

Abstract
The introduction of new targeted therapies to the treatment algorithm of acute myeloid leukemia (AML) offers new opportunities, but also presents new challenges. Patients diagnosed with AML receiving targeted therapies as part of lower intensity regimens will relapse inevitably due to primary or secondary resistance mechanisms. In this review, we summarize the current knowledge on the main mechanisms of resistance to targeted therapies in AML. Resistance to FLT3 inhibitors is mainly mediated by on target mutations and dysregulation of downstream pathways. Switching the FLT3 inhibitor has a potential therapeutic benefit. During treatment with IDH inhibitors resistance can develop due to aberrant cell metabolism or secondary site IDH mutations. As a unique resistance mechanism the mutated IDH isotype may switch from IDH1 to IDH2 or vice versa. Resistance to gemtuzumab-ozogamicin is determined by the CD33 isotype and the degradation of the cytotoxin. The main mechanisms of resistance to venetoclax are the dysregulation of alternative pathways especially the upregulation of the BCL-2-analogues MCL-1 and BCL-XL or the induction of an aberrant cell metabolism. The introduction of therapies targeting immune processes will lead to new forms of therapy resistance. Knowing those mechanisms will help to develop strategies that can overcome resistance to treatment.
Keywords: Primary resistance, FLT3 inhibitors, IDH inhibitors, Gemtuzumab-ozogamicin, Venetoclax
Between 2017 and 2022, no less than ten new drugs have been approved by the FDA for AML-treatment, which has significantly changed the therapeutic landscape previously dominated by 7 + 3 chemotherapy and hypomethylating agents. Several of these drugs are directed against a specific target in the leukemic cell and allow an individualized therapeutic approach. However, with the emergence of new targeted agents, new resistance mechanisms have been found. The clinical impact of the development of resistance on patients’ outcome is currently unclear and no concepts are available yet whether and how patient monitoring and treatment management should be adapted to emerging treatment resistance.
In this review, we will summarize available evidence on resistance mechanisms to targeted therapies in AML and discuss potential consequences for clinical management of patients and for translational research to develop new strategies that may overcome drug resistance.
Resistance to FLT3 inhibitors
FLT3 (Fms related receptor tyrosine kinase 3) is a transmembrane kinase located in the cell membrane and the membrane of the endoplasmic reticulum. Binding of the FLT3 ligand (FL) to FLT3 on its extramembraneous binding site leads to the dimerization of FLT3 at the juxtamembrane domain. The phosphorylation of its juxtamembrane domain allows for the binding of substrates which are activated by phosphorylation of tyrosine [1–3]. The activation signal is passed on to the nucleus through several downstream signalling pathways such as RAS/MAPK, PI3K or JAK/STAT, thereby enhancing cell proliferation [2, 4]. FLT3-ITD mutations can be found in 25–35% of adult AML patients [5, 6]. A distinct prognostic impact of FLT3-ITD mutations has been described for low and high allelic burden depending on the NPM1 co-mutation [7, 8]. FLT3-TKD mutations on the other hand have no known prognostic impact on its own but offer a therapeutic target [9]. FLT3-TKD mutations can be found in about 7% of adult AML-patients [6, 10]. FLT3 inhibitors can be classified into type I and type II inhibitors. Type I inhibitors bind to the ATP-binding site, making them more specific for FLT3 and active for FLT3-ITD as well as TKD-mutations [11]. Type I inhibitors currently approved by the FDA are gilteritinib and midostaurin, while crenolanib is evaluated in clinical trials. The type I inhibitor midostaurin is approved for combination therapy with induction and consolidation chemotherapy in patients with FLT3-ITD or TKD mutations [12]. In the Ratify trial midostaurin prolonged survival independently of the mutational burden of FLT3 or the co-mutation of NPM1 [13]. Gilteritinib is used as monotherapy in relapsed or refractory FLT3-mutated AML patients, where it was shown to prolong survival compared to conventional chemotherapy [14].
Type II inhibitors bind to the hydrophobic area next to the ATP-binding site [15]. They are only effective with FLT3-ITD mutations but are also cross-inhibiting other receptor tyrosine kinases [15]. The most commonly used type II inhibitor is sorafenib. Sorafenib is used as maintenance treatment after allogeneic hematopoietic stem cell transplantation (alloHSCT) and prolongs survival in FLT3-mutated patients in this setting [16].
Quizartinib, another type II inhibitor, was recently shown to significantly prolong survival in patients with FLT3-ITD positive relapsed or refractory AML in comparison to placebo [17].
Resistance to type I inhibitors
The addition of midostaurin to conventional chemotherapy significantly improves survival of FLT3-mutated patients in all risk groups including older patients between 60 and 70 years of age [13, 18]. In the Ratify trial, it was found that about half of the patients lost the FLT3 mutation at the time of relapse and acquired mutations in alternative pathways such as the MAPK-pathway, 11% showed an expansion of new FLT3 clones [19]. The F691L gatekeeper mutation confers resistance to midostaurin [20]. The missense mutation N676K also induces resistance to midostaurin in patients [21]. Additionally, a subgroup analysis of the Ratify trial indicated that an insertion in the transmembrane kinase domain is associated with an unfavourable outcome even when adding midostaurin to chemotherapy [22].
Resistance to gilteritinib is mainly mediated by acquired mutations in FLT3 itself or new mutations in other oncogenes. McMahon et al. analysed samples of patients who relapsed after gilteritinib monotherapy and found mutations in the MAPK/RAS-pathway in one third of the patients [23]. Twelve percent of the patients developed a FLT3-F691L mutation but these two mechanisms never coincided [23].
Resistance to type II inhibitors
Missense mutations in FLT3 also play a potential role in resistance to type II inhibitors. The on-target mutation D835Y changes the conformation of the binding site, thereby conveying resistance to all type II inhibitors, whereas type I inhibitors with their different binding site are still effective [24]. Other mutations at amino acids F691, D835 and Y842 lead to decreased effectiveness of therapy with type II inhibitors [25–28] (Table 1).
Table 1.
Overview of on-target mutations conferring resistance to FLT3 inhibition
| FLT3 inhibitor | Mutations mediating resistance to FLT3 inhibitors | References |
|---|---|---|
| Crenolanib |
K429E (extramembrane domain) F691L (gatekeeper residue) |
[29, 30] |
| Midostaurin |
F691L (gatekeeper residue) N676K (tyrosine kinase domain I) |
[19, 21] |
| Gilteritinib | F691L (gatekeeper residue) | [23, 31–33] |
| Quizartinib |
D835Y/V/F (activation loop) Y842C/H (activation loop) F691L (gatekeeper residue) |
[34, 35] |
| Sorafenib |
D835Y/V (activation loop) Y842C/H (activation loop) F691L (gatekeeper residue) |
[20, 25, 28, 36, 37] |
The activation loop includes sites for phosphorylation, while the gatekeeper residue controls binding of small molecules to the kinase. Most notably, the F691L mutation in the gatekeeper residue of FLT3 leads to resistance against all FLT3 inhibitors used in clinical practice
In contrast to type I inhibitors, acquired mutations in the MAPK/RAS-pathway rarely confer resistance to type II inhibitors, as the latter cross-inhibit several other components of the MAPK/RAS pathway. Table 1 gives an overview of the most common missense mutations that mediate resistance to FLT3 inhibitors.
Common resistance mechanisms
One main problem in AML treatment is the constant clonal evolution. Frequently, the blasts at relapse bear a different molecular pattern than at diagnosis. Loss of FLT3 mutations has been observed even without the selection pressure exerted by the use of FLT3 inhibitors [38, 39]. Additionally, primary resistance can be observed in approximately 30% of patients [40]. It is important to note that FLT3 mutated cells are usually mutated in one allele only and therefore may express unmutated FLT3 as well. Because the FL primarily binds to wildtype FLT3, it activates downstream pathways the stronger the more wildtype FLT3 is expressed [41, 42]. Although this mechanism does not cause complete resistance, it can be an explanation that depending on the FLT3 wt/mut ratio the effect of FLT3 inhibitors might differ, as the activation of downstream pathways via FL is more effective. Additionally, the level of FL increases in chemotherapy induced aplasia and a rising FL expression has been associated with a worse outcome [42, 43].
Soluble factors from stromal cells that activate downstream pathways of FLT3 are able to overcome the effect of FLT3 inhibitors. For example, FGF2 is able to activate the RAS/MAPK pathway independently of FLT3 in vitro and increased levels of FGF2 can be found in patients who relapsed under treatment with FLT3 inhibitors [44]. Although this effect has only been examined regarding resistance to quizartinib, it may be an universal mechanism. Similarly, upregulation of the chemokine CXCL12, which is secreted into the bone marrow stroma by osteoblasts, is able to induce proliferation independently of FLT3 in vitro and in mouse models [45, 46]. Interestingly, the acquisition of different JAK2-mutations was associated with FLT3 inhibitor-resistance and the non-canonical JAK2 V658F mutation was able to activate CSF2RB and circumvent FLT3-dependency in vitro [47]. Upregulation of the CCL5-CCR5-pathway, which is known to play a role in therapy resistance in other malignant diseases as well [48], also mediated FLT3 inhibitor resistance as downstream pathways such as RAS, AKT and STAT5 are upregulated independently of FLT3 activation [49]. Upregulation of STAT5 stands out as a key element of FLT3 inhibitor-resistance. It upregulates PIM1 independently of FLT3 and thereby circumvents FLT3-dependency [50]. Furthermore, STAT5 upregulates AXL-expression in vitro in primary AML-blasts via activation from cytokines from the bone marrow environment like TPO or GM-CSF, which conveys FLT3 inhibitor-resistance by activating PI3K and RAS/RAF [51, 52]. However, AXL can also be activated by hypoxia via HIF-1α independently of STAT5 [52]. As gilteritinib inhibits AXL as well as FLT3, it may still be effective in patients after activation of AXL via the STAT5-pathway or HIF-1α [31]. Although the understanding of these pathway modifications is still restricted to in vitro and mouse models, they may also be important in patients.
An increased intracellular pH is known to enhance cell proliferation and reduce apoptosis. An increase in pH via activation of the Na/H+ ion channel via tescalcin (TESC) overexpression leads to FLT3 inhibitor resistance in vitro in cell lines as well as in primary AML cells [53].
FLT3 inhibitors are metabolized by CYP3A4. Therefore, overexpression of this enzyme leads to increased drug degradation [54]. Similarly, high expression levels of the P-glycoprotein Pgp efflux pumps decrease the effectiveness of FLT3 inhibitor-therapy [55]. The efficacy of FLT3 inhibitors can also be reduced by increased binding to plasma proteins at least in vitro. The only FLT3 inhibitor which does not seem to be affected by binding to plasma proteins is gilteritinib [56].
The currently known resistance mechanisms against FLT3 inhibitors are summarized in Fig. 1.
Fig. 1.

Overview of resistance mechanisms to FLT3 inhibitors. 1: On-target mutations; 2: Activation of downstream pathways; 3: Increased drug elimination; 4: Upregulation of soluble factors to activate downstream pathways; 5: Activation of AXL-pathways; 6: Increase in intracellular pH 7: Upregulation of FLT3 ligand (FL). (Created with BioRender.com) C crenolanib, G gilteritinib, M midostaurin, Q quizartinib, S sorafenib, TESC tescalcin, FLT3i Flt3 inhibitor
As more and more patients receive FLT3 inhibitors as frontline therapy, it is important to address the question of efficacy of sequential application of different FLT3 inhibitors. A retrospective analysis of patients relapsing after front line therapy with FLT3 inhibitor-based regimens compared the rates of CR and CRi and highlighted that with each subsequent use of a new FLT3 inhibitor the percentage of patients responding decreased [57]. One obvious reason may be the selection of FLT3-inhibitor-resistant clones independently of the exact binding site of the FLT3 inhibitor such as upregulation of downstream pathways. However, with over 30% achieving CRc with the second FLT3 inhibitor and 25% with the third, the clinical effect is still relevant and may be increased by using combination therapies [57].
Resistance to IDH inhibitors
Isocitrate dehydrogenase 1 and 2 (IDH1 and IDH2) are vital for the energy metabolism and biosynthesis in the cell as they catalyse the synthesis of α-ketoglutarate from isocitrate as part of the Krebs-Cycle. Mutations in either of these enzymes lead to increased production of R-2-hydroxyglutarate, which inhibits cellular differentiation [58, 59]. In the context of AML, mutations of IDH1 are found in 6–7% of AML patients while IDH2 mutations are found in approximately 15% of AML patients [60]. Ivosidenib as IDH1 inhibitor and enasidenib as IDH2 inhibitor are approved by the FDA for monotherapy in patients with relapsed/refractory AML and a proven mutation in the respective enzyme inducing CR in approximately 20% of patients [61, 62]. In 2019, the approval of ivosidenib was broadened to allow first line therapy as a single agent for unfit patients with IDH1-mutation. For the IDH1 inhibitor ivosidenib the combination with azacitidine has been shown to be superior to azacitidine monotherapy, inducing CR in 47% vs. 15% in newly diagnosed AML patients, which led to approval by the FDA in May 2022 [63].
The common mutation sites in IDH1 are R132 [64, 65] and in IDH2 R140 and R172 [58, 66].
Regarding primary resistance, Wang et al. described that cells with a stem cell-like gene expression profile show a poor response to treatment with IDH inhibitors [67]. This phenotype may be at least partly induced by downregulation of TET2 leading to DNA hypermethylation, which is characteristic for leukemic stem cells (LSC) [68]. Lower response rates to treatment with enasidenib have been associated with NRAS mutations [69]. It was also highlighted that at relapse after treatment with IDH inhibitors dominant clones within the blast population were likely to show mutations in RUNX1 or NRAS as well as FLT3 [69, 70]. It was pointed out as well that one mutation alone may not be sufficient to induce therapy resistance [71].
Newly acquired missense mutations within IDH1 or IDH2 are another central resistance mechanism against IDH1/2 inhibitors. Intlekofer et al. analysed two patients who relapsed under treatment with IDH2 inhibitors by acquiring additional mutations. Of note, the newly acquired mutations in these cases were located on the allele which did not carry the initial R140Q mutation. Both Q316E and I319M mutations inhibit the binding of enasidenib and are pathogenic only in combination with the R140Q mutation. In vivo these mutations have only been identified on the allele not bearing the initial IDH2 mutation but in vitro experiments hint that the acquisition of these mutations on the same allele would also confer resistance [72]. For IDH1 inhibitors the additional acquisition of a S280F or H315D mutation leads to therapy resistance due to changes to the substrate or NADPH binding site [71]. R119P, G131A, D279N and G289D mutations prevent binding due to steric interference [71].
A unique mechanism of resistance to IDH inhibitors is the switching of the mutated IDH isotype, e.g. from IDH1 to IDH2 or vice versa. This new isotype is either a completely new clone or the expansion of a pre-existing clone [73]. The use of IDH inhibitors, which are able to inhibit both IDH1 and IDH2 may be a strategy to overcome resistance mediated by this isotype switch. Co-targeting agents currently being investigated in AML and glioma are vorasidenib (AG-881) [74] and Ly3410738 [75].
Given its prominent role in energy metabolism, it is not surprising that therapy resistance to IDH inhibitors has been linked to changes in energy metabolism. R-2-hydroxyglutarate leads to an increase in fatty acid oxidation and oxidative phosphorylation via CEBPα. Interestingly, treatment with IDH inhibitors does not diminish fatty acid oxidation (FAO) as it is maintained via PGC1a and its associated pathways in vitro [76]. Upon relapse under IDH inhibitor treatment patients exhibit genetic changes associated with high activation levels of oxidative phosphorylation, which may be a resistance mechanism independent of acquired mutations in the IDH enzymes [76].
One way to improve the response to IDH inhibition may be the combination with hypomethylating agents or standard chemotherapy. In the AGILE-study the combination of ivosidenib with azacitidine induced responses in patients who harboured co-mutations in NRAS, KRAS and PTPN11, who reportedly do not respond as well to ivosidenib monotherapy [77].
Based on our current understanding resistance to IDH inhibition is thus conveyed by on-target mutations, isotype switching and changes in cell metabolism (Fig. 2).
Fig. 2.

Overview of resistance mechanisms to IDH inhibitors. Treatment with IDH inhibitors mainly blocks the synthesis of the oncometabolite R-2-hydroxyglutarate, which inhibits hypermethylation and induces differentiation of the cell. The upregulation of fatty acid oxidation is not inhibited. The main resistance mechanisms are 1: Mutations at binding site; 2: Isotype switch; 3: Aberrant regulation of cell metabolism. (Created with BioRender.com)
Resistance to gemutzumab-ozogamicin (GO)
Gemtuzumab-ozogamicin is a drug-antibody conjugate consisting of an anti-CD33 antibody linked with the cytotoxic drug calicheamicin. Upon internalisation calicheamicin leads to DNA-damage and subsequent apoptosis [78]. It is approved for induction therapy in patients with favourable or intermediate risk CD33 + AML in combination with conventional chemotherapy [79].
Primary resistance to GO is not associated with CD33-expression levels but rather polymorphisms in the gene encoding CD33. This mechanism of primary resistance was firstly described by Lamba et al. in a cohort of pediatric patients treated with standard chemotherapy and GO. The consequence of the polymorphism rs12459419 is an exchange of alanin to valin (C to T) in CD33 [80, 81]. This change leads to a different configuration of the antibody binding-site in CD33. All patients who were homozygous for this polymorphism showed lower response rates than those who were heterozygous [81]. In a larger cohort of patients the mean incidence of each genotype was 51% for CC, 39% for CT and 10% for TT [80]. However, the predictive impact of this polymorphism was variable in different cohorts. Two studies reported no difference in adult cohorts [82, 83], whereas in patients with NPM1 mutations the polymorphism predicted therapy response [84]. Another polymorphism of CD33, rs35112940, leads to a conformational change in the immunoreception tyrosine-based inhibitory motif. This form of CD33 is less likely to be internalized, thus reducing the cytotoxic effect of GO [85]. However, the prognostic effect of this polymorphism is unknown. So far, it has not been evaluated prospectively, whether CD33 polymorphisms can be used as a predictive marker for GO treatment.
The polymorphism rs1045642 in the gene ABCB1 encoding P-glycoprotein (Pgp) (C to T) also has predictive value for response to GO. This polymorphism has been associated with inferior response rates to GO in patients who are homozygous for CC [86].
Another mechanism of therapy resistance is associated with an MDR-phenotype and increased expression of Pgp. Early work by Walter et al. analysed the outcome of patients treated with GO monotherapy after AML relapse. Higher expression levels of ABCB1 correlated with a poor response to treatment with GO [87]. However, it is unclear whether this is a mechanism of primary resistance or if ABCB1 expression is upregulated in response to treatment. Furthermore, this mechanism is not specific for response to GO, but rather shows that ABCB1 expression is a marker for poor response to chemotherapy in general.
In addition to increased drug efflux, leukemic cells are able to increase CD33 degradation. The vital protein in this mechanism is SOCS3, which binds to phosphorylated CD33 and leads to internalisation and degradation of CD33 [88, 89]. Low levels of promoter methylation of SOCS3 and therefore increased expression levels correlated with lower response rates to GO treatment as mono- or combination therapy [90].
Acquired resistance to CD33 is mainly conveyed by upregulation of downstream pathways. Rosen et al. could show that the cytotoxicity by calicheamicin is not sufficient to induce apoptosis. GO resistant cell lines showed upregulation of the important pro-proliferative PI3K pathway and the application of an AKT inhibitor was able to overcome acquired GO resistance in vitro [91]. GO resistance is therefore mainly caused by polymorphisms of the binding site, increased degradation of CD33 and activation of pro-proliferative pathways (Fig. 3).
Fig. 3.
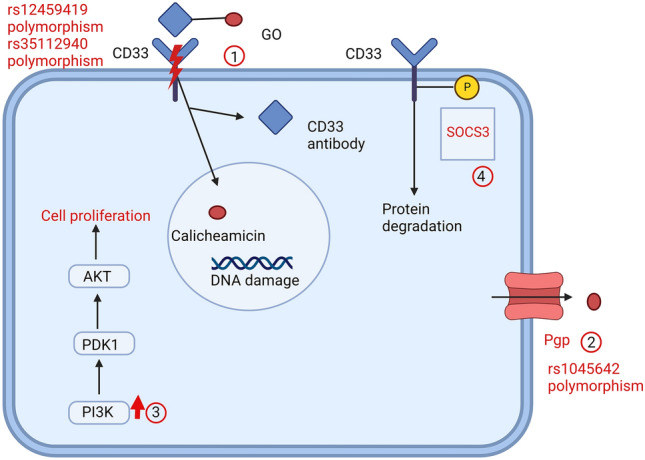
Overview of resistance mechanisms to GO. GO binds to CD33 and is internalized. Inside the cell, calicheamicin dissociates from the CD33-antibody and induces DNA-damage. Main mechanisms of resistance: 1: CD33 polymorphisms; 2: Increased drug degradation; 3: Upregulation of downstream pathways; 4: Phosphorylation of CD33 by SOCS3. (Created with BioRender.com)
Resistance to Venetoclax
Venetoclax is a BH3 mimetic, which acts as a BCL-2 inhibitor and induces apoptosis. It is widely used as first and second line therapy for CLL, and in combination therapies in AML. The combination of HMA with venetoclax in patients unfit for conventional chemotherapy has significantly improved the outcome of these patients [92].
However, 34% of newly diagnosed AML patients do not respond to the combination of venetoclax with azacitidine and all patients with an initial response will eventually relapse [92].
A resistance mechanism which is commonly found in CLL patients treated with venetoclax is a missense mutation G101V in BCL-2. It modifies the binding site of venetoclax [93]. However, this mechanism has so far not been observed in AML patients.
Retrospective analyses of patients who participated in the registration trials identified molecular markers that are associated with response to venetoclax.
Better response rates were seen in patients with NPM1 or IDH2 mutations, whereas at relapse selection of FLT3-mutated clones could be observed [94]. A deeper analysis showed that relapse in patients could not be pinpointed to a single mechanism, but rather the activation of several different pathways like kinase signalling, RNA-splicing, epigenetic modification, transcription and tumour suppression [95]. It is unclear whether the emerging aberrations are induced by the treatment or are selected from a pre-existing clone due to the selection pressure under treatment.
One central causal mechanism of acquired resistance to venetoclax treatment is the circumvention of pro-apoptotic signalling via BH3-mimetics. Several studies could show that BCL-XL and MCL-1 as homologues of BCL-2 can fulfil the same anti-apoptotic role as BCL-2, but are not inhibited by venetoclax [96–98]. Accordingly, blasts showing a monocytic phenotype were resistant to venetoclax as they—in analogy to healthy monocytes—depend on MCL-1 instead of BCL-2. Therefore, at relapse there is often a dominant monocytic clone [99]. Based on these findings, the in vitro use of MCL-1 inhibitors is able to overcome resistance to venetoclax [100, 101]. Other components of apoptosis may be associated with venetoclax resistance such as TP53, BAX, BAK, PMAIP1 and TFDP1 [102].
When looking at venetoclax resistance, it is important to note that the molecule does not only inhibit BCL-2, but the combination with hypomethylating agents also leads to a decrease in amino acid uptake that diminishes the cellular energy source [103, 104]. To overcome this inhibition leukemic cells activate fatty acid metabolism by upregulating several genes of this pathway [105]. Another way to recruit additional energy sources is an increased production of NADP + [106]. Thus, upregulating alternative pathways of energy resources seems to be a potential mechanism for acquired resistance to venetoclax [105, 106].
In summary, primary resistance to venetoclax is associated with variable, incompletely understood changes of cellular biology with high inter-patient variability. Acquired resistance to venetoclax mainly emerges by circumventing the pathways, which are inhibited by venetoclax, namely BCL-2 and amino acid metabolism (Fig. 4).
Fig. 4.

Overview of resistance mechanisms against venetoclax. Venetoclax is a BH3 mimetic, which inhibits BCL-2, an inhibitor of apoptosis. Common resistance mechanisms are: 1: Upregulation of MCL-1 or BCL-XL; 2: Upregulation of fatty acid oxidation; 3: Upregulation of nicotinamide leading to increased NADP + levels. (Created with BioRender.com)
Outlook: resistance to immunotherapy
In contrast to lymphoid malignancies, immunotherapy in AML is still facing many challenges. The bone marrow niche of AML provides an immunosuppressive environment protecting the leukemic cell from immune attack. For example, regulatory T-cells and myeloid derived suppressor cells are increased in the bone marrow niche [107, 108]. However, pathways involving the immune response appear to be vital for pathogenesis of AML and a valuable therapeutic target. Relapse after stem cell transplantation is in most cases not associated with new mutations in proliferative pathways, but with changes to the immune response. Patients with relapse after alloHSCT may show a downregulation of HLA class II expression and therefore fail to properly present antigens [109]. The importance of the immune system and its therapeutic potential is also underscored by the role of the graft-versus-leukemia-effect and the ability of donor lymphocyte infusions (DLI) to boost the graft vs leukemia effect as an important pillar of treating post-transplant relapses. Based on this central role of the immune response in relapse after alloHSCT, the application of ipilimumab in patients with relapse after alloHSCT activated T-cells and led to remission in some patients [110]. Other checkpoint inhibitors such as nivolumab and pembrolizumab are evaluated in combination with conventional chemotherapy [111, 112]. However, all these approaches have to overcome the immunosuppressive bone marrow environment and—for patients after transplantation—the higher risk of GvHD.
A novel mechanism targeting the immune system is the activation of macrophages. Several CD47-antibodies are being evaluated. CD47 is overexpressed in many cancer entities and acts as a signal to prevent phagocytosis by macrophages [113]. In a phase Ib study, the combination of the CD47-antibody magrolimab with azacitidine was able to induce a response in 64% of the patients, reaching CR/CRi in about 50% of the patients with similar response rates in patients with or without TP53 mutations [114]. Based on these promising results magrolimab is currently being evaluated in several clinical studies in AML patients.
Although resistance mechanisms to magrolimab have not been described so far, based on its mechanism of action one could consider several ways of resistance. One regulatory point may be the expression of CD47 on the cell surface. CD47 expression is regulated by a number of cytokines and pro-inflammatory mediators on hematopoietic stem cells (HSC), thus enabling migration from the bone marrow to the site of inflammation [115]. This mechanism may also apply to LSCs, as signals from the bone marrow niche may be able to considerably downregulate CD47 expression. CD47 can be located on the cell surface or in the membrane of the endoplasmic reticulum (ER). The localisation is determined by the length of the 3’ UTR: mRNA with a long 3’UTR encodes for CD47 located in the cell membrane, so the amount of CD47 on the cell surface can be regulated at a posttranscriptional level [116].
Binding between CD47 and the SIRPα-receptor on the macrophages is only possible, when there is polyglutamate added at the binding site via the glutaminyl-peptide cyclotransferase like protein [117]. Thus, any interference with binding of the antibody to its target may induce resistance.
Conclusion
Resistance to targeted therapies is either primary or secondary due to acquired mutations under the selective pressure of the treatment. Although the targets are different, there are some common mechanisms to resistance: On the one hand, there may be mutations at the binding site of the target protein; on the other hand, alternative pathways may be activated to allow cell proliferation and survival. Knowledge of common resistance mechanisms can help to develop strategies that may prevent resistance. For example, the activation of the MAPK-pathway is a common finding. However, currently available MEK inhibitors have limited activity in AML patients and are associated with considerable hematologic toxicity [118–120]. MCL-1- or BCL-XL-inhibition as an add-on to BCL-2-inhibition may be an option to overcome venetoclax resistance. On the other hand, added toxicity may be of concern and requires careful evaluation. Table 2 summarizes current knowledge on possible ways to overcome resistance. Most findings, however, are still limited to in vitro or mouse experiments. Clinical data on sequential therapies is scarce.
Table 2.
Known therapeutic strategies to overcome resistance to targeted therapies. Mainly evidence is restricted to findings in vitro or mice
| Mutation | Resistant drug | Theoretical treatment option | Clinical experience | References |
|---|---|---|---|---|
| Flt3 on-target mutations | FLT3 inhibitor |
Sequential use of FLT3i F691L confers resistance to all known FLT3i |
Exposure to another FLT3i after midostaurin induced CR in 29%, FLT3i after quizartinib induced CR in 20%; quizartinib after sorafenib induced CR in 25%, no experience with second FLT3i after gilteritinib | [57] |
| Upregulation of soluble downstream factors | FLT3 inhibitor |
FGF2 inhibitor CXCR4 inhibitor |
In vitro evidence only for FGF2 inhibition CXCR4 inhibitor AMD3465 in combination with cytarabine eliminated leukemic blasts in mice |
[44, 46] |
| Upregulation of downstream-pathways | FLT3 inhibitor |
AXL inhibitors/ gilteritinib STAT5 inhibitors PIM inhibitor JAK inhibitor |
The AXL inhibitor TP-0903 showed prolonged survival in combination with FLT3 inhibition in mice In vitro evidence only for PIM and JAK inhibitors |
[31, 47, 50–52] |
| Increased intracellular pH | FLT3 inhibitor | NHE1 inhibitor | In vitro evidence and engraftment experiments in mice only | [53] |
| Upregulation of FLT3-ligand | Flt3 inhibitor | Flt3 inhibitor with higher plasma levels, higher affinity to Flt3 | Not known | [42] |
| Binding site polymorphism | GO | Not known | ||
| Increased drug degradation | GO | Not known | ||
| Upregulation of PI3K-pathways | GO | AKT-inhibitor | In vitro evidence only | [91] |
| Mutation at binding site | IDH1/2 inhibitors | Not known | ||
| Isotype switch | IDH1/2 inhibitors |
Ivosidenib after enasidenib; Enasidenib after ivosidenib; Bivalent inhibitors e.g. Ly3410738 |
Clinical studies ongoing | [74, 75] |
| Upregulation of downstream pathways | IDH1/2 inhibitors | RAS inhibitor | Not known | [69] |
| Alternative energy sources | IDH1/2 inhibitors | OXPHOS inhibitor | Effect of combined IDH and OXPHOS inhibition in mice | [76] |
| Increased expression of MCL-1 or BCL-XL | Venetoclax | MCL-1 or BCL-XL inhibitors | In vitro evidence only | [101] |
| Utilization of alternative energy sources such as fatty acid metabolism/NADP + | Venetoclax | Inhibition of FAO | In vitro evidence only | [105] |
By using targeted therapies, resistant clones will have a proliferative advantage over sensitive clones and inhibiting one clone may not prevent other clones from expansion. Therefore, the sequential use of therapeutics that target different pathways may be an approach to overcome resistance. In addition, the dysregulation of the immune system offers a novel therapeutic area, which needs to be explored in much more detail to derive benefit of immunotherapy for AML patients.
Author contributions
RM and MH searched the literature, wrote the manuscript and designed the figures.
Funding
This work was supported by grant 70114478 from Deutsche Krebshilfe and DJCLS grant 16 R/2021. Open Access funding enabled and organized by Projekt DEAL.
Declarations
Competing interests
Michael Heuser has to declare the following conflicts of interest: Honoraria: Abbvie, Eurocept, Jazz Pharmaceuticals, Janssen, Novartis, Takeda Consultancy: Abbvie, Agios, BMS, Daiichi Sankyo, Glycostem, Jazz Pharmaceuticals, Kura Oncology, Novartis, Pfizer, PinotBio, Roche, Tolremo Research funding for institution: Abbvie, Agios, Astellas, Bayer Pharma AG, BergenBio, Daiichi Sankyo, Glycostem, Jazz Pharmaceuticals, Loxo Oncology, Novartis, Pfizer, PinotBio, Roche Rabea Mecklenbrauck has no conflicts of interest to declare.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Grafone T, Palmisano M, Nicci C, et al. An overview on the role of FLT3-tyrosine kinase receptor in acute myeloid leukemia: biology and treatment. Oncol Rev. 2012;6:e8. doi: 10.4081/oncol.2012.e8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hayakawa F, Towatari M, Kiyoi H, et al. Tandem-duplicated Flt3 constitutively activates STAT5 and MAP kinase and introduces autonomous cell growth in IL-3-dependent cell lines. Oncogene. 2000;19:624–631. doi: 10.1038/sj.onc.1203354. [DOI] [PubMed] [Google Scholar]
- 3.Griffith J, Black J, Faerman C, et al. The structural basis for autoinhibition of FLT3 by the juxtamembrane domain. Mol Cell. 2004;13:169–178. doi: 10.1016/s1097-2765(03)00505-7. [DOI] [PubMed] [Google Scholar]
- 4.Mizuki M, Fenski R, Halfter H, et al. Flt3 mutations from patients with acute myeloid leukemia induce transformation of 32D cells mediated by the Ras and STAT5 pathways. Blood. 2000;96:3907–3914. doi: 10.1182/blood.V96.12.3907. [DOI] [PubMed] [Google Scholar]
- 5.Kottaridis PD, Gale RE, Frew ME, et al. The presence of a FLT3 internal tandem duplication in patients with acute myeloid leukemia (AML) adds important prognostic information to cytogenetic risk group and response to the first cycle of chemotherapy: analysis of 854 patients from the United Kingdom Medical Research Council AML 10 and 12 trials. Blood. 2001;98:1752–1759. doi: 10.1182/blood.v98.6.1752. [DOI] [PubMed] [Google Scholar]
- 6.Thiede C, Steudel C, Mohr B, et al. Analysis of FLT3-activating mutations in 979 patients with acute myelogenous leukemia: association with FAB subtypes and identification of subgroups with poor prognosis. Blood. 2002;99:4326–4335. doi: 10.1182/blood.v99.12.4326. [DOI] [PubMed] [Google Scholar]
- 7.Döhner H, Estey E, Grimwade D, et al. Diagnosis and management of AML in adults: 2017 ELN recommendations from an international expert panel. Blood. 2017;129:424–447. doi: 10.1182/blood-2016-08-733196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gale RE, Green C, Allen C, et al. The impact of FLT3 internal tandem duplication mutant level, number, size, and interaction with NPM1 mutations in a large cohort of young adult patients with acute myeloid leukemia. Blood. 2008;111:2776–2784. doi: 10.1182/blood-2007-08-109090. [DOI] [PubMed] [Google Scholar]
- 9.Papaemmanuil E, Gerstung M, Bullinger L, et al. Genomic classification and prognosis in acute myeloid leukemia. N Engl J Med. 2016;374:2209–2221. doi: 10.1056/NEJMoa1516192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yamamoto Y, Kiyoi H, Nakano Y, et al. Activating mutation of D835 within the activation loop of FLT3 in human hematologic malignancies. Blood. 2001;97:2434–2439. doi: 10.1182/blood.v97.8.2434. [DOI] [PubMed] [Google Scholar]
- 11.Short NJ, Kantarjian H, Ravandi F, et al. Emerging treatment paradigms with FLT3 inhibitors in acute myeloid leukemia. Ther Adv Hematol. 2019;10:2040620719827310. doi: 10.1177/2040620719827310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Stone RM, Mandrekar SJ, Sanford BL, et al. Midostaurin plus chemotherapy for acute myeloid leukemia with a FLT3 mutation. N Engl J Med. 2017;377:454–464. doi: 10.1056/NEJMoa1614359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Döhner K, Thiede C, Jahn N, et al. Impact of NPM1/FLT3-ITD genotypes defined by the 2017 European LeukemiaNet in patients with acute myeloid leukemia. Blood. 2020;135:371–380. doi: 10.1182/blood.2019002697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Perl AE, Martinelli G, Cortes JE, et al. Gilteritinib or chemotherapy for relapsed or refractory FLT3-mutated AML. N Engl J Med. 2019;381:1728–1740. doi: 10.1056/NEJMoa1902688. [DOI] [PubMed] [Google Scholar]
- 15.Larrosa-Garcia M, Baer MR. FLT3 inhibitors in acute myeloid leukemia: current status and future directions. Mol Cancer Ther. 2017;16:991–1001. doi: 10.1158/1535-7163.MCT-16-0876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Burchert A, Bug G, Fritz LV, et al. Sorafenib maintenance after allogeneic hematopoietic stem cell transplantation for acute myeloid leukemia with FLT3-internal tandem duplication mutation (SORMAIN) J Clin Oncol. 2020;38:2993–3002. doi: 10.1200/JCO.19.03345. [DOI] [PubMed] [Google Scholar]
- 17.Cortes JE, Khaled S, Martinelli G, et al. Quizartinib versus salvage chemotherapy in relapsed or refractory FLT3-ITD acute myeloid leukaemia (QuANTUM-R): a multicentre, randomised, controlled, open-label, phase 3 trial. Lancet Oncol. 2019;20:984–997. doi: 10.1016/S1470-2045(19)30150-0. [DOI] [PubMed] [Google Scholar]
- 18.Döhner Hea Midostaurin Plus Intensive Chemotherapy for Younger and Older Patients with Acute Myeloid Leukemia and FLT3 Internal Tandem Duplications [DOI] [PMC free article] [PubMed]
- 19.Schmalbrock LK, Dolnik A, Cocciardi S, et al. Clonal evolution of acute myeloid leukemia with FLT3-ITD mutation under treatment with midostaurin. Blood. 2021;137:3093–3104. doi: 10.1182/blood.2020007626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Williams AB, Nguyen B, Li L, et al. Mutations of FLT3/ITD confer resistance to multiple tyrosine kinase inhibitors. Leukemia. 2013;27:48–55. doi: 10.1038/leu.2012.191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Heidel F, Solem FK, Breitenbuecher F, et al. Clinical resistance to the kinase inhibitor PKC412 in acute myeloid leukemia by mutation of Asn-676 in the FLT3 tyrosine kinase domain. Blood. 2006;107:293–300. doi: 10.1182/blood-2005-06-2469. [DOI] [PubMed] [Google Scholar]
- 22.Rücker FG, Du L, Luck TJ, et al. Molecular landscape and prognostic impact of FLT3-ITD insertion site in acute myeloid leukemia: RATIFY study results. Leukemia. 2022;36:90–99. doi: 10.1038/s41375-021-01323-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.McMahon CM, Ferng T, Canaani J, et al. Clonal selection with RAS pathway activation mediates secondary clinical resistance to selective FLT3 inhibition in acute myeloid leukemia. Cancer Discov. 2019;9:1050–1063. doi: 10.1158/2159-8290.CD-18-1453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Smith CC, Lin K, Stecula A, et al. FLT3 D835 mutations confer differential resistance to type II FLT3 inhibitors. Leukemia. 2015;29:2390–2392. doi: 10.1038/leu.2015.165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zhang W, Konopleva M, Shi Y, et al. Mutant FLT3: a direct target of sorafenib in acute myelogenous leukemia. J Natl Cancer Inst. 2008;100:184–198. doi: 10.1093/jnci/djm328. [DOI] [PubMed] [Google Scholar]
- 26.von Bubnoff N, Engh RA, Aberg E, et al. FMS-like tyrosine kinase 3-internal tandem duplication tyrosine kinase inhibitors display a nonoverlapping profile of resistance mutations in vitro. Cancer Res. 2009;69:3032–3041. doi: 10.1158/0008-5472.CAN-08-2923. [DOI] [PubMed] [Google Scholar]
- 27.Scholl S, Fleischmann M, Schnetzke U, et al. Molecular mechanisms of resistance to FLT3 inhibitors in acute myeloid leukemia: ongoing challenges and future treatments. Cells. 2020 doi: 10.3390/cells9112493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Baker SD, Zimmerman EI, Wang Y-D, et al. Emergence of polyclonal FLT3 tyrosine kinase domain mutations during sequential therapy with sorafenib and sunitinib in FLT3-ITD-positive acute myeloid leukemia. Clin Cancer Res. 2013;19:5758–5768. doi: 10.1158/1078-0432.CCR-13-1323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhang H, Savage S, Schultz AR, et al. Clinical resistance to crenolanib in acute myeloid leukemia due to diverse molecular mechanisms. Nat Commun. 2019;10:244. doi: 10.1038/s41467-018-08263-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Smith CC, Lasater EA, Zhu X, et al. Activity of ponatinib against clinically-relevant AC220-resistant kinase domain mutants of FLT3-ITD. Blood. 2013;121:3165–3171. doi: 10.1182/blood-2012-07-442871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lee LY, Hernandez D, Rajkhowa T, et al. Preclinical studies of gilteritinib, a next-generation FLT3 inhibitor. Blood. 2017;129:257–260. doi: 10.1182/blood-2016-10-745133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Tarver TC, Hill JE, Rahmat L, et al. Gilteritinib is a clinically active FLT3 inhibitor with broad activity against FLT3 kinase domain mutations. Blood Adv. 2020;4:514–524. doi: 10.1182/bloodadvances.2019000919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sharzehi S, Joshi SK, Pittsenbarger J, et al. The FLT3 F691L gatekeeper mutation promotes clinical resistance to Gilteritinib + Venetoclax (GILT + VEN) in AML. Blood. 2021;138:2235. doi: 10.1182/blood-2021-145762. [DOI] [Google Scholar]
- 34.Smith CC, Wang Q, Chin C-S, et al. Validation of ITD mutations in FLT3 as a therapeutic target in human acute myeloid leukaemia. Nature. 2012;485:260–263. doi: 10.1038/nature11016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Smith CC, Paguirigan A, Jeschke GR, et al. Heterogeneous resistance to quizartinib in acute myeloid leukemia revealed by single-cell analysis. Blood. 2017;130:48–58. doi: 10.1182/blood-2016-04-711820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wang Z, Hu B, An Y, et al. Exploring the resistance mechanisms of distal D835V mutation in FLT3 to inhibitors. Oxid Med Cell Longev. 2022;2022:3720026. doi: 10.1155/2022/3720026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Albers C, Leischner H, Verbeek M, et al. The secondary FLT3-ITD F691L mutation induces resistance to AC220 in FLT3-ITD+ AML but retains in vitro sensitivity to PKC412 and Sunitinib. Leukemia. 2013;27:1416–1418. doi: 10.1038/leu.2013.14. [DOI] [PubMed] [Google Scholar]
- 38.Cloos J, Goemans BF, Hess CJ, et al. Stability and prognostic influence of FLT3 mutations in paired initial and relapsed AML samples. Leukemia. 2006;20:1217–1220. doi: 10.1038/sj.leu.2404246. [DOI] [PubMed] [Google Scholar]
- 39.Shih L-Y, Huang C-F, Wu J-H, et al. Internal tandem duplication of FLT3 in relapsed acute myeloid leukemia: a comparative analysis of bone marrow samples from 108 adult patients at diagnosis and relapse. Blood. 2002;100:2387–2392. doi: 10.1182/blood-2002-01-0195. [DOI] [PubMed] [Google Scholar]
- 40.Stone RM, DeAngelo DJ, Klimek V, et al. Patients with acute myeloid leukemia and an activating mutation in FLT3 respond to a small-molecule FLT3 tyrosine kinase inhibitor, PKC412. Blood. 2005;105:54–60. doi: 10.1182/blood-2004-03-0891. [DOI] [PubMed] [Google Scholar]
- 41.Chen F, Ishikawa Y, Akashi A, et al. Co-expression of wild-type FLT3 attenuates the inhibitory effect of FLT3 inhibitor on FLT3 mutated leukemia cells. Oncotarget. 2016;7:47018–47032. doi: 10.18632/oncotarget.10147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sato T, Yang X, Knapper S, et al. FLT3 ligand impedes the efficacy of FLT3 inhibitors in vitro and in vivo. Blood. 2011;117:3286–3293. doi: 10.1182/blood-2010-01-266742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Haidar JH, Bazarbachi A, Mahfouz R, et al. Serum Flt3 ligand variation as a predictive indicator of hematopoietic stem cell mobilization. J Hematother Stem Cell Res. 2002;11:533–538. doi: 10.1089/15258160260090997. [DOI] [PubMed] [Google Scholar]
- 44.Traer E, Martinez J, Javidi-Sharifi N, et al. FGF2 from marrow microenvironment promotes resistance to FLT3 inhibitors in acute myeloid leukemia. Cancer Res. 2016;76:6471–6482. doi: 10.1158/0008-5472.CAN-15-3569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Jacobi A, Thieme S, Lehmann R, et al. Impact of CXCR4 inhibition on FLT3-ITD-positive human AML blasts. Exp Hematol. 2010;38:180–190. doi: 10.1016/j.exphem.2009.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Zeng Z, Shi YX, Samudio IJ, et al. Targeting the leukemia microenvironment by CXCR4 inhibition overcomes resistance to kinase inhibitors and chemotherapy in AML. Blood. 2009;113:6215–6224. doi: 10.1182/blood-2008-05-158311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Rummelt C, Gorantla SP, Meggendorfer M, et al. Activating JAK-mutations confer resistance to FLT3 kinase inhibitors in FLT3-ITD positive AML in vitro and in vivo. Leukemia. 2021;35:2017–2029. doi: 10.1038/s41375-020-01077-1. [DOI] [PubMed] [Google Scholar]
- 48.Aldinucci D, Borghese C, Casagrande N. The CCL5/CCR5 axis in cancer progression. Cancers (Basel) 2020 doi: 10.3390/cancers12071765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Waldeck S, Rassner M, Keye P, et al. CCL5 mediates target-kinase independent resistance to FLT3 inhibitors in FLT3-ITD-positive AML. Mol Oncol. 2020;14:779–794. doi: 10.1002/1878-0261.12640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Green AS, Maciel TT, Hospital M-A, et al. Pim kinases modulate resistance to FLT3 tyrosine kinase inhibitors in FLT3-ITD acute myeloid leukemia. Sci Adv. 2015;1:e1500221. doi: 10.1126/sciadv.1500221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Park I-K, Mundy-Bosse B, Whitman SP, et al. Receptor tyrosine kinase Axl is required for resistance of leukemic cells to FLT3-targeted therapy in acute myeloid leukemia. Leukemia. 2015;29:2382–2389. doi: 10.1038/leu.2015.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Dumas P-Y, Naudin C, Martin-Lannerée S, et al. Hematopoietic niche drives FLT3-ITD acute myeloid leukemia resistance to quizartinib via STAT5-and hypoxia-dependent upregulation of AXL. Haematologica. 2019;104:2017–2027. doi: 10.3324/haematol.2018.205385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Man CH, Lam SSY, Sun MKH, et al. A novel tescalcin-sodium/hydrogen exchange axis underlying sorafenib resistance in FLT3-ITD+ AML. Blood. 2014;123:2530–2539. doi: 10.1182/blood-2013-07-512194. [DOI] [PubMed] [Google Scholar]
- 54.Chang Y-T, Hernandez D, Alonso S, et al. Role of CYP3A4 in bone marrow microenvironment-mediated protection of FLT3/ITD AML from tyrosine kinase inhibitors. Blood Adv. 2019;3:908–916. doi: 10.1182/bloodadvances.2018022921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hunter HM, Pallis M, Seedhouse CH, et al. The expression of P-glycoprotein in AML cells with FLT3 internal tandem duplications is associated with reduced apoptosis in response to FLT3 inhibitors. Br J Haematol. 2004;127:26–33. doi: 10.1111/j.1365-2141.2004.05145.x. [DOI] [PubMed] [Google Scholar]
- 56.Young DJ, Nguyen B, Li L, et al. A method for overcoming plasma protein inhibition of tyrosine kinase inhibitors. Blood Cancer Discov. 2021;2:532–547. doi: 10.1158/2643-3230.BCD-20-0119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Yilmaz M, Alfayez M, DiNardo CD, et al. Outcomes with sequential FLT3-inhibitor-based therapies in patients with AML. J Hematol Oncol. 2020;13:132. doi: 10.1186/s13045-020-00964-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ward PS, Patel J, Wise DR, et al. The common feature of leukemia-associated IDH1 and IDH2 mutations is a neomorphic enzyme activity converting alpha-ketoglutarate to 2-hydroxyglutarate. Cancer Cell. 2010;17:225–234. doi: 10.1016/j.ccr.2010.01.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Dang L, White DW, Gross S, et al. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature. 2009;462:739–744. doi: 10.1038/nature08617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.DiNardo CD, Ravandi F, Agresta S, et al. Characteristics, clinical outcome, and prognostic significance of IDH mutations in AML. Am J Hematol. 2015;90:732–736. doi: 10.1002/ajh.24072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.DiNardo CD, Stein EM, de Botton S, et al. Durable remissions with ivosidenib in IDH1-mutated relapsed or refractory AML. N Engl J Med. 2018;378:2386–2398. doi: 10.1056/NEJMoa1716984. [DOI] [PubMed] [Google Scholar]
- 62.Stein EM, DiNardo CD, Pollyea DA, et al. Enasidenib in mutant IDH2 relapsed or refractory acute myeloid leukemia. Blood. 2017;130:722–731. doi: 10.1182/blood-2017-04-779405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Montesinos P, Recher C, Vives S, et al. Ivosidenib and azacitidine in IDH1-mutated acute myeloid leukemia. N Engl J Med. 2022;386:1519–1531. doi: 10.1056/NEJMoa2117344. [DOI] [PubMed] [Google Scholar]
- 64.Wagner K, Damm F, Göhring G, et al. Impact of IDH1 R132 mutations and an IDH1 single nucleotide polymorphism in cytogenetically normal acute myeloid leukemia: SNP rs11554137 is an adverse prognostic factor. J Clin Oncol. 2010;28:2356–2364. doi: 10.1200/JCO.2009.27.6899. [DOI] [PubMed] [Google Scholar]
- 65.Balss J, Meyer J, Mueller W, et al. Analysis of the IDH1 codon 132 mutation in brain tumors. Acta Neuropathol. 2008;116:597–602. doi: 10.1007/s00401-008-0455-2. [DOI] [PubMed] [Google Scholar]
- 66.Yan H, Parsons DW, Jin G, et al. IDH1 and IDH2 mutations in gliomas. N Engl J Med. 2009;360:765–773. doi: 10.1056/NEJMoa0808710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Wang F, Morita K, DiNardo CD, et al. Leukemia stemness and co-occurring mutations drive resistance to IDH inhibitors in acute myeloid leukemia. Nat Commun. 2021 doi: 10.1038/s41467-021-22874-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Figueroa ME, Abdel-Wahab O, Lu C, et al. Leukemic IDH1 and IDH2 mutations result in a hypermethylation phenotype, disrupt TET2 function, and impair hematopoietic differentiation. Cancer Cell. 2010;18:553–567. doi: 10.1016/j.ccr.2010.11.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Amatangelo MD, Quek L, Shih A, et al. Enasidenib induces acute myeloid leukemia cell differentiation to promote clinical response. Blood. 2017;130:732–741. doi: 10.1182/blood-2017-04-779447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Quek L, David MD, Kennedy A, et al. Clonal heterogeneity of acute myeloid leukemia treated with the IDH2 inhibitor enasidenib. Nat Med. 2018;24:1167–1177. doi: 10.1038/s41591-018-0115-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Choe S, Wang H, DiNardo CD, et al. Molecular mechanisms mediating relapse following ivosidenib monotherapy in IDH1-mutant relapsed or refractory AML. Blood Adv. 2020;4:1894–1905. doi: 10.1182/bloodadvances.2020001503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Intlekofer AM, Shih AH, Wang B, et al. Acquired resistance to IDH inhibition through trans or cis dimer-interface mutations. Nature. 2018;559:125–129. doi: 10.1038/s41586-018-0251-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Harding JJ, Lowery MA, Shih AH, et al. Isoform switching as a mechanism of acquired resistance to mutant isocitrate dehydrogenase inhibition. Cancer Discov. 2018;8:1540–1547. doi: 10.1158/2159-8290.CD-18-0877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Mellinghoff IK, Penas-Prado M, Peters KB, et al. Vorasidenib, a dual inhibitor of mutant IDH1/2, in recurrent or progressive glioma; results of a first-in-human phase I trial. Clin Cancer Res. 2021;27:4491–4499. doi: 10.1158/1078-0432.CCR-21-0611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Wouters BJ. Targeting IDH1 and IDH2 mutations in acute myeloid leukemia: emerging options and pending questions. Hemasphere. 2021;5:e583. doi: 10.1097/HS9.0000000000000583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Stuani L, Sabatier M, Saland E, et al. Mitochondrial metabolism supports resistance to IDH mutant inhibitors in acute myeloid leukemia. J Exp Med. 2021 doi: 10.1084/jem.20200924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.DiNardo CD, Stein AS, Stein EM, et al. mutant isocitrate dehydrogenase 1 inhibitor ivosidenib in combination with azacitidine for newly diagnosed acute myeloid leukemia. J Clin Oncol. 2021;39:57–65. doi: 10.1200/JCO.20.01632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hamann PR, Hinman LM, Beyer CF, et al. An anti-CD33 antibody-calicheamicin conjugate for treatment of acute myeloid leukemia. Choice of linker. Bioconjug Chem. 2002;13:40–46. doi: 10.1021/bc0100206. [DOI] [PubMed] [Google Scholar]
- 79.Burnett AK, Hills RK, Milligan D, et al. Identification of patients with acute myeloblastic leukemia who benefit from the addition of gemtuzumab ozogamicin: results of the MRC AML15 trial. J Clin Oncol. 2011;29:369–377. doi: 10.1200/JCO.2010.31.4310. [DOI] [PubMed] [Google Scholar]
- 80.Lamba JK, Chauhan L, Shin M, et al. CD33 splicing polymorphism determines gemtuzumab ozogamicin response in de novo acute myeloid leukemia: report from randomized phase III children's oncology group trial AAML0531. J Clin Oncol. 2017;35:2674–2682. doi: 10.1200/JCO.2016.71.2513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Lamba JK, Pounds S, Cao X, et al. Coding polymorphisms in CD33 and response to gemtuzumab ozogamicin in pediatric patients with AML: a pilot study. Leukemia. 2009;23:402–404. doi: 10.1038/leu.2008.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Gale RE, Popa T, Wright M, et al. No evidence that CD33 splicing SNP impacts the response to GO in younger adults with AML treated on UK MRC/NCRI trials. Blood. 2018;131:468–471. doi: 10.1182/blood-2017-08-802157. [DOI] [PubMed] [Google Scholar]
- 83.Short NJ, Richard-Carpentier G, Kanagal-Shamanna R, et al. Impact of CD33 and ABCB1 single nucleotide polymorphisms in patients with acute myeloid leukemia and advanced myeloid malignancies treated with decitabine plus gemtuzumab ozogamicin. Am J Hematol. 2020;95:E225–E228. doi: 10.1002/ajh.25854. [DOI] [PubMed] [Google Scholar]
- 84.Teich K, Krzykalla J, Kapp-Schwoerer S, et al. Cluster of differentiation 33 single nucleotide polymorphism rs12459419 is a predictive factor in patients with nucleophosmin1-mutated acute myeloid leukemia receiving Gemtuzumab ozogamicin. Haematologica. 2021;106:2986–2989. doi: 10.3324/haematol.2021.278894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Gbadamosi MO, Shastri VM, Meshinchi S, et al. The rs35112940 CD33 polymorphism reduces CD33 internalization and efficacy of CD33-directed Gemtuzumab Ozogamicin. Blood. 2021;138:2247. doi: 10.1182/blood-2021-150927. [DOI] [Google Scholar]
- 86.Rafiee R, Chauhan L, Alonzo TA, et al. ABCB1 SNP predicts outcome in patients with acute myeloid leukemia treated with Gemtuzumab ozogamicin: a report from Children's Oncology Group AAML0531 Trial. Blood Cancer J. 2019;9:51. doi: 10.1038/s41408-019-0211-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Walter RB, Gooley TA, van der Velden VHJ, et al. CD33 expression and P-glycoprotein-mediated drug efflux inversely correlate and predict clinical outcome in patients with acute myeloid leukemia treated with Gemtuzumab ozogamicin monotherapy. Blood. 2007;109:4168–4170. doi: 10.1182/blood-2006-09-047399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Ball ED. Pairing SOCS with CD33. Blood. 2007;109:852. doi: 10.1182/blood-2006-11-056838. [DOI] [Google Scholar]
- 89.Orr SJ, Morgan NM, Elliott J, et al. CD33 responses are blocked by SOCS3 through accelerated proteasomal-mediated turnover. Blood. 2007;109:1061–1068. doi: 10.1182/blood-2006-05-023556. [DOI] [PubMed] [Google Scholar]
- 90.Middeldorf I, Galm O, Osieka R, et al. Sequence of administration and methylation of SOCS3 may govern response to gemtuzumab ozogamicin in combination with conventional chemotherapy in patients with refractory or relapsed acute myelogenous leukemia (AML) Am J Hematol. 2010;85:477–481. doi: 10.1002/ajh.21723. [DOI] [PubMed] [Google Scholar]
- 91.Rosen DB, Harrington KH, Cordeiro JA, et al. AKT signaling as a novel factor associated with in vitro resistance of human AML to gemtuzumab ozogamicin. PLoS ONE. 2013;8:e53518. doi: 10.1371/journal.pone.0053518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.DiNardo CD, Jonas BA, Pullarkat V, et al. Azacitidine and venetoclax in previously untreated acute myeloid leukemia. N Engl J Med. 2020;383:617–629. doi: 10.1056/NEJMoa2012971. [DOI] [PubMed] [Google Scholar]
- 93.Birkinshaw RW, Gong J-N, Luo CS, et al. Structures of BCL-2 in complex with venetoclax reveal the molecular basis of resistance mutations. Nat Commun. 2019;10:2385. doi: 10.1038/s41467-019-10363-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.DiNardo CD, Tiong IS, Quaglieri A, et al. Molecular patterns of response and treatment failure after frontline venetoclax combinations in older patients with AML. Blood. 2020;135:791–803. doi: 10.1182/blood.2019003988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Yamatani K, Tabe Y, Maiti A, et al. Diverse mechanisms of resistance to decitabine and venetoclax therapy in newly diagnosed and relapsed/refractory AML inferred by transcriptome analysis. Blood. 2021;138:2244. doi: 10.1182/blood-2021-149432. [DOI] [Google Scholar]
- 96.Grant S. Rational combination strategies to enhance venetoclax activity and overcome resistance in hematologic malignancies. Leuk Lymphoma. 2018;59:1292–1299. doi: 10.1080/10428194.2017.1366999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Deng J, Carlson N, Takeyama K, et al. BH3 profiling identifies three distinct classes of apoptotic blocks to predict response to ABT-737 and conventional chemotherapeutic agents. Cancer Cell. 2007;12:171–185. doi: 10.1016/j.ccr.2007.07.001. [DOI] [PubMed] [Google Scholar]
- 98.Punnoose EA, Leverson JD, Peale F, et al. Expression profile of BCL-2, BCL-XL, and MCL-1 predicts pharmacological response to the BCL-2 selective antagonist venetoclax in multiple myeloma models. Mol Cancer Ther. 2016;15:1132–1144. doi: 10.1158/1535-7163.MCT-15-0730. [DOI] [PubMed] [Google Scholar]
- 99.Pei S, Pollyea DA, Gustafson A, et al. Monocytic subclones confer resistance to venetoclax-based therapy in patients with acute myeloid leukemia. Cancer Discov. 2020;10:536–551. doi: 10.1158/2159-8290.CD-19-0710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Bose P, Gandhi V, Konopleva M. Pathways and mechanisms of venetoclax resistance. Leuk Lymphoma. 2017;58:1–17. doi: 10.1080/10428194.2017.1283032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Tahir SK, Smith ML, Hessler P, et al. Potential mechanisms of resistance to venetoclax and strategies to circumvent it. BMC Cancer. 2017;17:399. doi: 10.1186/s12885-017-3383-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Nechiporuk T, Kurtz SE, Nikolova O, et al. The TP53 apoptotic network is a primary mediator of resistance to BCL2 inhibition in AML cells. Cancer Discov. 2019;9:910–925. doi: 10.1158/2159-8290.CD-19-0125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Pollyea DA, Stevens BM, Jones CL, et al. Venetoclax with azacitidine disrupts energy metabolism and targets leukemia stem cells in patients with acute myeloid leukemia. Nat Med. 2018;24:1859–1866. doi: 10.1038/s41591-018-0233-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Jones CL, Stevens BM, D'Alessandro A, et al. Inhibition of amino acid metabolism selectively targets human leukemia stem cells. Cancer Cell. 2018;34:724–740.e4. doi: 10.1016/j.ccell.2018.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Stevens BM, Jones CL, Pollyea DA, et al. Fatty acid metabolism underlies venetoclax resistance in acute myeloid leukemia stem cells. Nat Cancer. 2020;1:1176–1187. doi: 10.1038/s43018-020-00126-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Jones CL, Stevens BM, Pollyea DA, et al. Nicotinamide metabolism mediates resistance to venetoclax in relapsed acute myeloid leukemia stem cells. Cell Stem Cell. 2020;27:748–764.e4. doi: 10.1016/j.stem.2020.07.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Kittang AO, Kordasti S, Sand KE, et al. Expansion of myeloid derived suppressor cells correlates with number of T regulatory cells and disease progression in myelodysplastic syndrome. Oncoimmunology. 2016;5:e1062208. doi: 10.1080/2162402X.2015.1062208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Williams P, Basu S, Garcia-Manero G, et al. The distribution of T-cell subsets and the expression of immune checkpoint receptors and ligands in patients with newly diagnosed and relapsed acute myeloid leukemia. Cancer. 2019;125:1470–1481. doi: 10.1002/cncr.31896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Christopher MJ, Petti AA, Rettig MP, et al. Immune escape of relapsed AML cells after allogeneic transplantation. N Engl J Med. 2018;379:2330–2341. doi: 10.1056/NEJMoa1808777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Davids MS, Kim HT, Bachireddy P, et al. Ipilimumab for patients with relapse after allogeneic transplantation. N Engl J Med. 2016;375:143–153. doi: 10.1056/NEJMoa1601202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Goswami M, Gui G, Dillon LW, et al. Pembrolizumab and decitabine for refractory or relapsed acute myeloid leukemia. J Immunother Cancer. 2022 doi: 10.1136/jitc-2021-003392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Daver NG, Garcia-Manero G, Konopleva MY, et al. Azacitidine (AZA) with Nivolumab (Nivo), and AZA with Nivo + Ipilimumab (Ipi) in relapsed/refractory acute myeloid leukemia: a non-randomized, prospective, phase 2 study. Blood. 2019;134:830. doi: 10.1182/blood-2019-131494. [DOI] [Google Scholar]
- 113.Oldenborg PA, Zheleznyak A, Fang YF, et al. Role of CD47 as a marker of self on red blood cells. Science. 2000;288:2051–2054. doi: 10.1126/science.288.5473.2051. [DOI] [PubMed] [Google Scholar]
- 114.Sallman DA, Al Malki M, Asch AS, et al. Tolerability and efficacy of the first-in-class anti-CD47 antibody magrolimab combined with azacitidine in MDS and AML patients: phase Ib results. J Clin Oncol. 2020;38:7507. doi: 10.1200/JCO.2020.38.15_suppl.7507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Jaiswal S, Jamieson CHM, Pang WW, et al. CD47 is upregulated on circulating hematopoietic stem cells and leukemia cells to avoid phagocytosis. Cell. 2009;138:271–285. doi: 10.1016/j.cell.2009.05.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Berkovits BD, Mayr C. Alternative 3' UTRs act as scaffolds to regulate membrane protein localization. Nature. 2015;522:363–367. doi: 10.1038/nature14321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Logtenberg MEW, Jansen JHM, Raaben M, et al. Glutaminyl cyclase is an enzymatic modifier of the CD47- SIRPα axis and a target for cancer immunotherapy. Nat Med. 2019;25:612–619. doi: 10.1038/s41591-019-0356-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Welsh SJ, Corrie PG. Management of BRAF and MEK inhibitor toxicities in patients with metastatic melanoma. Ther Adv Med Oncol. 2015;7:122–136. doi: 10.1177/1758834014566428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.LoRusso PM, Krishnamurthi SS, Rinehart JJ, et al. Phase I pharmacokinetic and pharmacodynamic study of the oral MAPK/ERK kinase inhibitor PD-0325901 in patients with advanced cancers. Clin Cancer Res. 2010;16:1924–1937. doi: 10.1158/1078-0432.CCR-09-1883. [DOI] [PubMed] [Google Scholar]
- 120.Burgess MR, Hwang E, Firestone AJ, et al. Preclinical efficacy of MEK inhibition in Nras-mutant AML. Blood. 2014;124:3947–3955. doi: 10.1182/blood-2014-05-574582. [DOI] [PMC free article] [PubMed] [Google Scholar]