

Abstract
Phenazines are a large group of heterocyclic nitrogen-containing compounds with demonstrated insecticidal, antimicrobial, antiparasitic, and anticancer activities. These natural compounds are synthesized by several microorganisms originating from diverse habitats, including marine and terrestrial sources. The most well-studied producers belong to the Pseudomonas genus, which has been extensively investigated over the years for its ability to synthesize phenazines. This review is focused on the research performed on pseudomonads’ phenazines in recent years. Their biosynthetic pathways, mechanism of regulation, production processes, bioactivities, and applications are revised in this manuscript.
Keywords: phenazines, Pseudomonas spp., bioprocess
1. Introduction
Phenazines are a large group of heterocyclic, colored, and nitrogen-containing compounds of biological and chemical origin with great stability in natural environments. These natural compounds were first referred to in 1859 by Fordos [1,2], who observed a blue–green pigment responsible for the coloration of the “blue pus” associated with severe wounds resulting from surgical procedures. This pigment was named pyocyanin (1, Table 1) and 65 years later it was identified as a phenazine derivative [3,4]. Since then, over 150 natural phenazine derivatives and more than 6000 synthetic derivatives with wide-ranging bioactivities have been reported [5,6].
The position and type of functional groups in the phenazines’ molecules dictate their chemical, physical, and biological properties that encompass antibiotic, antifungal, insecticidal, antitumor, antimalarial, and antiparasitic activities. This wide bioactivity spectrum is conferred by the high redox activity of phenazine molecules and their ability to reduce molecular oxygen to reactive oxygen species (ROS) [2,7].
Phenazine-producing bacteria are found all over nature in association with plant and animal hosts that live in terrestrial, freshwater, and marine habitats, presenting an important role in bacterial physiological processes, namely, in biofilm formation and in iron reduction. These metabolites play an important role in the ecosystems in which their producers and hosts live, such as in the survival and persistence of rhizobacteria [7,8].
The Pseudomonas genus, which is well-known for its ability to produce multiple secondary metabolites, includes producers of almost one-third of all known phenazines. Among them, 1-hydroxyphenazine (2, Table 1), 2-hydroxyphenazine (3, Table 1), phenazine-1-carboxylic acid (4, Table 1), and phenazine-1-carboxamide (5, Table 1) are the most common derivatives [9]. Phenazine-1,6-dicarboxylic acid (PDC) and PCA act as ‘‘core’’ phenazines that strain-specific genes convert to a different type of phenazine. These phenazines can suffer other natural modifications and be converted into different derivatives [7,9]. Hence, depending on the strain and the environment, PDC and PCA can be converted into several different phenazines (Table 1).
Table 1.
Different phenazine derivatives associated with their producing bacterial strain.
| Compound | Phenazine Derivative | Bacterial Strain | Reference |
|---|---|---|---|
| 1 | Pyocyanin (PYO) | Pseudomonas aeruginosa | [10] |
| 2 | 1-hydroxyphenazine (1-OH-PHZ) | Pseudomonas aeruginosa | [10] |
| 3 | 2-hydroxyphenazine (2-OH-PHZ) | Pseudomonas chlororaphis | [11] |
| 4 | phenazine-1-carboxylic acid (PCA) |
Pseudomonas chlororaphis | [11] |
| 5 | phenazine-1-carboxamide (PCN) |
Pseudomonas aeruginosa; Pseudomonas chlororaphis |
[11,12] |
Natural bacterial phenazines are highly studied because of their importance in interactions with other microorganisms, and also due to their beneficial interactions with plants and animals. Thus, besides the increase in production yields, its isolation from cultivation broths and characterization are being studied. This aims to achieve efficient strategies to recover the product, as well as to elucidate the structure and function to find suitable applications. This review will bring together numerous key aspects of phenazines, focusing on those produced by Pseudomonas spp., ranging from the biological origin of the producing strains and the properties of these versatile secondary metabolites to their uses as biocontrol agents in agriculture and human health.
2. Phenazine-Producing Strains
Phenazines are produced by diverse bacteria mainly from Pseudomonas spp. and Streptomyces spp., but other bacteria have also been reported, isolated from terrestrial and marine environments, namely, Actinomycetes spp., Vibrio spp., Burkholderia spp., and Brevibacterium spp., among others [5,13,14].
Among all phenazine producers, Pseudomonas is the most studied genus at the metabolic and genomic level for that secondary metabolite production, especially fluorescent Pseudomonas [5,11,12]. A diverse phenazine production is observed for each strain, depending on the biosynthetic capabilities for phenazine derivatization, but those isolated from Pseudomonas spp. are mostly simple hydroxyl and carboxyl structures [15]. P. chlororaphis has been reported to produce phenazine-1-carboxylic acid (PCA), 2-hydroxyphenazine-1-carboxylic acid (2-OH-PCA) (Figure 1), 2-hydroxyphenazine (2-OH-PHZ) (Figure 2), and phenazine-1-carboxamide (PCN) [16,17]. For instance, P. choloraphis HT66 and PCL1391 produces PCN, while P. chlororaphis GP72 produces PCA, 2-OH-PHZ like P. chlororaphis 30-84 which is also reported for 2-OH-PCA production [11,18,19,20,21,22]. P. aeruginosa can produce pyocyanin (PYO), PCA, PCN, 1-hydroxyphenazine (1-OH-PHZ), and Aeruginosin A and B [5,23]. Examples of these are P. aeruginosa PA1201 and M18 which produces PCA, like strain LV which, additionally, produces PCN, and P. aeruginosa JY21 which produces PYO [23,24,25,26,27,28]. P. fluorescens, strains fp-5 and 2-79, produce only PCA [11,29,30].
Figure 1.
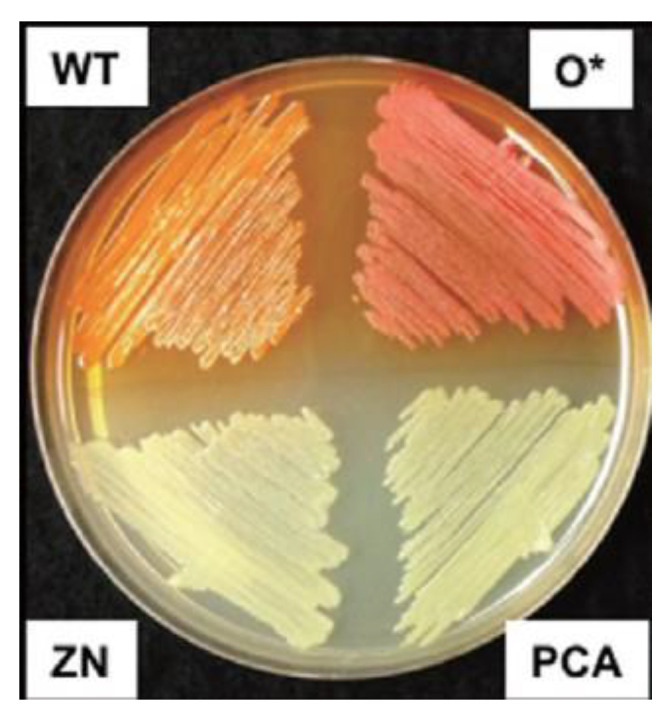
On agar plate, strains: P. chlororaphis 30-84 wildtype (WT), phenazine non-producer (ZN), PCA only producer (PCA), and 2-OH-PCA overproducer (O*) [18].
Figure 2.

Phenazine production on a bioreactor cultivation of Pseudomonas chlororaphis subsp. aurantiaca DSM 19603.
Phenazine production by wild-type strains present low yields to be used in large-scale applications, namely, as a biopesticide in agriculture. To improve phenazine production, based on genetic knowledge, recombinant engineered microorganisms are being developed and investigated as potential cell factories, as well as for growth medium optimization [31]. In fact, Peng et al. [12] achieved a PCN production above 9 g/L by engineering a P. chlororaphis HT66 strain. Zhou et al. [31] reported a PCA production of 6.4 g/L with an engineered Pseudomonas sp. M18 and Jin et al. (2015) have achieved 9.8 g/L of PCA with a P. aeruginosa PA1201 engineered strain (named PA-IV), while the production with the wild-type bacteria was reported to be only 180 mg/L (Table 2).
Table 2.
Phenazine production by Pseudomonas sp., including phenazine type and the amount produced.
| Strain Name | Strain Characteristics | Phenazine Type | Phenazine Production (mg/L) | Reference |
|---|---|---|---|---|
| Pseudomonas aeruginosa JY21 | Wild type | PYO | 311.1 | [28] |
| Pseudomonas aeruginosa LV | Wild type | PCA | 112.89 | [27] |
| PCN | 177.31 | |||
| Pseudomonas aeruginosa PA1201 | Wild type | PCA | 180 | [24] |
| Pseudomonas aeruginosa PA-IV | Delete phzS, phzM, phzH, pabB/C, trpE, and pchA; overexpression of aroG and phzC1; engineering the promoters of PCA biosynthetic gene clusters and the efflux pump, etc. | PCA | 9882 | [24] |
| Pseudomonas spp. MCC 3145 | Wild type | PYO | 313.94 | [34] |
| Pseudomonas sp. M18 | Wild type | PCA | 48.0 | [35] |
| Pseudomonas sp. M18 G | gacA deficient | PCA | 2597 | [33] |
| Pseudomonas sp. M18 GQ | gacA deficient; overexpression of phz gene cluster | PCA | 6365 | [33] |
| Pseudomonas chlororaphis HT66 | Wild type | PCN | 424.87 | [12] |
| Pseudomonas chlororaphis HT66 P3∆lon | Point mutations in 138 genes; deletion of lon gene | PCN | 9174 | [12] |
| Pseudomonas chlororaphis GP72 | Wild type | PCA | 22.0 | [20] |
| 2-OH-PHZ | 4.5 | |||
| Pseudomonas chlororaphis GP72AN | Inactivation of rpeA gene | PCA | 432/NR | [20,36] |
| 2-OH-PHZ | 170/258 | |||
| Pseudomonas fluorescens 2-79 | Wild type | PCA | 1010 | [37] |
Regarding media optimization, P. chlororaphis GP72 was engineered to utilize the glycerol pathway. The two genes, glpF and glpK, from the glycerol metabolism pathway were overexpressed in GP72ANO separately and after were co-expressed, resulting in a production improvement from 729.4 mg/L to 993.4 mg/L. To enhance the glycerol use, the shunt pathway was blocked, resulting in 1493.3 mg/L PCA production [32].
Li et al. [33] tested different strategies to enhance PCN production, namely, the knocking out of negative regulatory factors, the enhancement of the shikimate pathway by gene overexpression and gene knocking, and by using fed-batch cultivation. Finally, an engineered strain of P. chlororaphis achieved 11.45 g/L PCN.
3. Phenazines Biosynthesis
3.1. Metabolic Pathways
Phenazine biosynthesis is derived from the shikimate pathway, which is also known for the production of the amino acids: phenylalanine, tyrosine, and tryptophan in microorganisms [38,39,40]. From this pathway, the branch point for phenazine production is chorismic acid, giving rise to the basic phenazine structure.
The shikimate pathway (Figure 3) starts with the condensation of erythrose-4-phosphate (E4P) and phosphoenolpyruvate (PEP) to form D-arabino-heptulosonate-1-phosphate (DAHP), catalyzed by the enzyme PhzC, phospho-2-dehydro-3-deoxyheptonate aldolase [38,40]. DAHP is then converted into chorismate, the precursor of phenazines. The following step is the conversion of chorismate into 2-amino-4-deoxyhorismic acid (ADIC) by PhzE (anthranilate/para-aminobenzoate synthase). Subsequently, ADIC is hydrolyzed to pyruvate and trans-2,3-dihydro-3-hydroxyanthranlic acid (DHHA) by PhzD, an isochorismate hydrolase. DHHA is then converted to 6-amino-5-oxo-cyclohex-2-ene-1-carboxylic acid (AOCHC) by PhzF [40,41]. The next step is the formation of the phenazine tricycle, catalyzed by enzymes PhzB/A leading to hexahydrophenazine-1,6-dicarboxylic acid (HHPDC) [42,43,44]. This molecule suffers oxidative decarboxylation, giving rise to tetrahydrophenazine-1,6-carboxylic acid (THPCA). THPCA is transformed into 5,10-dihydrophenazine-1-carboxylix acid (DHPCA), catalyzed by PhzG (pyridoxamine 5′-phosphate oxidase) [42,43,44,45]. DHPCA is finally converted into PCA, by spontaneous oxidation by air. Figure 3 illustrates the biosynthetic pathway of Pseudomonas genus phenazines.
Figure 3.

Biosynthetic pathway for phenazine production by Pseudomonas spp. DAHP: 3-deoxy-d-arabino-heptulosonate 7-phosphate; ADIC: 2-amino-2-deoxyisochorismate; DHHA: trans 2,3-dihydro-3-hydroxyanthranilic acid; AOCHC: 6-amino-5-oxocyclohex-2-ene-1-carboxylic acid; HHPDC: hexahydro-phenazine-1,6-dicarboxylate; PHZ: phenazines; PDC: phenazine-1,6-dicarboxylic acid; PCA: 4 phenazine-1-carboxylic acid; 1-OH-PHZ: 2 1-hydroxy-phenazine; PCN: 5 phenazine-1-carboxamide; 2-OH-PHZ: 3 2-hydroxy-phenazine; 5-MPCA: 5-methylphenazine 1-carboxylato; PYO: 1 pyocianin.
The production of strain-specific phenazines is carried out by enzymes encoded by genes like phzH, phzM, phzO, and phzS, which usually flank the core gene cluster [24,46]. The production of these phenazines is coupled with the biosynthesis of the core molecule PCA. However, the modifications are thought to occur in DHPCA, instead of PCA, due to its higher reactivity [45]. The enzyme PhzM is a phenazine-specific methyltransferase and is reported to catalyze the formation of 5-methylphenazine-1-carboxylate (5MPCA) that is then converted to PYO by PhzS, a flavin-containing monooxygenase [23,46]. These two enzymes are required to obtain PYO and the genes that encode them are reported to be present in P. aeruginosa PAO1, for instance [23]. However, when only PhzS is present the phenazine derivative produced is 1-OH-PHZ [23]. PhzH, an asparagine synthase, is the enzyme involved in the conversion of DHPCA into PCN, resulting in a large accumulation of PCN compared with the amount of PCA detected [17,23,47,48]. This enzyme is found in P. chlororaphis PCL1391, P. chlororaphis HT66, and P. aeruginosa PAO1 [49,50]. The production of 2-OH-PCA is mediated by PhzO, a phenazine hydroxylase and afterward, a part of it suffers a spontaneous decarboxylation to form 2-OH-PHZ [20,46,47]. In this process, PhzO is believed to suffer substrate inhibition leading to a low yield of 2-OH-PCA and 2-OH-PHZ [51]. Such a modification pathway is reported for P. chlororaphis 30-84 and PCL1391 [46].
Phenazine-producing Pseudomonas spp. are reported to produce mainly PCA and PCA derivatives, while phenazine-1,6-dicarbooxylic acid (PDC) is typically produced by Streptomyces spp. [34,52]. The exception is Pseudomonas aeruginosa strain HRW.1-S3, which was reported to produce PDC when growing in the presence of crude oil as the sole carbon source [52].
In Pseudomonas spp. the gene cluster phzABCDEFG is responsible for converting the chorismate into PCA and it is highly conserved in all phenazine-producing pseudomonads [5,34,53]. Recent studies suggest that the PCA production by Pseudomonas is related to the presence of the phzA gene (that presents high homology with the phzB gene), whose presence is restricted to this type of phenazine producer, and the expression level of phzG gene [44,45]. Guo et al. [44] reported a significant PDC production by a P. chlororaphis HT66 phzA-disrupted mutant alongside PCA production. Later, Guo et al. [45] reported a spontaneous PDC synthesis by a P. chlororaphis GP72AN without the phzG gene.
3.2. Biological Regulation
In Pseudomonas spp., phenazine gene expression and consequent phenazine production is under the control of a two-component system, namely, the quorum-sensing (QS) mechanism and a small RNA (sRNAs) system [54,55,56,57].
In QS, bacteria release small molecules or peptide signals into the environment, which can then interact with bacteria of the same strain and activate the expression of genes under the control of this mechanism (Figure 4). Molecules with diverse structures have been associated with this behaviour, but acyl homoserine lactones (AHLs) are the most relevant for phenazine expression. Monitoring the number of compounds that are QS regulated, it becomes clear that they arise when a minimum cell concentration is reached. Therefore, those signals enable bacteria to sense their own population size and delay the expression of some genes until a specific cell density is achieved [54,55,58].
Figure 4.

Quorum-sensing regulation mechanism.
The most studied and best characterized QS mechanism regulates phenazine production mediated by N-acyl-L-homoserine lactones (AHL), as found in P. chlororaphis strains [12,59]. The genes phzI encodes for AHL-syntase, and phzR encodes for AHL-receptor, which precede the operon phz. The enzyme PhzI is responsible for producing the AHLs that accumulate in the external medium and, when it reaches a threshold level, binds to PhzR activating the transcription of the phz operon [50,59].
The sRNA signalling is controlled by GacS sensor kinase which is highly important since it gives bacteria the potential to directly understand the environmental signs and then modulate the activities of regulatory mechanisms or control phz expression. This occurs when environmental conditions change and the phosphorylation status of the sensor protein is altered through the binding of a small molecule or other environmental signals that activate conformational changes and affects the activity [7,56,57].
The two-component system most studied is the GacS/GacA where the membrane-bound protein GacS is regulated by GacA, a cytoplasmic regulatory protein, promoting the expression of small non-coding RNAs (e.g., RsmZ). These RNAs bind to translational repressors preventing the repression of the core biosynthetic genes phz in bacteria such as P. chlororaphis [60,61]. However, the regulation can be different among Pseudomonas spp. [61,62]. RpeA/RpeB is another two-component system found in P. chlororaphis, in which RpeA negatively regulates the phzR expression by negative regulation of pip and RpeB has the opposite effect [61]. Pip is an activator of phenazine production in P. chlororaphis that is also regulated by the GacS/GacA system [61,63]. P. chlororaphis is divided into four subspecies, Morohoshi et al. [63] investigated the relation between phenazine production and quorum sensing in the two subspecies that had not been studied yet, namely P. chlororaphis subsp. chlororaphis and piscium. It was found that the disruption of the phzI caused no production of phenazine-1-carboxylic acid (PCA) and phenazine-1-carboxamide (PCN). On the other hand, PCA and PCN production were not affected by the disruption of CsaR-CsaI, a second quorum-sensing system identified in P. chlororaphis subsp. aureofaciens 30-84. Moreover, Morohoshi et al. [63], showed that the PhzI/PhzR quorum-sensing system plays an important role in the production of phenazine derivatives in both strains.
4. Phenazine Production
Phenazine production is growth associated with and affected by environmental factors including temperature, culture medium composition, oxygen availability, and pH value [6,22]. In view of this, the improvement of fermentative conditions is being studied to understand and enhance phenazine production. However, different strains have different nutritional needs depending on their environment [26,30]. So, strategies such as the application of statistical methods for growth medium optimization, use of different feeding strategies, or evaluation at a genetic level, and factors that are associated with phenazine production are being studied to improve their biosynthesis [22,24,64,65]. Rij et al. [22] showed that PCN production by P. chlororaphis PCL 1391 is induced by some carbon sources, such as glucose, glycerol, and L-pyroglutamic acid. Similarly, other authors found glucose to be the best carbon source for phenazine production by engineered strains of Pseudomonas sp. M18 (M18G) and P. fluorescens 2-79, or glycerol for P. fluorescens fp-5, Pseudomonas spp. MCC3145, and the engineered strain P. chlororaphis P3∆lon [12,64,66,67]. Ethanol complemented with glucose was found to be the best carbon source for PCA production by the Pseudomonas sp. M18G and M18GQ strains [28,31]. Regarding nitrogen sources, ammonium ions and amino acids are reported to have a stimulatory effect on phenazine production, with peptone, tryptone, soybean meal, and corn steep liquor being the most significant [12,22,26,29,31,64]. Yuan et al. [65], using a surface response methodology, found the ideal concentrations of glucose and soytone to increase the PCA production by Pseudomonas sp. M18Q, the qscR-chromosomal inactivated mutant, from 750 mg/L to 1240 mg/L. Similarly, Peng et al. [12] improved the PCN production of the P. choloraphis P3∆lon mutant nearly three-fold to 9174 mg/L in an optimized medium, in which glycerol, tryptone, and soy peptone were identified as the most significant factors. For the genetically engineered Pseudomonas sp. M18G (a gac-inactivated mutant), the best results for PCA production were found with soybean meal complemented with soy peptone as a nitrogen source and ethanol complemented with glucose as a carbon source, having achieved 1.98 g/L, near a three-fold increase from the basal medium [26].
Temperature is also an important factor for phenazine production, and some enzymes involved in the phenazine production process are temperature dependent. That is the case of PhzM, which in P. aeruginosa PAO1 is expressed at higher levels at 37 °C, and in Pseudomonas sp. M18 at 28 °C, meaning that, depending on the temperature, the ratio between PCA and PYO will be different [23]. Similar results were achieved by Bedoya et al. [27] for the production of PCA and PCN by P. aeruginosa LV, where at 28 °C is possible to achieve a higher PCN production, than at 32 °C, favoring PCA production. On the other hand, a considerable decrease in PCN production was observed when the temperature decreased from 21 ºC to 16 °C. Moreover, acidic pH was found to negatively affect phenazine production, having observed a dramatic decrease in PCN synthesis when the pH was changed from 7 to 6 [23]. Similarly, the pH value was found to be a significant factor for PYO production by P. aeruginosa JY21 in Abo-Zaid et al. [28], being 8.2 as the best value for their process. Cui et al. [30] optimized the pH to 7.2 in a 1 L bioreactor, leading to an increase in AHLs that resulted in a higher PCN production (8.58 ± 0.25 g/L) by Pseudomonas chlororaphis H5ΔfleQΔrelA.
Nutrients such as oxygen, ferric iron, phosphate, sulphate, and magnesium were also reported to be significant in phenazine production [22,27,28,64]. Low concentrations of ferric iron, phosphate, and sulphate, as well as high magnesium and oxygen levels (above 1%), were found to reduce PCN levels in P. chlororaphis PCL1391 [22]. Jin et al. [64] reported that the downregulation of proteins involved in phosphate transport could result in an increase in PCN production, as well as the upregulation of proteins involved in iron homeostasis. The presence of ferric iron is also reported to enhance the conversion of PCA or DHPCA to 2-OH-PHZ since the reaction is Fe3+ dependent [51].
Operational strategies have been also studied to increase phenazine production. A fed-batch strategy was reported by Jin et al. [24] and Li et al. [33] for PCA production by the engineered Pseudomonas sp. M18G and P. aeruginosa PA-IV, respectively. Li et al. [33] found that glucose fed in a pulse manner to keep the concentration above the inhibitory level favored higher cellular growth. A different strategy was followed by Huang et al. [20] that added exogenous PCA (400 µg/mL) to the cultivation of the engineered P. chlororaphis GP72AN and favored PCA and 2-OH-PHZ production. Yue et al. [36] tried a two-stage cultivation strategy involving the use of the reducing agent DTT (Dithiothreitol), to create reducing conditions in the medium favoring a higher production of PCA. Posteriorly, they fed the system with the electron acceptors K3[Fe(CN)6] and H2O2 to generate an oxidative environment allowing the activity of PhzO to convert PCA into 2-OH-PHZ. This strategy was found to improve phenazine production, since phenazines are involved in ROS production that can be harmful to the producer itself, affecting phenazine yields. Aloui et al. [67] reported the improvement of co-production of medium-chain-length polyhydroxyalkanoates (mcl-PHA) and extracellular phenazines through a high cell density cultivation of Pseudomonas chlororaphis subsp. aurantiaca (DSM 19603) in a membrane bioreactor using crude glycerol as a feedstock.
5. Extraction and Purification of Phenazines
The most commonly reported method for extraction of phenazines involves the use of organic solvents applied in a cell-free supernatant that can suffer a previous acidification step. Organic solvents such as ethyl acetate, dichloromethane, chloroform, methylene dichloride, toluene, or diethyl ether are the most reported for this step of the purification process. Then, the organic phase where phenazines are dissolved can be evaporated and the dry solids dissolved in acetonitrile or methanol [19,20,22,23,27,29,32,52,68,69,70]. Some authors, after solvent utilization, include a chromatography step with silica gel to enhance the purification [29,71,72,73,74]. However, the use of organic solvents produces hazardous wastes requiring precautions for safe handling [75].
Rane et al. [76] reported a prior step in PCA extraction, the use of a XAD-4 resin column to remove impurities followed by the extraction of the phenazine compound with chloroform from the acidified solution followed by crystallization.
The separation and purification are mostly reported by thin layer chromatography (TLC) and preparative high-performance liquid chromatography (HPLC) which have good separation resolutions [68,71,75,76,77]. However, other techniques are being studied, such as capillary electrophoresis, free flow electrophoresis, and the use of microporous adsorbent resins as alternatives to reduce the price, the use of organic solvents and the time required for the separation [68,70,78,79,80]. Liu et al. [78] applied capillary zone electrophoresis to separate and quantify PCA and 2-OH-PHZ from the culture broth of P. chlororaphis GP72, after an extraction step, offering a rapid and sensitive detection. Free flow electrophoresis for low-concentration PCA purification from Pseudomonas sp. M18 cultivation broth was reported by Shao et al. [70]. This method, which requires a previous step for phenazine extraction, offers the advantages of a continuous process, a lower cost due to the consumption of the mobile phase, and high product recovery. Bilal et al. [68] reported the separation and purification of PCA from the cultivation broth of the engineered P. chlororaphis GP72AN with microporous adsorbent resins using methanol as a desorbing agent. This strategy is presented as a simple and efficient process that can be used for a scale-up process. Zhou et al. [81] developed electrochemical sensors based on laser-induced graphene for real-time monitoring of P. aeruginosa phenazines.
6. Phenazine Characterization
In recent years, different techniques to characterize phenazines have been studied taking into consideration the properties of phenazines, namely, their fluorescent properties, redox activity, and light adsorption properties.
TLC, HPLC, and UV-visible spectroscopy have been used by several authors to detect and characterize phenazines [13,20,32,69,72,76,79]. Further, structural characterization and identification of phenazines are performed, after extraction, separation, and purification, using mass spectroscopy (MS, liquid, or gas), nuclear magnetic resonance (NMR, 1H and 13C) (Figure 5), and Fourier Transform Infra-Red spectroscopy (FTIR) (Figure 6) [69,72,73,74,76,78].
Figure 5.

Spectra of different characterization analyses of phenazines, (A) Phase-reverse HPLC chromatogram of PCA and PCN [49]; (B) PCA and PCN peaks detected by LC-MS [18]; (C) FTIR spectrum of crystalline PCA [76].
Figure 6.

Spectra of PCN phenazine by (A) 1H NMR and by (B) 13C NMR [12].
Kern and Newman [82] reported the use of HPLC to separate the different phenazines, namely, PYO, PCA, and 1-hydroxyphenazine (1-OH-PHZ), which were then quantified by UV absorption. PCA and PCN were detected by Peng et al. [12] using HPLC with a C18-WR reversed-phase column at 254 nm. The retention times for PCA and PCN were approximately 9.523 and 17.217 min, respectively. Simionato et al. [83] purified and identified PCA produced by P. aeruginosa LV strains by extracting it with dichloromethane. Then, the purity degree of the extracted phenazine was determined using reversed-phase HPLC semi-preparative and the structure was confirmed by NMR and electrospray ionization mass spectroscopy.
PCA and 1-hydroxyphenazine were extracted and purified from a cultivation supernatant of P. aeruginosa JAAKPA using TLC and column chromatography. Then, the sample was subjected to gas chromatography–mass spectrometry (GC-MS), FTIR (Fourier Transform Infra-Red spectroscopy), and NMR were then identified [78]. Further, LC-MS was used by Yu et al. [18] to identify different phenazines (e.g., PCA, PCN) produced by P. chlororaphis 30-84 (Figure 5). Jasim et al. [69] identified a PCA produced by an endophytic P. aeruginosa isolated from Zingiber officinale. Firstly, they extracted the compound and then it was purified by TLC being obtained from 7 different fractions, then the presence of compounds was measured by UV scan. Further bioactivity was tested and the fraction that showed activity against Pythyum myriotylum was subjected to LC-MS (Light Scattering–Mass Spectroscopy) to identify the compound (Figure S1).
Extracellular metabolites were extracted using ethyl acetate and a UV spectrum was recorded to quantify phenazines. Further, the compound was purified using silica-gel column chromatography. The purified compound was characterized and identified using different techniques, namely, FTIR, NMR, and mass spectroscopy (supplementary material). IR spectra (Figure S2A) showed peaks related to the aromatic C–H stretching vibrations (2963, 2924, and 2853 cm−1). Moreover, an amide carbonyl group was also confirmed. The 1H NMR spectrum (Figure S2B) of the P. aeruginosa MML2212 purified compound showed two doublets, and two protons were also displayed. Further, three aromatic protons appeared as a multiplet in the range. The 13C NMR (Figure S2C) spectrum of the purified compound showed the presence of a carbonyl carbon of the amide group. Finally, the sample was subjected to mass spectroscopy (Figure S2D); the compound was identified as PCN (phenazine-1-carboxamide) [72].
P. aurantiaca produced secondary metabolites with antifungal activity. The metabolites were identified using TLC, UV spectra, and MALDI-TOFF spectra and were revealed to be PCA, 2-hydroxyphenazine (2-OH-PHZ), and N-hexanoyl homoserine lactone (HHL) (Figure S3) [71].
7. Properties, Applications, and Commercial Products
Phenazines present antifungal and antibacterial activities, being recognized as broad-spectrum antibiotics. Pierson and Pierson [84] reported the ability of phenazines to eliminate the disease of wheat root caused by the fungus Gaeumannomyces graminis var. graminis.
Pseudomonas phenazine species work also as biocontrol promoters of plant health, such as Macrophomina phaseolina, Fusarium graminerum, and Rosellinia necatrix. Macrophomina phaseolina is one of the most virulent phytopathogens that can infect more than 500 plant species, such as soybean, chickpea, or peanut, causing dry root and stem rot, known as charcoal rot. PCA shows a high antifungal activity against this fungus, presenting a MIC50 (Minimum Inhibitory Concentration 50%) value of 35 µg/mL [85,86].
Apart from antifungal activity, phenazines may also act as antibacterial agents [87]. Phenazines possess antibiotic activity against, for instance, Staphyloccoccus aureus, Escherichia coli, and other bacteria [88,89]. D-Alanylgriseoluteic acid, a potent antimicrobial phenazine compound, is a prime example of the antibacterial activity of these metabolites. This phenazine exhibits a MIC50 value of 0.25 µg/mL and a MIC90 value of 0.5 µg/mL for penicillin-resistant isolates of Streptococcus pneumoniae. Both MIC values of penicillin for these isolates were 4 µg/mL [90]. The antibiotic effect is attributed to the redox properties of phenazines and their capacity to promote the formation of toxic reactive oxygen species (ROS).
Several authors reported PCA activity against different fungi (e.g., Fusarium oxysporum, Penicillium expansum, Rhizoctonia solani), produced by different Pseudomonas spp. [74,91]. Pseudomonas sp., namely, P. piscium and P.aeruginosa, also produced phenzine 1HP that has activity against fungi, such as Fusarium graminearum, Colletotrichum gloeosporioides, Exserohilum turcicum [92,93,94]. Park et al. [91] reported activity against Rhizoctonia solani by phenazine 2-OH-PHZ produced by P. aurentiaca IB5-10. Karmegham et al. [95] also reported the antifungal activity of phenazine derivatives isolated from fluorescent pseudomonads (FPs). PCN was detected in the isolates of FPs, showing a prominent antifungal activity against R. solani and other tested fungal pathogens.
Antifungal activity was likewise reported against a major human pathogen fungus, Trichophyton rubrum, which could be responsible for causing athlete’s foot, jock itch, ringworm, and fingernail fungus infections [5,74]. Phenazines were tested against Candida species, which are responsible for candidiasis infections. Morales et al. [96] showed that 5-methylphenazine-1-carboxylic acid (5MPCA) produced by Pseudomonas aeruginosa has an antibiotic effect against Candida albicans. A synergistic effect between phenazines isolated from P. aeruginosa and azoles were observed against several species of genus Candida, and, further, no cytotoxicity against human cell lines was observed [74].
Furthermore, it was demonstrated by different authors that phenazines possess anticancer and neuroprotective activity. The anticancer ability is an outcome ensured by known mechanisms, such as polynucleotide interaction, topoisomerase inhibition, and radical scavenging [5,7,15,97]. Polynucleotide interaction occurs because the aromatic phenazine core has structural similarity to known intercalators-metabolites capable of inhibiting DNA replication and several studies of the interactions between phenazines and DNA/RNA have been made. In 1971, Hollstein et al. [98] reported the study of the interaction between Pseudomonad pyocyanin, two other phenazines, and various polynucleotides. Kennedy et al. [99] isolated a 5MPCA produced by Pseudomonas putida, which exhibited selective cytotoxicity regarding the cell lines of breast and lung cancer. The isolated phenazine inhibits cell viability, DNA synthesis, induced G cell cycle arrest, and apoptosis in cancer cells.
Topoisomerases (I and II) are enzymes responsible for topological changes in the DNA strand during cell division (translation and transcription). Proliferating cells, like cancer cells, contain large concentrations of topoisomerases, which, therefore, serve as obvious therapeutic targets in cancer treatment. Drugs with topoisomerase II inhibition properties are widely used in chemotherapy, whereas topoisomerase I inhibitors have proven clinically useful in the treatment of colon cancer. Topoisomerase inhibition has not been reported for any of the naturally occurring phenazines but has been intensively pursued in synthetic analogues [97,100].
Radical scavenging is probably the most important ability associated with phenazines since it is believed that free radicals are involved in the development and progression of a wide range of serious human diseases. Oxygen-delivered free radicals are known to induce irreversible damage to neuronal cells in diseases like Parkinson’s disease and, perhaps, dementia, atherosclerosis, and cerebral traumas or strokes [101]. They are also believed to be involved in inducing carcinogenesis, aging of cells, asthma, renal failure, rheumatoid arthritis, and inflammation [101]. Natural antioxidants like vitamins C and E have multiple physiological functions but are insufficient in the treatment of free-radical-induced diseases. Therefore, efficient synthetic radical scavengers or antioxidants are needed to reduce these types of damage, including the irreversible loss of neural tissue. In normal cells, free radicals are continuously produced by metabolic enzymatic processes in the mitochondria as a part of energy production. Likewise, the body utilizes free radicals in immune responses toward incoming pathogens and in the degeneration of toxins. However, the overproduction of radicals or other dysfunctions causes the accumulation of radicals inside the cell and, consequently, cell death. The antioxidant or radical scavenging character of the phenazines can, therefore, be confirmed by measuring their ability to inhibit lipid peroxidation in liver microsomes [101]. Pyocyanin isolated from P. aeruginosa demonstrated higher scavenging activities at much lower concentrations than ascorbic acid [102].
Fungal and oomycete are common plant diseases, such as Rhizoctonia solani Kuhn and Fusarium graminearum. These fungus pathogens cause sheath blight of rice and blight of wheat generating huge losses during production and postharvest storage [103]. The intervention of synthetic agrochemicals is the most effective method at present; however, the current chemical-control agents are not fully effective at inhibiting these fungi’s activity. Further, it is necessary to develop more effective novel agents to replace conventional agrochemicals that introduce massive environmental pollutants and cause soil-borne diseases [104,105].
An effective method to replace agrochemicals is the development of new “green agrochemicals”, which are active compounds contained in natural products. As it was previously mentioned, several phenazines present activity against several fungi. The most common active phenazines are PCA, PCN, 1-hydroxyphenazine (1HP), 2-hydroxyphenazine (2HP), and 2-hydroxyphenazine-1-carboxylic acid (2HPCA), with the former being the most common [8]. The phenazine concentrations are detected on a scale of µg/mL, however, the concentration differs depending on the strain [106], and, usually, the concentration of PCA is present in a higher concentration.
Despite all known phenazines with antifungal activity, PCA is the only commercialized fungicide product with the trade name Shenqinmycin noted for its high fungicidal efficiency, low toxicity to humans and animals, friendliness to the environment, and improvement of crop production [103]. However, in the market, there are other products based on Pseudomonas strains, namely, P. chlororaphis which has been commercialized by companies from Europe (Cedomon BioAgri AB, Uppsala, Sweden) and the USA (AtEze, Turf Science Laboratories). Further, P. fluorescens A506 is the main component of BlightBAn A506 (Nufarm Americas Inc., Houston, TX, USA). These products have activity against fungi such as Fusarium oxysporum, Aspergillus niger, and Botrytis cinerea [8,33,85].
As described above, phenazines can be widely used as medicine and pesticides. However, due to the phenazine’s non-selective DNA binding, they may present toxic risks. Hence, Ou et al. [107] studied the degradation, adsorption, and leaching behaviors of a PCN in the agricultural soil of China. The authors found that PCN is easily degraded, has high adsorption affinity, and low mobility in high organic matter content and clay soils, thus, resulting in lower risks of contamination to groundwater systems. However, the risk of contamination increases greatly in soils containing low organic matter and low clay content leading to low adsorption affinity and moderate mobility in soils [107]. Studies like this should be performed to provide interesting insights concerning the use of phenazines.
8. Final Remarks
This review focused on the work developed in the field of natural phenazines, in particular, Pseudomonas spp. phenazines. In recent years, the research in this area has increased greatly, contributing to much clearer knowledge regarding the biosynthetic pathways and the regulatory mechanisms of phenazine production by bacteria. However, more research is needed to better understand the reason why one or another phenazine is produced.
Moreover, the natural phenazines showed exciting bioactivities, including antifungal properties against plant pathogens but also as an anticancer agent and as an antibiotic for human diseases caused by different pathogens. However, despite the interesting features of phenazines, only one PCA is commercialized as a biofungicide. Such limitation could be related to low phenazine concentrations produced by most bacterial strains. PCA is the only phenazine produced in higher concentrations that also shows the best biocontrol ability in comparison with the rest of the natural Pseudomonas spp. phenazines. Hence, more research is needed in order to increase phenazine production and in discovering new phenazines with higher production by bacteria and higher antifungal activity, since it is of major importance to substitute the synthetic agrochemical with natural pesticides, in order to reduce the environmental burden. Moreover, future studies should be focused on the phenazine production process scale-up, considering all the challenges that a process scale-up may have, namely, in the downstream process. Therefore, more studies should be performed to optimize extraction and purification methods in order to enable the process scale-up, but also to reduce the use of organic solvents (e.g., chloroform) and substitute them with greener solvents, for instance, ethyl acetate or DES (Deep Euthetic Solvents).
Further, despite this review being about pseudomonad phenazines, it is known that many other microorganisms are able to produce phenazines, thus, the research about phenazine production by such bacteria should increase.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/molecules28031368/s1, Figure S1: Mass spectrum of purified active fraction using LC_MS analysis; Figure S2: Characterization and identification of the purified compound of P. aeruginosa MML2212 (A) IR spectrum; (B) 1H NMR; (C) 13C NMR and (D) Mass spectrum; Figure S3: Quantification and identification of PCA and 2-hydroxyphenazine (2-OH-PHZ) from P. aurantiaca PB-St2. (A) UV spectra of PCA and (B) 2-OH-PHZ; (C) Mass spectrum of “matrix A”; (D) Mass spectrum of “matrix A” spiked with PCA, (E) Mass spectrum of “matrix A” spiked with 2-OH-PHZ.
Author Contributions
Conceptualization, B.S., A.R.B., C.A.V.T. and F.F.; investigation, B.S. and C.A.V.T.; writing—original draft preparation, B.S. and A.R.B.; writing—review and editing, C.A.V.T. and F.F.; supervision and funding C.A.V.T. and F.F. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Funding Statement
This work was supported by the Associate Laboratory Institute for Health and Bioeconomy-i4HB (LA/P/0140/2020), the Applied Molecular Biosciences Unit–UCIBIO (UIDP/04378/2020 and UIDB/04378/2020), and the national project PTDC/CTM-CTM/29869/2017 and UID/DTP/04138/2019, which are financed by national funds from FCT-Fundação para a Ciência e a Tecnologia, I.P. (Portugal), Ana R. Bernardino acknowledge FCT, I.P. for financial support through Ph.D. fellowship SFRH/BD/138011/2018.
Footnotes
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
References
- 1.Fordos M.J. Recherches sur la matière colorante des suppurations bleues: Pyocyanine. CR Acad. Sci. 1860;51:215–217. [Google Scholar]
- 2.Guttenberger N., Blankenfeldt W., Breinbauer R. Recent developments in the isolation, biological function, biosynthesis, and synthesis of phenazine natural products. Bioorg. Med. Chem. 2017;25:6149–6166. doi: 10.1016/j.bmc.2017.01.002. [DOI] [PubMed] [Google Scholar]
- 3.Wrede F., Strack E. Über das pyocyanin, Den blauen Farbstoff des Bacillus pyocyaneus. Hoppe-Seyler´s Z. Physiol. Chem. 1924;140:1–15. doi: 10.1515/bchm2.1924.140.1-2.1. [DOI] [Google Scholar]
- 4.Gonçalves T., Vasconcelos U. Colour me blue: The history and the Biotechnological potential of pyocyanin. Molecules. 2021;26:927. doi: 10.3390/molecules26040927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Yan J., Liu W., Cai J., Wang Y., Li D., Hua H., Cao H. Advances in phenazines over the past decade: Review of their pharmacological activities, mechanisms of action, biosynthetic pathways and synthetic strategies. Mar. Drugs. 2021;19:610. doi: 10.3390/md19110610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Schmitz S., Rosenbaum M.A. Controlling the production of Pseudomonas phenazines by modulating the genetic repertoire. ACS Chem. Biol. 2020;15:3244–3252. doi: 10.1021/acschembio.0c00805. [DOI] [PubMed] [Google Scholar]
- 7.Chincholkar S., Thomashow L. Microbial Phenazines. 1st ed. Springer; Berlin/Heidelberg, Germany: 2013. [Google Scholar]
- 8.Wang S.-Y., Shi X.-C., Chen X., Laborda P., Zhao Y.-Y., Liu F.-Q., Laborda P. Biocontrol ability of phenazine-producing strains for the management of Fungal Plant Pathogens: A Review. Biol. Control. 2021;155:104548. doi: 10.1016/j.biocontrol.2021.104548. [DOI] [Google Scholar]
- 9.Wan Y., Liu H., Xian M., Huang W. Biosynthetic pathway construction and production enhancement of 1-hydroxyphenazine derivatives in Pseudomonas chlororaphis H18. J. Agric. Food Chem. 2022;70:1223–1231. doi: 10.1021/acs.jafc.1c07760. [DOI] [PubMed] [Google Scholar]
- 10.Kerr J.R., Taylor G.W., Rutman A., HOiby N., Cole P.J., Wilson R. Pseudomonas aeruginosa pyocianin and 1-hydroxyphenazine inhibit fungal growth. J. Clin. Pathol. 1999;52:385–387. doi: 10.1136/jcp.52.5.385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mavrodi D.V., Blankenfeldt W., Thomashow L.S. Phenazine compounds in fluorescent Pseudomonas spp. Biosynthesis and regulation. Annu. Rev. Phytopathol. 2006;44:417–445. doi: 10.1146/annurev.phyto.44.013106.145710. [DOI] [PubMed] [Google Scholar]
- 12.Peng H., Zhang P., Bilal M., Wang W., Hu H., Zhang X. Enhanced biosynthesis of phenazine-1-carboxamide by engineered Pseudomonas chlororaphis HT66. Microb. Cell Fact. 2018;17:1–12. doi: 10.1186/s12934-018-0962-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Blankenfeldt W., Parsons J.F. The structural biology of phenazines biosynthesis. Curr. Opin. Struct. Biol. 2014;29:26–33. doi: 10.1016/j.sbi.2014.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pierson L.S., Pierson E.A. Metabolism and function of phenazines in bacteria: Impacts on the behaviour of bacteria in the environment and Biotechnological Processes. Appl. Microbiol. Biotechnol. 2010;86:1659–1670. doi: 10.1007/s00253-010-2509-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bilal M., Guo S., Iqbal H.M., Hu H., Wang W., Zhang X. Engineering Pseudomonas for phenazine biosynthesis, regulation, and biotechnological applications: A Review. World J. Microbiol. Biotechnol. 2017;33:1–11. doi: 10.1007/s11274-017-2356-9. [DOI] [PubMed] [Google Scholar]
- 16.Laursen J.B., Nielsen J. Phenazine natural products: biosynthesis, synthetic analogues, and biological activity. Chem. Rev. 2004;104:1663–1686. doi: 10.1021/cr020473j. [DOI] [PubMed] [Google Scholar]
- 17.Chen Y., Shen X., Peng H., Hu H., Wang W., Zhang X. Comparative genomic analysis and phenazine production of Pseudomonas chlororaphis, a plant growth-promoting Rhizobacterium. Genom. Data. 2015;4:33–42. doi: 10.1016/j.gdata.2015.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yu J.M., Wang D., Pierson L.S., Pierson E.A. Effect of producing different phenazines on bacterial fitness and biological control in Pseudomonas chlororaphis 30-84. Plant Pathol. J. 2018;34:44–58. doi: 10.5423/PPJ.FT.12.2017.0277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yao R., Pan K., Peng H., Feng L., Hu H., Zhang X. Engineering and systems-level analysis of Pseudomonas chlororaphis for production of phenazine-1-carboxamide using glycerol as the cost-effective carbon source. Biotechnol. Biofuels. 2018;11:1–15. doi: 10.1186/s13068-018-1123-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Huang L., Chen M.-M., Wang W., Hu H.-B., Peng H.-S., Xu Y.-Q., Zhang X.-H. Enhanced production of 2-hydroxyphenazine in Pseudomonas chlororaphis GP72. App. Microbiol. Biotechnol. 2010;89:169–177. doi: 10.1007/s00253-010-2863-1. [DOI] [PubMed] [Google Scholar]
- 21.Liu K., Hu H., Wang W., Zhang X. Genetic engineering of Pseudomonas chlororaphis GP72 for the enhanced production of 2-hydroxyphenazine. Microb. Cell Fact. 2016;15:1–12. doi: 10.1186/s12934-016-0529-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Van Rij E.T., Wesselink M., Chin-A-Woeng T.F., Bloemberg G.V., Lugtenberg B.J. Influence of environmental conditions on the production of phenazine-1-carboxamide by Pseudomonas chlororaphis PCL1391. Mol. Plant Microbe Interact. 2004;17:557–566. doi: 10.1094/MPMI.2004.17.5.557. [DOI] [PubMed] [Google Scholar]
- 23.Jin K., Zhou L., Jiang H., Sun S., Fang Y., Liu J., Zhang X., He Y.-W. Engineering the central biosynthetic and secondary metabolic pathways of Pseudomonas aeruginosa strain PA1201 to improve phenazine-1-carboxylic acid production. Metab. Eng. 2015;32:30–38. doi: 10.1016/j.ymben.2015.09.003. [DOI] [PubMed] [Google Scholar]
- 24.Mavrodi D.V., Bonsall R.F., Delaney S.M., Soule M.J., Phillips G., Thomashow L.S. Functional analysis of genes for biosynthesis of pyocyanin and phenazine-1-carboxamide from Pseudomonas aeruginosa pao1. J. Bacteriol. 2001;183:6454–6465. doi: 10.1128/JB.183.21.6454-6465.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cheng C., Ma G., Deng L., Li R. A Kinetic Model for Phenazine-1-Carboxylic Acid Production by Pseudomonas sp. M18G. Int. J. Clin. Exp. Med. 2017;10:6468–6474. [Google Scholar]
- 26.Li Y., Jiang H., Xu Y., Zhang X. Optimization of nutrient components for enhanced phenazine-1-carboxylic acid production by GACA-inactivated Pseudomonas sp. M18G using response surface method. App. Microbiol. Biotechnol. 2008;77:1207–1217. doi: 10.1007/s00253-007-1213-4. [DOI] [PubMed] [Google Scholar]
- 27.Bedoya J.C., Dealis M.L., Silva C.S., Niekawa E.T.G., Navarro M.O.P., Simionato A.S., Modolon F., Chryssafidis A.L., Andrade G. Enhanced Production of Target Bioactive Metabolites Produced by Pseudomonas aeruginosa LV Strain. Biocatal. Agric. Biotechnol. 2019;17:545–556. doi: 10.1016/j.bcab.2018.12.024. [DOI] [Google Scholar]
- 28.Abo-Zaid G.A., Wagih E.E., Matar S.M., Ashmawy N.A., Hafez E.E. Optimization of Pyocyanin Production from Pseudomonas aeruginosa JY21 Using Statistical Experimental Designs. Int. J. ChemTech Res. 2015;8:137–148. [Google Scholar]
- 29.Haggag W.M., Abo El Soud M. Production and optimization of Pseudomonas fluorescens biomass and metabolites for biocontrol of Strawberry Grey Mould. Am. J. Plant Sci. 2012;03:836–845. doi: 10.4236/ajps.2012.37101. [DOI] [Google Scholar]
- 30.Cui J., Wang W., Hu H., Zhang H., Zhang X. Enhanced phenazine-1-carboxamide production in Pseudomonas chlororaphis h5△fleq△rela through fermentation optimization. Fermentation. 2022;8:188. doi: 10.3390/fermentation8040188. [DOI] [Google Scholar]
- 31.Zhou Q., Su J., Jiang H., Huang X., Xu Y. Optimization of Phenazine-1-Carboxylic Acid Production by a GacA/QscR-Inactivated Pseudomonas sp. M18GQ Harboring PME6032Phz Using Response Surface Methodology. App. Microbiol. Biotechnol. 2010;86:1761–1773. doi: 10.1007/s00253-010-2464-z. [DOI] [PubMed] [Google Scholar]
- 32.Song C., Yue S.-J., Liu W.-H., Zheng Y.-F., Zhang C.-H., Feng T.-T., Hu H.-B., Wang W., Zhang X.-H. Engineering of glycerol utilization in Pseudomonas chlororaphis GP72 for enhancing phenazine-1-carboxylic acid production. World J. Microbiol. Biotechnol. 2020;36:1–8. doi: 10.1007/s11274-020-02824-3. [DOI] [PubMed] [Google Scholar]
- 33.Li Y., Jiang H., Du X., Huang X., Zhang X., Xu Y., Xu Y. Enhancement of phenazine-1-carboxylic acid production using batch and fed-batch culture of GacA inactivated Pseudomonas sp. M18G. Biores. Technol. 2010;101:3649–3656. doi: 10.1016/j.biortech.2009.12.120. [DOI] [PubMed] [Google Scholar]
- 34.Patil S., Nikam M., Patil H., Anokhina T., Kochetkov V., Chaudhari A. Bioactive pigment production by Pseudomonas spp. MCC 3145: Statistical media optimization, biochemical characterization, fungicidal and DNA intercalation-based cytostatic activity. Proc. Biochem. 2017;58:298–305. doi: 10.1016/j.procbio.2017.05.003. [DOI] [Google Scholar]
- 35.Ge Y., Huang X., Wang S., Zhang X., Xu Y. Phenazine-1-carboxylic acid is negatively regulated and pyoluteorin positively regulated by GacA in Pseudomona ssp. M18. FEMS Microbiol. Lett. 2004;237:41–47. doi: 10.1111/j.1574-6968.2004.tb09676.x. [DOI] [PubMed] [Google Scholar]
- 36.Yue S.-J., Huang P., Li S., Jan M., Hu H.-B., Wang W., Zhang X.-H. Enhanced production of 2-hydroxyphenazine from glycerol by a two-stage fermentation strategy in Pseudomonas chlororaphis GP72AN. J. Agric. Food Chem. 2019;68:561–566. doi: 10.1021/acs.jafc.9b05033. [DOI] [PubMed] [Google Scholar]
- 37.Slininger P.J., Shea-Wilbur M.A. Liquid-culture pH, temperature, and carbon (not nitrogen) source regulate phenazine productivity of the take-all biocontrol agent Pseudomonas fluorescens 2-79. App. Microbiol. Biotechnol. 1995;43:794–800. doi: 10.1007/BF02431910. [DOI] [PubMed] [Google Scholar]
- 38.McDonald M., Mavrodi D.V., Thomashow L.S., Floss H.G. Phenazine Biosynthesis in Pseudomonas fluorescens: Branchpoint from the Primary Shikimate Biosynthetic Pathway and Role of Phenazine-1,6-Dicarboxylic Acid. J. Am. Chem. Soc. 2001;123:9459–9460. doi: 10.1021/ja011243+. [DOI] [PubMed] [Google Scholar]
- 39.Wang S., Fu C., Bilal M., Hu H., Wang W., Zhang X. Enhanced biosynthesis of Arbutin by engineering shikimate pathway in Pseudomonas chlororaphis p3. Microb. Cell Fact. 2018;17:1–14. doi: 10.1186/s12934-018-1022-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wang S., Liu D., Bilal M., Wang W., Zhang X. Uncovering the role of phzc as DAHP synthase in shikimate pathway of Pseudomonas chlororaphis HT66. Biology. 2022;11:86. doi: 10.3390/biology11010086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Blankenfeldt W., Kuzin A.P., Skarina T., Korniyenko Y., Tong L., Bayer P., Janning P., Thomashow L.S., Mavrodi D.V. Structure and function of the phenazine biosynthetic protein PhzF from Pseudomonas fluorescens. Proc. Natl. Acad. Sci. USA. 2004;101:16431–16436. doi: 10.1073/pnas.0407371101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Parsons J.F., Song F., Parsons L., Calabrese K., Eisenstein E., Ladner J.E. Structure and function of the phenazine biosynthesis protein PhzF from Pseudomonas fluorescens 2-79. Biochemistry. 2004;43:12427–12435. doi: 10.1021/bi049059z. [DOI] [PubMed] [Google Scholar]
- 43.Ahuja E.G., Janning P., Mentel M., Graebsch A., Breinbauer R., Hiller W., Costisella B., Thomashow L.S., Mavrodi D.V., Blankenfeldt W. PhzA/B catalyzes the formation of the tricycle in phenazine biosynthesis. J. Am. Chem. Soc. 2008;130:17053–17061. doi: 10.1021/ja806325k. [DOI] [PubMed] [Google Scholar]
- 44.Guo S., Wang Y., Dai B., Wang W., Hu H., Huang X., Zhang X. PhzA, the shunt switch of phenazine-1,6-dicarboxylic acid biosynthesis in Pseudomonas chlororaphis HT66. App. Microb. Biotechnol. 2017;101:7165–7175. doi: 10.1007/s00253-017-8474-3. [DOI] [PubMed] [Google Scholar]
- 45.Guo S., Wang Y., Bilal M., Hu H., Wang W., Zhang X. Microbial synthesis of antibacterial phenazine-1,6-dicarboxylic acid and the role of PhzG in Pseudomonas chlororaphis GP72AN. J. Agric. Food Chem. 2020;68:2373–2380. doi: 10.1021/acs.jafc.9b07657. [DOI] [PubMed] [Google Scholar]
- 46.Delaney S.M., Mavrodi D.V., Bonsall R.F., Thomashow L.S. PhzO, a gene for biosynthesis of 2-hydroxylated phenazine compounds in Pseudomonas aureofaciens 30-84. J. Bacteriol. 2001;183:318–327. doi: 10.1128/JB.183.1.318-327.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Xu N., Ahuja E.K., Janning P., Mavrodi D.V., Thomashow L.S., Blankenfeldt W. Trapped intermediates in crystals of the FMN-dependent oxidase PhzG provide insight into the final steps of phenazine biosynthesis. Acta Crystallog. D Biol. Crystallogr. 2013;69:1403–1413. doi: 10.1107/S0907444913008354. [DOI] [PubMed] [Google Scholar]
- 48.Huang L., Huang Y., Lou Y., Qian H., Xu D., Ma L., Jiang C., Zhang D. Pyocyanin-modifying genes phzM and phzS regulated the extracellular electron transfer in microbiologically-influenced corrosion of X80 carbon steel by Pseudomonas aeruginosa. Corros. Sci. 2020;164:108355. doi: 10.1016/j.corsci.2019.108355. [DOI] [Google Scholar]
- 49.Chin-A-Woeng T.F., Thomas-Oates J.E., Lugtenberg B.J., Bloemberg G.V. Introduction of the phzH gene of Pseudomonas chlororaphis PCL1391 extends the range of biocontrol ability of phenazine-1-carboxylic acid-producing Pseudomonas spp. strains. Mol. Plant Microbe Interact. 2001;14:1006–1015. doi: 10.1094/MPMI.2001.14.8.1006. [DOI] [PubMed] [Google Scholar]
- 50.Peng H., Ouyang Y., Bilal M., Wang W., Hu H., Zhang X. Identification, synthesis and regulatory function of the N-acylated homoserine lactone signals produced by Pseudomonas chlororaphis HT66. Microb. Cell Fact. 2018;17:1–11. doi: 10.1186/s12934-017-0854-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Chen M., Cao H., Peng H., Hu H., Wang W., Zhang X. Reaction kinetics for the biocatalytic conversion of phenazine-1-carboxylic acid to 2-hydroxyphenazine. PLoS ONE. 2014;9:e98537. doi: 10.1371/journal.pone.0098537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Dasgupta D., Kumar A., Mukhopadhyay B., Sengupta T.K. Isolation of phenazine 1,6-di-carboxylic acid from Pseudomonas aeruginosa strain HRW.1-S3 and its role in biofilm-mediated crude oil degradation and cytotoxicity against bacterial and cancer cells. App. Microbiol. Biotechnol. 2015;99:8653–8665. doi: 10.1007/s00253-015-6707-x. [DOI] [PubMed] [Google Scholar]
- 53.Mavrodi D.V., Peever T.L., Mavrodi O.V., Parejko J.A., Raaijmakers J.M., Lemanceau P., Mazurier S., Heide L., Blankenfeldt W., Weller D.M., et al. Diversity and evolution of the phenazine biosynthesis pathway. App. Environ. Microbiol. 2010;76:866–879. doi: 10.1128/AEM.02009-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Pierson L.S., III., Wood D.W., Pierson E.A. Homoserine lactone-mediated gene regulation in plant-associated bacteria. Annu. Rev. Phytopathol. 1998;36:207–225. doi: 10.1146/annurev.phyto.36.1.207. [DOI] [PubMed] [Google Scholar]
- 55.De Maeyer K., D’aes J., Hua G.K., Perneel M., Vanhaecke L., Noppe H., Höfte M. N-acylhomoserine lactone quorum-sensing signalling in antagonistic phenazine-producing Pseudomonas isolates from the red cocoyam rhizosphere. Microbiology. 2011;157:459–472. doi: 10.1099/mic.0.043125-0. [DOI] [PubMed] [Google Scholar]
- 56.Haas D., Défago G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005;3:307–319. doi: 10.1038/nrmicro1129. [DOI] [PubMed] [Google Scholar]
- 57.Bourret R.B., Silversmith R.E. Two-component signal transduction. Curr. Opin. Microbiol. 2010;13:113–115. doi: 10.1016/j.mib.2010.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Bauer J.S., Hauck N., Christof L., Mehnaz S., Gust B., Gross H. The systematic investigation of the Quorum Sensing system of the biocontrol strain Pseudomonas chlororaphis subsp. aurantiaca PB-ST2 unveils auri to be a biosynthetic origin for 3-oxo-homoserine lactones. PLoS ONE. 2016;11:e0167002. doi: 10.1371/journal.pone.0167002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Morohoshi T., Yamaguchi T., Xie X., Wang W.Z., Takeuchi K., Someya N. Complete genome sequence of Pseudomonas chlororaphis subsp. aurantiaca reveals a triplicate quorum-sensing mechanism for regulation of phenazine production. Microbes Environ. 2017;32:47–53. doi: 10.1264/jsme2.ME16162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Heeb S., Haas D. Regulatory roles of the GacS/GacA two-component system in plant-associated and other gram-negative bacteria. Mol. Plant Microbe Interact. 2001;14:1351–1363. doi: 10.1094/MPMI.2001.14.12.1351. [DOI] [PubMed] [Google Scholar]
- 61.Wang D., Yu J.M., Pierson L.S., Pierson E.A. Differential regulation of phenazine biosynthesis by RPEA and RpeB in Pseudomonas chlororaphis 30-84. Microbiology. 2012;158:1745–1757. doi: 10.1099/mic.0.059352-0. [DOI] [PubMed] [Google Scholar]
- 62.Girard G., van Rij E.T., Lugtenberg B.J., Bloemberg G.V. Regulatory roles of PSRA and rpoS in phenazine-1-carboxamide synthesis by Pseudomonas chlororaphis PCL1391. Microbiology. 2006;152:43–58. doi: 10.1099/mic.0.28284-0. [DOI] [PubMed] [Google Scholar]
- 63.Morohoshi T., Yabe N., Yaguchi N., Xie X., Someya N. Regulation of phenazine-1-carboxamide production by quorum sensing in type strains of Pseudomonas chlororaphis subsp. chlororaphis and Pseudomonas chlororaphis subsp. piscium. J. Biosci. Bioeng. 2022;133:541–546. doi: 10.1016/j.jbiosc.2022.03.004. [DOI] [PubMed] [Google Scholar]
- 64.Jin X.-J., Peng H.-S., Hu H.-B., Huang X.-Q., Wang W., Zhang X.-H. ITRAQ-based quantitative proteomic analysis reveals potential factors associated with the enhancement of phenazine-1-carboxamide production in Pseudomonas chlororaphis p3. Sci. Rep. 2016;6:27393. doi: 10.1038/srep27393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Yuan L.-L., Li Y.-Q., Wang Y., Zhang X.-H., Xu Y.-Q. Optimization of critical medium components using response surface methodology for phenazine-1-carboxylic acid production by Pseudomonas sp. M-18Q. J. Biosci. Bioeng. 2008;105:232–237. doi: 10.1263/jbb.105.232. [DOI] [PubMed] [Google Scholar]
- 66.He L., Xu Y.-Q., Zhang X.-H. Medium factor optimization and fermentation kinetics for phenazine-1-carboxylic acid production by Pseudomonas sp. M18G. Biotechnol. Bioeng. 2008;100:250–259. doi: 10.1002/bit.21767. [DOI] [PubMed] [Google Scholar]
- 67.Aloui H., Khomlaem C., Torres C.A.V., Freitas F., Reis M.A.M., Kim B.S. Enhanced co-production of medium-chain-length polyhydroxyalkanoates and phenazines from crude glycerol by high cell density cultivation of Pseudomonas chlororaphis in membrane bioreactor. Int. J. Biol. Macromol. 2022;211:545–555. doi: 10.1016/j.ijbiomac.2022.05.089. [DOI] [PubMed] [Google Scholar]
- 68.Bilal M., Yue S.-J., Hu H.-B., Wang W., Zhang X.-H. Adsorption/desorption characteristics, separation and purification of phenazine-1-carboxylic acid from fermentation extract by macroporous adsorbing resins. J. Cheml. Technol. Biotechnol. 2018;93:3176–3184. doi: 10.1002/jctb.5673. [DOI] [Google Scholar]
- 69.Jasim B., Anisha C., Rohini S., Kurian J.M., Jyothis M., Radhakrishnan E.K. Phenazine carboxylic acid production and rhizome protective effect of endophytic Pseudomonas aeruginosa isolated from Zingiber officinale. World J. Microbiol. Biotechnol. 2013;30:1649–1654. doi: 10.1007/s11274-013-1582-z. [DOI] [PubMed] [Google Scholar]
- 70.Shao J., Fan L.-Y., Zhang W., Guo C.-G., Li S., Xu Y.-Q., Cao C.-X. Purification of low-concentration phenazine-1-carboxylic acid from fermentation broth of Pseudomonas sp. M18 via free flow electrophoresis with Gratis gravity. Electrophoresis. 2010;31:3499–3507. doi: 10.1002/elps.201000213. [DOI] [PubMed] [Google Scholar]
- 71.Mehnaz S. Characterization of a phenazine and hexanoyl homoserine lactone producing Pseudomonas aurantiaca strain PB-st2, isolated from sugarcane stem. J. Microbiol. Biotechnol. 2009;19:1688–1694. doi: 10.4014/jmb.0904.04022. [DOI] [PubMed] [Google Scholar]
- 72.Shanmugaiah V., Mathivanan N., Varghese B. Purification, crystal structure and antimicrobial activity of phenazine-1-carboxamide produced by a growth-promoting biocontrol bacterium, Pseudomonas aeruginosa MML2212. J. App. Microbiol. 2010;108:703–711. doi: 10.1111/j.1365-2672.2009.04466.x. [DOI] [PubMed] [Google Scholar]
- 73.Aziz L.M., Hamza S.J. Isolation and characterization of phenazine produced from mutant Pseudomonas aeruginosa. Al-Anbar J. Vet. Sci. 2012;5:42–53. [Google Scholar]
- 74.Gorantla J.N., Nishanth Kumar S., Nisha G.V., Sumandu A.S., Dileep C., Sudaresan A., Sree Kumar M.M., Lankalapalli R.S., Kumar B.S. Purification and characterization of antifungal phenazines from a fluorescent Pseudomonas strain FPO4 against medically important fungi. J. Mycol. Médicale. 2014;24:185–192. doi: 10.1016/j.mycmed.2014.02.003. [DOI] [PubMed] [Google Scholar]
- 75.Joshi D.R., Adhikari N. An overview on common organic solvents and their toxicity. J. Pharm. Res. Int. 2019;28:1–18. doi: 10.9734/jpri/2019/v28i330203. [DOI] [Google Scholar]
- 76.Rane M.R., Sarode P.D., Chaudhari B.L., Chincholkar S.B. Detection, isolation and identification of phenazine-1-carboxylic acid produced by biocontrol strains of Pseudomonas Aeruginosa. J. Sci. Ind. Res. 2007;66:627–631. [Google Scholar]
- 77.Chen K., Hu H., Wang W., Zhang X., Xu Y. Metabolic degradation of phenazine-1-carboxylic acid by the strain Sphingomonas sp. DP58: The identification of two metabolites. Biodegradation. 2007;19:659–667. doi: 10.1007/s10532-007-9171-1. [DOI] [PubMed] [Google Scholar]
- 78.Liu H., He Y., Jiang H., Peng H., Huang X., Zhang X., Thomashow L.S., Xu Y. Characterization of a phenazine-producing strain Pseudomonas chlororaphis GP72 with broad-spectrum antifungal activity from green pepper rhizosphere. Curr. Microbiol. 2007;54:302–306. doi: 10.1007/s00284-006-0444-4. [DOI] [PubMed] [Google Scholar]
- 79.Thacharodi A., Priyadharshini R., Karthikeyan G., Jeganathan C., Reghu A.P., Pugazhendhi A. Extraction, purification and characterization of phenazine from Pseudomonas aeruginosa isolate of wastewater sources: A panacea towards clinical pathogens. App. Nanosci. 2021 doi: 10.1007/s13204-021-01944-y. [DOI] [Google Scholar]
- 80.Liu H., Zhang X., Huang X., Cao C., Xu Y. Rapid quantitative analysis of phenazine-1-carboxylic acid and 2-hydroxyphenazine from fermentation culture of Pseudomonas chlororaphis GP72 by capillary zone electrophoresis. Talanta. 2008;76:276–281. doi: 10.1016/j.talanta.2008.02.034. [DOI] [PubMed] [Google Scholar]
- 81.Zhou K., Kammarchedu V., Butler D., Soltan Khamsi P., Ebrahimi A. Electrochemical sensors based on MOS x -functionalized laser-induced graphene for real-time monitoring of phenazines produced by Pseudomonas aeruginosa. Adv. Health Mat. 2022;11:2200773. doi: 10.1002/adhm.202200773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kern S.E., Newman D.K. Measurement of phenazines in bacterial cultures. Methods Mol. Biol. 2014;1149:303–310. doi: 10.1007/978-1-4939-0473-0_25. [DOI] [PubMed] [Google Scholar]
- 83.Simionato A.S., Navarro M.O., de Jesus M.L., Barazetti A.R., da Silva C.S., Simões G.C., Balbi-Peña M.I., de Mello J.C., Panagio L.A., de Almeida R.S., et al. The effect of phenazine-1-carboxylic acid on mycelial growth of botrytis cinerea produced by Pseudomonas aeruginosa LV strain. Front. Microbiol. 2017;8:1102. doi: 10.3389/fmicb.2017.01102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Pierson L.S., Pierson E.A. Phenazine antibiotic production in Pseudomonas aureofaciens: Role in rhizosphere ecology and pathogen suppression. FEMS Microbiol. Lett. 1996;136:101–108. doi: 10.1111/j.1574-6968.1996.tb08034.x. [DOI] [Google Scholar]
- 85.Castaldi S., Masi M., Sautua F., Cimmino A., Isticato R., Carmona M., Tuzi A., Evidente A. Pseudomonas fluorescens showing antifungal activity against Macrophomina phaseolina, a severe pathogenic fungus of soybean, produces phenazine as the main active metabolite. Biomolecules. 2021;11:1728. doi: 10.3390/biom11111728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Raio A., Puopolo G. Pseudomonas chlororaphis metabolites as biocontrol promoters of plant health and improved crop yield. World J. Microbiol. Biotechnol. 2021;37:99. doi: 10.1007/s11274-021-03063-w. [DOI] [PubMed] [Google Scholar]
- 87.Halicki P.C.B., da Silva E.N., Jardim G.A.D.M., Almeida R.G.D., Vicenti J.R.D.M., Gonçalves B.L., da Silva P.E.A., Ramos D.F. Benzo[a]phenazine derivatives: Promising scaffolds to combat resistant Mycobacterium tuberculosis. Chem. Biol. Drug Des. 2021;98:352–362. doi: 10.1111/cbdd.13853. [DOI] [PubMed] [Google Scholar]
- 88.Huigens R.W., Abouelhassan Y., Yang H. Phenazine antibiotic-inspired discovery of bacterial biofilm-eradicating agents. Chem. Bio. Chem. 2019;20:2885–2902. doi: 10.1002/cbic.201900116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Saosoong K., Wongphathanakul W., Poasiri C., Ruangviriyachai C. Isolation and analysis of antibacterial substance produced from P. aeruginosa TISTR 781. KKU Sci. J. 2009;37:163–172. [Google Scholar]
- 90.Giddens S.R., Bean D.C. Investigations into the in vitro antimicrobial activity and mode of action of the phenazine antibiotic D-alanylgriseoluteic acid. Int. J. Antimicrob. Agents. 2007;29:93–97. doi: 10.1016/j.ijantimicag.2006.08.028. [DOI] [PubMed] [Google Scholar]
- 91.Park G.K. Elucidation of antifungal metabolites produced by Pseudomonas aurantiaca IB5-10 with broad-spectrum antifungal activity. J. Microbiol. Biotechnol. 2014;22:326–330. doi: 10.4014/jmb.1106.06042. [DOI] [PubMed] [Google Scholar]
- 92.George E., Kumar S.N., Jacob J., Bommasani B., Lankalapalli R.S., Morang P., Kumar B.S. Characterization of the bioactive metabolites from a plant growth-promoting rhizobacteria and their exploitation as antimicrobial and plant growth-promoting agents. App. Biochem. Biotechnol. 2015;176:529–546. doi: 10.1007/s12010-015-1593-3. [DOI] [PubMed] [Google Scholar]
- 93.Chen Y., Norrman A., Ponomarenko S.A., Friberg A.T. Coherence lattices in surface plasmon Polariton Fields. Opt. Lett. 2018;43:3429. doi: 10.1364/OL.43.003429. [DOI] [PubMed] [Google Scholar]
- 94.Liu T.T., Ye F.C., Pang C.P., Yong T.Q., Tang W.D., Xiao J., Shang C.H., Lu Z.J. Isolation and identification of bioactive substance 1-hydroxyphenazine from Pseudomonas aeruginosa and its antimicrobial activity. Lett. App. Microbiol. 2020;71:303–310. doi: 10.1111/lam.13332. [DOI] [PubMed] [Google Scholar]
- 95.Karmegham N., Vellasamy S., Natesan B., Sharma M.P., Al Farraj D.A., Elshikh M.S. Characterization of antifungal metabolite phenazine from rice rhizosphere fluorescent pseudomonads (FPS) and their effect on sheath blight of rice. Saudi J. Biol. Sci. 2020;27:3313–3326. doi: 10.1016/j.sjbs.2020.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Morales D.K., Jacobs N.J., Rajamani S., Krishnamurthy M., Cubillos-Ruiz J.R., Hogan D.A. Antifungal mechanisms by which a novel pseudomonas aeruginosa phenazine toxin kills Candida albicans in biofilms. Mol. Microbiol. 2010;78:1379–1392. doi: 10.1111/j.1365-2958.2010.07414.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Liu X.-W., Liu N.-Y., Deng Y.-Q., Wang S., Liu T., Tang Y.-C., Chen Y.-D., Lu J.-L. Anticancer activity, topoisomerase I inhibition, DNA ‘light switch’ behavior and molecular docking of two ruthenium complexes containing phenazine ring. J. Biomol. Struct. Dyn. 2020;39:5953–5962. doi: 10.1080/07391102.2020.1796806. [DOI] [PubMed] [Google Scholar]
- 98.Hollstein U., Van Gemert R.J. Interaction of phenazines with Polydeoxyribonucleotides. Biochemistry. 1971;10:497–504. doi: 10.1021/bi00779a023. [DOI] [PubMed] [Google Scholar]
- 99.Kennedy R.K., Naik P.R., Veena V., Lakshmi B.S., Lakshmi P., Krishna R., Sakthivel N. 5-methyl phenazine-1-carboxylic acid: A novel bioactive metabolite by a rhizosphere soil bacterium that exhibits potent antimicrobial and anticancer activities. Chem. Biol. Interact. 2015;231:71–82. doi: 10.1016/j.cbi.2015.03.002. [DOI] [PubMed] [Google Scholar]
- 100.Vicker N., Burgess L., Chuckowree I.S., Dodd R., Folkes A.J., Hardick D.J., Hancox T.C., Miller W., Milton J., Sohal S., et al. Novel angular benzophenazines: dual Topoisomerase I and Topoisomerase II inhibitors as potential anticancer agents. J. Med. Chem. 2002;45:721–739. doi: 10.1021/jm010329a. [DOI] [PubMed] [Google Scholar]
- 101.Kato S., Shindo K., Yamagishi Y., Matsuoka M., Kawai H., Mochizuki J. Phenazoviridin, a novel free radical scavenger from Streptomyces sp. taxonomy, fermentation, isolation, structure elucidation and biological properties. J. Antibiot. 1993;46:1485–1493. doi: 10.7164/antibiotics.46.1485. [DOI] [PubMed] [Google Scholar]
- 102.Laxmi M., Bhat S.G. Characterization of pyocyanin with radical scavenging and antibiofilm properties isolated from Pseudomonas aeruginosa strain BTRY1. 3 Biotech. 2016;6:1–5. doi: 10.1007/s13205-015-0350-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Xiong Z., Niu J., Liu H., Xu Z., Li J., Wu Q. Synthesis and bioactivities of phenazine-1-carboxylic acid derivatives based on the modification of PCA Carboxyl Group. Bioorg. Med. Chem. Lett. 2017;27:2010–2013. doi: 10.1016/j.bmcl.2017.03.011. [DOI] [PubMed] [Google Scholar]
- 104.Peng D., Li S., Wang J., Chen C., Zhou M. Integrated biological and chemical control of rice sheath blight by bacillus subtilisNJ-18 and pingyangmycin. Pest Manag. Sci. 2013;70:258–263. doi: 10.1002/ps.3551. [DOI] [PubMed] [Google Scholar]
- 105.Zhu X., Yu L., Hsiang T., Huang D., Xu Z., Wu Q., Du X., Li J. The influence of steric configuration of phenazine-1-carboxylic acid-amino acid conjugates on fungicidal activity and systemicity. Pest Manag. Sci. 2019;75:3323–3330. doi: 10.1002/ps.5455. [DOI] [PubMed] [Google Scholar]
- 106.Ali S., Hameed S., Shahid M., Iqbal M., Lazarovits G., Imran A. Functional characterization of potential PGPR exhibiting broad-spectrum antifungal activity. Microbiol. Res. 2020;232:126389. doi: 10.1016/j.micres.2019.126389. [DOI] [PubMed] [Google Scholar]
- 107.Ou J., Li H., Ou X., Yang Z., Chen M., Liu K., Teng Y., Xing B. Degradation, adsorption and leaching of phenazine-1-carboxamide in agrigulture soils. Ecotoxicol. Environ. Saf. 2020;205:111375. doi: 10.1016/j.ecoenv.2020.111374. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Not applicable.