

ABSTRACT
Candida albicans (C. albicans) is a ubiquitous fungal commensal component of the human microbiota, and under certain circumstances, such as during an immunocompromised state, it may initiate different types of infection. Moreover, C. albicans continuously and reciprocally interacts with the host immune system as well as with other elements of the gut microbiota, thus contributing significantly to both gut homeostasis and host immunity. People living with HIV (PLWH), including those receiving antiretroviral therapy, are characterized by a depletion of CD4 + T-cells and dysbiosis in their gut. C. albicans colonization is frequent in PLWH, causing both a high prevalence and high morbidity. Gut barrier damage and elevated levels of microbial translocation are also fairly common in this population. Herein, we take a closer look at the reciprocity among C. albicans, gut microbiota, HIV, and the host immune system, thus throwing some light on this complex interplay.
KEYWORDS: Candida albicans, HIV, gut microbiota, dysbiosis, systemic infections, microbial translocation
Introduction
There are approximately 200 known species within the Candida genus, and only 20 of these species have been reported to be responsible for disease development in humans 1. These Candida species are the most common human fungal pathogens, and cause (i) superficial infections on both skin and mucosal surfaces, which may be unpleasant but are relatively innocuous; however superficial infections can potentially impair an individual’s quality of life, and (ii) systemic infections resulting from the dissemination of the fungus through the bloodstream.2 In this context, it is known that systemic infections are life-threatening, as their mortality rates are greater than 40%.3 Clinical Candida infection rates increase yearly 4,5 due to the increasing number of newly-diagnosed immunocompromised patients,6 the widespread utilization of solid organ transplantation to treat organ failure,7 and the extensive and routine use of immunosuppressant drugs and broad-spectrum antibiotics, particularly in cancer chemotherapy.8 It is worth noting that within the 20 Candida species responsible for disease development in humans, the vast majority of infections are caused by Candida albicans (C. albicans),1,9 which is a diploid, polymorphic fungus, described as a commensal member of the normal human microbiome.10 Reports suggest that C. albicans is detected in 70%~90% of cases of candidiasis-related fungal disease.11–13
Among people living with HIV (PLWH), C. albicans is one of the most common opportunistic pathogens found during the course of HIV disease progression. It has been estimated that more than 90% of PLWH have ever developed oropharyngeal candidiasis (OPC).14 In one study, monitoring data derived from HIV-infected outpatients indicated that more than 50% of PLWH had been colonized by Candida species, and 12% of these patients developed symptomatic candidiasis.15 Out of the 262 strains that were isolated in the preceding study, 218 (83.2%) strains were identified as Candida albicans and only 44 (16.8%) strains were observed to be non-albicans species.15
In recent times, a number of studies have reported that the gut microbiota plays a major role in overall host health, and that microbial dysbiosis is purported to be associated with various diseases, including HIV, diabetes, and some cancers.16 As a constituent part of the normal human gut microbiota, C. albicans and its associated fungal virulence factors actively interact with the host immune system through adherence, invasion, and cellular damage which induces secretion of proinflammatory cytokines, and activation of antifungal activities.17 At the same time, there are ongoing complex interactions between C. albicans and other components of the gut microbiota, which impacts on the proliferation of each of the microbial components of the gut microbiota. In PLWH, Candida spp. are significantly more prevalent in the gut, compared to healthy controls.18 Moreover, PLWH also have to contend with CD4 + T-cell depletion, microbiota dysbiosis, and gut barrier damage, resulting in a high level of microbial translocation and systemic inflammation, which has previously been assumed to be one of the contributors to morbidity and mortality in this population.19 In this scenario, the pathogenicity of C. albicans is amplified, further disrupting the usually prevailing microbiological milieu of the intestine, thus enhancing gut damage and promoting microbial translocation. Herein, we review advances in the understanding of the biological and pathophysiological characteristics of C. albicans, its interactions with host immunity and the gut microbiota, and we critically discuss the implications of C. albicans infection in PLWH.
Candida albicans: knowledge review
Life cycle and epidemiology
The genus Candida is classified in the family of Saccharomycetales incertae sedis. Evolutionary studies have indicated that one species of Candida dubliniensis has the closest phylogenetic relationship to C. albicans, with both these organisms diverging from their common ancestor (a species of Candida tropicalis) approximately 20 million years ago.20,21 Clinical isolates of C. albicans are generally diploid, and the genome consists of eight chromosomes, which harbors roughly 6,100 genes.22–24 The reference genome of the standard C. albicans strain, SC5314, is 14.3 megabases in length.25 Although no accurate and specific life cycle has been described for C. albicans, it is currently universally acknowledged that C. albicans is a largely asexual, diploid pleomorphic fungus (Figure 1), that is able to grow either as a budding yeast, as a pseudomycelium (or pseudohyphae; elongated and conjoined yeast cells), or as hyphae (formed from conjugated parallel-sided tip-growing filaments).26 The transition from a budding yeast to the filamentous hyphal form has been suggested to be critical for pathogenicity.27 Indeed, the smaller yeast cells are thought to play a role in dissemination during an infection, while the hyphal forms [triggered by expression of genes associated with virulence factors, such as hyphal wall protein 1 (Hwp1), agglutinin-like sequence protein 3 (Als3), secreted aspartic proteases 4, 5, and 6 (Sap4, Sap5, and Sap6), and hyphal-associated proteins Ece1 and Hyr1 28] are more invasive and contribute to host tissue damage.29–31 Furthermore, C. albicans can manifest as a non-sexual form via a process referred to as phenotypic switching, whereby it can generate stable cell and colony variants displaying distinct properties.32 Such changes are spontaneous, reversible, and possibly controlled by regulatory gene expression. Based on phenotype, C. albicans exists in two forms: (1) white, round cells in smooth colonies and (2) opaque, rod-shaped cells in flat, gray colonies. These two forms have different tissue affinities, and antigen expression is also altered in each form. This inherent flexibility makes it highly adaptable to environmental changes.33 For instance, the white-GUT (gastrointestinally indUced transition) switch (adapted to survival in the digestive tract) is triggered after passage in the gut, leading to upregulation of the gene encoding for white-opaque switching (WOR1), in addition to other opaque-specific genes, as observed by Fox et al.34 (in vitro) and Pande et al. 35 (in vivo).
Figure 1.
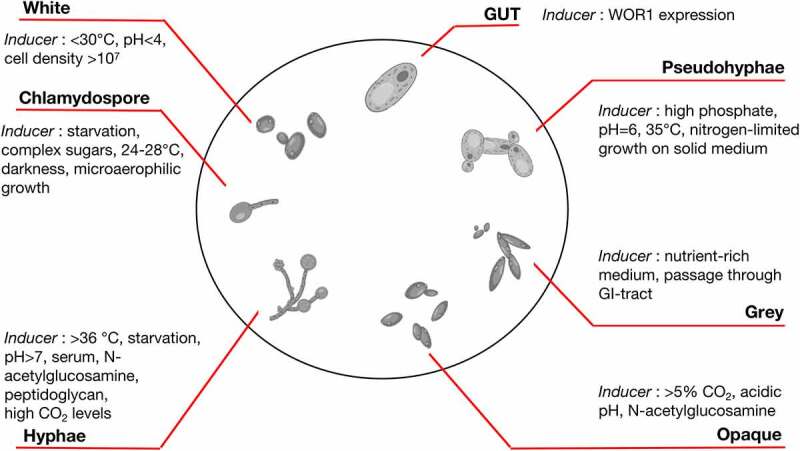
Candida albicans existing forms. Adapted from the review by d’Enfert et al.26
It is also acknowledged that C. albicans undergoes a parasexual life cycle, meaning that diploid cells of opposite mating types (different from conventional male/female organism forms) undergo cell fusion to create a tetraploid cell. This cellular unit subsequently undergoes a reductional division to return to a diploid state. During this division, chromosomes are randomly lost and are not exchanged in a conventional manner, although some gene conjugation occurs, to ensure genetic diversity.36 Furthermore, Bennett and Johnson have shown that the mating is efficient only if C. albicans switches from the white form to the opaque form (which is 106 times more efficient for mating than the white form).37
C. albicans can successfully adapt to different organ system niches (vagina, oral mucosa, skin, respiratory tract, and/or gut) without causing disease, which suggests that the organism is highly adapted for commensalism. Indeed, it is known that C. albicans is carried commensally on the mucosal membranes of 67% of humans without causing any tissue damage or disease.38 At the same time, contemporary studies suggest that C. albicans is the most common serious fungal pathogen in humans, as it causes between 250,000 and 400,000 deaths per annum worldwide. Also, an estimated 100 million cases of recurrent Candida vulvovaginitis manifests in women throughout the world annually.39,40 C. albicans behaves like a classical opportunistic pathogen,39 and its presence within the host is normally under close surveillance by the host immune system and the protective bacterial microbiota in the gut or other mucosal surfaces.41 Due to its adaptative capabilities, C. albicans has the capacity to evade immune system surveillance by switching from one form to another. For example, Uwamahoro et al. have demonstrated that C. albicans transformation from yeast to hypha can help fungi evade phagocytosis by macrophages,42 and subsequently invade host tissues. Thus, the hyphal form is invariably linked to pathological processes (invasion through intercellular junctions, small ridges, and grooves associated with weakened surface/mucosal integrity, in order to take advantage of disrupted epithelial integrity 43,44), while the budding yeast can disseminate within the vagina or the intestine without causing overt symptoms 45.
C. albicans pathogenic sites
C. albicans is commonly commensally found within the oral cavity, the gastrointestinal tract (GI tract), the urogenital mucosa, and on the skin. The organism may also be found in the paranasal sinus mucosa, in blood, and within various internal organs. Within each host location (Table 1), C. albicans must adapt to the various prevailing local environmental conditions, which differ significantly between different anatomical locations in terms of nutrient availability, pH, O2, and CO2 concentrations, etc.61,62 Furthermore, C. albicans must adapt to co-exist within and as part of the community of the individually unique microbiomes present at each anatomical location.63 Occasionally, however, C. albicans can take advantage of some of the particular conditions described in Table 1., and can become pathogenic at a specific site.
Table 1.
Common candidiasis and their associated risk factors.
| Infection Site | Study approach | Year | Population | N | Risk factors | Ref |
|---|---|---|---|---|---|---|
| Oral candidiasis | Case control study | 2020 | HIV/AIDS patients | 207 | Old age, male gender, xerostomia, smoking, alcohol consumption, antibiotic use, low CD4 + T-cells count, HIV infection | 45 |
| Oral candidiasis | Cross-sectional study | 2020 | HIV/AIDS patients | 378 | Pregnancy, oral hygiene, antibiotic use, low CD4 + T-cells count | 46 |
| Oral candidiasis | Retrospective cohort study | 2018 | Patients receiving radiotherapy for head and neck cancer | 300 | Lymphocyte count, severity of oral mucositis during radiotherapy | 47 |
| Oral candidiasis | Prospective cohort study | 2013 | Patients with renal transplants | 1001 | Mycophenolate mofetil use, dentures, tobacco | 48 |
| Oropharyngeal | Prospective observational study | 2012 | Patients in intensive care unit | 110 | Proton pump inhibitor use, diabetes, lower BMI | 49 |
| Vulvovaginal candidiasis | Retrospective cohort study | 2017 | Women with clinical signs of vulvovaginal candidiasis or pregnancy | 2160 | Pregnancy | 50 |
| Vulvovaginal candidiasis | Cross-sectional Study | 2018 | Women of reproductive age | 97 | Sedentary life style, wearing tights, frequent intravaginal douching, first sexual encounter when under 20 years old, number of sexual partners, curettage, not cleaning the vulva before or after sexual encounters | 51 |
| Esophageal candidiasis | Retrospective cohort study | 2021 | Immunocompetent patients | 7736 | Diabetes, proton pump inhibitor use, atrophic gastritis, advanced gastric cancer, gastrectomy | 52 |
| Vulvovaginal candidiasis | Meta-analysis | 2021 | Iranian women | 10,536 | Oral contraceptive pills use, pregnancy, antibiotics use | 53 |
| Candidemia | Case-control study. | 2006 | Pediatric patients with congenital heart disease | 28 | Severity of clinical condition, long antibiotic treatment | 54 |
| Candidemia | Case-control study | 2016 | Non-neutropenic critical patients | 81 | Long hospital stay, abdominal surgery, the use of meropenem, hemodialysis | 55 |
| Candidaemia | Case-control study | 2021 | Patients with liver cirrhosis | 90 | Acute-on-chronic liver failure, gastrointestinal endoscopy, long antibiotic treatment, central venous catheter, total parenteral nutrition, long in-hospital stay | 56 |
| Candidaemia | Case-control study | 2021 | Adult | 118 | Neutropenia, solid organ transplant, significant liver, respiratory or cardiovascular disease, recent gastrointestinal infection, biliary or urological surgery, central venous access device, intravenous drug use, urinary catheter, carbapenem receipt | 57 |
| Pulmonary candidiasis | Case-control study | 2005 | Preterm infants | 20 | Male gender, preterm premature rupture of membranes, duration of ventilation | 58 |
| Disseminated candidiasis | Case-control study | 2004 | Children with candidemia | 153 | Persistently positive blood cultures for Candida with a central venous catheter, immunosuppression | 59 |
| Invasive candidiasis | Meta-analysis | 2022 | Critically ill patients | 1652 | Broad-spectrum antibiotics, blood transfusion, Candida colonization, central venous catheter, total parenteral nutrition | 60 |
N: Sample size; Ref: References; BMI: Body mass index.
As can be seen from Table 1, several organ systems and/or sites may be susceptible to C. albicans infection, thus illustrating the adaptive and immunopathological complexity and niche specificity that applies to host–pathogen interactions that mediate the commensal-to-pathogen shift by C. albicans. The present review focuses mainly on the presence of pathogenic C. albicans in the gut, and its interactions with the host gastrointestinal microbiome.
Pathogenic Candida albicans in the gut: interactions with the host microbiota
C. albicans can become a pathogen in circumstances where nutrient availability, required pH, and CO2 concentrations are not necessarily ideal. Since the host gut microbiome is rich in heterogeneous carbon sources, C. albicans can use these alternative carbon sources to enhance its survivability and virulence.62 This adaptability leads to the absence of catabolite inactivation,64 underlining the metabolic flexibility of C. albicans, and also contributes to alterations of its cellular proteome and secretome,65 which explains its ability to switch from yeast to hyphal forms, from white to opaque units,66 to form biofilm, to display adhesion characteristics,65,67 and to remodel its cell wall.62,68 Remodeling of the cell wall 62,68 has the capacity to (i) modify the pathogen’s sensitivity to environmental stress and to antifungal drugs,61,68 and (ii) alter the expression and the presentation of critical pathogen-associated molecular patterns (PAMPs). Consequently, C. albicans becomes a continually changing target for the host immune system.62,64
Evidence suggests that the gut is the main reservoir from which C. albicans translocates through the intestinal barrier, causing bloodstream infections.69 Normally, C. albicans inhabits the intestinal tract together with other fungal and bacterial members of the microbiome, preferentially as the yeast form; however, as shown in Table 1, C. albicans can initiate an infection in the gut and subsequently promote gut inflammation, especially under certain specific circumstances, including during HIV infection. Thereafter, alteration of the prevailing conventional microbial and environmental equilibrium can lead to microbial dysbiosis and Candida overgrowth in the gut, which thus becomes a potential source for emergence and development of systemic candidiasis.70 Although systemic dissemination of Candida (occurring via transepithelial transport of the yeast form) does not rely on morphological transition, it may be mediated by indirect mechanisms such as lumen sampling dendritic cells or microfold cell transcytosis, as suggested by Vautier et al.71
To fully understand the impact C. albicans has on an individual’s (both HIV-negative and HIV-positive) immune hemostasis, a substantially more detailed and accurate knowledge of the complex interactions that C. albicans has with host cells and bacteria and other elements of the gut microbiome, is ultimately warranted.
Interactions with host cells and host immune system
In the mouth, the vagina, and the gut, C. albicans interacts first with local epithelial and endothelial cells. Epithelial cells usually act as a passive physical barrier preventing C. albicans from invading through to underlying tissue.72 Epithelial integrity is usually maintained via tight junctions (interepithelial cell connections) which effectively seal off the gaps between the cell surface and the mucosal lamina propria, and which also prevents intraepithelial invasion of C. albicans.72 Intestinal epithelial cells also secrete protective substances (referred to as mucins) to form a mucus layer which restricts and impedes direct contact between C. albicans and epithelial cellular surfaces.73 Gastrointestinal epithelial cells can also secrete peptides or antimicrobial agents (lysozymes and β-defensins) which, combined with gastric acid, bile, and other digestive substances, work at controlling the proliferation of C. albicans within the gastrointestinal tract.72 However, two complementary mechanisms (both triggered by hyphal-associated factors 74) have been described with regards to C. albicans invasion through host epithelial cells. The initial stages of invasion are characterized by fungal-induced endocytosis, which is a process mediated by host-produced pseudopods which surround the fungal organism, and subsequently encapsulates and incorporates the fungus into the host cell. The second mechanism is the active penetration of fungal hyphae through or in between epithelial/endothelial cells.75 Furthermore, to overcome the antifungal influence exerted by the previously-mentioned peptides in the gut, C. albicans can (i) secrete peptide effectors (Sap9, Sap10, Msb2) to cleave host antifungal peptides 76 and inactivate a broad range of different antimicrobial peptides (AMPs) in a dose-dependent manner,77 and (ii) expel host antifungal peptides from the cytoplasm using efflux transporters such as Flu1.78
Recent investigations have shown that candidalysin, a cytolytic peptide toxin secreted by hyphae (the invasive form of C. albicans), is also of significant importance. Candidalysin originates from the ECE1 gene, which encodes the Ece1p protein. The Ece1p protein is then processed by Kex2p (after arginine residues are inserted at positions 61 and 93) to generate immature candidalysin. Finally, mature candidalysin is generated via Kex1p, which removes the terminal Arg93.79 Candidalysin is a critical virulence factor, capable of host cell activation, neutrophil recruitment, and Th17 cell-mediated immunity. It is now acknowledged that in healthy individuals, C. albicans which is undergoing a commensal lifestyle produces low levels of candidalysin.79 Then, in circumstances allowing C. albicans proliferation, increased candidalysin levels cause host cell and tissue damage.80,81 Concurrently, high levels of candidalysin activate protective innate responses through neutrophil recruitment, and Th17 immunity, altogether contributing to fungal clearance. In a similar manner, high levels of candidalysin can trigger overreactions of the immune response in certain infections (e.g., severe vulvovaginal candidiasis) and/or when the immune response is dysregulated.82
Once C. albicans is able to cope with and overcome the physical barrier formed by epithelial and endothelial cells, innate and adaptive defenses against C. albicans infection are required to be brought into play. The innate immune response is responsible for recognizing C. albicans via host cell pattern recognition receptors (PRRs) [Toll-like receptors (TLRs), C-type lectin receptors (CLRs), and Nod-like receptors (NLRs) 83,84] which interact with pathogen-associated molecular patterns (PAMPs) on the microbial cells (mostly cell-wall components such as β-glucan and mannan, as well as nucleic acid).1 Specifically, the recognition of Candida PAMPs is mediated by several TLRs, among which are TLR2 for the recognition of the phospholipomannan component of the cell wall, TLR4 for the recognition of short linear O-linked mannans, and TLR9 for the recognition of fungal DNA.85 This recognition (PRRs-PAMPs) therefore activates host countermeasures, including the secretion of AMPs (β-defensins, histatin, and cathelicidin LL-37), which are small molecules comprising 10–50 amino acids that are able to either kill C. albicans or inhibit its growth,86 and ultimately clear the fungal infection. However, C. albicans has the ability to also avoid recognition through the host PRRs by shielding its important PAMPs, such as β-glucan, with an outer mannoprotein layer of the cell wall, and therefore expose only a limited number of cell wall recognition sites.87 This strategy, however, renders C. albicans vulnerable to phagocytosis by macrophages.88–90 Furthermore, the innate immune system responds via epithelial cells which release proinflammatory mediators [cytokines/chemokines, particularly interleukin- (IL-) 8] in order to induce the recruitment of polymorphonuclear leucocytes (PMNs, such as neutrophils, macrophages, and dendritic cells, which are crucial for host defense against mucosal and disseminated candidiasis 91) from the circulating blood and direct them to the site of infection.92,93 According to Weindl et al., the addition of neutrophils to an epithelial model prevents both infection and cell damage by C. albicans.94 The preceding research team has demonstrated that this protective effect seems to be mediated by an upregulation of epithelial TLR4 upon stimulation with the PMN-derived tumor necrosis factor (TNF)-α. However, the precise underlying mechanisms responsible for such protection, mediated via TLR4 against C. albicans (and participating in the clearance of C. albicans), remains to be elucidated. Also, epithelial cells can themselves secrete granulocyte-macrophage colony-stimulating factor (GM-CSF), which stimulates polymorphonuclear cells and leukocytes to produce their own cytokines, including TNF-α.75 Once at the site of infection, PMNs can directly eliminate C. albicans through phagocytosis and degranulation.95 However, C. albicans does have the ability to switch from the white to the opaque form, rendering it effectively invisible to neutrophils.96 This invisibility subsequent to morphological ‘switching’ can be explained by the release of a chemoattractant only discovered to be present in the white form of C. albicans.97
Apart from form switching, C. albicans also degrades the complement factor C3b (well-known for its role in opsonization of microbial cells), and therefore inhibits recognition and uptake by phagocytic cells.98 However, after being phagocytosed, C. albicans yeasts has the capacity to morph into hyphae and thus escape from the phagocytic cell through activation of Efg1P via the cAMP/PKA pathway, resulting in the production of carbon dioxide and an alkaline pH within macrophages, for instance. This process has the potential to eventually kill the phagocytic cell, as has been demonstrated by past investigations.99–101 Similarly, macrophages have been found to be of particular importance with respect to fluid absorption in the colon. Indeed, Chikina et al., 102 observed that the innate immune system [via distal colon macrophages (CD11chigh)] allows a rapid quality check of absorbed fluids to avoid causing toxicity to colonocytes. These specific macrophages use “balloon-like” protrusions (which positively correlates with the level of toxins) to sample the fluids absorbed through epithelial cells. In cases where the fluids are enriched with fungal metabolites/toxins (candidalysin for instance), the macrophages (through the secretion of prostaglandin 2) instruct epithelial cells to cease absorption (prostaglandin 2 induces the diminution of aquaporin localization at the apical membrane of epithelial cells 103), and this prevents epithelial cell poisoning and death. In the absence of this species of macrophage (with their “balloon-like” protrusions), colonic barrier integrity may well be compromised, as epithelial cells are known to undergo apoptosis subsequent to absorption of fungal metabolites/toxins. C. albicans interactions with innate immune cells are complex; however, dendritic cells, by phagocytosing and presenting Candida-specific antigens, can be considered as a link between the innate and the adaptive response to C. albicans.104,105
With respect to the adaptive response, it has been demonstrated that T-helper cells may differentiate into a specific T-helper (Th) subset depending on the nature of the antigen presented, to assist in the clearance of the infection.106 Indeed, the adaptive immune response can result in the induction of either the Th1, Th2, Th17, or the Treg response,75,85,107 depending on various different factors, which include specific molecular structures and the morphology of the fungus.108 Thus, each of the preceding Th phenotypes obtained from the naïve CD4+ Th precursor cell produces a distinct set of cytokines, resulting in Th cell type-specific effector mechanisms.109 It has now been established that the Th1 response (induced by the inflammasome, which mediates the production of IL-18 and IL-12, thus stimulating the secretion of IFN-γ, which is the prototypal cytokine of the Th1 response 110) and Th2 response (induced via a chitin-dependent stimulation 88) are more likely present during systemic mycotic infection. While the Th1 response plays a crucial role in the defense against C. albicans 111 (due to IFN-γ, which activates neutrophils, leading to pathogen elimination via phagocytosis), the Th2 response is considered to be non-protective as it (i) induces a class switch to non-opsonizing antibody subclasses and IgE 112 and (ii) produces IL-4, IL-5, and IL-13, which promotes allergic responses and protection against helminths.109 With respect to Th1ʹs proinflammatory activity, characterized by IFN-γ release, C. albicans has been demonstrated to be able to interfere with or modulate cytokine release, and can thus inhibit the Th1 response by avoiding recognition by TLR4.113 Of particular concern is the fact that C. albicans can, conversely, favor TLR2 activation, which drives the immune system into a non-protective Th2 response and leads to the production of anti-inflammatory cytokines. Consequently, C. albicans can prevent macrophage activation and also the oxidative burst used to neutralize internalized pathogens.114 Aside from the Th1 and Th2 responses, Th17 cells mainly assist in controlling initial growth of the pathogen and preventing invasion of tissues.109,115 The Th17 response can be generated via several CLRs (dectin-1, dectin-2, and mannose receptor), the NLRP3 inflammasome, the induction of IL-23, IL-1β, IL-6, and transforming growth factor (TGF)-β.109 Thus, the Th17 response is directed to mucosal areas (through chemokine receptors CCR6 and CCR4) where Th17 cells release IL-17A, IL-17 F, and IL-22 (Th17-characterizing cytokines) to (i) induce the recruitment of neutrophils to the site of infection and (ii) drive the production of proinflammatory cytokines, chemokines and AMPs by epithelial cells.116 Yet, C. albicans (in the hyphal form) can evade the Th17 response by releasing tryptophan metabolic byproducts (5-hydroxytryptophan metabolites, which inhibit IL-17 production).117 Nevertheless, Th17 cell maturation takes time, and in the quest to ensure a permanent protection against C. albicans in the gut, several different immune cells [such as mucosa-associated group 3 innate lymphoid cells (ILCs)] can produce IL-17 until mature Th17 cells are recruited to the site of infection. In the absence of an immune response from Th17 cells, an excessive growth of C. albicans can occur on intestinal mucosa. This has been demonstrated in patients with hyper IgE syndrome (in which a mutation in the transcription factor STATA3 blocks important steps of the Th17-development pathway 109), chronic mucocutaneous candidiasis, or an autosomal recessive deficiency of the IL-17 cytokine receptor, IL-17RA,88 thus highlighting the importance of the CD4 + T-cell subset in the modulation of protection against C. albicans.92
As illustrated in Figure 2, host cells play an important role in interacting with C. albicans.
Figure 2.
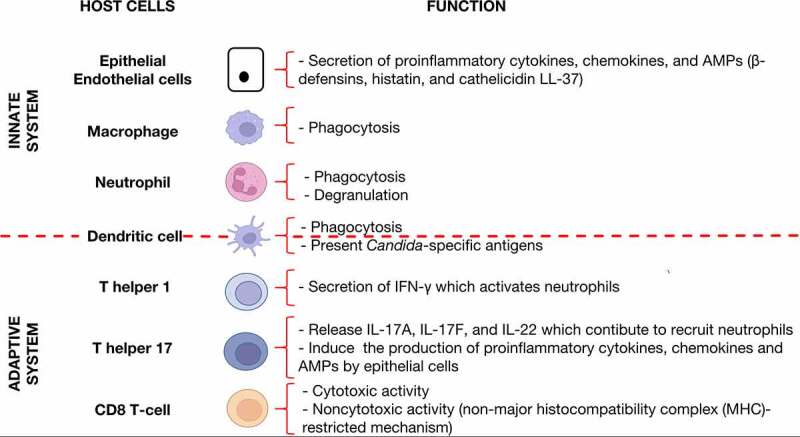
Specific host cells which contribute significantly to controlling C. albicans abundance in the gut.
Interactions with bacteria
In humans, it is generally accepted that C. albicans is a ubiquitous commensal component of the gut microbiome. This fungal organism resides in the intestines, within which continual and enduring inter-kingdom interactions occur with hundreds, and perhaps thousands, of bacterial taxa.118 Ultimately, the interactions between bacteria and C. albicans can be seen as interdependencies where bacterial functional and metabolic activities and products impinge upon C. albicans, and vice versa. In other words, fungi and bacteria influence the proliferation of each other in the gut. Interactions between C. albicans and different bacteria are summarized in Table 2, including synergistic and antagonistic effects.
Table 2.
The interactions between C. albicans and bacteria.
| Trend | Bacteria | Year | Model | Major findings | Ref |
|---|---|---|---|---|---|
| Synergistic | Staphylococcus aureus | 2019 | Mice | S. aureus can strongly adhere to C. albicans hyphae. Such adhesion is mediated by the Als3p protein of C. albicans, thereby promoting disseminated S. aureus disease. | 119 |
| 2017 | In vitro | C. albicans fungal film supports the adhesion and colonization of S. aureus through close interaction with hyphal elements, forming a polymicrobial biofilm, and enhancing miconazole resistance. | 120 | ||
| Streptococcus gordonii | 2014 | In vitro | In addition to C. albicans’ Als3 and S. gordonii’ SspB mediating co-aggregation between fungal and bacterial cells, Als1 was also found to bind S. gordonii. | 121 | |
| 2009 | In vitro | S. gordonii AgI/II proteins (SspA and SspB) mediate adhesion to C. albicans. S. gordonii alleviates the inhibitory effect of the quorum-sensing molecule farnesol on C. albicans hyphae and biofilm production. | 122 | ||
| Antagonistic | Klebsiella pneumoniae | 2021 | Mice | C. albicans antagonizes K. pneumonia, whereas Staphylococcus spp. may antagonize Candida. | 123 |
| Enterococcus faecalis | 2019 | Mice | E. faecalis peptide EntV requires disulfide bond formation and is cleaved by proteases to produce peptides that inhibit C. albicans proliferation. | 124 | |
| 2017 | Mice | E. faecalis competes for overlapping niches by producing EntV, a 68 amino acid peptide that inhibits hyphal morphogenesis, biofilm formation, and virulence in C. albicans. | 125 | ||
| Lactobacilli | 2022 | In vitro | Lactobacillus johnsonii MT4 exhibits pH-dependent and pH-independent antagonistic interactions against C. albicans by acidifying the local environment and producing soluble metabolites, thereby inhibiting C. albicans planktonic growth and biofilm formation. | 126 | |
| 2022 | In vitro | Lactobacillus plantarum inhibits the growth of C. albicans and Streptococcus mutans and disrupts S. mutans-C. albicans cross-kingdom biofilms. | 127 | ||
| 2013 | Mice | Lactobacilli use endogenous tryptophan as a carbon source to amplify and produce the aryl hydrocarbon receptor (AhR) ligand, indole-3 aldehyde (3-IAld), which triggers the production of IL-22 in the gut. This results in colonization resistance to C. albicans and protection of the mucosa from inflammation. | 128 | ||
| Salmonella enterica serovar Typhimurium | 2011 | In vitro | Killing of C. albicans filaments by S. typhimurium is mediated by sopB effectors. | 129 | |
| Anaerobic bacteria | 2015 | Mice | They limit the proliferation of C. albicans by stimulating the production of intestinal mucosal immune defenses, particularly cathelicidin-related antimicrobial peptide (CRAMP). | 130 | |
| Streptococcus mutans | 2010 | In vitro | Mutanobactin A, a secondary metabolite of S. mutans, affects the transformation of C. albicans from yeast to mycelium. | 131 | |
| 2010 | In vitro | S. mutans secretes trans-2-decenoic acid, a diffusible signaling factor, that inhibits filamentation of C. albicans. | 132 | ||
| 2009 | In vitro | S. mutans production capacity-stimulating peptide (CSP) inhibits C. albicans embryo tube formation and yeast-to-hyphal transition. | 133 | ||
| Pseudomonas aeruginosa | 2013 | In vitro | The action of phenazine produced by P. aeruginosa inhibits C. albicans filamentation, intercellular adhesion, and biofilm development. | 134 | |
| 2004 | In vitro | Inhibition of C. albicans filamentous formation by secretion of 3-oxo-C12 homoserine lactone and induction of filamentous reversion to yeast morphology in a N-acetylglucosamine-containing medium. | 135 |
Ref: References
It is well recognized that antibiotic treatments induce C. albicans overgrowth.136 In an identical manner, antifungal drug administration can lead to gut bacterial dysbiosis.137 Shotgun metagenomic sequencing has revealed that certain bacterial strains in the human gut inhibit C. albicans, and this process might be controlled naturally by bacterial metabolites such as propionate or 5-dodecenoate.136 It also appears that C. albicans has the capacity to mediate significant ecological changes in gut bacterial composition,138 given that it has been demonstrated that C. albicans is actively involved in the reconstitution of gut bacterial communities subsequent to antibiotic treatment.139 Recently, Rao et al., 123 have provided a clearer understanding of the existing interdependencies between specific fungi and bacterial taxa. Indeed, they have shown that C. albicans antagonizes Klebsiella pneumoniae whereas Staphylococcus spp. may antagonize Candida. Furthermore, an article by Sun et al., 140 has described the existence of a positive correlation between fungal and bacterial diversity in the gut. Although they failed to reveal specific correlations between fungi and bacteria at the species level, the observations of Sun et al., suggest a mutualism between these kingdoms of organisms. Peroumal et al., using 16S and 18S ribosomal DNA (rDNA) sequence analyses,141 reported that colonization of the gut by C. albicans modulates microbiome dynamics and alleviates uncontrolled body weight gain and metabolic hormonal imbalances in a murine high-fat-diet model.
Interactions with bacterial metabolites
Past reports have indicated that structural, functional, and metabolic products of gut bacteria may directly influence the proliferation of C. albicans. As such, the release of molecules such as cell surface components, peptides, or metabolic products may directly impact the biology of the fungus. Certain structural bacterial peptidoglycan (a major component of the bacterial cell wall) subunits (such as 1,6-anhydro-N-acetylmuramyl peptides) are highly effective hypha-inducing agents in C. albicans.142 In contrast, it has been demonstrated that EntV (a 68-amino-acid peptide), which is a product of the degradation of a 170-amino-acid prepropeptide produced by Enterococcus faecalis 124 (a commensal bacterium inhabiting the gastrointestinal tract of humans), inhibits the formation of C. albicans filaments, and thus prevents the formation of biofilms.125 Garcia et al., 143 have demonstrated, in vitro, that the human gut microbial metabolome (metabolites produced by a consortium of 60 bacterial strains derived from human feces) inhibits the proliferation of C. albicans to some extent, and also its ability to switch from yeast to filamentous forms. However, the identity of the specific metabolites involved as well as the identity of the actual bacterial species that produce these products are yet to be determined. Nevertheless, short-chain fatty acids (SCFAs), particularly acetic, butyric, and propionic acids have been shown to be able to inhibit germ tube formation, reduce metabolic activity in biofilms, and impair growth of C. albicans in vitro.144 Moreover, Nguyen et al., 145 have demonstrated that butyrate strongly inhibits yeast growth, virulence traits, and biofilm formation. Seelbinder et al., 136 have also demonstrated that acetic, propionic, and cis-5-dodecenoic acids reduce C. albicans-induced damage of cultured human epithelial cells. Also, in vitro evidence suggests that C. albicans enhances the growth of the strict anaerobes, Bacteroides fragilis and Bacteroides vulgatus.146 Authors of the studies reporting these findings have postulated that carbohydrates (such as α-mannan,147 located on the fungal surface) could fuel bacterial growth. Indeed, mannan digestion by Bacteroides enzymes may fuel the proliferation of Bacteroides species.
In addition to their direct effects on C. albicans, gut bacteria (via their products) can also indirectly influence the proliferation of C. albicans. This involves bacterial molecules (or the actual microbes themselves) which elicit responses from host cells, and the microbe-induced host response thus targets the fungus. This approach is known to be used by commensal bacteria to protect the mammalian host from invading pathogens.148 For example, Fan et al., 130 have shown that several Bacteroidetes and Clostridial Firmicutes restrict the proliferation of C. albicans by stimulating the production of gut mucosal immunological defenses, in particular the production of cathelin-related antimicrobial peptide (CRAMP).
Interactions with other fungi
Aside from bacteria, viruses, and archaea, numerous other varieties of fungi also reside in the human gut. Fungi can produce a variety of extracellular enzymes and mycotoxins which are harmful to other microbes, including C. albicans. Various fungi, such as Aspergillus, Trichoderma, Penicillium, and yeast strains show potential antagonism against C. albicans.149 In turn, studies have also reported that C. albicans can inhibit various species of filamentous fungi, such as Mucor spp and Chrysonilia sitophila.150 Rajasekharan et al., reported that zearalenone, a mycotoxin produced by several Fusarium and Gibberella species, inhibits C. albicans biofilm formation and hyphal morphogenesis.151 Similarly, deoxynivalenol, a trichothecene mycotoxin secreted by Fusarium, has been observed to reduce biofilm formation and metabolic activities of C. albicans.152 Moreover, antagonistic effects of yeasts against C. albicans has also been reported. Indeed, Türkel et al. have demonstrated that Metschnikowia pulcherrima strains, isolated from grapes, have strong antagonistic activity against C. albicans.153 Murzyn et al. also reported that Saccharomyces boulardii yeast cells and their extract inhibit virulence factors, hyphae formation, adhesion, and biofilm development in C. albicans.
The preceding observations indicate that, despite the mutual effects of C. albicans and bacteria on each other, interactions and functional crosstalk between C. albicans and other commensal fungi may also play an important role in the gut microbial ecosystem. However, our understanding of the complex regulation of, and factors involved in, these interactions remains limited, especially in the gut, and particularly in the context of HIV-infected people. Much more future research is warranted to clearly elucidate the underlying mechanisms governing these intra- and inter-mycotic relationships.
C. albicans immunopathogenesis during HIV infection
C. albicans is an integral commensal component of the healthy human gut microbiome; however, C. albicans can also be an opportunistic pathogen in the immunocompromised or in patients where structural and functional barriers that ordinarily prevent dissemination become disrupted or compromised.154 Because of its interactions with the gut microbiome, its ability to induce gut dysbiosis, and its ability to avoid immune system control, C. albicans infection should be considered as a potentially serious public health issue, especially in people living with HIV, where C. albicans pathogenicity may be significantly enhanced. In the following section, we further discuss C. albicans infection in the HIV-infected population.
HIV induces immunodeficiency
Once an individual is infected with HIV, their ability to competently orchestrate effective and efficient immune responses against various microorganisms becomes significantly compromised. HIV causes systemic CD4 + T-cell destruction, and results in impaired cell-mediated immunity, which leads to the inevitable emergence of various opportunistic infections,155 one example of which is the development of candidiasis (Figure 3). It is worth noting that Candida infections tend to manifest when CD4 + T-cell counts reduce to between 200 and 500 cells/µl, and may represent the first overt indication of a systemic immunodeficiency.156 Thus, Candida infections may be seen even in individuals on ART. Nevertheless, the initial step to either treating or preventing candidiasis in people living with HIV is an attempt to reconstitute the individual’s immune function by promptly initiating ART,157 as treating Candida infection in isolation in these individuals (with antifungal drugs such as fluconazole, clotrimazole, nystatin, or ketoconazole) does not either prevent recurrence or guarantee re-emergence of infection.
Figure 3.
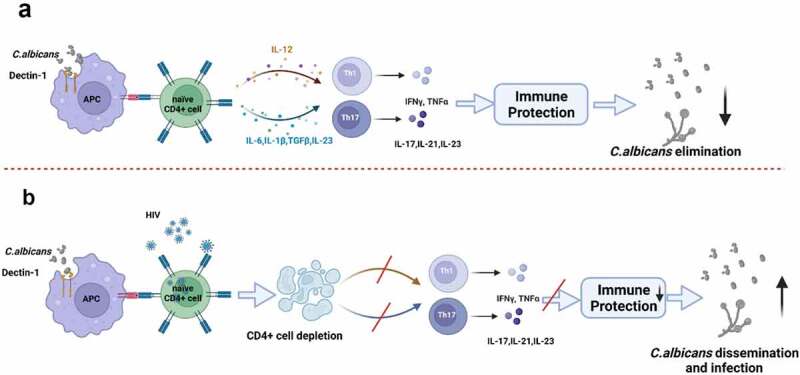
Changes of adaptive immunity for C. albicans in HIV-infected individuals. (a) In healthy individuals, commensal C. albicans colonize the host, preferably as a yeast form. They can be recognized, processed, and presented to CD4 + T cells by antigen presenting cells (APCs). Upon activation, naïve CD4+ Th precursor cells differentiate into protective effector T helper cells, Th1 and Th17, which produce multiple cytokines. These cytokines, such as IFN-γ, IL-17, and IL-21, activate immune responses to eliminate or control the growth and dissemination of C. albicans. (b) In HIV-infected individuals, CD4 + T-cells are invaded and depleted by HIV, especially in the intestine where CD4 + T-cells express high levels of the CCR5 co-receptor. Once CD4 + T-cell counts are reduced to threshold values, the effects of immune protection mediated by CD4 + T-cells decrease. In such a scenario, C. albicans would initiate overgrowth and disseminate, thus inducing different types of infection in the human host.
From an early stage of infection, HIV is known to subvert dendritic cell and macrophage activities to enhance its own replication at mucosal locations.158 In addition, intestinal CD4 + T-cells (which express high levels of the CCR5 co-receptor which facilitates entry of HIV virions into these cells) are significantly depleted.159 At both the mucosal level and within the gut, particularly, HIV contributes to progressive degradation of the immune system, and thus enables evasion by several microorganisms from routine immune system surveillance. CD4 + T-cell depletion in the gut occurs inexorably and rapidly after HIV infection,160 regardless of the route of exposure, and long before CD4 + T-cell reductions occur in blood or lymph nodes.161 Th1, Th17, and Th22 cells, which are specific subsets of CD4 + T-cells, critically important (i) for initiation of primary immune responses and (ii) for maintenance of mucosal integrity, are selectively targeted and eliminated first 160 (Figure 3). For example, Th1 cells (pro-inflammatory immune cells) are known to produce IFN-γ, TNF-α, and TNF-β, which primarily stimulate and promote cellular immune responses.162 Th17 cells, the preferential target for HIV reservoirs (as reported by Renault et al.,163), are abundant in mucosal-associated lymphoid tissues, and are recognized for their vital role in the immunological defense against intestinal bacterial and fungal infections.3,164 Th22 cells exclusively produce IL-22, and similar to Th17 cells, Th22 cells appear to have the capacity to acquire functional attributes of Th1 cells,165 which thus make them important immune cells for controlling C. albicans expansion in the gut.
In the relative absence of CD4 + T-cells (subsequent to their precipitous depletion by HIV/AIDS), CD8 + T-cells and epithelial cells become important in Candida-host interactions (Figure 2). Tissue biopsies from HIV positive patients [with or without oropharyngeal candidiasis (OPC)] have demonstrated an accumulation of activated CD8 + T-cells at the epithelium-lamina propria interface. However, it is worth noting that in the context of proven cases of OPC, CD8 + T-cells have been shown to be dysfunctional as they exhibit relatively small amounts of the adhesion molecule E-cadherin (which, in contrast, remain normal in OPC-negative individuals). This suggests that CD8 + T-cells may be recruited at the epithelial-lamina propria interface, where they normally have the ability to protect HIV positive individuals from developing oropharyngeal candidiasis if they normally express adhesion molecules such as E-cadherin. The preceding findings have been summarized in a past review by Fidel,166 who discusses the implications of HIV disease for oropharyngeal candidiasis. Instead of the recognized cytotoxic T-lymphocyte role played by CD8 + T-cells, Fidel has postulated that the effector activity of CD8 + T-cells in this context occurs via a non-major histocompatibility complex (MHC)-restricted mechanism (also referred to as the CD8 + T-cell noncytotoxic anti-HIV response),167 similar to a mechanism reported to be present in murine CD8 + T-cells cultured with IL-2.168,169 Although the preceding mechanisms have been demonstrated in the context of oropharyngeal candidiasis, they do not fully explain mechanisms which potentially occur within the gut during HIV infection. Therefore, further investigations are required in this regard.
In the absence of CD4 + T-cells the integrity of the gut cannot be ensured, and eventually leads to intestinal cell apoptosis and the progressive degradation of the tight junctions which seal the gaps between epithelial cell surfaces, both these processes contributing to the onset of leaky gut syndrome.170 In this context, HIV accelerates and exacerbates establishment of C. albicans infection, which may then easily disseminate and initiate blood stream infections. Apart from key immune cell depletion and subversion, HIV also actively participates in disruption of the natural bacterial composition and diversity within the gut, which ultimately further enhances C. albicans growth.
C. albicans interactions with gut bacteria during HIV infection
In an HIV positive individual’s gut, the interactions between C. albicans and other microorganisms are profoundly disturbed. Indeed, a reduction in abundance of some “beneficial” bacteria, including Akkermansia muciniphila, Bacteroides faecalis, Bacteroides vulvae, Diplococcus, and Arbuscular roseus 171 may be noted in HIV positive individuals. Interestingly, it has been reported that potentially pathogenic microorganisms such as Proteus, Enterococcus, Klebsiella, Shigella, and Streptococcus 171,172 tend to become predominant in these patients. In addition, Vujkovic-Cvijin and Somsouk,19 in an extensive review on gut microbiome perturbations in HIV infection (covering more than twenty studies), have reported that the intestines of HIV-infected individuals are enriched with Erysipelotrichaceae, Enterobacteriaceae, Desulfovibrionaceae, and Fusobacteria, but are depleted in Lachnospiraceae, Ruminococceae, Bacteroides, and Rikenellaceae. Altogether, these changes and the atypical balance of microorganisms within the gut impact on the growth and well-being of C. albicans, as described above. However, it remains unclear whether the changes to gut bacterial composition subsequent to HIV infection influence C. albicans growth positively or negatively. Thus, further investigation will help in providing a clearer understanding of precisely what occurs in this specific scenario. Nonetheless, the knowledge gleaned from contemporary literature allows for informed speculation around this. Indeed, Rao et al., 123 have demonstrated that C. albicans antagonizes Klebsiella pneumoniae whereas Staphylococcus spp. may antagonize Candida in infants. We believe that these findings may not be able to be reliably extrapolated to an adult context, and furthermore, may be unreasonable to extrapolate to an HIV infection context. Thus, it is possible that during HIV infection, the existing interdependencies between specific fungal and bacterial taxa exhibit different and changing trends. Otherwise, it would be difficult to explain the relatively high proportion of Candida in the guts of HIV positive individuals (39.5% as reported by Awoyeni et al., 173 and 52.5% as observed by Esebelahie et al., 174) despite the predominance of potentially pathogenic bacteria such as Klebsiella and Staphylococcus within the gut. Further investigations are warranted in this specific area of inquiry.
Results from previous investigations have pointed out the critical role of A. muciniphila in the gut, supporting intestinal mucosal homeostasis by modulating mucus thickness,175 leading academics to consider A. muciniphila as a potential “sentinel of the gut”.176 Indeed, A. muciniphila has the ability to (i) stimulate mucin production within the gut, (ii) improve enterocyte monolayer integrity, (iii) counteract inflammation, and (iv) induce intestinal adaptive immune responses.177,178 During HIV infection, a significantly lower intestinal abundance of A. muciniphila has been observed in PLWH. However, this is not an established conclusion, as the results of metagenomic studies have been heterogeneous, and not all of these studies have observed a reduction in this specific bacterial species in PLWH. Nevertheless, the lower abundance of A. muciniphila depends neither on whether or not ART treatment has been initiated, nor on prevailing CD4 + T-cell counts or viral loads. To our knowledge, there is no study in the published literature which clearly explains the possible mutual reciprocity existing between A. muciniphila and C. albicans, especially during HIV infection. However, in the light of available information gleaned from contemporary literature, it is possible to speculate on a negative association between these organisms, meaning that when A. muciniphila is abundant, C. albicans growth is inhibited, and vice versa. Indeed, since a major role of A. muciniphila is to prevent bacterial translocation in the context of the leaky gut syndrome, it is difficult to conceive of a positive association between these two organisms knowing that pathogenic C. albicans, in order to establish and disseminate infection, is likely to induce the leaky gut syndrome, as explained by Hofs et al.88 However, further investigations are required to clearly establish this possibility. In the relative absence of commensal bacteria (as seen during HIV infection), C. albicans is able to colonize epithelial surfaces and form hyphae. The subsequent inter- and intracellular invasion, therefore, facilitates the release of hydrolytic enzymes which profoundly damage epithelial cells, resulting in the loss of epithelial integrity (Figure 4).
Figure 4.
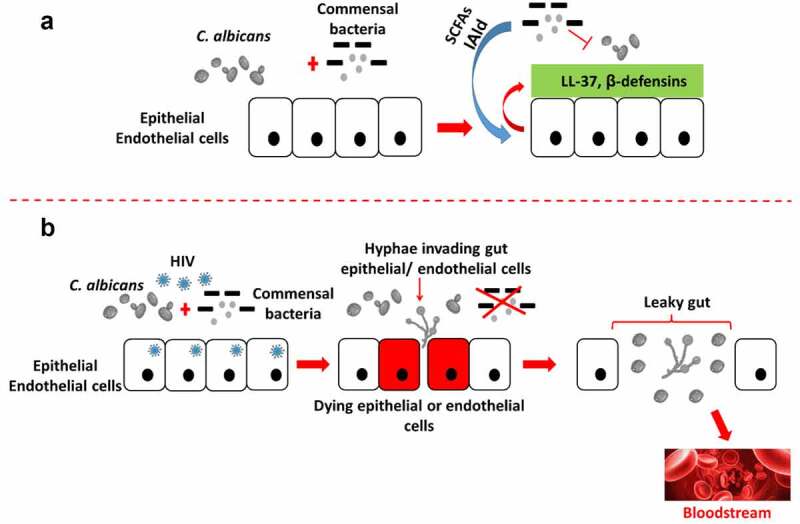
C. albicans interactions with host bacteria in the gut. In the absence of HIV infection, commensal bacterial actions (competition for adhesion sites, secretion of IAld and SCFAs) combined with host epithelial/endothelial cells secretion of β-defensins and LL-37 contribute to control C. albicans proliferation (a). However, during HIV infection, the reduced abundance of commensal bacteria favors C. albicans proliferation, which in turn develops hyphal forms and biofilm on and in-between epithelial/endothelial cells. Consequently, epithelial/endothelial cell integrity is disrupted, leading to their destruction, which further contributes to the establishment of the leaky gut syndrome (b). The epithelial/endothelial disruption eventually represents an “open gate”, so to speak, for C. albicans to disseminate into the bloodstream and thus initiate systemic infection and inflammation. SCFAs, short-chain fatty acids; IAld, indole-3-aldehyde.
Despite the evidence presented in the previous paragraphs, it is important to understand that the diversity and composition of the gut microbiome is also intimately linked to sexual preference. A study by Noguera-Julian et al., 179 has shown that men who have sex with men (MSM) have a Prevotella-rich enterotype, while non-MSM men are enriched in Bacteroides. In the preceding study, HIV negative MSM have been reported to possess a significantly richer and more diverse fecal microbiome compared to HIV negative non-MSM subjects. The preceding study also reported that HIV infection induced a diminution of bacterial richness in both categories of people (i.e., MSM and non-MSM). This said, we believe that the predominance of Bacteroides (which include Bacteroides faecalis and Bacteroides vulvae, well-known for their commensal role in the gut) in non-MSM subjects may indicate their predisposition to superior gut integrity, compared to MSM. In this regard, C. albicans proliferation in the non-MSM gut may likely be under relatively more stringent surveillance, compared to gay men. Therefore, much deeper investigation will be necessary to understand the biology of C. albicans in the gut (HIV-positive or not) based on sexual preference. Notwithstanding sexual preference, Rosas-Plaza et al., 180 have independently demonstrated that industrialized populations present less bacterial diversity in the gut microbiome than that from non-industrialized populations. Notably, Bacteroidaceae, Lanchospiraceae, and Rickenellaceae are predominant in the gut of populations living in modern societies. Their study involved comparing data collected from 14 studies, which represented the gut microbiome of 568 individuals from hunter-gatherers, agricultural, agropastoral, pastoral, and urban populations. Furthermore, they were able to establish that different lifestyles foster a distinct bacterial population diversity in the gut, and this supports the hypothesis that urbanization influences bacterial diversity in the human gut microbiome. Thus, urban populations and rural populations are likely to possess distinctly different microbiomes. In this regard, the influence of C. albicans on the gut of HIV positive individuals from either rural and urban populations remains to be further studied and clarified.
C. albicans takes advantage of microbiota dysbiosis and favors microbial translocation in PLWH
Normally, in an HIV negative individual, C. albicans may become pathogenic and colonize the gut subsequent to antibiotic drug treatment. Antibiotics non-selectively eliminate commensal bacteria that may inhibit the overgrowth of C. albicans through (i) a competition for adhesion sites, (ii) the secretion of inhibitory molecules [indole-3-aldehyde (IAld), SCFAs and lactic acid for example, which can inhibit the transition from yeast to hypha, and ensure a low environmental pH], and (iii) the induction of specific secretions from epithelial cells, such as β-defensins and LL-37. Thus, C. albicans is free to proliferate and switch from yeast to hyphal forms expressing adhesins/invasins, which are essential in the process of adhesion to and invasion of epithelial cells. In addition, C. albicans produces hydrolytic enzymes such as secreted aspartic proteases (Saps) to further facilitate the invasion of epithelial cells, to critically damage cells, and compromise epithelial integrity.88 In the absence of effective treatment, a progressive leaky gut syndrome is established. This scenario can be likened to C. albicans being a hidden ‘time-bomb’ waiting for a favorable opportunity to wreak havoc on the human body. Indeed, in the HIV infection context, in addition to impairment of the immune system’s ability to comprehensively respond to infections, there is a relative reduction of the abundance of commensal bacteria, including SCFA-producing bacteria (particularly the Ruminococcaceae and Lachnospiraceae taxa, which comprise the primary producers of SCFA).19 Furthermore, the bacterial microbiome of ART-treated PLWH can be depleted due to administration of preventive antibiotic drug treatment,181 especially in those with low CD4 + T-cell counts (also referred to as immunological nonresponders). Consequently, in the gut, a reduction of total SCFA production may promote the activation of intestinal T-lymphocytes, enhancing HIV replication, and the induction of further immune cell depletion.182
The microbial dysbiosis and the severe gut-associated immune cell depletion that takes place in the gut during HIV infection are now recognized to be associated with a degraded intestinal barrier, impaired mucosal immunological function, increased microbial translocation, and long-term immune activation in PLWH.19,176,183 In other words, HIV itself induces gut-associated microbiota dysbiosis, and increases susceptibility to the development of leaky gut syndrome. HIV infection therefore favors the pathogenicity of C. albicans, which in turn can easily exacerbate damage to the gut epithelial barrier and promote microbial translocation. Indeed, as HIV infection progresses, C. albicans (i) has no further competition for adhesion sites, (ii) can easily proliferate, and (iii) can proceed unchecked with its invasion of the epithelial barrier (Figure 4). HIV positive individuals are consistently found to have evidence of microbial translocation, even those on ART, with a suppressed viral replication and load.184,185 As such, microbial translocation markers such as LPS remain substantially high after ART initiation, compared to HIV-negative individuals.186 With the combined influence of C. albicans proliferation and HIV infection, the circulating levels of such markers cannot return to normal. Furthermore, it has been demonstrated that plasma LPS and bacterial DNA levels in ART-treated patients inversely correlate with CD4 + T-cell reconstitution in the gut-associated lymphoid tissue (GALT).187,188 Thus, we speculate that therapeutic interventions to block microbial translocation in ART-treated patients may be important for complete reconstitution of gastrointestinal immunity. To illustrate this, it is pertinent to refer to the immune status of immunological nonresponders, who maintain high levels of microbial translocation and immune activation, and fail to restore their CD4 + T-cell population, despite long-term ART,187,189 suggesting that microbial translocation can potentially thwart the likelihood of ART success. Thus, C. albicans in the gut can, indeed, be regarded as a mycopathological ‘time-bomb’ which may jeopardize immune reconstitution and also ART success in HIV positive individuals.
Finally, the influence of HIV on distal colonic macrophages (CD11chigh) and their “balloon like” protrusions, as defined by Chikina et al., 102 as regulators of fluid absorption by the colon, is worth exploring. Infection of these cells by HIV is likely to (i) decrease the population of these specific macrophages, (ii) dysregulate their protrusion formation, (iii) dysregulate the rapid quality check of absorbed fluids in the colon, (iv) induce toxicity of colonocytes by candidalysin generated by C. albicans, and (v) favor epithelial cell death (apoptosis). In such a context, C. albicans will further degrade gut integrity, increase microbial translocation, and aggravate the HIV-positive individuals’ clinical state.
Conclusions
To conclude, it appears from study of the available literature that the immunopathogenesis of C. albicans in humans has now been relatively well described. Overt Candida infections, resulting from pathological activity of C. albicans in particular locations of the body, are inextricably linked to interactions and interdependencies of C. albicans with other members of the local communal microbiome. In this article, we have explored how C. albicans, a commensal colonizer of most humans, and an opportunistic pathogen in the immunocompromised or in patients in which barriers that prevent dissemination have been disrupted, can evade immunological and commensal bacterial surveillance and regulation, to become pathogenic. As such, Candida represents an ongoing threat to every individual. In the particular context of HIV infection, where both the immune system and commensal bacterial populations are profoundly destabilized, C. albicans can therefore be considered to be a mycological ‘time-bomb’, waiting the perfect moment to strike and exacerbate microbial dysbiosis, leading to the leaky gut syndrome, subsequent microbial translocation, and the undesirable and potentially deadly consequences thereof. Thus, C. albicans should be considered as an important public health threat, even in this era of affordable and widely available ART, as microbial translocation may prevent the reconstitution of gastrointestinal immunity and the overall success of ART treatment. Lastly, we believe a close monitoring of C. albicans abundance within the gut of HIV positive individuals, particularly at the immunological level (using CD4 + T-cell counts, CD4+/CD8+ ratios, and measures of chronic systemic inflammation), and its associations, correlations, and interdependencies with both pathogenic and non-pathogenic commensal microbiome in the gut could assist in fostering a more comprehensive understanding and prevention of the consequences of C. albicans translocation in humans.
Funding Statement
This work was supported by the Chongqing Talent Cultivation Program (cstc2021ycjh-bgzxm0275), the Joint Medical Research Project (2020GDRC010) of Chongqing Science & Technology Bureau and Chongqing Health Commission, and the Chinese Federation of Public Health foundation (GWLM202024).
Disclosure statement
The authors report no conflicts of interest.
Author contributions
SZ and JO wrote the first draft of the manuscript. SI, XZ, VH, and HC provided critical revision of the manuscript. YC conceived and designed the study. All authors read and approved the manuscript, and agreed to its submission for publication.
Data availability statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
References
- 1.Williams DW, Jordan RP, Wei XQ, CT Alves, MP Wise, MJ Wilson, MA Lewis. Interactions of Candida albicans with host epithelial surfaces. J Oral Microbiol. 2013;5. doi: 10.3402/jom.v5i0.22434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Pfaller MA, Diekema DJ.. Epidemiology of invasive mycoses in North America. Crit Rev Microbiol. 2010;36(1):1–24. doi: 10.3109/10408410903241444. [DOI] [PubMed] [Google Scholar]
- 3.Khader SA, Gaffen SL, Kolls JK. Th17 cells at the crossroads of innate and adaptive immunity against infectious diseases at the mucosa. Mucosal Immunol. 2009;2(5):403–411. doi: 10.1038/mi.2009.100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Tati S, Davidow P, McCall A, Hwang-Wong E, Rojas IG, Cormack B, Edgerton M. Candida glabrata binding to Candida albicans hyphae enables its development in oropharyngeal candidiasis. PLoS Pathog. 2016;12(3):e1005522. doi: 10.1371/journal.ppat.1005522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lewis MAO, Williams DW. Diagnosis and management of oral candidosis. Br Dent J. 2017;223(9):675–681. doi: 10.1038/sj.bdj.2017.886. [DOI] [PubMed] [Google Scholar]
- 6.McCullough MJ, Clemons KV, Stevens DA. Molecular epidemiology of the global and temporal diversity of Candida albicans. Clin Infect Dis. 1999;29(5):1220–1225. doi: 10.1086/313455. [DOI] [PubMed] [Google Scholar]
- 7.Chen H, Zhou X, Ren B, Cheng L. The regulation of hyphae growth in Candida albicans. Virulence. 2020;11(1):337–348. doi: 10.1080/21505594.2020.1748930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lalla RV, Latortue MC, Hong CH, Ariyawardana A, D’Amato-Palumbo S, DJ Fischer, Martof A, Nicolatou-Galitis O, LL Patton, LS Elting, FK Spijkervet. A systematic review of oral fungal infections in patients receiving cancer therapy. Support Care Cancer. 2010;18(8):985–992. doi: 10.1007/s00520-010-0892-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hajjeh RA, Sofair AN, Harrison LH, GM Lyon, BA Arthington-Skaggs, SA Mirza, Phelan M, Morgan J, Lee-Yang W, MA Ciblak, LE Benjamin. Incidence of bloodstream infections due to Candida species and in vitro susceptibilities of isolates collected from 1998 to 2000 in a population-based active surveillance program. J Clin Microbiol. 2004;42(4):1519–1527. doi: 10.1128/jcm.42.4.1519-1527.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gow NA, van de Veerdonk FL, Brown AJ, Netea MG. Candida albicans morphogenesis and host defence: discriminating invasion from colonization. Nat Rev Microbiol. 2011;10(2):112–122. doi: 10.1038/nrmicro2711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhou PR, Hua H, Liu XS. Quantity of Candida colonies in saliva: a diagnostic evaluation for oral candidiasis. Chin J Dent Res. 2017;20(1):27–32. doi: 10.3290/j.cjdr.a37739. [DOI] [PubMed] [Google Scholar]
- 12.Kauffman CA. Fungal infections. Proc Am Thorac Soc. 2006;3(1):35–40. doi: 10.1513/pats.200510-110JH. [DOI] [PubMed] [Google Scholar]
- 13.Sobel JD. Vulvovaginal candidosis. Lancet. 2007;369(9577):1961–1971. doi: 10.1016/s0140-6736(07)60917-9. [DOI] [PubMed] [Google Scholar]
- 14.de Repentigny L, Lewandowski D, Jolicoeur P. Immunopathogenesis of oropharyngeal candidiasis in human immunodeficiency virus infection. Clin Microbiol Rev. 2004;17(4):729–759. doi: 10.1128/cmr.17.4.729-759.2004. table of contents [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hung CC, Yang YL, Lauderdale TL, McDonald LC, Hsiao C-F, Cheng -H-H, Ho YA, Lo H-J. Colonization of human immunodeficiency virus-infected outpatients in taiwan with candida species. J Clin Microbiol. 2005;43(4):1600–1603. doi: 10.1128/jcm.43.4.1600-1603.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ouyang J, Isnard S, Lin J, Fombuena B, Marette A, Routy B, Chen Y, Routy J-P. Metformin effect on gut microbiota: insights for HIV-related inflammation. AIDS Res Ther. 2020;17(1):10. doi: 10.1186/s12981-020-00267-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tong Y, Tang J. Candida albicans infection and intestinal immunity. Microbiol Res. 2017;198:27–35. doi: 10.1016/j.micres.2017.02.002. [DOI] [PubMed] [Google Scholar]
- 18.Hager CL, Ghannoum MA. The mycobiome in HIV. Curr Opin HIV AIDS. 2018;13(1):69–72. doi: 10.1097/coh.0000000000000432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Vujkovic-Cvijin I, Somsouk M. HIV and the gut microbiota: composition, consequences, and avenues for amelioration. Curr HIV/AIDS Rep. 2019;16(3):204–213. doi: 10.1007/s11904-019-00441-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jackson AP, Gamble JA, Yeomans T, Moran GP, Saunders D, Harris D, Aslett M, Barrell JF, Butler G, Citiulo F, et al. Comparative genomics of the fungal pathogens Candida dubliniensis and Candida albicans. Genome Res. 2009;19(12):2231–2244. doi: 10.1101/gr.097501.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mishra PK, Baum M, Carbon J. Centromere size and position in Candida albicans are evolutionarily conserved independent of DNA sequence heterogeneity. Mol Genet Genomics. 2007;278(4):455–465. doi: 10.1007/s00438-007-0263-8. [DOI] [PubMed] [Google Scholar]
- 22.Wang JM, Bennett RJ, Anderson MZ, Nielsen K. The genome of the human pathogen Candida albicans is shaped by mutation and cryptic sexual recombination. mBio. 2018;9(5):5. doi: 10.1128/mBio.01205-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Butler G, Rasmussen MD, Lin MF, Santos MAS, Sakthikumar S, Munro CA, Rheinbay E, Grabherr M, Forche A, Reedy JL, et al. Evolution of pathogenicity and sexual reproduction in eight Candida genomes. Nature. 2009;459(7247):657–662. doi: 10.1038/nature08064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hirakawa MP, Martinez DA, Sakthikumar S, MZ Anderson, Berlin A, Gujja S, Zeng Q, Zisson E, JM Wang, JM Greenberg, Berman J. Genetic and phenotypic intra-species variation in Candida albicans. Genome Res. 2015;25(3):413–425. doi: 10.1101/gr.174623.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.National Institutes of Health . Candida albicans SC5314. 2016. https://www.ncbi.nlm.nih.gov/data-hub/taxonomy/237561/. Accessed 16 Dec 2022.
- 26.d’Enfert C, Kaune AK, Alaban LR, Chakraborty S, Cole N, Delavy M, Kosmala D, Marsaux B, Fróis-Martins R, Morelli M, Rosati D. The impact of the Fungus-Host-Microbiota interplay upon Candida albicans infections: current knowledge and new perspectives. FEMS Microbiol Rev. 2021;45:3. doi: 10.1093/femsre/fuaa060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Soll DR. The role of phenotypic switching in the basic biology and pathogenesis of Candida albicans. J Oral Microbiol. 2014:6. doi: 10.3402/jom.v6.22993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jacobsen ID, Wilson D, Wächtler B, Brunke S, Naglik JR, Hube B. Candida albicans dimorphism as a therapeutic target. Expert Rev Anti Infect Ther. 2012;10(1):85–93. doi: 10.1586/eri.11.152. [DOI] [PubMed] [Google Scholar]
- 29.Mayer FL, Wilson D, Hube B. Candida albicans pathogenicity mechanisms. Virulence. 2013;4(2):119–128. doi: 10.4161/viru.22913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Berman J, Sudbery PE. Candida Albicans: a molecular revolution built on lessons from budding yeast. Nat Rev Genet. 2002;3(12):918–930. doi: 10.1038/nrg948. [DOI] [PubMed] [Google Scholar]
- 31.Saville SP, Lazzell AL, Monteagudo C, Lopez-Ribot JL. EngineeredControl of cell morphology in vivo reveals distinct roles for yeast and filamentous forms of Candida albicans duringInfection. Eukaryot Cell. 2003;2(5):1053–1060. doi: 10.1128/ec.2.5.1053-1060.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Netea MG, Joosten LA, van der Meer JW, Kullberg B-J, van de Veerdonk FL. Immune defence against Candida fungal infections. Nat Rev Immunol. 2015;15(10):630–642. doi: 10.1038/nri3897. [DOI] [PubMed] [Google Scholar]
- 33.Calderone RA, Fonzi WA. Virulence factors of Candida albicans. Trends Microbiol. 2001;9(7):327–335. doi: 10.1016/s0966-842x(01)02094-7. [DOI] [PubMed] [Google Scholar]
- 34.Fox EP, Cowley ES, Nobile CJ, Hartooni N, Newman D, Johnson A. Anaerobic bacteria grow within Candida albicans biofilms and induce biofilm formation in suspension cultures. Curr Biol. 2014;24(20):2411–2416. doi: 10.1016/j.cub.2014.08.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pande K, Chen C, Noble SM. Passage through the mammalian gut triggers a phenotypic switch that promotes Candida albicans commensalism. Nat Genet. 2013;45(9):1088–1091. doi: 10.1038/ng.2710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Forche A, Alby K, Schaefer D, Johnson AD, Berman J, Bennett RJ. The parasexual cycle in Candida albicans provides an alternative pathway to meiosis for the formation of recombinant strains. PLoS Biol. 2008;6(5):e110. doi: 10.1371/journal.pbio.0060110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bennett RJ, Johnson AD. Completion of a parasexual cycle in Candida albicans by induced chromosome loss in tetraploid strains. Embo J. 2003;22(10):2505–2515. doi: 10.1093/emboj/cdg235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kett DH, Azoulay E, Echeverria PM, Vincent J-L. Candida bloodstream infections in intensive care units: analysis of the extended prevalence of infection in intensive care unit study. Crit Care Med. 2011;39(4):665–670. doi: 10.1097/CCM.0b013e318206c1ca. [DOI] [PubMed] [Google Scholar]
- 39.Kullberg BJ, Arendrup MC. Invasive Candidiasis. N Engl J Med. 2015;373(15):1445–1456. doi: 10.1056/NEJMra1315399. [DOI] [PubMed] [Google Scholar]
- 40.Brown GD, Denning DW, Gow NA, SM Levitz, MG Netea, White TC. Hidden killers: human fungal infections. Sci Transl Med. 2012;4(165):165rv13. doi: 10.1126/scitranslmed.3004404. [DOI] [PubMed] [Google Scholar]
- 41.Erwig LP, Gow NA. Interactions of fungal pathogens with phagocytes. Nat Rev Microbiol. 2016;14(3):163–176. doi: 10.1038/nrmicro.2015.21. [DOI] [PubMed] [Google Scholar]
- 42.Uwamahoro N, Verma-Gaur J, Shen HH, Qu Y, Lewis R, Lu J, Bambery K, Masters SL, Vince JE, Naderer T, et al. The pathogen Candida albicans hijacks pyroptosis for escape from macrophages. mBio. 2014;5(2):e00003–14. doi: 10.1128/mBio.00003-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gow NA, Brown AJ, Odds FC. Fungal morphogenesis and host invasion. Curr Opin Microbiol. 2002;5(4):366–371. doi: 10.1016/s1369-5274(02)00338-7. [DOI] [PubMed] [Google Scholar]
- 44.Kumamoto CA, Vinces MD. Alternative Candida albicans lifestyles: growth on surfaces. Annu Rev Microbiol. 2005;59(1):113–133. doi: 10.1146/annurev.micro.59.030804.121034. [DOI] [PubMed] [Google Scholar]
- 45.Suryana K, Suharsono H, Antara I. Factors associated with oral candidiasis in people living with HIV/AIDS: a case control study. HIV AIDS (Auckl). 2020;12:33–39. doi: 10.2147/hiv.s236304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ambe NF, Longdoh NA, Tebid P, Bobga TP, Nkfusai CN, Ngwa SB, Nsai FS, Cumber SN. The prevalence, risk factors and antifungal sensitivity pattern of oral candidiasis in HIV/AIDS patients in Kumba District Hospital, South West Region, Cameroon. Pan Afr Med J. 2020;36:23. doi: 10.11604/pamj.2020.36.23.18202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kawashita Y, Funahara M, Yoshimatsu M, Kawashita Y, Funahara M, Yoshimatsu M, Nakao N, Soutome S, Saito T, Umeda M. A retrospective study of factors associated with the development of oral candidiasis in patients receiving radiotherapy for head and neck cancer: is topical steroid therapy a risk factor for oral candidiasis? Medicine (Baltimore). 2018;97(44):e13073. doi: 10.1097/md.0000000000013073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.López-Pintor RM, Hernández G, de Arriba L, de Andres A. Oral candidiasis in patients with renal transplants. Med Oral Patol Oral Cir Bucal. 2013;18(3):e381–7. doi: 10.4317/medoral.18658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mojazi Amiri H, Frandah W, Colmer-Hamood J, Raj R, Nugent K. Risk factors of Candida colonization in the oropharynx of patients admitted to an intensive care unit. J Mycol Med. 2012;22(4):301–307. doi: 10.1016/j.mycmed.2012.08.001. [DOI] [PubMed] [Google Scholar]
- 50.Mtibaa L, Fakhfakh N, Kallel A, Belhadj S, Belhaj Salah N, Bada N, Kallel K. Vulvovaginal candidiasis: etiology, symptomatology and risk factors. J Mycol Med. 2017;27(2):153–158. doi: 10.1016/j.mycmed.2017.01.003. [DOI] [PubMed] [Google Scholar]
- 51.Zeng X, Zhang Y, Zhang T, Xue Y, Xu H, An R. Risk factors of vulvovaginal candidiasis among women of reproductive age in Xi’an: a cross-sectional study. Biomed Res Int. 2018;2018:9703754. doi: 10.1155/2018/9703754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ogiso H, Adachi S, Mabuchi M, Horibe Y, Ohno T, Suzuki Y, Yamauchi O, Kojima T, Takada E, Iwama M, et al. Risk factors for the development of esophageal candidiasis among patients in community hospital. Sci Rep. 2021;11(1):20663. doi: 10.1038/s41598-021-00132-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Sasani E, Rafat Z, Ashrafi K, Salimi Y, Zandi M, Soltani S, Hashemi F, Hashemi SJ. Vulvovaginal candidiasis in Iran: a systematic review and meta-analysis on the epidemiology, clinical manifestations, demographic characteristics, risk factors, etiologic agents and laboratory diagnosis. Microb Pathog. 2021;154:104802. doi: 10.1016/j.micpath.2021.104802. [DOI] [PubMed] [Google Scholar]
- 54.Garcia-San Miguel L, Cobo J, Martos I, Otheo E, Muriel A, Pintado V, Moreno S. Risk factors for candidemia in pediatric patients with congenital heart disease. Infect Control Hosp Epidemiol. 2006;27(6):576–580. doi: 10.1086/505094. [DOI] [PubMed] [Google Scholar]
- 55.Ortíz Ruiz G, Osorio J, Valderrama S, Álvarez D, Elías Díaz R, Calderón J, Ballesteros D, Franco A. Risk factors for candidemia in non-neutropenic critical patients in Colombia. Med Intensiva. 2016;40(3):139–144. doi: 10.1016/j.medin.2015.08.001. [DOI] [PubMed] [Google Scholar]
- 56.Bartoletti M, Rinaldi M, Pasquini Z, et al. Risk factors for candidaemia in hospitalized patients with liver cirrhosis: a multicentre case-control-control study. Clin Microbiol Infect. 2021;27(2):276–282. doi: 10.1016/j.cmi.2020.04.030. [DOI] [PubMed] [Google Scholar]
- 57.Keighley CL, Pope A, Marriott DJE, Chapman B, Bak N, Daveson K, Hajkowicz K, Halliday C, Kennedy K, Kidd S, et al. Risk factors for candidaemia: a prospective multi-centre case-control study. Mycoses. 2021;64(3):257–263. doi: 10.1111/myc.13211. [DOI] [PubMed] [Google Scholar]
- 58.Frezza S, Maggio L, De Carolis MP, Gallini F, Puopolo M, Polimeni V, Costa S, Vento G, Tortorolo G. Risk factors for pulmonary candidiasis in preterm infants with a birth weight of less than 1250 g. Eur J Pediatr. 2005;164(2):88–92. doi: 10.1007/s00431-004-1571-1. [DOI] [PubMed] [Google Scholar]
- 59.Zaoutis TE, Greves HM, Lautenbach E, Bilker WB, Coffin SE. Risk factors for disseminated candidiasis in children with candidemia. Pediatr Infect Dis J. 2004;23(7):635–641. doi: 10.1097/01.inf.0000128781.77600.6f. [DOI] [PubMed] [Google Scholar]
- 60.Thomas-Rüddel DO, Schlattmann P, Pletz M, Kurzai O, Bloos F. Risk factors for invasive Candida infection in critically Ill patients: a systematic review and meta-analysis. Chest. 2022;161(2):345–355. doi: 10.1016/j.chest.2021.08.081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Brown AJ, Budge S, Kaloriti D, Tillmann A, Jacobsen MD, Yin Z, Ene IV, Bohovych I, Sandai D, Kastora S, et al. Stress adaptation in a pathogenic fungus. J Exp Biol. 2014;217(Pt 1):144–155. doi: 10.1242/jeb.088930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hall RA. Dressed to impress: impact of environmental adaptation on the Candida albicans cell wall. Mol Microbiol. 2015;97(1):7–17. doi: 10.1111/mmi.13020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Morgan XC, Segata N, Huttenhower C. Biodiversity and functional genomics in the human microbiome. Trends Genet. 2013;29(1):51–58. doi: 10.1016/j.tig.2012.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Childers DS, Raziunaite I, Mol Avelar G, Mackie J, Budge S, Stead D, Gow NAR, Lenardon MD, Ballou ER, MacCallum DM, et al. The rewiring of ubiquitination targets in a pathogenic yeast promotes metabolic flexibility, host colonization and virulence. PLoS Pathog. 2016;12(4):e1005566. doi: 10.1371/journal.ppat.1005566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ene IV, Heilmann CJ, Sorgo AG, Walker LA, de Koster CG, Munro CA, Klis FM, Brown AJP. Carbon source-induced reprogramming of the cell wall proteome and secretome modulates the adherence and drug resistance of the fungal pathogen C andida albicans. Proteomics. 2012;12(21):3164–3179. doi: 10.1002/pmic.201200228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Lan CY, Newport G, Murillo LA, Jones T, Scherer S, Davis RW, Agabian N. Metabolic specialization associated with phenotypic switching in Candida albicans. Proc Natl Acad Sci U S A. 2002;99(23):14907–14912. doi: 10.1073/pnas.232566499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Ene IV, Adya AK, Wehmeier S, Brand AC, MacCallum DM, Gow NAR, Brown AJP. Host carbon sources modulate cell wall architecture, drug resistance and virulence in a fungal pathogen. Cell Microbiol. 2012;14(9):1319–1335. doi: 10.1111/j.1462-5822.2012.01813.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Brown AJ, Brown GD, Netea MG, Gow NAR. Metabolism impacts upon Candida immunogenicity and pathogenicity at multiple levels. Trends Microbiol. 2014;22(11):614–622. doi: 10.1016/j.tim.2014.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Nucci M, Anaissie E. Revisiting the source of candidemia: skin or gut? Clin Infect Dis. 2001;33(12):1959–1967. doi: 10.1086/323759. [DOI] [PubMed] [Google Scholar]
- 70.Alonso-Monge R, Gresnigt MS, Román E, Hube B, Pla J. Candida albicans colonization of the gastrointestinal tract: a double-edged sword. PLoS Pathog. 2021;17(7):e1009710. doi: 10.1371/journal.ppat.1009710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Vautier S, Drummond RA, Chen K, Murray GI, Kadosh D, Brown AJP, Gow NAR, MacCallum DM, Kolls JK, Brown GD, et al. C andida albicans colonization and dissemination from the murine gastrointestinal tract: the influence of morphology and Th17 immunity. Cell Microbiol. 2015;17(4):445–450. doi: 10.1111/cmi.12388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Yan L, Yang C, Tang J. Disruption of the intestinal mucosal barrier in Candida albicans infections. Microbiol Res. 2013;168(7):389–395. doi: 10.1016/j.micres.2013.02.008. [DOI] [PubMed] [Google Scholar]
- 73.Vulvovaginal CA. Candida albicans infections: pathogenesis, immunity and vaccine prospects. Bjog. 2015;122(6):785–794. doi: 10.1111/1471-0528.12994. [DOI] [PubMed] [Google Scholar]
- 74.Yang W, Yan L, Wu C, Zhao X, Tang J. Fungal invasion of epithelial cells. Microbiol Res. 2014;169(11):803–810. doi: 10.1016/j.micres.2014.02.013. [DOI] [PubMed] [Google Scholar]
- 75.Naglik JR, Moyes DL, Wächtler B, Hube B. Candida albicans interactions with epithelial cells and mucosal immunity. Microbes Infect. 2011;13(12–13):963–976. doi: 10.1016/j.micinf.2011.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Meiller TF, Hube B, Schild L, Shirtliff ME, Scheper MA, Winkler R, Ton A, Jabra-Rizk MA. A novel immune evasion strategy of candida albicans: proteolytic cleavage of a salivary antimicrobial peptide. PLoS One. 2009;4(4):e5039. doi: 10.1371/journal.pone.0005039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Szafranski-Schneider E, Swidergall M, Cottier F, Tielker D, Román E, Pla J, Ernst JF. Msb2 shedding protects Candida albicans against antimicrobial peptides. PLoS Pathog. 2012;8(2):e1002501. doi: 10.1371/journal.ppat.1002501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Li R, Kumar R, Tati S, Puri S, Edgerton M. Candida albicans flu1-mediated efflux of salivary histatin 5 reduces its cytosolic concentration and fungicidal activity. Antimicrob Agents Chemother. 2013;57(4):1832–1839. doi: 10.1128/aac.02295-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Naglik JR, Gaffen SL, Hube B. Candidalysin: discovery and function in Candida albicans infections. Curr Opin Microbiol. 2019;52:100–109. doi: 10.1016/j.mib.2019.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Blagojevic M, Camilli G, Maxson M, Hube B, DL Moyes, JP Richardson, JR Naglik. Candidalysin triggers epithelial cellular stresses that induce necrotic death. Cell Microbiol. 2021;23(10):e13371. doi: 10.1111/cmi.13371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Mogavero S, Sauer FM, Brunke S, Allert S, Schulz D, Wisgott S, Jablonowski N, Elshafee O, Krüger T, Kniemeyer O, AA Brakhage. Candidalysin delivery to the invasion pocket is critical for host epithelial damage induced by Candida albicans. Cell Microbiol. 2021;23(10):e13378. doi: 10.1111/cmi.13378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Wu Y, Zeng Z, Guo Y, Song L, JE Weatherhead, Huang X, Zeng Y, Bimler L, CY Chang, JM Knight, Valladolid C. Candida albicans elicits protective allergic responses via platelet mediated T helper 2 and T helper 17 cell polarization. Immunity. 2021;54(11):2595–2610.e7. doi: 10.1016/j.immuni.2021.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Hollmig ST, Ariizumi K, Cruz PD Jr. Recognition of non-self-polysaccharides by C-type lectin receptors dectin-1 and dectin-2. Glycobiology. 2009;19(6):568–575. doi: 10.1093/glycob/cwp032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Kumagai Y, Takeuchi O, Akira S. Pathogen recognition by innate receptors. J Infect Chemother. 2008;14(2):86–92. doi: 10.1007/s10156-008-0596-1. [DOI] [PubMed] [Google Scholar]
- 85.Cambi A, Netea MG, Mora-Montes HM, NA Gow, SV Hato, DW Lowman, BJ Kullberg, Torensma R, DL Williams, CG Figdor. Dendritic cell interaction with Candida albicans critically depends on N-linked mannan. J Biol Chem. 2008;283(29):20590–20599. doi: 10.1074/jbc.M709334200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Peschel A, Sahl HG. The co-evolution of host cationic antimicrobial peptides and microbial resistance. Nat Rev Microbiol. 2006;4(7):529–536. doi: 10.1038/nrmicro1441. [DOI] [PubMed] [Google Scholar]
- 87.Gantner BN, Simmons RM, Underhill DM. Dectin-1 mediates macrophage recognition of Candida albicans yeast but not filaments. Embo J. 2005;24(6):1277–1286. doi: 10.1038/sj.emboj.7600594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Höfs S, Mogavero S, Hube B. Interaction of Candida albicans with host cells: virulence factors, host defense, escape strategies, and the microbiota. J Microbiol. 2016;54(3):149–169. doi: 10.1007/s12275-016-5514-0. [DOI] [PubMed] [Google Scholar]
- 89.Gow NA, Netea MG, Munro CA, Ferwerda G, Bates S, HM Mora-Montes, Walker L, Jansen T, Jacobs L, Tsoni V, GD Brown. Immune recognition of Candida albicans beta-glucan by dectin-1. J Infect Dis. 2007;196(10):1565–1571. doi: 10.1086/523110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Wheeler RT, Kombe D, Agarwala SD, GR Fink. Dynamic, morphotype-specific Candida albicans beta-glucan exposure during infection and drug treatment. PLoS Pathog. 2008;4(12):e1000227. doi: 10.1371/journal.ppat.1000227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.van ‘t Wout JW, Linde I, Leijh PC, Van Furth R. Contribution of granulocytes and monocytes to resistance against experimental disseminated Candida albicans infection. Eur J Clin Microbiol Infect Dis. 1988;7(6):736–741. doi: 10.1007/bf01975039. [DOI] [PubMed] [Google Scholar]
- 92.van der Meer JW, van de Veerdonk FL, Joosten LA, BJ Kullberg, MG Netea. Severe Candida spp. infections: new insights into natural immunity. Int J Antimicrob Agents. 2010;(36 Suppl 2):S58–62. doi: 10.1016/j.ijantimicag.2010.11.013. [DOI] [PubMed] [Google Scholar]
- 93.Hebecker B, Naglik JR, Hube B, ID Jacobsen. Pathogenicity mechanisms and host response during oral Candida albicans infections. Expert Rev Anti Infect Ther. 2014;12(7):867–879. doi: 10.1586/14787210.2014.916210. [DOI] [PubMed] [Google Scholar]
- 94.Weindl G, Naglik JR, Kaesler S, Biedermann T, Hube B, HC Korting, Schaller M. Human epithelial cells establish direct antifungal defense through TLR4-mediated signaling. J Clin Invest. 2007;117(12):3664–3672. doi: 10.1172/jci28115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Urban CF, Ermert D, Schmid M, Abu-Abed U, Goosmann C, Nacken W, Brinkmann V, PR Jungblut, Zychlinsky A. Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans. PLoS Pathog. 2009;5(10):e1000639. doi: 10.1371/journal.ppat.1000639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Geiger J, Wessels D, Lockhart SR, DR Soll. Release of a potent polymorphonuclear leukocyte chemoattractant is regulated by white-opaque switching in Candida albicans. Infect Immun. 2004;72(2):667–677. doi: 10.1128/iai.72.2.667-677.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Sasse C, Hasenberg M, Weyler M, Gunzer M, Morschhäuser J. White-opaque switching of Candida albicans allows immune evasion in an environment-dependent fashion. Eukaryot Cell. 2013;12(1):50–58. doi: 10.1128/ec.00266-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Gropp K, Schild L, Schindler S, Hube B, PF Zipfel, Skerka C. The yeast Candida albicans evades human complement attack by secretion of aspartic proteases. Mol Immunol. 2009;47(2–3):465–475. doi: 10.1016/j.molimm.2009.08.019. [DOI] [PubMed] [Google Scholar]
- 99.Marcil A, Harcus D, Thomas DY, Whiteway M. Candida albicans killing by RAW 264.7 mouse macrophage cells: effects of Candida genotype, infection ratios, and gamma interferon treatment. Infect Immun. 2002;70(11):6319–6329. doi: 10.1128/iai.70.11.6319-6329.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Ghosh S, Navarathna DH, Roberts DD, Cooper JT, Atkin AL, Petro TM, Nickerson KW. Arginine-induced germ tube formation in Candida albicans is essential for escape from murine macrophage line RAW 264.7. Infect Immun. 2009;77(4):1596–1605. doi: 10.1128/iai.01452-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Vylkova S, Carman AJ, Danhof HA, Collette JR, Zhou H, Lorenz MC. The fungal pathogen Candida albicans autoinduces hyphal morphogenesis by raising extracellular pH. mBio. 2011;2(3):e00055–11. doi: 10.1128/mBio.00055-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Chikina AS, Nadalin F, Maurin M, San-Roman M, Thomas-Bonafos T, Li XV, Lameiras S, Baulande S, Henri S, Malissen B, et al. Macrophages maintain epithelium integrity by limiting fungal product absorption. Cell. 2020;183(2):411–428.e16. doi: 10.1016/j.cell.2020.08.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Ikarashi N, Baba K, Ushiki T, Kon R, Mimura A, Toda T, Ishii M, Ochiai W, Sugiyama K. The laxative effect of bisacodyl is attributable to decreased aquaporin-3 expression in the colon induced by increased PGE 2 secretion from macrophages. Am J Physiol Gastrointest Liver Physiol. 2011;301(5):G887–95. doi: 10.1152/ajpgi.00286.2011. [DOI] [PubMed] [Google Scholar]
- 104.Hickey DK, Patel MV, Fahey JV, Wira CR. Innate and adaptive immunity at mucosal surfaces of the female reproductive tract: stratification and integration of immune protection against the transmission of sexually transmitted infections. J Reprod Immunol. 2011;88(2):185–194. doi: 10.1016/j.jri.2011.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Cheng SC, Joosten LA, Kullberg BJ, Netea MG. Interplay between Candida albicans and the mammalian innate host defense. Infect Immun. 2012;80(4):1304–1313. doi: 10.1128/iai.06146-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Sallusto F, Lanzavecchia A. The instructive role of dendritic cells on T-cell responses. Arthritis Res. 2002;4(4 Suppl 3 (Suppl3)):S127–32. doi: 10.1186/ar567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Cambi A, Gijzen K, de Vries L J, Torensma R, Joosten B, Adema G, Netea M, Kullberg B-J, Romani L, Figdor C, et al. The C-type lectin DC-SIGN (CD209) is an antigen-uptake receptor for Candida albicans on dendritic cells. Eur J Immunol. 2003;33(2):532–538. doi: 10.1002/immu.200310029. [DOI] [PubMed] [Google Scholar]
- 108.Feller L, Khammissa RA, Chandran R, Altini M, Lemmer J. Oral candidosis in relation to oral immunity. J Oral Pathol Med. 2014;43(8):563–569. doi: 10.1111/jop.12120. [DOI] [PubMed] [Google Scholar]
- 109.Conti HR, Gaffen SL. Host responses to Candida albicans: th17 cells and mucosal candidiasis. Microbes Infect. 2010;12(7):518–527. doi: 10.1016/j.micinf.2010.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.van de Veerdonk FL, Joosten LA, Netea MG. The interplay between inflammasome activation and antifungal host defense. Immunol Rev. 2015;265(1):172–180. doi: 10.1111/imr.12280. [DOI] [PubMed] [Google Scholar]
- 111.Weindl G, Wagener J, Schaller M. Epithelial cells and innate antifungal defense. J Dent Res. 2010;89(7):666–675. doi: 10.1177/0022034510368784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Netea MG, Maródi L. Innate immune mechanisms for recognition and uptake of Candida species. Trends Immunol. 2010;31(9):346–353. doi: 10.1016/j.it.2010.06.007. [DOI] [PubMed] [Google Scholar]
- 113.Luo S, Skerka C, Kurzai O, Zipfel PF. Complement and innate immune evasion strategies of the human pathogenic fungus Candida albicans. Mol Immunol. 2013;56(3):161–169. doi: 10.1016/j.molimm.2013.05.218. [DOI] [PubMed] [Google Scholar]
- 114.Netea MG, Sutmuller R, Hermann C, Van der Graaf CAA, Van der Meer JWM, van Krieken JH, Hartung T, Adema G, Kullberg BJ. Toll-like receptor 2 suppresses immunity against Candida albicans through induction of IL-10 and regulatory T cells. J Immunol. 2004;172(6):3712–3718. doi: 10.4049/jimmunol.172.6.3712. [DOI] [PubMed] [Google Scholar]
- 115.De Luca A, Zelante T, D’Angelo C, Zagarella S, Fallarino F, Spreca A, Iannitti RG, Bonifazi P, Renauld J-C, Bistoni F, et al. IL-22 defines a novel immune pathway of antifungal resistance. Mucosal Immunol. 2010;3(4):361–373. doi: 10.1038/mi.2010.22. [DOI] [PubMed] [Google Scholar]
- 116.Onishi RM, Gaffen SL. Interleukin-17 and its target genes: mechanisms of interleukin-17 function in disease. Immunology. 2010;129(3):311–321. doi: 10.1111/j.1365-2567.2009.03240.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Cheng SC, van de Veerdonk F, Smeekens S, Joosten LAB, van der Meer JWM, Kullberg B-J, Netea MG. Candida albicans dampens host defense by downregulating IL-17 production. J Immunol. 2010;185(4):2450–2457. doi: 10.4049/jimmunol.1000756. [DOI] [PubMed] [Google Scholar]
- 118.Pérez JC. Candida albicans dwelling in the mammalian gut. Curr Opin Microbiol. 2019;52:41–46. doi: 10.1016/j.mib.2019.04.007. [DOI] [PubMed] [Google Scholar]
- 119.Allison DL, Scheres N, Willems HME, Bode CS, Krom BP, Shirtliff ME. The host immune system facilitates disseminated staphylococcus aureus disease due to phagocytic attraction to Candida albicans during coinfection: a case of bait and switch. Infect Immun. 2019;87(11):11. doi: 10.1128/iai.00137-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Kean R, Rajendran R, Haggarty J, Townsend EM, Short B, Burgess KE, Lang S, Millington O, Mackay WG, Williams C, et al. Candida albicans mycofilms support staphylococcus aureus colonization and enhances miconazole resistance in dual-species interactions. Front Microbiol. 2017;8:258. doi: 10.3389/fmicb.2017.00258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Hoyer LL, Oh SH, Jones R, Cota E. A proposed mechanism for the interaction between the Candida albicans Als3 adhesin and streptococcal cell wall proteins. Front Microbiol. 2014;5:564. doi: 10.3389/fmicb.2014.00564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Bamford CV, d’Mello A, Nobbs AH, Dutton LC, Vickerman MM, Jenkinson HF. Streptococcus gordonii modulates Candida albicans biofilm formation through intergeneric communication. Infect Immun. 2009;77(9):3696–3704. doi: 10.1128/iai.00438-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Rao C, Coyte KZ, Bainter W, Geha RS, Martin CR, Rakoff-Nahoum S. Multi-kingdom ecological drivers of microbiota assembly in preterm infants. Nature. 2021;591(7851):633–638. doi: 10.1038/s41586-021-03241-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Brown AO, Graham CE, Cruz MR, Singh KV, Murray BE, Lorenz MC, Garsin DA. Antifungal activity of the enterococcus faecalis peptide entv requires protease cleavage and disulfide bond formation. mBio. 2019;10(4):4. doi: 10.1128/mBio.01334-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Graham CE, Cruz MR, Garsin DA, Lorenz MC. Enterococcus faecalis bacteriocin EntV inhibits hyphal morphogenesis, biofilm formation, and virulence of Candida albicans. Proc Natl Acad Sci U S A. 2017;114(17):4507–4512. doi: 10.1073/pnas.1620432114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Vazquez-Munoz R, Thompson A, Russell JT, Sobue T, Zhou Y, Dongari-Bagtzoglou A. Insights from the lactobacillus johnsonii genome suggest the production of metabolites with antibiofilm activity against the pathobiont Candida albicans. Front Microbiol. 2022;13:853762. doi: 10.3389/fmicb.2022.853762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Zeng Y, Fadaak A, Alomeir N, Wu TT, Rustchenko E, Qing S, Bao J, Gilbert C, Xiao J. Lactobacillus plantarum disrupts S. mutans-C. albicans cross-kingdom biofilms. Front Cell Infect Microbiol. 2022;12:872012. doi: 10.3389/fcimb.2022.872012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Zelante T, Iannitti RG, Cunha C, De Luca A, Giovannini G, Pieraccini G, Zecchi R, D’Angelo C, Massi-Benedetti C, Fallarino F, et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity. 2013;39(2):372–385. doi: 10.1016/j.immuni.2013.08.003. [DOI] [PubMed] [Google Scholar]
- 129.Kim Y, Mylonakis E. Killing of Candida albicans filaments by salmonella enterica serovar typhimurium is mediated by sopB effectors, parts of a type III secretion system. Eukaryot Cell. 2011;10(6):782–790. doi: 10.1128/ec.00014-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Fan D, Coughlin LA, Neubauer MM, Kim J, Kim MS, Zhan X, Simms-Waldrip TR, Xie Y, Hooper LV, Koh AY, et al. Activation of HIF-1α and LL-37 by commensal bacteria inhibits Candida albicans colonization. Nat Med. 2015;21(7):808–814. doi: 10.1038/nm.3871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Joyner PM, Liu J, Zhang Z, Merritt J, Qi F, Cichewicz RH. Mutanobactin A from the human oral pathogen Streptococcus mutans is a cross-kingdom regulator of the yeast-mycelium transition. Org Biomol Chem. 2010;8(24):5486–5489. doi: 10.1039/c0ob00579g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Vílchez R, Lemme A, Ballhausen B, Thiel V, Schulz S, Jansen R, Sztajer H, Wagner-Döbler I. Streptococcus mutans inhibits Candida albicans hyphal formation by the fatty acid signaling molecule trans-2-decenoic acid (SDSF). Chembiochem. 2010;11(11):1552–1562. doi: 10.1002/cbic.201000086. [DOI] [PubMed] [Google Scholar]
- 133.Jarosz LM, Deng DM, van der Mei HC, Crielaard W, Krom BP. Streptococcus mutans Competence-Stimulating Peptide Inhibits Candida albicans Hypha Formation. Eukaryot Cell. 2009;8(11):1658–1664. doi: 10.1128/ec.00070-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Morales DK, Grahl N, Okegbe C, Dietrich LEP, Jacobs NJ, Hogan DA. Control of Candida albicans metabolism and biofilm formation by pseudomonas aeruginosa phenazines. mBio. 2013;4(1):e00526–12. doi: 10.1128/mBio.00526-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Hogan DA, Vik A, Kolter R. A Pseudomonas aeruginosa quorum-sensing molecule influences Candida albicans morphology. Mol Microbiol. 2004;54(5):1212–1223. doi: 10.1111/j.1365-2958.2004.04349.x. [DOI] [PubMed] [Google Scholar]
- 136.Seelbinder B, Chen J, Brunke S, Vazquez-Uribe R, Santhaman R, Meyer A-C, de Oliveira Lino FS, Chan K-F, Loos D, Imamovic L, et al. Antibiotics create a shift from mutualism to competition in human gut communities with a longer-lasting impact on fungi than bacteria. Microbiome. 2020;8(1):133. doi: 10.1186/s40168-020-00899-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Wheeler ML, Limon JJ, Bar AS, Leal C, Gargus M, Tang J, Brown J, Funari V, Wang H, Crother T, et al. Immunological consequences of intestinal fungal dysbiosis. Cell Host Microbe. 2016;19(6):865–873. doi: 10.1016/j.chom.2016.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.van Tilburg Bernardes E, Pettersen VK, Gutierrez MW, Laforest-Lapointe I, Jendzjowsky NG, Cavin J-B, Vicentini FA, Keenan CM, Ramay HR, Samara J, et al. Intestinal fungi are causally implicated in microbiome assembly and immune development in mice. Nat Commun. 2020;11(1):2577. doi: 10.1038/s41467-020-16431-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Erb Downward JR, Falkowski NR, Mason KL, Muraglia R, GB Huffnagle. Modulation of post-antibiotic bacterial community reassembly and host response by Candida albicans. Sci Rep. 2013;3(1):2191. doi: 10.1038/srep02191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Sun Y, Zuo T, Cheung CP, Gu W, Wan Y, Zhang F, Chen N, Zhan H, Yeoh YK, Niu J, et al. Population-level configurations of gut mycobiome across 6 ethnicities in urban and rural China. Gastroenterology. 2021;160(1):272–286.e11. doi: 10.1053/j.gastro.2020.09.014. [DOI] [PubMed] [Google Scholar]
- 141.Peroumal D, Sahu SR, Kumari P, Utkalaja BG, Acharya N. Commensal fungus Candida albicans maintains a long-term mutualistic relationship with the host to modulate gut microbiota and metabolism. Microbiol Spectr. 2022;10(5):e0246222. doi: 10.1128/spectrum.02462-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Xu XL, Lee RT, Fang HM, YM Wang, Li R, Zou H, Zhu Y, Wang Y. Bacterial peptidoglycan triggers Candida albicans hyphal growth by directly activating the adenylyl cyclase Cyr1p. Cell Host Microbe. 2008;4(1):28–39. doi: 10.1016/j.chom.2008.05.014. [DOI] [PubMed] [Google Scholar]
- 143.García C, Tebbji F, Daigneault M, Liu -N-N, Köhler JR, Allen-Vercoe E, Sellam A. The human gut microbial metabolome modulates fungal growth via the TOR signaling pathway. mSphere. 2017;2(6):6. doi: 10.1128/mSphere.00555-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Guinan J, Wang S, Hazbun TR, Yadav H, Thangamani S. Antibiotic-induced decreases in the levels of microbial-derived short-chain fatty acids correlate with increased gastrointestinal colonization of Candida albicans. Sci Rep. 2019;9(1):8872. doi: 10.1038/s41598-019-45467-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Nguyen LN, Lopes LC, Cordero RJ, Nosanchuk JD. Sodium butyrate inhibits pathogenic yeast growth and enhances the functions of macrophages. J Antimicrob Chemother. 2011;66(11):2573–2580. doi: 10.1093/jac/dkr358. [DOI] [PubMed] [Google Scholar]
- 146.Valentine M, Benadé E, Mouton M, Khan W, Botha A. Binary interactions between the yeast Candida albicans and two gut-associated Bacteroides species. Microb Pathog. 2019;135:103619. doi: 10.1016/j.micpath.2019.103619. [DOI] [PubMed] [Google Scholar]
- 147.Cuskin F, Lowe EC, Temple MJ, Zhu Y, Cameron EA, Pudlo NA, Porter NT, Urs K, Thompson AJ, Cartmell A, et al. Human gut Bacteroidetes can utilize yeast mannan through a selfish mechanism. Nature. 2015;517(7533):165–169. doi: 10.1038/nature13995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Khosravi A, Mazmanian SK. Disruption of the gut microbiome as a risk factor for microbial infections. Curr Opin Microbiol. 2013;16(2):221–227. doi: 10.1016/j.mib.2013.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Li H, Yang J, Zhang X, Xu X, Song F, Li H. Biocontrol of Candida albicans by antagonistic microorganisms and bioactive compounds. Antibiotics (Basel). 2022;11:9. doi: 10.3390/antibiotics11091238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Janabi AA. Ali D, NA Mohammed, Rahem F. Vitro Antagonistic Activity of Candida Albicans against Filamentous Fungi. Med Mycol Open Access. 2015;1:2. doi: 10.21767/2471-8521.100002. [DOI] [Google Scholar]
- 151.Rajasekharan SK, Lee JH, Zhao Y, Lee J. The mycotoxin zearalenone hinders Candida albicans biofilm formation and hyphal morphogenesis. Indian J Microbiol. 2018;58(1):19–27. doi: 10.1007/s12088-017-0690-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Rajasekharan SK, Byun J, Lee J. Inhibitory effects of deoxynivalenol on pathogenesis of Candida albicans. J Appl Microbiol. 2018;125(5):1266–1275. doi: 10.1111/jam.14032. [DOI] [PubMed] [Google Scholar]
- 153.Türkel S, Ener B. Isolation and characterization of new Metschnikowia pulcherrima strains as producers of the antimicrobial pigment pulcherrimin. Z Naturforsch C J Biosci. 2009;64(5–6):405–410. doi: 10.1515/znc-2009-5-618. [DOI] [PubMed] [Google Scholar]
- 154.da Silva Dantas A, Lee KK, Raziunaite I, Schaefer K, Wagener J, Yadav B, Gow NA. Cell biology of Candida albicans-host interactions. Curr Opin Microbiol. 2016;34:111–118. doi: 10.1016/j.mib.2016.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Lucas S, Nelson AM. Nelson AM. HIV and the spectrum of human disease. J Pathol. 2015;235(2):229–241. doi: 10.1002/path.4449. [DOI] [PubMed] [Google Scholar]
- 156.Bharathi M, Rani AU. Pathogenic fungal isolates in sputum of HIV positive patients. Journal of AIDS and HIV Research. 2011;3:107–113. [Google Scholar]
- 157.Masur H, Brooks JT, Benson CA, Holmes KK, Pau AK, Kaplan JE. Prevention and treatment of opportunistic infections in HIV-infected adults and adolescents: updated guidelines from the centers for disease control and prevention, national institutes of health, and hiv medicine association of the infectious diseases society of America. Clin Infect Dis. 2014;58(9):1308–1311. doi: 10.1093/cid/ciu094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Weinberg A, Tugizov S, Pandiyan P, Jin G, Rakshit S, Vyakarnam A, JR Naglik. Innate immune mechanisms to oral pathogens in oral mucosa of HIV-infected individuals. Oral Dis. 2020;26(26 Suppl 1 (Suppl1)):69–79. doi: 10.1111/odi.13470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Nwosu FC, Avershina E, Wilson R, Rudi K. Gut microbiota in HIV infection: implication for disease progression and management. Gastroenterol Res Pract. 2014;2014:803185. doi: 10.1155/2014/803185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Veazey RS. Intestinal CD4 depletion in HIV/SIV infection. Curr Immunol Rev. 2019;15(1):76–91. doi: 10.2174/1573395514666180605083448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Connor S. HIV is active even in latency, say scientists. Bmj. 1993;306(6883):949. doi: 10.1136/bmj.306.6883.949. [DOI] [PubMed] [Google Scholar]
- 162.Wan YY, Flavell RA. How diverse–CD4 effector T cells and their functions. J Mol Cell Biol. 2009;1(1):20–36. doi: 10.1093/jmcb/mjp001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Renault C, Veyrenche N, Mennechet F, Bedin A-S, Routy J-P, Van de Perre P, Reynes J, Tuaillon E. Th17 CD4+ T-cell as a preferential target for HIV reservoirs. Front Immunol. 2022;13:822576. doi: 10.3389/fimmu.2022.822576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Peck A, Mellins ED, Peck A, Mellins ED. Precarious balance: th17 cells in host defense. Infect Immun. 2010;78(1):32–38. doi: 10.1128/iai.00929-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Sallusto F, Zielinski CE, Human LA. Th17 subsets. Eur J Immunol. 2012;42(9):2215–2220. doi: 10.1002/eji.201242741. [DOI] [PubMed] [Google Scholar]
- 166.Fidel PL Jr. Candida-host interactions in HIV disease: implications for oropharyngeal candidiasis. Adv Dent Res. 2011;23(1):45–49. doi: 10.1177/0022034511399284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Mackewicz CE, Patterson BK, Lee SA, Levy JA. CD8(+) cell noncytotoxic anti-human immunodeficiency virus response inhibits expression of viral RNA but not reverse transcription or provirus integration. J Gen Virol. 2000;81(Pt 5):1261–1264. doi: 10.1099/0022-1317-81-5-1261. [DOI] [PubMed] [Google Scholar]
- 168.Beno DW, Mathews HL. Growth inhibition of Candida albicans by interleukin-2-activated splenocytes. Infect Immun. 1992;60(3):853–863. doi: 10.1128/iai.60.3.853-863.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Beno DW, Stöver AG, Mathews HL. Growth inhibition of Candida albicans hyphae by CD8+ lymphocytes. J Immunol. 1995;154(10):5273–5281. doi: 10.4049/jimmunol.154.10.5273. [DOI] [PubMed] [Google Scholar]
- 170.Ouyang J, Zaongo SD, Zhang X, Qi M, Hu A, Wu H, Chen Y. Microbiota-mediated immunity abnormalities facilitate hepatitis B virus co-infection in people living with HIV: a review. Front Immunol. 2021;12:755890. doi: 10.3389/fimmu.2021.755890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Geng ST, Zhang ZY, Wang YX, Lu D, Yu J, Zhang J-B, Kuang Y-Q, Wang K-H. Regulation of gut microbiota on immune reconstitution in patients with acquired immunodeficiency syndrome. Front Microbiol. 2020;11:594820. doi: 10.3389/fmicb.2020.594820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Zhou Y, Ou Z, Tang X, Zhou Y, Xu H, Wang X, Li K, He J, Du Y, Wang H, et al. Alterations in the gut microbiota of patients with acquired immune deficiency syndrome. J Cell Mol Med. 2018;22(4):2263–2271. doi: 10.1111/jcmm.13508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Awoyeni A, Olaniran O, Odetoyin B, Hassan-Olajokun R, Olopade B, Afolayan D, Adekunle O. Isolation and evaluation of Candida species and their association with CD4+ T cells counts in HIV patients with diarrhoea. Afr Health Sci. 2017;17(2):322–329. doi: 10.4314/ahs.v17i2.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Esebelahie NO, Enweani IB, Omoregie R. Candida colonisation in asymptomatic HIV patients attending a tertiary hospital in Benin City, Nigeria. Libyan J Med. 2013;8(1):20322. doi: 10.3402/ljm.v8i0.20322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Microbiome: HU. A mucus colonizer manages host metabolism. Nat Rev Microbiol. 2013;11(7):430–431. doi: 10.1038/nrmicro3051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Ouyang J, Lin J, Isnard S, Fombuena B, Peng X, Marette A, Routy B, Messaoudene M, Chen Y, Routy J-P, et al. The bacterium akkermansia muciniphila: a sentinel for gut permeability and its relevance to HIV-related inflammation. Front Immunol. 2020;11:645. doi: 10.3389/fimmu.2020.00645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Ansaldo E, Slayden LC, Ching KL, Koch MA, Wolf NK, Plichta DR, Brown EM, Graham DB, Xavier RJ, Moon JJ, et al. Akkermansia muciniphila induces intestinal adaptive immune responses during homeostasis. Science. 2019;364(6446):1179–1184. doi: 10.1126/science.aaw7479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Huck O, Mulhall H, Rubin G, Kizelnik Z, Iyer R, Perpich JD, Haque N, Cani PD, Vos WM, Amar S, et al. Akkermansia muciniphila reduces porphyromonas gingivalis -induced inflammation and periodontal bone destruction. J Clin Periodontol. 2020;47(2):202–212. doi: 10.1111/jcpe.13214. [DOI] [PubMed] [Google Scholar]
- 179.Noguera-Julian M, Rocafort M, Guillén Y, Rivera J, Casadellà M, Nowak P, Hildebrand F, Zeller G, Parera M, Bellido R, et al. Gut microbiota linked to sexual preference and HIV infection. EBioMedicine. 2016;5:135–146. doi: 10.1016/j.ebiom.2016.01.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Rosas-Plaza S, Hernández-Terán A, Navarro-Díaz M, Escalante AE, Morales-Espinosa R, Cerritos R. Human gut microbiome across different lifestyles: from hunter-gatherers to urban populations. Front Microbiol. 2022;13:843170. doi: 10.3389/fmicb.2022.843170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Sadiq U, Shrestha U, Guzman N. Prevention of opportunistic infections in HIV/AIDS. In: StatPearls. Treasure Island (FL): StatPearls Publishing; 2022. May 23. [PubMed] [Google Scholar]
- 182.Dillon SM, Kibbie J, Lee EJ, Guo K, Santiago ML, Austin GL, Gianella S, Landay AL, Donovan AM, Frank DN, et al. Low abundance of colonic butyrate-producing bacteria in HIV infection is associated with microbial translocation and immune activation. Aids. 2017;31(4):511–521. doi: 10.1097/qad.0000000000001366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Rocafort M, Noguera-Julian M, Rivera J, Pastor L, Guillén Y, Langhorst J, Parera M, Mandomando I, Carrillo J, Urrea V, et al. Evolution of the gut microbiome following acute HIV-1 infection. Microbiome. 2019;7(1):73. doi: 10.1186/s40168-019-0687-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Estes J, Baker JV, Brenchley JM, Khoruts A, Barthold J, Bantle A, Reilly C, Beilman G, George M, Douek D, et al. Collagen deposition limits immune reconstitution in the gut. J Infect Dis. 2008;198(4):456–464. doi: 10.1086/590112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Mehandru S, Poles MA, Tenner-Racz K, Jean-Pierre P, Manuelli V, Lopez P, Shet A, Low A, Mohri H, Boden DMehandru S, Poles MA, Tenner-Racz K, et al . Lack of mucosal immune reconstitution during prolonged treatment of acute and early HIV-1 infection. PLoS Med. 2006;3(12):e484. doi: 10.1371/journal.pmed.0030484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Brenchley JM, Price DA, Schacker TW, Asher TE, Silvestri G, Rao S, Kazzaz Z, Bornstein E, Lambotte O, Altmann D, et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med. 2006;12(12):1365–1371. doi: 10.1038/nm1511. [DOI] [PubMed] [Google Scholar]
- 187.Jiang W, Lederman MM, Hunt P, Sieg S, Haley K, Rodriguez B, Landay A, Martin J, Sinclair E, Asher A, et al. Plasma levels of bacterial DNA correlate with immune activation and the magnitude of immune restoration in persons with antiretroviral-treated HIV infection. J Infect Dis. 2009;199(8):1177–1185. doi: 10.1086/597476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Baroncelli S, Galluzzo CM, Pirillo MF, Mancini MG, Weimer LE, Andreotti M, Amici R, Vella S, Giuliano M, Palmisano L, et al. Microbial translocation is associated with residual viral replication in HAART-treated HIV+ subjects with <50copies/ml HIV-1 RNA. J Clin Virol. 2009;46(4):367–370. doi: 10.1016/j.jcv.2009.09.011. [DOI] [PubMed] [Google Scholar]
- 189.Marchetti G, Bellistrì GM, Borghi E, Tincati C, Ferramosca S, La Francesca M, Morace G, Gori A, Monforte AD. Microbial translocation is associated with sustained failure in CD4+ T-cell reconstitution in HIV-infected patients on long-term highly active antiretroviral therapy. Aids. 2008;22(15):2035–2038. doi: 10.1097/QAD.0b013e3283112d29. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.