

Abstract
Most adult organs contain regenerative stem cells, often organized in specific niches. Stem cell function is critical for tissue homeostasis and repair upon injury, and it is dependent on interactions with the niche. During ageing, stem cells decline in their regenerative potential and ability to give rise to differentiated cells in the tissue, which is associated with a deterioration of tissue integrity and health. Ageing-associated changes in regenerative tissue regions include defects in maintenance of stem cell quiescence, differentiation ability and bias, clonal expansion and infiltration of immune cells in the niche. In this Review, we discuss cellular and molecular mechanisms underlying ageing in the regenerative regions of different tissues as well as potential rejuvenation strategies. We focus primarily on brain, muscle and blood tissues, but also provide examples from other tissues, such as skin and intestine. We describe the complex interactions between different cell types, non-cell-autonomous mechanisms between ageing niches and stem cells, and the influence of systemic factors. We also compare different interventions for the rejuvenation of old regenerative regions. Future outlooks in the field of stem cell ageing are discussed, including strategies to counter ageing and age-dependent disease.
Ageing is characterized by a decline in all organ function and a striking increase in multi-organ diseases1. Different organs and tissues in the adult body — brain, heart, blood, liver, gut, muscle and so on — have vastly divergent functions and contain many diverse cell types. To understand whether organ and cell function can be restored or at least improved in old individuals, it is essential to uncover the mechanisms underlying ageing-associated deterioration of the different organs and cell types they contain.
The cell types that build each organ are subject to different turnover rates. Some differentiated cell types can last a lifetime, such as neurons in the brain or cardiomyocytes in the heart. Other differentiated cell types are continuously lost and replenished quickly, such as red blood cells, monocytes and epithelial cells of the skin and intestine2. Cells with rapid turnover rates are largely replaced by the progeny of very active stem cells (often referred to as ‘adult stem cells’, ‘tissue stem cells’ or ‘somatic stem cells’). For example, haematopoietic stem cells (HSCs) located in the bone marrow can generate all cells in the blood, and intestinal stem cells (ISCs) located in crypts of the intestine produce epithelial cells lining the gut (FIG. 1a). By contrast, cells with a low or very low turnover rate (neurons and muscle fibres) are regenerated with smaller populations of stem cells (FIG. 1a). In addition to somatic stem cells, the gonad, or at least the male gonad in mammals, contains germ line stem cells that have the unique property of giving rise to gametes. Understanding how stem cells age compared with differentiated progeny and whether there are common mechanisms of ageing in different types of stem cells, with high-turnover and low-turnover progeny, is pivotal to counter tissue ageing.
Fig. 1 |. Organs with stem cells and stem cell niches.
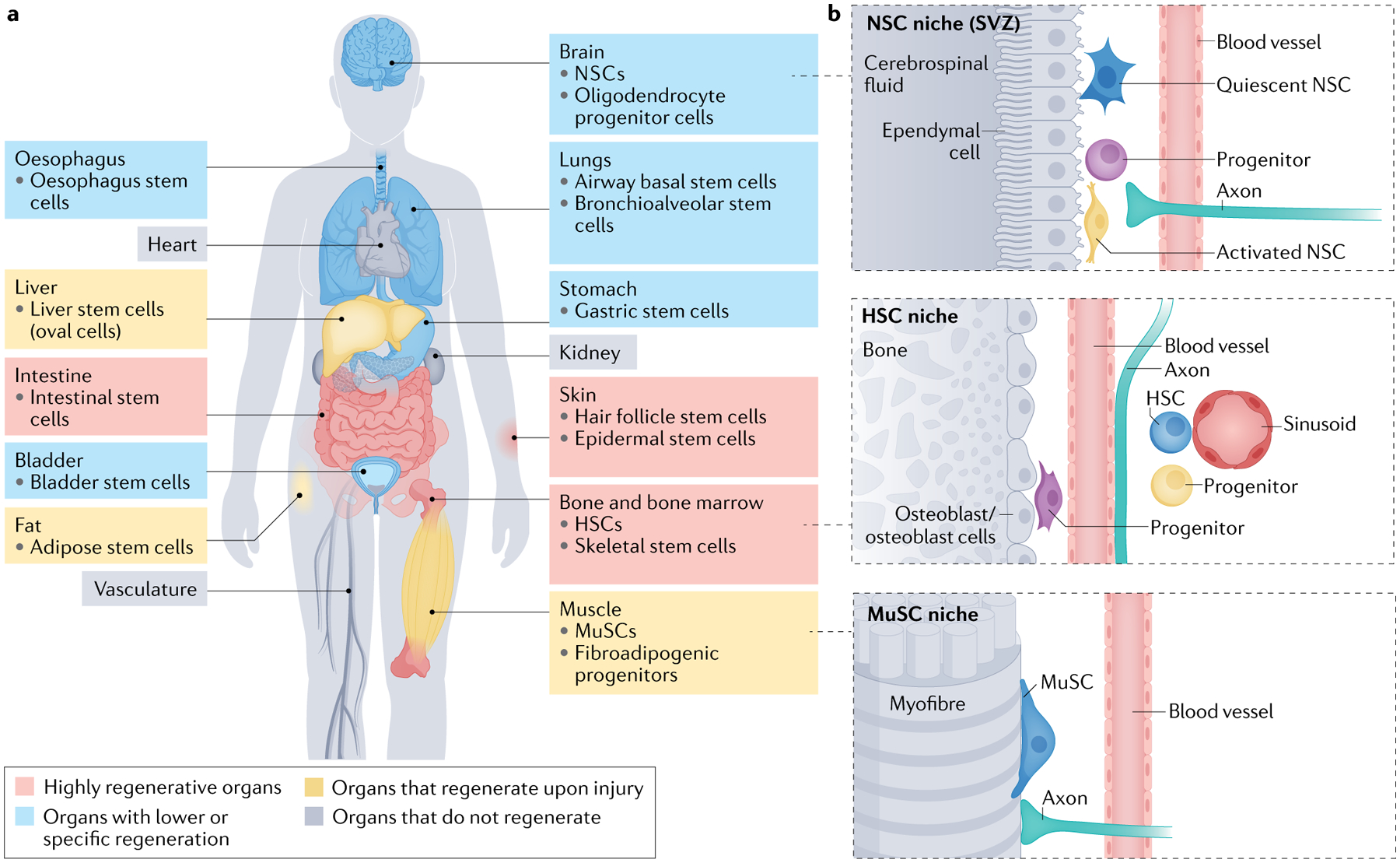
a | Several organs in humans and other mammals contain stem cells and stem cell niches. The figure depicts organs that are highly regenerative, with very active stem cells; organs that have restricted regeneration, sometimes with stem cells restricted to specialized regions; and organs that are regenerative upon injury, with dormant stem cells that are mostly activated in response to injury. Some organs do not regenerate and have no stem cells (or the presence of stem cells is controversial). b | Organization of adult stem cell niches in the brain, blood and muscle. Examples of markers for adult neural stem cells (NSCs) include SOX2, Nestin, glial fibrillary acid protein (GFAP) and CD133. Examples of markers for adult haematopoietic stem cells (HSCs) in humans include the presence of CD34 and the absence of CD38. Example of markers for muscle stem cells (MuSCs) include PAX7, CD34 and CD56 (human). SVZ, subventricular zone.
Stem cells have a crucial role for tissue homeostasis, and they are maintained and regulated by their micro-environment (the ‘niche’), which forms a regenerative unit or region (FIG. 1b). These regenerative regions can drastically respond to environmental stimuli such as injury. Their function is also affected by environmental stimuli known to impact ageing, including diet and exercise. Adult stem cells originate from their embryonic counterparts. Whereas embryonic stem cells proliferate actively, a crucial characteristic of most adult stem cells is their quiescent state. Therefore, quiescent stem cells constitute a reservoir of cells that can be induced to proliferate and that could be tapped into for regenerative purposes and repair during ageing.
Several reviews have discussed stem cell ageing in species ranging from invertebrates to mammals3–5. In this Review, we focus on mammalian somatic stem cell ageing. We discuss recent work that has increased our understanding of ageing mechanisms in the brain, muscle and blood — tissues in which much stem cell ageing research has been done — but we also provide some examples from other tissues, including skin and intestine. We describe ageing-associated changes in somatic stem cells and their niches. We specify which niche is being discussed for each mechanism, and we highlight similarities and differences between stem cells in diverse tissues. Furthermore, we analyse non-cell-autonomous mechanisms that link stem cells and their proximal and distal environments, and how these extrinsic mechanisms are connected to cell-intrinsic changes. An important goal of this Review is to discuss how different tissue stem cells respond to a variety of rejuvenation strategies, such as dietary restriction, exercise, young blood factors and cellular reprogramming.
Defects in stem cells during ageing
Ageing leads to a decline in tissue stem cell function3–5. The number and function of most somatic stem cells decline with ageing, with the exception of HSCs. The number of HSCs (defined by phenotype) increases but their function decreases with age. The age-dependent decline in stem cell function is associated with a deterioration in tissue function and repair. Technological advances — single-cell transcriptomics, lineage tracing and clonal analysis — have enabled more unbiased and quantitative analysis of stem cell changes during ageing. These studies highlight shared defects in stem cell ageing across tissues but also reveal some differences among various tissue stem cells.
Reduced ability to produce committed progeny.
A main feature of ageing is a reduced ability of somatic stem cells to produce new differentiated cells that are essential for homeostasis and repair in brain, blood, muscle, fat, skin, lung and so on. In recent years, cell-based approaches, including single-cell RNA sequencing (RNA-seq) and lineage tracing, have highlighted stem cell decline during ageing6–16. For example, in the mouse brain, single-cell RNA-seq analyses indicate that the number of activated neural stem cells (NSCs) and neuroblasts (progeny) decreases with ageing in both the subventricular zone (SVZ) NSC niche and the hippocampal NSC niche6–9 — the main locations of NSCs. In humans, the timing of decline in the ability of NSCs to generate new neurons (neurogenesis) in the hippocampal niche is still debated. Some immunostaining studies show that hippocampal neurogenesis declines relatively early in adolescence17,18, consistent with what is observed in single-nucleus RNA-seq19. However, other studies indicate that neurogenesis in the hippocampus and other regions declines much later in the human brain20–22. In blood, lineage tracing, single-cell RNA-seq and large-scale clonal analysis studies all indicate that old HSCs generate fewer lymphoid cells and are skewed towards the myeloid lineage23–26. In muscle, single-cell RNA-seq combined with machine learning modelling revealed that the activation and differentiation trajectories of old and young muscle stem cells (MuSCs) differ13, with slower kinetics in old MuSCs. Together, these data corroborate aberrant transition to progeny during ageing (FIG. 2).
Fig. 2 |. Changes in stem cells and their niches during ageing.
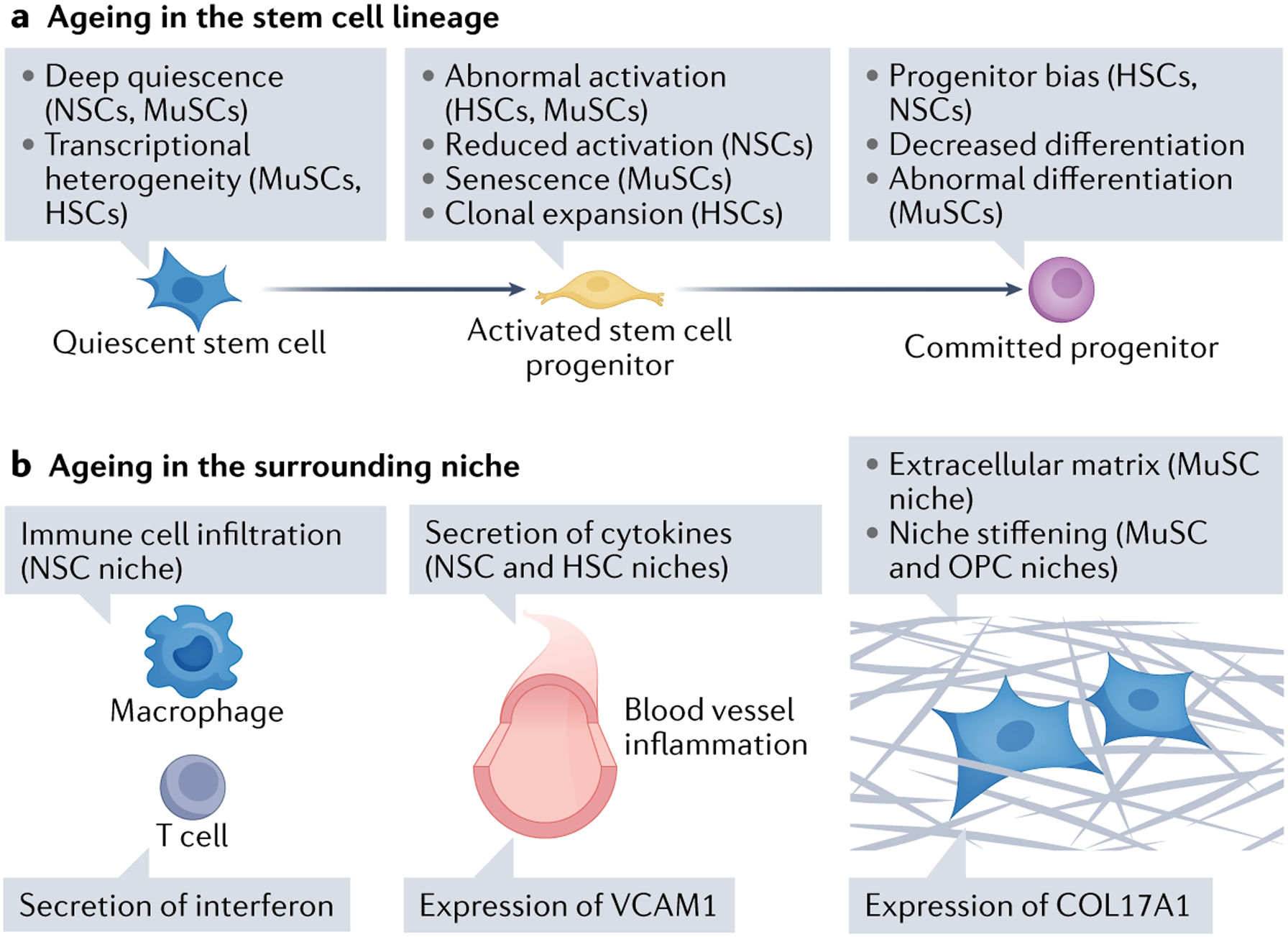
a | Ageing changes in the stem cell lineage. During ageing, subpopulations of stem cells can enter deep quiescence, and their ability to activate is reduced. Other subpopulations of stem cells can exhibit aberrant activation, reduced activation and clonal expansion (haematopoietic stem cells (HSCs)). There is also a bias in progenitor commitment and abnormal differentiation during ageing. b | Ageing changes in the niche. Ageing leads to immune cell infiltration, inflammation, secretion of cytokines (for example, interferon), expression of cell adhesion molecules (vascular cell adhesion molecule 1 (VCAM1)) by blood vessel cells, extracellular matrix secretion (collagen type XVII α1 chain (COL17A1)) and increased stiffening of the niche. MuSC, muscle stem cell; NSC, neural stem cell.
Heterogeneity of quiescent stem cell pools with ageing.
Another hallmark of ageing is increased stem cell heterogeneity, notably increased heterogeneity in the dormant (quiescent) subpopulation of stem cells in aged individuals (FIG. 2). In the brain, old age is accompanied by an increase in the number of quiescent NSCs and a decrease in their ability to activate6–8. Lineage tracing uncovered a subset of early-declining NSCs in the hippocampal neurogenic niche9. Such stem cell heterogeneity during ageing may be due to a decrease in symmetrical divisions and an increase in asymmetrical divisions in old NSCs27,28. With increasing age, a subset of NSCs enter deep quiescence instead of a shallower, reversible quiescence, which is characteristic of young NSCs29. Interestingly, a small pool of quiescent NSCs (maybe overlapping with those in deep quiescence) appears to be protected from ageing defects8.
In muscle, MuSCs also exist as a heterogeneous quiescent population in adult mice30–33, with changes in heterogeneity with ageing32,34,35. MuSCs enter a deeper state of quiescence with ageing as reflected by their slower kinetics of activation34. A subpopulation of MuSCs (identified by higher levels of CD34) appears to be preserved during ageing32. Another subset of MuSCs (identified by inducible Mx1–Cre lineage tracing) is particularly resistant to stress36, perhaps also representing a population that is protected from ageing37. Likewise, in bone marrow from old mice, a pool of quiescent HSCs located close to sinusoids (FIG. 1b) may be protected from some ageing effects38.
Abnormal fate with ageing.
The fate and behaviour of subsets of stem cells can become abnormal with ageing. Stem cells can enter a state of senescence or excessive activation or can undergo aberrant differentiation (FIG. 2). For example, some old MuSCs become senescent, a state characterized by irreversible cell cycle arrest and secretion of cytokines and other proteins (senescence-associated secreted phenotype)39–43. Other stem cell subpopulations lose quiescence and become abnormally activated, particularly in blood and muscle12,35,44. This abnormal activation is accompanied by a bias in specific progenitors. For example, subpopulations of MuSCs become more ‘primed’ (that is, poised towards a myogenic fate) with ageing32, whereas others adopt a more fibrogenic state16. Similarly, some HSCs become more poised towards myeloid progenitors during ageing26. In the brain, old progenitors exhibit reduced differentiation dynamics during ageing45, and their fate potential is predominantly skewed towards astrocytes46.
Thus, stem cells pools are heterogeneous, and different subpopulations arise with increasing age (FIG. 2). It will be important to further finely map stem cell subpopulations in different tissues in old individuals. Moreover, the decline of stem cell function is not a continuous and monotonous process throughout the lifespan — it may instead reflect different phases and demands in life. For example, during the transition to adulthood (when mating and pregnancy occur in females), the pressure for mechanisms to maintain a large pool of stem cells is strong, whereas later in adulthood, this pressure diminishes and a new equilibrium is reached. Consistent with this possibility, machine learning models of HSC division as a function of increasing age suggested that HSC division is intrinsically regulated in young adults and old adults, but is more extrinsically regulated in middle age47. Understanding stem cell heterogeneity during ageing will be critical to identify new ways to tap into pristine reservoirs of stem cells for late-life tissue regeneration.
Somatic mutations and clonal competition in stem cell pools with ageing.
Heterogeneity in stem cell pools leads to competition among stem cells, in at least some tissues (FIG. 2). In humans, this manifests itself as ‘clonal haematopoiesis’, where one or a few HSC clones disproportionately contribute to blood production over time48,49. This phenomenon is ubiquitous in aged individuals, and it is probably the result of variant stem cells having a survival and proliferation advantage. Stem cells that are expanded in clonal haematopoiesis often have somatic mutations in genes encoding epigenetic regulators such DNA methyltransferase 3A (DNMT3A)50,51. Broadly, even partial loss of function of these epigenetic regulators biases the stem cells towards self-renewal and reduced differentiation50,51. Clonal haematopoiesis is associated with a tenfold increased risk of malignancy development, thus underlying the increase in the incidence of blood cancer with ageing in humans.
Clonal expansion has also been found in other human tissues, including skin, colon and oesophagus52–54, and is thought to be a normal, possibly ubiquitous, process that is associated with ageing in all tissues, in particular those that experience significant cellular turnover. In muscle, clonal expansion of MuSCs has been reported during tissue regeneration in response to acute injury55, but has not yet been thoroughly examined during ageing. The degree to which variant clones expand owing to intrinsic differences driven by acquired somatic mutations56 versus environmental influences such as diet or infection57–59 remains to be determined. In humans, several stem cell types, including those of the colon and liver, have been shown to acquire somatic mutations at a steady rate during ageing60; this feature is likely universal. Animal models are needed to study age-associated clonal expansion. While clonal expansion during normal ageing has not yet been observed in mice, it has been reported in mouse models for disease61 and in physiologically aged macaques62. Ultimately, animal models will be critical for understanding the mechanisms of clonal expansion during ageing and developing interventions to prevent transformation to cancer.
Accumulation of ageing markers.
During ageing, stem cells also accumulate specific ageing-associated transcriptomic and proteomic signatures. Changes in transcriptional profiles may reflect the accumulation of cellular damage and ageing markers. Interestingly, activated NSCs exhibit fewer overall transcriptomic changes with ageing than quiescent NSCs6–9,63,64, perhaps because activated NSCs are recently generated and do not have the time to accumulate damage, or because they ‘dilute’ the damage as they proliferate. In muscle, however, activated MuSCs exhibit more transcriptional changes than quiescent MuSCs13. This difference between NSCs and MuSCs is not understood but could reflect differences in subpopulations of stem cells during ageing. Transcriptional changes in old MuSCs involve metabolic pathways and genes that regulate chromatin accessibility12,34,65. Moreover, proteomic studies in old MuSCs reveal changes in integrin signalling during ageing66. In muscle, single-cell multi-omic studies reveal that ageing is accompanied by transcriptional heterogeneity, especially in genes that impact stem cell–niche interactions67. In the skin, stem cells also exhibit transcriptional changes with increased expression of extracellular matrix (ECM) proteins11. In the bone marrow, ageing HSCs display increased transcriptional heterogeneity, but a set of markers is consistently upregulated with ageing (for example, the cell adhesion molecule P-selectin). Upregulation of cell adhesion markers in HSCs may be a direct indicator of their inflammatory state and may correlate with the bias towards myeloid cell production68.
Thus, a common feature of ageing stem cells is a reduction in their ability to give rise to a functional progeny. However, the specific changes in proliferative and differentiation capacity, the timing of decline and the type of molecular defects with ageing may differ depending on tissue and age time points.
Defects in stem cell niches during ageing
Recent technological advances have provided a systematic understanding of age-related defects not only in the stem cells themselves but also in niche cells (other than the stem cells) and in the niche milieu and its physical properties.
Age-dependent changes in niche cells during ageing.
Single-cell RNA-seq technologies have enabled the analysis of stem cells in the context of the entire regenerative niche during ageing. These types of analysis have revealed profound changes in the composition of the niche (that is, the proportion of different cell types other than the stem cells themselves) as well as transcriptomic shifts in specific niche cells during ageing (FIG. 2). For example, in the SVZ neurogenic niche, RNA-seq studies uncovered the presence of 11 cell types, including endothelial cells, microglia, oligodendrocytes and pericytes7. Ageing is accompanied by strong transcriptional changes in niche cells such as endothelial cells (which form blood vessels) and microglia (the immune cells of the brain), consistent with inflammation7. In muscle, quantitative proteomics studies have also identified changes in the tissue, notably in the ECM composition66. Niche cells in many tissues also experience drastic transcriptomic changes with ageing69, which could indirectly impair stem cell function.
The infiltration of different types of immune cells in stem cell niches is a key hallmark of ageing. Indeed, single-cell RNA-seq indicated that ageing is accompanied by infiltration of clonally expanded cytotoxic CD8+ T cells in old neurogenic niches7. Immunofluorescence staining confirmed the infiltration of T cells in the regenerative regions of aged mice and humans7,70. The infiltration of clonally expanded T cells and B cells is also observed in the brain and other tissues with ageing in mice and humans69,71–73, and it is accentuated in age-related diseases74,75. Conversely, other types of immune cells, such as regulatory T cells, are reduced in number in aged skin11, and this might contribute to the unbalance in tissue regulation for this regenerative niche. In the future, it will be important to systematically identify the immune cells present in stem cell niches and determine whether their clonal expansion is due to the presence of age-specific antigens.
These ageing-dependent changes in the composition of the stem cell niche and infiltration of immune cells influence the extracellular milieu, but also impact the ageing of stem cells in a non-cell-autonomous manner (see later).
Age-dependent changes in soluble factors and the stem cell milieu.
The milieu that surrounds stem cells also changes with ageing, with differences in many soluble factors (cytokines, growth factors, metabolites and nutrients) (FIG. 2). For example, owing to the infiltration of immune cells, the activation of macrophages and microglia, and the presence of senescent cells, the NSC niche milieu likely becomes extremely inflammatory during ageing6–8. inflammatory cytokines, including interferon, and inflammatory cytokines (interleukin-33 (IL-33) and CXCL10), are upregulated during ageing in the NSC niche7,8. Inflammatory cytokines, including TNF76, transforming growth factor-β1 (TGF1β)77, IL-6 (REF.78) and IL-1β (REF.79), have also been found to be upregulated in aged bone marrow (HSC niche).
Age-dependent changes in the biophysical properties of the niche.
Age-dependent changes in the biophysical properties of the niche have been uncovered (FIG. 2). Stem cell niches become stiffer during ageing, and they exhibit drastic changes in ECM composition. Increased stiffness was observed in ageing brain tissue80. In the bone marrow, the lymphoid progenitor cell niche contains mechanosensitive cells also involved in osteogenesis. Reduction in physical activity during ageing may be transmitted via these cells, contributing to a loss of lymphoid progenitors over time81. In skeletal muscle, ECM components also change with ageing66. Decreased collagen tortuosity and increased stiffness in the muscle niche are associated with aberrant MuSC differentiation82. In the skin, ageing is accompanied by increased ECM proteins in the bulge — a crucial change with ageing11. The ageing skin is characterized by niche stiffening and hair shaft miniaturization, which are accompanied by defects in hair follicle stem cells (HFSCs)83,84. Hence, the general milieu of the niche may become more inflamed and stiffer, which may contribute to stem cell defects.
Molecular mechanisms of stem cell ageing
In this section, we discuss mechanisms that are most likely to mediate the changes in cell–cell interactions and systemic factors in stem cell ageing and rejuvenation. These include signalling pathways and transcription factors, epigenomic mechanisms involving chromatin modulation, protein homeostasis and metabolism.
Signalling pathways and transcription factors.
A key mechanism by which cell–cell interactions and systemic factors influence stem cell function during ageing includes signalling pathways and transcription factor regulation (FIG. 3). For example, FOXO transcription factors integrate growth factors and stress signal transduction pathways, and may represent a ‘hub’ for the action of various systemic factors in stem cells in ageing brain, muscle, blood and intestine32,85–90. FOXO factors are activated by the lack of systemic or paracrine factors (for example, insulin and insulin-like growth factor 1 (IGF1)). Consistent with previous reports of the role of FOXO factors in HSC, MuSC and NSC quiescence85–90, constitutive deletion of three of the FOXO genes (FOXO1, FOXO3 and FOXO4) in the MuSC lineage during development results in the impairment of postnatal establishment of the MuSC population and a more rapid MuSC depletion with age32. FOXO factors may be critical in distinguishing different states of quiescence, perhaps by connecting quiescence to clearing mechanisms, including autophagy91,92. FOXO factors are also activated by oxidative stress and can induce interferon production in NSCs via the STING pathway93. Thus, FOXO factors may integrate systemic or paracrine signals to orchestrate transcriptional changes that maintain stem cell quiescence during ageing and regulate inflammation.
Fig. 3 |. Cell-autonomous and non-cell-autonomous mechanisms of stem cell ageing.
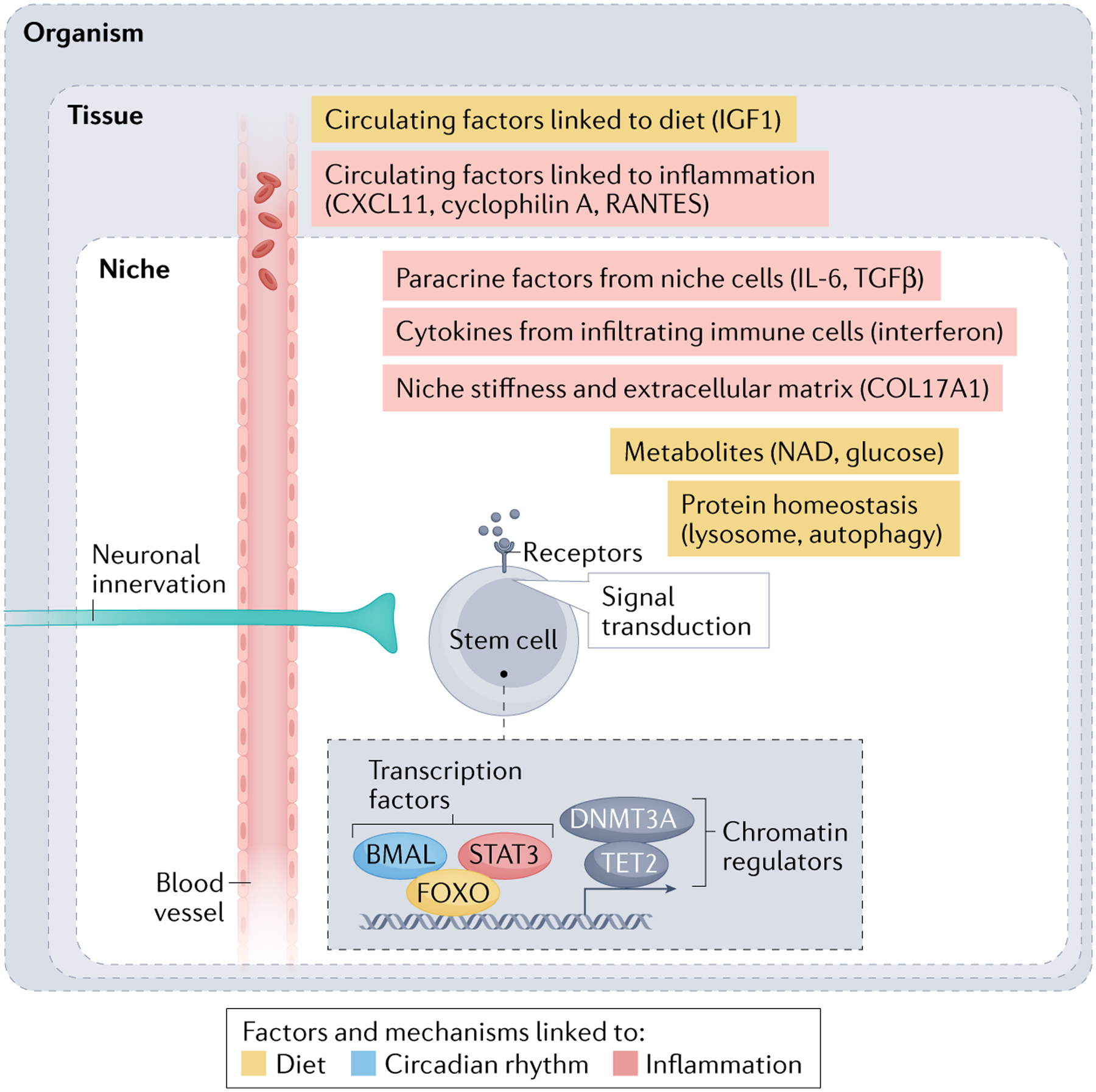
During ageing, stem cells undergo cell-autonomous changes, including modulations of signalling cascades, transcription factors (BMAL, STAT3 and FOXO) and chromatin regulators (DNA methyltransferase 3A (DNMT3A) and TET2). Many of these cell-autonomous changes are affected by non-cell-autonomous mechanisms originating from various sources. Stem cells experience non-cell-autonomous influences from proximal niche cells, involving growth factors such as transforming growth factor-β (TGFβ) and cytokines such as interleukin-6 (IL-6) secreted by niche cells, cytokines such as interferon secreted by infiltrated immune cells, and extracellular matrix proteins (collagen type XVII α1 chain (COL17A1)) secreted by niche cells. Stem cells are also modulated by long-range factors such as the nervous system or blood (with circulating growth factors such as insulin-like growth factor 1 (IGF1) and cytokines and chemokines (CXCL11, RANTES and cyclophilin A), which are themselves under the influence of the environment (diet and circadian rhythm). Yellow indicates factors and mechanisms linked to the diet (for example, nicotinamide adenine dinucleotide (NAD)). BMAL (shown in blue) is a transcription factor linked to circadian rhythm. Red indicates factors and mechanisms linked to inflammation.
Other signalling pathways and transcription factors implicated in inflammation signalling may also function downstream of systemic factors in stem cell niches during ageing. STAT3 — a transcription factor activated by inflammatory cytokines — is at least in part responsible for NSC differentiation being biased towards glial cells (rather than neurons) during ageing46,94. STAT3 is modified by O-linked β-N-acetylglucosamine, and this post-translational modification is lost during ageing, contributing to the ageing-dependent bias towards glia46.
Signalling pathways and transcription factors that respond to oxidative or genomic stress (for example, DNA damage) could also impact stem cells during ageing. For example, old MuSCs exhibit increased cell death owing to a reduction in activity of the transcription factor p53, in a manner that is dependent on activation of the Notch pathway95. Cell death is also increased in ageing HSCs, and this is dependent on the DNA-damage signalling pathway ATM96. Moreover, the zinc-finger transcription factor SLUG can activate a DNA damage response in epithelial stem cells during ageing71. Thus, these transcription factors are good candidates for mediating stem cell function changes during ageing.
Identifying transcription factors that could be manipulated to reprogramme stem cells into younger ones is a topic of active investigation. However, the transcription factors that induce stem cell rejuvenation remain largely unknown.
Epigenomic mechanisms.
DNA methylation, histone modifications and the enzymes that regulate them — all of which can induce changes in chromatin accessibility and gene expression — could respond to ageing-associated changes in cell–cell interactions and systemic factors. Several chromatin-modifying enzymes have been implicated in stem cell regulation during ageing (FIG. 3). In the brain, expression of TET2, the enzyme responsible for the oxidation of 5-methylcytosine to 5-hydroxymethylcytosine, decreases during ageing in the hippocampal neurogenic niche97, and Tet2 knockout mimics aspects of ageing, whereas its overexpression prevents the decline in neuronal differentiation during ageing97. In the human haematopoietic system, somatic mutations in epigenetic regulators, including DNMT3A and TET2, have consistently been associated with clonal expansion with ageing (reviewed in48). In muscle, DNA methylation changes with ageing in MuSCs, with increased heterogeneity (accompanied by transcriptional heterogeneity), suggesting an epigenetic drift with ageing33,67.
Many aspects of chromatin state and regulation change during ageing. A comprehensive analysis of murine HSC ageing has found increased chromatin accessibility and an overall increase in transcription68. Primary cultures of old quiescent NSCs also exhibit increased chromatin accessibility at genes involved in quiescence, a potential mechanism underlying deep quiescence during ageing98. Histone marks also change with ageing. In old quiescent MuSCs, the repressive mark trimethylated histone H3 at lysine 27 (H3K27me3) increases, whereas the activating mark trimethylated histone H3 at lysine 4 (H3K4me3) decreases65. By contrast, in old HSCs, H3K4me3 increases across genes regulating self-renewal, in line with functional changes99. Consistently, the H3K27me3 demethylase UTX helps maintain a youthful phenotype in HSCs, with UTX-deficient HSCs acquiring an early-ageing phenotype100. In addition, the lysine acetyltransferase KAT6B, whose expression declines with age, is important for maintaining a youthful HSC differentiation profile101. The global disruption of heterochromatin by deletion of both H3K9me3 methyltransferases SUV39H1 and SUV39H2 has a ‘pro-ageing’ effect on HSCs and the immune system102. Finally, expression of lamin B1, a key component of the nuclear architecture involved in epigenomic maintenance103, declines in old NSCs, and restoration of lamin B1 expression is sufficient to increase old NSC proliferation and neurogenesis104. It will be interesting to determine whether the three-dimensional chromatin structure, which is particularly important for stem and progenitor states105, is affected by ageing.
Epigenomic changes have the potential to connect stem cell changes to external stimuli. For example, in HFSCs, ageing is associated with decreased chromatin accessibility, notably silencing of ‘bivalent’ promoters (that is, marked by both H3K27me3 and H3K4me3)84. These age-dependent chromatin changes in HFSCs are affected by stiffening of the niche84, and could mediate the impact of external and biophysical properties on stem cells during ageing.
Together, these results illustrate the critical nature of chromatin regulators for long-lasting changes during ageing, and their potential to integrate extrinsic stimuli. These findings also highlight the need for a better understanding of the reversible nature of chromatin states during ageing.
Protein homeostasis.
Protein homeostasis is influenced by the external milieu, and could connect cell–cell interaction and systemic factors to key cellular function for stem cells (FIG. 3). For example, lysosomes are large and abundant in quiescent NSCs64. Lysosome clearing properties decline with age, resulting in protein aggregate accumulation64. Interestingly, age-dependent NSC decline can be reversed by expression of the master transcription factor for lysosomes TFEB and by nutrient deprivation64. These results suggest that inducing the lysosome and autophagy pathways can boost old NSC activation out of quiescence64. Autophagy, which helps degrade large aggregates or damaged organelles, is critical to maintain HSCs during ageing106. HSC maintenance by autophagy is mediated at least in part by a membrane ubiquitin ligase called ‘MAEA’ (macrophage–erythroblast attacher), although it is not yet clear whether this mechanism contributes to HSC ageing107. In MuSCs, autophagy is essential for the maintenance of MuSC quiescence, via the process of mitophagy (degradation of damaged mitochondria), and this declines with ageing108. Autophagy is also required for activation out of quiescence109, and defects in autophagic activity with ageing lead to increased cell death during activation110. Finally, a particular form of autophagy — chaperone-mediated autophagy — is essential to preserve HSCs throughout the lifespan, not only by clearing proteins but also by altering lipid metabolism111. Hence, lysosomes and autophagy, which clear damaged proteins and organelles, are both critical to maintain stem cells and ameliorate their function during ageing.
Other protein homeostasis mechanisms (for example, protein chaperones) play a role in stem cell ageing and could respond to changes in the external milieu. Indeed, the protein chaperone network is entirely rewired when NSCs give rise to differentiated progeny, and this rewiring is impaired during ageing112. The transcription factor HSF1, which promotes protein homeostasis by inducing many chaperone proteins, is necessary for protein homeostasis in HSCs during ageing113. It will be interesting to test how the various components of the protein homeostasis machinery (for example, proteasome, lysosome, autophagy and chaperones) respond to cell–cell interaction and environmental stimuli that are prevalent during ageing, such as inflammation.
Metabolism.
Various metabolites (for example, nicotinamide adenine dinucleotide (NAD), adenosine triphosphate, glucose, amino acids and lipids), and the enzymes that use or regulate them, are pivotal for the integration of external factors and the regulation of stem cell function114,115.
The sirtuin family of NAD-dependent protein deacetylases plays an important role in connecting metabolic state to stem cell function. Sirtuin activity and NAD levels both decline with age116,117. SIRT1, a member of the sirtuin family, protects MuSC progeny from premature differentiation by regulating histone acetylation118, and this protective effect could deteriorate with ageing. SIRT1 was also found to promote autophagy in quiescent MuSCs, and in this case, a loss of SIRT1 leads to a delay in MuSC activation out of quiescence — similar to what is observed in aged muscles109. Thus, SIRT1 appears to regulate multiple processes along the MuSC lineage, and the age-dependent decline in SIRT1 levels could have pleiotropic effects that contribute to impaired muscle regeneration with ageing. Additionally, SIRT2, another member of the sirtuin family, is necessary to prevent ageing in HSCs, and can inhibit the inflammation (inflammasome) response that occurs in old HSCs119. Finally, NAD precursors such as nicotinamide riboside can restore ageing defects in ISCs120, as well as in MuSCs, NSCs and melanocyte stem cells39.
Glycolysis and mitochondrial function also decline during ageing. Aged MuSCs and HSCs have increased glucose use and higher levels of glycolysis121–123. Quiescent MuSCs exhibit reduced energetic demands and perturbed oxidative phosphorylation during ageing122. Glucose metabolism is critical for MuSCs and influences histone acetylation124. Knockout of Tfam (a central transcription factor for mitochondria) mimics ageing in NSCs, whereas enhancement of mitochondrial function by piracetam boosts neurogenesis in aged mice125. Thus, changes in glucose metabolism and mitochondrial function could be critical for the integration of external longevity signals, notably diet.
Other metabolites, such as fumarate, succinate and α-ketoglutarate, as well as oxygen levels, could play a key role in improving old tissue stem cell function and connecting stem cells with changes in their environment114,115. For example, fumarate affects young HSC function126, and might restore defects in aged HSCs. Experiments are needed to understand how metabolites and enzymes that use or regulate them are themselves affected by extrinsic factors during ageing and how this impacts stem cell function.
Overall, it will be important to compare in a systematic manner how different pathways (protein homeostasis, metabolism, chromatin and transcription factors) impact stem cells and determine how these pathways may interact in response to external stimuli to modulate stem cell ageing and rejuvenation.
Interactions within the niche
The ageing process in stem cells involves non-cell-autonomous mechanisms (FIG. 3). Here we focus on paracrine factors secreted from other stem cells, niche cells, endothelial cells, immune cells and factors present in the ECM.
Role of paracrine factors produced by other stem cells and niche cells.
Paracrine signalling is altered during ageing, and this change contributes to stem cell defects in a non-cell-autonomous manner (FIG. 3). One stem or progenitor cell type can influence another stem or progenitor cell type within the same tissue via alteration of paracrine signalling. For example, in muscle, fibroadipogenic progenitors are cells of mesenchymal origin that are critical for MuSC support and expansion10,127. During ageing, the number the of fibroadipogenic progenitors declines, and so does their ability to produce paracrine factors such as WNT1-inducible signalling pathway protein 1 (WISP1), a non-structural protein of the ECM10. This leads to diminished MuSC support and contributes to the decline in MuSC function during ageing10. Fibroadipogenic progenitors can also alter many proteins in the ECM, including the protein SMOC2, which can in turn affect muscle regeneration via an integrin–MAPK signalling pathway66. In the bone marrow, skeletal stem cells become inflammatory with ageing, which impedes HSC function in old mice128. Cell composition of the bone marrow shifts with ageing, with the proportion of adipocytes markedly increasing, creating an inflammatory environment through secretion of soluble factors129. Thus, lack of supporting paracrine factors and enhanced inflammatory signalling from other progenitor/stem cells during ageing can promote the demise of stem cells and the deterioration of the tissue.
Niche cells can also produce factors that regulate stem cell function during ageing (FIG. 3). For example, MuSCs normally receive quiescence-promoting signals (for example, oncostatin M) from skeletal myofibres35,130,131, which are part of their niche. During ageing, skeletal myofibres start secreting other factors, such as FGF2, which induce MuSC activation and ultimately decrease their self-renewing properties35. In the lung, airway basal stem cells are responsible for glandular-like epithelial invaginations that appear de novo during ageing, and these new structures are dependent on WNT ligands14. In the intestine, old Paneth cells, which are niche cells, can secrete increased amounts of NOTUM, which in turn attenuates WNT signalling in ISCs and contributes to stem cell dysfunction and intestinal decline132. In the bone marrow, TNF (likely produced by niche cells) induces the expression of IL-27 receptor, and ablation of IL-27 receptor blunts the negative impact of ageing on HSCs76. Reduced levels of IGF1 in the bone marrow environment (probably because of reduced production by old niche cells) also promote HSC ageing133, although low IGF1 levels lead to increased longevity at the organismal level. Reduced production of IGF1 by stromal fibroblasts can also impair skin stem cells during ageing134. Thus, the deterioration and inflammation of niche cells and the lack of production of growth factors negatively impact stem cell function during ageing.
Together, these observations indicate that non-cell-autonomous mechanisms mediated by paracrine signalling emanating from adjacent cells have important roles in stem cell ageing. Understanding the spatial localization and inflammation status of different stem or progenitor cells within regenerative regions with respect to niche cells will be instrumental to better tease out the paracrine influences on stem cells during ageing.
Role of soluble factors secreted by other lineages — endothelial and immune cells.
Factors (for example, soluble cytokines) originating from cells from a different lineage (for example, endothelial cells or immune cells) also contribute to stem cell and tissue defects during ageing (FIG. 3).
Blood vessels are integral components of regenerative niches, and blood vessel cells (endothelial cells) and their secreted factors can impact stem cell proliferation during ageing. In the SVZ neurogenic niche, stromal cell-derived factor 1 (SDF1) is secreted from a subpopulation of blood vessels (capillaries) with ageing, and SDF1 from capillaries is associated with NSC proliferation135. During ageing, SDF1 signalling (via CXCR4 receptors in NSCs) is reduced, and this decline is in part responsible for impaired NSC proliferation during ageing135. Similarly, brain endothelial cells express vascular cell adhesion molecule 1 (VCAM1), and VCAM1 expression is upregulated with ageing and contributes to the observed increase of VCAM1 levels in old plasma136. VCAM1 facilitates vascular system–immune system interaction (for example, entry of T cells and macrophages into the brain). Although VCAM1 does not affect NSCs directly, it negatively impacts old proliferating cells (for example, neural progenitor cells). Interestingly, this decline in neural progenitors and defects in cognition can both be blocked by anti-VCAM1 antibodies or genetic ablation of Vcam1 in brain endothelial cells136. Hence, endothelial cells that form capillaries or blood vessels in the niche can negatively impact progenitor function and cognitive function during ageing.
Immune cells infiltrate old regenerative niches and can affect stem cells by releasing inflammatory cytokines, including interferons. In the brain SVZ neurogenic region, T cells can secrete interferon-γ (IFNγ), and IFNγ signalling is increased in the entire neurogenic region. NSCs respond to IFNγ by decreased proliferation, which is restored (at least in vitro) by blocking antibodies to IFNγ7. Interferon signalling is also implicated in maintaining stem cell quiescence in the SVZ during ageing8. Although IFNγ can also have positive effects, IFNα has an overall negative impact on the brain137. Chronic inflammation and IFNα convert αNSCs into ΩNSCs, which are more quiescent138. In the bone marrow, megakaryocytes appear to show increased production of the inflammatory cytokine IL-6 (REF.78), which is modulated by both β-adrenergic signalling (from sympathetic nerves) and inducible nitric oxide synthase signalling78. Indeed, β3-adrenergic receptor agonist treatment reduces the myeloid shift of HSCs in prematurely aged mice, caused by germ line mutations in lamin A78.
Thus, many different cell lineages in the microenvironment play a role in controlling stem cells during ageing via paracrine and soluble factors. The remodelling of the niche during ageing also results in changes in the spatial localization of cells, which could in turn influence tissue ageing by altering the physical proximity between stem cells and other supporting cells. Dissecting the importance of different cell types (for example, infiltration of immune cells or innervation in the tissues) with regard to the stem cell niche will be a key challenge for unravelling the mechanisms of tissue ageing.
ECM and biophysical properties.
ECM deposited by different cell types is emerging as a key player in stem cell ageing. In the brain, proteomic studies in the adult NSC niche have identified specific ECM components in the SVZ niche (compared with the olfactory bulb and the rest of the brain)139. ECM can be deposited by different cell types and impact ageing stem/progenitor cells. For example, ECM deposited by microglia in response to TGFβ redirects the fate of oligodendrocyte progenitor cells (OPCs) and renders them biased towards astrocytes during ageing140. In muscle, quantitative proteomic studies also reveal striking remodelling in ECM during ageing66,82, and those changes can influence MuSC function. In the skin, HFSCs exhibit significant transcriptomic changes in ECM proteins, and this is correlated with structural defects in the ageing stem cell niche11. Secreted ECM factors can also cause competition between stem cells and favour subclones during ageing. For example, during ageing, expression of collagen type XVII α1 chain (COL17A1) decreases owing to genomic stress (UV) in a heterogeneous manner in some epithelial stem cells (epidermal stem cells) but not others141. This results in competition and advantage of the subpopulation of COL17A1-positive stem cells during ageing and expansion of these cells141.
Importantly, general niche biophysical properties (for example, niche stiffness) are important and can act as a signalling mechanism via mechanosensing. In the brain, the ECM is stiffer in the SVZ neurogenic niche than in other regions139. In the central nervous system, transplantation experiments in rats indicate that ECM stiffness has a strong effect on OPC proliferation and differentiation during ageing80. OPC niche stiffness drastically increased with age, as determined by atomic force microscopy in the prefrontal cortex80. A softer niche is sufficient to restore OPC proliferation. Interestingly, niche stiffness is mediated by the mechanosensitive ion channel PIEZO1 in OPCs80. In muscle, ECM stiffness also increases with age and is associated with decreased collagen tortuosity82. These ECM changes induce MuSCs to adopt a more fibrogenic fate, thereby diminishing their myogenic potential. In the skin, niche stiffening and persistent hair shaft miniaturization during ageing can also contribute to HFSC ageing via a reduction in promoter bivalency in HFSCs84 and via a PIEZO1–TNF mechanism83. Thus, niche stiffness has a prominent effect on stem cell function during ageing. In the bone marrow, a specialized periarteriolar niche supports lymphoid progenitors and osteogenesis. The niche is maintained by mechanosensation, is transduced via PIEZO1 and is responsive to exercise. This niche is depleted with ageing, likely contributing to a reduced immune response81.
Finally, emerging results highlight the importance of fluid flow in stem cell regulation. In the SVZ of the adult brain, NSCs are bathed by cerebrospinal fluid. In a reconstituted slice system, increased fluid flow promotes proliferation of NSCs, and this is dependent on a sodium channel142. Thus, fluid flow could be an important component, at least in some stem cell niches, to integrate environmental changes.
Extrinsic signals from old niches appear to dominate over intrinsic function of young stem cells in transplantation experiments143,144. By contrast, signals from young niches may have only limited rejuvenation effects on old stem cells, at least for HSCs145. It will be interesting to identify strategies to bypass the sensitivity of stem cells to extrinsic signals from old niches, perhaps by conducting genetic or pharmacological screens in stem cells under conditions where the extrinsic signals are strongly inhibitory to identify genes or pathways whose manipulation renders stem cells insensitive to extrinsic inhibitory signals.
Interactions across systems
While many studies have focused on the role of local niche factors in regulating stem cell function, tissue crosstalk and communication at the system level are also important (FIG. 3). Here we discuss how stem cells in one tissue are influenced by long-range interactions from other tissues and systems altogether (for example, nervous system and circulating blood cells), with signals arising in completely different regions of the body. In addition, we review emerging studies on how the stem cells and their niches are influenced by broad systemic influences such as circadian rhythms.
System–system regulation of stem cells.
System–system long-range interactions that impact stem cells can involve circulating soluble factors (proteins and metabolites), circulating cells (immune cells), innervation and, potentially, circulating extracellular vesicles. Components of the blood — both cells and soluble factors, which circulate nearly everywhere — are easiest to envision as influencing ageing of distal stem cells. Indeed, as described earlier herein, some types of immune cells influence regenerative niches7. In addition, blood factors that can influence NSC ageing and rejuvenation include inflammatory cytokines (for example, CXCL11) (likely circulating in the blood), circulating enzymes produced by the liver and complement factors (likely circulating in the blood146–148) (see later). Interestingly, a number of factors associated with ageing have been traced to old HSCs. Some of these factors are found in greater abundance in the aged milieu and are linked to ageing, such as cyclophilin A149 and CCL5 (also known as RANTES)150, and others are found to be depleted with ageing, such as IGF1 (REF.133). While all such factors have the capacity to affect distal tissues, only a subset has been explored for their impact. Thus, a systematic study of factors that are commonly altered in human serum with ageing and how their absence or presence affects ageing of distal stem cell populations is warranted.
In addition to soluble molecules, the blood and other fluids also contain extracellular vesicles that are produced from different cells and could communicate with stem cells151. Extracellular vesicles contain RNAs and proteins, and could thereby influence tissues at a distance, depending on concentration and potency151. Interestingly, extracellular vesicles from young mice can enhance old MuSC function in culture, and extracellular vesicles in the serum are important for muscle regeneration in vivo152. Extracellular vesicles are heterogeneous, and their RNA content changes with age152. Age-dependent modulations of specific mRNAs (for example, Klotho mRNA) in extracellular vesicles could contribute to MuSC function and old muscle regeneration152. Likewise, extracellular vesicles from humans vary in content with age and can influence HSC proliferation in vitro153, although the impact of extracellular vesicles on HSCs in vivo remains unclear153. Further investigation will be needed to untangle the role of extracellular vesicles (and their various components) versus other blood components in the long-distance regulation of stem cell ageing in vivo.
Interestingly, stem cell niches are innervated, providing possible crosstalk between regenerative regions and the nervous system. In the bone marrow, nociceptive innervation has been shown to directly regulate HSC mobilization via calcitonin gene-related peptide (CGRP)154. Given the important role of CGRP in mammalian lifespan155, this could be an important mechanism for modulating HSC ageing. HSCs are also indirectly regulated by the sympathetic nervous system via the niche, and this mechanism is important for ageing156. In the brain, hypothalamic neurons have long-range interactions with a subset of NSCs (NKX2 positive) in the SVZ neurogenic niche157. Given the essential role of the hypothalamus in regulating body responses to diet, stress and even ageing, this connection between hypothalamic neurons and the NSC niche could help NSCs adapt to a changing environment. In muscle, muscle fibres are innervated by motor neurons, forming neuromuscular junctions. MuSCs contribute to the regeneration of neuromuscular junctions, and this is diminished during ageing158,159, but how motor neurons directly impact MuSCs during ageing is not yet known. Thus, the role of innervation to modulate stem cells and tissue repair with ageing is relatively unexplored and represents a potential therapeutic opportunity.
Circadian clocks.
Most if not all physiological responses are modulated by circadian clocks, which are mostly entrained by light (FIG. 3). Transcriptomic analysis shows that epidermal stem cells and MuSCs exhibit a circadian rhythm even in old animals, but that this rhythm is reprogrammed during ageing160,161. Caloric restriction can prevent this reprogramming of the circadian network with ageing in epidermal stem cells and MuSCs, whereas a high-fat diet makes it worse160,161. Circadian clock genes also play a key role in tissue homeostasis; for example, the clock gene Bmal1 (also known as Arntl) is required to prevent premature epithelial differentiation in the skin162. In the brain, the impact of the circadian clock with ageing is not yet known, but NSCs exhibit a circadian modulation, notably a differential ability to proliferate during the day that is calcium dependent163. In the haematopoietic system, normal HSCs are known to express a number of clock genes164. The influence of circadian rhythms on the haematopoietic system has been well documented. Circadian rhythms modulate HSC differentiation and maintenance165, HSC daily circulation166 and even survival of leukaemia cells167. Some of the effects of the circadian clocks are coordinated through the autonomic nervous system (reviewed in168). Given the disruption of circadian rhythms known to occur with ageing, and the documented effect of circadian clocks on at least some stem cell populations, age-associated changes in clock control very likely influence stem cell function and regeneration. This area has not yet been thoroughly examined, and represents an important research opportunity.
Stem cells in tissue ageing and disease
The decline in stem cell function is clearly a central feature of mammalian tissue ageing3–5, but tissues are composed of both stem cells and differentiated cells. A key question is to what extent does the decline in stem cells and their niches contribute to organ deterioration during ageing. Another important question is the role of stem cells in age-related diseases. This section reviews the current evidence and highlights remaining studies needed to understand the functional role of stem cell niches in the decline of tissue function in ageing and disease.
Forced depletion of stem cells in tissues.
Forced depletion of stem cells in a tissue usually results in impaired homeostasis and repair, suggesting that decrease in stem cell number may indeed contribute to decline that occurs during ageing. For example, depletion of fibroadipogenic progenitors in skeletal muscle leads to the ageing tissue phenotype of sarcopenia127. By contrast, forced depletion of MuSCs in muscle is not sufficient in and of itself to lead to muscle decline (sarcopenia)169, although it might participate in repair deficiencies. In the brain, glial cells and neurons originating from NSCs (or cells in the NSC region) can migrate to the site of injury and improve repair170–172. It will be interesting to more systematically test the role of stem cells in various aspects of tissue function and repair during ageing. However, it is important to note that the decline in stem cell numbers in tissues during ageing is more gradual than with genetic manipulations. In addition, in some regenerative regions (HSC niches), the number of stem cells increases with age173,174. Thus, genetic manipulations that more closely mimic the magnitude and type of the changes that occur with ageing will be needed to test the contribution of stem cells to the decline in homeostasis and repair.
Interventions that strongly impair stem cell function also exacerbate age-related phenotypes in tissues. For example, genetic deletion of ATR leads to defects in stem cell functions and ageing phenotypes175. Such interventions are also more drastic than the changes in stem cell function during ageing, and interventions that more closely mimic the changes in cell function during ageing will be needed. Another way of testing the role of stem cell function in tissue decline during ageing is transplantation. Introducing aged stem cells into tissues can result in organ impairments. For example, intraperitoneal transplantation of old, but not young, adipose-derived mesenchymal stem cells into old mice leads to an increase in signs of frailty, such as decreased walking speed and grip strength176.
Connection between stem cell depletion and age-related diseases.
Reduced stem cell number and function may also underlie aspects of age-related diseases. In the haematopoietic system, age-associated myelodysplastic syndrome is thought to be due to dysfunction of HSCs — driven by somatic or germ line mutations — that is exacerbated by an aged inflammatory milieu177. In addition, decline in stem cell number and function caused by germ line mutations can confer a host of phenotypes that resemble ageing-associated diseases. For example, germline mutations in genes of the telomerase complex cause dyskeratosis congenita, a disease characterized by age-associated phenotypes in multiple tissues, including bone marrow, skin and liver178. These tissue defects in dyskeratosis congenita are thought to result from inadequate tissue maintenance due to stem cell dysfunction178. Finally, as discussed earlier herein, somatic mutations in HSCs can lead to clonal haematopoiesis. These somatic mutations can be propagated through all the downstream blood lineages and result in abnormal circulating blood, which in turn impacts the function of distal tissues and contributes to cardiovascular and other age-associated diseases179,180.
Although there is debate about the formation of new neurons in the adult human hippocampus17,18,20,181–183, the decline in hippocampal neurogenesis appears to be accelerated in Alzheimer disease compared with physiological ageing20,184. Likewise, in most mouse models of Alzheimer disease based on familial mutations (for example, germ line mutations in the genes encoding amyloid precursor protein or presenilin), neurogenesis is decreased in both the SVZ and the hippocampus184,185. While some effects of Alzheimer disease mutations can be recapitulated in cultured NSCs, suggesting a direct effect of Alzheimer disease mutations on NSC function184, other effects of the mutations could be indirect, via niche cells. Indeed, risk factor genes involved in sporadic Alzheimer disease (for example, APOE) impair niche cells (astrocytes and microglia)186,187, which could in turn alter NSC function. Human patients with other neurodegenerative diseases due to mutations (for example, Parkinson disease, Huntington disease and amyotrophic lateral sclerosis) show abnormal increases in the numbers of astrocytes and microglia (and aberrant morphology of newborn neurons), although the number of NSCs is not strongly affected183. These studies, while correlative, raise the possibility that decline in the neurogenic niche and NSC function, especially in brain regions known to be important for Alzheimer disease, could play a role in disease progression.
Less is known about the impact of disease mutations on stem cells in muscle and other tissues. Mutations in key genes (for example, the dystrophin gene) involved in congenital muscle disease result in MuSC defects that could mimic aspects of stem cell ageing for tissue regeneration188,189. Overall, a more systematic understanding of the causal role of stem cells in age-related diseases will be a key step for innovative strategies for treatment of these diseases (BOX 1).
Box 1 |. Therapeutic strategies involving stem cells.
The ability of stem cells to generate a wide range of differentiated cells raises the tantalizing possibility that transplants of stem cells and their progeny could be used as therapeutic strategies for degenerative age-related diseases or to help repair after injury. another possible therapeutic avenue would be to activate the endogenous pool of stem cell within a tissue.
Transplantation of neural stem cells (NsCs) has been considered for neurodegenerative diseases and stroke. For example, transplantation of fetal mouse NsCs in an alzheimer disease mouse model limits amyloid deposition and reverses aspects of synaptic dysfunction239. Furthermore, transplanting human NsCs in an alzheimer disease mouse model also diminishes amyloid deposits and improves cognition240. Moreover, transplanting human NsCs from fetal or embryonic origin into the hippocampus of a mouse model of stroke decreases inflammation and ameliorates the detrimental effects of stroke241,242. this effect can be potentiated by a small-molecule compound called ‘3K3a-activated protein C’243. For Parkinson disease, transplanting dopamine neurons generated from embryonic stem cells has shown promise as a safe therapy244,245. However, transplanting exogenous stem cells could also have a detrimental impact as newly differentiated cells may not integrate properly into the already formed tissue.
Transplantation of muscle stem cells has been pursued in preclinical studies, primarily in the context of childhood diseases such as the muscular dystrophies246,247 and in the treatment of traumatic injuries such as volumetric muscle loss248,249. the loss of muscle mass that accompanies ageing, sarcopenia, has not been a focus of studies of stem cell therapeutics in part because there is little evidence to suggest that stem cells could ameliorate this form of muscle wasting and in part because of the challenge that would be faced in treating all affected muscles throughout the body, including limb and trunk muscles250. However, other muscle disorders that are associated with ageing, such as inclusion body myositis or sphincter dysfunction, might be more amenable to a stem cell transplantation approach because specific muscles or muscle groups are primarily affected. Preclinical studies of incontinence have led to early trials in patients using autologous myogenic progenitors251.
Transplantation of haematopoietic stem cells (HsCs) or their progeny (via bone marrow transplantation or mobilized peripheral blood progenitor cells) is used for treatment of cancers such as leukaemias or myelofibrosis. However, the need to ablate the endogenous stem cells for bone marrow transplantation makes it a procedure that is used only in the most dire circumstances (for example, chemotherapy-resistant cancers). the ablation of endogenous stem cells is so far the main barrier to the use of HsC transplantation as a therapeutic strategy for age-related diseases. Less traumatic strategies for ablation of host HsCs and/or better engraftment methods for donor HsCs will need to be developed. approaches using antibodies to deplete endogenous host HsCs appear promising252,253, but require optimization before they are transferred to the clinic. Given the central importance of blood for tissue function and during ageing, developing new strategies for HsC transplantation would be very interesting as these might have multitissue benefits.
It could be more efficient, and less risky, in terms of therapeutic strategies to tap into the pool of endogenous stem cells to ameliorate tissue function in disease. For example, increasing the levels of brain-derived neurotrophic factor (BDNF) and boosting neurogenesis improves cognition in an alzheimer disease mouse model, although the effects may not be solely due to NsC function254. it is important to note that differentiated cells originating from stem cells are ‘new’, so they may not have the same properties as differentiated cells that originated during development, may also not integrate well in existing tissues and may not migrate to the correct location. it is also possible that a diseased or aged niche will override the benefits originating from the rejuvenated (or transplanted) stem cells. thus, it will be critical to better understand the cues that allow better integration of newly born cells into diseased tissues and maintenance of the young phenotype for targeted therapies.
Enhanced stem cell function in health and lifespan.
Evidence for the contribution of enhanced stem cell function to tissue health and lifespan is so far most compelling in cases when endogenous, old cells are replaced by young cells or are rejuvenated in situ (see later). Further studies of multiple stem cell compartments should help discern the extent to which the functional decline in the resident stem cells contributes to the age-related changes in tissue or organ function, and the extent to which stem cells have broader roles in organismal ageing.
Overall, in tissues that heavily rely on stem cells (for example, blood and intestine), stem cell dysfunction may strongly contribute to tissue decline during ageing and disease. By contrast, in tissues that have more limited stem cell reservoirs (for example, muscle and brain), stem cell dysfunction may be more important in more extreme cases such as disease or decline in injury repair with ageing. Finally, in tissues that have no stem cells (for example, heart and spinal cord), ageing might have different overall characteristics (at the molecular level and the cellular level). Thus, it will be important to establish which stem cell features might be most critical for tissue ageing, disease and repair and how tissue ageing differs in tissues with and without stem cells.
Rejuvenation strategies impacting the niche
Some interventions can delay the onset of stem cell ageing as well as reverse age-dependent changes, restoring youthful molecular and functional features to the stem cells. Such ‘rejuvenating’ interventions can restore youthful properties to aged stem cells by modulating the stem cell environment or directly impacting stem cell states.
Three interventions have been reported to impact aged stem cell function via systemic signalling: heterochronic parabiosis, exercise and diet (TABLE 1).
Table 1 |.
Rejuvenation strategies and their impact on old stem cells in the brain, blood and muscle
| Dietary restriction and metabolites | Exercise | Blood factors | Reprogramming (with OCT4, SoX2, KLF4 and MYc) |
|---|---|---|---|
| Effects on aged brain | |||
| Increased NSC number (rapamycin) | Increased NSC number | Increased NSC number | Increased newborn neuron migration |
| Increased cognition (dietary restriction) | Increased cognition | Increased cognition | Increased cognition |
| Regeneration of axons from retinal ganglion cells | |||
| Effects on aged blood | |||
| No improvement of old HSCs (dietary restriction), but improvement of young HSCs | No improvement of HSCs | No improvement of HSC function | Transcriptional ‘rejuvenation' of blood |
| Improvement of HSCs (nicotinamide riboside) | Improvement of HSC niche and lymphoid progenitors | Transcriptional ‘rejuvenation' of HSCs | Improvement of old HSCs using other transcription factors (for example, XBP1) |
| Increased HSC function (rapamycin) | |||
| Effects on aged muscle | |||
| Improvement of MuSCs (NR and NMN) | Increased MuSC function | Increased MuSC number | Increased regenerative potential of MuSCs |
| Increased MuSC resilience (fasting) | Increased tissue function | Youthful state is promoted in MuSCs | |
| Effects on other aged organs | |||
| Multi-organ improvement (dietary restriction) | Multi-organ improvement | Improvement of liver, pancreas, bone and spinal cord | Transcriptional ‘rejuvenation' of pancreas, liver and spleen |
| Increased ISC regenerative function (fasting) and differentiation (dietary restriction) | No improvement of skeletal stem cells | ||
| Increased repair ability of ISCs (NR) | |||
HSC, haematopoietic stem cell; ISC, intestinal stem cell; MuSC, muscle stem cell; NMN, nicotinamide mononucleotide; NR, nicotinamide riboside; NSC, neural stem cell.
Heterochronic parabiosis and blood factors.
Studies of heterochronic parabiosis — the sharing of blood circulation between two individuals of different ages — have revealed that circulatory factors can impact aged stem cell function (TABLE 1). For this approach, a young animal is surgically connected by the flank to an older animal (heterochronic pair)190. As controls, two young animals or two old animals can also be fused by the flank (isochronic pairs). After 3 weeks to 1 month, blood circulation is established by capillaries between both animals. Initial work showed that stem cell and progenitor cell populations in muscle, liver and brain of aged individuals acquire more youthful functional and molecular states in heterochronic parabiosis16,191,192. Rejuvenation of aged stem or progenitor cells has since been confirmed in these tissues193,194, and extended to other tissues, including pancreas, bone and spinal cord195–197.
At least some aspects of heterochronic parabiosis can be recapitulated by repeated injection of young or old blood148,192. Thus, the beneficial effects of parabiosis are unlikely to be due to the engraftment of young cells from the circulation into the aged host, and are more likely to be due to non-cellular factors in the blood16,148,191,196,197. This is corroborated by heterochronic plasma transfer from young animals to aged animals or vice versa192,198. Indeed, transfusion of young plasma can partially recapitulate the effects of heterochronic parabiosis on aged tissues and, conversely, the transfusion of aged plasma has suppressive effects on young tissues. Interestingly, in studies of single blood exchanges, the ‘pro-ageing’ effect of old plasma is greater than the ‘anti-ageing’ effect of young plasma198. On the basis of these heterochronic parabiosis and plasma transfer studies, there has been much interest in identifying the factors that change with age in the blood and that have the potential to contribute to either the pro-ageing effects of aged plasma or the rejuvenating effects of young plasma199. Factors that have been suggested included potential rejuvenating factors (for example, TIMP2 from umbilical cord plasma)200 and potential ageing factors (for example, CXCL11)148, and there are likely additional factors, including non-protein factors (for example, metabolites). Not all stem cells may respond in the same way to the effect of parabiosis. For example, two studies showed that HSCs and skeletal stem cells were not rejuvenated by parabiosis128,201, while another study showed that HSCs were transcriptionally very responsive to young blood202. Perhaps stem cells that are needed for constant tissue replenishment are less functionally responsive to external blood factors than stem cells needed for tissue repair.
Exercise.
While the benefits of exercise on health are well known203, less is known about the molecular mediators of those benefits and whether and how those benefits are conferred to stem cell function. Exercise results in many physiological changes, including increases in blood flow, alterations in energy utilization and secretion of factors from muscle itself into the circulation, all of which have the potential to impact all the cells in the body204. In mice, exercise is often achieved by providing a free-spinning wheel in the cage for several weeks to a month. Mice naturally will use the wheel and run relatively long distances. Several recent studies have specifically tested for beneficial effects of exercise conferred by the transfer of plasma from exercised mice to non-exercised mice, with a specific emphasis on reversing age-related declines in stem cell function146,147 (TABLE 1).
Exercise has been found to rejuvenate aged MuSCs, in part by reversing the age-dependent increase in cyclin D1 levels to the levels observed in young cells34. Knockdown of cyclin D1 can prevent the exercise-induced enhancement of MuSC function34. Furthermore, aged MuSC function can be rejuvenated by induction of cyclin D1 expression specifically in those cells34. Intriguingly, transfer of plasma from exercised aged animals to non-exercised aged animals confers measurable benefits of exercise itself34. A rejuvenating effect of exercised aged animals has also been observed in NSCs205. Transfer of plasma from exercised mice to non-exercised aged mice enhanced neurogenesis and cognitive function146,147. There are likely several mechanisms at play. First, exercise can increase the levels of inhibitors of the complement cascade, including clusterin146. Depletion of clusterin abrogates the beneficial effects of plasma from exercised mice on aged cognitive function. Second, blood levels of the liver enzyme glycosylphosphatidylinositol-specific phospholipase D1 (GPLD1) increase in response to exercise, and the administration of GPLD1 to aged mice enhanced performance on cognitive tests147. However, old HSCs seem to be resistant to the effect of exercise201, although the HSC niche and lymphoid progenitors can respond positively to exercise81. It will be important to dissect in a systematic manner the beneficial effect of exercise on different tissue stem cells and their progeny.
Dietary restriction.
Alterations in the diet, particularly dietary restriction, have long been used to delay the ageing process206,207. Recently, there has been interest in other dietary regimens that confer similar health benefits but without the challenges of lifelong dietary restriction208,209. A frequently used dietary restriction regimen is restriction of food by a certain percentage (for example, 40%) without malnutrition, compared with ad libitum (no limit in food or a 10% reduction in food intake). Intermittent fasting (for example, every-other-day feeding) has also been implemented as a dietary restriction-related regimen. While animals tend to ‘binge’ on the day of feeding, this binging does not compensate for the fasting period. Finally, another dietary restriction-related regimen is the ‘fasting-mimicking diet’, which alternates 5 days of fasting and 2 days of feeding, repeated for several cycles208. While dietary restriction and related regimens extend lifespan and healthspan, a key question is whether they have a ‘rejuvenating’ effect and reverse ageing features in already old individuals. Among different regimens, the fasting-mimicking diet has been shown to have a bona fide rejuvenation effect on multiple organs in mice208, which may be due to alternating cycles of fasting and feeding.
Dietary restriction regimens have profound effects on many cells and tissues (TABLE 1). Dietary restriction can generally ‘reprogramme’ both the transcriptome and the proteome in ageing rats and mice210,211. The effects of dietary interventions are ultimately conveyed to cells, including stem cells, throughout the body via changes in the blood and then the local cellular milieu.
Dietary interventions have been tested for their potential to restore youthful function. In the blood, dietary restriction enhances the repopulating ability of HSCs, but also suppresses differentiation along the lymphoid lineage212. Old HSCs appear to be resistant to the beneficial effect of dietary restriction 201. In the intestine, ageing is associated with impaired ISC function that can lead to malabsorption syndromes. Dietary restriction followed by refeeding restores aged epithelial function by promoting stem cell differentiation along the secretory lineage211. The levels of the rate-limiting enzyme for ketogenesis, 3-hydroxy-3-methylglutaryl coenzyme A synthetase 2 (HMGCS2), are increased with dietary restriction and impact cell fate decisions of the stem cell211. Various regimens of intermittent fasting have also been found to impact stem and progenitor cells in aged animals213. In aged mice, the fasting-mimicking diet with cycles of fasting and refeeding enhances neurogenesis in the hippocampus208. Thus, dietary restriction has a beneficial effect on stem cell function, but may not affect all stem cell niches (or all cells in the niche) in the same manner.
Interestingly, complete fasting (nutrient deprivation for 24 h) has beneficial effects on ISC function — measured in this case by the ability of ISCs to regenerate in organoids — in both young mice and old mice214. The beneficial effect of fasting on ISCs is in part mediated by the induction of fatty acid oxidation214. Indeed, the critical enzyme for fatty acid oxidation, carnitine palmitoyltransferase 1A (CPT1A), is necessary for this improvement of ISCs upon fasting214. In muscle, fasting, while impairing the regenerative activity of MuSCs, enhances their resilience to a variety of stresses215. The beneficial effects of fasting on MuSC resilience can be mimicked by a ketogenic diet or by direct administration of ketone bodies215. Collectively, these studies indicate that dietary restriction and even complete fasting have a beneficial effect on stem cell pools. It will be important to test which specific components of the diet are most important for stem cell function.
Dietary restriction mimetics and metabolites.
In addition to dietary interventions, dietary restriction mimetics (for example, pharmacological compounds or metabolites that mimic dietary restriction) have also been shown to reverse age-related declines in stem cell function209 (TABLE 1). These include, but are not limited to, compounds that inhibit IGF1 signalling, that inhibit the mTOR pathway (for example, rapamycin) or that activate sirtuins and AMPK (for example, metformin).
For example, rapamycin treatment in old mice increases the percentage of activated NSCs64 and rejuvenates HSCs216. Metformin, an activator of AMPK and sirtuins (as well as other metabolic pathways), can restore youthful remyelination in the aged spinal cord following a demyelinating injury, and this effect is mediated by the promotion of OPC differentiation217. Activators of sirtuins and other metabolic pathways have been tested in studies to restore youthful function to aged stem cells. Among these approaches is the administration of precursors of NAD+, a cofactor not only for sirtuins but for a variety of enzymes116. The levels of NAD decline with ageing in both serum and tissues218. The administration of NAD precursors, in particular nicotinamide riboside and nicotinamide mononucleotide, has shown benefits in terms of healthspan219 and lifespan39. Nicotinamide riboside administration on ISCs revealed a reversal of the age-related impairment of the ability of these cells to repair injury in the gut120. The benefits of nicotinamide riboside can be suppressed by treatment of mice with inhibitors of either SIRT1 or mTOR complex 1 (mTORC1), which is paradoxical as inhibition of mTORC1 signalling normally extends healthspan and lifespan220. Nicotinamide riboside also has a beneficial effects on old MuSCs, NSCs and melanocyte stem cells, in part by rescuing mitochondrial function39. Likewise, nicotinamide riboside improves HSC function in young mice221 and ameliorates the lymphoid potential of HSCs in old mice222.
As described above, specific metabolites could play a key role in improving old tissue stem cell function114,115. For example, fumarate, which impacts young HSC function126, might also have beneficial effects on aged HSCs (although this has not been tested). In addition, metabolites that are key for stem cells, such as succinate and α-ketoglutarate, as well as oxygen levels, could restore old stem cell function114,115. In the future, it will be important to dissect the role of compounds and metabolites in stem cell and tissue function during ageing, especially as they have therapeutic applications. It is also worth noting that in most cases compounds or metabolites have not yet been used in a cyclical manner. Cyclical use of compounds and metabolites may increase their beneficial impact on aged stem cells.
Stem cell rejuvenation: partial reprogramming
Another interesting avenue towards rejuvenation is epigenetic reprogramming (TABLE 1). Full expression of the Yamanaka factors (OCT4, SOX2, KLF4 and MYC (OSKM)) in differentiated cells results in induction of pluripotency (induced pluripotent stem cells) and a resetting of cellular ageing characteristics223–225. Whether the rejuvenating effects of the reprogramming factors could be uncoupled from the dedifferentiating effects has been of great interest for enhancing the function of aged cells226. Initial studies of conditional expression of OSKM factors in vivo resulted in the development of teratomas, as differentiated cells acquired pluripotent characteristics227. Hence, continuous OSKM factor expression and reprogramming activity appeared to be incompatible with the notion of uncoupling dedifferentiation from rejuvenation. In contrast, cyclical induction of OSKM can restore youthful properties to aged cells and tissues, including stem and progenitor cells, without loss of cellular differentiation characteristics228. In this approach, OSKM are expressed in a cyclical manner using doxycycline: 5 days on (expression of OSKM), 2 days off (no expression of OSKM), repeated for several cycles (3–16)228,229. Alternatively, OSKM (or OSK) have been delivered via RNA delivery or adeno-associated viruses230,231.
Beneficial effects of partial reprogramming are observed in stem and progenitor cells in muscle and pancreas228. Transient transduction with mRNAs of reprogramming factors also rejuvenates different cell types in vitro, including MuSCs230. In transplantation assays, reprogrammed MuSCs from both aged mice and aged humans are almost as effective in their regenerative potential as their young counterparts230. The beneficial effects of OSKM on muscle function may be not cell autonomous: indeed expression of OSKM in myofibres, but not in MuSCs, promotes regeneration of old muscles232. Transient reprogramming also improves cells in the brain. For example, cyclical expression of OSKM in all tissues of middle-aged mice improves migration of a subtype of newly born hippocampal cells, and this is associated with improved cognitive function229. Reprogramming factors also affect differentiated cells in the brain: viral transduction of cells in vivo with OSK (no MYC) restores a youthful regenerative potential of axons emanating from aged mouse retinal ganglion cells231. It will be interesting to dissect the mechanisms by which reprogramming factors impact ageing hallmarks. Studies so far have shown that reprogramming factors ‘reset’ chromatin modifications in old cells228 and rejuvenate the DNA methylation and transcriptome in several organs of old mice (pancreas, liver, spleen and blood)233. Another key question will be to determine how long term the effect of reprogramming factors is in tissue ageing.
In addition to the Yamanaka factors, other forms of cellular reprogramming could also prove efficient to counter ageing features. HSCs lose function when maintained ex vivo, even in the presence of niche cells that sustain HSC potency in vivo234. While this in vitro assay does not directly reflect ageing, it can help identify new molecular players. With use of this system, a screen identified five transcription factors (including KLF7 and XBP1) that, when expressed in niche cells ex vivo, sustain HSC function for subsequent competitive reconstitution assays235. Although they do not induce reprogramming of cellular age per se, these transcription factors may allow restoration of youthful function that is observed with aged cells. Indeed, expression of these transcription factors prevents the accumulation of DNA damage, as occurs during ageing in vivo, in HSCs ex vivo. Identification of additional transcription factors, or other molecules236, involved in ‘reprogramming’ of old ageing features will be a key step in stem cell and tissue rejuvenation.
Other promising rejuvenation interventions for stem cells include senolytics (compounds that kill senescent cells within tissues), microbiota exchange (where a young microbiota is transferred to an old animals) and anti-inflammatory compounds237. It will be important to determine the effects of these interventions on different tissue stem cells during ageing.
Conclusion and perspectives
Recent advances in single-cell transcriptomics and lineage tracing technology have provided a systematic understanding of the ageing mechanisms in regenerative regions, highlighting the importance of cell–cell interactions and external factors such as inflammation.
Rejuvenating strategies have shown promising effects on stem cell function. In the future, it will be important to identify the main molecular pathways underlying these global rejuvenation strategies. It will also be intriguing to determine whether other emerging rejuvenation strategies (for example, microbiota replacement238 or circadian changes) could impact stem cell function.
For all stem cell rejuvenation strategies, it is crucial to understand how to balance rejuvenating effects and the risk of triggering tumorigenesis, especially in the case of reprogramming that involves an oncogene (for example, MYC), and young blood that could include factors that favour tumorigenesis. Furthermore, some rejuvenation strategies may have only transient benefits or even have negative long-term consequences. One possible explanation for such negative effects is that these rejuvenation interventions activate a pool of stem cells but in so doing deplete this pool prematurely. Thus, it will be essential to understand the mechanisms that control the balance between stem cell rejuvenation and stem cell exhaustion.
Most of the studies discussed in this Review were conducted in mice (sometimes in rats), and it is important to note that mouse and rat lifespan is relatively short (~3 years) and that mechanisms of stem cell maintenance may differ from those in humans and other long-lived species. Studying long-lived species for comparison (for example, comparing the naked mole rat and mice) and human stem cells or even generating stem cell chimaera (mouse–naked mole rat and chimpanzee–human) could shed light on the differences in stem cell maintenance mechanisms between species. Similarly, organoid or cellular models of human tissues might provide insights into specific mechanisms of stem cell ageing and rejuvenation, although the extent to which such in vitro models recapitulate the in vivo ageing features remains to be determined.
We have discussed current knowledge of the importance of stem cells for tissue homeostasis, tissue repair and disease development during ageing, as well as the potential of manipulating stem cells for therapy (BOX 1), but an efficient and safe stem cell therapy to slow down or reverse tissue ageing has not been identified. Several barriers will need to be overcome for such therapies to be used, including proper cell fate and integration in the tissue, maintenance of the youthful state of stem cells and their progeny in old tissues, and the absence of tumorigenesis. It will be important to determine which mechanisms have the potential to provide better therapeutic avenues — using stem cell transplantation or manipulating endogenous stem cell pools.
Developing and leveraging new technologies to follow and perturb stem cells will be a key area for future research. For example, longitudinal measurements of stem cell function throughout the lifespan will be crucial. It will also be important to identify in a systematic manner the gene networks involved in functional changes (for example, by performing CRISPR–Cas9-based screens). Identifying age-dependent changes in molecules other than RNAs and proteins (for example, metabolites and lipids) will likely uncover new players in the regulation of stem cell ageing, and possibly rejuvenation. Finally, as the spatial organization of regenerative niches is important for their function, a better understanding of such organization and cell interactions will be essential to better understand the normal role of stem cells in tissue function and repair and their changes during ageing. A systematic understanding of the mechanisms underlying stem cell niche ageing should also inform new strategies to reverse aspects of ageing in stem cells and counter tissue decline during ageing and disease.
Acknowledgements
The authors apologize for not including all relevant studies due to space constraints. They thank the reviewers for helpful comments. This work was supported by National Institutes of Health/National Institute on Aging P01AG036695 (A.B., M.A.G. and T.A.R.).
Glossary
- Regenerative regions
Specific regions of the tissue that contain stem cells. in some tissues (for example, brain and intestine), these regions are well delineated. in other tissues (for example, muscle), stem cells are distributed throughout.
- Neuroblasts
Progenitor cells that are committed to give rise to neurons.
- Lymphoid cells
B lymphocytes, T lymphocytes and natural killer cells.
- Myeloid lineage
Lineage giving rise to platelets, red blood cells, monocytes, neutrophils, basophils and eosinophils.
- CD34
Sialomucin, a glycosylated transmembrane protein that is expressed by several stem cells, notably muscle stem cells and, in humans, haematopoietic stem cells.
- Fibrogenic state
State characterized by abnormal deposition of extracellular matrix proteins and scarring of the tissue.
- DNA methyltransferase 3A
(DNMT3A). Enzyme that adds methyl groups to cytosines in DNA.
- Inflammatory cytokines
Signalling molecules secreted from immune cells or other cell types that promote inflammation.
- Mechanosensitive cells
Cells that respond to mechanical stress or mechanical stimulation.
- Bulge
Region of the outer root sheath of the hair which contains hair follicle stem cells.
- Paracrine
Refers to the effect of factors in the vicinity of cells that secrete these factors.
- ATM
Serine and threonine protein kinase that is recruited at and activated by double-strand breaks in DNA.
- Extracellular vesicles
Lipid-bound vesicles, including exosomes, secreted into the extracellular space.
- Nociceptive innervation
Innervation by sensory neurons that respond to damaging stimuli and send signals to the spinal cord and the brain.
- ATR
Serine and threonine protein kinase that is activated by single-strand breaks in DNA.
Footnotes
Competing interests
The authors declare no competing interests.
References
- 1.Rando TA & Wyss-Coray T Asynchronous, contagious and digital aging. Nat. Aging 1, 29–35 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sender R & Milo R The distribution of cellular turnover in the human body. Nat. Med 27, 45–48 (2021). [DOI] [PubMed] [Google Scholar]
- 3.Goodell MA & Rando TA Stem cells and healthy aging. Science 350, 1199–1204 (2015). [DOI] [PubMed] [Google Scholar]
- 4.Navarro Negredo P, Yeo RW & Brunet A Aging and rejuvenation of neural stem cells and their niches. Cell Stem Cell 27, 202–223 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Oh J, Lee YD & Wagers AJ Stem cell aging: mechanisms, regulators and therapeutic opportunities. Nat. Med 20, 870–880 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Artegiani B et al. A single-cell RNA sequencing study reveals cellular and molecular dynamics of the hippocampal neurogenic niche. Cell Rep. 21, 3271–3284 (2017). [DOI] [PubMed] [Google Scholar]
- 7.Dulken BW et al. Single-cell analysis reveals T cell infiltration in old neurogenic niches. Nature 571, 205–210 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kalamakis G et al. Quiescence modulates stem cell maintenance and regenerative capacity in the aging brain. Cell 176, 1407–1419.e14 (2019). [DOI] [PubMed] [Google Scholar]
- 9.Ibrayeva A et al. Early stem cell aging in the mature brain. Cell Stem Cell 28, 955–966.e7 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lukjanenko L et al. Aging disrupts muscle stem cell function by impairing matricellular WISP1 secretion from fibro-adipogenic progenitors. Cell Stem Cell 24, 433–446.e7 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ge Y et al. The aging skin microenvironment dictates stem cell behavior. Proc. Natl Acad. Sci. USA 117, 5339–5350 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Shcherbina A et al. Dissecting murine muscle stem cell aging through regeneration using integrative genomic analysis. Cell Rep. 32, 107964 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kimmel JC, Hwang AB, Scaramozza A, Marshall WF & Brack AS Aging induces aberrant state transition kinetics in murine muscle stem cells. Development 147, dev183855 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Aros CJ et al. Distinct spatiotemporally dynamic Wnt-secreting niches regulate proximal airway regeneration and aging. Cell Stem Cell 27, 413–429.e4 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chambers SM et al. Aging hematopoietic stem cells decline in function and exhibit epigenetic dysregulation. PLoS Biol. 5, e201 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Brack AS et al. Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science 317, 807–810 (2007). [DOI] [PubMed] [Google Scholar]
- 17.Sorrells SF et al. Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature 555, 377–381 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sorrells SF et al. Positive controls in adults and children support that very few, if any, new neurons are born in the adult human hippocampus. J. Neurosci 41, 2554–2565 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Franjic D et al. Transcriptomic taxonomy and neurogenic trajectories of adult human, macaque, and pig hippocampal and entorhinal cells. Neuron 110, 452–469.e14 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Moreno-Jimenez EP et al. Adult hippocampal neurogenesis is abundant in neurologically healthy subjects and drops sharply in patients with Alzheimer’s disease. Nat. Med 25, 554–560 (2019). [DOI] [PubMed] [Google Scholar]
- 21.Boldrini M et al. Human hippocampal neurogenesis persists throughout aging. Cell Stem Cell 22, 589–599.e5 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Durante MA et al. Single-cell analysis of olfactory neurogenesis and differentiation in adult humans. Nat. Neurosci 23, 323–326 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mann M et al. Heterogeneous responses of hematopoietic stem cells to inflammatory stimuli are altered with age. Cell Rep. 25, 2992–3005.e5 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Leins H et al. Aged murine hematopoietic stem cells drive aging-associated immune remodeling. Blood 132, 565–576 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sawen P et al. Murine HSCs contribute actively to native hematopoiesis but with reduced differentiation capacity upon aging. Elife 7, e41258 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yamamoto R et al. Large-scale clonal analysis resolves aging of the mouse hematopoietic stem cell compartment. Cell Stem Cell 22, 600–607.e4 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Obernier K et al. Adult neurogenesis is sustained by symmetric self-renewal and differentiation. Cell Stem Cell 22, 221–234.e8 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bast L et al. Increasing neural stem cell division asymmetry and quiescence are predicted to contribute to the age-related decline in neurogenesis. Cell Rep. 25, 3231–3240.e8 (2018). [DOI] [PubMed] [Google Scholar]
- 29.Harris L et al. Coordinated changes in cellular behavior ensure the lifelong maintenance of the hippocampal stem cell population. Cell Stem Cell 28, 863–876.e6 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kuang S, Kuroda K, Le Grand F & Rudnicki MA Asymmetric self-renewal and commitment of satellite stem cells in muscle. Cell 129, 999–1010 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Rocheteau P, Gayraud-Morel B, Siegl-Cachedenier I, Blasco MA & Tajbakhsh S A subpopulation of adult skeletal muscle stem cells retains all template DNA strands after cell division. Cell 148, 112–125 (2012). [DOI] [PubMed] [Google Scholar]
- 32.Garcia-Prat L et al. FoxO maintains a genuine muscle stem-cell quiescent state until geriatric age. Nat. Cell Biol 22, 1307–1318 (2020). [DOI] [PubMed] [Google Scholar]
- 33.Evano B et al. Transcriptome and epigenome diversity and plasticity of muscle stem cells following transplantation. PLoS Genet. 16, e1009022 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Brett JO et al. Exercise rejuvenates quiescent skeletal muscle stem cells in old mice through restoration of cyclin D1. Nat. Metab 2, 307–317 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chakkalakal JV, Jones KM, Basson MA & Brack AS The aged niche disrupts muscle stem cell quiescence. Nature 490, 355–360 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Scaramozza A et al. Lineage tracing reveals a subset of reserve muscle stem cells capable of clonal expansion under stress. Cell Stem Cell 24, 944–957.e5 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Collins CA, Zammit PS, Ruiz AP, Morgan JE & Partridge TA A population of myogenic stem cells that survives skeletal muscle aging. Stem Cell 25, 885–894 (2007). [DOI] [PubMed] [Google Scholar]
- 38.Sacma M et al. Haematopoietic stem cells in perisinusoidal niches are protected from ageing. Nat. Cell Biol 21, 1309–1320 (2019). [DOI] [PubMed] [Google Scholar]
- 39.Zhang H et al. NAD+ repletion improves mitochondrial and stem cell function and enhances life span in mice. Science 352, 1436–1443 (2016). [DOI] [PubMed] [Google Scholar]
- 40.Sousa-Victor P et al. Geriatric muscle stem cells switch reversible quiescence into senescence. Nature 506, 316–321 (2014). [DOI] [PubMed] [Google Scholar]
- 41.Le Roux I, Konge J, Le Cam L, Flamant P & Tajbakhsh S Numb is required to prevent p53-dependent senescence following skeletal muscle injury. Nat. Commun 6, 8528 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zhu P et al. The transcription factor Slug represses p16Ink4a and regulates murine muscle stem cell aging. Nat. Commun 10, 2568 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chiche A et al. Injury-induced senescence enables in vivo reprogramming in skeletal muscle. Cell Stem Cell 20, 407–414.e4 (2017). [DOI] [PubMed] [Google Scholar]
- 44.Yanai H & Beerman I Proliferation: driver of HSC aging phenotypes? Mech. Ageing Dev 191, 111331 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhao X et al. 4D imaging analysis of the aging mouse neural stem cell niche reveals a dramatic loss of progenitor cell dynamism regulated by the RHO-ROCK pathway. Stem Cell Rep. 17, 245–258 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.White CW III et al. Age-related loss of neural stem cell O-GlcNAc promotes a glial fate switch through STAT3 activation. Proc. Natl Acad. Sci. USA 117, 22214–22224 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Arai F et al. Machine learning of hematopoietic stem cell divisions from paired daughter cell expression profiles reveals effects of aging on self-renewal. Cell Syst. 11, 640–652.e5 (2020). [DOI] [PubMed] [Google Scholar]
- 48.Challen GA & Goodell MA Clonal hematopoiesis: mechanisms driving dominance of stem cell clones. Blood 136, 1590–1598 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Jaiswal S & Ebert BL Clonal hematopoiesis in human aging and disease. Science 366, eaan4673 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Jeong M et al. Loss of Dnmt3a immortalizes hematopoietic stem cells in vivo. Cell Rep. 23, 1–10 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Tovy A et al. Tissue-biased expansion of DNMT3A-mutant clones in a mosaic individual is associated with conserved epigenetic erosion. Cell Stem Cell 27, 326–335.e4 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lee-Six H et al. The landscape of somatic mutation in normal colorectal epithelial cells. Nature 574, 532–537 (2019). [DOI] [PubMed] [Google Scholar]
- 53.Martincorena I et al. Somatic mutant clones colonize the human esophagus with age. Science 362, 911–917 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Martincorena I et al. High burden and pervasive positive selection of somatic mutations in normal human skin. Science 348, 880–886 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Tierney MT, Stec MJ, Rulands S, Simons BD & Sacco A Muscle stem cells exhibit distinct clonal dynamics in response to tissue repair and homeostatic aging. Cell Stem Cell 22, 119–127.e3 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Watson CJ et al. The evolutionary dynamics and fitness landscape of clonal hematopoiesis. Science 367, 1449–1454 (2020). [DOI] [PubMed] [Google Scholar]
- 57.Hormaechea-Agulla D et al. Chronic infection drives Dnmt3a-loss-of-function clonal hematopoiesis via IFNgamma signaling. Cell Stem Cell 28, 1428–1442.e6 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Dharan NJ et al. HIV is associated with an increased risk of age-related clonal hematopoiesis among older adults. Nat. Med 27, 1006–1011 (2021). [DOI] [PubMed] [Google Scholar]
- 59.Bhattacharya R et al. Association of diet quality with prevalence of clonal hematopoiesis and adverse cardiovascular events. JAMA Cardiol. 6, 1069–1077 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Blokzijl F et al. Tissue-specific mutation accumulation in human adult stem cells during life. Nature 538, 260–264 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Heyde A et al. Increased stem cell proliferation in atherosclerosis accelerates clonal hematopoiesis. Cell 184, 1348–1361.e22 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yu KR et al. The impact of aging on primate hematopoiesis as interrogated by clonal tracking. Blood 131, 1195–1205 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ximerakis M et al. Single-cell transcriptomic profiling of the aging mouse brain. Nat. Neurosci 22, 1696–1708 (2019). [DOI] [PubMed] [Google Scholar]
- 64.Leeman DS et al. Lysosome activation clears aggregates and enhances quiescent neural stem cell activation during aging. Science 359, 1277–1283 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Liu L et al. Chromatin modifications as determinants of muscle stem cell quiescence and chronological aging. Cell Rep. 4, 189–204 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Schuler SC et al. Extensive remodeling of the extracellular matrix during aging contributes to age-dependent impairments of muscle stem cell functionality. Cell Rep. 35, 109223 (2021). [DOI] [PubMed] [Google Scholar]
- 67.Hernando-Herraez I et al. Ageing affects DNA methylation drift and transcriptional cell-to-cell variability in mouse muscle stem cells. Nat. Commun 10, 4361 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Svendsen AF et al. A comprehensive transcriptome signature of murine hematopoietic stem cell aging. Blood 138, 439–451 (2021). [DOI] [PubMed] [Google Scholar]
- 69.Tabula Muris C A single-cell transcriptomic atlas characterizes ageing tissues in the mouse. Nature 583, 590–595 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Moreno-Valladares M et al. CD8+ T cells are increased in the subventricular zone with physiological and pathological aging. Aging Cell 19, e13198 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Gross KM et al. Loss of slug compromises dna damage repair and accelerates stem cell aging in mammary epithelium. Cell Rep. 28, 394–407.e6 (2019). [DOI] [PubMed] [Google Scholar]
- 72.Mogilenko DA et al. Comprehensive profiling of an aging immune system reveals clonal GZMK+ CD8+ T cells as conserved hallmark of inflammaging. Immunity 54, 99–115.e12 (2021). [DOI] [PubMed] [Google Scholar]
- 73.Groh J et al. Accumulation of cytotoxic T cells in the aged CNS leads to axon degeneration and contributes to cognitive and motor decline. Nat. Aging 1, 357–367 (2021). [DOI] [PubMed] [Google Scholar]
- 74.Gate D et al. Clonally expanded CD8 T cells patrol the cerebrospinal fluid in Alzheimer’s disease. Nature 577, 399–404 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Gate D et al. CD4+ T cells contribute to neurodegeneration in Lewy body dementia. Science 374, 868–874 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.He H et al. Aging-induced IL27Ra signaling impairs hematopoietic stem cells. Blood 136, 183–198 (2020). [DOI] [PubMed] [Google Scholar]
- 77.Valletta S et al. Micro-environmental sensing by bone marrow stroma identifies IL-6 and TGFbeta1 as regulators of hematopoietic ageing. Nat. Commun 11, 4075 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Ho YH et al. Remodeling of bone marrow hematopoietic stem cell niches promotes myeloid cell expansion during premature or physiological aging. Cell Stem Cell 25, e6 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Frisch BJ et al. Aged marrow macrophages expand platelet-biased hematopoietic stem cells via interleukin-1B. JCI Insight 5, e124213 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Segel M et al. Niche stiffness underlies the ageing of central nervous system progenitor cells. Nature 573, 130–134 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Shen B et al. A mechanosensitive peri-arteriolar niche for osteogenesis and lymphopoiesis. Nature 591, 438–444 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Stearns-Reider KM et al. Aging of the skeletal muscle extracellular matrix drives a stem cell fibrogenic conversion. Aging Cell 16, 518–528 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Xie Y et al. Hair shaft miniaturization causes stem cell depletion through mechanosensory signals mediated by a Piezo1-calcium-TNF-alpha axis. Cell Stem Cell 29, 70–85.e6 (2022). [DOI] [PubMed] [Google Scholar]
- 84.Koester J et al. Niche stiffening compromises hair follicle stem cell potential during ageing by reducing bivalent promoter accessibility. Nat. Cell Biol 23, 771–781 (2021). [DOI] [PubMed] [Google Scholar]
- 85.Renault VM et al. FoxO3 regulates neural stem cell homeostasis. Cell Stem Cell 5, 527–539 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Gopinath SD, Webb AE, Brunet A & Rando TA FOXO3 promotes quiescence in adult muscle stem cells during the process of self-renewal. Stem Cell Rep. 2, 414–426 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Paik JH et al. FoxOs cooperatively regulate diverse pathways governing neural stem cell homeostasis. Cell Stem Cell 5, 540–553 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Tothova Z et al. FoxOs are critical mediators of hematopoietic stem cell resistance to physiologic oxidative stress. Cell 128, 325–339 (2007). [DOI] [PubMed] [Google Scholar]
- 89.Miyamoto K et al. Foxo3a is essential for maintenance of the hematopoietic stem cell pool. Cell Stem Cell 1, 101–112 (2007). [DOI] [PubMed] [Google Scholar]
- 90.Yalcin S et al. Foxo3 is essential for the regulation of ataxia telangiectasia mutated and oxidative stress-mediated homeostasis of hematopoietic stem cells. J. Biol. Chem 283, 25692–25705 (2008). [DOI] [PubMed] [Google Scholar]
- 91.Schaffner I et al. FoxO function is essential for maintenance of autophagic flux and neuronal morphogenesis in adult neurogenesis. Neuron 99, e6 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Audesse AJ et al. FOXO3 directly regulates an autophagy network to functionally regulate proteostasis in adult neural stem cells. PLoS Genet. 15, e1008097 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Hwang I et al. Cellular stress signaling activates type-I IFN response through FOXO3-regulated lamin posttranslational modification. Nat. Commun 12, 640 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Wheatley EG et al. Neuronal O-GlcNAcylation improves cognitive function in the aged mouse brain. Curr. Biol 29, 3359–3369 e4 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Liu L et al. Impaired notch signaling leads to a decrease in p53 activity and mitotic catastrophe in aged muscle stem cells. Cell Stem Cell 23, 544–556.e4 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Gutierrez-Martinez P et al. Diminished apoptotic priming and ATM signalling confer a survival advantage onto aged haematopoietic stem cells in response to DNA damage. Nat. Cell Biol 20, 413–421 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Gontier G et al. Tet2 rescues age-related regenerative decline and enhances cognitive function in the adult mouse brain. Cell Rep. 22, 1974–1981 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Maybury-Lewis SY et al. Changing and stable chromatin accessibility supports transcriptional overhaul during neural stem cell activation and is altered with age. Aging Cell 20, e13499 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Sun D et al. Epigenomic profiling of young and aged HSCs reveals concerted changes during aging that reinforce self-renewal. Cell Stem Cell 14, 673–688 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Sera Y et al. UTX maintains functional integrity of murine hematopoietic system by globally regulating aging-associated genes. Blood 137, 908–922 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Khokhar ES et al. Aging-associated decrease in the histone acetyltransferase KAT6B is linked to altered hematopoietic stem cell differentiation. Exp. Hematol 82, 43–52.e4 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Keenan CR et al. Extreme disruption of heterochromatin is required for accelerated hematopoietic aging. Blood 135, 2049–2058 (2020). [DOI] [PubMed] [Google Scholar]
- 103.Guerreiro I & Kind J Spatial chromatin organization and gene regulation at the nuclear lamina. Curr. Opin. Genet. Dev 55, 19–25 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Bin Imtiaz MK et al. Declining lamin B1 expression mediates age-dependent decreases of hippocampal stem cell activity. Cell Stem Cell 28, 967–977.e8 (2021). [DOI] [PubMed] [Google Scholar]
- 105.Dall’Agnese A et al. Transcription factor-directed re-wiring of chromatin architecture for somatic cell nuclear reprogramming toward trans-differentiation. Mol. Cell 76, 453–472.e8 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Ho TT et al. Autophagy maintains the metabolism and function of young and old stem cells. Nature 543, 205–210 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Wei Q et al. MAEA is an E3 ubiquitin ligase promoting autophagy and maintenance of haematopoietic stem cells. Nat. Commun 12, 2522 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Garcia-Prat L et al. Autophagy maintains stemness by preventing senescence. Nature 529, 37–42 (2016). [DOI] [PubMed] [Google Scholar]
- 109.Tang AH & Rando TA Induction of autophagy supports the bioenergetic demands of quiescent muscle stem cell activation. EMBO J. 33, 2782–2797 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.White JP et al. The AMPK/p27Kip1 axis regulates autophagy/apoptosis decisions in aged skeletal muscle stem cells. Stem Cell Rep. 11, 425–439 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Dong S et al. Chaperone-mediated autophagy sustains haematopoietic stem-cell function. Nature (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Vonk WIM et al. Differentiation drives widespread rewiring of the neural stem cell chaperone network. Mol. Cell 78, 329–345 e9 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Kruta M et al. Hsf1 promotes hematopoietic stem cell fitness and proteostasis in response to ex vivo culture stress and aging. Cell Stem Cell 28, 1950–1965.e6 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Chandel NS, Jasper H, Ho TT & Passegue E Metabolic regulation of stem cell function in tissue homeostasis and organismal ageing. Nat. Cell Biol 18, 823–832 (2016). [DOI] [PubMed] [Google Scholar]
- 115.Meacham CE, DeVilbiss AW & Morrison SJ Metabolic regulation of somatic stem cells in vivo. Nat. Rev. Mol. Cell Biol 23, 428–443 (2022). [DOI] [PubMed] [Google Scholar]
- 116.Imai S & Guarente L NAD+ and sirtuins in aging and disease. Trends Cell Biol. 24, 464–471 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Schultz MB & Sinclair DA Why NAD+ declines during aging: it’s destroyed. Cell Metab. 23, 965–966 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Ryall JG et al. The NAD+-dependent SIRT1 deacetylase translates a metabolic switch into regulatory epigenetics in skeletal muscle stem cells. Cell Stem Cell 16, 171–183 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Luo H et al. Mitochondrial stress-initiated aberrant activation of the NLRP3 inflammasome regulates the functional deterioration of hematopoietic stem cell aging. Cell Rep. 26, 945–954.e4 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Igarashi M et al. NAD+ supplementation rejuvenates aged gut adult stem cells. Aging Cell 18, e12935 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Poisa-Beiro L et al. Glycogen accumulation, central carbon metabolism, and aging of hematopoietic stem and progenitor cells. Sci. Rep 10, 11597 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Pala F et al. Distinct metabolic states govern skeletal muscle stem cell fates during prenatal and postnatal myogenesis. J. Cell Sci 131, jcs212977 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Hennrich ML et al. Cell-specific proteome analyses of human bone marrow reveal molecular features of age-dependent functional decline. Nat. Commun 9, 4004 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Yucel N et al. Glucose metabolism drives histone acetylation landscape transitions that dictate muscle stem cell function. Cell Rep. 27, 3939–3955.e6 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Beckervordersandforth R et al. Role of mitochondrial metabolism in the control of early lineage progression and aging phenotypes in adult hippocampal neurogenesis. Neuron 93, 560–573.e6 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Guitart AV et al. Fumarate hydratase is a critical metabolic regulator of hematopoietic stem cell functions. J. Exp. Med 214, 719–735 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Wosczyna MN et al. Mesenchymal stromal cells are required for regeneration and homeostatic maintenance of skeletal muscle. Cell Rep. 27, 2029–2035.e5 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Ambrosi TH et al. Aged skeletal stem cells generate an inflammatory degenerative niche. Nature 597, 256–262 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Ambrosi TH et al. Adipocyte accumulation in the bone marrow during obesity and aging impairs stem cell-based hematopoietic and bone regeneration. Cell Stem Cell 20, 771–784 e6 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Sampath SC et al. Induction of muscle stem cell quiescence by the secreted niche factor Oncostatin M. Nat. Commun 9, 1531 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Kann AP, Hung M & Krauss RS Cell-cell contact and signaling in the muscle stem cell niche. Curr. Opin. Cell Biol 73, 78–83 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Pentinmikko N et al. Notum produced by Paneth cells attenuates regeneration of aged intestinal epithelium. Nature 571, 398–402 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Young K et al. Decline in IGF1 in the bone marrow microenvironment initiates hematopoietic stem cell aging. Cell Stem Cell 28, 1473–1482.e7 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Maity P et al. Persistent JunB activation in fibroblasts disrupts stem cell niche interactions enforcing skin aging. Cell Rep. 36, 109634 (2021). [DOI] [PubMed] [Google Scholar]
- 135.Zhu C, Mahesula S, Temple S & Kokovay E Heterogeneous expression of SDF1 retains actively proliferating neural progenitors in the capillary compartment of the niche. Stem Cell Rep. 12, 6–13 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Yousef H et al. Aged blood impairs hippocampal neural precursor activity and activates microglia via brain endothelial cell VCAM1. Nat. Med 25, 988–1000 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Baruch K et al. Aging-induced type I interferon response at the choroid plexus negatively affects brain function. Science 346, 89–93 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Martin-Suarez S, Valero J, Muro-Garcia T & Encinas JM Phenotypical and functional heterogeneity of neural stem cells in the aged hippocampus. Aging Cell 18, e12958 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Kjell J et al. Defining the adult neural stem cell niche proteome identifies key regulators of adult neurogenesis. Cell Stem Cell 26, 277–293.e8 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Baror R et al. Transforming growth factor-beta renders ageing microglia inhibitory to oligodendrocyte generation by CNS progenitors. Glia 67, 1374–1384 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Liu N et al. Stem cell competition orchestrates skin homeostasis and ageing. Nature 568, 344–350 (2019). [DOI] [PubMed] [Google Scholar]
- 142.Petrik D et al. Epithelial sodium channel regulates adult neural stem cell proliferation in a flow-dependent manner. Cell Stem Cell 22, 865–878.e8 (2018). [DOI] [PubMed] [Google Scholar]
- 143.Carlson BM & Faulkner JA Muscle transplantation between young and old rats: age of host determines recovery. Am. J. Physiol 256, C1262–C1266 (1989). [DOI] [PubMed] [Google Scholar]
- 144.Popplewell LL & Forman SJ Is there an upper age limit for bone marrow transplantation? Bone Marrow Transpl. 29, 277–284 (2002). [DOI] [PubMed] [Google Scholar]
- 145.Kuribayashi W et al. Limited rejuvenation of aged hematopoietic stem cells in young bone marrow niche. J. Exp. Med 218 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.De Miguel Z et al. Exercise plasma boosts memory and dampens brain inflammation via clusterin. Nature 600, 494–499 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Horowitz AM et al. Blood factors transfer beneficial effects of exercise on neurogenesis and cognition to the aged brain. Science 369, 167–173 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Villeda SA et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature 477, 90–94 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Smith LK et al. The aged hematopoietic system promotes hippocampal-dependent cognitive decline. Aging Cell 19, e13192 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Ergen AV, Boles NC & Goodell MA Rantes/Ccl5 influences hematopoietic stem cell subtypes and causes myeloid skewing. Blood 119, 2500–2509 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Kalluri R & LeBleu VS The biology, function, and biomedical applications of exosomes. Science 367, eaau6977 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Sahu A et al. Regulation of aged skeletal muscle regeneration by circulating extracellular vesicles. Nat. Aging 1, 1148–1161 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Grenier-Pleau I et al. Blood extracellular vesicles from healthy individuals regulate hematopoietic stem cells as humans age. Aging Cell 19, e13245 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Gao X et al. Nociceptive nerves regulate haematopoietic stem cell mobilization. Nature 589, 591–596 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Riera CE et al. TRPV1 pain receptors regulate longevity and metabolism by neuropeptide signaling. Cell 157, 1023–1036 (2014). [DOI] [PubMed] [Google Scholar]
- 156.Maryanovich M et al. Adrenergic nerve degeneration in bone marrow drives aging of the hematopoietic stem cell niche. Nat. Med 24, 782–791 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Paul A, Chaker Z & Doetsch F Hypothalamic regulation of regionally distinct adult neural stem cells and neurogenesis. Science 356, 1383–1386 (2017). [DOI] [PubMed] [Google Scholar]
- 158.Liu W et al. Loss of adult skeletal muscle stem cells drives age-related neuromuscular junction degeneration. Elife 6, e26464 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Larouche JA et al. Murine muscle stem cell response to perturbations of the neuromuscular junction are attenuated with aging. Elife 10, e66749 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Solanas G et al. Aged stem cells reprogram their daily rhythmic functions to adapt to stress. Cell 170, 678–692.e20 (2017). [DOI] [PubMed] [Google Scholar]
- 161.Sato S et al. Circadian reprogramming in the liver identifies metabolic pathways of aging. Cell 170, 664–677.e11 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Welz PS et al. BMAL1-driven tissue clocks respond independently to light to maintain homeostasis. Cell 177, 1436–1447.e12 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Gengatharan A et al. Adult neural stem cell activation in mice is regulated by the day/night cycle and intracellular calcium dynamics. Cell 184, 709–722.e13 (2021). [DOI] [PubMed] [Google Scholar]
- 164.Chambers SM et al. Hematopoietic fingerprints: an expression database of stem cells and their progeny. Cell Stem Cell 1, 578–591 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Golan K, Kollet O, Markus RP & Lapidot T Daily light and darkness onset and circadian rhythms metabolically synchronize hematopoietic stem cell differentiation and maintenance: the role of bone marrow norepinephrine, tumor necrosis factor, and melatonin cycles. Exp. Hematol 78, 1–10 (2019). [DOI] [PubMed] [Google Scholar]
- 166.Garcia-Garcia A et al. Dual cholinergic signals regulate daily migration of hematopoietic stem cells and leukocytes. Blood 133, 224–236 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Puram RV et al. Core Circadian clock genes regulate leukemia stem cells in AML. Cell 165, 303–316 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Garcia-Garcia A & Mendez-Ferrer S The autonomic nervous system pulls the strings to coordinate circadian HSC functions. Front. Immunol 11, 956 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Fry CS et al. Inducible depletion of satellite cells in adult, sedentary mice impairs muscle regenerative capacity without affecting sarcopenia. Nat. Med 21, 76–80 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Adams KL & Gallo V The diversity and disparity of the glial scar. Nat. Neurosci 21, 9–15 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Kernie SG & Parent JM Forebrain neurogenesis after focal ischemic and traumatic brain injury. Neurobiol. Dis 37, 267–274 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Jin K et al. Evidence for stroke-induced neurogenesis in the human brain. Proc. Natl Acad. Sci. USA 103, 13198–13202 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.de Haan G & Van Zant G Dynamic changes in mouse hematopoietic stem cell numbers during aging. Blood 93, 3294–3301 (1999). [PubMed] [Google Scholar]
- 174.Rossi DJ et al. Cell intrinsic alterations underlie hematopoietic stem cell aging. Proc. Natl Acad. Sci. USA 102, 9194–9199 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Ruzankina Y et al. Deletion of the developmentally essential gene ATR in adult mice leads to age-related phenotypes and stem cell loss. Cell Stem Cell 1, 113–126 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Wang B et al. Transplanting cells from old but not young donors causes physical dysfunction in older recipients. Aging Cell 19, e13106 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Muto T et al. TRAF6 functions as a tumor suppressor in myeloid malignancies by directly targeting MYC oncogenic activity. Cell Stem Cell 29, 298–314.e9 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Beier F, Foronda M, Martinez P & Blasco MA Conditional TRF1 knockout in the hematopoietic compartment leads to bone marrow failure and recapitulates clinical features of dyskeratosis congenita. Blood 120, 2990–3000 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Jaiswal S et al. Age-related clonal hematopoiesis associated with adverse outcomes. N. Engl. J. Med 371, 2488–2498 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Genovese G et al. Clonal hematopoiesis and blood-cancer risk inferred from blood DNA sequence. N. Engl. J. Med 371, 2477–2487 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Kempermann G et al. Human adult neurogenesis: evidence and remaining questions. Cell Stem Cell 23, 25–30 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Snyder JS Recalibrating the relevance of adult neurogenesis. Trends Neurosci. 42, 164–178 (2019). [DOI] [PubMed] [Google Scholar]
- 183.Terreros-Roncal J et al. Impact of neurodegenerative diseases on human adult hippocampal neurogenesis. Science 374, 1106–1113 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Babcock KR, Page JS, Fallon JR & Webb AE Adult Hippocampal neurogenesis in aging and Alzheimer’s disease. Stem Cell Rep. 16, 681–693 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Hamilton LK et al. Aberrant lipid metabolism in the forebrain niche suppresses adult neural stem cell proliferation in an animal model of Alzheimer’s disease. Cell Stem Cell 17, 397–411 (2015). [DOI] [PubMed] [Google Scholar]
- 186.Sadick JS et al. Astrocytes and oligodendrocytes undergo subtype-specific transcriptional changes in Alzheimer’s disease. Neuron 110, 1788–1805.e10 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Parhizkar S & Holtzman DM APOE mediated neuroinflammation and neurodegeneration in Alzheimer’s disease. Semin Immunol. 10.1016/j.smim.2022.101594 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Dumont NA et al. Dystrophin expression in muscle stem cells regulates their polarity and asymmetric division. Nat. Med 21, 1455–1463 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Wang YX et al. EGFR-Aurka signaling rescues polarity and regeneration defects in dystrophin-deficient muscle stem cells by increasing asymmetric divisions. Cell Stem Cell 24, 419–432.e6 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Conboy MJ, Conboy IM & Rando TA Heterochronic parabiosis: historical perspective and methodological considerations for studies of aging and longevity. Aging Cell 12, 525–530 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Conboy IM et al. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature 433, 760–764 (2005). [DOI] [PubMed] [Google Scholar]
- 192.Villeda SA et al. Young blood reverses age-related impairments in cognitive function and synaptic plasticity in mice. Nat. Med 20, 659–663 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Katsimpardi L et al. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science 344, 630–634 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Sinha I, Sinha-Hikim AP, Wagers AJ & Sinha-Hikim I Testosterone is essential for skeletal muscle growth in aged mice in a heterochronic parabiosis model. Cell Tissue Res. 357, 815–821 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Salpeter SJ et al. Systemic regulation of the age-related decline of pancreatic beta-cell replication. Diabetes 62, 2843–2848 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Baht GS et al. Exposure to a youthful circulaton rejuvenates bone repair through modulation of beta-catenin. Nat. Commun 6, 7131 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Ruckh JM et al. Rejuvenation of regeneration in the aging central nervous system. Cell Stem Cell 10, 96–103 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Rebo J et al. A single heterochronic blood exchange reveals rapid inhibition of multiple tissues by old blood. Nat. Commun 7, 13363 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Ashapkin VV, Kutueva LI & Vanyushin BF The effects of parabiosis on aging and age-related diseases. Adv. Exp. Med. Biol 1260, 107–122 (2020). [DOI] [PubMed] [Google Scholar]
- 200.Castellano JM et al. Human umbilical cord plasma proteins revitalize hippocampal function in aged mice. Nature 544, 488–492 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Ho TT et al. Aged hematopoietic stem cells are refractory to bloodborne systemic rejuvenation interventions. J. Exp. Med 218, e20210223 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Palovics R et al. Molecular hallmarks of heterochronic parabiosis at single-cell resolution. Nature 603, 309–314 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Warburton DE, Nicol CW & Bredin SS Health benefits of physical activity: the evidence. CMAJ 174, 801–809 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Neufer PD et al. Understanding the cellular and molecular mechanisms of physical activity-induced health benefits. Cell Metab. 22, 4–11 (2015). [DOI] [PubMed] [Google Scholar]
- 205.van Praag H, Shubert T, Zhao C & Gage FH Exercise enhances learning and hippocampal neurogenesis in aged mice. J. Neurosci 25, 8680–8685 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.McCay CM, Cromwell MF & Maynard LA The effect of retarded growth upon the length of life span and upon ultimate body size. J. Nutr 10, 63–79 (1935). [PubMed] [Google Scholar]
- 207.McDonald RB & Ramsey JJ Honoring Clive McCay and 75 years of calorie restriction research. J. Nutr 140, 1205–1210 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Brandhorst S et al. A periodic diet that mimics fasting promotes multi-system regeneration, enhanced cognitive performance, and healthspan. Cell Metab. 22, 86–99 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Green CL, Lamming DW & Fontana L Molecular mechanisms of dietary restriction promoting health and longevity. Nat. Rev. Mol. Cell. Biol 23, 56–73 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Ma S et al. Caloric restriction reprograms the single-cell transcriptional landscape of rattus norvegicus aging. Cell 180, 984–1001.e22 (2020). [DOI] [PubMed] [Google Scholar]
- 211.Gebert N et al. Region-specific proteome changes of the intestinal epithelium during aging and dietary restriction. Cell Rep. 31, 107565 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Tang D et al. Dietary restriction improves repopulation but impairs lymphoid differentiation capacity of hematopoietic stem cells in early aging. J. Exp. Med 213, 535–553 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Mattson MP, Longo VD & Harvie M Impact of intermittent fasting on health and disease processes. Ageing Res. Rev 39, 46–58 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Mihaylova MM et al. Fasting activates fatty acid oxidation to enhance intestinal stem cell function during homeostasis and aging. Cell Stem Cell 22, 769–778 e4 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Benjamin DI et al. Fasting induces a highly resilient deep quiescent state in muscle stem cells via ketone body signaling. Cell Metab. 34, 902–918.e6 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Chen C, Liu Y, Liu Y & Zheng P mTOR regulation and therapeutic rejuvenation of aging hematopoietic stem cells. Sci. Signal 2, ra75 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Neumann B et al. Metformin restores CNS remyelination capacity by rejuvenating aged stem cells. Cell Stem Cell 25, 473–485.e8 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Covarrubias AJ, Perrone R, Grozio A & Verdin E NAD+ metabolism and its roles in cellular processes during ageing. Nat. Rev. Mol. Cell Biol 22, 119–141 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Yoshino J, Baur JA & Imai SI NAD+ intermediates: the biology and therapeutic potential of NMN and NR. Cell Metab. 27, 513–528 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Johnson SC, Rabinovitch PS & Kaeberlein M mTOR is a key modulator of ageing and age-related disease. Nature 493, 338–345 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Vannini N et al. The NAD-booster nicotinamide riboside potently stimulates hematopoiesis through increased mitochondrial clearance. Cell Stem Cell 24, 405–418.e7 (2019). [DOI] [PubMed] [Google Scholar]
- 222.Zong L et al. NAD+ augmentation with nicotinamide riboside improves lymphoid potential of Atm−/− and old mice HSCs. NPJ Aging Mech. Dis 7, 25 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Takahashi K & Yamanaka S Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663–676 (2006). [DOI] [PubMed] [Google Scholar]
- 224.Izpisua Belmonte JC Reprogramming development and aging: cell differentiation as a malleable process. Curr. Opin. Cell Biol 24, 713–715 (2012). [DOI] [PubMed] [Google Scholar]
- 225.Mahmoudi S & Brunet A Aging and reprogramming: a two-way street. Curr. Opin. Cell Biol 24, 744–756 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Rando TA & Chang HY Aging, rejuvenation, and epigenetic reprogramming: resetting the aging clock. Cell 148, 46–57 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Abad M et al. Reprogramming in vivo produces teratomas and iPS cells with totipotency features. Nature 502, 340–345 (2013). [DOI] [PubMed] [Google Scholar]
- 228.Ocampo A et al. In vivo amelioration of age-associated hallmarks by partial reprogramming. Cell 167, 1719–1733 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Rodriguez-Matellan A, Alcazar N, Hernandez F, Serrano M & Avila J In vivo reprogramming ameliorates aging features in dentate gyrus cells and improves memory in mice. Stem Cell Rep. 15, 1056–1066 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Sarkar TJ et al. Transient non-integrative expression of nuclear reprogramming factors promotes multifaceted amelioration of aging in human cells. Nat. Commun 11, 1545 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Lu Y et al. Reprogramming to recover youthful epigenetic information and restore vision. Nature 588, 124–129 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Wang C et al. In vivo partial reprogramming of myofibers promotes muscle regeneration by remodeling the stem cell niche. Nat. Commun 12, 3094 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Chondronasiou D et al. Multi-omic rejuvenation of naturally aged tissues by a single cycle of transient reprogramming. Aging Cell 21, e13578 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Isern J et al. Self-renewing human bone marrow mesenspheres promote hematopoietic stem cell expansion. Cell Rep. 3, 1714–1724 (2013). [DOI] [PubMed] [Google Scholar]
- 235.Nakahara F et al. Engineering a haematopoietic stem cell niche by revitalizing mesenchymal stromal cells. Nat. Cell Biol 21, 560–567 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Guan J et al. Chemical reprogramming of human somatic cells to pluripotent stem cells. Nature 605, 325–331 (2022). [DOI] [PubMed] [Google Scholar]
- 237.Mahmoudi S, Xu L & Brunet A Turning back time with emerging rejuvenation strategies. Nat. Cell Biol 21, 32–43 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Seidel J & Valenzano DR The role of the gut microbiome during host ageing. F1000Res 10.12688/f1000research.15121.1 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Kim JA et al. Neural stem cell transplantation at critical period improves learning and memory through restoring synaptic impairment in Alzheimer’s disease mouse model. Cell Death Dis. 6, e1789 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.McGinley LM et al. Human neural stem cell transplantation improves cognition in a murine model of Alzheimer’s disease. Sci. Rep 8, 14776 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Eckert A et al. Bystander effect fuels human induced pluripotent stem cell-derived neural stem cells to quickly attenuate early stage neurological deficits after stroke. Stem Cell Transl. Med 4, 841–851 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Huang L, Wong S, Snyder EY, Hamblin MH & Lee JP Human neural stem cells rapidly ameliorate symptomatic inflammation in early-stage ischemic-reperfusion cerebral injury. Stem Cell Res. Ther 5, 129 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Wang Y et al. 3K3A-activated protein C stimulates postischemic neuronal repair by human neural stem cells in mice. Nat. Med 22, 1050–1055 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Piao J et al. Preclinical efficacy and safety of a human embryonic stem cell-derived midbrain dopamine progenitor product, MSK-DA01. Cell Stem Cell 28, 217–229.e7 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Barker RA, Parmar M, Studer L & Takahashi J Human trials of stem cell-derived dopamine neurons for Parkinson’s disease: dawn of a new era. Cell Stem Cell 21, 569–573 (2017). [DOI] [PubMed] [Google Scholar]
- 246.Sun C, Serra C, Lee G & Wagner KR Stem cell-based therapies for Duchenne muscular dystrophy. Exp. Neurol 323, 113086 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Biressi S, Filareto A & Rando TA Stem cell therapy for muscular dystrophies. J. Clin. Invest 130, 5652–5664 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Mihaly E, Altamirano DE, Tuffaha S & Grayson W Engineering skeletal muscle: Building complexity to achieve functionality. Semin. Cell Dev. Biol 119, 61–69 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Eugenis I, Wu D & Rando TA Cells, scaffolds, and bioactive factors: Engineering strategies for improving regeneration following volumetric muscle loss. Biomaterials 278, 121173 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Boyer O et al. Myogenic cell transplantation in genetic and acquired diseases of skeletal muscle. Front. Genet 12, 702547 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Boyer O et al. Autologous myoblasts for the treatment of fecal incontinence: results of a phase 2 randomized placebo-controlled study (MIAS). Ann. Surg 267, 443–450 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Kwon HS et al. Anti-human CD117 antibody-mediated bone marrow niche clearance in nonhuman primates and humanized NSG mice. Blood 133, 2104–2108 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Chhabra A et al. Hematopoietic stem cell transplantation in immunocompetent hosts without radiation or chemotherapy. Sci. Transl. Med 8, 351ra105 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Choi SH et al. Combined adult neurogenesis and BDNF mimic exercise effects on cognition in an Alzheimer’s mouse model. Science 361, eaan8821 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]