

Abstract
Proteinoids (thermal proteins) are produced by heating amino acids to their melting point and initiation of polymerisation to produce polymeric chains. Amino acid-like molecules, or proteinoids, can condense at high temperatures to create aggregation structures called proteinoid microspheres, which have been reported to exhibit strong electrical oscillations. When the amino acids L-glutamic acid (L-Glu) and L-aspartic acid (L-Asp) were combined with electric fields of varying frequencies and intensities, electrical activity resulted. We recorded electrical activity of the proteinoid microspheres’ ensembles via a pair of differential electrodes. This is analogous to extracellular recording in physiology or EEG in neuroscience but at micro-level. We discovered that the ensembles produce spikes of electrical potential, an average duration of each spike is 26 min and average amplitude is 1 mV. The spikes are typically grouped in trains of two spikes. The electrical activity of the ensembles can be tuned by external stimulation because ensembles of proteinoid microspheres can generate and propagate electrical activity when exposed to electric fields.
Subject terms: Biophysics, Materials science
Introduction
Thermal proteins (proteinoids)1 are produced by heating amino acids to their melting point and initiation of polymerisation to produce polymeric chains. In aqueous solution proteinoids swell forming microsphere1. The proteinoid microspheres have been considered as proto-neurons2–4. Emerging cells had similar to proteinoid microspheres’ structure. Artificial fossilization of the laboratory products closely resemble ancient microfossils from ancient strata2.
The findings on propagation of excitation waves in ensembles of proteinoid microspheres will open ways for studying coupling of biochemical and other phenomena in ways that cannot be accomplished with other models such as bilayer membranes. The proteinoids are sufficiently accurate prototypes of terrestrial protocells with bioelectrical properties5,6.
Having most characteristics of excitable cells proteinoid microspheres have been considered as proto-neurons7. By studying proteinoid oscillations, we can learn more about their molecular dynamics and how they interact with one another8. This information is essential for the search for alien life and for understanding how life first developed on Earth9.
In a 1985, paper published in the journal Biosystems, Przybylski et al.7 proposed the concept of electrical oscillations in proteinoid cells. Although they originate in different cell types, the action potentials of crayfish stretch receptor neurons and proteinoid microspheres are very similar (Fig. 1).
Figure 1.
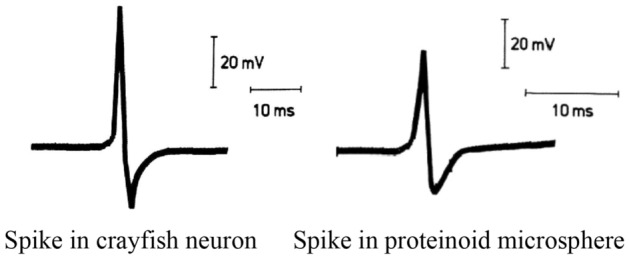
A comparison of two distinct electrical signal types is illustrated in the figure. The first form of nerve impulse is the action potential of a crayfish stretch receptor neuron. These neurons are specialised to sense changes in either length or pressure. Proteinoid microspheres, a form of artificial cell created by heat polymerisation of amino acids, generate the second electrical signal. Modified from2.
In a paper by Margel et al., glutamic acid was found to be essential for the proper folding of proteinoids10. Glutamic acid is one of the most important amino acids in the polymerization of proteinoids. It acts as a solvent and condensation agent, and is also a key component of pyroglutamic acid, which is essential for cross-linking proteinoids. Glutamic acid is also involved in the formation of disulfide bonds11,12 which are essential for the stability of proteinoids. The process of self-assembly is a key factor in the formation of both polystyrene formation13,14 and proteinoids15,16. However, there are some key differences in the way the two materials form. Polystyrene form via a process of self-assembly in the presence of surfactants. The surfactants help to lower the surface tension of the polystyrene, allowing to self assembly into nanoparticles. In contrast, proteinoids form through self assembly in the absence of surfactants. Instead the amino acids that make up proteinoids are able to self assemble due to their hydrophobic and hydrophilic interactions17.
A consensus was achieved that proteinoids directly led into neurons, which then self-associated into brains1. Thus ensembles of proteinoid microspheres can be seen as proto-brains, potentially, capable for sensorial fusion, information transmission and processing. In18, we proposed to design and prototype unconventional computing devices from ensembles of proteinoid microspheres. There are two primary tenets upon which this idea rests. The first method borrows ideas and prototypes from excitable medium computers, such as their algorithms. To simulate the self-organising behaviour of living systems, scientists turn to computers based on excitable media. This method paves the way for a dynamic and adaptable structure that can quickly respond to shifting requirements. As for the second, it makes use of reservoir computing methods19. The machine learning approach known as reservoir computing makes use of a huge pool of neurons with predetermined connections that may be easily tweaked to process new types of data. These computers have the potential to transform the way we handle data and address difficult problems by integrating the two major methodologies. Moreover, this idea may have far-reaching consequences for the future of artificial intelligence.
All prior experiments on the electrical activity of proteinoid microspheres recorded a trans-membrane potential7,20,21, similar to intra-cellular or patch-clamp approaches. In our studies we decided to test an analog of ‘extra-cellular’ recording by inserting a pair of differential electrodes into an ensemble of proteinoid micro-spheres. Our approach is somewhat similar to EEG recording of an integral neural activity: the electrical potential waves recorded from human brain are manifestations of the summarily activity of hundreds of thousands of neurons while the electrical activity of the proteinoid microspheres’ ensembles is an integration of electrical discharges of thousands of proteinoids.
Methods
The preparation of the proteinoid L-Glu:L-Asp (Fig. 2) was achieved using L-aspartic acid and L-glutamic acid with reagent grade of greater than 98% and 99% , respectively. These compounds were purchased from Sigma-Aldrich and used without further purification. The proteinoid was prepared by combining the two amino acids in a 1:1 molar ratio to a magnetically stirred 35 ml vial. The reaction vessel was then heated to 290 C for 2 days before being cooled to room temperature.
Figure 2.

The ionised carboxyl and amino groups of an L-Glu:L-Asp polypeptide are depicted here, illustrating the zwitterionic structure of the peptide. Zwitterion molecules have a net electrical charge of zero due to the orientation of their polar groups22.
At 80 C, 100 mg of the proteinoid was added to 10 ml of water in a vial. This made it possible for the proteinoid to dissolve in water. After thoroughly stirring the product, it was dialyzed using Thermo Scientific Slide-A-Lyzer Mini Dialysis Devices with a 3500 Da MWCO cellulose membrane. This allowed for the removal of any excess reagents, leaving the proteinoid in its purest form. The proteinoid solution was sealed and kept undisturbed for 12–24 h to permit dialysis. The product was then collected once the membrane was removed23.
An experimental approach was carried out to record the proteinoid voltage utilising a PicoLog USB data logger ADC 24 with a range of up to ± 2.5 V. A centimetre separated the platinum iridium electrodes, which were also connected to the data recorder. The 1-s sampling interval was chosen. Mixing an adequate amount of proteinoid powder with distilled water while stirring until the powder was completely dissolved yielded the proteinoid solution. The solution was then drawn up with a 5 ml syringe and injected into the space between the two electrodes in a 5 ml vial.
The voltage was recorded in the input range to ± 130 mV and the sample rate was 1 s.
This work used the field emission scanning electron microscope (FESEM) FEI Quanta 650 to explore the proteinoids L-Glu:L-Asp.To acquire a precise SEM image of the proteinoid, samples were initially coated with a thin layer of gold. Gold plating supplies the electron beam with a conductive surface, resulting in a high-quality image. Additionally, the gold coating protects the sample from damage induced by the electron beam, which might result in sample degradation. The particle size distribution was estimated using the Image analysis software ImageJ24. Electrical activity was plotted using DataGraph, we have used standard Matlab functions provided in Statistics toolbox.
Results
Spheroid clusters with a diameter of less than 100 nm may be seen forming in the Scanning Electron Microscopy (SEM) image of the proteinoid ensembles L-Glu:L-Asp (Fig. 3a). Agglomeration of proteinoid molecules leads to the production of these microspheres. The Lorentzian distribution of particle size and relative-molecular-mass dispersity of proteinoid nanoparticles is shown in Fig. 3b. A relative-molecular-mass dispersity of 99.1 % was calculated from the fitted data, which indicated that the average particle diameter was 25 nm25. This is evidence of remarkable consistency.The ratio of the standard deviation of the fit data to the average particle size was used to figure out the relative molecular mass dispersity. In addition, two scanning electron micrograph (SEM) images of connected nanostructures are displayed in Fig. 3c,d. Nanoparticles of proteinoids are microscopic spheres that can aggregate into bigger structures. The Lorentzian distribution shows the nanoparticles’ size and relative-molecular-mass dispersity and sheds light on how the particles clump together. When the majority of the particles fall to the left side of the curve, we say that the distribution is skewed to the left. In other words, little particles are more robust than big ones. What this means is that the median particle size is larger than the mean particle size. Particle size distributions typically have a left-skewed shape because small particles prefer to cluster together, increasing the proportion of small particles in the distribution. Larger particles are less likely to concentrate in one location since they are heavier and more difficult to shift.
Figure 3.

Characterisation of proteinoid ensembles. (a) SEM image shows the formation of proteinoid L-Glu:L-Asp nanoparticles, which are created by the self-assembly of amino acids and monomers. (b) In a Lorentzian fit to the SEM data, the nanoparticles’ small diameter of 25 nm and a relative-molecular-mass dispersity of 99.1 % are both indicative of a highly uniform size distribution. (c) This figure shows how the proteinoid’s microsphers are interconnected by a complex system of channels. Proteinoids rely on these channels for signal transmission and reception, allowing them to function properly. (d) With a scale bar of 177 nm, a magnified picture of proteinoid microspheres exhibiting their detailed structure.
Figure 4a shows electrical potential, recorded in a pair of differential electrodes inserted in the ensemble of proteinoid microspheres made of L-Glu:L-Asp. Pronounced oscillation in the electrical output indicates that there is significant electrical activity between the microspheres. A typical spike of electrical potential is shown in Fig. 4b.
Figure 4.

Electrical activity recorded in the ensemble of proteinoid microspheres via a pair of differential electrodes. (a) The activity recorded during 21 h. Few characteristic voltage spikes are magnified in the inserts. (b) A typical spike of electrical potential. The proteinoid aqueous solution L-Glu:L-Asp created its own electrical activity via self-assembly. During this process, an electrical charge was produced when the proteinoid molecules spontaneously arranged themselves into highly organised and stable structures. This mechanism is driven by the presence of electrostatic dipoles in proteinoid molecules, which originate from the breakdown of hydrogen bonds between their side chains. These electrostatic interactions between proteinoid molecules generate an electrical potential throughout the solution, resulting in electrical activity.
Of twenty spikes measured we found that average spike duration is 1574 s, , median 1360 s, minimum 830 s, maximum 4130 s. That is a typical spike recorded lasts for c. 26 min. Spike duration distribution is show in Fig. 5a. The distribution has relatively short right tail but still more spread to the right, as skewness = 2.79568212 and kurtosis =13.2915541.
Figure 5.

Characteristic of spiking. (a) Distribution of spike lengths, bin size is 200 s. (b) Distribution of spike amplitude, bin size is 0.3 mV.
Average spike amplitudes varied from minimum 0.3 mV to maximum 1.9 mV, average amplitude 1 mV, , median 1 mV. Distribution of spike amplitudes is shown in Fig. 5b. The distribution is almost symmetric with very small leaning to the right: skewness = 0.052873856 and kurtosis = 2.80547952.
We say spikes form a train when a distance between consecutive spikes does not exceed a double of average spike width. All trains of spikes observed have just two spikes each. An example of a train of spikes is shown in magnifying in insert at the top right of the plot Fi(g. 4a).
Ensembles of proteinoid micropheres, made of L-Glu:L-Asp, can be stimulated to create electrical signals of varying amplitudes and frequencies by being subjected to a range of frequencies, from 0.001 Hz to 100 kHz, for 1 min. Exposed to these frequencies, the proteinoid will begin to generate electrical oscillations with a period of 1 h and voltages ranging from − 2.29 to 12.771 mV. After 1 min of stimulation at frequencies ranging from 0.001 Hz to 100 kHz, the electrical oscillations of a proteinoid containing L-Glu:L-Asp are depicted in Fig. 6. This suggests that the proteinoid can be stimulated by electrical oscillations of varying frequencies. Therefore, it appears that the proteinoid can pick up on the frequencies and react appropriately. These findings add to the growing body of evidence that proteinoids can sense and react to their environment.
Figure 6.

The pronounced electrical oscillations of the L-Glu:L-Asp proteinoid microspheres ensembles after being stimulated for 1 min with frequencies between 0.001 Hz and 100 kHz.
Discussion
In laboratory experiments, we demonstrated that ensembles of L-Glu:L-Asp proteinoid microspheres generated oscillations of electrical potential recorded via pair of differential electrodes. The spikes of the electrical potential reflect propagation of electrical activity in proteinoid ensembles. An average duration of a voltage spike is 26 min and average amplitude 1 mV. The value is much smaller than electrical potential amplitudes, ranging from 20 to 70 mV, recorded ‘intra-spherically’, between interior and exterior of proteinoid microspheres20, this is because strength of a signal is substantially smaller in case of ‘extra-spherical’ recording. We can speculate that the spiking recorded is manifestation of the coordinated electrical activity propagating in the microspheres’ ensembles. Reported duration of ‘intra-spherically’ recorded voltage spikes are around 1 s. Distance between electrodes in our experiments was 10 mm. Media duration of spikes recorded in our experiments is 1360 s. Assuming the activity propagates along a shortest path connecting the electrodes we can propose that there are c. 1360 proteinoid microspheres along the path, which might give an indicated diameter of a microsphere as c. 7 m. This is less than the sizes of the microspheres, 20–200 m1, reported previously. Therefore we can also speculate that it might take some time, at least 3 s, exact value to be investigated in further experiments, for electrical activity to propagate between the proteinoid microspheres.
Why do ensembles of proteinoid microspheres form?
Electrostatic interactions between the charged amino acids in the proteinoid are most likely responsible for the clustering. Since proteinoids are made up of amino acids, which are the foundation of proteins, their formation is significant. Proteinoids are able to organise themselves into a more sophisticated structure by aggregating into nanoparticles. This higher level of complexity has the potential to improve the nanostructure’s stability, which in turn could have future applications. The creation of proteinoid microspheres can be broken down into three distinct stages, each of which is seen in the image (Fig. 7) below. First, proteinoids cluster together by a process called nucleation. Next, the molecules will cluster together to create a larger aggregate. The clumps eventually coalesce into microscopic spheres. Below, each of these steps will be discussed in detail.
Figure 7.

The three-step production process of proteinoid microspheres: nucleation, clustering, formation of microspheres.
Nucleation: The amino acid building blocks of proteins, called proteinoids, are heated in a watery solution. Due to this, 1–2 nm sized nano-spheres are formed.
Clustering: Following this, the nanospheres coalesce into bigger clusters of 50 nm in size. Hydrophobic interactions, van der Waals forces, and hydrogen bonding all play a role in facilitating this clustering.
Microspheres formation: When additional molecules, including phospholipids and carbohydrates, are present in the solution, the clustered proteinoid molecules are further stabilised and form larger microspheres. This procedure produces microspheres of proteinoid several hundred nanometers in diameter that are extremely stable.
What are possible mechanisms of the oscillations?
An external stimulus, like as light or an electrical current, can trigger electrical oscillations in proteinoids. It is assumed that electrostatic interactions between the proteinoids and their environment are responsible for generating the oscillations, which in turn depend on the physical and chemical properties of the proteinoids as well as their concentrations. The interaction between L-Glu and L-Asp is likely responsible for the fluctuations in electrical activity, which in turn imply a dynamic exchange of ions between the microspheres. L-Glu is a cation with a single positive charge at a pH of 7.4. Since the amino group gets an extra proton. On the other hand, L-Asp has a negative charge26,27. This results in an electrostatic relationship between the two amino acids, which results in an electrical current and electrical oscillations. The presence of water molecules in the proteinoid solution further enhances the oscillations. The water molecules serve as electrolytes, so facilitating the flow of electrical current and enhancing the oscillations. The inclusion of other amino acids in the solution, such as L-Arginine, can further amplify the oscillations and amplify the electrical current.
Proteinoid aqueous solutions can be understood in terms of the behaviour of the proteinoid molecules, which causes electrical oscillations. Every molecule of a proteinoid in solution carries an electrical charge, which changes as the molecule shifts position and interacts with others. The proteinoids store and release electrical energy like a capacitor due to their changing charge. The solution’s oscillatory behaviour is the result of the energy being released as electrical oscillations. Proteinoids can also form complex interactions with other solute molecules. Clusters of molecules can form as a result of this interaction, increasing the power of the electrical oscillation produced by the proteinoids. Clusters of proteinoid molecules contribute further to the solution’s oscillatory behaviour by storing and releasing energy more efficiently than individual molecules.
Proteinoids interacting with other particles, such electrons, results in electrical oscillations. Charged proteinoids generate an electric field when they come into contact with electrons. The electrical waves originate from an underlying electrical field that oscillates at the same frequency as the proteinoids.
Proteinoids are sensitive to the frequencies of their environmental inputs, which impacts the period of their electrical oscillations. Proteinoids, which are made up of amino acid chains that interact with one another to form a polypeptide, may be electrically stimulated at frequencies ranging from 0.001 Hz to 100 kHz. Stimulating the proteinoid with these frequencies causes its period of electrical oscillations to grow from 25 min to a maximum of 2 h and 45 min. The ability of the polypeptide to absorb and store energy from the environmental inputs is linked to the lengthening of the oscillation period. The polypeptide’s structure becomes more ordered and stable when it is exposed to a frequency within its range and absorbs the energy, storing it in its bonds. Longer electrical oscillation periods come from the polypeptide’s enhanced stability, which allows it to absorb and store more energy.
Moreover, the structure of polypeptides is related to their sensitivity to particular frequencies. Proteinoid structure and amino acid sequences are two elements that influence how proteinoids respond to specific frequencies. Some proteinoids respond better to high frequencies whereas others respond better to low frequencies, for example. The period of oscillations increases as frequency increases because the structure of polypeptides adapts to meet it.
Conclusions
The electrical activity propagation in ensembles of proteinoid microspheres is a complicated process that depends on a wide range of variables, such as the kind and concentration of proteinoids, the resistivity of the medium, and the gap junctions between microspheres. Ion exchange between proteinoid microspheres and the conductive media is responsible for their neuron like spiking activity. It is shown that proteinoid microsphere ensembles have the potential to be used as a model system for studying the propagation of electrical activity. While previous patch clamp and ‘intra-microsphere’ research demonstrated that proteinoids produce action-potential likes spikes, our research demonstrated it is possible to recorded EEG like pattern from the ensembles of proteinoid microspheres. Thus the proteinoid ensembles may be seen as nano-brains composed of hundred of neurons where each neuron is physically represented by a proteinoid microsphere.
Supplementary Information
Acknowledgements
The research was supported by EPSRC Grant EP/W010887/1 “Computing with proteinoids”. Authors are grateful to David Paton for helping with SEM imaging and to Neil Phillips for helping with instruments.
Author contributions
P.M. and A.A. wrote the main manuscript text and prepared figures. All authors reviewed the manuscrpit.
Data availibility
All data generated or analysed during this study are included in this published article (and its supplementary information files).
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-023-29067-0.
References
- 1.Fox SW. Evolution of Information Processing Systems. Springer; 1992. Thermal proteins in the first life and in the “mind-body” problem; pp. 203–228. [Google Scholar]
- 2.Fox SW, Bahn PR, Dose K, Harada K, Hsu L, Ishima Y, Jungck J, Kendrick J, Krampitz G, Lacey JC, et al. Experimental retracement of the origins of a protocell. J. Biol. Phys. 1995;20(1–4):17–36. doi: 10.1007/BF00700418. [DOI] [Google Scholar]
- 3.Rizzotti M, Crisma M, DeLuca F, Iobstraibizer P, Mazzei P. Exobiology: Matter, Energy, and Information in the Origin and Evolution of Life in the Universe. Springer; 1998. Did the first cell emerge from a microsphere? pp. 199–202. [Google Scholar]
- 4.Matsuno K. Molecular Evolution and Protobiology. Springer; 2012. [Google Scholar]
- 5.Follmann H. Deoxyribonucleotide synthesis and the emergence of dna in molecular evolution. Naturwissenschaften. 1982;69(2):75–81. doi: 10.1007/BF00441226. [DOI] [PubMed] [Google Scholar]
- 6.VanGestel J, Tarnita CE. On the origin of biological construction, with a focus on multicellularity. Proc. Natl. Acad. Sci. 2017;114(42):11018–11026. doi: 10.1073/pnas.1704631114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Przybylski AT. Excitable cell made of thermal proteinoids. Biosystems. 1985;17(4):281–288. doi: 10.1016/0303-2647(85)90044-9. [DOI] [PubMed] [Google Scholar]
- 8.Jerman I. Biological, Physical and Technical Basics of Cell Engineering. Springer; 2018. Emergence of organisms from ordered mesoscopic states of water (liquids)-physical instead of chemical origin of life; pp. 321–338. [Google Scholar]
- 9.Fox SW. Life from an orderly cosmos. Naturwissenschaften. 1980;67(12):576–581. doi: 10.1007/BF00396536. [DOI] [PubMed] [Google Scholar]
- 10.Kolitz-Domb M, Margel S. Engineered narrow size distribution high molecular weight proteinoids, proteinoid-poly (l-lactic acid) copolymers and nano/micro-hollow particles for biomedical applications. J. Nanomed. Nanotechnol. 2014;5(216):2. [Google Scholar]
- 11.Leqraa, N. & Vallée, Y. From amino acids to peptides before the coming of ribosomes. In Prebiotic Chemistry and Life’s Origin, pp. 177–214 (2022)
- 12.Matsuo M, Kurihara K. Proliferating coacervate droplets as the missing link between chemistry and biology in the origins of life. Nat. Commun. 2021;12(1):1–13. doi: 10.1038/s41467-021-25530-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Junfan C, Cong Z, Jie Z, Yong Z. Raman enhancement of large-area silver grating arrays based on self-assembled polystyrene microspheres. Opt. Mater. Express. 2021;11(4):1234–1246. doi: 10.1364/OME.422627. [DOI] [Google Scholar]
- 14.Dahle S, Meuthen J, Gustus R, Prowald A, Viöl W, Maus-Friedrichs W. Superhydrophilic coating of pine wood by plasma functionalization of self-assembled polystyrene spheres. Coatings. 2021;11(2):114. doi: 10.3390/coatings11020114. [DOI] [Google Scholar]
- 15.Itzhaki E, Hadad E, Moskovits N, Stemmer SM, Margel S. Tumor-targeted fluorescent proteinoid nanocapsules encapsulating synergistic drugs for personalized cancer therapy. Pharmaceuticals. 2021;14(7):648. doi: 10.3390/ph14070648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lugasi, L., Otis, G., Oliel, M., Margel, S., & Mastai, Y. Chirality of proteinoid nanoparticles made of lysine and phenylalanine. Polymers for Advanced Technologies.
- 17.Bamnolker H, Nitzan B, Gura S, Margel S. New solid and hollow, magnetic and non-magnetic, organic-inorganic monodispersed hybrid microspheres: Synthesis and characterization. J. Mater. Sci. Lett. 1997;16(16):1412–1415. doi: 10.1023/A:1018565428355. [DOI] [Google Scholar]
- 18.Adamatzky A. Towards proteinoid computers. Hypothesis paper. Biosystems. 2021;208:104480. doi: 10.1016/j.biosystems.2021.104480. [DOI] [PubMed] [Google Scholar]
- 19.Han X, Zhao Y. Reservoir computing dissection and visualization based on directed network embedding. Neurocomputing. 2021;445:134–148. doi: 10.1016/j.neucom.2021.02.029. [DOI] [Google Scholar]
- 20.Przybylski AT, Stratten WP, Syren RM, Fox SW. Membrane, action, and oscillatory potentials in simulated protocells. Naturwissenschaften. 1982;69(12):561–563. doi: 10.1007/BF00396351. [DOI] [PubMed] [Google Scholar]
- 21.Przybylski AT. Molecular Evolution and Protobiology. Springer; 1984. Physical background of excitability: Synthetic membranes and excitable cells; pp. 253–266. [Google Scholar]
- 22.Hanwell MD, Curtis DE, Lonie DC, Vandermeersch T, Zurek E, Hutchison GR. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012;4(1):1–17. doi: 10.1186/1758-2946-4-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Przybylski AT, Fox SW. Excitable artificial cells of proteinoid. Appl. Biochem. Biotechnol. 1984;10(1):301–307. doi: 10.1007/BF02783764. [DOI] [PubMed] [Google Scholar]
- 24.Collins TJ. Imagej for microscopy. Biotechniques. 2007;43(S1):S25–S30. doi: 10.2144/000112517. [DOI] [PubMed] [Google Scholar]
- 25.Stepto RF. Dispersity in polymer science (iupac recommendations 2009) Pure Appl. Chem. 2009;81(2):351–353. doi: 10.1351/PAC-REC-08-05-02. [DOI] [Google Scholar]
- 26.Neuberger A. Dissociation constants and structures of glutamic acid and its esters. Biochem. J. 1936;30(11):2085. doi: 10.1042/bj0302085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Voet D, Voet JG, Pratt CW. Fundamentals of Biochemistry: Life at the Molecular Level. Wiley; 2016. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data generated or analysed during this study are included in this published article (and its supplementary information files).