growth, since it affects the area available for photosynthesis, which in turn corresponds positively to plant biomass accumulation [30, 31]. In the current study, Pseudomonas inoculation significantly increases the total biomass, silique-, and leaf number per plant, in Arabidopsis (Fig. 4). In addition, leaf chlorophyll content is an important physiological trait that directly correlates to photosynthetic rate in plants [32]. Previous studies show that plants grown under Cd stress, synthesize less chlorophyll, abnormal grana structure, and low dry matter than those without Cd exposure [3]. Several studies confirm that PGPRs augment photosynthetic capacity by increasing photosynthetic efficiency and chlorophyll content on exposure to abiotic stress [6, 10, 33]. In our investigation, Pseudomonas also remarkably enhances leaf chlorophyll content and biomass on exposure to Cd stress conditions (1 and 2 mM CdCl2) (Fig. 5). In context with this, PCR2 over-expressing transgenic T3 lines under CaMV35 promoter (Fig. 6B) exhibit higher accumulation of PCR2 protein than WT along with robust stress tolerance (Fig. 7A–D). Among the 10 isoforms of PCR gene family, the PCR2 has a vital role in heavy metal detoxification in yeast as well as in Arabidopsis [34]. Taken together, our results elucidate the efficacy of P. fluorscens in imparting heavy metal stress tolerance via inducing high expression of AtPCR2 gene. Conclusively, our results offer in-depth insights into the possible mechanisms of PGPR-Pseudomonas in combating cadmium stress in plants.
Conclusion
Although we proposed that P. fluorscens in imparting heavy metal stress tolerance via inducing high expression of AtPCR2 gene, complete molecular mechanism is yet to be elucidated to describe how it is mitigating other abiotic stress conditions.
Supplementary Information
Additional file 1: Fig. S1. PCR2 gene regulatory network analyzed by co-expression analysis.
Additional file 2: Fig. S2. Expression analysis of Arabidopsis PCR2 gene responses todifferentstress conditions in the shoot and root tissues. Microarray expression data for AtPCR2 gene was retrieved from TAIR (ver 9) during various abiotic stresses, i.e. salt, drought, osmotic, cold, heat, oxidative, genotoxic, wounding and UV/B stress. The datasets obtained for various time points of stress, namely 0.5, 1, 3, 6, 12 and 24 h, were analyzed with respect to the control. The colour bar below represents relative expression values; wherein green represents lowest, black represents medium and red signifies highest expression levels. The hierarchical clustering is performed, and heat maps have been generated using TIGR MeV software package.
Additional file 3: Fig. S3. Controlled condition grown Col-0, T3-12, T3-18 Arabidopsis thaliana plants.
Additional file 4: Fig. S4. Various abiotic stress conditions imposed to control vs overexpressed lines and their root length and biomass data. Values are means and bars indicate SDs (n=6) statistically significant difference at P< 0.05 (t-test).
Acknowledgements
This work was supported by National Research foundation of Korea (NRF-2020R1I1A1A01069595).
Abbreviations
- AtPCR2
Arabidopsis thaliana plant cadmium resistance-2 gene
- Cd
Cadmium
- mM
Milli molar
Authors’ contributions
Study conception and design: KK and CSR; data collection analysis and interpretation of the results: CSR, MC, TK, and J. JJ. The authors reviewed the results and approved the final version of the manuscript.
Funding
Not applicable.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Song WY, Hörtensteiner S, Tomioka R, Lee Y, Martinoia E. Common functions or only phylogenetically related? The large family of PLAC8 motif-containing/PCR genes. Molecules and cells. 2011;31(1):1–7. doi: 10.1007/s10059-011-0024-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gill SS, Hasanuzzaman M, Nahar K, Macovei A, Tuteja N. Importance of nitric oxide in cadmium stress tolerance in crop plants. Plant Physio and Biochem. 2013;63:254–261. doi: 10.1016/j.plaphy.2012.12.001. [DOI] [PubMed] [Google Scholar]
- 3.Siedlecka A, Krupa Z. Cd/Fe interaction in higher plants-its consequences for the photosynthetic apparatus. Photosynthetica. 1999;36(3):321–331. doi: 10.1023/A:1007097518297. [DOI] [Google Scholar]
- 4.White PJ, Brown PH. Plant nutrition for sustainable development and global health. Annals of botany. 2010;105(7):1073–1080. doi: 10.1093/aob/mcq085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Van Hulten M, Pelser M, Van Loon LC, Pieterse CM, Ton J. Costs and benefits of priming for defense in Arabidopsis. Proc Natl Acad Sci. 2006;103(14):5602–5607. doi: 10.1073/pnas.0510213103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zhang H, Kim S, Sun Y, Dowd SE, Shi H, Paré PW. Soil bacteria confer plant salt tolerance by tissue-specific regulation of the sodium transporter HKT1. Mol Plant-Microbe Interact. 2008;21(6):737–744. doi: 10.1094/MPMI-21-6-0737. [DOI] [PubMed] [Google Scholar]
- 7.Rudrappa T, Biedrzycki ML, Kunjeti SG, Donofrio NM, Czymmek KJ, Paul WP, Bais HP. The rhizobacterial elicitor acetoin induces systemic resistance in Arabidopsis thaliana Commun Integr Biol. 2010;3(2):130–138. doi: 10.4161/cib.3.2.10584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Medeiros FH, Souza RM, Medeiros FC, Zhang H, Wheeler T, Payton P, Ferro HM, Paré PW. Transcriptional profiling in cotton associated with Bacillus subtilis (UFLA285) induced biotic-stress tolerance. Plant and soil. 2011;347(1):327–337. doi: 10.1007/s11104-011-0852-5. [DOI] [Google Scholar]
- 9.De Zelicourt A, Al-Yousif M, Hirt H. Rhizosphere microbes as essential partners for plant stress tolerance. Molecular plant. 2013;6(2):242–245. doi: 10.1093/mp/sst028. [DOI] [PubMed] [Google Scholar]
- 10.Han QQ, Lü XP, Bai JP, Qiao Y, Paré PW, Wang SM, Zhang JL, Wu YN, Pang XP, Xu WB, Wang ZL. Beneficial soil bacterium Bacillus subtilis (GB03) augments salt tolerance of white clover. Front Plant Sci. 2014;5:525. doi: 10.3389/fpls.2014.00525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Weller DM. Pseudomonas biocontrol agents of soil borne pathogens: looking back over 30 years. Phytopathology. 2007;97(2):250–256. doi: 10.1094/PHYTO-97-2-0250. [DOI] [PubMed] [Google Scholar]
- 12.Hernández-León R, Rojas-Solís D, Contreras-Pérez M, del Carmen O-MM, Macías-Rodríguez LI, Reyes-delaCruz H, Valencia-Cantero E, Santoyo G. Characterization of the antifungal and plant growth-promoting effects of diffusible and volatile organic compounds produced by Pseudomonas fluorescens strains. Biol Control. 2015;81:83–92. doi: 10.1016/j.biocontrol.2014.11.011. [DOI] [Google Scholar]
- 13.Nadeem SM, Zahir ZA, Naveed M, Arshad M. Preliminary investigations on inducing salt tolerance in maize through inoculation with rhizobacteria containing ACC deaminase activity. Can J Microbiol. 2007;53(10):1141–1149. doi: 10.1139/W07-081. [DOI] [PubMed] [Google Scholar]
- 14.Saravanakumar D, Samiyappan R. ACC deaminase from Pseudomonas fluorescens mediated saline resistance in groundnut (Arachis hypogea) plants. J Appl Microbiol. 2007;102(5):1283–1292. doi: 10.1111/j.1365-2672.2006.03179.x. [DOI] [PubMed] [Google Scholar]
- 15.Wang Y, Ohara Y, Nakayashiki H, Tosa Y, Mayama S. Microarray analysis of the gene expression profile induced by the endophytic plant growth-promoting rhizobacteria, Pseudomonas fluorescens FPT9601-T5 in Arabidopsis. Mol Plant-Microbe Interact. 2005;18(5):385–396. doi: 10.1094/MPMI-18-0385. [DOI] [PubMed] [Google Scholar]
- 16.Islam T, Manna M, Kaul T, Pandey S, Reddy CS, Reddy MK. Genome-wide dissection of arabidopsis and rice for the identification and expression analysis of glutathione peroxidases reveals their stress-specific and overlapping response patterns. Plant Mol Biol Rep. 2015;33(5):1413–1427. doi: 10.1007/s11105-014-0846-6. [DOI] [Google Scholar]
- 17.Eisen MB, Spellman PT, Brown PO, Botstein D. Cluster analysis and display of genome-wide expression patterns. Proc Natl Acad Sci. 1998;95(25):14863–14868. doi: 10.1073/pnas.95.25.14863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Khare E, Kangmin K, Kui-Jae L. Rice OsPBL1 (ORYZA SATIVA ARABIDOPSIS PBS1-LIKE 1) enhanced defense of Arabidopsis against Pseudomonas syringae DC3000. Eur J Plant Pathol. 2016;146(4):901–910. doi: 10.1007/s10658-016-0968-9. [DOI] [Google Scholar]
- 19.Hwang I, Sheen J. Two-component circuitry in Arabidopsis cytokinin signal transduction. Nature. 2001;413(6854):383–389. doi: 10.1038/35096500. [DOI] [PubMed] [Google Scholar]
- 20.Clough SJ, Bent AF. Floral dip: a simplified method for agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1988;16(6):735–743. doi: 10.1046/j.1365-313x.1998.00343.x. [DOI] [PubMed] [Google Scholar]
- 21.Reddy CS, Vijayalakshmi M, Kaul T, Islam T, Reddy MK. Improving flavour and quality of tomatoes by expression of synthetic gene encoding sweet protein monellin. Mol Biotechnol. 2015;57(5):448–453. doi: 10.1007/s12033-015-9838-5. [DOI] [PubMed] [Google Scholar]
- 22.Paré PW, Zhang H, Aziz M, Xie X, Kim MS, Shen X, Zhang J. Biocommunication in soil microorganisms. Berlin, Heidelberg: Springer; 2011. Beneficial rhizobacteria induce plant growth: mapping signaling networks in Arabidopsis; pp. 403–412. [Google Scholar]
- 23.Park YS, Dutta S, Ann M, Raaijmakers JM, Park K. Promotion of plant growth by Pseudomonas fluorescens strain SS101 via novel volatile organic compounds. Biochem Biophys Res. 2015;461(2):361–365. doi: 10.1016/j.bbrc.2015.04.039. [DOI] [PubMed] [Google Scholar]
- 24.Mayak S, Tirosh T, Glick BR. Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol Biochem. 2004;42(6):565–572. doi: 10.1016/j.plaphy.2004.05.009. [DOI] [PubMed] [Google Scholar]
- 25.Mhadhbi H, Jebara M, Limam F, Aouani ME. Rhizobial strain involvement in plant growth, nodule protein composition and antioxidant enzyme activities of chickpea-rhizobia symbioses: modulation by salt stress. Plant Physiol Biochem. 2004;42(9):717–722. doi: 10.1016/j.plaphy.2004.07.005. [DOI] [PubMed] [Google Scholar]
- 26.Ibragimova MV, Rumyantseva ML, Onishchuk OP, Belova VS, Kurchak ON, Andronov EE, Dzyubenko NI, Simarov BV. Symbiosis between the root-nodule bacterium Sinorhizobiummeliloti and alfalfa (Medicago sativa) under salinization conditions. Microbiology. 2006;75(1):77–81. doi: 10.1134/S0026261706010140. [DOI] [PubMed] [Google Scholar]
- 27.Sahoo RK, Rani V, Tuteja N. Azotobacter vinelandii helps to combat chromium stress in rice by maintaining antioxidant machinery. 3 Biotech. 2021;11(6):1–11. doi: 10.1007/s13205-021-02835-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Afzal S, Begum N, Zhao H, Fang Z, Lou L, Cai Q. Influence of endophytic root bacteria on the growth cadmium tolerance and uptake of switchgrass (Panicumvirgatum L.) J Appl Microbiol. 2017;123(2):498–510. doi: 10.1111/jam.13505. [DOI] [PubMed] [Google Scholar]
- 29.Sinha S, Mukherjee SK. Pseudomonas aeruginosa KUCd1, a possible candidate for cadmium bioremediation. Braz J Microbiol. 2009;40(3):655–662. doi: 10.1590/S1517-83822009000300030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gutiérrez-Boem FH, Thomas GW. Phosphorus nutrition affects wheat response to water deficit. J Agron. 1998;90(2):166–171. doi: 10.2134/agronj1998.00021962009000020008x. [DOI] [Google Scholar]
- 31.Battie-Laclau P, Laclau JP, de Cassia PM, Arenque BC, Beri C, Mietton L, Muniz MR, Jordan-Meille L, Buckeridge MS, Nouvellon Y, Ranger J. Influence of potassium and sodium nutrition on leaf area components in Eucalyptus grandis trees. Plant and soil. 2013;371(1):19–35. doi: 10.1007/s11104-013-1663-7. [DOI] [Google Scholar]
- 32.Ma Q, Yue LJ, Zhang JL, Wu GQ, Bao AK, Wang SM. Sodium chloride improves photosynthesis and water status in the succulent xerophyte Zygophyllum xanthoxylum. Tree Physiol. 2012;32(1):4–13. doi: 10.1093/treephys/tpr098. [DOI] [PubMed] [Google Scholar]
- 33.Pishchik VN, Vorobyev NI, Chernyaeva II, Timofeeva SV, Kozhemyakov AP, Alexeev YV, Lukin SM. Experimental and mathematical simulation of plant growth promoting rhizobacteria and plant interaction under cadmium stress. Plant and Soil. 2002;243(2):173–186. doi: 10.1023/A:1019941525758. [DOI] [Google Scholar]
- 34.Won-Yong S, Martinoia E, Lee J, Kim D. A novel family of Cys-rich membrane proteins mediates cadmium resistance in arabidopsis 1. Plant Physiol. 2004;135(2):1027. doi: 10.1104/pp.103.037739. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Fig. S1. PCR2 gene regulatory network analyzed by co-expression analysis.
Additional file 2: Fig. S2. Expression analysis of Arabidopsis PCR2 gene responses todifferentstress conditions in the shoot and root tissues. Microarray expression data for AtPCR2 gene was retrieved from TAIR (ver 9) during various abiotic stresses, i.e. salt, drought, osmotic, cold, heat, oxidative, genotoxic, wounding and UV/B stress. The datasets obtained for various time points of stress, namely 0.5, 1, 3, 6, 12 and 24 h, were analyzed with respect to the control. The colour bar below represents relative expression values; wherein green represents lowest, black represents medium and red signifies highest expression levels. The hierarchical clustering is performed, and heat maps have been generated using TIGR MeV software package.
Additional file 3: Fig. S3. Controlled condition grown Col-0, T3-12, T3-18 Arabidopsis thaliana plants.
Additional file 4: Fig. S4. Various abiotic stress conditions imposed to control vs overexpressed lines and their root length and biomass data. Values are means and bars indicate SDs (n=6) statistically significant difference at P< 0.05 (t-test).
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.