

Abstract
Parasites are integral parts of ecosystem function and important drivers of evolutionary processes. Characterizing ectoparasite diversity is fundamental to studies of host–parasite interactions, evolution, and conservation, and also for understanding emerging disease threats for some vector borne pathogens. With more than 1400 species, bats represent the second most speciose mammalian clade, but their ectoparasite fauna are poorly known for most species. We sequenced mitochondrial Cytochrome Oxidase C subunit I and nuclear 18S ribosomal gene fragments, and used Bayesian phylogenetic analyses to characterize ectoparasite taxon identity and diversity for 17 species of parasitized bats sampled along the Baja California peninsula and in Northwestern Mexico. The sequence data revealed multiple novel lineages of bat bugs (Cimicidae), flies (Nycteribiidae and Streblidae), and ticks (Argasidae). Within families, the new linages showed more than 10% sequence divergence, which is consistent with separation at least at the species level. Both families of bat flies showed host specificity, particularly on Myotis species. We also identified new records for the Baja peninsula of one tick (Carios kelleyi), and of five Streblid bat fly species. One Nycteribiid bat fly haplotype from Pallid bat (Antrozous pallidus) hosts was found throughout the peninsula, suggesting potential long distance co‐dispersal with hosts. Different bat bug and tick communities were found in the north and south of the peninsula. This study is the first systematic survey of bat ectoparasites in the Baja California peninsula, revealing novel lineages that are highly genetically differentiated from other parts of North America. For some ectoparasite species, haplotype distributions may reflect patterns of bat migration. This work is a first step in characterizing ectoparasite diversity over the Baja California peninsula, and understanding how ecological and evolutionary interactions shape bat ectoparasite communities among host species in different parts of their ranges.
Keywords: 18S, Argasidae, Chiroptera, Cimicidae, COI, DNA barcoding, ectoparasites, Nycteribiidae, phylogeny, Streblidae
Parasites are integral parts of the ecosystem. Using phylogenetic techniques, we found multiple novel lineages of bat bugs, flies, and ticks in Northwestern Mexico. For some ectoparasite species, haplotype distribution may reflect patterns of bat dispersal.
1. INTRODUCTION
Characterizing ectoparasite diversity is fundamental to studies of host–parasite interactions, parasite evolution and conservation, and for understanding emerging disease threats for some vector borne pathogens (Morand et al., 2006; Poulin, 2014; Spencer & Zuk, 2016). Bat ectoparasites are of particular interest as model systems for host–parasite co‐evolution and parasite community structure studies (Gómez & Nichols, 2013; Spencer & Zuk, 2016). Bat–ectoparasite relationships are also important for understanding bat dispersal, pathogen transmission, and zoonotic disease risks (Klimpel & Mehlhorn, 2014; Speer et al., 2019; Wilder et al., 2015). Despite the widely recognized need to increase sampling effort for pathogen/parasite discovery in bats, bat ectoparasites are still understudied in most parts of the world (Gay et al., 2014; Reinhardt & Siva‐Jothy, 2007). Bat‐associated ectoparasites include Insecta, comprising bat bugs (Hemiptera), fleas (Siphonaptera), and flies (Diptera); and Arachnida comprising ticks (Ixodida) and mites (orders Mesostigmata and Trombidiformes) (Seneviratne et al., 2009; Ter Hofstede & Fenton, 2005). All these ectoparasites are hematophagous (blood feeding) organisms, creating potential zoonotic disease transmission risks for humans and domestic animals (Poulin, 2014).
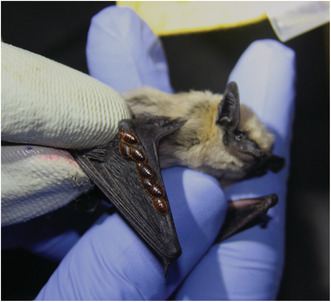
Bat bugs (family Cimicidae), have a worldwide distribution and comprise approximately 110 known species within 24 genera, the majority of which are ecologically and biologically associated with bats (Hornok et al., 2017; Ossa et al., 2019). Figure 1 shows a Parastrellus hesperus individual sampled for this study in northwestern Mexico, infested by bat bugs. A second, less studied family, Polyctenidae also includes bat‐associated bugs, with tropical and subtropical distributions (Klimpel & Mehlhorn, 2014).
FIGURE 1.

Specimen of canyon bat (Parastrellus hesperus) infested by bat bugs, sampled for this study in northwestern Mexico. Image credits: The authors, University of Leeds.
Bat flies comprise two main families, Nycteribiidae and Streblidae, which have a common origin from a single lineage coevolving with bats (Dittmar et al., 2006; Wenzel & Tipton, 1966). As of 2006, there were approximately 520 species described (Dittmar et al., 2006). With new records added regularly (Saldaña‐Vázquez et al., 2019; Szentiványi et al., 2019), taxonomy at the species level is often ambiguous, and an updated global review and synthesis is yet to be produced.
Ticks are grouped in three families: Argasidae (soft ticks), Ixodidae (hard ticks), and Nuttalliellidae. The latter family comprises a single known species, Nuttalliella namaqua (Burger et al., 2014; Mans et al., 2012), which has been suggested as the basal tick lineage (Mans et al., 2012). There are approximately 894 known species of ticks worldwide, with 32 species of Argasidae and 68 species of Ixodidae in Mexico (Guglielmone et al., 2010; Pérez et al., 2014). Bat tick phylogenetic studies have mainly focused on old world species (Hornok et al., 2017), with a limited number for the Americas and other parts of the world (Black et al., 1997; Burger et al., 2014), and suggest bat‐associated ticks commonly exhibit host‐specificity (Sándor et al., 2019). Classification of taxonomic relationships among soft ticks are controversial (Burger et al., 2014), and more studies are needed to accurately describe their taxonomic status (Burger et al., 2014; Estrada‐Pena et al., 2010).
Previous bat ectoparasite studies in North America have described the distribution and taxonomic status of bat flies, bugs, and ticks (Bradshaw & Ross, 1961; Graciolli et al., 2007; Jobling, 1938; Usinger, 1966), and their medical importance (Dick et al., 2003; Gill et al., 2004; Steinlein et al., 2001), from a limited number of bat host species. In Mexico, publications have focused on describing species diversity of flies, ticks, and mites (Bolaños‐García et al., 2018; Guzmán‐Cornejo et al., 2017; Pérez et al., 2014); new records of bat flies (Cuxim‐Koyoc et al., 2015; Ramírez‐Martínez et al., 2016; Trujillo‐Pahua & Ibáñez‐Bernal, 2018); ecological relationships between bats and bat flies (Saldaña‐Vázquez et al., 2019; Salinas‐Ramos et al., 2018; Zamora‐Mejías et al., 2020); and Rickettsia presence in soft ticks (Sánchez‐Montes et al., 2016). To our knowledge, there have been no studies of bat bugs in Mexico.
Baja California (Figure 2) is an isolated peninsula, 1300 km long, in northwestern Mexico. Its complex geological history means it has a mosaic of habitats, with high levels of biodiversity and endemism (González‐Abraham et al., 2010; Nájera‐Cortazar et al., 2015; Riddle et al., 2000), with approximately 25 species of bats recorded (Álvarez‐Castañeda et al., 2015; Medellín et al., 2007). While there are several studies describing Baja bat diversity, distributions, and ecology (Álvarez‐Castañeda et al., 2015; Frick et al., 2008; Nájera‐Cortazar et al., 2015), there are no studies describing the bat ectoparasite fauna.
FIGURE 2.
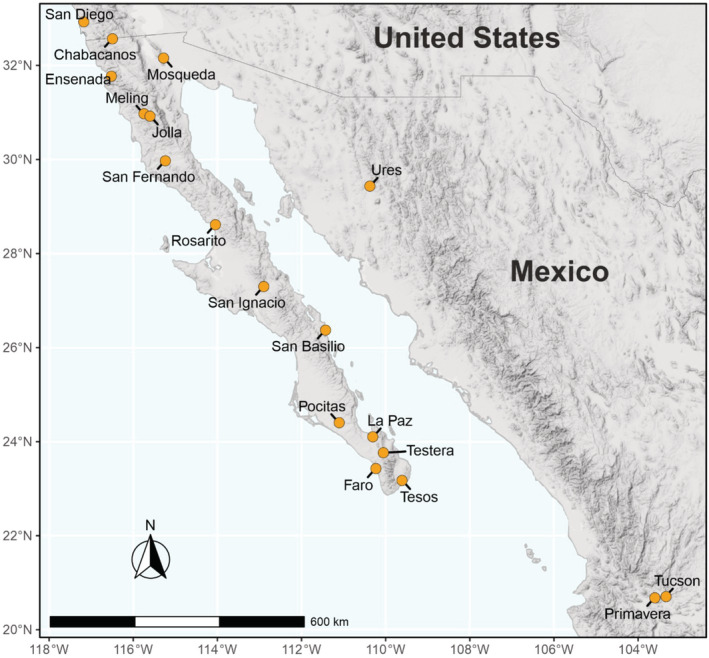
Map of sampling locations for bats and bat ectoparasites reported in this study.
Here, we use a phylogenetic approach to characterize ectoparasite (bat bugs, flies, and ticks) identity and diversity for 17 species of bat resident along the Baja California peninsula, and northwestern Mexico, based on mitochondrial Cytochrome Oxidase C subunit I (COI) and nuclear 18S ribosomal (18S) DNA amplicon sequences. Ectoparasites are assigned as known or novel lineages within Baja and the rest of the Americas, and we evaluate potential distributions with respect to Baja, and their hosts' wider ranges. The results are relevant for increasing knowledge of bat and ectoparasite distributions across western North America, and to provide a basis for understanding how ecological and evolutionary interactions shape parasite community structure along environmental gradients such as those found in northwestern Mexico.
2. METHODS
2.1. Sample collection
Bat sampling and collection of ectoparasites were conducted at 14 sites along the Baja California peninsula, and at three sites in western continental Mexico during 2016–2018 (Figure 2). Bats were identified to species level in the field following published identification guides (Álvarez‐Castañeda et al., 2015; Medellín et al., 2007), and subsequently, Myotis bats were identified by molecular assays (Najera‐Cortazar, 2020). A list of sites and bat species sampled is given in Table 1. Ectoparasites were collected manually from bats using forceps and transferred to labeled Eppendorf tubes with 96% ethanol for storage. Ectoparasites collected from the same bat but from different taxonomic families were stored in separate tubes for each family, with corresponding labels and appropriate specimen source identifiers. All samples were kept on ice during fieldwork, until their arrival to the laboratory where they were stored at −20°C. Ectoparasites were photographed during fieldwork using a portable Maozua 5MP 20‐300X USB microscope, taking ventral and dorsal views. Preliminary classifications of ectoparasites were assigned according to morphological characters specified in keys by McDaniel (1979), Usinger (1966), Knee and Proctor (2006) and Dick and Miller (2010), and adapted for North American ectoparasites by this study. In addition, 10 specimens of bat bugs from Parastrellus hesperus hosts, captured at Ramona, California, U.S.A. (Site 0), were donated from the San Diego Natural History Museum, California U.S.A. Since the focus of this study was a molecular barcoding and phylogenetic approach, morphologies are not described in detail, and will be presented elsewhere.
TABLE 1.
Left section: List of sites sampled showing number, name, abbreviation (site label) and geographic coordinates. Right section: List of bat species sampled and their abbreviated species labels. Myotis sp. refers to any unidentified Myotis morphologically resembling M. velifer, found at Ures site.
| Site number | Site name | Site label | Latitude | Longitude | Bat species | Species label |
|---|---|---|---|---|---|---|
| Baja Caliifornia Peninsula | ||||||
| 0 | San Diego | SanDi | 32.927 | −117.176 | Antrozous pallidus | ANPA |
| 1 | Chabacanos | Chaba | 32.566 | −116.493 | Artibeus hirsutus | ARHI |
| 2 | Mosqueda | Mosq | 32.156 | −115.279 | Choeronycteris mexicana | CHME |
| 3 | Ensenada | Ense | 31.770 | −116.520 | Eptesicus fuscus | EPFU |
| 4 | Meling | Meli | 30.972 | −115.744 | Glossophaga soricina | GLSO |
| 5 | Jolla | Jolla | 30.920 | −115.601 | Leptonycteris yerbabuenae | LEYE |
| 6 | San Fernando | SanFe | 29.971 | −115.237 | Macrotus californicus | MACA |
| 7 | Rosarito | Rosa | 28.613 | −114.047 | Mormoops megalophylla | MOME |
| 8 | San Ignacio | SanIg | 27.297 | −112.898 | Myotis californicus | MYCA |
| 9 | San Basilio | SanBa | 26.371 | −111.429 | Myotis peninsularis | MYPE |
| 10 | Pocitas | Poza | 24.403 | −111.104 | Myotis sp | MYsp |
| 11 | La Paz | LaPaz | 24.103 | −110.306 | Myotis velifer | MYVE |
| 12 | Testera | Teste | 23.764 | −110.055 | Myotis vivesi | MYVI |
| 13 | Faro | Faro | 23.427 | −110.233 | Myotis volans | MYVO |
| 14 | Tesos | Teso | 23.175 | −109.611 | Myotis yumanensis | MYYU |
| Continental | Parastrellus hesperus | PAHE | ||||
| 15 | Ures | Ures | 29.433 | −110.376 | Sturnira parvidens | STPA |
| 16 | Tucson | Tucs | 20.705 | −103.336 | ||
| 17 | Primavera | Prima | 20.679 | −103.602 | ||
2.2. DNA extraction, primer selection, and PCR amplification
Individual ectoparasite bodies were crushed in Eppendorf tubes using sterile pestles, followed by digestion with Proteinase K at 56°C for 18 h. DNA was extracted using either a Thermo‐Fisher DNA extraction and purification kit (Thermo Fisher Scientific Inc.) or QuickExtract kit (Epicenter, Illumina), following the manufacturers' protocols. PCR conditions were optimized separately for each ectoparasite group, and according to each primer set. Primers LCO1490 (5′‐GGTCAACAAATCATAAAGATATTGG‐3) and HCO2198 (5′‐TAAACTTCAGGGTGACCAAAAAATCA‐3′) (Folmer et al., 1994) were used to amplify an approximately 700 bp fragment of the mitochondrial COI gene in a 25 μl reaction containing: 1 U of Flexi GoTaq Taq Polymerase, 5× GoTaq reaction buffer, 50 mM MgCl2 (1.5–2.5 mM final concentration), 0.5 μl PCR nucleotide Mix (0.2 mM each), 0.5 μl of each set of primers (1 μM final concentration), 15.8 μl ddH2O, and 8 μl template DNA (extractions diluted at 1:10 with sterile distilled water). Thermal cycling parameters were as follows on a TECHNE thermocycler model TC‐512: initial denaturation step at 95°C for 5 min, followed by 40 cycles of denaturation at 94°C for 40 s, annealing at 53°C to 56°C for 1 min and extension at 72°C for 1 min. Final extension was performed at 72°C for 10 min.
For the 18S rDNA gene, approximately 800 bp amplicons were amplified using the primers 18S‐1F (5′ CTGGTTGATCCTGCCAGTAGT 3′) and 18S‐3R (5′ GGTTAGAACTAGGGCGGTATCT 3′) for bat bugs (Campbell et al., 1995); a0.7‐F (5′ ATTAAAGTTGTTGCGGTT 3′) and 7R (5′ GCATCACAGACCTGTTATTGC 3′) for flies (Whiting, 2002); and D‐F (5′ GGCCCCGTAATTGGAATGAGTA 3′) and C‐R (5′ CTGAGATCCAACTACGAGCTT 3′) for ticks (Mangold et al., 1997). The same reaction mix quantities were used as for the COI gene, with MgCl2 at a final concentration of 1.5 mM for all primer sets. Thermal cycling parameters were: initial denaturation at 95°C for 5 min, followed by 30 cycles of denaturation at 94°C for 40 s, annealing at 53°C for 1 min and extension at 72°C for 1 min. Final extension was performed at 72°C for 10 min.
PCR products were visualized by gel electrophoresis using 1% agarose with GelRed® (Biotium Ltd) staining, and then sent to Genewiz Inc. (Azenta Life Sciences), for PCR product purification and Sanger sequencing, with each amplicon sequenced in both forward and reverse directions.
2.3. Quality control and reference sequences
Sequence quality for both the forward and reverse strands of each amplicon was evaluated in BioEdit 7.2.5 (Hall, 2005). Trimmed forward and reverse sequences were combined to generate a consensus sequence for each amplicon, and then analyzed in BLAST (The National Library of Medicine, 2018) to generate initial taxon identities and identify reference sequences. Additionally, COI sequences were also analyzed in the BOLD system platform (Ratnasingham & Hebert, 2007). Reference sequences for phylogenetic analyses were compiled from previous studies on each ectoparasite group (Burger et al., 2014; Dittmar et al., 2006; Mans et al., 2012; Tortosa et al., 2013), and by performing a systematic search using AnnotationBustR 1.2 package (Borstein, 2018) in RStudio 1.1.456 (RStudio Team, 2021), searching for the closest genus for the sequences generated in this study and for those obtained by BLAST. Alignments combining reference sequences and those from this study were generated using CLUSTAL W (Thompson et al., 1994), implemented in BioEdit, and were reviewed by eye, with manual correction of potential errors as required.
2.4. Sequence summary statistics and phylogenetic analyses
Genetic distance estimates among haplotypes and best fit sequence evolution models for the COI and 18S datasets were evaluated using MEGA 10.1.7 (Sudhir et al., 2018). The Barcode Index Number system was followed to delimit a lineage, where intraspecific variation at COI is generally considered as groups of sequences with less than 2% divergence, and exhibiting more than 4% divergence from neighboring lineages (Hebert et al., 2003; Ratnasingham & Hebert, 2013; Salinas‐Ramos et al., 2018).
Bayesian phylogenetic analyses were performed using the best fit evolution model identified for each group and each marker implemented with BEAST 1.10.4 (Suchard et al., 2018). A MCMC chain length of 10,000,000 was used and priors specific to each parasite group and sequence evolution model were selected using the program BEAUti 1.10.4 (Suchard et al., 2018). Phylogenetic analyses were performed separately for each ectoparasite group and each marker. Nycteribiidae and Streblidae families of bat flies were analyzed separately in order to improve sequence alignment quality and phylogenetic resolution. For each family‐marker combination, two separate runs using the same Bayesian settings file generated by BEAUti were run in BEAST, with a burn in of the 10% of the total number of iterations. After, stationarity of BEAST results were assessed in Tracer 1.7.1 (Rambaut et al., 2018). Both files for each ectoparasite family‐marker combination were combined using LogCombiner 1.10.4 (Suchard et al., 2018), generating a single .log file and a single .tree file. A majority‐rule consensus tree was inferred using TreeAnnotator 1.10.4 (Suchard et al., 2018) with the combined .tree file, and using a posterior probability limit of 0.6, median nodes heights were summarized. Phylogenetic trees were annotated, edited, and visualized using iTOL 5.6.2 (Letunic & Bork, 2016).
Following phylogenetic analysis, haplotypes for each family were grouped by lineage, and haplotype diversity and genetic summary statistics were calculated in DNAsp 5.10.01 (Librado & Rozas, 2009). Outgroups were chosen based on previous phylogenetic studies of each group, using Bucimex chilensis, Primicimex cavernis, and Anthocoris flavipes for bat bugs (Ossa et al., 2019); Drosophila melanogaster, Chrysops niger, Musca domestica, and Sarcophaga bullata (Dittmar et al., 2006), for both bat fly families, plus Ornithomya avicularia for the family Streblidae only; and Nuttalliella namaqua (Mans et al., 2012) for ticks.
2.5. Ethics approval
All bat capture, handling and sampling was carried out with the approval of the ethics committee of the Faculty of Biological Sciences, University of Leeds (AWCNRW170615); and following the Guidelines of the American Society of Mammalogists (Sikes et al., 2016). Sampling was carried out under the permits SEMARNAT‐DGVS‐008972‐16 and SEMARNAT‐DGVS‐001642‐18 issued by Secretaría del Medio Ambiente y Recursos Naturales (SEMARNAT) in Mexico. The latter included two Myotis bat species listed in the Mexican Official Norm for the protection of native species of flora and fauna in Mexico [NOM‐059‐SEMARNAT‐2010, (Secretaría de Medio Ambiente y Recursos Naturales, 2010)], under the Pr (under special protection) and P (in danger of extinction) categories (M. evotis and M. vivesi, respectively); and sampling on protected reserves. When required, permission was also solicited and granted from private land owners. All samples were imported into the UK under permission of the Department for Environment, Food and Rural Affairs (DEFRA) from the Animal and Plant Health Agency (permit ITIMP19.0036).
3. RESULTS
3.1. Sampling and field identifications
A total of 1988 ectoparasites were collected within the orders Diptera (flies), Hemiptera (bugs), Ixodida (ticks), Mesostigmata (mites), Siphonaptera (fleas), and Trombidiformes (chiggers). Fleas, chiggers, mites, and any unclassified specimens (e.g., where it was not possible to differentiate small tick larvae and mites) were excluded for this study, to be evaluated separately. The remaining samples comprised 90 bat bugs, 213 bat flies, and 126 ticks, collected from 138 individual bat hosts of 17 species (Figure 3).
FIGURE 3.
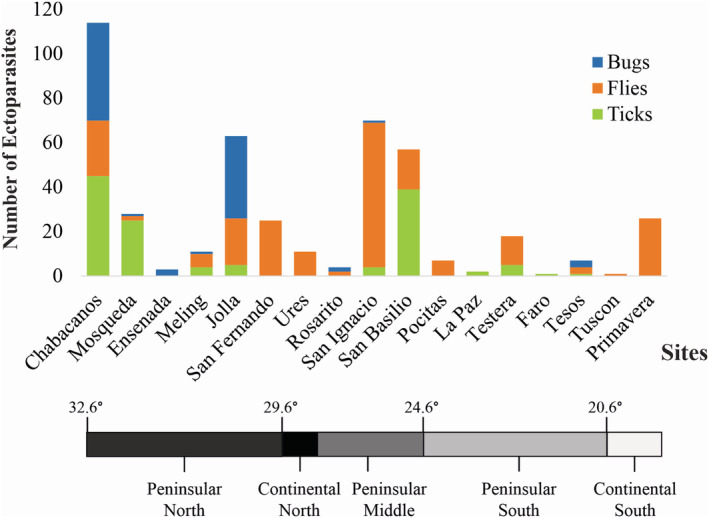
Number of bat bugs, flies, and ticks captured per site in 2016–2018 fieldwork.
Flies were the predominant ectoparasite type, found in 13 of 17 sites, followed by ticks and bugs, present in eight sites each. Initial field morphological evaluations suggested most bug specimens were related to Cimex pilosellus (Usinger, 1966). Most bat flies could be identified to the genus level, which were later corroborated with molecular data. On the basis of morphology, all bat ticks were identified as family Argasidae (soft ticks), with at least six different morphotypes present. Most of these were tentatively attributed to the genus Ornithodoros.
3.2. BLAST and BOLD sequence matches
Amplicon sequences were generated from 145 specimens for mitochondrial COI and from 147 specimens for 18S rDNA across the three groups: bugs n = 30/30 (COI/18S, respectively); flies n = 76/73; and ticks n = 39/44. When there was more than one ectoparasite specimen available per ectoparasite family per bat, one specimen for sequencing was selected based on morphological similarity, sequencing one individual of each morphotype of each family. No haplotypes from bugs and most tick specimens matched existing sequences deposited in GenBank or BOLD at the species level (divergence >4%), with the exception of one of the tick lineages showing similarity to Carios kelleyi (96.95% in GenBank, 97.07% in BOLD). Similarly for nycteribiid bat flies, there were no species level matches for neither COI nor 18S markers. In the case of streblid bat flies, 14 sequences matched (~99%–100% in GenBank‐BOLD databases) with five known species for COI (see Table 2); but there were no matches for 18S. These marker differences are likely attributable to database coverage, since where reference sequences were available for COI and 18S from the same specimen/study, BLAST results for our sequences were consistent for both markers. For brevity and to provide comparability with larger numbers of reference sequences further reporting of diversity, divergence statistics, and taxonomic identity will be focused on COI results.
TABLE 2.
Molecular summary statistics for the ectoparasites lineages identified in this study
| Lineage/putative genus | N | S | H | H d | N d | Host(s) | Closest reference species | Min. % div. | Known congeneric ectoparasite species for host(s) when no molecular match | |
|---|---|---|---|---|---|---|---|---|---|---|
| Bat bugs: Cimicidae | Cimex 1 | 1 | 0 | 1 | 0 | 0 | Antrozous pallidus | Cimex latipennis | 5.1 | C. incrassatus, C. pilosellus 1 |
| Cimex 2 | 2 | 4 | 2 | 1 | 0.006 | Myotis californicus | Cimex latippenis | 3.2 | Cimex adjunctus, C. antennatus, C. brevis, C. pilosellus, C. latippenis 1 | |
| Cimex 3 | 5 | 2 | 2 | 0.4 | 0.001 | Antrozous pallidus, Myotis californicus | Cimex adjuntus | 5.9 | C. incrassatus, Paracimex cavernis 1 | |
| Cimex 4 | 22 | 14 | 17 | 0.97 | 0.005 | Parastrellus hesperus, Antrozous pallidus | Cimex antennatus | 4.4 | C. antennatus, C. incrassatus, C. pilosellus 1,8 | |
| Bat flies: Nycteribiidae | Nycteribiid 1 | 5 | 6 | 5 | 1 | 0.003 | Myotis sp Myotis californicus Parastrellus hesperus | Nycteribia pygmaea | 9.4 | Basilia antrozoi, B. corynborhini, B. forcipata, B. rondani 6,8 |
| Nycteribiid 2 | 12 | 22 | 8 | 0.89 | 0.007 | Myotis volans Antrozous pallidus Myotis californicus Myotis yumanensis | Nycteribia pygmaea | 9.4 | Basilia antrozoi, B. corynborhini, B. forcipata, B. jellisoni, B. rondani 5,6,8 | |
| Basilia 1 | 3 | 0 | 1 | 0 | 0 | Myotis yumanensis | Basilia boardmani | 5.0 | Basilia forcipata, B. jellisoni, B. rondani 5, 6 | |
| Basilia 2a | 3 | 1 | 2 | 0.67 | 0.0009 | Myotis vivesi | Basilia ortizi | 9.5 | Basilia plaumanni, B. pynzonix, B. producto 6 | |
| Basilia 2b | 26 | 4 | 5 | 0.58 | 0.001 | Antrozous pallidus | Basilia ortizi. | 8.6 | Basilia antrozoi, B. rondani 6 | |
| Bat flies: Streblidae | Aspidoptera phyllostomatis | 2 | 0 | 1 | 0 | 0 | Sturnira parvidens | Aspidoptera phyllostomatis | 2.3 | – |
| Megistopoda aranea | 1 | 0 | 1 | 0 | 0 | Sturnira parvidens | Megistopoda aranea | 0.8 | – | |
| Megistopoda sp | 2 | 1 | 2 | 1 | 0.0015 | Artibeus hirsutus | Megistopoda aranea | 3.0 | Aspidoptera phyllostomatis, Paratrichobius longicrus 8 | |
| Nycterophilia coxata | 9 | 9 | 4 | 0.75 | 0.006 | Leptonycteris yerbabuenae Macrotus californicus | Nycterophilia coxata | 0.0 | – | |
| Paratrichobius 1 | 1 | 0 | 1 | 0 | 0 | Artibeus hirsutus | Paratrichobius longicrus | 4.3 | Aspidoptera phyllostomatis, Paratrichobius longicrus 3 | |
| Streblid 1 | 4 | 15 | 3 | 0.83 | 0.011 | Macrotus californicus | Trichobius sparsus | 10.3 | Trichobius adamsi, Nycterophilia coxata 8 | |
| Trichobius 1 | 2 | 6 | 2 | 1 | 0.008 | Mormoops megalophylla | Trichobius sphaeronotus | 6.0 | Trichobius sphaeronotus, T. galei, T. leionotus, Nycterophilia mormoopsis 7 | |
| Trichobius 2 | 2 | 0 | 1 | 0 | 0 | Choeronycteris mexicana | Trichobius dugesii | 5.6 | Trichobius mixtus, Paratrichobius longicrus 8 | |
| Trichobius intermedius | 1 | 0 | 1 | 0 | 0 | Artibeus hirsutus | Trichobius intermedius | 3.1 | – | |
| Trichobius dugesii | 1 | 0 | 1 | 0 | 0 | Glossophaga soricina | Trichobius dugesii | 0.2 | – | |
| Trichobius sphaeronotus | 2 | 0 | 1 | 0 | 0 | Leptonycteris yerbabuenae | Trichobius sphaeronotus | 0.0 | – | |
| Bat Ticks: Argasidae | Antricola 1 | 1 | 0 | 1 | 0 | 0 | Macrotus californicus Myotis vivesi | Antricola marginatus | 10.7 | Ornithodoros sp.8 |
| Carios kelleyi | 10 | 33 | 9 | 0.98 | 0.011 | Antrozous pallidus | Carios kelleyi | 1.9 | – | |
| Tick 1 | 3 | 2 | 3 | 1 | 0.002 | Antrozous pallidus Myotis peninsularis | Ornithodoros turicata | 12.3 | Ornithodoros sp., O. kelleyi, O. rossi 8,9 | |
| Tick 2 | 2 | 1 | 2 | 1 | 0.001 | Antrozous pallidus Myotis yumanensis | Ornithodoro faccinii | 13.2 | Ornithodoros kelleyi, O. yumatensis, O. rossi 5,8,9 | |
| Tick 3 | 6 | 15 | 4 | 0.8 | 0.008 | Myotis yumanensis | Carios vespertilionis | 13.1 | Ornithodoros kelleyi, O. yumatensis 5,8 | |
| Tick 4 | 3 | 72 | 3 | 1 | 0.071 | Antrozous pallidus Eptesicus fuscus | Ornithodoro faccinii | 11.6 | Ornithodoros sp., O. kelleyi, O. rossi 8,9 | |
| Tick 5 | 3 | 16 | 8 | 0.93 | 0.005 | Myotis yumanensis Antrozous pallidus Parastrellus hesperus Eptesicus fuscus | Antricola marginatus | 11.8 | Ornithodoros sp., O. kelleyi, O. rossi, O. yumatensis 8,9 | |
| Tick 6 | 3 | 1 | 2 | 0.67 | 0.001 | Myotis peninsularis | Ornithodoros yumatensis | 9.9 | No previous record to our knowledge | |
Note: N, number of sequences within each lineage; S, number of segregating sites; H, number of haplotypes; H d, haplotype diversity; N d, nucleotide diversity, and Min. % Div., minimum percentage sequence divergence from closest reference sequence. 1. Usinger (1966), 2. Cuxim‐Koyoc et al. (2015), 3. Trujillo‐Pahua and Ibáñez‐Bernal (2018), 4. Zamora‐Mejías et al. (2020), 5. Braun et al. (2015), 6. Graciolli et al. (2007), 7. Ramírez‐Martínez et al. (2016), 8. Bradshaw and Ross (1961), 9. Steinlein et al. (2001).
3.3. Genetic diversity
Genetic diversity statistics for each COI lineage in each ectoparasite family are summarized in Table 2. Excluding four lineages represented by single individuals (e.g., Cimex 1), and three lineages including only individuals with the same haplotype (e.g., Basilia 1), nucleotide diversity (N d) ranged from 0.001 to 0.006, with the highest value 0.071, presented by the Tick 4 lineage, with three haplotypes (H) in three sequences (Table 2). In general, the number of haplotypes were close to or the same as the number of sequences tested per lineage.
3.4. Phylogenetic assessment of Baja peninsula bat bug sequences
The best fit evolution model for the COI gene set was GTR + G + I, and K2 + G + I for 18S. For each marker, 30 sequences were generated, forming four novel lineages with respect to reference sequences (Figure 4). Genetic divergence between the four peninsular lineages ranged from 9.9% to 17.1% (Appendix 1), and between 7.2% and 20.9% against reference sequences, with C. latipennis, C. antennatus, and C. adjuntus presenting the lowest divergence values. Phylogenetically, all the lineages sit within the Pilosellus complex, of North American members of the Cimex genus.
FIGURE 4.
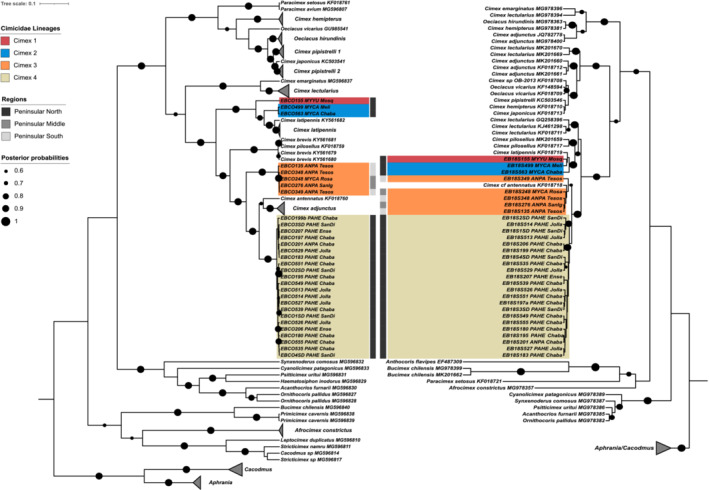
Bayesian phylogenetic trees for bat bugs of the family Cimicidae, obtained using mitochondrial COI (left) and the ribosomal 18S (right) sequences. Posterior probability support is indicated by the size of black circles at tree nodes. Where available, information for location and host is written next to each reference sequence label. To improve clarity of the tree, collapsed clades of reference sequences are shown as gray triangles. GenBank accession numbers for reference sequences are given in the sequence labels.
For COI, Cimex 1 is represented by a single specimen (EBCO155) obtained from a Myotis yumanensis host at Mosqueda, and forms a sister lineage to Cimex 2, represented by two haplotypes from two bugs parasitizing M. californicus hosts, which were also sampled from northern sites. The 18S analysis also placed EB18S155 in a separate clade, but Cimex 2 is paraphyletic. Both markers placed Cimex latipennis, which is distributed from Canada to the north western USA (Usinger, 1966), as the closest named molecular reference species to these 2 lineages. Cimex 3 is represented by a monophyletic clade for COI, consisting of two haplotypes, sampled from specimens collected from four Antrozous pallidus and one M. californicus hosts, all distributed in the southern half of the peninsula (sites Rosarito, San Ignacio and Tesos). For 18S, a C. cf antennatus reference sequence nests within the clade with greater than 60% of posterior probability support. For COI, reference sequences for C. pilosellus and C. brevis formed a clade which appeared to be ancestral to Cimex 3, with 13% divergence (Appendix 1). There is no molecular reference for C. brevis in the 18S analysis, and Cimex 4 is the closest sister lineage.
The Cimex 4 COI lineage includes 17 haplotypes derived from 22 specimens, where 21 of the bugs were found parasitizing Parastrellus hesperus individuals, and one from A. pallidus (EBCO201/EB18S201), all from northern sites (Figure 2), with C. antennatus as the closet sequence match (9.3%; Figure 4), followed by C. adjunctus. Sequences of C. adjunctus in the 18S topology clustered in a clade with other Cimex species with mixed origins, including some non‐Palearctic species. This may represent misidentification of those specimens, or mistakes in the annotation of sequences submitted to GenBank. Cimex incrassatus is reported as occurring on A. pallidus hosts but no reference sequences are available. Therefore, one of the novel bug lineages may represent this species, but further morphological assessments are required for confirmation.
3.5. Phylogenetic assessment of bat fly sequences
The best fit sequence evolution models were GTR + G for the COI gene set, and T92 + G + I for the 18S data. In total, 77 bat flies were sequenced, yielding 76 sequences for COI and 73 for 18S amplicons respectively. Individual sequences for two specimens for COI, and three for 18S did not pass sequence quality thresholds and were discarded. In the final sequence set for the COI marker, there were 49 sequences from the Nycteribiidae family (wingless bat flies) and 27 for the Streblidae family (winged bat flies), representing 10 novel lineages and six new species records for Baja. For the 18S marker, 46 sequences were generated for the Nycteribiidae family, and 26 sequences for the Streblidae family. For this marker, nine novel lineages were observed (18S sequences corresponding to the Basilia 2a COI lineage could not be amplified), along with six new species records for Baja.
In the COI phylogenetic analysis, the 49 Nycteribiidae sequences formed five lineages, all of which appeared to be novel with respect to GenBank references. Genetic divergence among Baja lineages ranged from 2.9% to 14.5%, and up to 16% against reference sequences (Appendix 2). For COI, Nycteribiid 1 and 2, formed sister clades with 4.4% divergence, and 10% divergence from the closest reference haplotypes derived from Asian Nycteribia, species. For 18S, the closest references to Nycteribiid 1 and 2 were North American species not available for COI, Basilia corynorhini and Basilia forcipata, respectively. Nycteribiid 1 and 2 lineages were primarily associated with Myotis bat hosts, but with one Nycteribiid 2 haplotype recovered from a Parastrellus hesperus host in the 18S dataset (specimen EN18S514, Figure 5).
FIGURE 5.

Bayesian phylogenetic trees for bat flies of the family Nycteribiidae obtained using mitochondrial COI (left) and the ribosomal 18S (right) sequences. Posterior probability support is indicated by the size of black circles at tree nodes. Where available, information on location and host is written next to each reference sequence label. To improve clarity of the tree, collapsed clades of reference sequences are shown as gray triangles. GenBank accession numbers for reference sequences are given in the sequence labels.
The three other Baja lineages formed clades associated with Basilia reference sequences from species recorded in Madagascar, USA, and Panama. Basilia 1 has 4.9% divergence from Basilia boardmani, a bat fly distributed throughout the United States parasitizing Myotis bats (Graciolli et al., 2007). Specimens with Basilia 1 haplotypes were sampled at mid‐peninsula, parasitizing Myotis yumanensis, which is widely distributed throughout western North America.
Genetic divergence among lineages Basilia 2a and 2b was 2.9% (Table 2), representing the threshold for intra/inter interspecific values (Hebert et al., 2003; Ratnasingham & Hebert, 2013). We assign them as distinct lineages due to their different host species, where Basilia 2a parasitized M. vivesi, and Basilia 2b appeared to be restricted to Antrozous pallidus (Figure 5). The spatial distribution of Basilia 2b haplotypes across the peninsula, suggests potential long distance co‐dispersal with their hosts A. pallidus (Figure 5), providing evidence supporting bat movement along Baja. Basilia 2b may represent B. antrozoi, which has been previously reported to parasitize A. pallidus (Graciolli et al., 2007).
The 11 Streblidae lineages had genetic divergence for COI of 0.01%–18.9% for sequences sampled within this study, and up to 21.6% against their reference sequences (Appendix 3). For the five novel lineages from this family four were named according to the closest genus in the phylogenetic analysis (Trichobius 1, Trichobius 2, Paratrichobius 1, and Megistopoda sp., Figure 6), with genetic divergence values ranging from 3% to 8.4% from each of the closest corresponding clades. The Streblid 1 lineage presented 10.8% divergence against its closest reference, T. sphaeronotus (Appendix 3, Figure 6), and is therefore not attributed to an existing genus.
FIGURE 6.
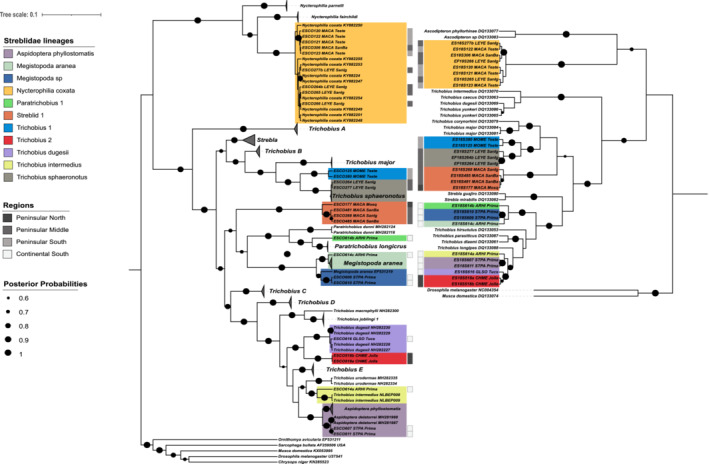
Bayesian phylogenetic trees for bat flies of the family Streblidae obtained using mitochondrial COI (left) and the ribosomal 18S (right) sequences. Posterior probability support is indicated by the size of black circles at tree nodes. Where available, information on location and host is written next to each reference sequence label. To improve clarity of the tree, collapsed clades of reference sequences are shown as gray triangles. Genbank accession numbers for reference sequences are given in the sequence labels.
The other streblid clades matched sequences from six known species: Aspidoptera phyllostomatis, Megistopoda aranea, Nycterophilia coxata, Trichobius dugesii, T. intermedius, and T. sphaeronotus (Figure 6), representing new records for these species in Baja and western Mexico. For the specimens classified as N. coxata, T. dugesii, T. intermedius, and T. sphaeronotus (Figure 6, Appendix 3), there was less than 1.5% divergence against the GenBank references. The fly lineages A. phyllostomatis matched with two reference sequences classified as A. delatorrei (Figure 6). However, A. phyllostomatis and A. delatorrei reference sequences only differed by 2.1%–2.3%, which falls at the threshold for species level differentiation. Without further information on the source A. delatorrei specimens, we decided to retain the A. phyllostomatis classification for our specimens. In the case of the M. aranea and Megistopoda sp lineages, there were two separate clades of M. aranea reference sequences present. The Megistopoda sp specimens (ESCO606 and ESCO610, dark blue clade, Figure 6) were grouped closer to M. aranea EF531219 reference sequence, with 2.9% and 3.1% divergence, suggesting a different species from the M. aranea EF531219 reference, as well as from the other M. aranea clade (bright blue clade, Figure 6), with 4.4% genetic divergence. Supporting this, it is noted that each lineage had different host species, Sturnira parvidens and Artibeus hirsutus, respectively (Table 2). Most streblid lineages were parasitizing fruit‐nectar feeding bats (Phyllostomidae) over the mid and northern peninsula, with the exception of Trichobius 1 found on Mormoops megalophylla hosts, (family Mormoopidae), an insectivore. Trichobius 1 was the only streblid fly lineage found in the south of Baja. The other fly lineages were distributed in the southern continental sites (Tucson and Primavera, Figure 2, Table 1). Outside of the closest sequence matches, none of the other species reported as parasitizing hosts in Baja (Table 2) appear to have reference sequences in GenBank. Genetic divergence within groups from the lineages in this study showed levels between 0.0% and 1.1%, with the exception of A. phyllostomatis, which showed a 3.3% divergence within its group (Appendix 3).
3.6. Phylogenetic assessment of Baja peninsula bat tick sequences
The best fit sequence evolution models were GTR + G + I for the COI gene set, and K2 + G + I for the 18S gene set. There were 45 ticks sequenced from the Argasidae family (soft ticks), with 39 sequences generated for COI, and 44 for 18S, where six specimens failed to amplify for COI (specimens ETCO157, ETCO306, ETCO338, ETCO480b, ETCO480c, and ETCO485), and one specimen failed to amplify for 18S (specimen ET18S457). One new Baja species record and seven potential novel lineages were obtained (Figure 7). The only match with GenBank and BOLD COI records (95.95% and 97.07%, respectively) was the soft tick Carios kelleyi, for 11 specimens (parasitizing A. pallidus hosts) with intra‐clade divergence of 1.94%–2.47% (Figure 7, Appendix 4). For COI data, the Antricola 1 lineage had 10.6% divergence from Antricola marginatus, and 14.6% from A. mexicanus (Appendix 4). Ornithodoros faccini had a divergence of around 11% for clades Carios kelleyi, Tick 1 to 5, and O. yumatensis showed divergence of around 9.9% with respect to Tick 6 lineage. Ticks of lineage Tick 6 were recovered from M. peninsularis from which there are no previous records of bat ticks to our knowledge.
FIGURE 7.
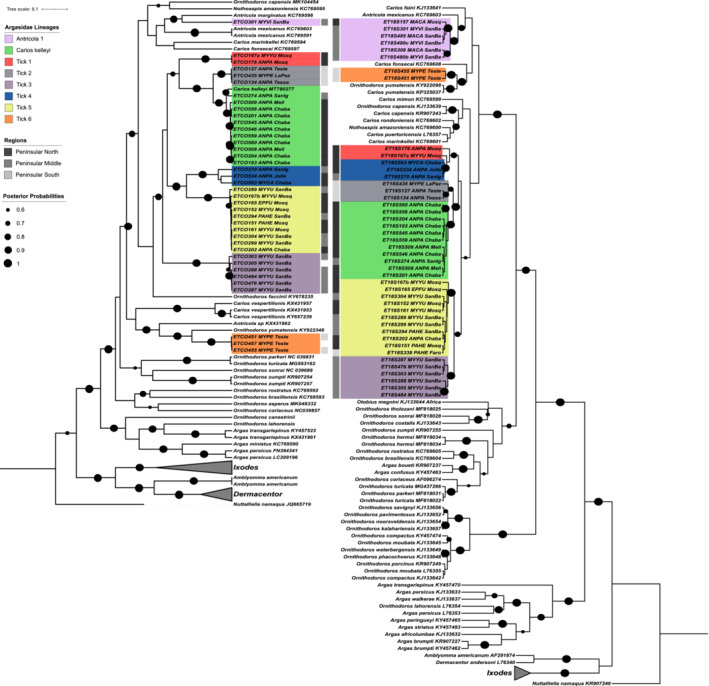
Bayesian phylogenetic trees for bat ticks of the family Argasidae obtained using mitochondrial COI (left) and the ribosomal 18S (right) sequences. Posterior probability support is indicated by the size of black circles at tree nodes. Where available, information on location and host is written next to each reference sequence label. To improve clarity of the tree, collapsed clades of reference sequences are shown as gray triangles. GenBank accession numbers for reference sequences are given in the sequence labels.
All sequences from this study generated the same lineages for both COI and 18S genes (Figure 7). Lineages C. kelleyi and Tick 1 to 5 formed a monophyletic group with respect to the reference sequences with posterior support greater than 0.6 for both markers, while Antricola 1 and Tick 6 lineages were positioned in separate clades. The topology of sister clade relationships varied slightly between markers, particularly around deeper nodes which had posterior probability support less than 0.85. This suggests more data is required to resolve deeper taxonomic relationships among species. Antricola 1 and Tick 6 were separated around shallow deep nodes in both analyses, where the absence or presence of reference sequences influenced their topological proximity (Figure 7). None of the reference sequences from Antricola, Carios, and Ornithodoros genera, form monophyletic groupings with respect to genus nomenclature.
Tick lineages 2 and 6 (gray and orange, Figure 7) were only observed in the south of the peninsula, with hosts A. pallidus and M. peninsularis, and M. peninsularis, respectively. These two species of bats share at least one confirmed roosting site within the study region, at Tesos (Figure 2), suggesting a potential interchange of host species for Tick lineage 2. Evidence of singular host specificity was observed for the lineages C. kelleyi (A. pallidus), Tick 3 (M. yumanensis), and Tick 6 (M. peninsularis), all observed exclusively on the same hosts across sites and field seasons. The Antricola 1 lineage (Figure 7, lilac clade, ETCO301_MYVI) was recovered from one specimen on a M. vivesi host using the COI marker, and grouped with Antricola marginatus (found in the South East of Mexico), followed by A. mexicanus. In the 18S topology, the 18S sequence from this sample formed a clade with five additional sequences from M. vivesi and Macrotus californicus hosts, grouped with A. mexicanus, as no A. marginatus reference was available. The two host species for ticks of this lineage were, sampled mostly at mid‐peninsula (Figures 2 and 3). Tick 5 lineage had the most diverse host and spatial distribution, being found on four bat species, and at sites from the north to the south of the peninsula (yellow clade, Figure 7).
4. DISCUSSION
This study represents the first molecular characterization and phylogenetic analysis of bat bug, bat fly, and bat tick diversity along the Baja California peninsula and northwestern Mexico, a region where the bat ectoparasite fauna is largely undescribed. From a total of 292 ectoparasite specimens, from 17 bat species, evaluated for COI and 18S markers, 21 novel genetic lineages, plus seven new species records for the region, were found. Some of the novel lineages may derive from previously recorded species with no reference sequences available, while others are likely to represent new species. Overall, the work demonstrates that the northwestern region of Mexico hosts a high diversity of previously unknown bat ectoparasites.
4.1. Bat bugs
Four novel lineages of Cimex bugs were identified. The lineages showed evidence of host preference, being found primarily on M. yumanensis, M. californicus, A. pallidus, and P. hesperus, respectively, for lineages 1–4. A threshold of 4% sequence divergence is typically taken as representing species level differentiation for COI in arthropods (Hebert et al., 2003), indicating Cimex 1, 3, and 4, could be classed as novel species under this criterion. Genetic divergence of Cimex 2 compared with C. latippenis was 3.2%, but C. latippenis has not previously been recorded parasitizing M. yumanensis (Braun et al., 2015), which might also suggest Cimex 2 as a potential new cryptic species.
Cimicid bugs have low inherent dispersal capacity, generally feeding for a few days, before dropping from the bat host to digest the blood in the roost, where they can survive without feeding for approximately 1.5 years (Ossa et al., 2019). The population structure of bat bugs is mainly influenced by bat movements (Ossa et al., 2019; Talbot et al., 2016, 2017; Usinger, 1966). While the current data indicate the new Cimex lineages may have distinct regional distributions within Baja, their host species' ranges extend further through western North America, suggesting these bugs may also have wider distributions, or come into contact with hosts which migrate over long distances at roost sites. For example, M. yumanensis individuals captured in the northern peninsula in our study had mitochondrial haplotypes matching reference sequences from bats sampled in Alaska (Najera‐Cortazar, 2020). This raises the possibility of long‐range mixing of microbial pathogen communities in bat bugs along the west coast of North America.
4.2. Bat flies
Ten novel genetic lineages and six new records of bat flies for the study region were found. Nycteribiid flies were more abundant in the northern temperate sites, while streblids were more abundant in the southern and subtropical sites, supporting trends noted by Dittmar et al. (2006). To our knowledge, molecular records of Basilia sp. species have not previously been reported from bats with ranges in the peninsula. Streblid flies were present on bats from the Phyllostomidae, which in general are fruit and nectar feeders, with the exception of the insectivorous Macrotus californicus; as well as Mormoops megallophyla, from the family Mormoopidae. Nycteribiids parasitized only Vespertilionid bats, which include insectivores and omnivores.
In previous studies it was found that host‐specificity can vary according to the diversity and geographic distribution of hosts (de Vasconcelos et al., 2016; Graciolli et al., 2007; Saldaña‐Vázquez et al., 2019). We found that most of our Nycteribiid lineages exhibited host specificity, despite having hosts with overlapping ranges and which may share roost sites (e.g., Myotis species in northern Baja). The Nycteribiid 2 lineage was found on multiple Myotis species, which could potentially indicate specificity at a genus level. Streblid winged flies have previously been described as mostly non host‐specific (Dittmar et al., 2006), but more recent studies suggest most species to be host specific (de Vasconcelos et al., 2016; Estrada‐Villegas et al., 2018), a change attributable to methodological improvements in sample collection and taxonomic assignments of flies and hosts. In our data N. coxata was found parasitizing Macrotus californicus and Leptonycteris yerbabuenae, both Phyllostomidae species, which are known to share roost site in Baja (Álvarez‐Castañeda et al., 2015), implying potential horizontal transmission.
Lineages Basilia 2a and Basilia 2b (COI divergence 2.9%) are restricted to hosts Myotis vivesi and Antrozous pallidus, respectively. M. vivesi is endemic to the Gulf of Cortes, and restricted to coastal habitats because of its piscivorous diet (Blood & Clark, 1998; Herrera‐Montalvo et al., 2017). A. pallidus is sympatric with M. vivesi on the Gulf coast but has a wider distribution across western North America. It is primarily an insectivorous feeder, but also includes scorpions and nectar in its diet (Frick et al., 2009). Basilia fly records previously reported for M. vivesi, but without reference sequences, include B. plaumanni, B. pynzonix, and B. producto (Graciolli et al., 2007), while flies parasitizing A. pallidus have been described as B. antrozoi (Table 2). The threshold level of divergence between the Basilia 2a and Basilia 2b lineages may indicate recent divergence from a common ancestor, and potential incipient speciation driven by association with sympatric but ecologically differentiated hosts.
4.3. Bat ticks
A new record of Carios kelleyi and seven novel tick lineages belonging to the Argasidae family were found. For both COI and 18S tick lineages 1–5 and Carios kelleyi formed a clade with more than 0.6 posterior support, suggesting they form a Baja or western North America endemic lineage of bat ticks derived from a common ancestor.
The genera Antricola, Carios, Nothoaspis, and Ornithodoros are associated with bats and their roosting sites in Mexico (Sánchez‐Montes et al., 2016). Compared with Ixodidae, the family Argasidae has few molecular studies and reference sequences (Porter & Hajibabaei, 2018). Classifications at genus level are controversial, with many genera being paraphyletic in existing phylogenies, and debates over synonymous use of genus names such as Carios and Ornithodros (Burger et al., 2014). Such ambiguities are also reflected in our phylogenies. Furthermore, while many key internal nodes had posterior support greater than 0.6, differences in reference sequence availability made it challenging to interpret the consistency of inter‐clade placement between makers.
Argasid ticks have previously been reported for the bat species in this study, primarily Ornithodoros species (Table 2), but with the exception of Carios (Ornithodros) kelleyi, none are close molecular matches for our sequences. In ticks, for COI, the threshold for between genus divergence is considered to be above 10% (Hebert et al., 2003). The observed COI divergence among lineages of this study (6.1% up to 19.3%), and to reference sequences (9.9%–22.8%), suggests that the Baja lineages could represent novel species and potentially novel genera.
We found apparent host‐specific and generalist lineages for the ticks reported here. Lineages that appeared host‐specific were restricted to single sites, while generalists were found across multiple sampling locations. For example, Tick 3 parasitizing M. yumanensis was found only in San Basilio, and Tick 6 was found only on M. peninsularis sampled in Teste. Carios kelleyi was found to only parasitise A. pallidus in this study, and was found at three sites in the mid‐ and north peninsula. However, C. kelleyi is known to parasitise multiple species across North America and Cuba (Gill et al., 2004), and the reference COI sequence used here derives from an Eptesicus fuscus bat sampled in New Jersey, eastern USA (GenBank accession code: MT780277). Argasid ticks show a continuum of hosts‐specificity (Cumming, 1998; Esser et al., 2016), but tick stage‐cycle must be considered, with immature ticks being more generalist than adult conspecifics (Esser et al., 2016; Nava & Guglielmone, 2013). In this study, life stage was not assessed while collecting ticks; therefore, it is possible that there are gaps in host range regarding unidentified larvae that were not sequenced.
The Tick 5 lineage was found on multiple species in the mid‐ and north peninsula, but the only specimen recorded for the south peninsula was collected from Parastrellus hesperus (specimen 338, Faro site, 18S only). Since P. hesperus is rare in southern Baja, the presence of this tick indicates potential dispersal of P. hesperus from the north to the south peninsula.
4.4. Limits on ectoparasite sampling and identification
The present study identified 21 novel genetic lineages, plus 7 new ectoparasite species records, from 138 bats of 17 species, sampled across 18 sites and 2 years. This suggests a diverse ectoparasite fauna in this previously unsurveyed region of Mexico, but it is also likely to be an underestimate of the true diversity, due to constraints around sampling effort. Bat sampling was limited to May–August and conducted at relatively accessible locations with water sources to facilitate bat capture. While our sampling sites were chosen to be representative of habitat types across Baja, increasing the spatial and temporal scope of sampling would be likely to increase the number of ectoparasite species discovered. Assessment of ectoparasite fauna found in roosting sites against those found feeding directly from their hosts and expanding seasonal coverage will be important for future work.
Although previous studies report limited data on bat ectoparasites from North Western Mexico and South‐Western USA (Bradshaw & Ross, 1961; Braun et al., 2015; Pérez et al., 2014; Usinger, 1966), they do not integrate morphological and molecular information. For many species, no reference sequence is available from voucher specimens, or there are errors in species identifications and incorrect annotation of reference sequences. Therefore, for all the ectoparasite groups in this study, further work is needed to unify molecular and morphological characterization, to fully confirm which lineages represent previously undescribed species, and which are species with morphological descriptions but no previous molecular record. This is particularly important for bat tick lineages where input from expert morphologists is needed to account for different life stages.
5. CONCLUSIONS
This work presents an initial description of bat ectoparasite diversity relevant to western North America, providing resources useful for future ectoparasite surveys and studies of host‐ectoparasite ecology, and evaluation of zoonotic disease risks. Future work should focus on expanding spatial and bat host species coverage, integrating morphological and molecular characterization of ectoparasite species, profiling ectoparasite microbiomes and viromes, and understanding the ecological and environmental factors that influence host–parasite community structure and evolution. Bat populations and habitat in Baja California are vulnerable to anthropogenic pressures, and such knowledge will be vital for informing assessments of population status and extinction risks of both hosts and parasites.
AUTHOR CONTRIBUTIONS
Laura A. Najera‐Cortazar: Conceptualization (equal); data curation (lead); formal analysis (lead); funding acquisition (equal); investigation (lead); methodology (lead); project administration (lead); resources (equal); software (lead); supervision (supporting); validation (equal); visualization (lead); writing – original draft (lead); writing – review and editing (equal). Alex Keen: Data curation (supporting); formal analysis (supporting); methodology (supporting); writing – review and editing (supporting). Thomas Kitching: Data curation (supporting); validation (supporting); writing – review and editing (supporting). Drew Stokes: Resources (supporting); writing – review and editing (supporting). Simon J. Goodman: Conceptualization (equal); data curation (supporting); funding acquisition (equal); investigation (supporting); methodology (supporting); project administration (equal); resources (equal); supervision (lead); validation (equal); visualization (supporting); writing – original draft (supporting); writing – review and editing (equal).
FUNDING INFORMATION
LANC was supported by a PhD studentship from CONACYT, Mexico (Mexican Council for Science and Technology; reference 411432), held at the School of Biology, University of Leeds, UK, with additional financial support from the University of Leeds Faculty of Biological Sciences Graduate School. Fieldwork season of 2018 was carried out thanks to a Rufford Foundation Small Grant (reference 25403‐1).
CONFLICT OF INTEREST
The authors declare no competing interests.
ACKNOWLEDGMENTS
We thank Dr. Sergio Ticul Alvarez Castañeda for help with field logistics and for sharing his vast knowledge of bat distribution and behavior over the peninsula; and to Dr. Alfredo Ortega and Izmene Rojas, all from Centro de Investigaciones Biológicas del Noroeste, for their support and collaboration throughout this project. We also thank Dr. Winifred Frick for recommending bat sampling locations and for sharing other vital information and contacts that supported the development of the project. Dr. Eduardo F. Aguilera Miller assisted with fieldwork over the 20016‐17 field seasons, and Alejandro Najera, Halley O'Connor and Natasha Kebala during the 2018 season. Alan Harper, Paul Heady and Quintin Frick also provided additional invaluable help with fieldwork. Finally, thanks to Kieran Neal for support with some DNA extractions and generation of some PCR amplicons; and to Dr Michael Siva‐Jothy for providing bed bug samples used for optimisation of PCR conditions and positive controls. We thank Dr. Sergio Guerrero Vazquez from Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, for his support and collaboration for sampling. We thank CONACYT and the Rufford Foundation for the funding provided, as well as the University of Leeds for all the resources provided for this research.
APPENDIX 1.
Estimates of percentage divergence (COI) over sequence‐pairs between groups (diagonal left matrix), and within groups (rightmost column) of bat bugs (family Cimicidae), showing the lineages obtained in this study (row label shaded) versus closely related reference sequences from GenBank. n/c ‐ not calculated as only a single sequence available.
| Cimex 1 | Cimex 2 | Cimex 3 | Cimex 4 | C. adjuntus | C. antennatus | C. brevis | C. hemipterus | C. latipennis | C. lectularius | C. pipistrelle | Cimex Asia | Oeciacus | Within groups | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cimex 1 | n/c | |||||||||||||
| Cimex 2 | 9.9 | 0.59 | ||||||||||||
| Cimex 3 | 16.3 | 16.0 | 0.12 | |||||||||||
| Cimex 4 | 17.1 | 16.1 | 13.5 | 0.50 | ||||||||||
| C. adjuntus | 19.9 | 17.1 | 12.6 | 10.5 | 0.70 | |||||||||
| C. antennatus | 16.9 | 15.2 | 12.1 | 9.3 | 9.7 | n/c | ||||||||
| C. brevis | 14.2 | 13.1 | 13.5 | 14.6 | 15.8 | 14.3 | n/c | |||||||
| C. hemipterus | 19.3 | 17.9 | 20.9 | 18.1 | 19.6 | 19.7 | 17.7 | 0.10 | ||||||
| C. latipennis | 10.6 | 7.2 | 13.9 | 16.2 | 17.1 | 15.2 | 13.7 | 19.1 | 2.01 | |||||
| C. lectularius | 18.7 | 17.3 | 19.8 | 18.0 | 19.1 | 17.9 | 17.4 | 18.5 | 18.6 | 3.51 | ||||
| C. pipistrelli | 19.5 | 16.8 | 20.0 | 17.5 | 18.1 | 17.3 | 16.6 | 15.3 | 17.8 | 15.0 | 1.68 | |||
| Cimex Asia | 19.1 | 16.4 | 20.1 | 17.8 | 17.8 | 17.4 | 16.6 | 15.3 | 17.5 | 15.2 | 2.3 | n/c | ||
| Oeciacus | 20.1 | 17.7 | 20.7 | 18.2 | 18.1 | 17.8 | 17.0 | 15.2 | 18.7 | 15.7 | 6.9 | 7.2 | 6.26 |
APPENDIX 2.
Estimates of evolutionary divergence (COI) in percentage over sequence‐pairs between groups (diagonal left matrix), and within groups (rightmost column) of bat flies (family Nycteribiidae), showing the lineages obtained in this study (row label shaded) versus closely related reference sequences from GenBank.
| Nycteribiid 1 | Nycteribiid 2 | Nycteribia | Penicillidia | Basilia 1 | Basilia 2a | Basilia 2b | Basilia boardmani | Basilia America | Basilia Africa | Within groups | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nycteribiid 1 | 0.35 | ||||||||||
| Nycteribiid 2 | 4.4 | 0.69 | |||||||||
| Nycteribia | 10.7 | 10.5 | 5.91 | ||||||||
| Penicillidia | 11.7 | 11.4 | 9.8 | 5.75 | |||||||
| Basilia 1 | 14.1 | 13.0 | 13.3 | 13.6 | 0.0 | ||||||
| Basilia 2a | 13.5 | 11.4 | 12.9 | 14.6 | 9.8 | 0.10 | |||||
| Basilia 2b | 12.9 | 12.7 | 13.6 | 15.1 | 9.2 | 2.9 | 0.09 | ||||
| Basilia boardmani | 14.5 | 13.9 | 13.0 | 13.2 | 4.9 | 10.3 | 9.9 | 0.00 | |||
| Basilia America | 13.4 | 12.7 | 11.9 | 12.6 | 8.5 | 10.5 | 10.6 | 8.8 | 6.54 | ||
| Basilia Africa | 15.8 | 14.9 | 13.3 | 14.0 | 13.3 | 12.9 | 13.5 | 14.3 | 13.7 | 11.34 |
APPENDIX 3.
Estimates of percentage divergence (COI) over sequence‐pairs between groups (diagonal left matrix), and within groups (rightmost column) of bat flies (family Streblidae), showing the lineages obtained in this study (row label shaded) versus closely related reference sequences from GenBank. Names in the first column have the genus abbreviated. n/c ‐ not calculated as only a single sequence available.
| Aspidoptera phyllostomatis | Megistopoda aranea | Megistopoda sp | Nycterophilia coxata | Paratrichobius 1 | Streblid 1 | Trichobius 1 | Trichobius 2 | Trichobius dugesii | Trichobius intermedius | Trichobius sphaeronotus | Megistopoda aranea 1 | Megistopoda aranea 2 | Nycterophilia fairchildi | Nycterophilia parnelli | Paratrichobius dunni | Paratrichobius longicrus | Strebla sp. | Trichobius major | Trichobius sp | Trichobius urodermae | Within groups | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A. phyllostomatis | 3.3 | |||||||||||||||||||||
| M. aranea | 15.1 | n/c | ||||||||||||||||||||
| M. sp | 14.8 | 4.4 | 0.2 | |||||||||||||||||||
| N. coxata | 17.0 | 18.6 | 18.4 | 0.5 | ||||||||||||||||||
| Paratrichobius 1 | 13.2 | 11.8 | 13.3 | 18.9 | n/c | |||||||||||||||||
| Streblid 1 | 15.1 | 14.2 | 15.2 | 15.5 | 11.9 | 1.1 | ||||||||||||||||
| Trichobius 1 | 13.6 | 17.7 | 17.3 | 16.3 | 13.6 | 12.1 | 0. 9 | |||||||||||||||
| Trichobius 2 | 9.6 | 14.2 | 15.0 | 18.1 | 12.8 | 15.5 | 13.5 | 0.0 | ||||||||||||||
| T. dugesii | 8.8 | 13.6 | 13.7 | 17.3 | 12.5 | 14.2 | 13.1 | 5.7 | 0. 3 | |||||||||||||
| T. intermedius | 10.9 | 15.6 | 16.0 | 17.3 | 14.8 | 15.9 | 14.8 | 11.0 | 9.2 | 1.1 | ||||||||||||
| T. sphaeronotus | 13.9 | 15.9 | 16.9 | 17.8 | 13.3 | 10.8 | 6.2 | 13.0 | 15.1 | 15.4 | 0.0 | |||||||||||
| M. aranea 1 a | 16.0 | 3.1 | 3.0 | 19.0 | 13.4 | 14.8 | 19.3 | 15.1 | 14.4 | 15.5 | 18.2 | n/c | ||||||||||
| M. aranea 2 | 14.8 | 0.8 | 4.0 | 18.8 | 12.0 | 14.3 | 17.7 | 14.3 | 13.5 | 15.3 | 15.8 | 3.0 | 0. 2 | |||||||||
| N. fairchildi | 17.1 | 16.7 | 17.5 | 7.1 | 19.6 | 14.8 | 15.3 | 16.5 | 16.6 | 17.0 | 14.6 | 17.9 | 16.8 | 0. 6 | ||||||||
| N. parnelli | 17.6 | 17.3 | 17.6 | 8.1 | 17.1 | 14.9 | 15.8 | 17.8 | 16.3 | 16.5 | 15.8 | 17.5 | 17.7 | 7.4 | 4.7 | |||||||
| P. dunni | 13.5 | 14.4 | 15.4 | 20.2 | 8.4 | 14.5 | 13.5 | 12.4 | 13.5 | 15.6 | 13.6 | 15.9 | 14.4 | 18.9 | 18.7 | 0.2 | ||||||
| P. longicrus | 13.4 | 13.1 | 14.1 | 21.0 | 4.5 | 14.3 | 13.5 | 14.3 | 13.9 | 15.7 | 13.9 | 14.4 | 13.2 | 19.7 | 18.6 | 8.9 | 1.2 | |||||
| Strebla sp. | 15.0 | 14.6 | 15.0 | 18.2 | 14.5 | 13.3 | 13.3 | 14.4 | 13.3 | 15.6 | 13.2 | 15.5 | 14.5 | 16.5 | 17.2 | 15.2 | 15.9 | 9.1 | ||||
| T. major | 13.4 | 20.3 | 20.0 | 13.7 | 15.3 | 13.1 | 10.8 | 13.3 | 17.0 | 15.8 | 11.9 | 21.6 | 20.2 | 14.5 | 17.5 | 16.0 | 15.6 | 16.1 | 0.0 | |||
| Trichobius sp | 14.2 | 15.4 | 15.8 | 17.6 | 14.3 | 13.7 | 13.4 | 13.2 | 12.4 | 14.1 | 13.4 | 16.0 | 15.1 | 16.7 | 16.8 | 15.2 | 15.3 | 14.6 | 15.1 | 13.3 | ||
| T. urodermae | 7.7 | 14.2 | 14.9 | 19.6 | 13.9 | 15.3 | 13.9 | 8.7 | 7.5 | 9.3 | 14.7 | 15.0 | 14.0 | 18.6 | 18.1 | 13.7 | 14.1 | 14.3 | 15.1 | 13.1 | 0.2 |
Megistopoda aranea 1: GenBank accession number EF531219.
APPENDIX 4.
Estimates of evolutionary divergence (COI) in percentage over sequence‐pairs between groups (diagonal left matrix), and within groups (rightmost column) of bat ticks (family Argasidae), showing the lineages obtained in this study (row label shaded) versus closely related reference sequences from GenBank. n/c ‐ not calculated as only a single sequence available.
| Antricola 1 | Carios kelleyi | Tick 1 | Tick 2 | Tick 3 | Tick 4 | Tick 5 | Tick 6 | Antricola sp. | Carios sp. | Nothoaspis sp. | Ornithodoros Asia | Ornithodoros America | Ornithodoros Africa | Within groups | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Antricola 1 | 0.99 | ||||||||||||||
| Carios kelleyi | 13.9 | 0.10 | |||||||||||||
| Tick 1 | 13.1 | 11.8 | 0.13 | ||||||||||||
| Tick 2 | 13.9 | 6.1 | 11.4 | 0.56 | |||||||||||
| Tick 3 | 13.3 | 12.5 | 12.9 | 12.7 | 4.97 | ||||||||||
| Tick 4 | 13.7 | 11.9 | 11.9 | 11.5 | 12.1 | 0.33 | |||||||||
| Tick 5 | 12.7 | 12.7 | 12.5 | 11.3 | 12.6 | 11.4 | 0.07 | ||||||||
| Tick 6 | 13.7 | 14.5 | 13.4 | 14.2 | 15.0 | 14.3 | 15.4 | 11.83 | |||||||
| Antricola sp. | 13.7 | 14.6 | 15.2 | 14.4 | 15.0 | 14.8 | 14.2 | 14.3 | 11.12 | ||||||
| Carios | 14.0 | 14.0 | 14.3 | 13.9 | 14.4 | 14.7 | 14.1 | 15.2 | 15.1 | n/c | |||||
| Nothoaspis | 14.4 | 13.5 | 14.8 | 13.7 | 15.8 | 14.6 | 14.6 | 16.8 | 14.2 | 15.0 | 18.70 | ||||
| Ornithodoros Asia | 17.6 | 18.3 | 18.5 | 17.1 | 17.9 | 17.2 | 17.9 | 19.3 | 17.8 | 19.1 | 17.7 | 14.74 | |||
| Ornithodoros America | 15.3 | 14.6 | 15.1 | 14.2 | 15.5 | 14.7 | 15.7 | 13.9 | 15.0 | 15.2 | 15.9 | 17.7 | 14.67 | ||
| Ornithodoros Africa | 16.2 | 16.2 | 16.4 | 15.2 | 16.3 | 16.4 | 16.3 | 16.5 | 16.6 | 15.9 | 16.3 | 18.1 | 16.4 | 0.99 |
Najera‐Cortazar, L. A. , Keen, A. , Kitching, T. , Stokes, D. , & Goodman, S. J. (2023). Phylogenetic analyses reveal bat communities in Northwestern Mexico harbor a high diversity of novel cryptic ectoparasite species. Ecology and Evolution, 13, e9645. 10.1002/ece3.9645
Contributor Information
Laura A. Najera‐Cortazar, Email: l.a.najera@leeds.ac.uk.
Simon J. Goodman, Email: s.j.goodman@leeds.ac.uk.
DATA AVAILABILITY STATEMENT
Sequences for the haplotypes reported in this study are available under GenBank accession numbers OP963052–OP963192 for the COI set, and OP941218–OP941364 for the 18S set.
REFERENCES
- Álvarez‐Castañeda, S. T. , Álvarez, T. , & González‐Ruíz, N. (2015). Guía para la identificación de los mamíferos de México en campo y laboratorio. Centro de Investigaciones Biológicas del Noroeste, S. C. y Asociacion Mexicana de Mastozoología, A. C. [Google Scholar]
- Black, W. C., IV , Klompen, J. S. H. , & Keirans, J. E. (1997). Phylogenetic relationships among tick subfamilies (Ixodida: Ixodidae: Argasidae) based on the 18S nuclear rDNA gene. Molecular Phylogenetics and Evolution, 7(1), 129–144. [DOI] [PubMed] [Google Scholar]
- Blood, B. R. , & Clark, M. K. (1998). Myotis vivesi . Mammalian Species, 588, 1–5. [Google Scholar]
- Bolaños‐García, R. , Rodríguez‐Estrella, R. , & Guzmán‐Cornejo, C. (2018). Ectoparásitos asociados a polluelos del Búho Cornudo (Aves: Strigidae) en un paisaje fragmentado de la península de Baja California, México. Acta Zoológica Mexicana, 34(1), 1–15. 10.21829/azm.2018.3412142 [DOI] [Google Scholar]
- Borstein, S. (2018). AnnotationBustR: An R package to extract sub‐sequences from GenBank annotations . https://cran.r‐project.org/web/packages/AnnotationBustR/vignettes/AnnotationBustR‐vignette.html [DOI] [PMC free article] [PubMed]
- Bradshaw, G. V. R. , & Ross, A. (1961). Ectoparasites of Arizona bats. Journal of the Arizona Academy of Science, 1(4), 109. 10.2307/40026556 [DOI] [Google Scholar]
- Braun, J. K. , Yang, B. , González‐Pérez, S. B. , & Mares, M. A. (2015). Myotis yumanensis (Chiroptera: Vespertilionidae). Mammalian Species, 47(918), 1–14. 10.1093/mspecies/sev001 [DOI] [Google Scholar]
- Burger, T. D. , Shao, R. , Labruna, M. B. , & Barker, S. C. (2014). Molecular phylogeny of soft ticks (Ixodida: Argasidae) inferred from mitochondrial genome and nuclear rRNA sequences. Ticks and Tick‐Borne Diseases, 5(2), 195–207. 10.1016/j.ttbdis.2013.10.009 [DOI] [PubMed] [Google Scholar]
- Campbell, B. C. , Steffen‐Campbell, J. D. , Sorensen, J. T. , & Gill, R. J. (1995). Paraphyly of Homoptera and Auchenorrhyncha inferred from 18S rDNA nucleotide sequences. Systematic Entomology, 20(3), 175–194. 10.1111/j.1365-3113.1995.tb00090.x [DOI] [Google Scholar]
- Cumming, G. S. (1998). Host preference in African ticks (Acari: Ixodida): A quantitative data set. Bulletin of the Entomological Research, 88(4), 379–406. [Google Scholar]
- Cuxim‐Koyoc, A. , Reyes‐Novelo, E. , Morales‐Malacara, J. B. , Bolívar‐Cimé, B. , & Laborde, J. (2015). Streblidae (Diptera: Hippoboscoidea) from Yucatan and updated species list for Mexico. Journal of Medical Entomology, 52(5), 947–961. 10.1093/jme/tjv117 [DOI] [PubMed] [Google Scholar]
- de Vasconcelos, P. F. , Falcão, L. A. D. , Graciolli, G. , & Borges, M. A. Z. (2016). Parasite–host interactions of bat flies (Diptera: Hippoboscoidea) in Brazilian tropical dry forests. Parasitology Research, 115(1), 367–377. 10.1007/s00436-015-4757-8 [DOI] [PubMed] [Google Scholar]
- Dick, C. W. , Gannon, M. R. , Little, W. E. , & Patrick, M. J. (2003). Ectoparasite associations of bats from Central Pennsylvania. Journal of Medical Entomology, 40(6), 813–819. 10.1603/0022-2585-40.6.813 [DOI] [PubMed] [Google Scholar]
- Dick, C. W. , & Miller, J. A. (2010). Streblidae (bat flies). In Brown B. V., Borkent A., Cumming J. M., Wood D. M., Woodley N. E., & Zumbado M. (Eds.), Manual of central American Diptera. (Vol. 2, pp. 1249–1260). NRC Research Press. [Google Scholar]
- Dittmar, K. , Porter, M. L. , Murray, S. , & Whiting, M. F. (2006). Molecular phylogenetic analysis of nycteribiid and streblid bat flies (Diptera: Brachycera, Calyptratae): Implications for host associations and phylogeographic origins. Molecular Phylogenetics and Evolution, 38(1), 155–170. 10.1016/j.ympev.2005.06.008 [DOI] [PubMed] [Google Scholar]
- Esser, H. J. , Herre, E. A. , Blüthgen, N. , Loaiza, J. R. , Bermúdez, S. E. , & Jansen, P. A. (2016). Host specificity in a diverse Neotropical tick community: An assessment using quantitative network analysis and host phylogeny. Parasites and Vectors, 9(1), 1–14. 10.1186/s13071-016-1655-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Estrada‐Pena, A. , Mangold, A. J. , Nava, S. , Venzal, J. M. , Labruna, M. , & Guglielmone, A. A. (2010). A review of the systematics of the tick family argasidae (Ixodida). Acarologia, 50(3), 317–333. 10.1051/acarologia/20101975 [DOI] [Google Scholar]
- Estrada‐Villegas, S. , Halczok, T. K. , Tschapka, M. , Page, R. A. , Brändel, S. D. , & Hiller, T. (2018). Bats and their bat flies: Community composition and host specificity on a Pacific Island archipelago. Acta Chiropterologica, 20(1), 161–176. 10.3161/15081109ACC2018.20.1.012 [DOI] [Google Scholar]
- Folmer, O. , Black, M. , Hoeh, W. , Lutz, R. , & Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3(5), 294–299. 10.1371/journal.pone.0013102 [DOI] [PubMed] [Google Scholar]
- Frick, W. F. , Hayes, J. P. , & Heady, P. A. (2008). Island biogeography of bats in Baja California, Mexico: Patterns of bat species richness in a near‐shore archipelago. Journal of Biogeography, 35(2), 353–364. 10.1111/j.1365-2699.2007.01798.x [DOI] [Google Scholar]
- Frick, W. F. , Hayes, J. P. , & Heady, P. A. (2009). Nestedness of desert bat assemblages: Species composition patterns in insular and terrestrial landscapes. Oecologia, 158(4), 687–697. 10.1007/s00442-008-1168-x [DOI] [PubMed] [Google Scholar]
- Gay, N. , Olival, K. J. , Bumrungsri, S. , Siriaroonrat, B. , Bourgarel, M. , & Morand, S. (2014). Parasite and viral species richness of Southeast Asian bats: Fragmentation of area distribution matters. International Journal for Parasitology: Parasites and Wildlife, 3(2), 161–170. 10.1016/j.ijppaw.2014.06.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gill, J. , Rowley, W. , Bush, P. , Viner, J. , & Gilchrist, M. (2004). Detection of human blood in the bat tick Carios (Ornithodoros) kelleyi (Acari: Argasidae) in Iowa. Journal of Medical Entomology, 41(6), 1179–1181. 10.1603/0022-2585-41.6.1179 [DOI] [PubMed] [Google Scholar]
- Gómez, A. , & Nichols, E. (2013). Neglected wild life: Parasitic biodiversity as a conservation target. International Journal for Parasitology: Parasites and Wildlife, 2(1), 222–227. 10.1016/j.ijppaw.2013.07.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- González‐Abraham, C. E. , Garcillán, P. , & Ezcurra, E. (2010). Ecorregiones de la península de Baja California: una síntesis. Boletín de la Sociedad Botánica de México, 87, 69–82. [Google Scholar]
- Graciolli, G. , Autino, A. G. , & Claps, G. L. (2007). Catalogue of American Nycteribiidae (Diptera, Hippoboscoidea). Revista Brasileira de Entomologia, 51(2), 142–159. 10.1590/S0085-56262007000200004 [DOI] [Google Scholar]
- Guglielmone, A. A. , Robbins, R. G. , Apanaskevich, D. A. , Petney, T. N. , Estrada‐Peña, A. , Horak, I. G. , Shao, R. , & Barker, S. C. (2010). The Argasidae, Ixodidae and Nuttalliellidae (Acari: Ixodida) of the world: A list of valid species names. Zootaxa, 2528, 1–28. 10.1023/A:1025381712339 [DOI] [Google Scholar]
- Guzmán‐Cornejo, C. , García‐Prieto, L. , Rebollo‐Hernández, A. , Venzal, J. M. , Nava, S. , & Sánchez‐Montes, S. (2017). Molecular evidence and additional morphological characters to distinguish Ornithodoros brodyi and Ornithodoros yumatensis (Ixodida: Argasidae) in their different developmental stages. Acta Parasitologica, 62(2), 432–448. 10.1515/ap-2017-0051 [DOI] [PubMed] [Google Scholar]
- Hall, T. (2005). BioEdit. Biological sequence editor . http://www.mbio.ncsu.edu/BioEdit/bioedit.html
- Hebert, P. D. N. , Ratnasingham, S. , & DeWaard, J. R. (2003). Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proceedings of the Royal Society B: Biological Sciences, 270, 96–99. 10.1098/rsbl.2003.0025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrera‐Montalvo, L. G. , Flores‐Martínez, J. J. , & Sánchez‐Cordero, V. (2017). Geographical distribution and conservation status of an endemic insular mammal: The vulnerable fish‐eating bat Myotis vivesi . Oryx, 53(2), 388–393. 10.1017/S0030605317000874 [DOI] [Google Scholar]
- Hornok, S. , Szoke, K. , Boldogh, S. A. , Sándor, A. D. , Kontschán, J. , Tu, V. T. , Halajian, A. , Takács, N. , Görföl, T. , & Estók, P. (2017). Phylogenetic analyses of bat‐associated bugs (Hemiptera: Cimicidae: Cimicinae and Cacodminae) indicate two new species close to Cimex lectularius . Parasites and Vectors, 10(1), 1–10. 10.1186/s13071-017-2376-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jobling, B. (1938). A revision of the species of the genus Trichobius: (Diptera Acalypterae, Streblidae). Parasitology, 30(3), 358–387. 10.1017/S0031182000025932 [DOI] [Google Scholar]
- Klimpel, S. , & Mehlhorn, H. (Eds.). (2014). Bats (Chiroptera) as vectors of diseases and parasites. Parasitology Research Monographs, 5 (pp. 187). Springer‐Verlag. 10.1007/978-3-642-39333-4 [DOI] [Google Scholar]
- Knee, W. , & Proctor, H. (2006). Keys to the families and genera of blood and tissue feeding mites associated with albertan birds. Canadian Journal of Arthropod Identification, 2(2), 1–18. 10.3752/cjai.2006.02 [DOI] [Google Scholar]
- Letunic, I. , & Bork, P. (2016). Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Research, 28(3), 1–4. 10.1093/nar/gkw290 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Librado, P. , & Rozas, J. (2009). DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics, 25(11), 1451–1452. [DOI] [PubMed] [Google Scholar]
- Mangold, A. J. , Bargues, M. D. , & Mas‐Coma, S. (1997). 18S rRNA gene sequences and phylogenetic relationships of European hard‐tick species (Acari: Ixodidae). Parasitology Research, 84(1), 31–37. 10.1007/s004360050352 [DOI] [PubMed] [Google Scholar]
- Mans, B. J. , de Klerk, D. , Pienaar, R. , de Castro, M. H. , & Latif, A. A. (2012). The mitochondrial genomes of Nuttalliella namaqua (Ixodoidea: Nuttalliellidae) and Argas africolumbae (Ixodoidae: Argasidae): Estimation of divergence dates for the major tick lineages and reconstruction of ancestral blood‐feeding characters. PLoS One, 7(11), e49461. 10.1371/journal.pone.0049461 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDaniel, B. (1979). How to know the mites and ticks. W.C. Brown Co. [Google Scholar]
- Medellín, R. A. , Arita, H. T. , & Sánchez, O. (2007). Identificación de los murciélagos de México. Claves de Campo (2nd ed.). Universidad Nacional Autonóma de México, Instituto Nacional de Ecología. [Google Scholar]
- Morand, S. , Krasnov, B. R. , & Poulin, R. (2006). Micromammals and macroparasites: From evolutionary ecology to management. Springer Publishing. [Google Scholar]
- Najera‐Cortazar, L. A. (2020). Ecological genomics of Myotis bats and their ectoparasites in the Baja California peninsula . PhD thesis, University of Leeds, UK.
- Nájera‐Cortazar, L. A. , Álvarez‐Castañeda, S. T. , & De Luna, E. (2015). An analysis of Myotis peninsularis (Vespertilionidae) blending morphometric and genetic datasets. Acta Chiropterologica, 17(1), 37–47. 10.3161/15081109ACC2015.17.1.003 [DOI] [Google Scholar]
- Nava, S. , & Guglielmone, A. A. (2013). A meta‐analysis of host specificity in Neotropical hard ticks (Acari: Ixodidae). Bulletin of the Entomological Research, 103(2), 216–224. [DOI] [PubMed] [Google Scholar]
- Ossa, G. , Johnson, J. S. , Puisto, A. I. E. , Rinne, V. , Sääksjärvi, I. E. , Waag, A. , Vesterinen, E. J. , & Lilley, T. M. (2019). The Klingon batbugs: Morphological adaptations in the primitive bat bugs, Bucimex chilensis and Primicimex cavernis, including updated phylogeny of Cimicidae. Ecology and Evolution, 9(4), 1736–1749. 10.1002/ece3.4846 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pérez, T. M. , Guzmán‐Cornejo, C. , Montiel‐Parra, G. , Paredes‐León, R. , & Rivas, G. (2014). Biodiversidad de ácaros en México. Revista Mexicana de Biodiversidad, 85(Suppl), 399–407. 10.7550/rmb.36160 [DOI] [Google Scholar]
- Porter, T. M. , & Hajibabaei, M. (2018). Over 2.5 million COI sequences in GenBank and growing. PLoS One, 13(9), e0200177. 10.1371/journal.pone.0200177 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poulin, R. (2014). Parasite biodiversity revisited: Frontiers and constraints. International Journal for Parasitology, 44(9), 581–589. 10.1016/j.ijpara.2014.02.003 [DOI] [PubMed] [Google Scholar]
- Rambaut, A. , Drummond, A. J. , Xie, D. , Baele, G. , & Suchard, M. A. (2018). Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Systematic Biology, 67(5), 901–904. 10.1093/sysbio/syy032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramírez‐Martínez, M. , Ibarra‐López, M. P. , Iniguez‐Dávalos, L. I. , Yuill, T. , Orlova, M. V. , & Reeves, W. K. (2016). New records of ectoparasitic Acari (Arachnida) and Streblidae (Diptera) from bats in Jalisco, Mexico. Journal of Vector Ecology, 41(2), 309–313. 10.1111/jvec.12228 [DOI] [PubMed] [Google Scholar]
- Ratnasingham, S. , & Hebert, P. D. N. (2007). BOLD: The barcode of life data system (www.barcodinglife.org). Molecular Ecology Notes, 7, 355–364. 10.1111/j.1471-8286.2006.01678.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ratnasingham, S. , & Hebert, P. D. N. (2013). A DNA‐based registry for all animal species: The Barcode Index Number (BIN) system. PLoS One, 8(8), e66213. 10.1371/journal.pone.0066213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reinhardt, K. , & Siva‐Jothy, M. T. (2007). Biology of the bed bugs (Cimicidae). Annual Review of Entomology, 52(1), 351–374. 10.1146/annurev.ento.52.040306.133913 [DOI] [PubMed] [Google Scholar]
- Riddle, B. R. , Hafner, D. J. , Alexander, L. F. , & Jaeger, J. R. (2000). Cryptic vicariance in the historical assembly of a Baja California peninsular desert biota. Proceedings of the National Academy of Sciences of the United States of America, 97(26), 14438–14443. 10.1073/pnas.250413397 [DOI] [PMC free article] [PubMed] [Google Scholar]
- RStudio Team . (2021). RStudio: Integrated development for R. RStudio, PBC; http://www.rstudio.com/ [Google Scholar]
- Saldaña‐Vázquez, R. A. , Sandoval‐Ruiz, C. A. , Veloz‐Maldonado, O. S. , Durán, A. A. , & Ramírez‐Martínez, M. M. (2019). Host ecology moderates the specialization of Neotropical bat‐fly interaction networks. Parasitology Research, 118(10), 2919–2924. 10.1007/s00436-019-06452-1 [DOI] [PubMed] [Google Scholar]
- Salinas‐Ramos, V. B. , Zaldívar‐Riverón, A. , Rebollo‐Hernández, A. , & Herrera‐M, L. G. (2018). Seasonal variation of bat‐flies (Diptera: Streblidae) in four bat species from a tropical dry forest. Mammalia, 82(2), 133–143. 10.1515/mammalia-2016-0176 [DOI] [Google Scholar]
- Sánchez‐Montes, S. , Guzmán‐Cornejo, C. , Martínez‐Nájera, Y. , Becker, I. , Venzal, J. M. , & Labruna, M. B. (2016). Rickettsia lusitaniae associated with Ornithodoros yumatensis (Acari: Argasidae) from two caves in Yucatan, Mexico. Ticks and Tick‐Borne Diseases, 7(6), 1097–1101. 10.1016/j.ttbdis.2016.09.003 [DOI] [PubMed] [Google Scholar]
- Sándor, A. D. , Corduneanu, A. , Péter, Á. , Mihalca, A. D. , Barti, L. , Csősz, I. , Szőke, K. , & Hornok, S. (2019). Bats and ticks: Host selection and seasonality of bat‐specialist ticks in eastern Europe. Parasites & Vectors, 12(1), 605. 10.1186/s13071-019-3861-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT) . (2010). Norma Oficial Mexicana NOM‐059‐SEMARNAT‐2010. Protección ambiental‐especies nativas de México de flora y fauna silvestres‐Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio‐lista de especies en riesgo .
- Seneviratne, S. S. , Fernando, H. C. , & Udagama‐Randeniya, P. V. (2009). Host specificity in bat ectoparasites: A natural experiment. International Journal for Parasitology, 39(9), 995–1002. 10.1016/j.ijpara.2008.12.009 [DOI] [PubMed] [Google Scholar]
- Speer, K. A. , Luetke, E. , Bush, E. , Sheth, B. , Gerace, A. , Quicksall, Z. , Miyamoto, M. , Dick, C. W. , Dittmar, K. , Albury, N. , & Reed, D. L. (2019). A fly on the cave wall: Parasite genetics reveal fine‐scale dispersal patterns of bats. Journal of Parasitology, 105(4), 555. 10.1645/19-20 [DOI] [PubMed] [Google Scholar]
- Spencer, H. G. , & Zuk, M. (2016). For host's sake: The pluses of parasite preservation. Trends in Ecology and Evolution, 31(5), 341–343. 10.1016/j.tree.2016.02.021 [DOI] [PubMed] [Google Scholar]
- Steinlein, D. B. , Durden, L. A. , & Cannon, W. L. (2001). Tick (Acari) infestations of bats in New Mexico. Journal of Medical Entomology, 38(4), 609–611. 10.1603/0022-2585-38.4.609 [DOI] [PubMed] [Google Scholar]
- Suchard, M. A. , Lemey, P. , Baele, G. , Ayres, D. L. , Drummond, A. J. , & Rambaut, A. (2018). Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evolution, 4(1), vey016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudhir, K. , Stecher, G. , Li, M. , Knyaz, C. , & Tamura, K. (2018). MEGA X: Molecular evolutionary genetics analysis across computing platforms. Molecular Biology and Evolution, 35, 1547–1549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szentiványi, T. , Christe, P. , & Glaizot, O. (2019). Bat flies and their microparasites: Current knowledge and distribution. Frontiers in Veterinary Science, 6, 1–12. 10.3389/fvets.2019.00115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talbot, B. , Vonhof, M. J. , Broders, H. G. , Fenton, B. , & Keyghobadi, N. (2016). Range‐wide genetic structure and demographic history in the bat ectoparasite Cimex adjunctus . BMC Evolutionary Biology, 16(1), 1–13. 10.1186/s12862-016-0839-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talbot, B. , Vonhof, M. J. , Broders, H. G. , Fenton, M. B. , & Keyghobadi, N. (2017). Population structure in two geographically sympatric and congeneric ectoparasites: Cimex adjuntus and Cimex lectularius, in the North American Great Lakes region. Canadian Journal of Zoology, 95, 901–907. [Google Scholar]
- Ter Hofstede, H. M. , & Fenton, M. B. (2005). Relationships between roost preferences, ectoparasite density, and grooming behaviour of neotropical bats. Journal of Zoology, 266(4), 333–340. 10.1017/S095283690500693X [DOI] [Google Scholar]
- The National Library of Medicine . (2018). The Basic Local Alignment Search Tool (BLAST) . The National Center for Biotechnology Information website: http://blast.ncbi.nlm.nih.gov/Blast.cgi
- Thompson, J. D. , Higgins, D. G. , & Gibson, T. J. (1994). CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position‐specific gap penalties and weight matrix choice. Nucleic Acids Research, 22, 4673–4680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tortosa, P. , Dsouli, N. , Gomard, Y. , Ramasindrazana, B. , Dick, C. W. , & Goodman, S. M. (2013). Evolutionary history of Indian Ocean Nycteribiid bat flies mirroring the ecology of their hosts. PLoS One, 8(9), e75215. 10.1371/journal.pone.0075215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trujillo‐Pahua, L. , & Ibáñez‐Bernal, S. (2018). New geographical records of bat flies (Diptera: Streblidae) associated with phyllostomid bats (Chiroptera: Phyllostomidae) in the West Highlands of Mexico. Journal of Medical Entomology, 56, 18–28. 10.1093/jme/tjy166 [DOI] [PubMed] [Google Scholar]
- Usinger, R. L. (1966). Monograph of Cimicidae (Hemiptera–Heteroptera). Entomological Society of America, VII, 572. http://apps.webofknowledge.com/full_record.do?product=UA&search_mode=GeneralSearch&qid=32&SID=Y2m6OyGYg7txy2Oc2bE&page=1&doc=3 [Google Scholar]
- Wenzel, R. , & Tipton, V. J. (1966). Some relationships between mammal hosts and their ectoparasites. In Wenzel R. & Tipton V. J. (Eds.), Ectoparasites of Panama (pp. 677–723). Field Museum of Natural History. [Google Scholar]
- Whiting, M. F. (2002). Mecoptera is paraphyletic: Multiple genes and phylogeny of Mecoptera and Siphonaptera. Zoologica Scripta, 31(1), 93–104. 10.1046/j.0300-3256.2001.00095.x [DOI] [Google Scholar]
- Wilder, A. P. , Kunz, T. H. , & Sorenson, M. D. (2015). Population genetic structure of a common host predicts the spread of white‐nose syndrome, an emerging infectious disease in bats. Molecular Ecology, 24, 5495–5506. 10.1111/mec.13396 [DOI] [PubMed] [Google Scholar]
- Zamora‐Mejías, D. , Morales‐Malacara, J. B. , Rodríguez‐Herrera, B. , Ojeda, M. , & Medellín, R. A. (2020). Does latitudinal migration represent an advantage in the decrease of ectoparasite load in Leptonycteris yerbabuenae (Chiroptera)? Journal of Mammalogy, 101, 979–989. 10.1093/jmammal/gyaa075 [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Sequences for the haplotypes reported in this study are available under GenBank accession numbers OP963052–OP963192 for the COI set, and OP941218–OP941364 for the 18S set.