

Abstract
Cardiovascular disease (CVD) is the leading cause of death worldwide, in both developed and developing countries. According to the WHO report, the morbidity and mortality caused by CVD will continue to rise with the estimation of death going up to 22.2 million in 2030. NADPH oxidase (NOX)-derived reactive oxygen species (ROS) production induces endothelial nitric oxide synthase (eNOS) uncoupling and mitochondrial dysfunction, resulting in sustained oxidative stress and the development of cardiovascular diseases. Seven distinct members of the family have been identified of which four (namely, NOX1, 2, 4 and 5) may have cardiovascular functions. Currently, the treatment and management plan for patients with CVDs mainly depends on the drugs. However, prolonged use of prescribed drugs may cause adverse drug reactions. Therefore, it is crucial to find alternative treatment options with lesser adverse effects. Natural products have been gaining interest as complementary therapy for CVDs over the past decade due to their wide range of medicinal properties, including antioxidants. These might be due to their potent active ingredients, such as flavonoid and phenolic compounds. Numerous natural compounds have been demonstrated to have advantageous effects on cardiovascular disease via NADPH cascade. This review highlights the potential of natural products targeting NOX-derived ROS generation in treating CVDs. Emphasis is put on the activation of the oxidases, including upstream or downstream signalling events.
Keywords: natural products, cardiovascular diseases, oxidative stress, NADPH signalling pathway
1. Introduction
Cardiovascular disease (CVD) remains the major cause of mortality and premature death worldwide [1]. In 2019, according to the World Health Organization, 17.9 million deaths were reported from cardiovascular disease, accounting for 32% of all global fatalities, and 85% of these deaths are the result of a heart attack or a stroke [2]. It has been reported that 58% of the 17.9 million CVD fatalities that occurred globally were in Asia [3]. Risk factors for cardiovascular disease include sedentary lifestyle, unhealthy diet, smoking, diabetes, age, and genetics [4]. These risk factors unite behind a convergence of mechanisms involving reactive oxygen species (ROS), as reported in vitro, in vivo, and clinical studies [4]. Oxidative stress is a result of a change in redox equilibrium that leads to an imbalance between ROS formation and endogenous antioxidant systems [5]. Oxidative stress elicits deleterious effects by inducing damage to macromolecules, such as deoxyribonucleic acid (DNA), ribonucleic acid (RNA), protein, and lipids. The imbalance in ROS metabolism has been reported in the pathogenesis of several CVDs, including heart failure, stroke, hypertension, atherosclerosis, etc. [6]. In fact, oxidative stress is no longer focused on the imbalance between ROS production and scavenging, but on the relevant enzyme dysfunctions [7]. In biological systems, several major factors that contribute to the production of ROS are an increase in mechanoreceptor activation [8], mitochondrial electron transport chain, lipoxygenases, cytochrome P450 oxidases, nitric oxide (NO) synthase, xanthine oxidase (XO), and nicotinamide adenine dinucleotide phosphate (NADPH) oxidase [6]. Among these, NADPH oxidase (NOX) is the primary source of ROS in the cardiovascular system, which affects a variety of signalling pathways [9,10,11] and certain transcription factors [12] to modify cardiovascular cell function.
Statins and metformin are examples of prescribed drugs that function as antioxidants, scavenging free radicals and ROS superoxide (O2−) in CVD [13]. In order to overcome undesirable side effects or any harmful effects of prescribed cardiovascular drugs with prolonged usage [14], research is shifting gears towards natural products due to their greater safety and potent various medicinal properties, including antioxidant [15]. Natural products, such as herbs and spices, are the major source of development of active drugs and are used in traditional medicine [15]. Furthermore, the preparation of these natural products is easier and low cost [16]. This is due to the availability of these natural products, which can be sourced locally and are easily available throughout the year [16]. In addition, due to the low toxicity beneficial properties of natural products, they have long been used as therapeutic agents to combat cardiovascular disease [17]. Thus, this review is aimed at highlighting natural products which are being used against cardiovascular disease by inhibiting NADPH signalling pathways. We hypothesise that natural products are effective in inhibiting specific subunits of NADPH in cardiovascular conditions, such as cardiac damage, arteriosclerosis, hypertension, aortic valve disease, myocardial ischemic-reperfusion injury, hypercholesterolemia, and cardiac hypertrophy.
2. Target Pathway: NADPH Oxidases
2.1. The NOX Family of NADPH Oxidases
NADPH oxidases (NOXs) are a family of multi subunit enzymes that produce superoxide anion radical (O2−) by oxygen reduction using NADPH or nicotinamide adenine dinucleotide (NADH). They are found in phagocytic cells (neutrophils, monocytes, and macrophages) and non-phagocytic cells, such as vascular smooth muscle cells (VSMC) and endothelial cells [18]. NOXs are also implicated in redox-sensitive signalling pathways that are crucial for physiological cellular functions, including differentiation, growth, and proliferation [19,20]. NOXs are acknowledged as the primary generator of ROS in cells, along with a respiratory chain’s mitochondrial enzymes. Other than function of cell signalling and innate immune response, numerous diseases, including cancer, neurodegeneration, and muscular dystrophy, are exacerbated by these enzymes [21]. NOX has the qualities of a second messenger other than its responsibilities in host defence; the production of ROS by NOXs is highly regulated, and while NOXs are not ubiquitous, they are specified in certain membrane tissue and compartments. Superoxide is produced when NOXs transfer electrons from NADPH to heme groups, flavin adenine dinucleotide (FAD), and eventually to molecular oxygen. Genetic instability is associated with high amounts of ROS produced by exudative NOX activity, followed by proliferative senescence, DNA damage response activation, hyperproliferation, and occasionally, apoptosis [22]. NOXs are important in the pathophysiology of many CVDs. They have been recognised for their complexity, regulation, and specific function in the underlying molecular mechanisms of the pathogenic processes in CVDs [6].
Seven components have been identified; based on the isoform, they are produced in various cardiovascular signalling pathways and cell compartments and control a variety of processes, including inflammatory responses, migration, proliferation, differentiating, senescence and apoptosis. Each isoform contains up to seven regulatory subunits in addition to a catalytic subunit, known as NOX (NOX-1–5) or dual oxidase (DUOX); (DUOX–1-2, also known as NOX–6-7) [23]. NOX1, NOX2, NOX4, and NOX5 have been shown to have a considerable expression in the cardiovascular system, which will be the focus of this review (Table 1).
2.1.1. NOX1
NOX1 was discovered to be the first homolog of NOX2 and shared 60% amino acid similarity [23,24,25]. Despite the lack of information on NOX1 subcellular location, it has been hypothesised that this protein is membrane-localised and may be found in caveolar rafts [26]. NOX1 is broadly expressed in various cell types, with a particularly high level of expression in the colon epithelium [27,28]. The identification of NOX organiser 1 (NOXO1), a homolog of cytosolic protein p47phox, and NOX activator 1 (NOXA1), a homolog of cytosolic protein p67phox, provided evidence that NOX1 action is dependent on cytosolic subunits [29,30]. NOX1 requires the p22phox (membrane subunit), cytosolic subunits, and the Rac guanosine triphosphatase (GTPase) to be activated [31,32]. In short, the combination of several subunits is necessary for NOX1 activation [33] where it required p47phox, which then is transported to the membrane in which it binds to p22phox [33].
2.1.2. NOX2
NOX2 was the first enzyme from the NOX family to be identified [33]. NOX2, also known as gp91phox, has six transmembrane domains, with the C- and N-termini facing the cytoplasm. Furthermore, NOX2 is stabilised by constitutive interaction with p22phox [34,35]. NOX2 activation needs translocation and interaction with phosphorylated p47phox in order for other cytosolic components, such as p67phox and p40phox, to attach to the NOX2/p22phox complex [36,37,38]. Upon complex formation, the Rac GTPase first connects with NOX2 and then with p67phox, resulting in an active complex for superoxide generation via electron transfer from cytosolic NADPH to oxygen on the luminal or extracellular area [39,40].
2.1.3. NOX4
Geiszt et al. discovered that NOX4 has 39% sequence homology with NOX2 [41,42]. NOX4 activity is substantially influenced by p22phox [43], but not by cytosolic subunits [44]. Furthermore, the role of Rac in NOX4 activation remains debatable [45]. Polymerase delta-interacting protein 2 (Poldip2), a polymerase (DNA-directed) delta-interacting protein, acts as a positive regulator in vascular smooth muscle cells by interacting with NOX4/p22phox [46]. The connection of Poldip2 with p22phox was demonstrated by using glutathione S-transferase (GST) pull-down assays [46]. The use of Western blotting, real-time quantitative reverse transcription PCR (qRT-PCR), and immunohistochemistry on NOX4-rich tissues (e.g., aorta, lung, and kidney) revealed enhanced Poldip2 expression [46,47]. The role of Poldip2 as a positive regulator of NOX4 in conjunction with p22phox has been established in vascular smooth muscle cells utilising short interfering RNA (siRNA) against Poldip2 [46].
2.1.4. NOX5
NOX5, a protein with five isoforms, was found by two separate research groups. Banfi et al. [48] found isoforms a, b, c, and d, while Cheng et al. [49] discovered a fifth isoform, NOX5e or NOX5-S. NOX5 isoforms a-d have a lengthy, intracellular N-terminal domain with a Ca21-binding EF-hand region [49,50], but the fifth isoform lacks the EF-hand region and is structurally identical to NOX1-4 [49]. NOX5 isoforms a-d are activated by cytosolic calcium rather than p22phox or cytosolic subunits, indicating the presence of a Ca21-binding domain [51]. In contrast, the NOX5e isoform, which lacks a Ca21-binding domain, relies on the cAMP response element binding protein to function [52]. The N-terminal calmodulin-like domain with four Ca2+ binding sites makes NOX5 distinct from other NOX isoforms. NOX5 stimulation is Ca2+-dependent and independent of engagement with designated subunits. The fundamental structure of DUOX1 and DUOX2 is similar to NOX5; however, they are linked to an extracellular N-terminus and transmembrane domain [33].
Table 1.
NOX family and its regulatory subunit and expression in various tissues or cells.
| NOX Family | Regulatory Subunit | Expression Observed in | |||
|---|---|---|---|---|---|
| Tissue | Reference | Cells | Reference | ||
| 1 | NOXO1, NOXA1, p22phox, Rac | Smooth muscle Colon epithelial Uterus and placenta |
[53] [54] [55] |
Endothelial cells | [56] |
| Cardiomyocytes | [57] | ||||
| Vascular smooth muscle cells | [58] | ||||
| Human aortic smooth muscle cells | [59] | ||||
| Smooth muscle cells | [60] | ||||
| 2 | p22phox, p47phox, p40phox, and p67phox, Rac |
Heart tissue | [61] | Endothelial cells, adventitial cells | [62] |
| Cardiomyocytes, smooth muscle cells of the arteries | [63] | ||||
| Coronary microvascular endothelial cells | [64] | ||||
| Human umbilical vein endothelial cells | [65] | ||||
| 4 | p22phox, Poldip2 | Lung tissues Ovary and eye Fetal tissues Kidney |
[66] [67] [49] [68] |
Endothelial cells | [69] |
| Cardiomyocytes, adventitial cells | [70] | ||||
| Vascular smooth muscle cells | [71] | ||||
| Human aortic smooth muscle cells | [72] | ||||
| Mesangial cells | [73] | ||||
| Hepatocytes | [74] | ||||
| 5 | calmodulin-like domain with four Ca2+ | Fetal, spleen and uterus tissues Lung, heart, thymus, liver, kidney, skeletal muscle |
[49] [75] |
Vascular smooth muscle cells Cardiomyocyte |
[76] [70] |
2.2. Expression of NOX Isoforms-Derived ROS in CVD
Numerous pathways related to CVD are associated with NADPH oxidase-derived ROS produced by vascular and phagocytic cells. The vasculature expresses certain NADPH oxidases in a constitutive manner [77]. In CVD, imbalanced ROS production causes vascular injury by stimulating several processes, such as vascular cell proliferation, migration, the deposition of extracellular matrix (ECM) proteins, or inflammation, sequentially enhancing vascular remodelling. The NADPH oxidase family regulates vasculature remodelling and is a significant generator of ROS in the arterial wall during CVD.
In various CVDs, dysfunctional mitochondria are believed to be the intracellular producer of ROS. It has been demonstrated that NOX-derived ROS can reach the mitochondria and induce electron leakage and the formation of mitochondrial ROS, indicating that defective mitochondria are connected downstream of NOXs [33]. On the other hand, eNOS uncoupling activity is also modulated by mitochondrial ROS production. Peroxynitrite, a harmful radical formed when the O2− interacts with NO, can damage mitochondria by oxidising membrane lipids and electron transport chain complexes, resulting in defective cardiac contractile activity and impaired cardiac mitochondrial function [33]. Additionally, research has shown xanthine oxidase (XO) as a contributor of ROS in developing a range of CVDs. The enzyme xanthine oxidoreductase is first produced in the dehydrogenase form (xanthine dehydrogenase; XDH), which is then quickly turned by oxidation into the oxidase form (XO) [78]. The ROS, O2−, and hydrogen peroxide (H2O2) are produced via XO by directly delivering electrons to molecular oxygen (O2) through one-electron and two-electron reductions, respectively [79].
In general, different NOX subunits are related to various pathophysiology of CVD. NOX1 and NOX4 play a role in the remodelling of the vascular system in various clinical situations, including atherosclerosis, pulmonary hypertension, restenosis aortic stenosis, and hypertension [58]. NOX1, NOX2, and NOX4 are expressed in endothelial cells, vascular smooth muscle cells exhibit NOX1, NOX4, and NOX5, cardiomyocytes express NOX1, NOX2, NOX4, and NOX5, whereas NOX2 and NOX4 are mainly expressed by adventitial cells [33]. NOX1-derived ROS promote vascular remodelling upon triggering vascular smooth muscle cell dedifferentiation, as well as promoting its migration and proliferation with other activities, such as the deposition and remodelling of ECM proteins. Although NOX4 is shown to play a part in migration and proliferation of VSMC, it has also been demonstrated to maintain the smooth muscle phenotype. The role of NOX4 is debatable, but NOX1 plays a harmful part in vascular remodelling in several diseases. Therefore, NOX4 may have advantageous or detrimental effects based on the disease model or the specific cell site.
Studies using cell-specific NOX deletion will clarify the precise function of each NOX isoform, particularly in vascular remodelling models [77]. NOX1 and NOX2 overexpression has been shown in studies to promote inflammation and oxidative stress, which can lead to cardiologic conditions [80,81]. NOX4 has an impact on a variety of cellular processes related to vascular remodelling, including cell proliferation, apoptosis, senescence, cell differentiation, cell migration, and cell cycle regulation [46,82,83]. NOX4 expression has been found to increase in the heart in response to pressure overload induced by transverse aortic constriction (TAC) over 2–4 weeks, as well as after phenylephrine or angiotensin II infusion [84,85,86,87]. NOX5 is involved in platelet-derived growth factor (PDGF) and capillary-like structure formation. NOX5 downregulation has been shown to reduce the production of thrombin-stimulated growth factor and tube formation. In contrast, NOX5 upregulation increased the endothelial nitric oxide synthase (eNOS) activity, but decreased NO bioavailability [88]. Given these, NOX5 overexpression disrupts the acetylcholine-induced relaxation and phenylephrine-induced contractile effects [81,88,89,90].
There is evidence that angiotensin II (Ang II) is crucial in the development of cardiovascular disorders linked to hypertension. In cultured human aorta smooth muscle cells, angiotensin II increased the expression of the NOX subunit’s mRNA for NOX1, NOX4, p67phox, p47phox, and p22phox [59]. This finding raised the possibility that NOX1 and NOX4 are involved in the oxidative stress that angiotensin II causes in human vascular smooth muscle cells. Moreover, angiotensin II enhanced the protein expression of NOX subunits (NOX2, p22phox, p47phox, p40phox, and p67phox) in smooth muscle cells of the arteries obtained from gluteal biopsies of healthy individuals.
In atherosclerotic lesions of apolipoprotein E-deficient (ApoE/) mice and humans, the literature findings showed that expression of the NOX1 and/or NOXA1 subunits increased, suggesting that the enzyme plays a role in this disease state [91] (Figure 1). There is evidence that Nox1/ApoE/double knockout mice have a higher reduction in the formation of atherosclerotic lesions in the aortic arch than ApoE/single knockout animals [92]. NOX2 appears to be involved in the redox signalling involved in the start and progression of atherosclerosis. Overexpression of NOX2 in ApoE/mice boosted macrophage recruitment, but did not affect atherosclerosis progression [93].
Figure 1.

Schematic diagram of the NOX isoform-derived ROS production in cardiovascular diseases and natural products targeting those NOX isoforms leading to changes in intracellular targets. Natural products are divided into cardioprotective compounds targeting NOX2, NOX4, and p47phox, and vasculorprotective compound targeting NOX1, NOX2, NOX4, p47phox, and p22phox. The reduction in intercellular protein expression is noted in red font, while the upregulation of proteins is noted in green font in the diagram.
Furthermore, a previous study showed that global NOX2 deletion mice increased activity of NADPH oxidase isoforms contributes to worsening outcomes following stroke, at least in males [94], and administration of apocynin before cerebral ischaemia improves outcome in wild-type, but not NOX2-deficient mice, suggesting apocynin’s protective effects occur via NOX2 inhibition (Figure 1) [94]. Neuronal apoptosis and blood–brain barrier leakage, which are pathophysiological characteristics of ischemic stroke, are caused by NOX4-mediated oxidative stress and leads to neuronal damage [95]. Neither sex of NOX4-deficient mice was largely protected against oxidative stress, blood–brain barrier leakage, and neuronal death after temporary and permanent cerebral ischemia. However, NOX1 and NOX2-deficient mice did not show this protection.
In heart failure, ROS regulate fibroblast proliferation, collagen synthesis, and matrix metalloproteinase activation, resulting in cardiac hypertrophy, fibrosis, and necrosis, which can lead to endothelial and myocardial dysfunction [96]. When pressure overload was induced by constriction of the ascending aorta in NOX2-deficient mice, a large increase in messenger (m)RNA expression of p22phox and a slight increase in mRNA expression of p47phox was reported, whereas, in wild-type mice, there was a moderate increase in p22phox and no increase in p47phox. This was reflected by the ROS levels in the myocardium, which was twice as high compared to wild-type animals. Similarly, NOX4 expression was raised in cardiomyocytes in response to pressure overload and after myocardial infarction in NOX4−/− mice and a cardiomyocyte-targeted NOX4 transgenic model. NOX4−/− mice, on the other hand, exhibited much greater cardiac dilatation and contractile degradation than wild-type mice, but NOX4 transgenic mice had less hypertrophy and fibrosis than wild-type mice. These suggest that the NOX2 and NOX4 isoform and the subunits of the enzyme complex associated with it were identified as a critical source of ROS in pressure-induced left ventricular hypertrophy, and they contributed to pathophysiologic alterations such as redox-sensitive kinase activation and heart failure development (Figure 1).
Using NOX isoform-specific knockout (KO) mice, researchers studied the activities of endogenous NOX1, NOX2, and NOX4 in myocardial ischemic-reperfusion (I/R) damage. Systemic NOX1, NOX2, and NOX1/NOX2 double KO mice showed a significant reduction in myocardial infarct size following I/R, while systemic NOX4 KO mice did not [97]. Furthermore, in response to I/R, cardiac-specific NOX4 transgenic mice showed increases in ROS generation and infarct size [39]. These data imply that NOX2 and NOX4 play a role in myocardial I/R damage (Figure 1).
3. Natural Products Targeting NADPH Oxidase Pathway in Cardiovascular Disease
The use of medicinal herbs as an alternate treatment option has gained an incredible interest in modern medical systems, especially in CVDs. This review provides a comprehensive summary of the published scientific data on the regulation of NADPH oxidase pathways and its related molecular mechanisms of natural bioactive compounds in its therapeutic approach for the prevention and treatment of various CVDs experimental models (Table 2).
3.1. Berberine
Berberine or 5,6-dihydro-9,10-dimethoxy-benzo[g]-1,3-benzodioxolo [5,6-a]quinolizinium is found as an active component in the roots, stem bark, and rhizomes of Hydrastis canadensis (goldenseal) plants [98]. Berberine has been used as an Ayurvedic and Chinese medicine for over 3000 years due to its potent antimicrobial, antiprotozoal, and antidiarrheal properties [99]. Berberine is recognised as one of the most promising natural compounds for treating metabolic illnesses, such as hyperlipidemia [100], obesity [101], gout [102], non-alcoholic fatty liver disease (NAFLD) [103], and type 2 diabetes [104].
Cheng et al. demonstrated berberine improves endothelial function in human study and in vitro human umbilical vein endothelial cells (HUVECs) by reducing endothelial microparticle (EMP)-mediated oxidative stress [105]. In this study, healthy middle-aged people who received berberine therapy for one month demonstrated increased endothelium-dependent vasodilation, but not endothelium-independent vasodilation measured by flow-mediated vasodilation (FMD) and sublingual nitroglyceride-mediated vasodilation. Additionally, after berberine treatment, serum MDA and circulating CD31+/CD42 microparticles were considerably reduced. The same research group also discovered that pre-treatment with the antioxidants berberine and apocynin decreased NOX4 protein expression in HUVECs, while endothelial microparticles enhanced it. These findings suggest that NOX4 may play a significant role in enhancing EMP-mediated ROS generation, resulting in decreased NO bioavailability and endothelial cell damage, which can be reversed by berberine treatment.
In another study, Zhang et al. demonstrated that by increasing eNOS expression and decreasing NOX4 expression, berberine reduces palmitate-induced endothelial dysfunction, and this regulatory impact of berberine may be connected to the activation of AMP-activated protein kinase (AMPK) [106]. In cardiovascular cells, AMPK plays a significant role as an NADPH oxidase inhibitor. When activated, AMPK lowers ROS production by reducing NADPH oxidase activity, which, in turn, prevents the endothelial cell death caused by palmitic acid in cultured HUVECs. Berberine treatment activated eNOS and inhibited NOX4-derived ROS accumulation in endothelial cells, which will help reduce oxidative stress through activation of AMPK, which may help explain why berberine has protective effects on endothelial function. Berberine significantly increased AMPK and p-AMPK protein expression levels in palmitate-induced endothelial dysfunction in HUVECs; however, Akt and p-Akt protein expression was unchanged. The study suggests that berberine may be a novel therapeutic medication for the prevention and treatment of CVD, warranting further clinical research.
3.2. Paeonol
Paeonia suffruticosa has a variety of medicinal properties and has been used for thousands of years in traditional oriental medicine [107]. The main component isolated from the root bark of Paeonia suffruticosa is paeonol [107]. Paeonol, also known as 1-(2-hydroxy-4-methoxyphenyl) ethenone, is insoluble in water [108]. Paeonol has been shown in numerous in vitro and in vivo studies to be an antiinflammatory compound [109,110,111]. Paeonol has been linked to neuroprotection in diabetic encephalopathy [112], Parkinson’s disease [113], and Alzheimer’s disease [114].
In mice induced by tunicamycin, chronic treatment with paeonol protected endothelial function and stabilised blood pressure by inhibiting endoplasmic reticulum (ER) stress-associated reactive oxygen species (ROS) [115]. Tunicamycin (1 mg/kg/week) was injected intraperitoneally into male C57BL/6J mice for two weeks to induce ER stress. Their study demonstrated that tunicamycin-treated mice had higher blood pressure, loss of weight, and had impaired endothelium-dependent relaxations of the aorta, all of which were improved by co-treatment with paeonol, TUDCA (ER stress inhibitor), or tempol (antioxidant). Co-treatment with paeonol or tempol reduced tunicamycin-stimulated up-regulation of NADPH subunits, NOX2, and nitrotyrosine (marker for peroxynitrate, an indication of enhanced oxidative stress) in mice, compared to the control group as demonstrated via Western blot. An increased ROS formation in en face endothelium and O2− level was also observed in mice treated for 2 weeks with tunicamycin compared to the control group, as reflected by the intensity of DHE fluorescence staining and lucigenin-enhanced chemiluminescence (LEC). Additionally, chronic paeonol, TUDCA, and tempol treatment increased the phosphorylation of eNOS at Ser1176 in the aortas, whereas tunicamycin treatment decreased it. Thus, paeonol enhanced nitric oxide bioavailability in mouse aorta via inhibition of ROS, specifically on NOX2 [115].
Methotrexate (MTX) is known to increase reactive oxygen species (ROS) by increasing homocysteine, which is rapidly autoxidized and activates NOX, which generates ROS. In another separate study, paeonol reduced methotrexate (MTX)-induced cardiac damage in rats by decreasing NOX2 levels, increasing glutathione (GSH) levels, and elevating superoxide dismutase (SOD) activity [116]. Finally, paeonol protects against MTX-induced cardiac damage by decreasing oxidative and nitrosative stress, which, subsequently, suppressed the toll-like receptor 4 (TLR4) inflammatory pathway and also restored the histological structure.
These findings add to the growing evidence that paeonol could be used as a novel therapeutic drug or health supplement for patients with ER-stress-related cardiovascular disorders and methotrexate (MTX)-induced cardiac damage.
3.3. Thymoquinone
Thymoquinone (TQ) is a monoterpene compound identified chemically as 2-methyl-5-isopropyl-1, 4-benzoquinone and is the most prominent constituent of Nigella sativa seeds essential oil [117,118]. TQ has been studied for a wide range of pharmacological activities, which include antioxidant [119] anti-inflammatory [120], immunomodulatory [121], antihistamine [122], antimicrobial [123], and antitumour properties [124].
Zhang et al. reported thymoquinone protects apolipoprotein E-deficient (ApoE−/−) mice against angiotensin II (Ang II)-induced heart injury [125]. The ApoE−/− mice treated for 4 weeks with Ang II and TQ demonstrated a decreased area of cardiac fibrosis from Masson trichrome-stained sections compared to those treated with Ang II alone. This was further supported by reduced collagen I and III expression in the heart tissue Ang II-treated mice given TQ. Gene and protein expression studies showed elevated pro-inflammatory cytokines, tumour necrosis factor (TNF)- α, interleukin (IL)-1β and IL-6 in Ang II group, while TQ treatment suppressed this increase. Additionally, TQ decreased the expression of NOX4 and p53 in the cardiac tissue compared to those treated with Ang II alone. This group also showed NOX4, p53, collagen I and III, IL-1, IL-6 and TNF- α, protein expression levels were increased when in rat cardiac H9c2 cells treated with Ang II, which were reversed by TQ. Therefore, these results suggested that TQ could protect the heart from Ang II-induced injury suppressing oxidative stress, particularly NOX4.
Moreover, Guo et al. demonstrated that TQ inhibited inflammatory cell infiltration, the production of pro-inflammatory cytokines, apoptosis, oxidative stress, and PI3K/AKT pathway activation, all of which are indicators of sepsis-induced cardiac damage [126]. Four groups of male BALB/c mice were created randomly: the control, TQ, cecal ligation and puncture (CLP), and CLP + TQ groups. Following two weeks of TQ gavage on the mice, CLP was carried out. Histological alterations in the cardiac tissue after 48 h were evaluated. Markers linked to apoptosis, oxidative stress, inflammation, and the PI3K/AKT pathway were quantified. Compared to the control group, there were increases in NOX4 expression and decreases in Heme Oxygenase-1 (HO-1) and nuclear factor erythroid 2–related factor 2 (Nrf2) expression in the CLP group. However, NOX4 overexpression was normalised by TQ administration, followed by HO-1 and Nrf2 downregulation. In addition, TQ reversed the increase of IL-6, TNF- α, Bax, p-PI3K, Bcl-2, and p-AKT as well as decreased intestine histological changes. According to these findings, TQ is crucial in the treatment of sepsis-induced cardiac damage because it efficiently inhibited NOX4 mainly via PI3K/AKT pathways.
Furthermore, Chen et al. reported that through the activation of AMPK and suppression of MAPK signalling in vivo and in vitro, TQ has a protective impact on pathological heart hypertrophy [127]. Transverse aortic constriction (TAC) or a sham procedure was performed on male C57BL/6J mice, and both procedures were followed by six weeks of TQ therapy. Neonatal rat cardiomyocytes (NRCMs), used in in vitro investigations to stimulate cardiomyocyte hypertrophy, were treated to phenylephrine (PE) stimulation. The result also showed that ventricular tissues with 6 weeks of pressure overload had much higher ROS levels, while tissues treated with TQ had levels that were nearly identical to those of the group that underwent sham surgery. TQ improved the mRNA expression of genes associated to oxidative stress (NOX4, SOD1, and SOD2) in mice from the TAC group. Thus, TQ was shown to be a potential drug for the treatment of cardiac hypertrophy, since it protects against the condition by reducing NOX4 in an AMPK-dependent way.
3.4. Reinioside C
Reinioside C is the primary component extracted from Polygala fallax Hemsl. [128]. Polygala fallax Hemsl. is a Polygalaceae genus shrub or small tree that grows in the shade and humid environment of the valley forest and is primarily distributed in Jiangxi, Fujian, Hunan, Guangxi, and Yunnan provinces of China [129]. Polygala fallax Hemsl., a popular Chinese medicinal herb, has been used to treat various ailments, including infective inflammation and hypercholesterolemia [130].
Bai et al. demonstrated that the inhibition of NADPH oxidase-ROS-ERK1/2-NF-kB-AP-1 pathway by reinioside C reduces Ang II-induced proliferation of vascular smooth muscle cells (VSMCs) [131]. Rat aortic smooth muscle cells A10 (A10 VSMCs) were pre-treated with reinioside C (3, 10 or 30 μM), diphenyleneiodonium (DPI) a specific NADPH inhibitor (10 μM), the ERK1/2 inhibitor, PD98059 (40 μM), or the NF-κB inhibitor, pyrrolidine dithiocarbamate (PDTC) (10 μM), for 1 h before being cultured with Ang II (1 μM) for 24 h. Treatment with Ang II elevated mRNA expression of NADPH oxidase subunits (both NOX1 and NOX4) and intracellular ROS formation, and pre-treatment with reinioside C reduced significantly these effects of Ang II in a concentration-dependent manner. The phosphorylation of ERK1/2 (p-ERK1/2) elicited by Ang II was also significantly inhibited in a concentration-dependent manner after cells were pre-treated with reinioside C. Pre-treatment of the cells with DPI or the inhibitor PD98059 had the same effect. Ang II stimulation significantly increased IκBα degradation, NF-kB activity, AP-1 subunits (c-fos and c-jun), and c-myc of mRNA expression while in a concentration-dependent manner, pre-treatment of cells with reinioside C significantly reduced these effects of Ang II. Collectively, this suggest that the effect of reinioside C is by inhibition of NADPH oxidase-ROS, specifically NOX1 and NOX4 mRNA expression, and inhibition of ERK1/2-NF-κB-AP-1 pathway.
Bai et al. demonstrated oxidised low-density lipoprotein ox-(low-density lipoprotein (LDL) (100 µg/mL) for 24 h increased the adhesion of monocytes as well as elevated the expression of ICAM-1 and P-selectin in HUVECs and human monocytoid (THP-1) cells [132]. This effect by ox-LDL were attenuated by reinioside C. To determine the role of NADPH oxidase/ROS/ NF-κB pathway, NADPH oxidase subunit (NOX2 and p22phox) mRNA expression, intracellular ROS level, and NF-κB activity were measured on ox-LDL treated endothelial cells. The results revealed that reinioside C attenuated the elevated mRNA expression of NOX2 and p22phox, two NADPH oxidase subunits, reduced intracellular ROS levels and NF-kB activity caused on by ox-LDL. These findings imply that reinioside C reduces ox-LDL-induced adhesion molecule expression (P-selectin and ICAM-1) and monocyte adherence to endothelial cells via blocking the NADPH oxidase/ROS/NF-κB pathway.
3.5. Curcumin
Curcumin (CUR), the main ingredient of turmeric, is derived from the root of Curcuma longa Linn [133]. Turmeric is a spice that is made from the rhizomes of the Curcuma longa plant, which belongs to the ginger family (Zingiberaceae) [134]. Curcuma longa grows wild throughout the Indian subcontinent as well as in tropical areas, such as Indonesia and Malaysia [135]. Curcumin is still used as an alternative medicinal treatment in many parts of Southeast Asia today to cure a variety of diseases, including stomach upset [136], jaundice [137], arthritis, sprains, wounds, and skin infections [138].
Boonla et al. demonstrated CUR reduces oxidative stress and improves endothelial dysfunction and vascular remodelling in 2 kidneys 1 clip (2K-1C) hypertensive rats [139]. A model of 2K-1C renovascular hypertension in male Sprague–Dawley rats was used. The rats were treated with CUR at a dose of 50 or 100 mg/kg/day. After 6 weeks of treatment, elevated p47phox expression and higher superoxide production was observed in the vascular wall of 2K-1C rat’s aortas and carotid arteries, which was decreased with CUR treatment. Furthermore, 2K1C hypertension was associated with increased oxidative stress, which can be seen in elevated malondialdehyde (MDA) and protein carbonyl levels. CUR significantly reduced superoxide production, plasma MDA, and protein carbonyl levels in 2K-1C rats, and these negative effects were linked to a decrease in the p47phox NADPH oxidase subunit. CUR treatment also improved impaired endothelial function and this was related to increased nitrate/nitrite levels and higher eNOS expressions 2K-1C mice treated with CUR. CUR treatment prevented morphological alterations in the aorta wall found in 2K-1C hypertensive rats and significantly reduced matrix metalloproteinase-2 (MMP-2) and matrix metalloproteinase-9 (MMP-9) levels in the aortic walls generated by 2K-1C hypertension.
In the other study, CUR (30 mol/L) dramatically reduced the amount of p47phox that was overproduced by Lipopolysaccharides (LPS) in VSMCs [140]. To determine if LPS-induced cytokine upregulation in VSCMs was due to an increase in ROS, cells were pre-treated with or without DPI (20 mol/L), which inhibits NADPH oxidase before adding CUR for 1 h, and then cells were stimulated with LPS (1 g/mL) for 24 h. DPI and CUR both reduced LPS-induced Monocyte chemoattractant protein-1 (MCP-1), TNF-α, and NO upregulation. Pre-treatment of VSMCs with a combination of DPI and CUR reduced the LPS-induced rise in inflammatory cytokines synergistically. These findings imply that CUR’s anti-LPS-induced inflammatory activity is partially dependent on decreasing the formation of NADPH-mediated intracellular ROS in VSMCs.
3.6. Celastrol
Celastrol is one of the active compounds derived from Tripterygium wilfordii Hook F, a traditional Chinese medicinal plant which has been utilised for centuries [141]. Celastrol has been shown to have anti-inflammatory properties in animal models of lupus [142], Alzheimer’s disease [143], rheumatoid arthritis [144], and reduced oxidative stress injury by upregulation of Nrf2/anti-oxidant enzymes pathway [145,146].
Li M. et al. revealed that through activation of the Nrf2/ERK1/2/NOX2 signal pathway, celastrol reduces Ang II-mediated damage to HUVECs [147]. The cells were treated with arecelastrol, Ang II, Ang II + celastrol, or Ang II + celastrol+, a specific inhibitor of Nrf2 (brusatol), for 24 h. The fluorescence intensity of 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA) was used to estimate intracellular reactive oxygen species generation, and NADPH oxidase activity was measured using the AmpliteTM Fluorime NADPH test Kit. At a concentration of 50 nmol/L, celastrol was able to completely inhibit the activity of NADPH oxidase and reduced the production of ROS. NADPH oxidase activity and reactive oxygen species levels in HUVECs significantly increased when co-cultured with brusatol or PD98059 (a specific Nrf2 inhibitor) compared to the celastrol. Western blotting showed celastrol significantly decreased NOX2 expression elevated by Ang II, but this effect was reversed by brusatol and PD98059. Celastrol also effectively increased anti-oxidant enzymes activities (SOD, GPx) while decreasing MDA levels in comparison to the Ang II group.
In another study, Liu et. al. demonstrated that celastrol dramatically reduced aortic valvular interstitial cell (AVIC) calcification in vitro by reducing NOX2 activity, and it greatly reduced the severity of aortic valve fibrosis, calcification, and stenosis in a rabbit model of calcific aortic valve disease (CAVD) in vivo [148]. Endogenous NOX2 knockdown reduced AVIC calcification by 39%, with an inhibitory effect similar to celastrol therapy. Celastrol therapy decreased NOX2 levels in both AVICs with osteogenic stimulation and in calcified rabbit valves, suggesting that the effect may be due to less NOX2 being induced by reduced calcification rather than by the celastrol’s direct action. Celestrol treatment also reduced NOX2 expression and ROS generation in aortic valves (AVs) of high-cholesterol (HC) plus vitamin D2-fed rabbits. Celastrol greatly decreased aortic valve ROS production, fibrosis, calcification, and severity of aortic stenosis in a rabbit CAVD model, with less left ventricular dilatation and better retained contractile function. Celastrol is useful in the treatment of CAVD, most likely through inhibiting the NOX2 in AVICs.
3.7. Apocynin
Apocynin (4-hydroxy-3-methoxyacetophenone) was isolated from the roots of Apocynum cannabinum (Canadian hemp) and was used as an approved remedy for dropsy and cardiac problems [149]. Apocynin has the ability to scavenge free radicals and act as a non-specific NOX inhibitor [150]. Apocynin has anti-inflammatory actions in atherosclerosis [151], neuroinflammatory [152], respiratory [153], and renal diseases [154] as well as cancers [155], via a ROS-dependent mechanism.
Perassa et al. demonstrated in spontaneously hypertensive rats (SHR), apocynin lowers blood pressure and restores vascular endothelial function [156]. Their results showed that in SHR, apocynin treatment significantly lowers systemic ROS. Furthermore, systolic blood pressure (SBP) and diastolic blood pressure (DBP) in apocynin-treated SHR were lower than in untreated SHR. These findings imply that reduced ROS production generated by apocynin administration may play a crucial role in the antihypertensive effect of apocynin found in several experimental hypertension models, including SHR. In SHR treated with apocynin, overexpression of NOX2 and the subunit p47phox decreased, while expression of NOX1, NOXO1, and NOX4 remains unchanged. In comparison to untreated SHR, apocynin-treated SHR reported reduced NOX2 expression. Apocynin treatment decreased the expression of p47phox in the aorta of SHR as compared to an untreated SHR. The expression of p47phox was comparable in the treated SHR and untreated Wistar rats’ aortas. The direct interaction between apocynin and p47phox prevents p47phox translocation and inhibits NOX. In SHR, apocynin improved endothelial function in the resistance and conductance arteries and increased potency to SNP in intact resistance vasculature, which may be because apocynin enhanced eNOS expression in endothelial cells. SHR aortic endothelial cells (AEC) treated with apocynin had higher [Ca2+] compared to untreated. Moreover, stimulation with acetylcholine doubled the amount of nitric oxide released in the AEC of treated SHR compared to untreated SHR. The ROS levels in treated SHR AEC were lower than in untreated SHR AEC.
3.8. Oleanolic acid
Oleanolic acid (OA) is a natural compound found in a variety of foods and medicinal plants [157]. It is a pentacyclic triterpenoid found in abundance in the Oleaceae family of plants, such as the olive tree [158]. Traditionally, oleanolic acid has been taken as a hepatic medication in China for over 20 years due to its hepatoprotective properties [159]. OA has been linked to antioxidant [160], antimicrobial [161], anti-inflammatory, and antidiabetic properties [162].
Jiang Q et al. reported that the anti-atherosclerosis action of oleanolic acid is facilitated by the modulation of oxidized-LDL receptor-1 (LOX1) [163]. Sixteen male quails (Coturnix coturnix) fed a high-fat diet and OA or simvastatin were sacrificed, and the serum was collected while the aorta was dissected out to measure NO, MDA, SOD, catalase (CAT), glutathione (GSH), NADPH, and GSH-Px levels. Their results showed that a high-fat diet significantly deteriorated the serum and aorta lipid profile as observed with increased total cholesterol, triacylglycerol, and LDL and reduced high-density lipoprotein (HDL), and treatment with OA or simvastatin reversed these lipid profile. OA and simvastatin possess an antioxidant effect, which was demonstrated with decreased MDA levels and increased GSH and NADPH levels. The activities of SOD, CAT, and GSH-px were also increased in high-fat diet quails by OA or simvastatin.
The same group also treated HUVECs with ox-LDL 200 g/mL for 24 h, with or without pre-treatment with OA (5, 10, or 20 μM) or vitamin E 20 μM for 24h and determined cell viability by MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) tetrazolium reduction assay and ROS by DCFDA staining. Cell viability of HUVECs were decreased and production of ROS were increased after exposed by ox-LDL, which was reduced by pre-treatment with OA or vitamin E. The protein levels of gp91phox, p67phox, and p47phox (subunits of NADPH oxidase) were decreased in pre-treatment with OA after exposure to ox-LDL. Both Ho-1 and Nrf2 expression were significantly increased after 24 h of exposure to ox-LDL in HUVECs, whereas pre-treatment with vitamin E or OA further increased expression of Ho-1 and Nrf2. Additionally, LOX-1 expression was significantly increased, but pre-treatment with vitamin E or OA effectively reduced it in a concentration-dependent manner. Knockdown of LOX-1 effectively inhibited ox-LDL-induced cytotoxicity in HUVECs and also reduced ox-LDL-induced expression of Nrf2 and Ho-1 and NADPH oxidase (gp91phox, p67phox, and p47phox). These findings show NADPH oxidase may contribute to cytotoxicity, as evidenced by the fact that cytoprotection was followed by a reduction in the expression of NADPH oxidase subunits.
3.9. Quercetin
Quercetin (3, 5, 7, 3′, 4′-pentahydroxyflavone) is a member of the flavonoids family and one of the most prominent dietary antioxidants [164]. It can be found in various meals, including vegetables, fruits, tea, and wine, as well as a variety of healthy goods [165]. In addition, several clinical studies have shown that quercetin supplementation can help prevent and treat a variety of chronic conditions, including cardiovascular issues [166], diabetes mellitus [167], cancer [168], and obesity [169].
Galindo P et al. reported in SHRs that quercetin has cardiovascular protective effects when given orally or intraperitoneally (i.p.) [170]. SHR were randomly assigned to four experimental treatments, which are: (1) control group; (2) 10 mg kg−1 quercetin, single dose; (3) 10 mg kg−1 quercetin by oral gavage, two daily doses; and (4) 10 mg kg−1 quercetin i.p. injection. The results showed that when compared to normotensive Wistar–Kyoto rats, SHR aortas showed significantly reduced endothelium-dependent relaxation (EDR) and nitric oxide (NO)-dependent vasodilator responses to ACh in arteries stimulated by phenylephrine, which was reversed with chronic oral quercetin administration. The aortic superoxide formation was also decreased with chronic oral quercetin treatment. protein expression of the NADPH subunits p47phox, NOX1, and NOX4 were reduced by quercetin treatment in the aorta via oral, but via i.p. administration. Intraperitoneal quercetin treatment failed to ameliorate endothelial dysfunction and reduced both NADPH oxidase activity and protein expression of any NADPH oxidase subunits (p47phox, NOX1, and NOX4), suggesting that oral quercetin was superior to intraperitoneal administration for the protection from cardiovascular complications in SHR.
Wan et al. explored the effects of quercetin on NOX2, eNOS, and inducible NOS (iNOS) after myocardial ischemia-reperfusion injury (MIRI) in rabbits [171]. NOX2 mRNA expression was higher in I/R hearts after 30 min of coronary ligation followed by 12 h of reperfusion, as determined by real-time PCR, compared to control hearts. Western blotting tests, which provide a real assessment of protein expression, were also done to explore the relative expression of NOX2 in both strains, and NOX2 protein expression was found to be significantly increased in I/R hearts. NOX2 mRNA and protein expression in I/R hearts following quercetin administration were lower in I/R + quercetin rabbits than in I/R rabbits. Similarly, NOX2 mRNA and protein levels in quercetin treated animals were significantly lower than in control rabbits. Quercetin suppressed not only NOX2 but also iNOS mRNA, eNOS mRNA, and protein expression induced by MIRI [171]. The current study provides the first evidence that quercetin, a flavonoid, inhibits cardiac ischemia-reperfusion-induced NOX2, iNOS, eNOS mRNA, and protein expression. As a result, quercetin could be a potential antioxidant for cardioprotection.
3.10. Delphinidin-3-Glucoside
Delphinidin, a prominent anthocyanins found in pigmented fruits and vegetables such as pomegranates, berries, dark grapes, eggplant, tomatoes, and carrots, is one of the most common anthocyanins [172]. Delphinidin is a potent radical scavenger capacity for superoxide [173]. Delphinidin has been shown to have anti-inflammatory [174], antioxidant [175], anticancer [176], cardiovascular protection [177], and neuroprotection [178] properties.
Xie et al. demonstrated delphinidin-3-glucoside effects on ox-LDL-induced apoptosis and oxidative stress in cultured porcine aortic endothelial cells (PAEC) [179]. PAEC were treated with ox-LDL, delphinidin alone, and a co-treatment of delphinidin and ox-LDL. Their findings revealed the amount of intracellular superoxide in PAEC rose more than two-fold after a 2 h treatment with 100 μg/mL ox-LDL as measured using 2,7-dichlorodihydrofluorescein diacetate (H2DCF-DA) and enhanced lucigenin assay, respectively. Compared to the control, delphinidin alone at 100μM decreased intracellular superoxide by more than 50%. Compared to ox-LDL alone, co-treatment with delphinidin and ox-LDL reduced the amount of superoxide elevated from ox-LDL in endothelial cells. Moreover, increased levels of NOX2, NOX4, p22phox, and caspase 3 produced by ox-LDL in PAEC were decreased by co-treatment with delphinidin. The data suggest that ox-LDL caused oxidative stress and apoptosis in endothelial cells, which was related with NOX activation, mitochondrial respiration chain enzyme impairment, and the disruption of critical apoptosis regulators.
Chen et al. evaluated the effect of delphinidin in a model of cardiac hypertrophy using the transverse aortic constriction (TAC)-induced pressure overload method and primary culture of neonatal rat cardiomyocytes. The result showed that delphinidin reduces pathological cardiac hypertrophy by suppressing the NOX/MAPK signalling pathway to counteract oxidative stress [180]. In comparison to the sham control group, there was also an increase in ROS levels and NOX activity. However, delphinidin at the high dosage (15 mg/kg/day) prevented these changes. In Ang II-induced hypertrophy of neonatal rat cardiomyocytes (NRCMs), the NOX subunit proteins p22phox, p47phox, p40phox, p67phox, and gp91phox were all considerably upregulated by Ang II stimulation, while delphinidin coadministration inhibited the upregulation of p47phox. Compound C, an AMPK inhibitor, prevented the delphinidin-mediated upregulation of Rac1 and lowered expression levels of p47phox. The expression of pathogenic genes, the formation of ROS, and NOX activity were all raised by compound C, which also removed the delphinidin-mediated reduction of the Ang II-induced hypertrophy of neonatal rat cardiomyocytes (NRCMs). Therefore, these findings revealed that delphinidin activated AMPK, which inhibited ROS production, particularly p47phox, and pathological hypertrophy by downregulating NOX.
Table 2.
List of natural products that target the NADPH oxidase pathway in CVD. The arrow 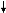 indicate down-regulation and
indicate down-regulation and  indicate up-regulation of the markers.
indicate up-regulation of the markers.
| Natural Product | Plant | Effective Dosage |
Induction of Cardiovascular Condition | Model | Result | NADPH Oxidase Subunit Involved | Reference |
|---|---|---|---|---|---|---|---|
Berberine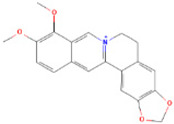
|
Rhizoma Coptidis | 10 μM | Endothelial microparticles (EMPs)-induced endothelial dysfunction | Human study In vitro using HUVECs |
ROS, NOX4, NO, MDA, CD31+/CD42 microparticles |
NOX4 | [105] |
| 5.0 μmol/L | Palmitate-induced endothelial dysfunction | In vitro using HUVECs |
eNOS, AMPK, NOX4 |
NOX4 | [106] | ||
Paeonol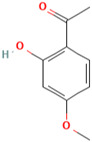
|
Paeonia suffruticosa | 20 mg/kg/day | Tunicamycin-induced endothelial dysfunction | In vivo |
ROS, GRP78, ATF6 and p-eIF2α, NOX2 eNOS, NO |
NOX2 | [115] |
| 100 mg/kg/day | Methotrexate (MTX)-induced | In vivo |
eNOS, NO, GSH, SOD NOX2, MDA, TLR4 |
NOX2 | [116] | ||
Thymoquinone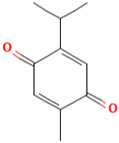
|
Nigella sativa | 50 mg/kg/day 20 µmol/L |
Angiotensin II-induced cardiac damage | In vivo In vitro (H9c2 cells) |
IL-1, IL-6, TNF, collagen I, collagen III NOX4 and p53 |
NOX4 | [125] |
| 100 mg/kg/day | Sepsis-induced cardiac damage | In vivo |
IL-6, TNF, Bax, NOX4, p-PI3K, and p-AKT Bcl-2, Ho-1, and Nrh2 expression |
NOX4 | [126] | ||
| 50 mg/kg/day 5 µM |
Transverse aortic constriction | In vivo In vitro (neonatal rat cardiomyocytes) |
mRNA levels of NOX4, SOD1, SOD2
|
NOX4 | [127] | ||
Reinioside C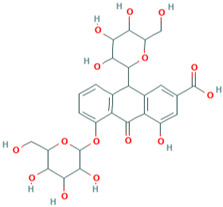
|
Polygala fallax Hemsl. | 30 µM | Arteriosclerosis | In vitro (VSMCs) |
ROS, NOX1, NOX4, IB degradation, NF-kB activity, and p-ERK1/2 |
NOX1 NOX4 |
[131] |
| 10 µM | Atherosclerosis | In vitro (HUVECs, THP-1) |
NOX2 and p22phox, ICAM-1, P-selectin, NF-κB |
NOX2 and p22phox | [132] | ||
Curcumin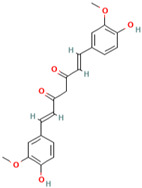
|
Curcuma longa Linn | 100 mg/kg/day | Renovascular hypertension | In vivo (2K-1C model) In vitro (VSMCs) |
MMP-2 and MMP-9, ROS, MDA and p47phox EDR, eNOS |
p47phox | [139] |
| 30 μmol/L | Atherosclerosis | In vitro (VSMCs) |
/> p47phox, MCP-1, TNF |
p47phox | [140] | ||
Celastrol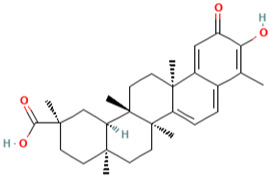
|
Tripterygium wilfordii Hook F | 50 nmol/L | Hypertension | In vitro (HUVECs) |
MDA, NOX2 SOD and GPx-SH |
NOX2 | [147] |
| 1 mg/kg/day 10 nmol/L |
Calcific aortic valve disease | In vivo (rabbit) In vitro (AVICs) |
NOX2 |
NOX2 | [148] | ||
Apocynin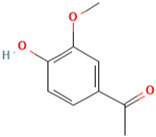
|
Apocynum cannabinum | (30 mg/kg) | Hypertension | In vivo (SHR rats) In vitro (endothelial cells) |
ROS, MAP, HR, SBP, DBP, NOX2 and p47phox eNOS, cytosolic calcium, and NO |
NOX2 and p47phox | [156] |
Oleanolic acid
|
Oleaceae | 100 mg/kg/day 20 µM |
Atherosclerosis | In vivo (high-fat diet-induced atherosclerosis in quails) In vitro (HUVECs) |
MDA, GSH, gp91phox, p67phox and p47phox, LDL HDL, SOD, CAT GPx-SH, LOX-1, Nrf2 and ho-1 |
gp91phox, p67phox and p47phox | [163] |
Quercetin
|
Berries, onions, and red wine | 10 mg/kg/day | Hypertension | In vivo (SHR) |
SBP, HR, NOX1, NOX4, p47phox EDR |
NOX1, NOX4, p47phox and p22phox | [170] |
| 1 mg/kg | Myocardial ischemia-reperfusion injury | In vivo (rabbit) |
NOX2, eNOS, iNOS |
NOX2 | [171] | ||
Delphinidin
|
Pigmented fruits and vegetables | 100 μM | Hypercholesterolemia | In vitro (ox-LDL-induced ROS in PAEC) |
NOX2, NOX4, and p22phox, caspase 3 |
NOX2, NOX4, and p22phox | [179] |
| 15 mg/kg/day 50 μM |
Cardiac hypertrophy | In vivo (TAC-induced pressure overload C57BL/6 mice) In vitro (neonatal rat cardiomyocytes) |
p47phox |
p47phox | [180] |
4. Conclusions
A variety of natural bioactive compounds have been shown to possess potent medicinal properties in treating and preventing cardiovascular diseases. It is demonstrated that natural bioactive compounds provide an advantage as a protective agent or supplementary or combination therapy against cardiovascular diseases. Our review concludes that various natural bioactive compounds provide protection against cardiovascular diseases by targeting the NADPH oxidase pathway. These include: berberine targeting NOX4; paeonol targeting NOX2; thymoquinone targeting NOX4; reinioside C targeting NOX1,2,4, and p22phox; curcumin targeting p47phox; celastrol targeting NOX2; apocynin targeting NOX2 and p47phox; oleanolic acid targeting gp91phox, p67phox, and p47phox; quercetin targeting NOX1,2,4, p47phox, and p22phox; and delphinidin targeting NOX2, NOX4, p22phox and p47phox. However, there are limited clinical, pharmacokinetic, and pharmacodynamic studies of these natural bioactive compounds; thus, restricting the potential use of the natural bioactive compounds for supplementary treatment and management plans for patients with cardiovascular diseases. Future research in both fundamental, pharmacokinetic, or pharmacodynamics profile and clinical trials are warranted to explore the underlying mechanism and the effectiveness of those listed natural bioactive compounds to prevent or reduce the occurrence of cardiovascular diseases, specifically coronary artery disease.
Author Contributions
Conceptualization, C.K.W.; methodology, C.K.W.; writing—original draft preparation, S.S.M.S., W.Y.S., S.Z.S.A.K., R.A., N.R.A.R. and C.K.W.; writing—review and editing, S.S.M.S., C.K.W., D.D.M., S.A.M. and N.H.Z.; supervision, C.K.W.; project administration, C.K.W. and S.S.M.S.; funding acquisition, C.K.W. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Not applicable.
Funding Statement
This research was funded by Geran Penyelidikan Khas (GPK) Universiti Teknologi MARA: 600-RMC/GPK 5/3 (269/2020) and Geran Insentif Penyeliaan (GIP) Fasa 3/2021. Teknologi MARA: 600-RMC/GIP 5/3 (079/2021).
Footnotes
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
References
- 1.Wright J.M., Musini V.M., Gill R. First-line drugs for hypertension. Cochrane Database Syst. Rev. 2018;4:CD001841. doi: 10.1002/14651858.CD001841.pub3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mahadir Naidu B., Mohd Yusoff M.F., Abdullah S., Musa K.I., Yaacob N.M., Mohamad M.S., Sahril N., Aris T. Factors associated with the severity of hypertension among Malaysian adults. PLoS ONE. 2019;14:e0207472. doi: 10.1371/journal.pone.0207472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Roth G. Global Burden of Disease Collaborative Network. Global Burden of Disease Study 2017 (GBD 2017) Results. Seattle, United States: Institute for Health Metrics and Evaluation (IHME), 2018. Lancet. 2018;392:1736–1788. doi: 10.1016/S0140-6736(18)32203-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sharifi-Rad J., Rodrigues C.F., Sharopov F., Docea A.O., Can Karaca A., Sharifi-Rad M., Kahveci Karıncaoglu D., Gülseren G., Şenol E., Demircan E., et al. Diet, Lifestyle and Cardiovascular Diseases: Linking Pathophysiology to Cardioprotective Effects of Natural Bioactive Compounds. Int. J. Environ. Res. Public Health. 2020;17:2326. doi: 10.3390/ijerph17072326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Madamanchi N.R., Runge M.S. Redox signaling in cardiovascular health and disease. Free Radic. Biol. Med. 2013;61:473–501. doi: 10.1016/j.freeradbiomed.2013.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sirker A., Zhang M., Shah A.M. NADPH oxidases in cardiovascular disease: Insights from in vivo models and clinical studies. Basic Res. Cardiol. 2011;106:735–747. doi: 10.1007/s00395-011-0190-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Schramm A., Matusik P., Osmenda G., Guzik T.J. Targeting NADPH oxidases in vascular pharmacology. Vasc. Pharmacol. 2012;56:216–231. doi: 10.1016/j.vph.2012.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Khavandi K., Baylie R.A., Sugden S.A., Ahmed M., Csato V., Eaton P., Hill-Eubanks D.C., Bonev A.D., Nelson M.T., Greenstein A.S. Pressure-induced oxidative activation of PKG enables vasoregulation by Ca2+ sparks and BK channels. Sci. Signal. 2016;9:ra100. doi: 10.1126/scisignal.aaf6625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Paravicini T.M., Montezano A.C., Yusuf H., Touyz R.M. Activation of vascular p38MAPK by mechanical stretch is independent of c-Src and NADPH oxidase: Influence of hypertension and angiotensin II. J. Am. Soc. Hypertens. JASH. 2012;6:169–178. doi: 10.1016/j.jash.2012.01.002. [DOI] [PubMed] [Google Scholar]
- 10.Burger D., Montezano A.C., Nishigaki N., He Y., Carter A., Touyz R.M. Endothelial microparticle formation by angiotensin II is mediated via Ang II receptor type I/NADPH oxidase/ Rho kinase pathways targeted to lipid rafts. Arterioscler. Thromb. Vasc. Biol. 2011;31:1898–1907. doi: 10.1161/ATVBAHA.110.222703. [DOI] [PubMed] [Google Scholar]
- 11.Meng F.G., Zhang Z.Y. Redox regulation of protein tyrosine phosphatase activity by hydroxyl radical. Biochim. Biophys. Acta. 2013;1834:464–469. doi: 10.1016/j.bbapap.2012.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Knock G.A., Ward J.P. Redox regulation of protein kinases as a modulator of vascular function. Antioxid. Redox Signal. 2011;15:1531–1547. doi: 10.1089/ars.2010.3614. [DOI] [PubMed] [Google Scholar]
- 13.Santilli F., D’Ardes D., Davì G. Oxidative stress in chronic vascular disease: From prediction to prevention. Vasc. Pharmacol. 2015;74:23–37. doi: 10.1016/j.vph.2015.09.003. [DOI] [PubMed] [Google Scholar]
- 14.Rossello X., Pocock S.J., Julian D.G. Long-Term Use of Cardiovascular Drugs: Challenges for Research and for Patient Care. J. Am. Coll. Cardiol. 2015;66:1273–1285. doi: 10.1016/j.jacc.2015.07.018. [DOI] [PubMed] [Google Scholar]
- 15.Lourenço S.C., Moldão-Martins M., Alves V.D. Antioxidants of Natural Plant Origins: From Sources to Food Industry Applications. Molecules. 2019;24:4132. doi: 10.3390/molecules24224132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Atanasov A.G., Zotchev S.B., Dirsch V.M., Orhan I.E., Banach M., Rollinger J.M., Barreca D., Weckwerth W., Bauer R., Bayer E.A., et al. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021;20:200–216. doi: 10.1038/s41573-020-00114-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ooi B.K., Chan K.G., Goh B.H., Yap W.H. The Role of Natural Products in Targeting Cardiovascular Diseases via Nrf2 Pathway: Novel Molecular Mechanisms and Therapeutic Approaches. Front. Pharmacol. 2018;9:1308. doi: 10.3389/fphar.2018.01308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Panday A., Sahoo M.K., Osorio D., Batra S. NADPH oxidases: An overview from structure to innate immunity-associated pathologies. Cell. Mol. Immunol. 2015;12:5–23. doi: 10.1038/cmi.2014.89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Coso S., Harrison I., Harrison C.B., Vinh A., Sobey C.G., Drummond G.R., Williams E.D., Selemidis S. NADPH oxidases as regulators of tumor angiogenesis: Current and emerging concepts. Antioxid. Redox Signal. 2012;16:1229–1247. doi: 10.1089/ars.2011.4489. [DOI] [PubMed] [Google Scholar]
- 20.Sies H., Jones D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020;21:363–383. doi: 10.1038/s41580-020-0230-3. [DOI] [PubMed] [Google Scholar]
- 21.Ryu J.K., Rafalski V.A., Meyer-Franke A., Adams R.A., Poda S.B., Rios Coronado P.E., Pedersen L.Ø., Menon V., Baeten K.M., Sikorski S.L. Fibrin-targeting immunotherapy protects against neuroinflammation and neurodegeneration. Nat. Immunol. 2018;19:1212–1223. doi: 10.1038/s41590-018-0232-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Magnani F., Mattevi A. Structure and mechanisms of ROS generation by NADPH oxidases. Curr. Opin. Struct. Biol. 2019;59:91–97. doi: 10.1016/j.sbi.2019.03.001. [DOI] [PubMed] [Google Scholar]
- 23.Drummond G.R., Selemidis S., Griendling K.K., Sobey C.G. Combating oxidative stress in vascular disease: NADPH oxidases as therapeutic targets. Nat. Rev. Drug Discov. 2011;10:453–471. doi: 10.1038/nrd3403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cifuentes-Pagano E., Csanyi G., Pagano P.J. NADPH oxidase inhibitors: A decade of discovery from Nox2ds to HTS. Cell. Mol. Life Sci. 2012;69:2315–2325. doi: 10.1007/s00018-012-1009-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kleniewska P., Piechota A., Skibska B., Gorąca A. The NADPH oxidase family and its inhibitors. Arch. Immunol. Ther. Exp. 2012;60:277–294. doi: 10.1007/s00005-012-0176-z. [DOI] [PubMed] [Google Scholar]
- 26.Helmcke I., Heumüller S., Tikkanen R., Schröder K., Brandes R.P. Identification of structural elements in Nox1 and Nox4 controlling localization and activity. Antioxid. Redox Signal. 2009;11:1279–1287. doi: 10.1089/ars.2008.2383. [DOI] [PubMed] [Google Scholar]
- 27.Balamayooran G., Batra S., Theivanthiran B., Cai S., Pacher P., Jeyaseelan S. Intrapulmonary G-CSF rescues neutrophil recruitment to the lung and neutrophil release to blood in Gram-negative bacterial infection in MCP-1−/− mice. J. Immunol. 2012;189:5849–5859. doi: 10.4049/jimmunol.1200585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Fukata M., Hernandez Y., Conduah D., Cohen J., Chen A., Breglio K., Goo T., Hsu D., Xu R., Abreu M.T. Innate immune signaling by Toll-like receptor-4 (TLR4) shapes the inflammatory microenvironment in colitis-associated tumors. Inflamm. Bowel Dis. 2009;15:997–1006. doi: 10.1002/ibd.20880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Bánfi B., Clark R.A., Steger K., Krause K.-H. Two novel proteins activate superoxide generation by the NADPH oxidase NOX1. J. Biol. Chem. 2003;278:3510–3513. doi: 10.1074/jbc.C200613200. [DOI] [PubMed] [Google Scholar]
- 30.Miyano K., Sumimoto H. N-Linked glycosylation of the superoxide-producing NADPH oxidase Nox1. Biochem. Biophys. Res. Commun. 2014;443:1060–1065. doi: 10.1016/j.bbrc.2013.12.086. [DOI] [PubMed] [Google Scholar]
- 31.Ueyama T., Geiszt M., Leto T.L. Involvement of Rac1 in activation of multicomponent Nox1-and Nox3-based NADPH oxidases. Mol. Cell. Biol. 2006;26:2160–2174. doi: 10.1128/MCB.26.6.2160-2174.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Miyano K., Ueno N., Takeya R., Sumimoto H. Direct involvement of the small GTPase Rac in activation of the superoxide-producing NADPH oxidase Nox1. J. Biol. Chem. 2006;281:21857–21868. doi: 10.1074/jbc.M513665200. [DOI] [PubMed] [Google Scholar]
- 33.Zhang Y., Murugesan P., Huang K., Cai H. NADPH oxidases and oxidase crosstalk in cardiovascular diseases: Novel therapeutic targets. Nat. Rev. Cardiol. 2020;17:170–194. doi: 10.1038/s41569-019-0260-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sumimoto H., Minakami R., Miyano K. NADPH Oxidases. Springer; Berlin/Heidelberg, Germany: 2019. Soluble regulatory proteins for activation of NOX family NADPH oxidases; pp. 121–137. [DOI] [PubMed] [Google Scholar]
- 35.Kitajima N., Numaga-Tomita T., Watanabe M., Kuroda T., Nishimura A., Miyano K., Yasuda S., Kuwahara K., Sato Y., Ide T. TRPC3 positively regulates reactive oxygen species driving maladaptive cardiac remodeling. Sci. Rep. 2016;6:37001. doi: 10.1038/srep37001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Brandes R.P., Weissmann N., Schröder K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014;76:208–226. doi: 10.1016/j.freeradbiomed.2014.07.046. [DOI] [PubMed] [Google Scholar]
- 37.Rastogi R., Geng X., Li F., Ding Y. NOX Activation by Subunit Interaction and Underlying Mechanisms in Disease. Front. Cell. Neurosci. 2017;10:301. doi: 10.3389/fncel.2016.00301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Pastori D., Pignatelli P., Carnevale R., Violi F. Nox-2 up-regulation and platelet activation: Novel insights. Prostaglandins Other Lipid Mediat. 2015;120:50–55. doi: 10.1016/j.prostaglandins.2015.03.010. [DOI] [PubMed] [Google Scholar]
- 39.Diebold B.A., Bokoch G.M. Molecular basis for Rac2 regulation of phagocyte NADPH oxidase. Nat. Immunol. 2001;2:211–215. doi: 10.1038/85259. [DOI] [PubMed] [Google Scholar]
- 40.O’Neill S., Brault J., Stasia M.-J., Knaus U.G. Genetic disorders coupled to ROS deficiency. Redox Biol. 2015;6:135–156. doi: 10.1016/j.redox.2015.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zana M. Functional Investigation of the Nox4 NADPH Oxidase. Hungarian Academy of Sciences; Budapest, Hungary: 2019. [Google Scholar]
- 42.Chocry M., Leloup L. The NADPH oxidase family and its inhibitors. Antioxid. Redox Signal. 2020;33:332–353. doi: 10.1089/ars.2019.7915. [DOI] [PubMed] [Google Scholar]
- 43.Sun J., Chen J., Li T., Huang P., Li J., Shen M., Gao M., Sun Y., Liang J., Li X. ROS production and mitochondrial dysfunction driven by PU. 1-regulated NOX4-p22phox activation in Aβ-induced retinal pigment epithelial cell injury. Theranostics. 2020;10:11637. doi: 10.7150/thno.48064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Waghela B.N., Vaidya F.U., Agrawal Y., Santra M.K., Mishra V., Pathak C. Molecular insights of NADPH oxidases and its pathological consequences. Cell Biochem. Funct. 2021;39:218–234. doi: 10.1002/cbf.3589. [DOI] [PubMed] [Google Scholar]
- 45.Manickam N., Patel M., Griendling K.K., Gorin Y., Barnes J.L. RhoA/Rho kinase mediates TGF-β1-induced kidney myofibroblast activation through Poldip2/Nox4-derived reactive oxygen species. Am. J. Physiol.-Ren. Physiol. 2014;307:F159–F171. doi: 10.1152/ajprenal.00546.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lyle A.N., Deshpande N.N., Taniyama Y., Seidel-Rogol B., Pounkova L., Du P., Papaharalambus C., Lassegue B., Griendling K.K. Poldip2, a novel regulator of Nox4 and cytoskeletal integrity in vascular smooth muscle cells. Circ. Res. 2009;105:249–259. doi: 10.1161/CIRCRESAHA.109.193722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Bedard K., Krause K.-H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007;87:245–313. doi: 10.1152/physrev.00044.2005. [DOI] [PubMed] [Google Scholar]
- 48.Laddha A.P., Kulkarni Y.A. NADPH oxidase: A membrane-bound enzyme and its inhibitors in diabetic complications. Eur. J. Pharmacol. 2020;881:173206. doi: 10.1016/j.ejphar.2020.173206. [DOI] [PubMed] [Google Scholar]
- 49.Cheng G., Cao Z., Xu X., Van Meir E.G., Lambeth J.D. Homologs of gp91phox: Cloning and tissue expression of Nox3, Nox4, and Nox5. Gene. 2001;269:131–140. doi: 10.1016/S0378-1119(01)00449-8. [DOI] [PubMed] [Google Scholar]
- 50.Wei C.-C., Hay E., Smith D., Lloyd L., Acharya G., Ngo R. Binding of Nox5’s EF-Hand domain to the peptides corresponding to the phosphorylatable region and regulatory inhibitory loop in its dehydrogenase domain. Biophys. Chem. 2020;262:106379. doi: 10.1016/j.bpc.2020.106379. [DOI] [PubMed] [Google Scholar]
- 51.Furmanik M., Chatrou M., Gorp R.v., Akbulut A., Willems B., Schmidt H., Eys G.v., Bochaton-Piallat M.-L., Proudfoot D., Biessen E., et al. Reactive Oxygen-Forming Nox5 Links Vascular Smooth Muscle Cell Phenotypic Switching and Extracellular Vesicle-Mediated Vascular Calcification. Circ. Res. 2020;127:911–927. doi: 10.1161/CIRCRESAHA.119.316159. [DOI] [PubMed] [Google Scholar]
- 52.Fu X., Beer D.G., Behar J., Wands J., Lambeth D., Cao W. cAMP-response element-binding protein mediates acid-induced NADPH oxidase NOX5-S expression in Barrett esophageal adenocarcinoma cells. J. Biol. Chem. 2006;281:20368–20382. doi: 10.1074/jbc.M603353200. [DOI] [PubMed] [Google Scholar]
- 53.Ago T., Kitazono T., Kuroda J., Kumai Y., Kamouchi M., Ooboshi H., Wakisaka M., Kawahara T., Rokutan K., Ibayashi S., et al. NAD(P)H oxidases in rat basilar arterial endothelial cells. Stroke. 2005;36:1040–1046. doi: 10.1161/01.STR.0000163111.05825.0b. [DOI] [PubMed] [Google Scholar]
- 54.Miyano K., Okamoto S., Yamauchi A., Kajikawa M., Kiyohara T., Taura M., Kwai C., Kuribayashi F. Constitutive activity of NADPH oxidase 1 (Nox1) that promotes its own activity suppresses the colon epithelial cell migration. Free. Radic. Res. 2020;54:640–648. doi: 10.1080/10715762.2020.1823383. [DOI] [PubMed] [Google Scholar]
- 55.Waller M.D.W. Master’s Thesis. University of Saskatchewan; Saskatoon, Canada: Oct, 2020. Characterization of NADPH Oxidases in Uterine Smooth Muscle during Pregnancy. [Google Scholar]
- 56.Matsushima S., Sadoshima J. Yin and Yang of NADPH Oxidases in Myocardial Ischemia-Reperfusion. Antioxidants. 2022;11:1069. doi: 10.3390/antiox11061069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Gimenez M., Schickling B.M., Lopes L.R., Miller F.J., Jr. Nox1 in cardiovascular diseases: Regulation and pathophysiology. Clin. Sci. 2016;130:151–165. doi: 10.1042/CS20150404. [DOI] [PubMed] [Google Scholar]
- 58.Konior A., Schramm A., Czesnikiewicz-Guzik M., Guzik T.J. NADPH oxidases in vascular pathology. Antioxid. Redox Signal. 2014;20:2794–2814. doi: 10.1089/ars.2013.5607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Manea A., Manea S.A., Gafencu A.V., Raicu M., Simionescu M. AP-1-dependent transcriptional regulation of NADPH oxidase in human aortic smooth muscle cells: Role of p22phox subunit. Arterioscler. Thromb. Vasc. Biol. 2008;28:878–885. doi: 10.1161/ATVBAHA.108.163592. [DOI] [PubMed] [Google Scholar]
- 60.Starke R.M., Thompson J.W., Ali M.S., Pascale C.L., Martinez Lege A., Ding D., Chalouhi N., Hasan D.M., Jabbour P., Owens G.K. Cigarette smoke initiates oxidative stress-induced cellular phenotypic modulation leading to cerebral aneurysm pathogenesis. Arterioscler. Thromb. Vasc. Biol. 2018;38:610–621. doi: 10.1161/ATVBAHA.117.310478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Heymes C., Bendall Jennifer K., Ratajczak P., Cave Alison C., Samuel J.-L., Hasenfuss G., Shah Ajay M. Increased myocardial NADPH oxidase activity in human heart failure. J. Am. Coll. Cardiol. 2003;41:2164–2171. doi: 10.1016/S0735-1097(03)00471-6. [DOI] [PubMed] [Google Scholar]
- 62.Violi F., Carnevale R., Loffredo L., Pignatelli P., Gallin J.I. NADPH oxidase-2 and atherothrombosis: Insight from chronic granulomatous disease. Arterioscler. Thromb. Vasc. Biol. 2017;37:218–225. doi: 10.1161/ATVBAHA.116.308351. [DOI] [PubMed] [Google Scholar]
- 63.Touyz R.M., Chen X., Tabet F., Yao G., He G., Quinn M.T., Pagano P.J., Schiffrin E.L. Expression of a functionally active gp91phox-containing neutrophil-type NAD (P) H oxidase in smooth muscle cells from human resistance arteries: Regulation by angiotensin II. Circ. Res. 2002;90:1205–1213. doi: 10.1161/01.RES.0000020404.01971.2F. [DOI] [PubMed] [Google Scholar]
- 64.Suliman H.B., Ryan L.K., Bishop L., Folz R.J. Prevention of influenza-induced lung injury in mice overexpressing extracellular superoxide dismutase. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2001;280:L69–L78. doi: 10.1152/ajplung.2001.280.1.L69. [DOI] [PubMed] [Google Scholar]
- 65.Snelgrove R.J., Edwards L., Rae A.J., Hussell T. An absence of reactive oxygen species improves the resolution of lung influenza infection. Eur. J. Immunol. 2006;36:1364–1373. doi: 10.1002/eji.200635977. [DOI] [PubMed] [Google Scholar]
- 66.Hecker L., Vittal R., Jones T., Jagirdar R., Luckhardt T.R., Horowitz J.C., Pennathur S., Martinez F.J., Thannickal V.J. NADPH oxidase-4 mediates myofibroblast activation and fibrogenic responses to lung injury. Nat. Med. 2009;15:1077–1081. doi: 10.1038/nm.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Heyworth P.G., Curnutte J.T., Nauseef W.M., Volpp B.D., Pearson D.W., Rosen H., Clark R.A. Neutrophil nicotinamide adenine dinucleotide phosphate oxidase assembly. Translocation of p47-phox and p67-phox requires interaction between p47-phox and cytochrome b558. J. Clin. Investig. 1991;87:352–356. doi: 10.1172/JCI114993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Nada M., Ohnishi H., Tochio H., Kato Z., Kimura T., Kubota K., Yamamoto T., Kamatari Y.O., Tsutsumi N., Shirakawa M. Molecular analysis of the binding mode of Toll/interleukin-1 receptor (TIR) domain proteins during TLR2 signaling. Mol. Immunol. 2012;52:108–116. doi: 10.1016/j.molimm.2012.05.003. [DOI] [PubMed] [Google Scholar]
- 69.Babior B.M. NADPH oxidase: An update. Blood. 1999;93:1464–1476. doi: 10.1182/blood.V93.5.1464. [DOI] [PubMed] [Google Scholar]
- 70.Hahn N.E., Musters R.J.P., Fritz J.M., Pagano P.J., Vonk A.B.A., Paulus W.J., van Rossum A.C., Meischl C., Niessen H.W.M., Krijnen P.A.J. Early NADPH oxidase-2 activation is crucial in phenylephrine-induced hypertrophy of H9c2 cells. Cell. Signal. 2014;26:1818–1824. doi: 10.1016/j.cellsig.2014.04.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Akaike T., Noguchi Y., Ijiri S., Setoguchi K., Suga M., Zheng Y.M., Dietzschold B., Maeda H. Pathogenesis of influenza virus-induced pneumonia: Involvement of both nitric oxide and oxygen radicals. Proc. Natl. Acad. Sci. USA. 1996;93:2448–2453. doi: 10.1073/pnas.93.6.2448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Guo X., Fan Y., Cui J., Hao B., Zhu L., Sun X., He J., Yang J., Dong J., Wang Y., et al. NOX4 expression and distal arteriolar remodeling correlate with pulmonary hypertension in COPD. BMC Pulm. Med. 2018;18:111. doi: 10.1186/s12890-018-0680-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Kamdar K., Nguyen V., DePaolo R.W. Toll-like receptor signaling and regulation of intestinal immunity. Virulence. 2013;4:207–212. doi: 10.4161/viru.23354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Bureau C., Bernad J., Chaouche N., Orfila C., Béraud M., Gonindard C., Alric L., Vinel J.-P., Pipy B. Nonstructural 3 protein of hepatitis C virus triggers an oxidative burst in human monocytes via activation of NADPH oxidase. J. Biol. Chem. 2001;276:23077–23083. doi: 10.1074/jbc.M100698200. [DOI] [PubMed] [Google Scholar]
- 75.Fink M.P. Intensive Care Medicine. Springer; New York, NY, USA: 2003. Ethyl pyruvate: A novel anti-inflammatory agent; pp. 604–614. [DOI] [PubMed] [Google Scholar]
- 76.Theivanthiran B., Batra S., Balamayooran G., Cai S., Kobayashi K., Flavell R.A., Jeyaseelan S. NOD2 signaling contributes to host defense in the lungs against Escherichia coli infection. Infect. Immun. 2012;80:2558–2569. doi: 10.1128/IAI.06230-11. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 77.García-Redondo A.B., Aguado A., Briones A.M., Salaices M. NADPH oxidases and vascular remodeling in cardiovascular diseases. Pharmacol. Res. 2016;114:110–120. doi: 10.1016/j.phrs.2016.10.015. [DOI] [PubMed] [Google Scholar]
- 78.Borghi C., Omboni S., Reggiardo G., Bacchelli S., Esposti D.D., Ambrosioni E. Effects of the concomitant administration of xanthine oxidase inhibitors with zofenopril or other ACE-inhibitors in post-myocardial infarction patients: A meta-analysis of individual data of four randomized, double-blind, prospective studies. BMC Cardiovasc. Disord. 2018;18:112. doi: 10.1186/s12872-018-0800-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Battelli M.G., Polito L., Bortolotti M., Bolognesi A. Xanthine Oxidoreductase-Derived Reactive Species: Physiological and Pathological Effects. Oxidative Med. Cell. Longev. 2016;2016:3527579. doi: 10.1155/2016/3527579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Altenhöfer S., Kleikers P.W., Radermacher K.A., Scheurer P., Rob Hermans J.J., Schiffers P., Ho H., Wingler K., Schmidt H.H. The NOX toolbox: Validating the role of NADPH oxidases in physiology and disease. Cell. Mol. Life Sci. CMLS. 2012;69:2327–2343. doi: 10.1007/s00018-012-1010-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Lassègue B., Griendling K.K. NADPH oxidases: Functions and pathologies in the vasculature. Arterioscler. Thromb. Vasc. Biol. 2010;30:653–661. doi: 10.1161/ATVBAHA.108.181610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Clempus R.E., Sorescu D., Dikalova A.E., Pounkova L., Jo P., Sorescu G.P., Lassègue B., Griendling K.K. Nox4 is required for maintenance of the differentiated vascular smooth muscle cell phenotype. Arterioscler. Thromb. Vasc. Biol. 2007;27:42–48. doi: 10.1161/01.ATV.0000251500.94478.18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Poznyak A.V., Grechko A.V., Orekhova V.A., Khotina V., Ivanova E.A., Orekhov A.N. NADPH Oxidases and Their Role in Atherosclerosis. Biomedicines. 2020;8:206. doi: 10.3390/biomedicines8070206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Ago T., Kuroda J., Pain J., Fu C., Li H., Sadoshima J. Upregulation of Nox4 by hypertrophic stimuli promotes apoptosis and mitochondrial dysfunction in cardiac myocytes. Circ. Res. 2010;106:1253–1264. doi: 10.1161/CIRCRESAHA.109.213116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Matsushima S., Kuroda J., Ago T., Zhai P., Park J.Y., Xie L.-H., Tian B., Sadoshima J. Increased oxidative stress in the nucleus caused by Nox4 mediates oxidation of HDAC4 and cardiac hypertrophy. Circ. Res. 2013;112:651–663. doi: 10.1161/CIRCRESAHA.112.279760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Burgoyne J.R., Mongue-Din H., Eaton P., Shah A.M. Redox signaling in cardiac physiology and pathology. Circ. Res. 2012;111:1091–1106. doi: 10.1161/CIRCRESAHA.111.255216. [DOI] [PubMed] [Google Scholar]
- 87.Zhao Q.D., Viswanadhapalli S., Williams P., Shi Q., Tan C., Yi X., Bhandari B., Abboud H.E. NADPH oxidase 4 induces cardiac fibrosis and hypertrophy through activating Akt/mTOR and NFκB signaling pathways. Circulation. 2015;131:643–655. doi: 10.1161/CIRCULATIONAHA.114.011079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Zhang Q., Malik P., Pandey D., Gupta S., Jagnandan D., Belin de Chantemele E., Banfi B., Marrero M.B., Rudic R.D., Stepp D.W., et al. Paradoxical activation of endothelial nitric oxide synthase by NADPH oxidase. Arterioscler. Thromb. Vasc. Biol. 2008;28:1627–1633. doi: 10.1161/ATVBAHA.108.168278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Nouel A., Rojano-Rada J., Storino-Farina M.A. Atherosclerosis and inflammatory response: New pathophysiological routes toward a therapeutic role. Rev. Mex. Cardiol. 2016;27:130–137. [Google Scholar]
- 90.Sumimoto H. Structure, regulation and evolution of Nox-family NADPH oxidases that produce reactive oxygen species. FEBS J. 2008;275:3249–3277. doi: 10.1111/j.1742-4658.2008.06488.x. [DOI] [PubMed] [Google Scholar]
- 91.Niu X.L., Madamanchi N.R., Vendrov A.E., Tchivilev I., Rojas M., Madamanchi C., Brandes R.P., Krause K.H., Humphries J., Smith A., et al. Nox activator 1: A potential target for modulation of vascular reactive oxygen species in atherosclerotic arteries. Circulation. 2010;121:549–559. doi: 10.1161/CIRCULATIONAHA.109.908319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Sheehan A.L., Carrell S., Johnson B., Stanic B., Banfi B., Miller F.J., Jr. Role for Nox1 NADPH oxidase in atherosclerosis. Atherosclerosis. 2011;216:321–326. doi: 10.1016/j.atherosclerosis.2011.02.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Douglas G., Bendall J.K., Crabtree M.J., Tatham A.L., Carter E.E., Hale A.B., Channon K.M. Endothelial-specific Nox2 overexpression increases vascular superoxide and macrophage recruitment in ApoE−/− mice. Cardiovasc. Res. 2012;94:20–29. doi: 10.1093/cvr/cvs026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Jackman K.A., Miller A.A., De Silva T.M., Crack P.J., Drummond G.R., Sobey C.G. Reduction of cerebral infarct volume by apocynin requires pretreatment and is absent in Nox2-deficient mice. Br. J. Pharmacol. 2009;156:680–688. doi: 10.1111/j.1476-5381.2008.00073.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Li G., Ye C., Zhu Y., Zhang T., Gu J., Pan J., Wang F., Wu F., Huang K., Xu K. Oxidative Injury in Ischemic Stroke: A Focus on NADPH Oxidase 4. Oxidative Med. Cell. Longev. 2022;2022:1148874. doi: 10.1155/2022/1148874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Rodiño-Janeiro B.K., Paradela-Dobarro B., Castiñeiras-Landeira M.I., Raposeiras-Roubín S., González-Juanatey J.R., Alvarez E. Current status of NADPH oxidase research in cardiovascular pharmacology. Vasc. Health Risk Manag. 2013;9:401–428. doi: 10.2147/vhrm.S33053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Braunersreuther V., Montecucco F., Ashri M., Pelli G., Galan K., Frias M., Burger F., Quinderé A.L.G., Montessuit C., Krause K.-H. Role of NADPH oxidase isoforms NOX1, NOX2 and NOX4 in myocardial ischemia/reperfusion injury. J. Mol. Cell. Cardiol. 2013;64:99–107. doi: 10.1016/j.yjmcc.2013.09.007. [DOI] [PubMed] [Google Scholar]
- 98.Tillhon M., Guamán Ortiz L.M., Lombardi P., Scovassi A.I. Berberine: New perspectives for old remedies. Biochem. Pharmacol. 2012;84:1260–1267. doi: 10.1016/j.bcp.2012.07.018. [DOI] [PubMed] [Google Scholar]
- 99.Singh I.P., Mahajan S. Berberine and its derivatives: A patent review (2009–2012) Expert Opin. Ther. Pat. 2013;23:215–231. doi: 10.1517/13543776.2013.746314. [DOI] [PubMed] [Google Scholar]
- 100.Li X.-Y., Zhao Z.-X., Huang M., Feng R., He C.-Y., Ma C., Luo S.-H., Fu J., Wen B.-Y., Ren L. Effect of Berberine on promoting the excretion of cholesterol in high-fat diet-induced hyperlipidemic hamsters. J. Transl. Med. 2015;13:278. doi: 10.1186/s12967-015-0629-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Zhang X., Zhao Y., Xu J., Xue Z., Zhang M., Pang X., Zhang X., Zhao L. Modulation of gut microbiota by berberine and metformin during the treatment of high-fat diet-induced obesity in rats. Sci. Rep. 2015;5:14405. doi: 10.1038/srep14405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Liu Y.-F., Wen C.-Y.-Z., Chen Z., Wang Y., Huang Y., Tu S.-H. Effects of berberine on NLRP3 and IL-1β expressions in monocytic THP-1 cells with monosodium urate crystals-induced inflammation. BioMed Res. Int. 2016;2016:2503703. doi: 10.1155/2016/2503703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Liang H., Wang Y. Berberine alleviates hepatic lipid accumulation by increasing ABCA1 through the protein kinase C δ pathway. Biochem. Biophys. Res. Commun. 2018;498:473–480. doi: 10.1016/j.bbrc.2018.03.003. [DOI] [PubMed] [Google Scholar]
- 104.Memon M.A., Khan R.N., Riaz S., Ain Q.U., Ahmed M., Kumar N. Methylglyoxal and insulin resistance in berberine-treated type 2 diabetic patients. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2018;23:110. doi: 10.4103/jrms.JRMS_1078_17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Cheng F., Wang Y., Li J., Su C., Wu F., Xia W.H., Yang Z., Yu B.B., Qiu Y.X., Tao J. Berberine improves endothelial function by reducing endothelial microparticles-mediated oxidative stress in humans. Int. J. Cardiol. 2013;167:936–942. doi: 10.1016/j.ijcard.2012.03.090. [DOI] [PubMed] [Google Scholar]
- 106.Zhang M., Wang C.M., Li J., Meng Z.J., Wei S.N., Li J., Bucala R., Li Y.L., Chen L. Berberine protects against palmitate-induced endothelial dysfunction: Involvements of upregulation of AMPK and eNOS and downregulation of NOX4. Mediat. Inflamm. 2013;2013:260464. doi: 10.1155/2013/260464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Zhang L., Li D.C., Liu L.F. Paeonol: Pharmacological effects and mechanisms of action. Int. Immunopharmacol. 2019;72:413–421. doi: 10.1016/j.intimp.2019.04.033. [DOI] [PubMed] [Google Scholar]
- 108.Adki K.M., Kulkarni Y.A. Chemistry, pharmacokinetics, pharmacology and recent novel drug delivery systems of paeonol. Life Sci. 2020;250:117544. doi: 10.1016/j.lfs.2020.117544. [DOI] [PubMed] [Google Scholar]
- 109.Zhai K.-F., Duan H., Luo L., Cao W.-G., Han F.-K., Shan L.-L., Fang X.-M. Protective effects of paeonol on inflammatory response in IL-1β-induced human fibroblast-like synoviocytes and rheumatoid arthritis progression via modulating NF-κB pathway. Inflammopharmacology. 2017;25:523–532. doi: 10.1007/s10787-017-0385-5. [DOI] [PubMed] [Google Scholar]
- 110.Xue P., Wang Y., Zeng F., Xiu R., Chen J., Guo J., Yuan P., Liu L., Xiao J., Lu H. Paeonol suppresses solar ultraviolet-induced skin inflammation by targeting T-LAK cell-originated protein kinase. Oncotarget. 2017;8:27093. doi: 10.18632/oncotarget.15636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Zhang L., Chen Z., Gong W., Zou Y., Xu F., Chen L., Huang H. Paeonol ameliorates diabetic renal fibrosis through promoting the activation of the Nrf2/ARE pathway via up-regulating Sirt1. Front. Pharmacol. 2018;9:512. doi: 10.3389/fphar.2018.00512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Liu J., Feng L., Ma D., Zhang M., Gu J., Wang S., Fu Q., Song Y., Lan Z., Qu R., et al. Neuroprotective effect of paeonol on cognition deficits of diabetic encephalopathy in streptozotocin-induced diabetic rat. Neurosci. Lett. 2013;549:63–68. doi: 10.1016/j.neulet.2013.06.002. [DOI] [PubMed] [Google Scholar]
- 113.Shi X., Chen Y.H., Liu H., Qu H.D. Therapeutic effects of paeonol on methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine/probenecid-induced Parkinson’s disease in mice. Mol. Med. Rep. 2016;14:2397–2404. doi: 10.3892/mmr.2016.5573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Zhou J., Zhou L., Hou D., Tang J., Sun J., Bondy S.C. Paeonol increases levels of cortical cytochrome oxidase and vascular actin and improves behavior in a rat model of Alzheimer’s disease. Brain Res. 2011;1388:141–147. doi: 10.1016/j.brainres.2011.02.064. [DOI] [PubMed] [Google Scholar]
- 115.Choy K.W., Lau Y.S., Murugan D., Mustafa M.R. Chronic treatment with paeonol improves endothelial function in mice through inhibition of endoplasmic reticulum stress-mediated oxidative stress. PLoS ONE. 2017;12:e0178365. doi: 10.1371/journal.pone.0178365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Al-Taher A.Y., Morsy M.A., Rifaai R.A., Zenhom N.M., Abdel-Gaber S.A. Paeonol Attenuates Methotrexate-Induced Cardiac Toxicity in Rats by Inhibiting Oxidative Stress and Suppressing TLR4-Induced NF-κB Inflammatory Pathway. Mediat. Inflamm. 2020;2020:8641026. doi: 10.1155/2020/8641026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Ahmad A., Mishra R.K., Vyawahare A., Kumar A., Rehman M.U., Qamar W., Khan A.Q., Khan R. Thymoquinone (2-Isoprpyl-5-methyl-1, 4-benzoquinone) as a chemopreventive/anticancer agent: Chemistry and biological effects. Saudi Pharm. J. SPJ Off. Publ. Saudi Pharm. Soc. 2019;27:1113–1126. doi: 10.1016/j.jsps.2019.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Khader M., Eckl P.M. Thymoquinone: An emerging natural drug with a wide range of medical applications. Iran. J. Basic Med. Sci. 2014;17:950–957. [PMC free article] [PubMed] [Google Scholar]
- 119.Mansour M.A., Nagi M.N., El-Khatib A.S., Al-Bekairi A.M. Effects of thymoquinone on antioxidant enzyme activities, lipid peroxidation and DT-diaphorase in different tissues of mice: A possible mechanism of action. Cell Biochem. Funct. 2002;20:143–151. doi: 10.1002/cbf.968. [DOI] [PubMed] [Google Scholar]
- 120.Badary O.A., Hamza M.S., Tikamdas R. Thymoquinone: A Promising Natural Compound with Potential Benefits for COVID-19 Prevention and Cure. Drug Des. Dev. Ther. 2021;15:1819–1833. doi: 10.2147/DDDT.S308863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Allemailem K.S., Almatroudi A., Alrumaihi F., Aljaghwani A., Alnuqaydan A.M., Khalilullah H., Younus H., El-Kady A.M., Aldakheel F.M., Khan A.A. Antimicrobial, Immunomodulatory and Anti-Inflammatory Potential of Liposomal Thymoquinone: Implications in the Treatment of Bacterial Pneumonia in Immunocompromised Mice. Biomedicines. 2021;9:1673. doi: 10.3390/biomedicines9111673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.El-Far A.H., Salaheldin T.A., Godugu K., Darwish N.H., Mousa S.A. Thymoquinone and its nanoformulation attenuate colorectal and breast cancers and alleviate doxorubicin-induced cardiotoxicity. Nanomedicine. 2021;16:1457–1469. doi: 10.2217/nnm-2021-0103. [DOI] [PubMed] [Google Scholar]
- 123.Chaieb K., Kouidhi B., Jrah H., Mahdouani K., Bakhrouf A. Antibacterial activity of Thymoquinone, an active principle of Nigella sativa and its potency to prevent bacterial biofilm formation. BMC Complement. Altern. Med. 2011;11:29. doi: 10.1186/1472-6882-11-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Imran M., Rauf A., Khan I.A., Shahbaz M., Qaisrani T.B., Fatmawati S., Abu-Izneid T., Imran A., Rahman K.U., Gondal T.A. Thymoquinone: A novel strategy to combat cancer: A review. Biomed. Pharmacother. 2018;106:390–402. doi: 10.1016/j.biopha.2018.06.159. [DOI] [PubMed] [Google Scholar]
- 125.Zhang L., Zhang H., Ma J., Wang Y., Pei Z., Ding H. Effects of thymoquinone against angiotensin II-induced cardiac damage in apolipoprotein E-deficient mice. Int. J. Mol. Med. 2022;49:1–12. doi: 10.3892/ijmm.2022.5119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Guo W., Long X., Lv M., Deng S., Liu D., Yang Q. Effect of thymoquinone on sepsis-induced cardiac damage via anti-inflammatory and anti-apoptotic mechanisms. J. Int. Med. Res. 2022;50:3000605221118680. doi: 10.1177/03000605221118680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Chen H., Zhuo C., Zu A., Yuan S., Zhang H., Zhao J., Zheng L. Thymoquinone ameliorates pressure overload-induced cardiac hypertrophy by activating the AMPK signalling pathway. J. Cell. Mol. Med. 2022;26:855–867. doi: 10.1111/jcmm.17138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Chao S., Xu Q., Dong S., Guo M., Liu X., Cheng X. Polygala fallax Hemsl combined with compound Sanqi granules relieves glomerulonephritis by regulating proliferation and apoptosis of glomerular mesangial cells. J. Int. Med. Res. 2020;48:0300060519894124. doi: 10.1177/0300060519894124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Yao Z., Li Y., Wang Z., Lan Y., Zeng T., Gong H., Zhu K., Tang H., Gu S. Research on anti-hepatocellular carcinoma activity and mechanism of Polygala fallax Hemsl. J. Ethnopharmacol. 2020;260:113062. doi: 10.1016/j.jep.2020.113062. [DOI] [PubMed] [Google Scholar]
- 130.Singh D., Chaudhuri P.K. Structural characteristics, bioavailability and cardioprotective potential of saponins. Integr. Med. Res. 2018;7:33–43. doi: 10.1016/j.imr.2018.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Hong D., Bai Y.P., Shi R.Z., Tan G.S., Hu C.P., Zhang G.G. Inhibitory effect of reinioside C on vascular smooth muscle cells proliferation induced by angiotensin II via inhibiting NADPH oxidase-ROS-ENK1/2-NF-kappaB-AP-1 pathway. Die Pharm. 2014;69:698–703. doi: 10.1136/heartjnl-2012-302920b.26. [DOI] [PubMed] [Google Scholar]
- 132.Bai Y.P., Hu C.P., Chen M.F., Xu K.P., Tan G.S., Shi R.Z., Li Y.J., Zhang G.G. Inhibitory effect of reinioside C on monocyte-endothelial cell adhesion induced by oxidized low-density lipoprotein via inhibiting NADPH oxidase/ROS/NF-kappaB pathway. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2009;380:399–406. doi: 10.1007/s00210-009-0450-8. [DOI] [PubMed] [Google Scholar]
- 133.Kim K.M., Pae H.-O., Zhung M., Ha H.-Y., Ha Y.A., Chai K.-Y., Cheong Y.-K., Kim J.-M., Chung H.-T. Involvement of anti-inflammatory heme oxygenase-1 in the inhibitory effect of curcumin on the expression of pro-inflammatory inducible nitric oxide synthase in RAW264.7 macrophages. Biomed. Pharmacother. 2008;62:630–636. doi: 10.1016/j.biopha.2008.01.008. [DOI] [PubMed] [Google Scholar]
- 134.Bar-Sela G., Epelbaum R., Schaffer M. Curcumin as an anti-cancer agent: Review of the gap between basic and clinical applications. Curr. Med. Chem. 2010;17:190–197. doi: 10.2174/092986710790149738. [DOI] [PubMed] [Google Scholar]
- 135.Kumar S., Singh N.N., Singh A., Singh N., Sinha R.K. Use of Curcuma longa L. extract to stain various tissue samples for histological studies. Ayu. 2014;35:447–451. doi: 10.4103/0974-8520.159027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Kim K.S., Kim N.Y., Son J.Y., Park J.H., Lee S.H., Kim H.R., Kim B., Kim Y.G., Jeong H.G., Lee B.M. Curcumin ameliorates benzo [a] pyrene-induced DNA damages in stomach tissues of Sprague-Dawley rats. Int. J. Mol. Sci. 2019;20:5533. doi: 10.3390/ijms20225533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Imam Z., Khasawneh M., Jomaa D., Iftikhar H., Sayedahmad Z. Drug induced liver injury attributed to a curcumin supplement. Case Rep. Gastrointest. Med. 2019;2019:6029403. doi: 10.1155/2019/6029403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Prasad S., Aggarwal B.B. Turmeric, the Golden Spice: From Traditional Medicine to Modern Medicine. In: Benzie I.F.F., Wachtel-Galor S., editors. Herbal Medicine: Biomolecular and Clinical Aspects. CRC Press/Taylor & Francis; Boca Raton, FL, USA: 2011. [PubMed] [Google Scholar]
- 139.Boonla O., Kukongviriyapan U., Pakdeechote P., Kukongviriyapan V., Pannangpetch P., Prachaney P., Greenwald S.E. Curcumin improves endothelial dysfunction and vascular remodeling in 2K-1C hypertensive rats by raising nitric oxide availability and reducing oxidative stress. Nitric Oxide Biol. Chem. 2014;42:44–53. doi: 10.1016/j.niox.2014.09.001. [DOI] [PubMed] [Google Scholar]
- 140.Meng Z., Yan C., Deng Q., Gao D.F., Niu X.L. Curcumin inhibits LPS-induced inflammation in rat vascular smooth muscle cells in vitro via ROS-relative TLR4-MAPK/NF-κB pathways. Acta Pharmacol. Sin. 2013;34:901–911. doi: 10.1038/aps.2013.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Hu H., Straub A., Tian Z., Bassler N., Cheng J., Peter K. Celastrol, a triterpene extracted from Tripterygium wilfordii Hook F, inhibits platelet activation. J. Cardiovasc. Pharmacol. 2009;54:240–245. doi: 10.1097/FJC.0b013e3181b21472. [DOI] [PubMed] [Google Scholar]
- 142.Venkatesha S.H., Moudgil K.D. Celastrol and Its Role in Controlling Chronic Diseases. Adv. Exp. Med. Biol. 2016;928:267–289. doi: 10.1007/978-3-319-41334-1_12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Allison A.C., Cacabelos R., Lombardi V.R., Alvarez X.A., Vigo C. Celastrol, a potent antioxidant and anti-inflammatory drug, as a possible treatment for Alzheimer’s disease. Prog. Neuro-Psychopharmacol. Biol. Psychiatry. 2001;25:1341–1357. doi: 10.1016/S0278-5846(01)00192-0. [DOI] [PubMed] [Google Scholar]
- 144.Tao X., Younger J., Fan F.Z., Wang B., Lipsky P.E. Benefit of an extract of Tripterygium Wilfordii Hook F in patients with rheumatoid arthritis: A double-blind, placebo-controlled study. Arthritis Rheum. 2002;46:1735–1743. doi: 10.1002/art.10411. [DOI] [PubMed] [Google Scholar]
- 145.Divya T., Dineshbabu V., Soumyakrishnan S., Sureshkumar A., Sudhandiran G. Celastrol enhances Nrf2 mediated antioxidant enzymes and exhibits anti-fibrotic effect through regulation of collagen production against bleomycin-induced pulmonary fibrosis. Chem.-Biol. Interact. 2016;246:52–62. doi: 10.1016/j.cbi.2016.01.006. [DOI] [PubMed] [Google Scholar]
- 146.Seo W.Y., Goh A.R., Ju S.M., Song H.Y., Kwon D.J., Jun J.G., Kim B.C., Choi S.Y., Park J. Celastrol induces expression of heme oxygenase-1 through ROS/Nrf2/ARE signaling in the HaCaT cells. Biochem. Biophys. Res. Commun. 2011;407:535–540. doi: 10.1016/j.bbrc.2011.03.053. [DOI] [PubMed] [Google Scholar]
- 147.Li M., Liu X., He Y., Zheng Q., Wang M., Wu Y., Zhang Y., Wang C. Celastrol attenuates angiotensin II mediated human umbilical vein endothelial cells damage through activation of Nrf2/ERK1/2/Nox2 signal pathway. Eur. J. Pharmacol. 2017;797:124–133. doi: 10.1016/j.ejphar.2017.01.027. [DOI] [PubMed] [Google Scholar]
- 148.Liu H., Wang L., Pan Y., Wang X., Ding Y., Zhou C., Shah A.M., Zhao G., Zhang M. Celastrol Alleviates Aortic Valve Calcification Via Inhibition of NADPH Oxidase 2 in Valvular Interstitial Cells. JACC Basic Transl. Sci. 2020;5:35–49. doi: 10.1016/j.jacbts.2019.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Luchtefeld R., Luo R., Stine K., Alt M.L., Chernovitz P.A., Smith R.E. Dose formulation and analysis of diapocynin. J. Agric. Food Chem. 2008;56:301–306. doi: 10.1021/jf072792n. [DOI] [PubMed] [Google Scholar]
- 150.Boshtam M., Kouhpayeh S., Amini F., Azizi Y., Najaflu M., Shariati L., Khanahmad H. Anti-inflammatory effects of apocynin: A narrative review of the evidence. All Life. 2021;14:997–1010. doi: 10.1080/26895293.2021.1990136. [DOI] [Google Scholar]
- 151.Hwang Y.-J., Nam S.-J., Chun W., Kim S.I., Park S.C., Kang C.D., Lee S.J. Anti-inflammatory effects of apocynin on dextran sulfate sodium-induced mouse colitis model. PLoS ONE. 2019;14:e0217642. doi: 10.1371/journal.pone.0217642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Chen H., He Y., Chen S., Qi S., Shen J. Therapeutic targets of oxidative/nitrosative stress and neuroinflammation in ischemic stroke: Applications for natural product efficacy with omics and systemic biology. Pharmacol. Res. 2020;158:104877. doi: 10.1016/j.phrs.2020.104877. [DOI] [PubMed] [Google Scholar]
- 153.Jin H.-Z., Yang X.-J., Zhao K.-L., Mei F.-C., Zhou Y., You Y.-D., Wang W.-X. Apocynin alleviates lung injury by suppressing NLRP3 inflammasome activation and NF-κB signaling in acute pancreatitis. Int. Immunopharmacol. 2019;75:105821. doi: 10.1016/j.intimp.2019.105821. [DOI] [PubMed] [Google Scholar]
- 154.Xin R., Sun X., Wang Z., Yuan W., Jiang W., Wang L., Xiang Y., Zhang H., Li X., Hou Y. Apocynin inhibited NLRP3/XIAP signalling to alleviate renal fibrotic injury in rat diabetic nephropathy. Biomed. Pharmacother. 2018;106:1325–1331. doi: 10.1016/j.biopha.2018.07.036. [DOI] [PubMed] [Google Scholar]
- 155.Yang T., Zang D.-W., Shan W., Guo A.-C., Wu J.-P., Wang Y.-J., Wang Q. Synthesis and evaluations of novel apocynin derivatives as anti-glioma agents. Front. Pharmacol. 2019;10:951. doi: 10.3389/fphar.2019.00951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Perassa L.A., Graton M.E., Potje S.R., Troiano J.A., Lima M.S., Vale G.T., Pereira A.A., Nakamune A.C., Sumida D.H., Tirapelli C.R., et al. Apocynin reduces blood pressure and restores the proper function of vascular endothelium in SHR. Vasc. Pharmacol. 2016;87:38–48. doi: 10.1016/j.vph.2016.06.005. [DOI] [PubMed] [Google Scholar]
- 157.Fai Y.M., Tao C.C. A review of presence of oleanolic acid in natural products. Nat. Proda Med. 2009;2:77–290. [Google Scholar]
- 158.Markhali F.S., Teixeira J.A., Rocha C.M. Olive tree leaves—A source of valuable active compounds. Processes. 2020;8:1177. doi: 10.3390/pr8091177. [DOI] [Google Scholar]
- 159.Zhao H., Zhou M., Duan L., Wang W., Zhang J., Wang D., Liang X. Efficient synthesis and anti-fungal activity of oleanolic acid oxime esters. Molecules. 2013;18:3615–3629. doi: 10.3390/molecules18033615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Wang X., Ye X.-L., Liu R., Chen H.-L., Bai H., Liang X., Zhang X.-D., Wang Z., Li W.-L., Hai C.-X. Antioxidant activities of oleanolic acid in vitro: Possible role of Nrf2 and MAP kinases. Chem.-Biol. Interact. 2010;184:328–337. doi: 10.1016/j.cbi.2010.01.034. [DOI] [PubMed] [Google Scholar]
- 161.Jesus J.A., Lago J.H.G., Laurenti M.D., Yamamoto E.S., Passero L.F.D. Antimicrobial activity of oleanolic and ursolic acids: An update. Evid.-Based Complement. Altern. Med. 2015;2015:620472. doi: 10.1155/2015/620472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Wang X., Liu R., Zhang W., Zhang X., Liao N., Wang Z., Li W., Qin X., Hai C. Oleanolic acid improves hepatic insulin resistance via antioxidant, hypolipidemic and anti-inflammatory effects. Mol. Cell. Endocrinol. 2013;376:70–80. doi: 10.1016/j.mce.2013.06.014. [DOI] [PubMed] [Google Scholar]
- 163.Jiang Q., Wang D., Han Y., Han Z., Zhong W., Wang C. Modulation of oxidized-LDL receptor-1 (LOX1) contributes to the antiatherosclerosis effect of oleanolic acid. Int. J. Biochem. Cell Biol. 2015;69:142–152. doi: 10.1016/j.biocel.2015.10.023. [DOI] [PubMed] [Google Scholar]
- 164.García-Pérez P., Gallego P.P. Lipid Oxidation in Food and Biological Systems. Springer; Cham, Switzerland: 2022. Plant phenolics as dietary antioxidants: Insights on their biosynthesis, sources, health-promoting effects, sustainable production, and effects on lipid oxidation; pp. 405–426. [Google Scholar]
- 165.Patel R.V., Mistry B.M., Shinde S.K., Syed R., Singh V., Shin H.S. Therapeutic potential of quercetin as a cardiovascular agent. Eur. J. Med. Chem. 2018;155:889–904. doi: 10.1016/j.ejmech.2018.06.053. [DOI] [PubMed] [Google Scholar]
- 166.Ulusoy H.G., Sanlier N. A minireview of quercetin: From its metabolism to possible mechanisms of its biological activities. Crit. Rev. Food Sci. Nutr. 2020;60:3290–3303. doi: 10.1080/10408398.2019.1683810. [DOI] [PubMed] [Google Scholar]
- 167.Ansari P., Choudhury S.T., Seidel V., Rahman A.B., Aziz M.A., Richi A.E., Rahman A., Jafrin U.H., Hannan J., Abdel-Wahab Y.H. Therapeutic potential of quercetin in the management of type-2 diabetes mellitus. Life. 2022;12:1146. doi: 10.3390/life12081146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Asgharian P., Tazekand A.P., Hosseini K., Forouhandeh H., Ghasemnejad T., Ranjbar M., Hasan M., Kumar M., Beirami S.M., Tarhriz V., et al. Potential mechanisms of quercetin in cancer prevention: Focus on cellular and molecular targets. Cancer Cell Int. 2022;22:257. doi: 10.1186/s12935-022-02677-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Carrasco-Pozo C., Cires M.J., Gotteland M. Quercetin and epigallocatechin gallate in the prevention and treatment of obesity: From molecular to clinical studies. J. Med. Food. 2019;22:753–770. doi: 10.1089/jmf.2018.0193. [DOI] [PubMed] [Google Scholar]
- 170.Galindo P., González-Manzano S., Zarzuelo M.J., Gómez-Guzmán M., Quintela A.M., González-Paramás A., Santos-Buelga C., Pérez-Vizcaíno F., Duarte J., Jiménez R. Different cardiovascular protective effects of quercetin administered orally or intraperitoneally in spontaneously hypertensive rats. Food Funct. 2012;3:643–650. doi: 10.1039/c2fo10268d. [DOI] [PubMed] [Google Scholar]
- 171.Wan L.L., Xia J., Ye D., Liu J., Chen J., Wang G. Effects of quercetin on gene and protein expression of NOX and NOS after myocardial ischemia and reperfusion in rabbit. Cardiovasc. Ther. 2009;27:28–33. doi: 10.1111/j.1755-5922.2009.00071.x. [DOI] [PubMed] [Google Scholar]
- 172.Jin X., Yi L., Chen M.-L., Chen C.-Y., Chang H., Zhang T., Wang L., Zhu J.-D., Zhang Q.-Y., Mi M.-T. Delphinidin-3-glucoside protects against oxidized low-density lipoprotein-induced mitochondrial dysfunction in vascular endothelial cells via the sodium-dependent glucose transporter SGLT1. PLoS ONE. 2013;8:e68617. doi: 10.1371/journal.pone.0068617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Noda Y., Kneyuki T., Igarashi K., Mori A., Packer L. Antioxidant activity of nasunin, an anthocyanin in eggplant peels. Toxicology. 2000;148:119–123. doi: 10.1016/S0300-483X(00)00202-X. [DOI] [PubMed] [Google Scholar]
- 174.Sari D.R.T., Cairns J.R.K., Safitri A., Fatchiyah F. Virtual Prediction of the Delphinidin-3-O-glucoside and Peonidin-3-O-glucoside as Anti-inflammatory of TNF-α Signaling. Acta Inform. Med. 2019;27:152. doi: 10.5455/aim.2019.27.152-157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Nas J.S.B., Roxas C.K.F., Acero R.R.G., Gamit A.L.P., Kim J.P., Rentutar J.A., Ching A.C., Saludares A.Q. Solanum melongena (eggplant) crude anthocyanin extract and delphinidin-3-glucoside protects Caenorhabditis elegans against Staphylococcus aureus and Klebsiella pneumoniae. Philipp. J. Health Res. Dev. 2019;23:17–24. [Google Scholar]
- 176.Park M., Sharma A., Lee H.-J. Anti-adipogenic effects of delphinidin-3-O-β-glucoside in 3T3-L1 preadipocytes and primary white adipocytes. Molecules. 2019;24:1848. doi: 10.3390/molecules24101848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Sharma A., Choi H.-K., Kim Y.-K., Lee H.-J. Delphinidin and Its Glycosides’ War on Cancer: Preclinical Perspectives. Int. J. Mol. Sci. 2021;22:11500. doi: 10.3390/ijms222111500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Sasaki K., Geribaldi-Doldan N., Szele F.G., Isoda H. Grape skin extract modulates neuronal stem cell proliferation and improves spatial learning in senescence-accelerated prone 8 mice. Aging. 2021;13:18131. doi: 10.18632/aging.203373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Xie X., Zhao R., Shen G.X. Influence of delphinidin-3-glucoside on oxidized low-density lipoprotein-induced oxidative stress and apoptosis in cultured endothelial cells. J. Agric. Food Chem. 2012;60:1850–1856. doi: 10.1021/jf204461z. [DOI] [PubMed] [Google Scholar]
- 180.Chen Y., Ge Z., Huang S., Zhou L., Zhai C., Chen Y., Hu Q., Cao W., Weng Y., Li Y. Delphinidin attenuates pathological cardiac hypertrophy via the AMPK/NOX/MAPK signaling pathway. Aging. 2020;12:5362–5383. doi: 10.18632/aging.102956. [DOI] [PMC free article] [PubMed] [Google Scholar]