

Abstract
Probiotics are defined as live microbial food elements that are beneficial to human health. Lacticaseibacillus casei T1 was considered to have potential as a bioactive ingredient in functional foods, which was isolated from kurut. Previous research by our group proved that L. casei T1 could prevent inflammatory responses caused by Helicobacter pylori. This study aimed to investigate whether treatment with L. casei T1 resulted in a suppressive effect on H. pylori-induced oxidative stress and inflammatory responses. The results showed that treatment with L. casei T1 could relieve H. pylori-induced overexpression of inflammatory cytokines in GES-1 cells. Experiments in animals suggested that taking long-term L. casei T1 could reduce oxidative stress and inflammatory cytokines and improve H. pylori-induced gastric mucosal damage. Furthermore, taking L. casei T1 could increase the relative abundance of beneficial intestinal bacterium (Lachnospiraceae and Odoribacter) of H. pylori-infected mice and help in maintaining the balance of intestinal microflora.
Collectively, L. casei T1 had certain degrees of therapeutic effect against H. pylori. In the future, it combined with antibiotics for H. pylori eradication deserves further study.
Keywords: H. pylori, Lacticaseibacillus casei, Probiotics, Inflammation, Oxidative stress
Introduction
Helicobacter pylori, a strain of microaerobic bacteria, is a curved spiral-shaped bacterium isolated from the stomach [1]. More than half of the world population lives with H. pylori, with an infection rate of about 25% in developed countries and 80% in developing countries [2]. And H. pylori is recognized as the most common cause of chronic gastritis, gastric ulcer, duodenal ulcer and is also an important pathogenic factor in gastric cancer [3, 4]. It is considered a class I human carcinogen [5, 6]. A triple or quadruple regimen is a routine treatment for H. pylori infection [7–9]. Along with extensive use of antibiotics in clinical and agricultural settings over the past few decades, the ever-increasing number of drug-resistant H. pylori and lack of alternative antibiotics lead to increasing difficulties in eradication therapy. Reinfection rates are also high in developing countries [10]. Moreover, misuse and overuse of antibiotics not only led to many adverse consequences, for instance diarrhea, nausea and drug eruption, but also resulted in an imbalance in the gastrointestinal microbiota [2, 11]. Therefore, it is necessary to develop new complementary therapeutic strategies. Currently, probiotics are extensively studied for their beneficial effects in prevention and treatment of many prevailing diseases [12]. Probiotics have antimicrobial activity, produce bacteriocins, have antitoxin effects, enhance the intestinal barrier function and exercise immune modulation [13]. Because of the multiple bactericidal mechanisms of probiotics, taking them does not cause bacterial resistance. Thus, probiotic adjuvant therapy may become a new choice in H. pylori infection [14–16].
Probiotics can colonize the gastrointestinal tract and beneficially affect the host by balancing the gastrointestinal flora, reducing inflammation and improving immune function [17]. In contrast to the currently used antibiotics, bacteriocins generated by lactic acid bacteria (LAB) are often considered natural and safe, because they have been naturally present in fermented foods [18]. LAB is also used as fermenting agent, and they are extensively used in the manufacture of fermented milk products, wine and pickled vegetables [19, 20]. The safety of these microorganisms has not been questioned, and reports of these bacteria being harmful are very rare [18]. Studies in vitro have documented two strains of Limosilactobacillus reuteri isolated from the human body have good viability under gastrointestinal conditions, as well as strong resistance to H. pylori and antioxidant activity [21]. Studies showed that H. pylori infection was effectively treated and controlled, when a fermented milk-based product which contains Bifidobacterium lactis Bb12 and Lactobacillus acidophilus Bb12 was consumed for six weeks [21]. Some research verified ingestion of probiotics alone could reduce colonization of H. pylori, but it couldn't be completely excluded from the stomach. The eradication rate of H. pylori could be enhanced by 10% when probiotics combined with antibiotic [22–24]. In addition, taking probiotics can reduce the side effects of antibiotic treatment [25]. It is well accepted that long-term consumption of food containing probiotics is beneficial to human health.
Lacticaseibacillus casei T1 is a probiotic strain isolated from kurut described from Qinghai-Tibet plateau, which has excellent acid tolerance and a high survival rate in the stomach [26, 27]. The acid generating capacity of L. casei T1 is strong and can reduce the pH of the fermentation broth to well below 4.0 after 48 h incubation. It also can produce an extracellular bacteriocin that possesses the activities against drug-resistant bacteria, which has high thermal and pH stability [26]. Our previous studies have confirmed that taking L. casei T1 in advance can contribute to ameliorates inflammatory response caused by H. pylori in vivo [28]. Nevertheless, the difference between prevention and treatment might be that the latter provides a more direct and visible effect. In the present study, we investigated the therapeutic effect of L. casei T1 after H. pylori infection in vivo.
Materials and methods
Bacterial strains and their cultivation
All the strains were derived from Key Laboratory of Biological Resource and Ecological Environment of Chinese Education Ministry. L. casei T1 was cultured on MRS agar solid medium or MRS liquid medium for 24 h at 37℃. The strain of H. pylori used in this paper is a clinical strain isolated from a patient with gastric ulceration and moderate gastritis at the People's Hospital of Sichuan Province.
H. pylori and the remaining strains were cultured according to the method reported by Wu et al. [28] and Luo et al. [26]. Escherichia coli (China Center of Industrial Culture Collection; CICC 10,354) and Staphylococcus aureus (CICC 10,201) were isolated from spoiled milk. Escherichia coli, Salmonella typhimurium, Staphylococcus aureus, Bacillus subtilis and Bacillus cereus were propagated in LB broth (Qingdao Haibo Biotechnology Co., Ltd, HB0128) at 37℃ for 24 h. MRS Broth medium (Qingdao Haibo Biotechnology Co., Ltd, HB0384-1) was used to culture Lactobacillus bulgaricus, Lactobacillus acidophilus, and Levilactobacillus brevis for 24 h at 37℃.
Bacteriostatic spectrum detection
L. casei T1 was cultured for 48 h at 37℃ in MRS liquid medium and the supernatant were collected by centrifugation at 8,000 g for 10 min. Then it was filtered with a 0.22um filter to remove residual organisms. Then it was tested for antimicrobial activities by the well-diffusion assay [29]. The melted MRS agar medium was first poured onto the plate and solidified as a bottom layer. Then, 1 mL of the bacterial solution (1 × 107 cfu/mL) was added, and LB agar medium was added at about 50℃. Shake the plate gently to mix and set horizontally until set. Holes were punched by Sterile hole punchers (d = 7 mm) and 100 µL L. casei T1 fermentation supernatant was added to each hole and cultured at 37℃ for 24 h. Bacteriostatic activity was determined by measuring the diameter of inhibitory zones.
Cell culture
GES-1 cells are used as a reliable in vitro model of human gastric epithelium. The source and culture conditions of GES-1 cells refer to the report of Wu et al. [28]. GES-1 cells (5 × 105 cells/well) were seeded in 6-well plates and cultured 24 h. Then, the GES-1 cells were co-cultured with H. pylori (5 × 107 Colony Forming Unit, CFU/well) or L. casei T1 (5 × 107 CFU/well) for 24 h. Afterwards, the GES-1 cells co-cultured with H. pylori were treated with L. casei T1 (5 × 107 CFU/well) or amoxicillin (5 mg/L) for 1 day. The gathered cells and supernatants were used for qPCR and Enzyme Linked Immunosorbent Assay (ELISA).
Animal experiments
Eight-ten weeks old female Balb/c mice were purchased from Chengdu Dashuo Experimental Animal Co., Ltd. (Chengdu, China). Mice were housed in standard conditions with food and water ad libitum in the conventional vivarium. After adaptive feeding for 7 days, 30 mice were randomized into 2 groups receiving the first pretreatment as described below: ① Group C(8 mice): 500 μL normal saline were orally perfused to mice once every other day for 4 weeks; ② Group H(22 mice): mice were administered 500 µl H. pylori suspension (2 × 109 cfu/mL) by intragastric gavage alternate day for 4 weeks. After the first pretreatment, 6 randomly selected mice in Group H were fasted overnight for gastric tissue sampling. Prior literature suggested DNA of H. pylori could be discovered in infected antrum stomach tissues [30]. In order to evaluate the status of H. pylori infection, gastric tissues were used for routine rapid urease testing and polymerase chain reaction (PCR). DNA was extracted by TIANamp Bacteria DNA Kit purchased from Tiangen Biotech Co., Ltd. The extracted DNA was subject to PCR using specific primers of the H. pylori vacA gene. Primers were F-vacA: GCCGATATGCAAATGAGCCGC, R-vacA: CAATCGTGTGGGTTCTGGAGC. The products were identified by 1% agarose gel electrophoresis and anticipated being 678 base pairs.
When the H. pylori infection model was successfully established, 8 mice were arbitrarily selected from Group H and assigned to Group H + T1. Meanwhile, the mice underwent the second treatment as described below: ① Group C(8 mice) and Group H(8 mice): 500 μL normal saline were orally perfused once daily for 2 weeks; ② Group H + T1(8 mice): 500 μL bacterial suspension of L. casei T1 (2 × 109 cfu/mL) was administered by oral gavages once a day for 2 weeks. Fecal samples were collected by placing the mice individually in aseptic cages on the last day of the gavage. Mice were euthanized after 12 h of fasting at the end point of the experiment to collect blood, and tissues. 100 ~ 200 μL of blood was collected in anticoagulant tubes (Ethylene Diamine Tetraacetic Acid, EDTA) for blood routine examinations. The rest of blood collected in anticoagulant-free tubes was allowed to clot overnight at 4℃. And serum was prepared by centrifugation at 3000 g at 4℃ for 20 min, stored at − 80℃. The gastric tissue was collected for qPCR, the rapid urease test, Hematoxylin and eosin (HE) staining and immunohistochemical (IHC) experiments.
Analysis of gene expression
Total RNA was extracted by an RNA extraction kit (Thermo Fisher Scientific, PureLink™ RNA, 12183018A). RNA was reverse transcribed by Reverse Transcription kit (TaKaRa, PrimeScript RT reagent Kit with gDNA Eraser, RR047A). SYBR Green real time PCR was performed using QuantStudio3. Reactions were run at 50℃ for 2 min; 95℃for 30 s; then 40 cycles at 95℃ for 15 s, 60℃ for 30 s and 72℃ for 10 s. The GAPDH gene was used as an internal reference gene. Primers of selecting genes for qPCR designed through Oligo7. The primers of GES-1 were as follows: F-GAPDH TGCACCACCAACTGCTTAGC, R-GAPDH GGCATGGACTGTGGTCATGAG, F- tumor necrosis factor (TNF-α) GAGGCCAAGCCCTGGTATG, R-TNF-α CGGGCCGATTGATCTCAGC, F- interleukin 6 (IL6) CTTCGGTCCAGTTGCCTTCT, R-IL6 TGGAATCTTCTCCTGGGGGT, F-IL8 ACACTGCGCCAACACAGAAA, R-IL8 CAACCCTCTGCACCCAGTTT. The primers of mice were as follows: F-GAPDH ATG-GTGAAGGTCGGTGTGAAC, R-GAPDHGGAGTCATACTGGAACAT-GTAGACC, F-TNF-α CCTGTAGCCCACGTCGTAG, R-TNF-α GGGAGTAGACAAGGTACAACC-C, F-MUC5AC CTGTGACATTATCCCATAAGCCC, R-MUC5AC AAGGGGTATAGCTGGCCTGA.
Analysis of protein expression
The cell supernatants were detected by Human IL6 ELISA KIT, Human TNF-α ELISA KIT, Human IL8 ELISA KIT and Human C–C motif chemokine ligand 2 (CCL2) ELISA KIT purchased from Chengdu Pengshida Experimental Supplies Co., Ltd. The stored serum samples were thawed on ice. Subsequently, the concentrations of CRP, MDA, GSH-PX and SOD in serum were quantified by MOUSE ELISA kits from the same company.
Routine blood examination
As mentioned earlier, the blood samples just collected in anticoagulant tubes (EDTA) was sent to Harmony Yijia Pet Hospital (chengdu, China) within two hours, and the routine blood examination was performed by Celltac Alpha MEK-6400 series hematology analyzers (Nihon Kohden, MEK-6400).
HE staining and IHC experiments
HE staining and IHC staining experiments were performed according to standard histological protocols. Anti NF-κB/p65 was purchased from Wanleibio Co., Ltd. Goat anti-rabbit secondary antibody was purchased from Beyotime Biotechnology Co., Ltd.
Gut microbiota analysis
The collected fecal samples were shipped to Beijing Baimaike Biotechnology Co., Ltd on dry ice for 16S ribosomal DNA sequencing and analyses. 338F and 806R were used to amplify the V3-V4 region of the 16S rRNA gene.
Statistics
Data are expressed as means ± SD. Prism 8.0.2 software was used for statistics analysis. To determine the statistical significance, P values were determined by ANOVA or Student’s t-test. P < 0.05 was considered as significant.
Results
The antibacterial spectrum of fermentation broth
The effect of the bacteriostatic experiment is shown in Table 1. The fermentation liquor of L. casei T1 exhibited comparatively good antibacterial activity against resistant strains of S. aureus and E. coli. Further analysis showed that L. casei T1 had antagonistic effects not only on lactic acid bacteria with a high degree of relatedness, but also on gram-positive bacteria and gram-negative bacteria. Excessive proliferation of harmful bacterium reduces food shelf life and increases foodborne illness [31]. Microbial contamination will bring health problems and huge economic losses [32]. Nevertheless, the public demands to reduce the use of chemical preservatives or additives in food, increasing attention is being paid to the natural additives [33, 34]. So the antimicrobial activity of L. casei T1 indicated it could be used in agriculture and food applications. The fermentation liquor of L. casei T1 was also active in repressing H. pylori growth. The above results showed broad spectrum antimicrobial activity of L. casei T1’s fermentation broth.
Table 1.
Antibacterial spectrum of L. casei T1’s fermentation broth
| Indicator bacteria | Bacteriostatic activity | Indicator bacteria | Bacteriostatic activity |
|---|---|---|---|
| H. pylori | + + | B. cereus | + + |
| Escherichia coli | + + | L. bulgaricus | + + |
| S. typhimurium | + + | L. acidophilus | + + |
| S. aureus | + + + | L. brevis | + |
| B. subtilis | + + + |
‘ + , + + , + + + ’ represents the size of the bacteriostatic circle diameter. ‘ + + + ’: the diameter is over 15 mm, ‘ + + ’: the diameter is 10-15 mm, ‘ + ’: the diameter is 8-10 mm
L. casei T1 inhibited the overexpression of inflammatory cytokines by H. pylori infection
By far, many reports have shown that H. pylori infection induces high expression of inflammatory factors such as IL6 and TNF-α [35, 36]. By Fig. 1, it was clearly seen the IL6 and TNF-α mRNA expression levels of Group HP + T1 were markedly reduced compared to Group H. CCL2, also known as MCP-1, was found to possess mighty chemotactic ability to recruit monocytes and macrophages [37]. It also had a significant influence in many acute and chronic inflammatory diseases [38, 39]. Studies showed that CCL2 could activate monocytes to unleash inflammatory cytokines and provoke respiratory burst that can lead to generate excess reactive oxygen species (ROS) and harm cells [40]. As presented in Fig. 2, the overexpression of inflammatory proteins IL6, IL-8, TNF-α and CCL2 were inhibited by L. casei T1 and amoxicillin (5 mg/L) treatment. These findings clearly indicated that L. casei T1 could attenuate the inflammatory response induced by H. pylori in vitro.
Fig. 1.

Impact of L. casei T1 on inflammatory cytokines gene expression in vitro. The results are the means and SDs (N = 3) and P values were calculated by t tests. P values: * P < 0.05; ** P < 0.01; *** P < 0.001. C: blank control; T1: treated with L. casei T1 for 48 h; HP: treated with H. pylori for 48 h; HP + T1: treated with H. pylori for 24 h and L. casei T1 for another 24 h
Fig. 2.
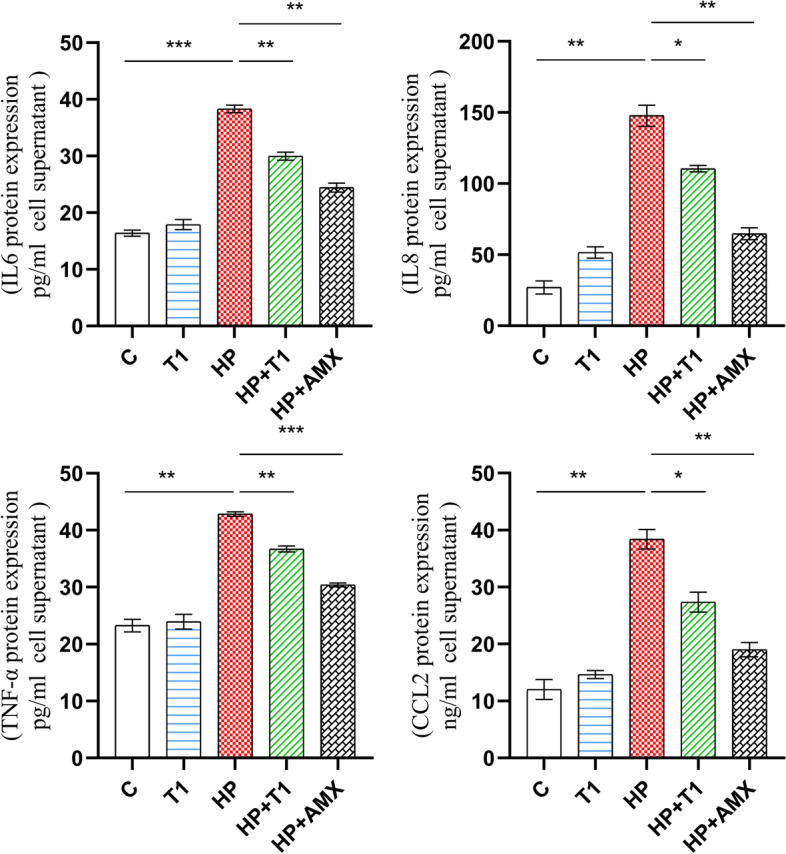
Impact of L. casei T1 on inflammatory protein in vitro. The results are the means and SDs (N = 3) and P values were calculated by t tests. P values: * P < 0.05; ** P < 0.01; *** P < 0.001. C: blank control; T1: treated with L. casei T1 for 48 h; HP: treated with H. pylori for 48 h; HP + T1: treated with H. pylori for 24 h and L. casei T1 for another 24 h; HP + AMX: treated with H. pylori for 24 h and amoxicillin (5 mg/L) for another 24 h
H. pylori infection model
Following the completion of the first pretreatment of mice, assessment of H. pylori colonization was identified by rapid urease test and PCR. As indicated in Fig. 3 (A), the dipsticks appeared red within a minute. After extracting the DNA of sampled gastric mucosae, PCR was performed with specific primers of H. pylori's vacA gene. After the reaction, the product was consistent with the expectation at 678 bp as indicated in Fig. 3 (B). Overall, we concluded that Balb/c mice of Group H were successfully infected with H. pylori.
Fig. 3.
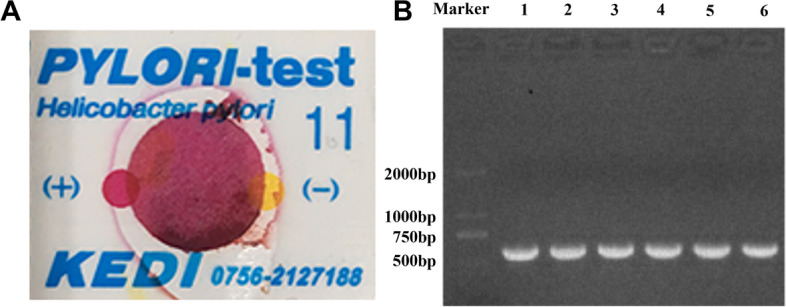
Detection of H. pylori infection model. A: Results of rapid urease test (turn red within a minute means positive, turn red within three minutes means light positive, yellow means negative); B: Electrophoresis of PCR-amplified products (vacA)
Rapid urease test
After the second treatment, all mice were euthanized and the gastric tissues were evaluated by the rapid urease test. It can be seen from Fig. 4 that gastric samples of Group C presented yellow color and Group H appeared red color. The color of Group H + T1 lies between the two, which means the severity of H. pylori infection of Group H + T1 is less serious than that of Group H.
Fig. 4.

Results of rapid urease test in mouse gastric tissues. Turn red within a minute means positive, turn red within three minutes means light positive, yellow means negative. A: Group C; B: Group H; C: Group H + T1
Routine blood tests
The results of the routine blood tests were given in Table 2. By comparing the experimental date, the numbers of white blood cells (WBC) and neutrophils (NEU) of Group H and Group H + T1 were substantially higher than Group C (*P < 0.05, # P < 0.05). However, there were no noticeable differences between Group H and Group H + T1. Overall, these results indicated that there were obvious inflammatory reactions in Group H and Group H + T1.
Table 2.
The number of white blood cell (WBC) and neutrophils (NEU)
| Numbers(109/L) | Group C | Group H | Group H + T1 |
|---|---|---|---|
| WBC | 4.75 ± 0.42 | 10.07 ± 0.88* | 9.73 ± 0.50* |
| NEU | 0.35 ± 0.05 | 0.98 + 0.08# | 0.95 ± 0.08# |
The results are the means and SDs (N = 3 ~ 6) and P values were calculated by one-way ANOVA
* P < 0.05, vs. Group C
# P < 0.05, vs. Group C
L. casei T1 alleviated H. pylori-induced gastric mucosal inflammation
Previous studies have indicated that mucin 5AC (MUC5AC) played a role in the adhesion of H pylori to the gastric mucosa [41, 42]. It was recently shown that MUC5AC was appreciably elevated in response to live H. pylori and had a positive association with H. pylori adhesion [43]. It can be seen from Fig. 5, Compared with Group H, the mRNA expression of TNF- α and MUC5AC in Group H + T1 decreased significantly. (P < 0.05). C-reactive protein (CRP) whose production is stimulated by IL6 is an acute reactive protein in humans and increases rapidly in the presence of inflammation [44–46]. As shown in Fig. 6 (A), the content of CRP in the serum of Groups C, H, and H + T1 was 235.40, 488.75 and 302.00 ng/mL, respectively. Compared to Group C, the CRP concentration of Group H and Group H + T1 was dramatically increased (P < 0.05). And the CRP concentration of Group H was obviously higher than Group H + T1 (P < 0.05). This could be illustrated as follows: L. casei T1 could effectively restrain H. pylori colonization in gastric mucosa and improve inflammation caused by H. pylori.
Fig. 5.

The TNF-α and MUC5AC mRNA expression in mice. The results are the means and SDs (N = 3 ~ 6) and P values were calculated by t tests. P values: * P < 0.05; ** P < 0.01; *** P < 0.001. C: Group C; H: Group H; H + T1: Group H + T1
Fig. 6.

Results of ELISA detection of mouse serum. The results are the means and SDs (N = 3 ~ 6) and P values were calculated using t tests. P values: * P < 0.05; ** P < 0.01; *** P < 0.001. C: Group C; H: Group H; H + T1: Group H + T1
Long-term inflammation process increases ROS production, causing oxidative stress the severity can be reflected by malondialdehyde (MDA) [47, 48]. As can be seen from Fig. 6 (B), the MDA of Group H + T1 was clearly lower than Group H (P < 0.05). Antioxidant enzymes are commonly used to scavenge ROS, such as glutathione peroxidase (GSH-PX) and superoxide dismutase (SOD) [49]. GSH-PX plays a crucial role in detoxifying H2O2, a major type of ROS [50]. SOD efficiently detoxifies superoxide in the cytoplasm and mitochondria and is a major cellular defense against ROS [51]. According to Fig. 6 (C and D), L. casei T1 could effectively improve the low expression of GSH-PX and SOD induced by H. pylori infection. Thus, the outcomes confirmed that L. casei T1 can alleviate oxidative stress.
H. pylori causes widespread gastric disease via directly injuring the gastric mucosa [30, 52]. HE staining results are shown in Fig. 7. The gastric mucosa of Group C was normal morphology and had no mucosal shedding and erosion (Fig. 7A). The result of Group H shows red blood cells can be seen in the gastric mucosa and there are cell necrosis and falling off the surface of the mucosa (Fig. 7B). Meanwhile, the gastric mucosa of Group H + T1 is visible with red blood cells and a slight amount of shedding cells (Fig. 7C). It follows that continuously feeding L. casei T1 alleviated mild gastric mucosal injury. NF-κB is seen to be a prototypical proinflammatory signaling pathway, greatly based on the activation of NF-kB by proinflammatory cytokines, for instance, TNF-a and IL-1 [53]. We could see from Fig. 8 that the NF-κB protein expression of Group H was 2.42 times of Group C, indicating H. pylori infection generated gastritis in mice. And, the NF-κB protein expression of Group H + T1 was as 0.67 times as Group H (P < 0.01), implying L. casei T1 had the capacity to directly resist H. pylori infection.
Fig. 7.

The results of gastric tissue HE staining. A: Group C; B: Group H; C: Group H + T1
Fig. 8.

The NF-κB protein expression. The results are the means and SDs (N = 3 ~ 6) and P values were calculated by t tests. P values: * P < 0.05; ** P < 0.01; *** P < 0.001.C: Group C; H: Group H; H + T1: Group H + T1
Effects of L. casei T1 on gut microbiota in mice
To identify microbial genera associated with L. casei T1 treatment, mouse fecal microbial composition was examined by 16S rRNA sequence. As can be seen from Fig. 9 (A), the 3D plot of PCoA provided a visual illustration of grouped samples. PC1 explained 37.78% of the variation, PC2 explained 21.53% and PC3 explained 15.13%. It demonstrated relatively significant differences in gut flora between groups. The differences of the intestinal microbial communities of groups at genus level was shown in Fig. 9 (D). Compared with Group C, the relative abundance of Bacteroidales_S24-7, Lactobacillus and Helicobacter of Group H increased, and the relative abundance of Lachnospiraceae_NK4A136, Lachnospiraceae, Desulfovibrio and Alistipes reduced in genus level. Compared to Group H, the relative abundance of Bacteroidales_S24-7, Lactobacillus and Alistipes of Group H + T1 reduced, and the relative abundance of Lachnospiraceae_NK4A136, Lachnospiraceae, Helicobacter and Odoribacter increased in genus level. The results of the significant differences analysis between groups were shown in Fig. 9 (B and C). Combined with relative species abundance, the main differential strains between the Group H and Group H + T1 were Bacteroides, Escherichia_Shigella, Enterobacter and Helicobacter. The main differential strains between Group C and Group H + T1 were Bifidobacterium and Helicobacter.
Fig. 9.

Effects of L. casei T1 treatment on gut microbiota of mice. A Principal coordinates analyses (PCoA) provide a visual illustration of grouped samples; B: Linear discriminant analysis (LDA) effect size (LEfSe) identified taxa most characteristic in H and HT1 groups; C: Linear discriminant analysis (LDA) effect size (LEfSe) identified taxa most characteristic in C and HT1 groups; D: Relative abundances of gut flora at genus level. C: Group C; H: Group H; HT1: Group H + T1
Bacteroides represent opportunistic pathogens in infectious diseases [54]. Escherichia_Shigella and Enterobacter are recognized in mammals as being both potentially pathogenic gut commensals [55]. After H. pylori infection in mice, the relative abundance of Bacteroides, Escherichia_Shigella and Enterobacter in the gut decreased. This showed that H. pylori could inhibit the reproduction of opportunistic pathogens. And treatment of H. pylori infection with L. casei T1 further suppressed the relative abundance of Bacteroides, Escherichia_Shigella and Enterobacter. Studies have identified Lachnospiraceae as potential promoters of gut health; for instance, it has been shown to have protective effects against Clostridioides difficile infection in preclinical studies [56]. Moreover, many Lachnospiraceae produce butyrate, which is a nutrient for the gut [57]. Odoribacter is generally considered to have a beneficial effect against inflammation [58]. In addition, decreased Odoribacter levels were found in insulin-resistant people [59]. In H. pylori infected mice, the relative abundance of Lachnospiraceae and Odoribacter decreased. However, after treating with L. casei T1, the relative abundance of Lachnospiraceae and Odoribacter not only increased, but also was higher than Group C. This indicated that treatment with L. casei T1 could improve the intestinal environment by promoting the growth of beneficial bacteria. Although the relative abundance of Bifidobacterium in Group H + T1 (0.091%) was significantly lower compared to Group C (2.99%), it was higher than that in Group H (0.030%). The elevated relative abundance of Helicobacter in Group H + T1 might be due to L. casei T1 inhibiting the colonization of H. pylori in the stomach, resulting in a large amount of H. pylori entering the intestine.
Discussion
It is widely recognized that diet affects the incidence of many diseases [60]. For instance, bearberry and cranberry juice were reported to treat urinary tract infections [61]. Foods that have sufficient effects to improve health or reduce disease are called functional foods. Functional foods, including probiotics, have become an established dietary trend. Probiotic foods not only preserve probiotic viability during product manufacture and shelf life, but also exert their beneficial effect in consumers’ gastrointestinal tract [62]. The bacteriostasis of probiotics is by producing acidic substances, inhibiting bacterial adherence, or producing antibacterial substances [63]. Enhancement of mucosal barrier could be another important pathway that probiotics confer benefits to the host [64]. Studies have shown that probiotics have a variety of health benefits, such as anti-obesity, anti-inflammatory, and immune-boosting [65]. It has been shown the antimicrobial effect of Lactobacillus species on H. pylori infection in vivo, achieved by the unleash of bacteriocins and the ability to decrease adherence of H. pylori to epithelial cells [66]. Our previous studies proved L. casei T1 possessed excellent acid tolerance and a high survival rate in the stomach [26, 27]. L. casei T1 also could suppress adhesion of H. pylori strains in vitro [28]. As a result, L. casei T1 helps to competitively inhibit the colonization and absorption of nutrients of H. pylori. Probiotics also can inhibit or kill H. pylori by producing substances in the stomach [67]. Earlier studies showed that bacteriocins secreted by L. casei T1 had broad-spectrum antimicrobial activities. In this study, we demonstrated L. casei T1’s fermentation broth containing bacteriocin and organic acid can effectively inhibit H. pylori proliferation and had a better bacteriostatic effect than bacteriocin only (Table 1) [26]. It is evident from Figs. 1 and 2 that L. casei T1 significantly attenuated the expression of inflammatory factors induced by H. pylori in vitro. This confirmed that L. casei T1 can attenuate or even suppress harmful inflammatory responses during H. pylori infection in vitro. We also constructed H. pylori-infected murine models, which were verified to be successful by rapid urease experiment and agarose gel electrophoresis. The capacity to improve infection symptoms of H. pylori was finally confirmed in vivo.
Recent findings have suggested that probiotics performed systemic anti-inflammatory effects [68, 69]. Our results of blood routine are consistent with it (Table 2). Group H and Group H + T1 had a much higher level of leukocytes and neutrophils than Group C, indicating successful modeling of H. pylori infection. H. pylori can be colonized in the stomach through the production of urease, which protects it from the acidic environment by converting urea into bicarbonate and ammonia [70, 71]. So, the deeper the color of rapid urease test, the more serious H. pylori infection. The test paper color of Group H was significantly redder than Group H + T1 (Fig. 4), showing L. casei T1 treatment could substantially improve the prognosis for H. pylori infection and inhibit colonization rate in vivo. Compared to Group H, the mRNA expression of MUC5AC of Group H + T1 was markedly reduced (Fig. 5), also illustrating L. casei T1 treatment could help the body struggle with H. pylori colonization. Excessive inflammation causes tissue damage and chronic disease, including autoinflammatory and cardiovascular diseases [72–74]. Studies have revealed that TNF-α was over-expressed in patients with H. pylori infection [75]. The TNF-α mRNA expression of Group H + T1 was markedly lower than Group H (Fig. 5, P < 0.01). Some comparative studies showed that H. pylori infection may promote the expression of CRP [76, 77]. The results showed that L. casei T1 could alleviate the high CRP level by H. pylori infection (Fig. 6A). HE staining was used for observing mucosal inflammation and tissue damage [78]. After 4 weeks of L. casei T1 treatment, HE staining (Fig. 7) showed Group H + T1 exhibited lighter inflammation than Group H. The immunohistochemistry results (Fig. 8) also indicated that L. casei T1 treatment inhibited the overexpression of NF-κB protein triggered by H. pylori infection. The results of 16S sequencing showed that gut microbiome diversity and composition after treating H. pylori with L. casei T1 were closer to the control group. These results showed that L. casei T1 treatment was effective in mitigating the inflammatory response caused by H. pylori. As known, oxidative stress and inflammation are mutually dependent and connected [79, 80]. The change of ROS also can reflect inflammatory conditions of the body [81]. Oxidative stress is a harmful process that can negatively affect several cellular structures, such as membranes, lipids, proteins, lipoproteins and DNA [82, 83]. Studies show that H. pylori infection increases oxidative stress in vivo [84–86]. Reducing oxidative stress is beneficial for preventing the development of H. pylori-associated gastric diseases [87]. We have tested the indicators of oxidative stress. Despite the level of MDA shows that both Group H and H + T1 produce oxidative stress, but the MDA concentration of Group H + T1 was significantly lower than Group H (p < 0.001). At the same time, GSH-PX and SOD which play as an antioxidant of Group H + T1 was more than Group H. These findings clearly indicated that L. casei T1 was able to attenuate H. pylori-induced ROS production in vivo.
Conclusions
In this paper, we revealed that L. casei T1 could alleviate H. pylori-induced inflammation and oxidative stress in vivo and in vitro. Although taking L. casei T1 cannot completely eradicate H. pylori, it can effectively reduce the damage caused by H. pylori infection. Therefore, further research could explore thoroughly eradicate H. pylori by L. casei T1 in conjunction with antibiotic and reveal the differences in prevention and treatment of H. pylori-induced gastric mucosal inflammation.
Acknowledgements
Not applicable.
Authors’ contributions
Jingshan Peng, Daoyan Wu, Andong Zhang and Jian Zhao conceived and conducted the experiment, Zhihao Yu and Mei Cao analyzed the results and wrote the manuscript. All authors reviewed the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (82160390), Sichuan Science and Technology Program (2023YFS0156), Open Project Program of Irradiation Preservation Technology Key Laboratory of Sichuan Province, (FZBC2021001), Training Plan 2.0 for Top Students in Basic Sciences (20212079), and Higher Education Personnel Training and Teaching Reform Project of Sichuan Province (JG2021-42).
Availability of data and materials
The dataset supporting the conclusions of this article are available in the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) repository, BioProject ID PRJNA857345 (http://www.ncbi.nlm.nih.gov/bioproject/857345). The data will be released to the public when the manuscript is formally accepted for publication.
Declarations
Ethics approval and consent to participate
Animal care and all experimental procedures were conducted in accordance with the Animal Care Committee guidelines and approved by the Ethics Committee of Sichuan University (ethics
approval number: 20201109002). The report of animal experiments is in accordance with the ARRIVE guidelines.
Consent for publication
Not applicable.
Competing interests
The authors confirm that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Zhihao Yu and Mei Cao contributed equally to this work.
References
- 1.Dunn BE, Cohen H, Blaser MJ. Helicobacter pylori. Clin Microbiol Rev. 1997;10(4):720–741. doi: 10.1128/cmr.10.4.720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Jonkers D, Stockbrugger R. Review article: Probiotics in gastrointestinal and liver diseases. Aliment Pharmacol Ther. 2007;26(Suppl 2):133–148. doi: 10.1111/j.1365-2036.2007.03480.x. [DOI] [PubMed] [Google Scholar]
- 3.Brawner KM, Morrow CD, Smith PD. Gastric microbiome and gastric cancer. Cancer J. 2014;20(3):211–216. doi: 10.1097/PPO.0000000000000043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lamb A, Chen LF. Role of the Helicobacter pylori-induced inflammatory response in the development of gastric cancer. J Cell Biochem. 2013;114(3):491–497. doi: 10.1002/jcb.24389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hsu PI, Lai KH, Hsu PN, Lo GH, Yu HC, Chen WC, et al. Helicobacter pylori infection and the risk of gastric malignancy. Am J Gastroenterol. 2007;102(4):725–730. doi: 10.1111/j.1572-0241.2006.01109.x. [DOI] [PubMed] [Google Scholar]
- 6.Hu Y, Zhang M, Lu B, Dai J. Helicobacter pylori and antibiotic resistance, a continuing and intractable problem. Helicobacter. 2016;21(5):349–363. doi: 10.1111/hel.12299. [DOI] [PubMed] [Google Scholar]
- 7.Herrera V, Parsonnet J. Helicobacter pylori and gastric adenocarcinoma. Clin Microbiol Infect. 2009;15(11):971–976. doi: 10.1111/j.1469-0691.2009.03031.x. [DOI] [PubMed] [Google Scholar]
- 8.Dore MP, Lu H, Graham DY. Role of bismuth in improving Helicobacter pylori eradication with triple therapy. Gut. 2016;65(5):870–878. doi: 10.1136/gutjnl-2015-311019. [DOI] [PubMed] [Google Scholar]
- 9.Garza-Gonzalez E, Perez-Perez GI, Maldonado-Garza HJ, Bosques-Padilla FJ. A review of Helicobacter pylori diagnosis, treatment, and methods to detect eradication. World J Gastroenterol. 2014;20(6):1438–1449. doi: 10.3748/wjg.v20.i6.1438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gisbert JP. The recurrence of Helicobacter pylori infection: incidence and variables influencing it. A critical review Am J Gastroenterol. 2005;100(9):2083–2099. doi: 10.1111/j.1572-0241.2005.50043.x. [DOI] [PubMed] [Google Scholar]
- 11.Scribano ML, Prantera C. Antibiotics and inflammatory bowel diseases. Dig dis. 2013;31(3–4):379–384. doi: 10.1159/000354704. [DOI] [PubMed] [Google Scholar]
- 12.Hou Q, Zhao F, Liu W, Lv R, Khine WWT, Han J, et al. Probiotic-directed modulation of gut microbiota is basal microbiome dependent. Gut microbes. 2020;12(1):1736974. doi: 10.1080/19490976.2020.1736974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mundula T, Ricci F, Barbetta B, Baccini M, Amedei A. Effect of probiotics on oral candidiasis: a systematic review and meta-analysis. Nutrients. 2019;11(10):2449. doi: 10.3390/nu11102449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Principi N, Cozzali R, Farinelli E, Brusaferro A, Esposito S. Gut dysbiosis and irritable bowel syndrome: The potential role of probiotics. J Infect. 2018;76(2):111–120. doi: 10.1016/j.jinf.2017.12.013. [DOI] [PubMed] [Google Scholar]
- 15.Lin TH, Lin CH, Pan TM. The implication of probiotics in the prevention of dental caries. Appl Microbiol Biotechnol. 2018;102(2):577–586. doi: 10.1007/s00253-017-8664-z. [DOI] [PubMed] [Google Scholar]
- 16.Mantegazza C, Molinari P, D'Auria E, Sonnino M, Morelli L, Zuccotti GV. Probiotics and antibiotic-associated diarrhea in children: A review and new evidence on Lactobacillus rhamnosus GG during and after antibiotic treatment. Pharmacol Res. 2018;128:63–72. doi: 10.1016/j.phrs.2017.08.001. [DOI] [PubMed] [Google Scholar]
- 17.Zhou Q, Xue B, Gu R, Li P, Gu Q. Lactobacillus plantarum ZJ316 attenuates Helicobacter pylori-induced gastritis in C57BL/6 mice. J Agric Food Chem. 2021;69(23):6510–6523. doi: 10.1021/acs.jafc.1c01070. [DOI] [PubMed] [Google Scholar]
- 18.Ba Lciunas EM, Martinez FC, Todorov SD, Franco B, Converti A, Oliveira R. Novel biotechnological applications of bacteriocins: A review. Food Control. 2013;32(1):134–142. [Google Scholar]
- 19.Devlieghere F, Vermeiren L, Debevere J. New preservation technologies: Possibilities and limitations. Int Dairy J. 2004;14(4):273–285. [Google Scholar]
- 20.Jelena L, Branko J, Milan K, et al. Molecular characterization of a novel bacteriocin and an unusually large aggregation factor of Lactobacillus paracasei subsp. paracasei BGSJ2-8, a natural isolate from homemade cheese. Curr Microbiol. 2007;55(3):266–271. doi: 10.1007/s00284-007-0159-1. [DOI] [PubMed] [Google Scholar]
- 21.Wang KY, Li SN, Liu CS, Perng DS, Su YC, Wu DC, et al. Effects of ingesting Lactobacillus- and Bifidobacterium-containing yogurt in subjects with colonized Helicobacter pylori. Am J Clin Nutr. 2004;80(3):737–741. doi: 10.1093/ajcn/80.3.737. [DOI] [PubMed] [Google Scholar]
- 22.Lionetti E, Miniello VL, Castellaneta SP, Magista AM, de Canio A, Maurogiovanni G, et al. Lactobacillus reuteri therapy to reduce side-effects during anti-Helicobacter pylori treatment in children: a randomized placebo controlled trial. Aliment Pharmacol Ther. 2006;24(10):1461–1468. doi: 10.1111/j.1365-2036.2006.03145.x. [DOI] [PubMed] [Google Scholar]
- 23.Ruggiero P. Use of probiotics in the fight against Helicobacter pylori. World J Gastrointest Pathophysiol. 2014;5(4):384–391. doi: 10.4291/wjgp.v5.i4.384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Emara MH, Mohamed SY, Abdel-Aziz HR. Lactobacillus reuteri in management of Helicobacter pylori infection in dyspeptic patients: a double-blind placebo-controlled randomized clinical trial. Therap Adv Gastroenterol. 2014;7(1):4–13. doi: 10.1177/1756283X13503514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lin WH, Wu CR, Fang TJ, Guo JT, Huang SY, Lee MS, et al. Anti-Helicobacter pylori activity of fermented milk with lactic acid bacteria. J Sci Food Agric. 2011;91(8):1424–1431. doi: 10.1002/jsfa.4327. [DOI] [PubMed] [Google Scholar]
- 26.Luo F, Feng S, Sun Q, Xiang W, Zhao J, Zhang J, et al. Screening for bacteriocin-producing lactic acid bacteria from kurut, a traditional naturally-fermented yak milk from Qinghai-Tibet plateau. Food Control. 2011;22(1):50–53. [Google Scholar]
- 27.Wang M, Chen C, Song L, Panpan QI, Zhenzhen YI, Feng S, et al. Biological properties and preservation of Lactobacillus casei T1. Journal of Anhui Agricultural University. 2016;43(1):73–76. [Google Scholar]
- 28.Wu DY, Cao M, Zhou J, Yan SY, Peng JS, Yu ZH, et al. Lactobacillus casei T1 from kurut against Helicobacter pylori-induced inflammation and the gut microbial disorder. J Funct Foods. 2021;85:104611.
- 29.Corsetti A, Settanni L, Van Sinderen D. Characterization of bacteriocin-like inhibitory substances (BLIS) from sourdough lactic acid bacteria and evaluation of their in vitro and in situ activity. J Appl Microbiol. 2004;96(3):521–534. doi: 10.1111/j.1365-2672.2004.02171.x. [DOI] [PubMed] [Google Scholar]
- 30.Wei MY, Shi S, Liang C, Meng QC, Hua J, Zhang YY, et al. The microbiota and microbiome in pancreatic cancer: more influential than expected. Mol Cancer. 2019;18(1):97. doi: 10.1186/s12943-019-1008-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Raspor P, Jevsnik M. Good nutritional practice from producer to consumer. Crit Rev Food Sci Nutr. 2008;48(3):276–292. doi: 10.1080/10408390701326219. [DOI] [PubMed] [Google Scholar]
- 32.Mu WM, Yu SH, Zhu LJ, Zhang T, Jiang B. Recent research on 3-phenyllactic acid, a broad-spectrum antimicrobial compound. Appl Microbiol Biotechnol. 2012;95(5):1155–1163. doi: 10.1007/s00253-012-4269-8. [DOI] [PubMed] [Google Scholar]
- 33.Bo Y, Hui J, Wei B, Yang T, Liu H, Ju X, et al. A new chitosan sub-micron and encapsulated Iturin A with enhanced antifungal activity against Ceratocystis fimbriata and Rhizopus strolonifer. Int J Biol Macromol. 2020;159:995–1003. doi: 10.1016/j.ijbiomac.2020.05.116. [DOI] [PubMed] [Google Scholar]
- 34.Szczepańska E, Colombo D, Tentori F, Olejniczak T, Brenna E, Monti D, et al. Ene-reductase transformation of massoia lactone to δ-decalactone in a continuous-flow reactor. Sci Rep. 2021;11(1):18794. doi: 10.1038/s41598-021-97585-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wustner S, Anderl F, Wanisch A, Sachs C, Steiger K, Nerlich A, et al. Helicobacter pylori gamma-glutamyl transferase contributes to colonization and differential recruitment of T cells during persistence. Sci Rep. 2017;7(1):13636. doi: 10.1038/s41598-017-14028-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mwangi C, Njoroge S, Tshibangu-Kabamba E, Moloo Z, Rajula A, Devani S, et al. Whole genome sequencing reveals virulence potentials of Helicobacter pylori strain KE21 isolated from a Kenyan patient with gastric signet ring cell carcinoma. Toxins. 2020;12(9):556. doi: 10.3390/toxins12090556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Jin J, Lin J, Xu A, Lou J, Qian C, Li X, et al. CCL2: an important mediator between tumor cells and host cells in tumor microenvironment. Front Oncol. 2021;11:722916. doi: 10.3389/fonc.2021.722916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Moadab F, Khorramdelazad H, Abbasifard M. Role of CCL2/CCR2 axis in the immunopathogenesis of rheumatoid arthritis: Latest evidence and therapeutic approaches. Life Sci. 2021;269:119034. doi: 10.1016/j.lfs.2021.119034. [DOI] [PubMed] [Google Scholar]
- 39.Ragin AB, Wu Y, Storey P, Cohen BA, Edelman RR, Epstein LG. Monocyte chemoattractant protein-1 correlates with subcortical brain injury in HIV infection. Neurology. 2006;66(8):1255–1257. doi: 10.1212/01.wnl.0000208433.34723.65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rollins BJ, Walz A, Baggiolini M. Recombinant human MCP-1/JE induces chemotaxis, calcium flux, and the respiratory burst in human monocytes. Blood. 1991;78(4):1112–1116. [PubMed] [Google Scholar]
- 41.Van den Brink GR, Tytgat KM, Van der Hulst RW, Van der Loos CM, Einerhand AW, Buller HA, et al. H pylori colocalises with MUC5AC in the human stomach. Gut. 2000;46(5):601–607. doi: 10.1136/gut.46.5.601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Van de Bovenkamp JH, Mahdavi J, Korteland-Van Male AM, Buller HA, Einerhand AW, Boren T, et al. The MUC5AC glycoprotein is the primary receptor for Helicobacter pylori in the human stomach. Helicobacter. 2003;8(5):521–532. doi: 10.1046/j.1523-5378.2003.00173.x. [DOI] [PubMed] [Google Scholar]
- 43.Gonciarz W, Walencka M, Moran AP, Hinc K, Obuchowski M, Chmiela M. Upregulation of MUC5AC production and deposition of LEWIS determinants by Helicobacter pylori facilitate gastric tissue colonization and the maintenance of infection. J Biomed Sci. 2019;26(1):23. doi: 10.1186/s12929-019-0515-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Black S, Kushner I, Samols D. C-reactive Protein. J Biol Chem. 2004;279(47):48487–48490. doi: 10.1074/jbc.R400025200. [DOI] [PubMed] [Google Scholar]
- 45.Del Giudice M, Gangestad SW. Rethinking IL6 and CRP: Why they are more than inflammatory biomarkers, and why it matters. Brain Behav Immun. 2018;70:61–75. doi: 10.1016/j.bbi.2018.02.013. [DOI] [PubMed] [Google Scholar]
- 46.Sproston NR, Ashworth JJ. Role of C-reactive protein at sites of inflammation and infection. Front Immunol. 2018;9:754. doi: 10.3389/fimmu.2018.00754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zhang C, Qiu Z, Zhang L, Pang Q, Yang Z, Qin JK, et al. Design and synthesis of a ratiometric photoacoustic imaging probe activated by selenol for visual monitoring of pathological progression of autoimmune hepatitis. Chem Sci. 2021;12(13):4883–4888. doi: 10.1039/d0sc06573k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Agita A, Alsagaff MT. Inflammation, immunity, and hypertension. Acta Med Indones. 2017;49(2):158–165. [PubMed] [Google Scholar]
- 49.Shen J, Yang D, Zhou X, Wang Y, Tang S, Yin H, et al. Role of autophagy in zinc oxide nanoparticles-induced apoptosis of mouse leydig cells. Int J Mol Sci. 2019;20(16):4042. doi: 10.3390/ijms20164042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Mo JL, Pan ZG, Chen X, Lei Y, Lv LL, Qian C, et al. MicroRNA-365 knockdown prevents ischemic neuronal injury by activating oxidation resistance 1-mediated antioxidant signals. Neurosci Bull. 2019;35(5):815–825. doi: 10.1007/s12264-019-00371-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wang X, Zhang H, Sapio R, Yang J, Wong J, Zhang X, et al. SOD1 regulates ribosome biogenesis in KRAS mutant non-small cell lung cancer. Nat Commun. 2021;12(1):2259. doi: 10.1038/s41467-021-22480-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Deen NS, Huang SJ, Gong L, Kwok T, Devenish RJ. The impact of autophagic processes on the intracellular fate of Helicobacter pylori: more tricks from an enigmatic pathogen? Autophagy. 2013;9(5):639–652. doi: 10.4161/auto.23782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lawrence T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb Perspect Biol. 2009;1(6):a001651. doi: 10.1101/cshperspect.a001651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Souai N, Zidi O, Mosbah A, Kosai I, Manaa JE, Mokhtar NB, et al. Impact of the post-transplant period and lifestyle diseases on human gut microbiota in kidney graft recipients. Microorganisms. 2020;8(11):1724. doi: 10.3390/microorganisms8111724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Funosas G, Triadó-Margarit X, Castro F, Villafuerte R, Delibes-Mateos M, Rouco C, et al. Individual fate and gut microbiome composition in the European wild rabbit (Oryctolagus cuniculus) Sci Rep. 2021;11(1):766. doi: 10.1038/s41598-020-80782-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Taur Y, Jenq RR, Perales MA, Littmann ER, Morjaria S, Ling L, et al. The effects of intestinal tract bacterial diversity on mortality following allogeneic hematopoietic stem cell transplantation. Blood. 2014;124(7):1174–1182. doi: 10.1182/blood-2014-02-554725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Freetly HC, Dickey A, Lindholm-Perry AK, Thallman RM, Keele JW, Foote AP, et al. Digestive tract microbiota of beef cattle that differed in feed efficiency. J Anim Sci. 2020;98(2):skaa008. doi: 10.1093/jas/skaa008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Silvestri C, Pagano E, Lacroix S, Venneri T, Cristiano C, Calignano A, et al. Fish oil, cannabidiol and the gut microbiota: an investigation in a murine model of colitis. Front Pharmacol. 2020;11:585096. doi: 10.3389/fphar.2020.585096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Fu Y, Yin R, Liu Z, Niu Y, Guo E, Cheng R, et al. Hypoglycemic effect of prolamin from cooked foxtail millet (setaria italic) on streptozotocin-induced diabetic mice. Nutrients. 2020;12(11):3452. doi: 10.3390/nu12113452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Arvanitoyannis IS, Van Houwelingen-Koukaliaroglou M. Functional foods: a survey of health claims, pros and cons, and current legislation. Crit Rev Food Sci Nutr. 2005;45(5):385–404. doi: 10.1080/10408390590967667. [DOI] [PubMed] [Google Scholar]
- 61.Viuda-Martos M, Ruiz-Navajas Y, Fernández-López J, Pérez-Alvarez JA. Spices as functional foods. Crit Rev Food Sci Nutr. 2011;51(1):13–28. doi: 10.1080/10408390903044271. [DOI] [PubMed] [Google Scholar]
- 62.Sumeri I, Arike L, Adamberg K, Paalme T. Single bioreactor gastrointestinal tract simulator for study of survival of probiotic bacteria. Appl Microbiol Biotechnol. 2008;80(2):317–324. doi: 10.1007/s00253-008-1553-8. [DOI] [PubMed] [Google Scholar]
- 63.Ng SC, Hart AL, Kamm MA, Stagg AJ, Knight SC. Mechanisms of action of probiotics: recent advances. Inflamm Bowel Dis. 2009;15(2):300–310. doi: 10.1002/ibd.20602. [DOI] [PubMed] [Google Scholar]
- 64.Meddings J. The significance of the gut barrier in disease. Gut. 2008;57(4):438. doi: 10.1136/gut.2007.143172. [DOI] [PubMed] [Google Scholar]
- 65.Lee CS, Kim SH. Anti-inflammatory and anti-osteoporotic potential of Lactobacillus plantarum A41 and L. fermentum SRK414 as probiotics. Probiotics Antimicrob Proteins. 2020;12(2):623–634. doi: 10.1007/s12602-019-09577-y. [DOI] [PubMed] [Google Scholar]
- 66.Gotteland M, Brunser O, Cruchet S. Systematic review: are probiotics useful in controlling gastric colonization by Helicobacter pylori? Aliment Pharmacol Ther. 2006;23(8):1077–1086. doi: 10.1111/j.1365-2036.2006.02868.x. [DOI] [PubMed] [Google Scholar]
- 67.Tannock GW. A special fondness for lactobacilli. Appl Environ Microbiol. 2004;70(6):3189–3194. doi: 10.1128/AEM.70.6.3189-3194.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Kaila M, Isolauri E, Soppi E, Virtanen E, Laine S, Arvilommi H. Enhancement of the circulating antibody secreting cell response in human diarrhea by a human Lactobacillus strain. Pediatr Res. 1992;32(2):141–144. doi: 10.1203/00006450-199208000-00002. [DOI] [PubMed] [Google Scholar]
- 69.Sheil B, McCarthy J, O'Mahony L, Bennett MW, Ryan P, Fitzgibbon JJ, et al. Is the mucosal route of administration essential for probiotic function? Subcutaneous administration is associated with attenuation of murine colitis and arthritis. Gut. 2004;53(5):694–700. doi: 10.1136/gut.2003.027789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Choi DS, Seo SI, Shin WG, Park CH. Risk for Colorectal Neoplasia in Patients With Helicobacter pylori Infection: A Systematic Review and Meta-analysis. Clin Transl Gastroenterol. 2020;11(2):e00127. doi: 10.14309/ctg.0000000000000127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Takahashi-Kanemitsu A, Knight CT, Hatakeyama M. Molecular anatomy and pathogenic actions of Helicobacter pylori CagA that underpin gastric carcinogenesis. Cell Mol Immunol. 2020;17(1):50–63. doi: 10.1038/s41423-019-0339-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Eberhardson M, Tarnawski L, Centa M, Olofsson PS. Neural control of inflammation: bioelectronic medicine in treatment of chronic inflammatory disease. Cold Spring Harb Perspect Med. 2020;10(3):a034181. doi: 10.1101/cshperspect.a034181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Raimondo TM, Mooney DJ. Anti-inflammatory nanoparticles significantly improve muscle function in a murine model of advanced muscular dystrophy. Sci Adv. 2021;7(26):eabh3693. doi: 10.1126/sciadv.abh3693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kay J, Thadhani E, Samson L, Engelward B. Inflammation-induced DNA damage, mutations and cancer. DNA Repair. 2019;83:102673. doi: 10.1016/j.dnarep.2019.102673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Lu W, Pan K, Zhang L, Lin D, Miao X, You W. Genetic polymorphisms of interleukin (IL)-1B, IL-1RN, IL-8, IL-10 and tumor necrosis factor alpha and risk of gastric cancer in a Chinese population. Carcinogenesis. 2005;26(3):631–636. doi: 10.1093/carcin/bgh349. [DOI] [PubMed] [Google Scholar]
- 76.Ishida Y, Suzuki K, Taki K, Niwa T, Kurotsuchi S, Ando H, et al. Significant association between Helicobacter pylori infection and serum C-reactive protein. Int J Med Sci. 2008;5(4):224–229. doi: 10.7150/ijms.5.224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Hou B, Zhang M, Liu M, Dai W, Lin Y, Li Y, et al. Association of active Helicobacter pylori infection and anemia in elderly males. BMC Infect Dis. 2019;19(1):228. doi: 10.1186/s12879-019-3849-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Ma QL, Zhao LZ, Liu RR, Jin BQ, Song W, Wang Y, et al. Improved implant osseointegration of a nanostructured titanium surface via mediation of macrophage polarization. Biomaterials. 2014;35(37):9853–9867. doi: 10.1016/j.biomaterials.2014.08.025. [DOI] [PubMed] [Google Scholar]
- 79.Trostel J, Truong LD, Roncal-Jimenez C, Miyazaki M, Miyazaki-Anzai S, Kuwabara M, et al. Different effects of global osteopontin and macrophage osteopontin in glomerular injury. Am J Physiol Renal Physiol. 2018;315(4):F759–F768. doi: 10.1152/ajprenal.00458.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Hsu YH, Chiu IJ, Lin YF, Chen YJ, Lee YH, Chiu HW. Lactoferrin contributes a renoprotective effect in acute kidney injury and early renal fibrosis. Pharmaceutics. 2020;12(5):434. doi: 10.3390/pharmaceutics12050434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Fernandes EFA, Ozcelik D. Imaging biomarkers for monitoring the inflammatory redox landscape in the brain. Antioxidants (Basel, Switzerland) 2021;10(4):528. doi: 10.3390/antiox10040528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Pizzino G, Irrera N, Cucinotta M, Pallio G, Mannino F, Arcoraci V, et al. Oxidative stress: harms and benefits for human health. Oxid Med Cell Longev. 2017;2017:8416763. doi: 10.1155/2017/8416763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Milne GL, Yin H, Hardy KD, Davies SS, Roberts LJ., 2nd Isoprostane generation and function. Chem Rev. 2011;111(10):5973–5996. doi: 10.1021/cr200160h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Peter S, Beglinger C. Helicobacter pylori and gastric cancer: the causal relationship. Digestion. 2007;75(1):25–35. doi: 10.1159/000101564. [DOI] [PubMed] [Google Scholar]
- 85.Sezikli M, Cetinkaya ZA, Sezikli H, Guzelbulut F, Tiftikci A, Ince AT, et al. Oxidative stress in Helicobacter pylori infection: does supplementation with vitamins C and E increase the eradication rate? Helicobacter. 2009;14(4):280–285. doi: 10.1111/j.1523-5378.2009.00686.x. [DOI] [PubMed] [Google Scholar]
- 86.Arslan E, Atilgan H, Yavasoglu I. The prevalence of Helicobacter pylori in obese subjects. Eur J Intern Med. 2009;20(7):695–697. doi: 10.1016/j.ejim.2009.07.013. [DOI] [PubMed] [Google Scholar]
- 87.Kyung S, Lim JW, Kim H. Alpha-lipoic acid inhibits IL-8 expression by activating Nrf2 signaling in Helicobacter pylori-infected gastric epithelial cells. Nutrients. 2019;11(10):2524. doi: 10.3390/nu11102524. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The dataset supporting the conclusions of this article are available in the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) repository, BioProject ID PRJNA857345 (http://www.ncbi.nlm.nih.gov/bioproject/857345). The data will be released to the public when the manuscript is formally accepted for publication.