

Abstract
Background
The mechanisms used by SARS-CoV-2 to induce major adverse cardiac events (MACE) are unknown. Thus, we aimed to determine if SARS-CoV-2 can infect the heart to kill cardiomyocytes and induce MACE in patients with severe COVID-19.
Methods
This observational prospective cohort study includes experiments with hamsters and human samples from patients with severe COVID-19. Cytokines and serum biomarkers were analyzed in human serum. Cardiac transcriptome analyses were performed in hamsters' hearts.
Results
From a cohort of 70 patients, MACE was documented in 26% (18/70). Those who developed MACE had higher Log copies/mL of SARS-CoV-2, troponin-I, and pro-BNP in serum. Also, the elevation of IP-10 and a major decrease in levels of IL-17 , IL-6, and IL-1r were observed. No differences were found in the ability of serum antibodies to neutralize viral spike proteins in pseudoviruses from variants of concern. In hamster models, we found a stark increase in viral titers in the hearts 4 days post-infection. The cardiac transcriptome evaluation resulted in the differential expression of ~ 9% of the total transcripts. Analysis of transcriptional changes of the effectors of necroptosis (mixed lineage kinase domain-like, MLKL) and pyroptosis (gasdermin D) showed necroptosis, but not pyroptosis, to be elevated. Active form of MLKL (phosphorylated MLKL, pMLKL) was elevated in hamster hearts and, most importantly, in the serum of MACE patients.
Conclusion
SARS-CoV-2 can reach the heart during severe COVID-19 and induce necroptosis in the heart of patients with MACE. Thus, pMLKL could be used as a biomarker of cardiac damage and a therapeutic target.
Keywords: COVID-19, Necroptosis, Heart injury
Introduction
COVID-19 has been the most devastating infectious disease since the 1918 influenza pandemic over 100 years ago [1], shaking healthcare systems and rattling the scientific community. It is projected to cause a cumulative worldwide loss of about USD 12.5 trillion and up to 8.76 million lives through 12/2024 [2, 3]. While infection with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), the causative agent of COVID-19, is often associated with respiratory and pulmonary-related diseases, it can also lead to detrimental effects on several organs and induce systemic complications. Major adverse cardiac events (MACE) (i.e., heart attacks, arrhythmias, heart failure, and strokes) are some of the most frequently diagnosed complications of COVID-19 [4-7]. MACE has been shown to occur during acute hospitalization and even once the patient is discharged from the hospital, leading to worse clinical outcomes, including higher mortality [8-10]. Despite these observations, the exact mechanism(s) by which infection with SARS-CoV-2 leads to MACE is unknown.
SARS-CoV-2 uses the angiotensin-converting enzyme 2 (ACE2) as the receptor to enter host cells. ACE2 is an enzyme within the renin-angiotensin system, involved in blood pressure regulation and electrolyte homeostasis [11, 12]. Baseline ACE2 expression is highest in the thyroid, heart, kidney, testis, and small intestine [13]. Furthermore, the expression of ACE2 is increased in cardiac tissue upon biological stress and chronic cardiovascular diseases such as heart failure [14] and respiratory infection by Streptococcus pneumoniae or influenza A virus [15]. These data suggest that the myocardium is susceptible to SARS-CoV-2 entry and can potentially lead to the development of MACE via direct viral infection of the cardiac tissue.
Several studies have documented the development of MACE in patients with COVID-19 [16, 17], mainly those with severe infection, comorbid conditions, and older age. Moreover, some researchers have documented that MACE is also frequent in patients that survive the acute infection and are now recognized as part of the long-COVID syndrome [18]. However, the underlying mechanisms of these complications still need to be better understood. Of importance, some studies have shown the presence of SARS-CoV-2 viral particles in the cardiac tissue of patients that succumbed to infection [16, 19-21]. Recent reports showed that SARS-CoV-2 could infect human cardiomyocytes via ACE2, replicate, and cause cell death in vitro [22, 23]. However, it is unknown whether cardiac infection with SARS-CoV-2 and the associated tissue injuries lead to the development of MACE. In addition, it is uncertain whether serum biomarkers could be used to identify patients at higher risk of developing MACE during acute COVID-19. Here we attempted to bridge these gaps to identify potential therapeutic targets to prevent MACE in patients infected with SARS-CoV-2.
Material And Methods
This observational prospective cohort study includes experiments with hamsters and human samples gathered of subjects admitted to the Clínica Universidad de La Sabana in Chía, Colombia, with confirmed COVID-19 diagnosed by reverse transcription polymerase chain reaction (RT-PCR). All consecutive patients admitted with severe disease to the participating center between November 2019 and May 2020 were included. Data were collected prospectively by the attending physicians by reviewing medical records, laboratory data, and blood samples within the first 24 hours of hospital admission were gathered to dissect the underlying mechanisms of MACE in these patients. This study was approved by the Institutional Review Board (IRB) of the Clínica Universidad de La Sabana, and all patients signed informed consent to participate in the study (CUS-LFR-012).
Subjects and data collection
The human cohort includes hospitalized patients older than 18 years. The defined disease severity was defined based on the World Health Organization criteria. Severe illness was diagnosed in patients with SpO2 ≤ 94% on room air, including patients on supplemental oxygen, oxygen through a high-flow device, or no-invasive ventilation. Critical illness was diagnosed in patients requiring invasive mechanical ventilation and/or extracorporeal membrane oxygenation or end-organ dysfunction.
During hospital admission, the following variables were collected: demographic data, comorbidities, symptoms, physiological variables collected during the first 24 hours of hospital admission, systemic complications, and laboratory reports. A retrospective chart review was conducted at hospital discharge to double-check the registered data.
Study definitions
MACE is a composite outcome [24, 25] that encompasses patients who develop any of the following clinical diagnoses. Cardiac arrhythmia (new or worsening): change from the sequence of electrical impulses in the electrocardiogram (EKG), compared to EKG at hospital admission or in past medical history [26]. Heart Failure (new or worsening): a clinical syndrome with symptoms and/or signs secondary to functional or structural cardiac abnormality, which may occur with or without previous cardiac disease documented through an echocardiogram, evidence of pulmonary or systemic congestion, and/or an increase of serum biomarkers such as Pro-BNP [27]. Myocardial injury: acute cardiac cell injury corroborated by the rise of serum troponin values with at least one value above the 99th percentile of the normal reference value of each local laboratory; development of pathological Q waves and/or new ischemic changes in EKG; evidence of coronary thrombus by angiography and/or new loss of viable myocardium, or regional wall motion abnormality identified in the echocardiogram [28, 29]. Finally, stroke was defined as a neurological deficit caused by an acute focal injury of the central nervous system by a vascular cause (i.e., cerebral infarction, intracerebral hemorrhage, or subarachnoid hemorrhage) [30].
Virus strain and hamster infection
SARS-CoV-2 isolate USA-WA-1/2020 was used for these studies. The reagent was deposited by the Centers for Disease Control and Prevention and obtained through BEI Resources, NIAID, NIH: SARS-Related Coronavirus 2, Isolate USA-WA-1/2020, NR-52281. Male golden Syrian hamsters of 3–4 weeks old were purchased from Charles River Laboratory and used at 5–6 weeks of age. All procedures were per approved IACUC (2020040AR and 2020048AR) and Institutional Biosafety Committee protocols (08496). Hamsters were infected with the indicated dose of virus intratracheally (i.t.) as previously described (31); animals were sedated with ketamine (100 mg/kg) and xylazine (5 mg/kg), and the tongue was pulled forward to visualize the trachea. Three hundred microliters of virus at a dose of 9 × 105 PFU of SARS-CoV-2, USA-WA-1/2020 suspension was applied to the trachea, and then the nose was covered to stimulate respiration.
Testing
All the specific performed testing, such as the quantification of cytokines and chemokines, RNA sequencing, neutralization assays, and western blots are explained in the Additional file 1.
Statistical analysis
Categorical variables are presented in counts (percentages) and were evaluated through the Chi-square test or Fisher's exact test. Continuous variables with normal distribution are expressed as means (standard deviation); variables with no normal distribution are expressed as median (interquartile ranges). For continuous variables with normal distribution, the t-Student test was performed, and for variables with no normal distribution Wilcoxon-Mann-Whitney test was used. Descriptive and bivariate analysis of the information was performed to determine the association between inflammatory profile and clinical outcomes, such as in-hospital mortality, the requirement of invasive mechanical ventilation, ICU admission, and hospital length of stay.
Results
A total of 70 patients with confirmed SARS-CoV-2 infection admitted to the intensive care unit (ICU) were included in the study (Fig. 1A). The median (IQR) age was 61.5 (50.5–69.0), and most of the patients were males (80% [56/70]). The main comorbidities were arterial hypertension (58.6% [41/70]), obesity (18.6% [13/70]), and diabetes mellitus (17.1% [12/70]) (Table 1). More than a quarter of patients developed MACE during ICU stay (25.7% [18/70]), and a total of 22 MACE were reported (Fig. 1B). The most frequent MACE diagnoses in the cohort were myocardial injury (50% [9/18]), followed by new onset arrhythmia (27.8% [5/18]), worsening heart failure (22.2% [4/18]), and myocardial infarction (11.1% [2/18]) (Fig. 1B). Interestingly, only one case of new-onset cardiac failure (5.6% [1/18]) and cardiovascular death (5.6% [1/18]) were documented (Fig. 1B).
Figure 1. Flowchart and diagnoses.

(A) Flowchart of patient selection and (B) Percentage diagnoses distribution among 18 patients that developed MACE.
Table 1.
Demographics characteristics of patients hospitalized due to COVID stratified by the presence of Major Adverse Cardiovascular Events (MACE)
| Characteristic | All n = 70 |
MACE n = 18 |
NO-MACE n = 52 |
P value |
|---|---|---|---|---|
| Demographic | ||||
| Male. n (%) | 56 (80.0) | 18 (100) | 38 (73.1) | 0.03 |
| Age. median (IQR) | 61.5 (50.5–69.0) | 67.5 (53.3–70.8) | 59.0 (49.8–68.0) | 0.40 |
| Comorbid conditions. n (%) | ||||
| Stroke | 1 (1.4) | 0 (0.0) | 1 (1.9) | 0.58 |
| Myocardial infarction | 4 (5.7) | 3 (16.7) | 1 (1.9) | 0.08 |
| Cardiac arrhythmia | 3 (4.3) | 3 (16.7) | 0 (0.0) | 0.02 |
| Asthma | 1 (1.4) | 0 (0.0) | 1 (1.0) | 0.58 |
| Diabetes mellitus | 12 (17.1) | 5 (27.8) | 7 (13.7) | 0.30 |
| Coronary disease | 2 (2.9) | 1 (5.6) | 1 (1.9) | 0.98 |
| Mental Illness | 1 (1.4) | 0 (0.0) | 1 (1.9) | 0.58 |
| Chronic kidney disease | 3 (4.3) | 2 (11.1) | 1 (1.9) | 0.33 |
| COPD | 8 (11.4) | 1 (5.6) | 7 (13.5) | 0.63 |
| Congestive cardiac failure | 2 (2.9) | 2 (11.1) | 0 (0.0) | 0.11 |
| Arterial hypertension | 41 (58.6) | 10 (55.6) | 31 (59.6) | 0.98 |
| Obesity | 13 (18.6) | 5 (27.8) | 8 (15.4) | 0.42 |
| OSAHS | 4 (5.7) | 0 (0.0) | 4 (7.7) | 0.53 |
| Smoking | 9 (12.9) | 2 (11.1) | 7 (13.5) | 0.88 |
| Dyslipidemia | 3 (4.3) | 3 (16.7) | 0 (0.0) | 0.02 |
| Physiological variables during the first 24 hours of admission. median (IQR) | ||||
| Heart rate. BPM | 93.5 (82.5–111.0) | 96.5 (83.3-116.5) | 92.0 (83.5-110.3) | 0.16 |
| Respiratory rate. BrPM | 22.0 (20.0–30.0) | 24.0 (20.0–30.0) | 22.0 (20.0–27.8) | 0.63 |
| Temperature. °C | 36.5 (36.3–37.0) | 36.7 (36.4–37.1) | 36.4 (36.2–37.0) | 0.16 |
| SBP. mmHg | 120.5 (108.5–131.0) | 124.5 (115.5-131.0) | 119.5 (104.5–131.0) | 0.55 |
| Demographic | ||||
| DBP. mmHg | 70.0 (64.0–79.0) | 70.5 (64.5–75.5) | 70.0 (64.0-80.25) | 0.63 |
| MAP. mmHg | 87.0 (79.5–98.0) | 87.5 (83.3–93.0) | 86.5 (78.0-98.3) | 1.00 |
| SPO2. (%) | 87.0 (72.75-91.0) | 80.5 (67.8–89.8) | 88.5 (80.5–91.0) | 0.12 |
| Glasgow | 15.0 (15.0–15.0) | 15.0 (15.0–15.0) | 15.0 (15.0–15.0) | 0.18 |
| Laboratory variables at admission. median (IQR) | ||||
| Presepsin, ng/L | 716.0 (471.8-1743.8) | 1080.5 (615.8-2458.8) | 683.5 (429.3–1596.0) | 0.14 |
| Pro-BNP, pg/mL | 715.5 (171.5-1443.3) | 1860.5 (939.8-4526.5) | 398.0 (132.0-1029.5) | < 0.001 |
| Troponin, ng/mL | 12.9 (4.9–89.7) | 106.5 (23.2–354.0) | 8.9 (3.7–21.5) | < 0.001 |
| D Dimer, mg/mL | 1.5 (0.9–3.3) | 1.2 (0.7–3.3) | 1.6 (1.0–3.2) | 0.45 |
| WBC, cell x 103 | 9.1 (7.1–12.4) | 8.6 (7.1–11.1) | 9.2 (7.2–13.2) | 0.51 |
| Neutrophils, (%) | 83.6 (78.4–89.4) | 85.4 (78.4–89.7) | 83.5 (79.1–89.3) | 0.84 |
| Hemoglobin, g/dL | 15.8 (14.4–17.5) | 16.5 (15.1–17.6) | 15.6 (14.3–17.2) | 0.42 |
| Platelet, cell x 103 | 236.5 (179.3-269.3) | 226.5 (178.3-263.8) | 237.0 (179.8-271.5) | 0.73 |
| Creatinine, mg/dL | 1.1 (0.8–1.3) | 1.1 (1.0–1.8) | 1.0 (0.8–1.2) | 0.07 |
| BUN, mg/dL | 19.9 (15.1–27.3) | 23.9 (14.5–29.1) | 19.1 (15.1–26.4) | 0.34 |
| Blood glucose, mg/dL | 138.1 (116.0-163.9) | 167.5 (129.5-209.3) | 129.9 (114.9–149.0) | 0.02 |
| Sodium, mEq/L | 136.9 (135.0-139.0) | 135.0 (133.0-136.8) | 137.0 (135.0-140.3) | 0.01 |
| Potassium, mEq/L | 4.3 (4.1–4.6) | 4.5 (4.1–5.0) | 4.3 (4.0–4.5) | 0.10 |
| Chloride, mEq/L | 98.6 (94.0-101.9) | 96.4 (89.4–99.2) | 99.0 (94.6–103.0) | 0.03 |
| Bilirubin max, mg/dL | 0.7 (0.5–1.3) | 0.7 (0.5–0.9) | 0.8 (0.5–1.4) | 0.26 |
| ALT, U/L | 60.5 (38.9–83.4) | 53.9 (38.9–83.0) | 60.9 (40.6–87.5) | 0.46 |
| AST, U/L | 62.9 (47.7–80.7) | 62.8 (48.9–74.9) | 62.9 (49.0-83.3) | 0.60 |
| pH | 7.5 (7.4–7.5) | 7.4 (7.4–7.5) | 7.5 (7.5–7.5) | 0.02 |
| Demographic | ||||
| PCO2, mmHg | 29 (26.3–33) | 29.0 (27.0–36.0) | 28.0 (26.0–32.0) | 0.44 |
| PaO2, mmHg | 59 (51–66) | 54.0 (45.3–59.8) | 61.0 (51.8–67.0) | 0.04 |
| HCO3, mmol/L | 21.5 (19.2–23.9) | 21.6 (17.4–25.3) | 21.5 (19.4–23.8) | 0.89 |
| Lactic acid, mmol/L | 1.6 (1.2–2.1) | 1.8 (1.3–2.5) | 1.6 (1.2–2.1) | 0.33 |
| CRP, mg/L | 204.4 (128.7-276.3) | 246.1 (174.7-274.7) | 191.2 (122.3-274.8) | 0.46 |
| PT, seconds | 12.6 (11.5–13.4) | 12.2 (11.2–12.7) | 12.7 (11.9–13.4) | 0.24 |
| PTT, seconds | 27.4 (26.1–29.8) | 28.5 (26.9–31.2) | 27.2 (25.9–28.6) | 0.11 |
| Outcomes | ||||
| Hospital LOS, days (IQR) | 12.5 (9.0–18.0) | 10.5 (9.0–14.5) | 14.0 (9.0–19.0) | 0.33 |
| In-Hospital Mortality (%) | 27 (38.6) | 11 (61.1) | 16 (30.8) | 0.02 |
Abbreviations: MACE: major cardiovascular events; IQR: interquartile range; COPD: chronic obstructive pulmonary disease; OSAHS: obstructive sleep apnea-hypopnea syndrome; BPM: beats per minute; BrPM: breaths per minute; SBP: systolic blood pressure; DPB: diastolic blood pressure; MAP: mean arterial pressure; SPO2: oxygen saturation; BNP: brain natriuretic peptide; WBC: white blood cell count; BUN: blood urea nitrogen; ALT: alanine aminotransferase; AST: aspartate aminotransferase; PCO2: partial pressure of carbon dioxide; PaO2: partial pressure of oxygen; HCO3: bicarbonate; CRP: C reactive protein; PT: prothrombin Time; PTT: thromboplastin time; LOS: length of stay.
Patients who developed MACE were older than those who did not (median [IQR], 67.5 years old [53.3–70.8] vs. 59.0 years old [49.8–68.0], p = 0.04). In addition, patients who developed MACE had more frequently a past medical history of cardiovascular diseases; such as myocardial infarction (16.7% [3/18] vs. 1.9% [1/52]; p = 0.08), cardiac arrhythmia (16.7% [3/18] vs. 0.0 [0/52]; p = 0.02), dyslipidemia (16.7 [3/18] vs. 0.0 [0/52]; p = 0.02), and congestive heart failure (1.1% [2/18] vs. 0.0 [0/52]; p = 0.11). Both groups (i.e., MACE and no-MACE) had similar physiological variables at admission; however, some laboratory results were different among the groups (Table 1). For instance, chloride, sodium, and pH levels were significantly lower in MACE patients than in no-MACE patients (96.4 mEq/L [89.4–99.2] vs. 99.0 mEq/L [94.6–103.0], p = 0.03; 135.0 mEq/L [133.0-136.8] vs. 137.0 mEq/L [135.0-140.3], p = 0.01; 7.4 [7.4–7.5] vs. 7.5 [7.5–7.5], p = 0.02; respectively) (Table 1).
Regarding the clinical outcomes, MACE patients had a shorter length of stay when compared with no-MACE patients; however, this difference was not statistically significant (10.5 days [9.0-14.5] vs. 14.0 days [9.0–19.0], p = 0.33). In contrast, we found that patients who developed MACE during hospital admission had a significantly higher mortality rate (61.1% [11/18] vs. 30.8% [16/52], p < 0.02). All demographic characteristics are presented in Table 1.
The relation between serum viral load and the development of MACE
To determine whether serum viral load was associated with the development of MACE, we assessed the amount of SARS-CoV-2 in the serum of the cohort at hospital admission. We found that the patients who developed MACE had higher Log copies/mL of SARS-CoV-2 (1.422 [0.000-6.971] vs. 0.363 [0.000-3.582], p < 0.05) when tested via quantitative RT-PCR in serum (Fig. 2A).
Figure 2. MACE patients increase markers of cardiac injury and SARS-CoV-2 presence.

A. Log copies per mL of SARS-CoV-2 tested via quantitative RT-PCR. Changes in B. Troponin I, C. Pro-BNP, D. D dimer, and E. Presepsin (ng per L of serum). As measured by multiplex analysis, F. Cytokine and chemokine changes in serum (pg per mL). Mean ± SEM analyzed by Kruskal-Wallis test with Dunn's multiple-comparison post-test (A-E), t-tests used in F. Asterisks denote the level of significance observed: * = p ≤ 0.05; ** = p ≤ 0.01; *** = p ≤ 0.001.
The relation of serum biomarkers and cytokines with MACE development
Different critical mediators were measured to define possible biomarkers for MACE development. MACE patients had increased serum concentration of cardiac biomarkers, principally Troponin I (106.5 ng per mL [23.2–354.0] vs. 8.9 ng per mL [3.7–21.5]; p < 0.001) (Fig. 2B) and pro-B-type natriuretic peptide (pro-BNP) (1860.5 [939.8-4526.5] vs. 398.0 [132.0-1029.5]; p < 0.001) at hospital admission (Fig. 2C). Notably, D-dimer and presepsin were not increased in MACE patients at hospital admission (Fig. 2D, Fig. 2E). Moreover, cytokines levels also showed differences among MACE and no-MACE patients, mainly by elevation of interferon-g-induced protein-10 (IP-10) (6689.22 ng per mL [1083.44-20327.7] vs. 10625.92 ng per mL [3492.59-18129.20]; p = 0.05) and interleukin (IL)-10 (105.56 ng per mL [28.83-243.87] vs. 46.68 ng per mL [3.58-271.29]; p < 0.05) or a decrease in levels of IL-17 (1.54 ng per mL [0.0000745-7.22] vs. 261.63 ng per mL [0.00000411-1822]; p < 0.05), IL-1β (0.025 ng per mL [0.00144-0.052] vs. 0.381 ng per mL [0.000356-6.68]; p < 0.05), Il-4 (3.34 ng per mL [0.412–8.97] vs. 26.16 ng per mL [0.006–149.76]; p < 0.05), and IL-6 (37.05 ng per mL [16.79–60.21] vs. 72.86 ng per mL [0.06-345.13]; p < 0.05) compared to the no-MACE group (Fig. 2F).
Neutralization of SARS-CoV-2 variants and the development of MACE
Spike-specific neutralizing antibodies are generally considered correlates of protection against COVID-19. We want to determine whether there were any significant differences in the neutralization titers between the MACE and no-MACE groups, which could explain the disease trajectory following ICU admission. No significant differences were observed in the ability to neutralize pseudoviruses from the variant of concerns: SARS-CoV-2 (D614) or Beta, Gamma, Delta, and Omicron subvariants (both BA.1 and BA.2). While a partial increase in the neutralization of D614 was observed in the no-MACE group and an increase of Delta neutralization in the MACE group, these were not statistically significant. Our results suggest that at the time of ICU admission, the robustness of neutralizing antibody activity does not appear to be involved in preventing MACE (Fig. 3). Moreover, these data also suggest that MACE was not associated with a particular viral variant of concern.
Figure 3. Plasma-neutralizing titers in patients with MACE show no significant changes.

pNT50 values against the four SARS-CoV-2 pseudo-viral variants Beta, Gamma, Delta, and Omicron BA.1 and BA.2 and the control early 2020 strain with the D614 mutation. They were measured in samples collected from patients experiencing MACE and those with no signs of MACE (No-MACE).
Hearts of hamsters infected with SARS-COV-2 show upregulation of major pathways associated with injury, cell death, antiviral immune responses, and metabolic changes.
To understand the underlying effects of severe SARS-CoV-2 infection in the heart, we used a recently established Golden Syrian Hamster model of severe infection [31]. Hamsters were infected intratracheally with SARS-CoV-2 strain USA-WA-1/2020 at a 9 X 10^5 PFU dose. Four days later, mice were sacrificed, and hearts were excised for assessment of viral titers and RNA sequencing (Fig. 4A). We observed a stark increase in viral titers (plaque forming units, PFU) in the hearts of hamsters 4 days post-infection when compared to mock-infected animals (Fig. 4B). At this time point, viral titers in the lungs were shown to start decreasing in a previous report by our group, using the same model [31]. Then, the cardiac transcriptome was evaluated by transcriptome sequencing (RNA-seq) of uninfected hearts and hearts infected with SARS-CoV-2 on day 4 post-infection. The infection resulted in the differential expression of 1,084 transcripts with a p-value of ≤ 0.05 out of 12,556 transcripts for a change of 8.63% of the total cardiac transcriptome. Gene ontology (biological processes) analysis for the significantly changed transcripts showed major changes in terms associated with programmed cell death (Fig. 4C), regulation of reactive oxygen species (ROS) (Fig. 4D), defense response to the virus (Fig. 4E), and carbohydrate metabolic processes (Fig. 4F). Of note, cell death and ROS activity in the heart have been linked to active pathogenesis of cardiovascular diseases [32] and cardiac damage in models of severe pneumococcal and pandemic influenza infections [15, 33-35]. The stark response to viral infection shown in the transcriptional increase of factors such as Ifitm2, Irf7, Stat1, and Eif2a, among others (Fig. 4E), indicates a heart actively mounting a host response to the invading SARS-CoV-2 (Fig. 4B).
Figure 4. Cardiac transcriptional changes in a hamster model of severe SARS-CoV-2.

A. Male 5-to-6-week-old golden Syrian hamsters were IT infected with WA1, or mock challenged with vehicle (PBS), and hearts excised at 4 days post-infection for transcriptomics. B. Log PFU per gram tissue of SARS-CoV-2 in hearts of hamsters (student t-test, p-value < 0.001 ****). C-F. Log fold change (log FC) of transcriptional changes in hearts of infected vs. uninfected hamsters. Representative gene ontology terms selected were C. programmed cell death, D.regulation of ROS, E. defense response to the virus, and F. carbohydrate metabolic processes.
Hamsters and humans show the activity of programmed necrosis, i.e., necroptosis in the heart and serum, respectively.
Analysis of transcriptional changes of the effectors of the two major programmed necrosis pathways, necroptosis (mixed lineage kinase domain-like; MLKL) and pyroptosis (gasdermin D; GSDMD), showed necroptosis to be starkly elevated in SARS-CoV-2 infected hamsters compared to the hearts of the uninfected group (Fig. 5A). We used immunoblots to confirm whether these transcription changes reflected protein and activity levels. Blots for the phosphorylated MLKL (pMLKL), the active form of MLKL, in hamster hearts showed significant activity of necroptosis in the hearts of SARS-CoV-2-infected hamsters (Fig. 5B).
Figure 5. Hamster hearts show increased necroptosis in model of severe SARS-CoV-2.

A-C. Male 5-to-6-week-old golden Syrian hamsters were IT infected with WA1 or mock challenged with vehicle (PBS) and hearts excised at 4 days post-infection for transcriptomics. A. Normalized Transcript levels of MLKL and gasdermin-D (GSDMD) in uninfected vs. SARS-CoV-2 infected hamsters. B. Immunoblot for pMLKL in homogenates of hamsters' hearts. C. Immunoblot for pMLKL in serum of hospitalized patients experiencing MACE or no-MACE. Mean ± SEM analyzed by (B) student T test or (C) Kruskal-Wallis test with Dunn's multiple-comparison post-test. Asterisks denote the level of significance observed: * = p ≤ 0.05; ** = p ≤ 0.01; *** = p ≤ 0.001; *** = p ≤ 0.0001.
While hamsters provide a pivotal model to study SARS-CoV-2 cardiac pathogenesis, we aimed to define if such a strong indicator of tissue injury was present in our MACE-experiencing human cohort. Immunoblots for pMLKL showed a distinct increase in the presence of this molecular marker of necroptotic cell death in the serum of MACE patients compared to those without MACE (Fig. 5C). These results suggest necroptosis activation in patients with MACE, which correlates with the findings described in the animal model. The complete uncropped gel and blot images can be found in the Additional File (Figure S1)
Markers of cardiac injury, inflammation, necroptosis, and circulating SARS-CoV-2 strongly correlate with the development of MACE
Using an annotated heatmap, we evaluated the association of MACE with all analyzed serum biomarkers (Fig. 6A). We observed that Troponin-I and pMLKL were the main variables associated with the development of MACE in humans. Then, single linear regressions were performed to analyze these relations further. We found a strong correlation between serum pMLKL and serum Troponin-I levels with a regression coefficient of 0.3589, p = 0.0086 (Fig. 6B) in MACE patients but not in the no-MACE (Fig. 6E). Notably, the serum viral burden evaluated by SARS-CoV-2 copies was associated with Troponin-I release in patients with MACE (Fig. 6C), with a regression coefficient of 0.2576, p = 0.03, but not in no-MACE patients (Fig. 6F). SARS-CoV-2 copies and pMLKL levels in serum were found not to correlate in both the MACE (Fig. 6D) and no-MACE (Fig. 6G) groups. Of note, the top four biomarkers for the development of MACE were observed using a mean accuracy plot and defined to be pMLKL, Troponin-I, serum presence of SARS-CoV-2, and Pro-BNP (Additional File Figure S2). These findings illustrate the relation between necroptosis, cardiac injury, and viral burden with the development of MACE.
Figure 6. Markers of cardiac-injury, inflammation, necroptosis, and SARS-CoV-2 correlate with MACE.
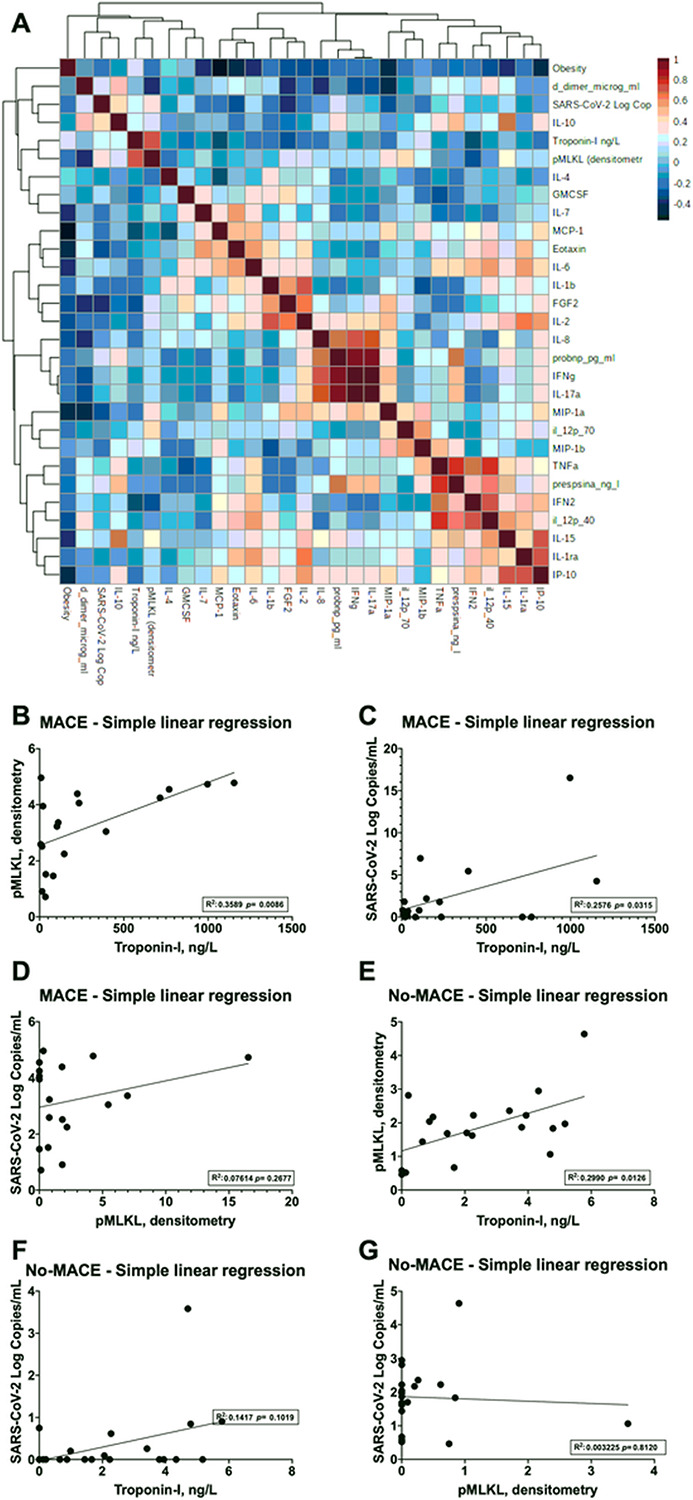
A. Correlation plot of serum cytokines, chemokines, biomarkers of cardiac injury, cell death effector MLKL, and SARS-CoV-2 (Log copies/mL). Linear regression plots for pMLKL - Troponin-I, SARS-CoV-2 (Log copies/mL) - Troponin-I and SARS-CoV-2 (Log copies/mL) - pMLKL (densitometry) in patients experiencing B-D. MACE or E-G.No-MACE.
Discussion
In this comprehensive assessment of the underlying mechanisms of MACE, we found that SARS-CoV-2 can infect hamster heart tissue, propagate, and induce cardiac damage by activating necroptosis. We also observed that human participants with COVID-19 who developed MACE had a higher viral burden, necroptosis activity, and markers of cardiac injury (troponin and pro-BNP) in the serum. In this cohort, we did not detect any significant difference in the neutralizing antibody breadth against SARS-CoV-2 variants. This observation suggests that antibody-neutralizing activity is not associated with protection from MACE progression at admission. These findings are novel and constitute an important advance in understanding MACE in patients with COVID-19 and propose some potential therapeutic targets.
Several epidemiological studies have extensively characterized MACE during and after COVID-19 [7, 18, 36, 37]. A recent systematic review of 150 studies with 33,805 patients showed that COVID-19 increased the risk of developing MACE. These complications were more frequently documented in patients with pre-existing cardiovascular comorbidities and disease severity [38]. However, a recent report by Xie et al. showed that even mild cases of COVID-19 have a high risk of developing MACE, such as heart failure and stroke, 1-year after recovering from the disease. Cardiac dysfunction has substantial implications for the treatment of COVID-19 but also demands deep mechanistic research to aid in reducing acute and long-lasting disease sequela (i.e., long-COVID) [10]. Of note, using a rodent model of infection Dhanyalayam et al. observed that SARS-CoV-2 could persist in the cardiac tissue of male and female humanized ACE2 of infected mice (USA-WA1/2020) at 10 days post-infection. The SARS-CoV-2 infection led to sex-dependent morphological changes in the hearts of mice. Males were found more susceptible to pathological changes such as immune cell infiltration, and a slight increase in fibrosis was observed in both sexes [39]. However, the molecular and immune determinants that modulate such pathology are unknown. The results from the presented study propose direct SARS-CoV-2-driven necroptosis and pyroptosis as a mechanism for cardiac injury and cardiovascular complications associated with COVID-19. An increase in circulatory SARS-CoV-2 signal (DNA copies/mL) is associated with increased markers of cardiac injury (Troponin-I and pro-BNP) and the presence of necroptosis effector pMLKL in serum. To corroborate this observation, we infected golden Syrian hamsters with SARS-CoV-2 and detected an increase in virus titers in the heart and the activity of necroptosis via transcriptomics and immunoblots. Taken together, our results propose a new mechanism for SARS-CoV-2-driven cardiac injury via direct cardiac infection and activation of necroptosis. It also provides a new therapeutic target to reduce the acute and long-term cardiac damage associated with COVID-19.
Most epidemiological and systematic reviews regarding MACE and COVID-19 have suggested Troponin-I increase as a major biomarker for MACE and prevalent disease outcomes, which was expected [5, 7, 22, 40, 41]. In this study, we found that viral burden and high concentrations of the necroptosis effector pMLKL in the serum were more accurate in predicting the development of MACE and possibly long-term cardiac events in convalescent COVID-19 patients. This is important as none of the MACE patients in this study had major changes in circulating cytokines and chemokines associated with cell death and injury or in biomarkers of cardiac damage driven by inflammation (i.e., presepsin). Inflammation has been proposed as the primary driver of severe COVID-19 [42-44] and, in some cases, of MACE [42-44]. However, here we found that the importance of systemic inflammation in the development of MACE is relatively marginal.
Necroptosis is a highly inflammatory form of cell death regulated by the receptor-interacting serine/threonine-protein kinases (RIPK)1 and RIPK3 and MLKL [45, 46]. Our study suggests that necroptosis can be a therapeutic target to reduce the acute and long-term effects of COVID-19 on the heart. Supporting this notion, our group has shown that inhibiting necroptosis or oxidative stress reduces cardiac injury observed during and after pneumococcal pneumonia [34]. Treatment with FDA-approved tyrosine kinase and necroptosis inhibitor "Ponatinib" (by inhibition of RIPK1 and RIPK3 [47]) was found to reduce collagen deposition, troponin release, and promote cardiac function up to 3 months after pneumococcal pneumonia when used as an adjunct therapy to antibiotics in a mouse model[34]. Notably, while beneficial during bacterial infection, Ponatinib blocks upstream effectors of necroptosis required in the cell defense against viruses [47, 48] ; thus, future studies should aim to use selective MLKL inhibitors to reduce cardiac injury driven by SARS-CoV-2.
While this study represents a comprehensive approach to dissecting some mechanisms of MACE, it has several limitations. First, we could not study the heart tissue of patients that developed MACE as our group lacked access to these tissues. However, we performed a robust clinical and paraclinical characterization of these patients that allowed us to identify necroptosis and viral burden as crucial factors for developing MACE. Second, the number of patients included in this study was small, which limits our capacity to develop high-accuracy predictive models using the serum biomarkers identified in our study. However, future studies will define how these markers directly correlate with MACE and if additional effectors can be linked to these phenotypes. Third, the kinetics of the study is not longitudinal; however, the data presented on Log copies/mL of SARS-CoV-2 were from samples collected upon admission to ICU. Thus, MACE patients showed more viral load persistence in serum.
In conclusion, the presented study suggests that necroptosis and circulating viral particles are primary drivers of MACE in severe COVID-19 patients. Notably, systemic inflammation and antibody responses to different viral variants were not associated with the development of MACE in humans. Herein we propose that pMLKL, along with Troponin-I and pro-BNP, can serve as more accurate biomarkers for acute and future development of MACE and disease outcomes. Finally, our data support the notion that inhibition of MLKL can be a possible new therapeutic or adjunct therapeutic approach to prevent the acute and long-term MACE caused by cardiac injury during severe COVID-19.
Funding:
LFR was supported by the Universidad de La Sabana and Clinica Universidad de La Sabana. N.G-J. was supported in part by the National Institutes for Health (NIH) awards AI148722-01A1, ES030227, and J. Craig Venter Institute Start-Up Funds. GST was supported in part by Conrad Prebys Foundation Grant 204. S.V. was partly supported by NIH awards R01AI137365, R03AI146632, R21AI146703, and R21AI149448 and the JCVI.
Footnotes
Ethics approval and consent to participate: For human participants and data, this study was approved by the Institutional Review Board (IRB) of the Clínica Universidad de La Sabana, and all patients signed informed consent to participate in the study (CUS-LFR-012). For animal study, golden Syrian hamsters of 3-4 weeks old were purchased from Charles River Laboratory and used at 5-6 weeks of age. All procedures were per approved IACUC (2020040AR and 2020048AR) and Institutional Biosafety Committee protocols (08496).
Availability of data and materials: The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Competing interests: We declare that none of the authors have competing financial or financial interests. LFR receives research funding and personal fees outside this manuscript's subject from Merck, Pfizer, and GSK
Contributor Information
Rosana Wiscovitch-Russo, J Craig Venter Institute.
Elsa D. Ibáñez-Prada, Universidad de la Sabana
Cristian C. Serrano-Mayorga, Universidad de la Sabana
Benjamin L. Sievers, J Craig Venter Institute
Maeve A. Engelbride, J Craig Venter Institute
Surya Padmanabhan, J Craig Venter Institute.
Gene S. Tan, J Craig Venter Institute
Sanjay Vashee, J Craig Venter Institute.
Ingrid G. Bustos, Universidad de la Sabana
Carlos Pachecho, Universidad de la Sabana.
Lina Mendez, Clinica Universidad de La Sabana.
Peter H. Dube, The University of Texas Health Science Center at San Antonio
Harinder Singh, J Craig Venter Institute.
Luis Felipe Reyes, Universidad de la Sabana.
Norberto Gonzalez-Juarbe, J Craig Venter Institute.
References
- 1.Simonetti O, Martini M, Armocida E: COVID-19 and Spanish flu-18: review of medical and social parallelisms between two global pandemics. J Prev Med Hyg 2021, 62(3):E613–E620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.IMF sees cost of COVID pandemic rising beyond $12.5 trillion estimate. [Google Scholar]
- 3.Ioannidis JPA: Global perspective of COVID-19 epidemiology for a full-cycle pandemic. Eur J Clin Invest 2020, 50(12):e13423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Carfi A, Bernabei R, Landi F, Gemelli Against C-P-ACSG: Persistent Symptoms in Patients After Acute COVID-19. JAMA 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chachques JC, Mazzini L, Mitrecic D, Zavan B, Rogante M, Latremouille C, Rustichelli F: Cardiovascular, hematological and neurosensory impact of COVID-19 and variants. Eur Rev Med Pharmacol Sci 2021, 25(8):3350–3364. [DOI] [PubMed] [Google Scholar]
- 6.Feng Y, Ling Y, Bai T, Xie Y, Huang J, Li J, Xiong W, Yang D, Chen R, Lu F et al. : COVID-19 with Different Severities: A Multicenter Study of Clinical Features. Am J Respir Crit Care Med 2020, 201(11):1380–1388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Long B, Brady WJ, Koyfman A, Gottlieb M: Cardiovascular complications in COVID-19. Am J Emerg Med 2020, 38(7):1504–1507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wu L, O'Kane AM, Peng H, Bi Y, Motriuk-Smith D, Ren J: SARS-CoV-2 and cardiovascular complications: From molecular mechanisms to pharmaceutical management. Biochem Pharmacol 2020, 178:114114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Unudurthi SD, Luthra P, Bose RJC, McCarthy JR, Kontaridis MI: Cardiac inflammation in COVID-19: Lessons from heart failure. Life Sci 2020, 260:118482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Xie Y, Xu E, Bowe B, Al-Aly Z: Long-term cardiovascular outcomes of COVID-19. Nat Med 2022, 28(3):583–590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bourgonje AR, Abdulle AE, Timens W, Hillebrands JL, Navis GJ, Gordijn SJ, Bolling MC, Dijkstra G, Voors AA, Osterhaus AD et al. : Angiotensin-converting enzyme 2 (ACE2), SARS-CoV-2 and the pathophysiology of coronavirus disease 2019 (COVID-19). J Pathol 2020, 251(3):228–248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Edmonston DL, South AM, Sparks MA, Cohen JB: Coronavirus Disease 2019 and Hypertension: The Role of Angiotensin-Converting Enzyme 2 and the Renin-Angiotensin System. Adv Chronic Kidney Dis 2020, 27(5):404–411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Li MY, Li L, Zhang Y, Wang XS: Expression of the SARS-CoV-2 cell receptor gene ACE2 In a wide variety of human tissues. Infect Dis Poverty 2020, 9(1):45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chamsi-Pasha MA, Shao Z, Tang WH: Angiotensin-converting enzyme 2 as a therapeutic target for heart failure. Curr Heart Fail Rep 2014, 11(1):58–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Platt MP, Lin YH, Wiscovitch-Russo R, Yu Y, Gonzalez-Juarbe N: Pandemic Influenza Infection Promotes Streptococcus pneumoniae Infiltration, Necrotic Damage and Proteomic Remodeling in the Heart. mBio 2022:e0325721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shi S, Qin M, Shen B, Cai Y, Liu T, Yang F, Gong W, Liu X, Liang J, Zhao Q et al. : Association of Cardiac Injury With Mortality in Hospitalized Patients With COVID-19 in Wuhan, China. JAMA Cardiol 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zou X, Chen K, Zou J, Han P, Hao J, Han Z: Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front Med 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Satterfield BA, Bhatt DL, Gersh BJ: Cardiac involvement in the long-term implications of COVID-19. Nat Rev Cardiol 2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lindner D, Fitzek A, Brauninger H, Aleshcheva G, Edler C, Meissner K, Scherschel K, Kirchhof P, Escher F, Schultheiss HP et al. : Association of Cardiac Infection With SARS-CoV-2 in Confirmed COVID-19 Autopsy Cases. JAMA Cardiol 2020, 5(11):1281–1285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Tavazzi G, Pellegrini C, Maurelli M, Belliato M, Sciutti F, Bottazzi A, Sepe PA, Resasco T, Camporotondo R, Bruno R et al. : Myocardial localization of coronavirus in COVID-19 cardiogenic shock. Eur J Heart Fail 2020, 22(5):911–915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yancy CW, Fonarow GC: Coronavirus Disease 2019 (COVID-19) and the Heart-Is Heart Failure the Next Chapter? JAMA Cardiol 2020. [DOI] [PubMed] [Google Scholar]
- 22.Bailey AL, Dmytrenko O, Greenberg L, Bredemeyer AL, Ma P, Liu J, Penna V, Winkler ES, Sviben S, Brooks E et al. : SARS-CoV-2 Infects Human Engineered Heart Tissues and Models COVID-19 Myocarditis. JACC Basic Transl Sci 2021, 6(4):331–345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Perez-Bermejo JA, Kang S, Rockwood SJ, Simoneau CR, Joy DA, Silva AC, Ramadoss GN, Flanigan WR, Fozouni P, Li H et al. : SARS-CoV-2 infection of human iPSC-derived cardiac cells reflects cytopathic features in hearts of patients with COVID-19. Sci Transl Med 2021, 13(590). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Africano HF, Serrano-Mayorga CC, Ramirez-Valbuena PC, Bustos IG, Bastidas A, Vargas HA, Gomez S, Rodriguez A, Orihuela CJ, Reyes LF: Major Adverse Cardiovascular Events During Invasive Pneumococcal Disease are Serotype Dependent. Clin Infect Dis 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bosco E, Hsueh L, McConeghy KW, Gravenstein S, Saade E: Major adverse cardiovascular event definitions used in observational analysis of administrative databases: a systematic review. BMC Med Res Methodol 2021, 21(1):241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Vukmir RB: Cardiac arrhythmia therapy. Am J Emerg Med 1995, 13(4):459–470. [DOI] [PubMed] [Google Scholar]
- 27.Bozkurt B, Coats AJ, Tsutsui H, Abdelhamid M, Adamopoulos S, Albert N, Anker SD, Atherton J, Bohm M, Butler J et al. : Universal Definition and Classification of Heart Failure: A Report of the Heart Failure Society of America, Heart Failure Association of the European Society of Cardiology, Japanese Heart Failure Society and Writing Committee of the Universal Definition of Heart Failure. J Card Fail 2021. [DOI] [PubMed] [Google Scholar]
- 28.Thygesen K, Alpert JS, White HD, Joint ESCAAHAWHFTFftRoMI: Universal definition of myocardial infarction. Eur Heart J 2007, 28(20):2525–2538. [DOI] [PubMed] [Google Scholar]
- 29.Chapman AR, Adamson PD, Mills NL: Assessment and classification of patients with myocardial injury and infarction in clinical practice. Heart 2017, 103(1):10–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Correction to: An Updated Definition of Stroke for the 21st Century: A Statement for Healthcare Professionals From the American Heart Association/American Stroke Association. Stroke 2019, 50(8):e239. [DOI] [PubMed] [Google Scholar]
- 31.Cantwell AM, Singh H, Platt M, Yu Y, Lin YH, Ikeno Y, Hubbard G, Xiang Y, Gonzalez-Juarbe N, Dube PH: Kinetic multi-omic analysis of responses to SARS-CoV-2 infection in a model of severe COVID-19. J Virol 2021:JVI0101021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Xu T, Ding W, Ji X, Ao X, Liu Y, Yu W, Wang J: Oxidative Stress in Cell Death and Cardiovascular Diseases. Oxid Med Cell Longev 2019, 2019:9030563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lin YH, Platt M, Gilley RP, Brown D, Dube PH, Yu Y, Gonzalez-Juarbe N: Influenza Causes MLKL-Driven Cardiac Proteome Remodeling During Convalescence. Circ Res 2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Beno SM, Riegler AN, Gilley RP, Brissac T, Wang Y, Kruckow KL, Jadapalli JK, Wright GM, Shenoy AT, Stoner SN et al. : Necroptosis inhibition prevents long-term cardiac damage during pneumococcal pneumonia and invasive disease. J Infect Dis 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Reyes LF, Restrepo MI, Hinojosa CA, Soni NJ, Anzueto A, Babu BL, Gonzalez-Juarbe N, Rodriguez AH, Jimenez A, Chalmers JD et al. : Severe Pneumococcal Pneumonia Causes Acute Cardiac Toxicity and Subsequent Cardiac Remodeling. Am J Respir Crit Care Med 2017, 196(5):609–620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Islam SJ, Malla G, Yeh RW, Quyyumi AA, Kazi DS, Tian W, Song Y, Nayak A, Mehta A, Ko YA et al. : County-Level Social Vulnerability is Associated With In-Hospital Death and Major Adverse Cardiovascular Events in Patients Hospitalized With COVID-19: An Analysis of the American Heart Association COVID-19 Cardiovascular Disease Registry. Circ Cardiovasc Qual Outcomes 2022, 15(8):e008612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Mai F, Del Pinto R, Ferri C: COVID-19 and cardiovascular diseases. J Cardiol 2020, 76(5):453–458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mahendran AJ, Gupta NK, Gupta N, Kumar R, Ish P: COVID-19 - deliberating beyond steroids in 2021. Adv Respir Med 2021, 89(6):612–614. [DOI] [PubMed] [Google Scholar]
- 39.Dhanyalayam D, Thangavel H, Lizardo K, Oswal N, Dolgov E, Perlin DS, Nagajyothi JF: Sex Differences in Cardiac Pathology of SARS-CoV2 Infected and Trypanosoma cruzi Co-infected Mice. Front Cardiovasc Med 2022, 9:783974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Fox SE, Akmatbekov A, Harbert JL, Li G, Quincy Brown J, Vander Heide RS: Pulmonary and cardiac pathology in African American patients with COVID-19: an autopsy series from New Orleans. Lancet Respir Med 2020, 8(7):681–686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kadosh BS, Garshick MS, Gaztanaga J, Moore KJ, Newman JD, Pillinger M, Ramasamy R, Reynolds HR, Shah B, Hochman J et al. : COVID-19 and the Heart and Vasculature: Novel Approaches to Reduce Virus-Induced Inflammation in Patients With Cardiovascular Disease. Arterioscler Thromb Vasc Biol 2020:ATVBAHA120314513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Brodin P: Immune determinants of COVID-19 disease presentation and severity. Nat Med 2021, 27(1):28–33. [DOI] [PubMed] [Google Scholar]
- 43.Gupta A, Madhavan MV, Sehgal K, Nair N, Mahajan S, Sehrawat TS, Bikdeli B, Ahluwalia N, Ausiello JC, Wan EY et al. : Extrapulmonary manifestations of COVID-19. Nat Med 2020, 26(7):1017–1032. [DOI] [PubMed] [Google Scholar]
- 44.Vabret N, Britton GJ, Gruber C, Hegde S, Kim J, Kuksin M, Levantovsky R, Malle L, Moreira A, Park MD et al. : Immunology of COVID-19: Current State of the Science. Immunity 2020, 52(6):910–941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bertheloot D, Latz E, Franklin BS: Necroptosis, pyroptosis and apoptosis: an intricate game of cell death. Cell Mol Immunol 2021, 18(5):1106–1121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Murphy JM, Czabotar PE, Hildebrand JM, Lucet IS, Zhang JG, Alvarez-Diaz S, Lewis R, Lalaoui N, Metcalf D, Webb AI et al. : The pseudokinase MLKL mediates necroptosis via a molecular switch mechanism. Immunity 2013, 39(3):443–453. [DOI] [PubMed] [Google Scholar]
- 47.Fauster A, Rebsamen M, Huber KV, Bigenzahn JW, Stukalov A, Lardeau CH, Scorzoni S, Bruckner M, Gridling M, Parapatics K et al. : A cellular screen identifies ponatinib and pazopanib as inhibitors of necroptosis. Cell Death Dis 2015, 6:e1767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nogusa S, Thapa RJ, Dillon CP, Liedmann S, Oguin TH 3rd, Ingram JP, Rodriguez DA, Kosoff R, Sharma S, Sturm O et al. : RIPK3 Activates Parallel Pathways of MLKL-Driven Necroptosis and FADD-Mediated Apoptosis to Protect against Influenza A Virus. Cell Host Microbe 2016, 20(1):13–24. [DOI] [PMC free article] [PubMed] [Google Scholar]