

Abstract
This review summarizes and systematizes the literature on the anti-HIV activity of plant coumarins with emphasis on isolation and the mechanism of their antiviral action. This review summarizes the information on the anti-HIV properties of simple coumarins as well as annulated furano- and pyranocoumarins and shows that coumarins of plant origin can act by several mechanisms: inhibition of HIV reverse transcriptase and integrase, inhibition of cellular factors that regulate HIV-1 replication, and transmission of viral particles from infected macrophages to healthy ones. It is important to note that some pyranocoumarins are able to act through several mechanisms or bind to several sites, which ensures the resistance of these compounds to HIV mutations. Here we review the last two decades of research on the anti-HIV activity of naturally occurring coumarins.
Keywords: coumarins, anti-HIV agents, reverse transcriptase inhibitors, integrase inhibitors, pyranocoumarins, furocoumarins, natural products
1. Introduction
Coumarin is a heterocyclic system of annulated pyrone and benzene rings. Coumarins are widely distributed in the plant kingdom, and are present in the roots, stems, bark, leaves, seeds, fruits, or flowers of various plant species. While unsubstituted coumarin can have significant hepatotoxicity [1], oxygenated coumarin derivatives generally have low toxicity [2] (expect aflatoxin coumarins), which may be due to the destructive opening of the pyrone ring [3] and hydroxylation [2] leading to the formation of easily excreted highly hydrophilic products. These properties offer great potential for combining coumarin derivatives with other compounds in the treatment of diseases [4,5]. This is especially important for the highly effective antiretroviral therapy proposed by David Ho in 1996 [6,7].
In addition, the synthesis of hydroxyl-containing coumarins by Pechmann condensation [8,9] is a convenient protocol, which makes this class of heterocyclic compounds particularly easy to access, both in terms of obtaining libraries of compounds to optimize the structure of the drug lead and in terms of large-scale industrial production.
Over the past 30 years, natural coumarin compounds have attracted great attention as lead compounds for developing low molecular weight antiviral agents, particularly anti-HIV agents [10,11]. Despite the fact that there are no approved coumarin derivatives in clinical practice, several natural and semi-synthetic coumarins have recently been discovered and are currently undergoing various phases of clinical trials [12].
The HIV replication cycle consists of thirteen steps (Figure 1) [13]; each step can be the target of chemotherapeutic interventions. The viral cycle begins with virion attachment to the membrane receptor of the T cell (step 1), followed by entry of viral components into the cell (step 2). Uncoated viral reverse transcriptase (RT) and viral RNA trigger reverse transcription (step 4), producing viral DNA, which is transported to the nucleus (step 5) and incorporated into the DNA of the infected cell (step 6). Viral RNA is synthesized in the nucleus (step 7), and the viral RNA leaves the nucleus (step 8) and serves to synthesize viral proteins (step 9). Then, the synthesized viral components assemble in new virions (step 10) and leave the infected cell (steps 11 and 12). After protease-catalyzed maturation (step 13), they may infect new cells.
Figure 1.

Steps of the HIV-1 replication cycle.
Currently, the vast majority of antiretroviral drugs target the steps of reverse transcription and integration (steps 4 and 6). In 2022, there were 14 FDA-approved reverse transcriptase and integrase inhibitors, while only 7 drugs that aimed to block the remaining 11 stages had been approved [14]. RT inhibitors can be nucleoside or non-nucleoside RT inhibitors (NRTIs and NNRTIs). NRTIs contain a nucleoside base, which is incorporated into the growing DNA chain, terminating its elongation, while NNRTIs block the active site of the RT itself.
In terms of the mechanism of antiviral action, anti-HIV coumarins can be divided into four groups (Figure 2): NNRTIs (this group includes simple coumarins I, linear furocoumarins IIa, as well as both angular and linear pyranocoumarins IIIa and IIIb, respectively), the largest group; integrase inhibitors (coumestans IIb); compounds inhibiting cellular factors that regulate HIV-1 replication, for example, by inhibition of the transcription factor NF-κB (simple coumarins I, pyranocoumarins IIIc); and inhibitors of the transmission of viral particles from infected macrophages to healthy ones, for example, inhibiting HIV-1 entry, and downregulating the expression of chemokine receptors CXCR4, CD4, and CCR5 (simple coumarins I, furocoumarins IIa, and pyranocoumarins IIIa). In addition, for some compounds there are still no reliable data on the mechanism of action [15]. Therefore, typical coumarins with anti-HIV activity are RT and integrase inhibitors, so they can potentially be used as components of highly effective antiretroviral therapy.
Figure 2.

Molecular targets for naturally occurring anti-HIV coumarins.
Naturally occurring anti-HIV coumarins may be classified by their structure into three main groups: simple coumarins I, furocoumarins (psoralene-type IIa and coumestan-type IIb), and pyranocoumarins (angular IIIa and linear IIIb) (Figure 2) [16].
This review provides a full account of simple coumarins I and their annulated analogs II and III as potential anti-HIV agents, with emphasis on recent articles published in the last 5 years. Since not all coumarins have a known mechanism of anti-HIV action but all have a known structure, in this review, we have systematized the antiviral coumarins according to their structure.
2. Simple Coumarins
In the group of simple coumarins, we included compounds that contain the benzopyrone system and do not contain other cycles annulated with it; substituted coumarins are also included in this section. Simple natural coumarins usually contain methoxy, hydroxy groups and/or 1–3 prenyl residues as substituents of benzo or pyran rings via an optional -O-linker.
Methoxy- and hydroxy-derivatives of coumarin represent a small group of anti-HIV coumarins with moderate antiviral activity. 5-, 6-, 7-, and 8-methoxy- as well as 6- and 7-hydroxycoumarins are known.
It was reported that aesculetin 1 (Figure 3) isolated from Fraxinus ornus (manna ash) [17], Manihot esculenta (cassava) [18] and Artemisia capillaries [19] was able to inhibit HIV replication in H9 cells with EC50 = 2.51 μM and a therapeutic index (TI) of 11.2 (cytotoxic concentration (CC50) = 28.1 μM) [19].
Figure 3.

Anti-HIV hydroxy- and methoxycoumarins 1–3.
Citropten (5,7-dimethoxycoumarin) 2 and 6,8-dimethoxycoumarin 3 (Figure 3), found in the plant families Campanulaceae [20] and Rutaceae [21,22], showed moderate anti-HIV activity with EC50 = 33.8 μM and CC50 = 417 μM (TI = 12.4) and EC50 = 100.9 μM and CC50 > 970.9 μM (TI > 7.36), respectively [22], while 5,7,8-trimethoxycoumarin 4 (Figure 3) from Zapota ailanthoides showed high activity with EC50 = 0.933 μM and TI = 107 [23].
Various derivatives of terpene coumarins and related compounds isolated from plants of the families Rutaceae [22,23,24,25,26,27,28], Umbelliferae [29,30], Fabaceae [31,32] were found to be active against HIV (Table 1) [22,23,24,32]. Among them, auraptene 5 and anisocoumarin B 6 inhibit the replication of HIV-1 (strain LAV-1) with EC50 activities of 5.7 µM and 23.2 µM, respectively, on peripheral blood mononuclear cells [24]. Regarding the HIV-1 IIIB strain, the activity of auraptene 5 was 10 times lower (EC50 = 59.7 μM) in the C8166 cell line [24]. Anisocoumarin B 6 inhibits HIV viral replication with EC50 18.3 μM, with a sufficiently high TI > 44.4 [22]. In Sup T1 cells, Isobtusitin 7 had a weak HIV inhibitory effect (about 10%) with a concentration close to CC50, namely 20 μg/mL [33].
Table 1.
O-terpenoid anti-HIV coumarins.
| Compound | Plant Source | Anti-HIV Activity and Toxicity |
|---|---|---|
 auraptene 5 |
Triphasia trifolia [24] Zanthoxylum schinifolium [25] |
EC50 = 5.7 μM CC50 = 46.8 μM TI = 8.21 [24] |
 anisocoumarin B 6 |
Poncirus trifoliata [22] Clausena anisata [31] |
EC50 = 18.3 μM CC50 = 813 μM TI = 44.4 [24] |
 isobtusitin 7 |
Psiadia dentata [33] | EC10 = 20 μM CC50 = 20 μM [33] |
 farnesiferol C 8 |
Ferula assa-foetida [35] Ferula szowitsiana [36] Ferula kopetdaghensis [37] |
EC50 = 30 μM [35] CC50 > 150 μM [34] |
Farnesiferol C 8 (Table 1) is a well-known sesquiterpene coumarin isolated from genus Ferula (Apiaceae) species such as Ferula assafoetida and Ferula szowitsiana. It has been reported to have cytotoxic, apoptotic, MDR reversal, antitumor, antimutagenic, and antiviral activity [34]. Recently, farnesiferol C 8 was identified as an HIV-1 RT inhibitor with a mixed inhibition mechanism (IC50 = 30 μM). Using theoretical and experimental methods, it was demonstrated that farnesiferol C 8 could quench the intrinsic fluorescence emission of HIV-1 RT through a static quenching mechanism, while molecular docking studies indicated that farnesiferol C 8 interacts with the enzyme through a hydrophobic pocket [35].
The low toxicity of coumarin O-terpenoids (up to 813 μM) allows for an acceptable TI in spite of the low EC50 (5 to 60 μM).
C-terpenoid anti-HIV coumarin derivatives are represented by a large number of compounds. In general, C-terpenoids are characterized by higher activity compared to their O- counterparts. Among them, it is possible to distinguish monoterpenoids and diterpenoids (Table 2).
Table 2.
C-terpenoid anti-HIV coumarins.
| Compound | Plant Source | Anti-HIV Activity and Toxicity |
|---|---|---|
| Monoterpenoids | ||
| C3-monoterpenoids 9 | ||

9 |
Poncirus trifoliata [22] | EC50 = 10.7 μM CC50 = 330 μM TI = 31 [22] |
| C6-monoterpenoids 10–16 | ||
 fipsomin 10 |
Ficus nipponica [38] Artocarpus heterophyllus [45] |
EC50 = 2.93 μM CC50 > 200 μM TI > 68.26 [45] |
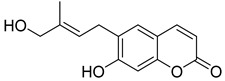 phellodenol C 11 |
Phellodendron amurense [39] Artocarpus heterophyllus [45] |
EC50 = 8.07 μM CC50 > 200 μM TI > 21.93 [45] |
 isophellodenol C 12 |
Heracleum candicans [40] Artocarpus heterophyllus [45] |
EC50 = 9.12 μM CC50 > 200 μM TI > 24.78 [45] |

13 |
Manilkara zapota [41] Ruta pinnata [42] Skimmia laureola [43] Acalypha indica [44] |
EC50 = 8.69 μM CC50 > 200 μM TI > 38.02 [41] |

14 |
Artocarpus heterophyllus [45] Brombya sp. Nova [46] |
EC50 = 5.68 μM CC50 > 200 μM TI > 35.21 [45] |
 (+)-trachypleuranin A 15 |
Manilkara zapota [41] Harbouria trachypleura [47] |
EC50 = 5.26 μM CC50 > 200 μM TI > 38.02 [41] |

16 |
Manilkara zapota [41] Skimmia laureola [43] Clausena lenis [48] |
EC50 = 6.73 μM CC50 > 200 μM TI > 29.72 [41] EC50 = 7.39 μM CC50 > 200 μM TI > 31.9 [48] |
| C8-monoterpenoids 17, 18 | ||
 osthol 17 |
Many species. For recent reviews, see [49,50] | EC50 = 0.64 μM CC50 = 48 μM TI = 75.5 [30] |
 O-methylcedrelopsin 18 |
Zanthoxylum ailanthoides [23] | EC50 = 7.66 μM CC50 = 77 μM TI = 36.6 [23] |
| Diterpenoids | ||
| C3,C6-diterpenoids 19–27 | ||
 gravelliferone 19 |
Manilkara zapota [41] Clausena lansium [51] Helietta apiculata [52] |
EC50 = 2.28 μM CC50 > 200 μM TI > 87.7 [41] |
 O-methyl gravelliferone 20 |
Manilkara zapota [41] Ruta pinnata [53] |
EC50 = 3.49 μM CC50 > 200 μM TI > 35.3 [41] |
 tanizin 21 |
Manilkara zapota [41] Artocarpus heterophyllus [45] Helietta apiculata [52] |
EC50 = 4.26 μM CC50 > 200 μM TI > 47.0 [41]EC50 = 0.56 μM CC50 > 200 μM TI > 357 [45] |
 manizapotin A 22 |
Manilkara zapota [41] | EC50 = 0.12 μM CC50 > 200 μM TI > 1667 [41] |
 manizapotin B 23 |
Manilkara zapota [41] | EC50 = 0.33 μM CC50 > 200 μM TI > 606 [41] |
 manizapotin C 24 |
Manilkara zapota [41] | EC50 = 0.42 μM CC50 > 200 μM TI > 467 [41] |
 clauselenin B 25 |
Clausena lenis [48] | EC50 = 0.68 μM CC50 > 200 μM TI > 294 [48] |
 artoheteronin 26 |
Artocarpus heterophyllus [45] | EC50 = 0.18 μM CC50 > 200 μM TI > 1111 [45] |
 anisocoumarin A 27 |
Manilkara zapota [41] Artocarpus heterophyllus [45] |
EC50 = 0.97 μM CC50 > 200 μMTI > 206 [41] EC50 = 0.33 μM CC50 > 200 μMTI > 606 [45] |
| C6,C8-diterpenoids 28 | ||
 6,8-diprenylumbelliferone 28 |
Clausena lenis [48] | EC50 = 1.59 μM CC50 > 200 μM TI > 125 [48] |
3-(1,1-Dimethylallyl)-8-hydroxy-7-methoxycoumarin 9 (Table 2), the only representative of the C3-monoterpenoids, inhibits HIV viral replication with EC50 10.7 μM, with a sufficiently high TI (31.0) [22].
C6-coumarin monoterpenoids include fipsomin 10 [38], phellodenol C 11 [39], isophellodenol C 12 [40], and coumarins 13-16 [41,42,43,44,45,46,47,48] (Table 2). C6-terpenoids are characterized by moderate anti-HIV activity (3–8 μM).
Osthol 17 and O-methylcedrelopsin 18, C8-coumarin monoterpenoids, exhibited high inhibitory activity against HIV replication in H9 cells with EC50 values of 0.155 μg/mL (0.64 μM) and 0.576 μg/mL (2.10 μM), respectively, and TIs of 75.5 [30] and 36.6 [23], respectively.
Diterpene derivatives represent a large class of terpene coumarins, which usually have the highest anti-HIV activity.
C3,C6-diterpenoids are characterized by high activity against HIV-1 RT, with IC50 values 4-0.12 μM, in combination with low toxicity (CC50 > 200 μM) [32,41,45] (Table 2). Coumarin diterpenoids have been isolated from a large variety of plant materials [32,41,45,51,52,53].
Ethanol extract of the fruits of Manilkara zapota contains at least 7 coumarin diterpenoids with high anti-HIV activity: tanizin 21 (EC50 = 0.56 μM), manizapotins A, B, and C 24, 25, and 26 (EC50 = 0.12, 0.33, and 0.42 μM, respectively), anisocoumarin A 27 (EC50 = 0.33 μM), gravelliferone 19 and its methyl ether 20 (EC50 = 2.28 and 3.49 μM), as well as monoterpenoid coumarins 15 and 16 (EC50 = 5.26 and 6.37 μM, respectively). Gravelliferone 19, O-methyl gravelliferone 20, and tanizin 21 were also isolated from Clausena lansium [51], Helietta apiculata [52], Ruta pinnata [53], Artocarpus heterophyllus [45] and Helietta apiculata [52] species.
Clauselenin B 25, together with diprenylated coumarin 28 isolated from stems of Clausena lenis, demonstrated high activity against HIV-1 RT (EC50 = 0.68 and 1.59 μM for compounds 25 and 28, respectively) [32].
Another diprenylated coumarin, artoheteronin 26, was isolated from fruits of Artocarpus heterophyllus. This compound demonstrated anti-HIV RT activity with EC50 = 0.18 μM [45].
Anisocoumarin E 29, a representative of the C,O-diterpene coumarins (Figure 4), was described in 2010 as an HIV RT inhibitor with IC50 = 23.2 μM and CC50 = 96.8 μM (TI = 4.17) [24].
Figure 4.

C,O-diterpenoid and phenyl-substituted mono- and diterpenoid coumarins with anti-HIV activity.
T. Okuda and colleagues demonstrated that 3-arylcoumarin, namely glycycoumarin 30 (Figure 4), isolated from licorice inhibits giant cell formation in HIV-infected cultures at 20 μg/mL without any observed cytotoxicity [32]. Later, J. Alcami et al. studied natural 4-phenyl-coumarins isolated from the Marila pluricostata plant (family Clusiaceae) for their activity against HIV transcription, which occurred by the mechanism of inhibition of the transcription factor NF-κB and by the function of transactivation of transcription (Tat). Among the studied coumarins, only two derivatives (disparinol A 31 and isodispar B 32 (Figure 4)) showed high inhibitory efficacy towards NF-κB proteins (81.0% and 55.4% at 25 μM respectively) and Tat (similar for both, ~40% at 25 μM and ~70% at 50 μM) [54].
In 2022, Fobofou et al. reported a new type of coumarin dimer with activity against HIV [15]. In particular, 8,8’-dicoumarin derivative (bichromonol) 33 (Figure 5) isolated from the stem bark of Hypericum roeperianum exhibits anti-HIV activity in infected MT-4 cells. Bichromonol demonstrated micromolar activity with EC50 = 6.6–12.0 µM against both wild-type HIV-1 and its clinically relevant mutant strains. Bichromonol is particularly more effective than the commercial drug nevirapine against the resistant variants A17 and EFVR, and might thus have the potential to serve as a new anti-HIV lead. Unfortunately, there are still no reliable data on the mechanism of anti-HIV action of bichromonol. Since simple coumarins typically inhibit HIV RT, it was suggested that bichromonol is an HIV RT inhibitor. However, preliminarily enzymatic assay reveals no RT inhibitory activity of compound 33.
Figure 5.

Structure of bichromonol 33, a coumarin dimer.
For the anti-HIV activity of simple coumarins, the key factor is the presence of certain substituents. Since all simple coumarins inhibit HIV RT, a structure–activity relationship seems possible for them. Coumarins with only hydroxy or methoxy groups (compounds 1–3) tend to have low anti-HIV activity; O-terpenoid and C-monoterpenoid coumarins 5–8 and 9–18 often have acceptable characteristics, while diterpenes 19–28 mostly have high activity (EC50 reaches the nanomolar range). The most active diterpene coumarins contain isoprene substituents at the C3 and C6 positions.
In addition, simple diterpene coumarins have a low molecular weight, and their activity is tolerant to the introduction of polar substituents (compounds 22–27). Therefore, in our opinion, diterpene coumarins are promising lead compounds for developing new anti-HIV agents. In development, attention should probably be paid to unsaturated double bonds, which pose a potential problem in terms of drug stability and drug metabolism.
3. Furocoumarins
Furocoumarins are a class of compounds of natural and synthetic origin with various biological activities. Among natural furocoumarins, two classes of anti-HIV agents are distinguished: psoralene-type IIa and coumestan-type IIb (Figure 6). Due to the plane structure of psoralene, it intercalates into the DNA helix, forming strong bonds with nitrogenous bases [55]. This property is used in the photochemical treatment of skin diseases [55,56,57].
Figure 6.

Psoralene (IIa)- and coumestan (IIb)-type natural furocoumarins.
It was demonstrated that furocoumarins 34–43 inhibit HIV replication in H9 cells with an EC50 in the range of up to 0.37 μM (Table 3) [30,58]. Among them, the most active are psoralen 33 (EC50 = 0.54 and TI = 191), bergapten 35 (EC50 = 1.63 and TI = 70), imperatorin 36 (EC50 < 0.37 and TI > 1000) and heraclenol 40 (EC50 = 0.38 and TI = 870) [30,58]. Subsequently, Sancho and colleagues showed that imperatorin 36 showed no activity against RT and integrase [59]. However, it inhibits the critical transcription factor Sp1 and blocks infected cells in the G1 phase of the cell cycle. This indicates its potential therapeutic role in the treatment of HIV infections by inhibiting cellular factors that regulate HIV-1 replication at the transcriptional level [59].
Table 3.
Linear furocoumarins with anti-HIV activity.
| Compound | Plant Source | Anti-HIV Activity and Toxicity |
|---|---|---|
 Psoralen 34 |
Prangos tschimganica [58], Ruta graveolens [60], Dorstenia foetida [61] |
EC50 = 0.54 μM CC50 = 103 μM TI = 191 [58] |
 Bergapten 35 |
Zanthoxylum ailanthoides [23], Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62,63] |
EC50 = 1.63 μM CC50 = 115 μM TI = 69.9 [58] |
 Imperatorin 36 |
Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62], ,Clausena lansium [64], |
EC50 <0.37 μM CC50 > 370 μM TI > 1000 [30] |
 Oxypeucedanin 37 |
Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62] |
EC50 = 3.67 μM CC50 = 81.8 μM TI = 22.2 [30] |
 Heraclenin 38 |
Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62] |
EC50 = 8.29 μM CC50 > 70.3 μM TI = 8.48 [30] |
 Oxypeucedanin Hydrate 39 |
Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62] |
EC50 = 3.29 μM CC50 = 6.94 μM TI = 2.11 [30] |
 Heraclenol 40 |
Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62] |
EC50 = 0.38 μM CC50 = 329 μM TI = 870 [30] |
 Oxypeucedanin methylate 41 |
Ferula sumbul [30], Prangos tschimganica [58] |
EC50 = 105 μM CC50 = 314 μM TI = 3.00 [30] |
 Saxalin 42 |
Prangos tschimganica [58], Angelica officinalis [65] |
EC50 = 6.99 μM CC50 = 81.7 μM TI = 11.7 [58] |
 Pabulenol 43 |
Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62] |
EC50 = 22.3 μM CC50 = 58.4 μM TI = 2.61 [58] |
Very recently, it was demonstrated that prenylated coumarins clauselenins A and C 44 and 45 (Figure 7) isolated from stems of Clausena lenis inhibit HIV RT with a half-effective concentration (EC50) of 290 and 170 nM, respectively. These coumarins 44 and 45 also showed good half-maximal inhibition (IC50) values of 0.52 and 1.06 μM. In addition, clauselenins A and C were non-toxic to normal C8166 cells (CC50 > 200 μM), and TI50 was > 689 [48]. Therefore, these compounds can serve as new leads in the development of new antiviral agents.
Figure 7.

Structure of clauselenins A and C 44 and 45.
The second group of natural furocoumarins is the coumestans. They are mainly distributed in plants of the Fabaceae family [26,66,67,68,69] and have a wide spectrum of biological activity [70]. However, the antiviral activity of coumestans is poorly studied.
Wedelolactone 46 (Figure 8) isolated from Eclipta prostrate plants also demonstrated selective inhibition of HIV-1 integrase with IC50 = 4 μM [71]. It should be noted that wedelolactone is the only coumarin that has activity against HIV integrase. In addition, wedelolactone has weak protease inhibition activity (32.7% at 100 μM) [71].
Figure 8.

Structure of wedelolactone, a coumestan derivative.
Thus, among furocoumarins, linear coumarins are the largest class of anti-HIV compounds. The antiviral action is mediated by host cell cycle inhibition. The only example of integrase inhibitors is wedelolactone.
4. Pyranocoumarins
Pyranocoumarins are an important subclass of coumarins with a wide spectrum of biological activity, among which anti-HIV activity can be distinguished [72,73,74].
In this section, linear and angular pyranocoumarins are discussed.
Linear pyranocoumarins 47–49 (Figure 9) isolated from the stems of Clausena lenis [48] and Ficus nervosa [75] demonstrated high to moderate anti-HIV activity in the micromolar range (EC50 = 1.87–3.19 μM) and TI = 63–107 [54]. As demonstrated by the enzyme assay, the anti-HIV activity of these pyranocoumarins is mediated by RT inhibition [48].
Figure 9.

Linear pyranocoumarins with anti-HIV activity.
In addition to anti-HIV activity, these pyranocoumarins demonstrate antimycobacterial [75] and anti-inflammatory activity [48].
Angular pyranocoumarins comprise tetracyclic calanolide and tricyclic seseline-type scaffolds.
Among the natural pyranocoumarins, tetracyclic compounds isolated from tropical plants of the genus Calophyllum occupy a special place and include calanolide, inophyllum, and cordatolide [73,76]. Compounds 50–60 and their anti-HIV activity are discussed in detail in reviews [12,73,76,77,78]. The main results are depicted in Table 4. Note that calanolide A 50, inophyllum B 54, and inophyllum P 58 were identified as non-nucleoside inhibitors of HIV-1 RT with IC50 in the nanomolar range. In addition, calanolide A 50 demonstrated activity against a wide range of HIV strains, including azidotmidine- and pyridinone-resistant HIV strains [73,79,80,81,82]. Currently, calanolide A passed the first phase of clinical trials [12,83,84].
Table 4.
Compounds 50–60 with anti-HIV-1 activity.
| Compound | Inhibition of HIV-1 PRP, μM | Anti-HIV Activity in CEM SS Cells and Toxicity |
|---|---|---|
 calanolide A 50 |
IC50 = 0.07 [78] | IC50 = 0.1 μM CC50 = 20 μM TI = 200 [73,79] |
 calanolide B 51 |
IC50 = 0.4 μM CC50 = 15 μM TI = 37 [73,76,79] |
|
 calanolide C 52 |
no significant activity CC50 = 30 μM [79] |
|
 inophyllum A 53 |
IC50 = 30 μM [80] | |
 inophyllum B 54 |
IC50 = 0.038 [73] | IC50 = 1.4 μM CC50 = 55 μM TI = 39 [73,80] |
 inophyllum C 55 |
low activity CC50 = 18 μM [80] |
|
 inophyllum D 56 |
low activity CC50 = 15 μM [80] |
|
 inophyllum E 57 |
low activity CC50 = 6.2 μM [80] |
|
 inophyllum P 58 |
IC50 = 0.130 [73] | IC50 = 1.6 μM CC50 = 55 μM TI = 16 [80,81] |
 (+)-oxocalanolide 59 |
IC50 HIV-1 (IIIB) = 1.0 μM, CC50 HIV-1 (IIIB) > 10 μM IC50 HIV-1 (RF) = 0.9 μM, CC50 HIV-1 (RF) > 10 μM IC50 HIV-1 (SK1) = 0.17 μM, CC50 HIV-1 (SK1) > 10 μM IC50 HIV-2 (ROD) = 15.9 μM, CC50 HIV-1 (ROD) > 13.8 μM [81] |
|
 (–)-oxocalanolide 60 |
IC50 HIV-1 (IIIB)= 1.88 μM, CC50 HIV-1 (IIIB) > 10 μM IC50 HIV-1 (RF) = 3.41 μM, CC50 HIV-1 (RF) > 10 μM IC50 HIV-1 (SK1) = 0.27 μM, CC50 (SK1) = 55 μM [81] |
In their review [81], Yu and colleagues considered the structure–activity relationships (SARs) of natural compounds isolated from plants of the genus Calophyllum:
-
(1)
the presence of bulky substituents at the C(4) position;
-
(2)
positions C(10), C(11) and C(12) of the chromanol ring are sensitive to modification;
-
(3)
position C(12) must contain a hydrogen bond acceptor.
Subsequently, a systematic study of the SARs of calanolide A analogs led to more active derivatives than parent calanolide A [85,86]. Thus, it was demonstrated that 11-demethyl-12-oxo calanolide A 61 (Figure 10), possessing only one chiral center, has similar inhibitory activity against HIV-1 and a better therapeutic index (EC50 = 0.11 μM and TI = 21–169) [87]. 10-Bromomethyl-11-demethyl-12-oxo-calanolide A 62 [85] and 10-chloromethyl-11-demethyl-12-oxo-calanolide A 63 [86] (Figure 10) showed greater anti-HIV-1 activity at the nanomolar level (EC50 = 2.85 nM and TI > 10526 and EC50 = 7.4 nM and TI = 1417, respectively). Compounds 61–63 demonstrate no detectable toxicity [85]. In addition, compound 63 demonstrated a druggable profile with 32.7% oral bioavailability in rats and extremely high potency against the wild-type HIV-1 and Y181C single mutation of HIV-1 [86].
Figure 10.

Synthetic calanolide A analogs with anti-HIV activity.
Pyranocoumarin GUT-70 64 (Figure 11) was isolated from Calophyllum brasiliense stem bark [88,89]. It stabilized plasma membrane fluidity, inhibited HIV-1 entry, and down-regulated the expression of chemokine receptors CXCR4, CD4, and CCR5. GUT-70 also had an inhibitory effect on viral replication through the inhibition of NF-κB [90]. Therefore, this compound may be a dual-functional and viral mutation-resistant reagent. Another important feature of this compound is that GUT-70 inhibited HIV-1 replication in both acutely and chronically infected cells [91]. The antiviral activity (determined in several cell lines) and toxicity (MTT assay) of GUT-70 are presented in Table 5. Unfortunately, despite several mechanisms of action, this compound is characterized by low therapeutic indices.
Figure 11.

Structure of the anti-HIV pyranocoumarin GUT-70.
Table 5.
Antiviral activity and toxicity of GUT-70.
| Cell Line | CC50, μM | EC50, μM | TI (CC50/EC50) |
|---|---|---|---|
| U1 (PMA) | 8.44 | 3.48 | 2.43 |
| U1 (TNF-α) | 8.44 | 4.32 | 1.95 |
| Molt-4 | >10 | 3.41 | >2.93 |
| TZM-bl | >10 | – | – |
Reprinted/adapted with permission from Ref. [91]. 2013, Elsevier.
The seseline scaffold is presented by seselin and suksdorfin-type coumarin, its oxygenated counterpart.
The parent compound, seselin 65 shows moderate anti-HIV activity with EC50 = 25.5 μM and CC50 = 329.0 μM (TI = 12.9) [22].
Lee and colleagues [72] isolated suksdorfin 66 from the fruit Lomatium Suksdorfii, which demonstrated inhibition of HIV-1 replication in H9 cells with EC50 = 1.3 μM and TI = 79. Subsequently, a number of suksdorfin 3’,4’-analogs were synthesized, among which 3’,4’-di-O-camphanoyl-cis-khellactone (DCK) 67 inhibits HIV-1 replication in H9 cells with EC50 = 0.41 nM and TI > 78,000 [74]. Subsequently, the SAR DCK 67 was investigated, which was considered in reviews [12,81]. Table 6 shows anti-HIV active compounds 66–67 (Figure 12) in H9 cells.
Table 6.
Anti-HIV activity of suksdorfin and DCK analogs in H9 cells.
| Compound | Anti-HIV Activity, EC50 μM (TI) |
Compound | Anti-HIV Activity, EC50 μM (TI) |
|---|---|---|---|
| Seselin 65 | 25.5 (12.9) [22] | 68c | 2.4 × 10–7 (4 × 108) [92] |
| Suksdorfin 66 | 1.3 (79) [72] | 68d | 7.2 × 10–6 (2.1 × 107) [93] |
| DCK 67 | 2.5 × 10–4 (1.4 × 105) [92] | 68e | 2.4 × 10–3 (1.5 × 104) [94] |
| 68a | 5.3 × 10–5 (2 × 106) [92] | 68f | 4 × 10–3 (6 × 103) [94] |
| 68b | 1.8 × 10–6 (6.9 × 107) [92] |
Figure 12.

Structures of seselin 65, suksdorfin 66, DCK 67, and its analogues 68.
Among the modified compounds 67 and 68, 3-cyanomethyl-DCK 68e showed weak toxicity (IC50 > 37 μM) against HIV-1IIIB in H9 cells. In addition, 68e has anti-HIV activity against wild-type and drug-resistant viral infections in CD4+ T cell lines as well as in peripheral blood mononuclear cells. Preclinical trials have shown that compound 68e has moderate oral bioavailability in rats and moderate cell permeability [93].
Thus, among linear and angular pyranocoumarins, the latter are the most promising in terms of developing anti-HIV agents. These compounds are characterized by good antiviral activity, but they have some solubility problems, which seem to be due to the presence of a condensed ring system.
Prenyl derivatives of simple coumarins, pyranocoumarins and furanocoumarins, are genetically related compounds since the biosynthesis of pyranocoumarins and furanocoumarins occurs through cyclization of the isoprene fragment with the aromatic cycle [95,96]. Furthermore, the mode of action of anti-HIV coumarins is often carried out through identical targets, such as RT inhibition.
The unique structure of coumarins determines their low toxicity compared to other heterocycles and, in combination with effective protocols for coumarin synthesis, provides good potential for developing anti-HIV drugs based on the coumarin scaffold.
5. Conclusions
Among natural coumarins, there are antiviral compounds with activity against viral enzymes, reverse transcriptase, and integrase. While several types of coumarins (simple coumarins, pyranocoumarins, and furanocoumarins) are active against reverse transcriptase, the only known coumarin with anti-integrase activity is wedelolactone. In addition, simple and angular coumarins are capable of inhibiting cellular factors that regulate HIV-1 replication. Finally, simple coumarins can block the transmission of viral particles from infected macrophages to healthy cells.
Among the anti-HIV coumarin derivatives, the pyranocoumarins—represented by the calanolide and suksdorfin families, as well as simple coumarin C3,C6-diterpenoids—are of the greatest interest. These compounds are characterized by effective inhibition of immunodeficiency virus replication in the nanomolar range, and, in addition, some of them act by several mechanisms. A slight tuning of the structure of the natural compounds allows for obtaining even more active compounds with a therapeutic index of up to 108 for the suksdorfin derivatives.
We reviewed the information on the anti-HIV properties of simple coumarins which allows for the consideration of the coumarin cycle as a privileged framework for the search and further development of anti-HIV compounds. Nevertheless, the unique properties are not limited to anti-HIV activities, but they can also target other RNA-based viruses, among which SARS-CoV-2 currently attracts the most significant attention [97].
Author Contributions
Conceptualization, R.F.F.; writing—original draft preparation, A.D.S.; writing—review and editing, I.A.K.; supervision, O.N.C., G.V.Z. and M.V.T. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Funding Statement
This research was funded by the Ministry of Science and Higher Education of the Russian Federation grant 075-15-2022-1118 (Section “Simple coumarins”) and the Russian Science Foundation, grant number 21-13-00382 (Sections “Furocoumarins” and “Pyranocoumarins”).
Footnotes
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
References
- 1.Abraham K., Wöhrlin F., Lindtner O., Heinemeyer G., Lampen A. Toxicology and risk assessment of coumarin: Focus on human data. Mol. Nutr. Food Res. 2010;54:228–239. doi: 10.1002/mnfr.200900281. [DOI] [PubMed] [Google Scholar]
- 2.Heghes S.C., Vostinaru O., Mogosan C., Miere D., Iuga C.A., Filip L. Safety Profile of Nutraceuticals Rich in Coumarins: An Update. Front. Pharmacol. 2022;13:803338. doi: 10.3389/fphar.2022.803338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lake B.G. Coumarin metabolism, toxicity and carcinogenicity. Food Chem. Toxicol. 1999;37:423–453. doi: 10.1016/S0278-6915(99)00010-1. [DOI] [PubMed] [Google Scholar]
- 4.Vella S., Schwartländer B., Sow S.P., Eholie S.P., Murphy R.L. The history of antiretroviral therapy and of its implementation in resource-limited areas of the world. AIDS. 2012;26:1231–1241. doi: 10.1097/QAD.0b013e32835521a3. [DOI] [PubMed] [Google Scholar]
- 5.Al-Abbasa N.S., Shaer N.A. Combination of coumarin and doxorubicin induces drug-resistant acute myeloid leukemia cell death. Heliyon. 2021;7:e06255. doi: 10.1016/j.heliyon.2021.e06255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zou J., Liu Y., Guo R., Tang Y., Shi Z., Zhang M., Wu W., Chen Y., Hou K. An In Vitro Coumarin-Antibiotic Combination Treatment of Pseudomonas aeruginosa Biofilms. Nat. Prod. Commun. 2021;16:1–7. doi: 10.1177/1934578X20987744. [DOI] [Google Scholar]
- 7.Ghosh A.K., Osswald H.L., Prato G. Recent Progress in the Development of HIV-1 Protease Inhibitors for the Treatment of HIV/AIDS. J. Med. Chem. 2016;59:5172–5208. doi: 10.1021/acs.jmedchem.5b01697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sharapov A.D., Fatykhov R.F., Khalymbadzha I.A., Sharutin V.V., Santra S., Zyryanov G.V., Chupakhin O.N., Ranu B.C. Mechanochemical synthesis of coumarins via Pechmann condensation under solvent-free conditions: An easy access to coumarins and annulated pyrano-[2,3-f] and [3,2-f]indoles. Green Chem. 2022;24:2429–2437. doi: 10.1039/D1GC04564D. [DOI] [Google Scholar]
- 9.Lončarić M., Gašo-Sokač D., Jokić S., Molnar M. Recent Advances in the Synthesis of Coumarin Derivatives from Different Starting Materials. Biomolecules. 2020;10:151. doi: 10.3390/biom10010151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Serna-Arbeláez M.S., Florez-Sampedro L., Orozco L.P., Ramírez K., Galeano E., Zapata W. Natural Products with Inhibitory Activity against Human Immunodeficiency Virus Type 1. Adv. Virol. 2021;2021:5552088. doi: 10.1155/2021/5552088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cos P., Maes L., Vlietinck A., Pieters L. Plant-Derived Leading Compounds for Chemotherapy of Human Immunodefiency Virus (HIV) Infection—An Update (1998–2007) Planta Med. 2008;74:1323–1337. doi: 10.1055/s-2008-1081314. [DOI] [PubMed] [Google Scholar]
- 12.Kostova I., Raleva S., Genova P., Argirova R. Structure-Activity Relationships of Synthetic Coumarins as HIV-1 Inhibitors. Bioinorg. Chem. Appl. 2006;2006:68274. doi: 10.1155/BCA/2006/68274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Engelman A., Cherepanov P. The structural biology of HIV-1: Mechanistic and therapeutic insights. Nat. Rev. Genet. 2012;10:279–290. doi: 10.1038/nrmicro2747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Arts E.J., Hazuda D.J. HIV-1 Antiretroviral Drug Therapy. Cold Spring Harb. Perspect. Med. 2012;2:a007161. doi: 10.1101/cshperspect.a007161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Fobofou S.A.T., Franke K., Brandt W., Manzin A., Madeddu S., Serreli G., Sanna G., Wessjohann L.A. Bichromonol, a dimeric coumarin with anti-HIV activity from the stem bark of Hypericum roeperianum. Nat. Prod. Res. 2022:1–7. doi: 10.1080/14786419.2022.2110094. [DOI] [PubMed] [Google Scholar]
- 16.Venugopala K.N., Rashmi V., Odhav B. Review on Natural Coumarin Lead Compounds for Their Pharmacological Activity. BioMed. Res. Int. 2013;2013:963248. doi: 10.1155/2013/963248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Galabov A.S., Iosifova T., Vassileva E., Kostova I. Antiviral Activity of Some Hydroxycoumarin Derivatives. Z. Für Nat. C. 1996;51:558–562. doi: 10.1515/znc-1996-7-815. [DOI] [PubMed] [Google Scholar]
- 18.Bayoumi S.A., Rowan M.G., Beeching J.R., Blagbrough I.S. Constituents and secondary metabolite natural products in fresh and deteriorated cassava roots. Phytochemistry. 2010;71:598–604. doi: 10.1016/j.phytochem.2009.10.012. [DOI] [PubMed] [Google Scholar]
- 19.Wu T.-S., Tsang Z.-J., Wu P.-L., Lin F.-W., Li C.-Y., Teng C.-M., Lee K.-H. New constituents and antiplatelet aggregation and anti-HIV principles of Artemisia capillaris. Bioorganic Med. Chem. 2001;9:77–83. doi: 10.1016/S0968-0896(00)00225-X. [DOI] [PubMed] [Google Scholar]
- 20.Kuo P.-C., Hwang T.-L., Lin Y.-T., Kuo Y.-C., Leu Y.-L. Chemical constituents from Lobelia chinensis and their anti-virus and anti-inflammatory bioactivities. Arch. Pharmacal Res. 2011;34:715–722. doi: 10.1007/s12272-011-0503-7. [DOI] [PubMed] [Google Scholar]
- 21.Chan Y.-Y., Li C.-H., Shen Y.-C., Wu T.-S. Anti-inflammatory Principles from the Stem and Root Barks of Citrus medica. Chem. Pharm. Bull. 2010;58:61–65. doi: 10.1248/cpb.58.61. [DOI] [PubMed] [Google Scholar]
- 22.Feng T., Wang R.-R., Cai X.-H., Zheng Y.-T., Luo X.-D. Anti-human Immunodeficiency Virus-1 Constituents of the Bark of Poncirus trifoliata. Chem. Pharm. Bull. 2010;58:971–975. doi: 10.1248/cpb.58.971. [DOI] [PubMed] [Google Scholar]
- 23.Cheng M.-J., Lee K.-H., Tsai I.-L., Chen I.-S. Two new sesquiterpenoids and anti-HIV principles from the root bark of Zanthoxylum ailanthoides. Bioorganic Med. Chem. 2005;13:5915–5920. doi: 10.1016/j.bmc.2005.07.050. [DOI] [PubMed] [Google Scholar]
- 24.Likhitwitayawuid K., Supudompol B., Sritularak B., Lipipun V., Rapp K., Schinazi R.F. Phenolics with Anti-HSV and Anti-HIV Activities from Artocarpus gomezianus., Mallotus pallidus., and Triphasia trifolia. Pharm. Biol. 2005;43:651–657. doi: 10.1080/13880200500383058. [DOI] [Google Scholar]
- 25.Epifano F., Genovese S., Marcotullio M., Curini M. Chemistry and Pharmacology of Collinin, Active Principle of Zanthoxylum spp. Mini-Rev. Med. Chem. 2008;8:1203–1208. doi: 10.2174/138955708786141089. [DOI] [PubMed] [Google Scholar]
- 26.Adianti M., Aoki C., Komoto M., Deng L., Shoji I., Wahyuni T.S., Lusida M.I., Fuchino S.H., Kawahara N., Hotta H. Anti-hepatitis C virus compounds obtained from Glycyrrhiza uralensis and other Glycyrrhiza species. Microbiol. Immunol. 2014;58:180–187. doi: 10.1111/1348-0421.12127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chang C.-T., Doong S.-L., Tsai I.-L., Chen I.-S. Coumarins and anti-HBV constituents from Zanthoxylum schinifolium. Phytochemistry. 1997;45:1419–1422. doi: 10.1016/S0031-9422(97)89023-1. [DOI] [PubMed] [Google Scholar]
- 28.Lv H.-N., Wang S., Zeng K.-W., Li J., Guo X.-Y., Ferreira D., Zjawiony J.K., Tu P.-F., Jiang Y. Anti-inflammatory Coumarin and Benzocoumarin Derivatives from Murraya alata. J. Nat. Prod. 2015;78:279–285. doi: 10.1021/np500861u. [DOI] [PubMed] [Google Scholar]
- 29.Lee C.-L., Chiang L.-C., Cheng L.-H., Liaw C.-C., El-Razek M.H.A., Chang F.-R., Wu Y.-C. Influenza A (H(1)N(1)) Antiviral and Cytotoxic Agents from Ferula assa-foetida. J. Nat. Prod. 2009;72:1568–1572. doi: 10.1021/np900158f. [DOI] [PubMed] [Google Scholar]
- 30.Zhou P., Takaishi Y., Duan H., Chen B., Honda G., Itoh M., Takeda Y., Kodzhimatov O.K., Lee K.-H. Coumarins and bicoumarin from Ferula sumbul: Anti-HIV activity and inhibition of cytokine release. Phytochemistry. 2000;53:689–697. doi: 10.1016/S0031-9422(99)00554-3. [DOI] [PubMed] [Google Scholar]
- 31.Ngadjui T.B., Ayafor J.F., Sondengam B.L., Connolly J.D. Coumarins from Clausena anisate. Phytochemistry. 1989;28:585–589. doi: 10.1016/0031-9422(89)80056-1. [DOI] [Google Scholar]
- 32.Hatano T., Yasuhara T., Miyamoto K., Okuda T. Anti-human immunodeficiency virus phenolics from licorice. Chem. Pharm. Bull. 1988;36:2286–2288. doi: 10.1248/cpb.36.2286. [DOI] [PubMed] [Google Scholar]
- 33.Fortin H., Tomasi S., Jaccard P., Robin V., Boustie J. A Prenyloxycoumarin from Psiadia dentata. Chem. Pharm. Bull. 2001;49:619–621. doi: 10.1248/cpb.49.619. [DOI] [PubMed] [Google Scholar]
- 34.Kasaian J., Mohammadi A. Biological activities of farnesiferol C: A review. J. Asian Nat. Prod. Res. 2017;20:27–35. doi: 10.1080/10286020.2017.1379997. [DOI] [PubMed] [Google Scholar]
- 35.Sistani P., Dehghan G., Sadeghi L. Structural and kinetic insights into HIV-1 reverse transcriptase inhibition by farnesiferol C. Int. J. Biol. Macromol. 2021;174:309–318. doi: 10.1016/j.ijbiomac.2021.01.173. [DOI] [PubMed] [Google Scholar]
- 36.Turabelidze D.G., Kemertelidze É.P. Farnesiferol C from the roots of Ferula szovitsiana. Chem. Nat. Compd. 1976;12:589–589. doi: 10.1007/BF00565199. [DOI] [Google Scholar]
- 37.Kamilov K.M., Nikonov G.K. Coumarins of Ferula kopetdaghensis and the structure of kopeolin and kopeoside. Chem. Nat. Compd. 1973;9:294–298. doi: 10.1007/BF00565684. [DOI] [Google Scholar]
- 38.Akihara Y., Ohta E., Nehira T., Ômura H., Ohta S. New Prenylated ortho-Dihydroxycoumarins from the Fruits of Ficus nipponica. Chem. Biodivers. 2017;14:e1700196. doi: 10.1002/cbdv.201700196. [DOI] [PubMed] [Google Scholar]
- 39.Wu T.S., Hsu M.Y., Kuo P.C., Sreenivasulu B., Damu A.G., Su C.R., Li C.Y., Chang H.C. Constituents from the leaves of Phellodendron amurense var. wilsonii and their bioactivity. J. Nat. Prod. 2003;66:1207–1211. doi: 10.1021/np030034v. [DOI] [PubMed] [Google Scholar]
- 40.Nakamori T., Taniguchi M., Shibano M., Wang N.-H., Baba K. Chemical studies on the root of Heracleum candicans WALL. J. Nat. Med. 2008;62:403–412. doi: 10.1007/s11418-008-0266-8. [DOI] [PubMed] [Google Scholar]
- 41.Liu Y.-P., Yan G., Guo J.-M., Liu Y.-Y., Li Y.-J., Zhao Y.-Y., Qiang L., Fu Y.-H. Prenylated Coumarins from the Fruits of Manilkara zapota with Potential Anti-inflammatory Effects and Anti-HIV Activities. J. Agric. Food Chem. 2019;67:11942–11947. doi: 10.1021/acs.jafc.9b04326. [DOI] [PubMed] [Google Scholar]
- 42.Gonzalez A.G., Reyes R.E., Espino M.R. Two new coumarins from Ruta pinnata. Phytochemistry. 1977;16:2033–2035. doi: 10.1016/0031-9422(77)80126-X. [DOI] [Google Scholar]
- 43.Atta-ur-Rahman, Sultana N., Khan M.R., Choudhary M.I. Triterpene and coumarins from Skimmia laureola. Nat. Prod. Lett. 2002;16:305–313. doi: 10.1080/10575630290020613. [DOI] [PubMed] [Google Scholar]
- 44.Rajkumar P., Sundari S., Selvaraj S., Natarajan A., Suganya R., Jayaprakash R., Kasthuri K., Kumaresan S. GC-MS, Phytochemical Analysis and In Silico Approaches of a Medicinal Plant Acalypha indica. J. Sci. Res. 2022;14:671–684. doi: 10.3329/jsr.v14i2.56648. [DOI] [Google Scholar]
- 45.Tao L., Zhuo Y.-T., Qiao Z.-H., Li J., Tang H.-X., Yu Q.-M., Liu Y.-Y., Liu Y.-P. Prenylated coumarins from the fruits of Artocarpus heterophyllus with their potential anti-inflammatory and anti-HIV activities. Nat. Prod. Res. 2022;36:2526–2533. doi: 10.1080/14786419.2021.1913590. [DOI] [PubMed] [Google Scholar]
- 46.Halstead C.W., Forster P.I., Waterman P.G. Novel Metabolites from the Stem Bark of Brombya sp. Nova (Gap Creek) (Rutaceae) Nat. Prod. Commun. 2006;1:351–356. doi: 10.1177/1934578X0600100502. [DOI] [Google Scholar]
- 47.Guz N.R., Lorenz P., Stermitz F.R. New coumarins from Harbouria trachypleura: Isolation and synthesis. Tetrahedron Lett. 2001;42:6491–6494. doi: 10.1016/S0040-4039(01)01355-7. [DOI] [Google Scholar]
- 48.Liu Y.-P., Yan G., Xie Y.-T., Lin T.-C., Zhang W., Li J., Wu Y.-J., Zhou J.-Y., Fu Y.-H. Bioactive prenylated coumarins as potential anti-inflammatory and anti-HIV agents from Clausena lenis. Bioorganic Chem. 2020;97:103699. doi: 10.1016/j.bioorg.2020.103699. [DOI] [PubMed] [Google Scholar]
- 49.Sun M., Sun M., Zhang J. Osthole: An overview of its sources, biological activities, and modification development. Med. Chem. Res. 2021;30:1767–1794. doi: 10.1007/s00044-021-02775-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zafar S., Sarfraz I., Rasul A., Shah M.A., Hussain G., Zahoor M.K., Shafiq N., Riaz A., Selamoglu Z., Sarker S.D. Osthole: A Multifunctional Natural Compound with Potential Anticancer, Antioxidant and Anti-inflammatory Activities. Mini-Rev. Med. Chem. 2021;21:2747–2763. doi: 10.2174/1389557520666200709175948. [DOI] [PubMed] [Google Scholar]
- 51.Kumar V., Vallipuram K., Adebajo A.C., Reisch J. 2,7-dihydroxy-3-formyl-1-(3′-methyl-2′-butenyl) carbazole from Clausena lansium. Phytochemistry. 1995;40:1563−1565. doi: 10.1016/0031-9422(95)00452-D. [DOI] [Google Scholar]
- 52.Fernandes T.S., Copetti D., Carmo G.D., Neto A.T., Pedroso M., Silva U.F., Mostardeiro M.A., Burrow R.E., Dalcol I.I., Morel A.F. Phytochemical analysis of bark from Helietta apiculata Benth and antimicrobial activities. Phytochemistry. 2017;141:131–139. doi: 10.1016/j.phytochem.2017.05.017. [DOI] [PubMed] [Google Scholar]
- 53.González A., Estévez R., Jaraiz I. Structure of sabandinin and other coumarins isolated from the roots of Ruta pinnata. Phytochemistry. 1971;10:1621–1626. doi: 10.1016/0031-9422(71)85036-7. [DOI] [Google Scholar]
- 54.Bedoya L.M., Beltrán M., Sancho R., Olmedo D.A., Sánchez-Palomino S., del Olmo E., López-Pérez J.L., Muñoz E., San Feliciano A., Alcamí J. 4-Phenylcoumarins as HIV transcription inhibitors. Bioorg. Med. Chem. Lett. 2005;15:4447–4450. doi: 10.1016/j.bmcl.2005.07.041. [DOI] [PubMed] [Google Scholar]
- 55.El-Gogary T.M., Koehler G.J. Interaction of psoralens with DNA-bases (II): An ab initio quantum chemical, density functional theory and second-order MØller-Plesset perturbational study. Mol. Struc. THEOCHEM. 2009;895:57–64. doi: 10.1016/j.theochem.2008.10.012. [DOI] [Google Scholar]
- 56.El-Gogary T.M. Interaction of angelicin with DNA-bases (III) DFT and ab initio second-order Moeller-Plesset study. Int. J. Quantum Chem. 2009;110:1445–1454. doi: 10.1002/qua.22250. [DOI] [Google Scholar]
- 57.Stern R.S., Nichols K.T., Väkevä L.H. Malignant Melanoma in Patients Treated for Psoriasis with Methoxsalen (Psoralen) and Ultraviolet A Radiation (PUVA) N. Engl. J. Med. 1997;336:1041–1045. doi: 10.1056/NEJM199704103361501. [DOI] [PubMed] [Google Scholar]
- 58.Shikishima Y., Takaishi Y., Honda G., Ito M., Takeda Y., Kodzhimatov O.K., Ashurmetov O., Lee K.-H. Chemical Constituents of Prangos tschimganica; Structure Elucidation and Absolute Configuration of Coumarin and Furanocoumarin Derivatives with Anti-HIV Activity. Chem. Pharm. Bull. 2001;49:877–880. doi: 10.1248/cpb.49.877. [DOI] [PubMed] [Google Scholar]
- 59.Sancho R., Márquez N., Gómez-Gonzalo M., Calzado M.A., Bettoni G., Coiras M., Alcami J., López-Cabrera M., Appendino G.B., Muñoz E. Imperatorin Inhibits HIV-1 Replication through an Sp1-dependent Pathway. J. Biol. Chem. 2004;279:37349–37359. doi: 10.1074/jbc.M401993200. [DOI] [PubMed] [Google Scholar]
- 60.Nebo L., Varela R.M., Molinillo J.M.G., Sampaio O.M., Severino V.G., Cazal C.M., Fernandes M.F.D.G., Fernandes J.B., Macías F.A. Phytotoxicity of alkaloids, coumarins and flavonoids isolated from 11 species belonging to the Rutaceae and Meliaceae families. Phytochem. Lett. 2014;8:226–232. doi: 10.1016/j.phytol.2014.02.010. [DOI] [Google Scholar]
- 61.Heinke R., Franke K., Porzel A., Wessjohann L.A., Awadh A.N.A., Schmidt J. Furanocoumarins from Dorstenia foetida. Phytochemistry. 2011;72:929–934. doi: 10.1016/j.phytochem.2011.03.008. [DOI] [PubMed] [Google Scholar]
- 62.Ban H.S., Lim S.S., Suzuki K., Jung S.H., Lee S., Lee Y.S., Shin K.H., Ohuchi K. Inhibitory Effects of Furanocoumarins Isolated from the Roots of Angelica dahurica on Prostaglandin E2 Production. Planta Med. 2003;69:408–412. doi: 10.1055/s-2003-39702. [DOI] [PubMed] [Google Scholar]
- 63.Kostova I., Ivanova A., Mikhova B., Klaiber I. Alkaloids and Coumarins from Ruta graveolens. Monatsh. Chem. 1999;130:703–707. doi: 10.1007/PL00010251. [DOI] [Google Scholar]
- 64.Liu H., Li F., Li C.-J., Yang J.-Z., Li L., Chen N.-H., Zhang D.-M. Bioactive furanocoumarins from stems of Clausena lansium. Phytochemistry. 2014;107:141–147. doi: 10.1016/j.phytochem.2014.08.002. [DOI] [PubMed] [Google Scholar]
- 65.Harkar S., Razdan T., Waight E. Steroids, chromone and coumarins from angelica officinalis. Phytochemistry. 1984;23:419–426. doi: 10.1016/S0031-9422(00)80344-1. [DOI] [Google Scholar]
- 66.Liu X., Nam J.-W., Song Y.S., Viswanath A.N.I., Pae A.N., Kil Y.-S., Kim H.-D., Park J.H., Seo E.-K., Chang M. Psoralidin, a coumestan analogue, as a novel potent estrogen receptor signaling molecule isolated from Psoralea corylifolia. Bioorganic Med. Chem. Lett. 2014;24:1403–1406. doi: 10.1016/j.bmcl.2014.01.029. [DOI] [PubMed] [Google Scholar]
- 67.Jang D.S., Kim J.M., Lee Y.M., Kim Y.S., Kim J.-H., Kim J.S. Puerariafuran, a New Inhibitor of Advanced Glycation End Products (AGEs) Isolated from the Roots of Pueraria lobata. Chem. Pharm. Bull. 2006;54:1315–1317. doi: 10.1248/cpb.54.1315. [DOI] [PubMed] [Google Scholar]
- 68.Yadav P.P., Ahmad G., Maurya R. Furanoflavonoids from Pongamia pinnata fruits. Phytochemistry. 2004;65:439–443. doi: 10.1016/j.phytochem.2003.09.011. [DOI] [PubMed] [Google Scholar]
- 69.Macías F.A., Simonet A.M., Galindo J.C., Castellano D. Bioactive phenolics and polar compounds from Melilotus messanensis. Phytochemistry. 1999;50:35–46. doi: 10.1016/S0031-9422(98)00453-1. [DOI] [Google Scholar]
- 70.Tuskaev V.A. Synthesis and biological activity of coumestan derivatives (Review) Pharm. Chem. J. 2013;47:1–11. doi: 10.1007/s11094-013-0886-5. [DOI] [Google Scholar]
- 71.Tewtrakul S., Subhadhirasakul S., Cheenpracha S., Karalai C. HIV-1 protease and HIV-1 integrase inhibitory substances from Eclipta prostrata. Phytotherapy Res. 2007;21:1092–1095. doi: 10.1002/ptr.2252. [DOI] [PubMed] [Google Scholar]
- 72.Lee T.T.-Y., Kashiwada Y., Huang L., Snider J., Cosentino M., Lee K.-H. Suksdorfin: An anti-HIV principle from Lomatium suksdorfii, its structure-activity correlation with related coumarins, and synergistic effects with anti-AIDS nucleosides. Bioorganic Med. Chem. 1994;2:1051–1056. doi: 10.1016/S0968-0896(00)82054-4. [DOI] [PubMed] [Google Scholar]
- 73.Ishikawa T. Anti HIV-1 Active Calophyllum Coumarins: Distribution, Chemistry, and Activity. Heterocycles. 2000;53:453–474. doi: 10.3987/REV-99-526. [DOI] [Google Scholar]
- 74.Huang L., Kashiwada Y., Cosentino L.M., Fan S., Lee K.-H. 3′,4′-Di-o-(−)-camphanoyl-(+)-ciskhellactone and related compounds: A. new class of potent anti-HIV agents. Bioorg. Med. Chem. Lett. 1994;4:593–598. doi: 10.1016/S0960-894X(01)80161-X. [DOI] [Google Scholar]
- 75.Chen L.-W., Cheng M.-J., Peng C.-F., Chen I.-S. Secondary Metabolites and Antimycobacterial Activities from the Roots of Ficus nervosa. Chem. Biodivers. 2010;7:1814–1821. doi: 10.1002/cbdv.200900227. [DOI] [PubMed] [Google Scholar]
- 76.Su X.-H., Zhang M.-L., Li L.-G., Huo C.-H., Gu Y.-C., Shi Q.-W. Chemical Constituents of the Plants of the Genus Calophyllum. Chem. Biodivers. 2008;5:2579–2608. doi: 10.1002/cbdv.200890215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Flavin M.T., Rizzo J.D., Khilevich A., Kucherenko A., Sheinkman A.K., Vilaychack V., Lin L., Chen W., Greenwood E.M., Pengsuparp T., et al. Synthesis, Chromatographic Resolution, and Anti-Human Immunodeficiency Virus Activity of (±)-Calanolide A and Its Enantiomers. J. Med. Chem. 1996;39:1303–1313. doi: 10.1021/jm950797i. [DOI] [PubMed] [Google Scholar]
- 78.Matthée G., Wright A.D., König G.M. HIV Reverse Transcriptase Inhibitors of Natural Origin. Planta Med. 1999;65:493–506. doi: 10.1055/s-1999-14004. [DOI] [PubMed] [Google Scholar]
- 79.Kashman Y., Gustafson K.R., Fuller R.W., Ii J.H.C., McMahon J.B., Currens M.J., Buckheit R.W., Hughes S.H., Cragg G.M., Boyd M.R. HIV inhibitory natural products. Part 7. The calanolides, a novel HIV-inhibitory class of coumarin derivatives from the tropical rainforest tree, Calophyllum lanigerum. J. Med. Chem. 1992;35:2735–2743. doi: 10.1021/jm00093a004. [DOI] [PubMed] [Google Scholar]
- 80.Patil A.D., Freyer A.J., Eggleston D.S., Haltiwanger R.C., Bean M.F., Taylor P.B., Caranfa M.J., Breen A.L., Bartus H.R. The inophyllums, novel inhibitors of HIV-1 reverse transcriptase isolated from the Malaysian tree, Calophyllum inophyllum Linn. J. Med. Chem. 1993;36:4131–4138. doi: 10.1021/jm00078a001. [DOI] [PubMed] [Google Scholar]
- 81.Yu D., Suzuki M., Xie L., Morris-Natschke S.L., Lee K.-H. Recent progress in the development of coumarin derivatives as potent anti-HIV agents. Med. Res. Rev. 2003;23:322–345. doi: 10.1002/med.10034. [DOI] [PubMed] [Google Scholar]
- 82.Cardellina J.H., Bokesch H.R., McKee T.C., Boyd M.R. Resolution and comparative anti-HIV evaluation of the enantiomers of calanolides A and B. Bioorganic Med. Chem. Lett. 1995;5:1011–1014. doi: 10.1016/0960-894X(95)00158-P. [DOI] [Google Scholar]
- 83.Usach I., Melis V., Peris J.-E. Non-nucleoside reverse transcriptase inhibitors: A review on pharmacokinetics, pharmacodynamics, safety and tolerability. J. Int. AIDS Soc. 2013;16:18567. doi: 10.7448/IAS.16.1.18567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Creagh T., Ruckle J.L., Tolbert D.T., Giltner J., Eiznhamer D.A., Dutta B., Flavin M.T., Xu Z.-Q. Safety and Pharmacokinetics of Single Doses of (+)-Calanolide A, a Novel, Naturally Occurring Nonnucleoside Reverse Transcriptase Inhibitor, in Healthy, Human Immunodeficiency Virus-Negative Human Subjects. Antimicrob. Agents Chemother. 2001;45:1379–1386. doi: 10.1128/AAC.45.5.1379-1386.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Ma T., Liu L., Xue H., Han C., Wang L., Chen Z., Liu G. Chemical Library and Structure–Activity Relationships of 11-Demethyl-12-oxo Calanolide A Analogues as Anti-HIV-1 Agents. J. Med. Chem. 2008;51:1432–1446. doi: 10.1021/jm701405p. [DOI] [PubMed] [Google Scholar]
- 86.Xue H., Lu X., Zheng P., Liu L., Han C., Hu J., Liu Z., Ma T., Li Y., Wang L., et al. Highly Suppressing Wild-Type HIV-1 and Y181C Mutant HIV-1 Strains by 10-Chloromethyl-11-demethyl-12-oxo-calanolide A with Druggable Profile. J. Med. Chem. 2010;53:1397–1401. doi: 10.1021/jm901653e. [DOI] [PubMed] [Google Scholar]
- 87.Wang L., Chen H.S., Tao P.Z., Li Y., Bai Y., Hu J.P., Ma T., Xing Z.T., Peng Z.G., Zhou C.M., et al. Improved synthesis and pharmaceutical evaluation of racemic 11-demethyl calanolide A. Acta Pharm. Sin. 2008;43:707–718. [PubMed] [Google Scholar]
- 88.Kimura S., Ito C., Jyoko N., Segawa H., Kuroda J., Okada M., Adachi S., Nakahata T., Yuasa T., Filho V.C., et al. Inhibition of leukemic cell growth by a novel anti-cancer drug (GUT-70) from Calophyllum Brasiliense that acts by induction of apoptosis. Int. J. Cancer. 2004;113:158–165. doi: 10.1002/ijc.20505. [DOI] [PubMed] [Google Scholar]
- 89.Ito C., Itoigawa M., Mishina Y., Filho V.C., Enjo F., Tokuda H., Nishino H., Furukawa H. Chemical Constituents of Calophyllum brasiliense. 2. Structure of Three New Coumarins and Cancer Chemopreventive Activity of 4-Substituted Coumarins. J. Nat. Prod. 2003;66:368–371. doi: 10.1021/np0203640. [DOI] [PubMed] [Google Scholar]
- 90.Matsuda K., Hattori S., Kariya R., Komizu Y., Kudo E., Goto H., Taura M., Ueoka R., Kimura S., Okada S. Inhibition of HIV-1 entry by the tricyclic coumarin GUT-70 through the modification of membrane fluidity. Biochem. Biophys. Res. Commun. 2015;457:288–294. doi: 10.1016/j.bbrc.2014.12.102. [DOI] [PubMed] [Google Scholar]
- 91.Kudo E., Taura M., Matsuda K., Shimamoto M., Kariya R., Goto H., Hattori S., Kimura S., Okada S. Inhibition of HIV-1 replication by a tricyclic coumarin GUT-70 in acutely and chronically infected cells. Bioorganic Med. Chem. Lett. 2012;23:606–609. doi: 10.1016/j.bmcl.2012.12.034. [DOI] [PubMed] [Google Scholar]
- 92.Xie L., Takeuchi Y., Cosentino L.M., Lee K.-H. Anti-AIDS Agents. 37. Synthesis and Structure−Activity Relationships of (3′R,4′R)-(+)-cis-Khellactone Derivatives as Novel Potent Anti-HIV Agents. J. Med. Chem. 1999;42:2662–2672. doi: 10.1021/jm9900624. [DOI] [PubMed] [Google Scholar]
- 93.Xie L., Takeuchi Y., Cosentino L.M., McPhail A.T., Lee K.-H. Anti-AIDS Agents. 42. Synthesis and Anti-HIV Activity of Disubstituted (3´R,4´R)-3´,4´-Di-O-(S)-camphanoyl-(+)-cis-khellactone Analogues. J. Med. Chem. 2001;44:664–671. doi: 10.1021/jm000070g. [DOI] [PubMed] [Google Scholar]
- 94.Xie L., Guo H.-F., Hong L., Zhuang X.-M., Zhang A.-M., Wu G., Ruan J.-X., Zhou T., Yu D., Qian K., et al. Development and Preclinical Studies of Broad-Spectrum Anti-HIV Agent (3′R,4′R)-3-Cyanomethyl-4-methyl-3′,4′-di-O-(S)-camphanoyl-(+)-cis-khellactone (3-Cyanomethyl-4-methyl-DCK) J. Med. Chem. 2008;51:7689–7696. doi: 10.1021/jm8003009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Amaral J.C., da Silva M.M., da Silva M.F.G.F., Alves T.C., Ferreira A.G., Forim M.R., Fernandes J.B., Pina E.S., Lopes A.A., Pereira M.S., et al. Advances in the Biosynthesis of Pyranocoumarins: Isolation and 13C-Incorporation Analysis by High-Performance Liquid Chromatography−Ultraviolet−Solid-Phase Extraction−Nuclear Magnetic Resonance Data. J. Nat. Prod. 2020;83:1409–1415. doi: 10.1021/acs.jnatprod.9b00607. [DOI] [PubMed] [Google Scholar]
- 96.Khandy M.T., Sofronova A.K., Gorpenchenko T.Y., Chirikova N.K. Plant Pyranocoumarins: Description, Biosynthesis, Application. Plants. 2022;11:3135. doi: 10.3390/plants11223135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Abdelmohsen U.R., Albohy A., Abdulrazik B.S., Bayoumi S.A.L., Malak L.G., Khallaf I.S.A., Bringmann G., Farag S.F. Natural coumarins as potential anti-SARS-CoV-2 agents supported by docking analysis. RSC Adv. 2021;11:16970–16979. doi: 10.1039/D1RA01989A. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.