

Abstract
Ovarian cancer (OC) is the most lethal gynecological malignancy due to tumor heterogeneity, the lack of reliable early diagnosis methods and the high incidence of chemoresistant recurrent disease. Although there are developments in chemotherapies and surgical techniques to improve the overall survival of OC patients, the 5-year survival of advanced OC patients is still low. To improve the prognosis of OC patients, it is important to search for novel therapeutic approaches. Cancer stem cells (CSCs) are a subpopulation of tumor cells that participate in tumor growth, metastasis and chemoresistance. It is important to study the role of CSCs in a highly heterogeneous disease such as OC, which may be significant to a better understanding of the oncogenetic and metastatic pathways of the disease and to develop novel strategies against its progression and platinum resistance. Here, we summarized the current findings about targeting methods against ovarian cancer stem cells, including related signaling pathways, markers and drugs, to better manage OC patients using CSC-based therapeutic strategies.
Keywords: Ovarian cancer, Cancer stem cells, Therapy
Introduction
Ovarian cancer (OC) is the fifth most common female cancer and the leading cause of mortality among gynecological tumors. In 2020, there will be approximately 221,750 new cases of OC diagnosed and 13,940 ovarian cancer deaths in the USA [1]. OC is a heterogeneous group of malignancies caused by multiple etiologies. Epithelial ovarian cancers (EOC) are accounting for 90% of OC. According to the new criteria by The World Health Organization (WHO) in 2014, EOC are divided to several morphological categories depending on cell type: high-grade serous (HGSC), low-grade serous (LGSC), mucinous carcinomas (MC), endometrioid carcinomas (EC), and clear-cell carcinomas (CCC) [2]. Except for WHO classification, there are 8 dichotomies for EOC: high grade versus low grade, ovarian versus extraovarian primary, extrauterine versus uterine primary, sporadic versus hereditary, orthodox versus alternative histology, p53 overexpression versus complete absence of immunophenotype, TP53-mutated versus intact precursor, and therapy responsive versus refractory. A better understanding of different dichotomies and molecular classifications is significant for the individual diagnosis and therapy for EOC [3].
Despite intensive investigation, the pathogenesis of OC is not completely understood. HGSC arises from the ovarian surface epithelium or from the fallopian tube and LGSC arises from fallopian tube. Endometrioid carcinoma and clear cell carcinoma arise from endometriosis. Mucinous carcinoma arises from germ cells [4].
The high mortality rate is partly because many cases are diagnosed in advanced stages. In the early stage of OC, there are no obvious symptoms or efficient screening means; thus, early diagnosis is difficult. Over the past decades, the standard treatment for OC has been the combination of radical surgery and platinum-based chemotherapy [5].Although OC patients often initially respond to platinum/paclitaxel-based chemotherapy, most of them still experience relapses and eventually develop chemoresistance [6]. There are several different inhibitors targeting different cancer signal pathways, such as tyrosine kinase inhibitors and monoclonal antibodies, which exert antitumor role in affecting angiogenesis, cell growth and metastasis. Among these emerging inhibitors, the most promising one is poly-ADP-ribose polymerase inhibitors (PARPi), with greater benefit observed in patients with BRCA mutated or BRCA wild-type homologous recombination-deficient (HRD) tumors. According to the recent studies, there are new therapy strategies in HRD tumors, like combined PARPi with antiangiogenic agents, retreated with PARPis after previous PARPi treatment—“PARPi after PARPi” [7, 8]. The therapeutic effects of PARPi are limited, as treatment benefits only show on extending survival by a few months but not long-term survival [9]. Except for these, immunotherapy has also rapidly developed. Although treatment with immune checkpoint inhibitors, chimeric antigen receptor engineered T cells is rapidly developing, immunotherapy response rates among OC patients remain modest [10]. A number of researches discussed other novel therapeutic methods, such as agents targeting metabolism and biosynthetic pathways in OC cells, antibody drug conjugates targeting OC surface molecules and some new compounds whose mechanisms are not yet fully understood. Several studies focused on the gene mutations, like missense mutations in p53 and mutations in ARID1A [11]. In spite of multiple therapy, most of OC patients eventually relapse and the 5-year survival rate for OC is less than 35% [12].
There are three main models to explain histological and molecular heterogeneity for most solid tumors: the clonal evolution or stochastic model, the cancer stem cells (CSCs) or hierarchical model and plasticity model linking the two models above [13]. CSCs are a minor population of tumor cells with self-renewal, pluripotency and limitless proliferative properties [14]. The properties of CSCs are shown in Fig. 1. The CSC model suggests that regardless of the cell origins, many cancers have the same hierarchical organization as normal tissues and CSCs have similar molecular properties as normal stem cells. This model also indicates that the same CSC populations can originate from different cancer subtypes and the frequency of CSCs vary greatly, resulting in tumor heterogeneity [13]. CSCs have been identified in hematologic and solid cancers [15–19]. Experimental evidence supporting the existence of ovarian CSCs was reported by Bapat in 2005 [20]. They cultured a single clone from ascites capable of tumorigenesis and tumor differentiation. CSCs express distinct cell surface markers, such as CD24, CD44, and CD133 [21–23]. CSCs secrete proinflammatory cytokines and chemokines such as IL-4, IL-6, and IL8, contributing to self-renewal ability [24, 25]. However, the mechanism of how ovarian CSCs generate and how cell surface markers function are not known.
Fig. 1.
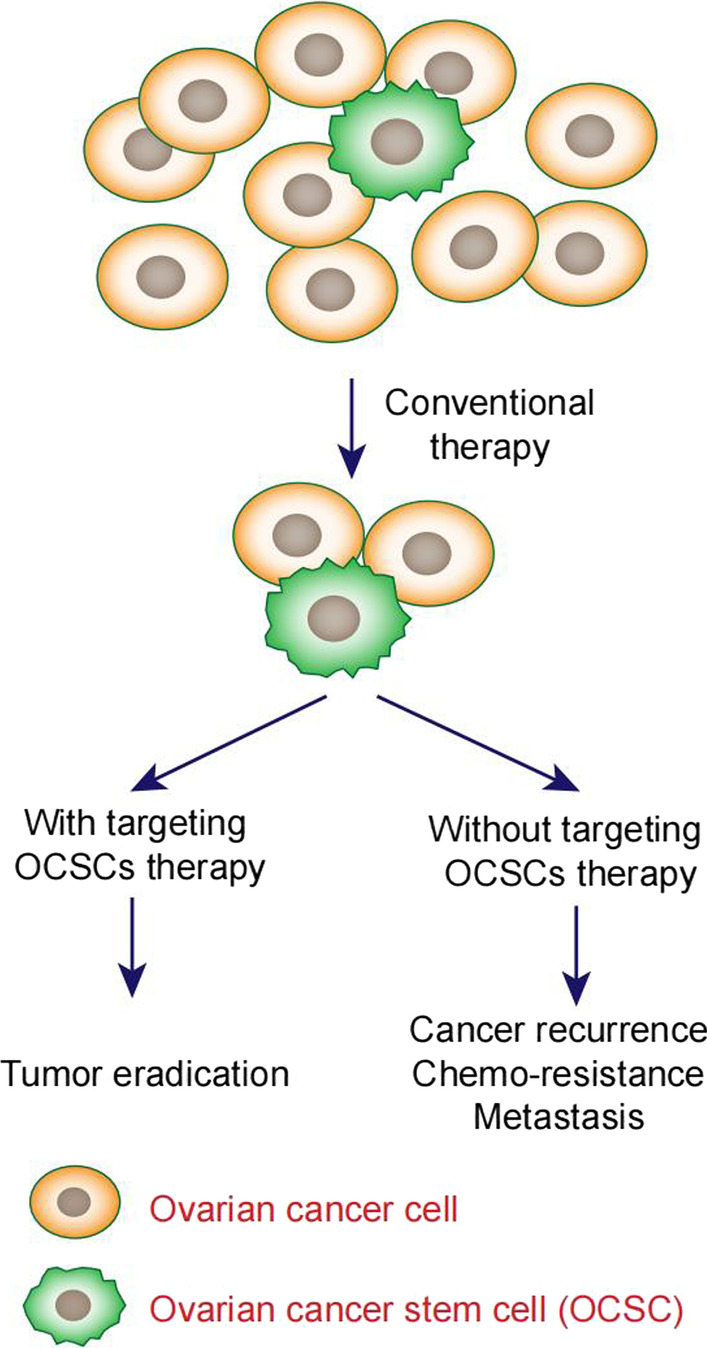
The properties of CSCs
Previous studies reported that CSCs play an important role not only in tumorigenesis but also in tumor metastasis and tumor relapses due to resistance to chemotherapy and radiotherapy [26, 27]. Therefore, targeting ovarian CSCs may offer a novel therapeutic strategy to overcome the present dilemma. The elimination of ovarian CSCs is challenging partly due to heterogeneity, resulting in limitations in therapeutic efficacy. Herein, we provide a review of primary research on targeted therapy for ovarian cancer stem cells (OCSCs).
Methods
The search strategy followed the methodological framework developed by Arksey and O’Malley, which was further clarified by Levac et al. and Joanna Briggs Institute [28–30]. Our review was conducted in five broad stages, each of which is outlined below.
Identifying the research questions
The central research question was “What is known from the existing literature about targeted therapy toward ovarian cancer stem cells?” As of this writing, multiple reviews have been published. We aim to gain a further understanding of all existing studies on ovarian CSCs through an analysis of published literature.
-
2.
Identifying and selecting relevant studies
Final terms were determined after an initial broad search using PubMed, which was used to identify MeSH headings and free words used in relevant papers. We searched four electronic databases (PubMed, Web of Science, ScienceDirect and Embase) with date limitations between 2000 and 2022 for relevant articles. Both MeSH terms and free words texts were used. Table 1 provides the inclusion criteria and PubMed search strategy. Equivalent search strings were used in the other databases.
Table 1.
Inclusion criteria and search strategy
| Inclusion criteria | • Written in English |
| • Articles that included study of both OC and CSCs in therapeutic aspect | |
| • Study date 2000 to 2022 | |
| Exclusion criteria | Review articles, case reports, conference reports, letters, editorial comments, opinions and non-English articles; studies of benign diseases, studies referring only to CSCs |
| Search strategy in PubMed | (("Ovarian Neoplasms" [Mesh]) OR ((((((((((((((((Neoplasm, Ovarian [Title/Abstract]) OR (Ovarian Neoplasm [Title/Abstract])) OR (Ovary Neoplasms [Title/Abstract])) OR (Neoplasm, Ovary [Title/Abstract])) OR (Neoplasms, Ovary [Title/Abstract])) OR (Ovary Neoplasm [Title/Abstract])) OR (Neoplasms, Ovarian [Title/Abstract])) OR (Ovary Cancer [Title/Abstract])) OR (Cancer, Ovary [Title/Abstract])) OR (Cancers, Ovary [Title/Abstract])) OR (Ovarian Cancer [Title/Abstract])) OR (Cancer, Ovarian [Title/Abstract])) OR (Cancers, Ovarian [Title/Abstract])) OR (Ovarian Cancers [Title/Abstract])) OR (Cancer of Ovary [Title/Abstract])) OR (Cancer of the Ovary [Title/Abstract]))) AND (("Stem Cells" [Mesh]) OR (((((((((((((((Cell, Stem [Title/Abstract]) OR (Cells, Stem [Title/Abstract])) OR (Stem Cell [Title/Abstract])) OR (Progenitor Cells [Title/Abstract])) OR (Cell, Progenitor [Title/Abstract])) OR (Cells, Progenitor [Title/Abstract])) OR (Progenitor Cell [Title/Abstract])) OR (Mother Cells [Title/Abstract])) OR (Cell, Mother [Title/Abstract])) OR (Cells, Mother [Title/Abstract])) OR (Mother Cell [Title/Abstract])) OR (Colony-Forming Unit [Title/Abstract])) OR (Colony Forming Unit [Title/Abstract])) OR (Colony-Forming Units [Title/Abstract])) OR (Colony Forming Units [Title/Abstract]))) |
-
3.
Charting the data
We recorded the following data from the selected studies on a data extraction sheet: authors, year of publication, aims of the study, study type, study design and study outcomes.
-
4.
Collating, summarizing and reporting the results
The selected articles were analyzed to address the research question. The data were collated into a summary of the study outcomes reported in narrative form.
Results and discussion
Overview of included studies
Initially, 15,717 scientific articles were found. After duplicate removal, 13,616 articles remained. A total of 13,160 articles were excluded after evaluation of the title and abstract. Two hundred forty further studies were excluded following full text review. As a result, 112 articles were included in the present review. Figure 2 shows the process of selecting studies. In Table 2, the relevant findings from some of included studies are summarized. In Table 3, the therapies targeting OCSCs showed in this review are summarized.
Fig. 2.

PRISMA flow diagram for review data evaluation
Table 2.
Characteristics and major findings of included studies
| Authors | Years | Targeting point or drugs | Major findings |
|---|---|---|---|
| Li [35] | 2011 | Notch pathway | Inhibition pf Notch signaling with the γ-secretase inhibitor DAPT inhibits OCSCs from self-renewing and proliferating |
| Shannon [32] | 2012 | Notch pathway | Notch3 siRNA knockdown depletes OCSCs and increases tumor sensitivity to platinum |
| Sham [36] | 2014 | Notch pathway | Withaferin A used alone or in combination with cisplatin targets putative OCSCs |
| Hyeok [37] | 2016 | Notch pathway | Galectin-3 maintains OCSCs by activating the Notch1 intracellular domain |
| Eun [129] | 2016 | Notch pathway | Hypoxia-Notch-SOX2 signaling axis is vital for activation of OCSCs |
| Zhu [130] | 2019 | Notch 1-c-Myc pathway | SNORD89 regulates Notch 1-c-Myc pathway to promote cell stemness and acts as an oncogene in ovarian tumorigenesis |
| Zhang [131] | 2020 | Notch 1-c-Myc pathway | Stemness transformation of OC cells can be activated by SNORA72 through the Notch 1-c-Myc pathway |
| Chau [43] | 2013 | Wnt/β-catenin pathway | The activation of Wnt/β-catenin and ATP-binding cassette G2 downstream of c-Kit could promote OCSCs |
| Mariya [132] | 2016 | Wnt/β-catenin pathway | MMP10 activate canonical Wnt signaling by inhibiting noncanonical Wnt signaling ligand Wnt5a |
| Chen [133] | 2017 | Wnt/β-catenin pathway | STAT3 signaling maintained stemness and interconnected Wnt/β-catenin signaling via the miR-92a/DKK1 regulatory pathways |
| Deng [45] | 2017 | Wnt/β-catenin pathway | Rb1 and compound K can chemosensitize OCSCs, inducing a synergistic cytotoxicity via Wnt/β-catenin pathway and epithelial-to-mesenchymal (EMT) transition regulation |
| Wen [114] | 2017 | Wnt/β-catenin pathway | SOX2 may directly bind to bcatenin and activate the Wnt/β-catenin pathway to maintain the stemness of ovarian spheroids |
| Srivastava [46] | 2018 | Wnt/β-catenin pathway | Calcitriol is able to deplete the OCSC population by inhibiting their Wnt signaling Pathway |
| Wu [134] | 2015 | Wnt/β-catenin pathway | MiR-1207 enhances the stem cell-like traits of OC cells by downregulating of multiple negative modulators of the Wnt/β-catenin pathway |
| Pan [44] | 2018 | Wnt/β-catenin pathway | Theaflavin-3, 3′-digallate inhibits OCSCs through Wnt/β-catenin signaling pathway |
| Deng [50] | 2019 | PI3K/Akt/mTOR pathway | EMT and enhanced CSC marker expression triggered by activated PI3K/Akt/mTOR signaling are involved in the chemoresistance of EOC |
| Ning [50, 52] | 2016 | PI3K/Akt/mTOR pathway | The reduced OVCSLC oncogenicity by DFOG through inhibitory effects on AKT and/or ERK and/or NF-κB pathways requires both FoxO3a and FoxM1 expression |
| Leizer [52] | 2010 | NF-κB pathway | EriB can inhibit NF-κB activity by down-regulating the level of nuclear p65 |
| Jiang [52] | 2022 | NF-κB pathway | PFKFB3 is a key process of glycolysis in OCSCs and its inhibitor can impede stem properties |
| Xia [52] | 2014 | Hippo pathway | The self-renewal and chemoresistance properties of OCSCs depend on YAP and TEADs |
| Feng [52] | 2016 | Hippo pathway | VP can increase YAP expression, resulting in the suppression of OCSCs progression |
| Casagrande [71] | 2011 | CD44 | Intraperitoneal CPE administration could eradicate CD44 + OCSCs after conventional therapy |
| Cheng [72] | 2011 | CD44 | MiR-199a targets CD44 and reduces the proliferation and invasion of CD44 +/CD117 + ovarian cancer stem cells in vitro and in vivo |
| Wu [135] | 2015 | CD44 | Human SKOV3 CD117 + CD44 + CSC-based vaccine may induce the anti-OCimmunity against tumor growth by reducing the CD117 + CD44 + CSC population |
| Skubitz [78] | 2013 | CD133 | dCD133KDEL is a novel deimmunized toxin that appears to be targeting and eliminating CD133 + tumor initiating cells |
| Xiang [80] | 2013 | CD133 | The stimulation function of IL-17 on self-renewal of CD133 + OCSCs might be mediated by NF-kB and MAPK signaling pathway |
| Wang [81] | 2016 | CD133 | IL-23 contribute to ovarian cancer malignancy through promoting the self-renewal of CD133 + ovarian cancer stem-like cells |
| Long [79] | 2016 | CD133 | The Cre/LoxP system-mediated tBid overexpression activated the pro-apoptotic signaling pathway and augmented the cytotoxic effect of cisplatin in CD133 + OCSCs |
| Kim [91] | 2018 | ALDH | ATRA suppressed ALDH1 expression, inhibiting NRF2 activation, which led to the attenuation of CSC-like properties in ALDH-H cells but not in ALDH-L cells |
| Choi [96] | 2015 | ALDH | BMP2 promotes the expansion of ALDH + C133 + OCSCs and restricting the growth of progenitor |
| Kakar [94] | 2017 | ALDH | WFA alone or when combined with CIS resulted in a significant suppression of tumorigenic function of isolated ALDH1 positive cancer stem cells in vitro |
| Young [92] | 2014 | ALDH | ATRA downregulates ALDH1/FoxM1/Notch1 signaling and suppresses tumor formation in OC cells |
| Cui [93] | 2018 | ALDH | DDB2, a transcription repressor, can abrogate ovarian CSC properties by downregulating ALDH1A1 expression |
| Shank [103] | 2012 | Metformin | Metformin can restrict the growth and proliferation of ovarian cancer stem cells in vitro and in vivo |
| Zhang [106] | 2015 | Metformin | Metformin at low dose inhibits selectively CD44 + CD117 + OCSCs through inhibition of EMT and potentiates the effect of cisplatin |
| Lee [115] | 2017 | Salinomycin | Combining salinomycin with other anti-cancer therapeutic agents can target OCSCs |
| Mi [116] | 2017 | Salinomycin | Salinomycin-loaded poly(lactic-co-glycolic acid)-poly(ethylene glycol) nanoparticles conjugated with CD133 antibodies can eliminate CD133 + OCSCs |
| Lee [123] | 2020 | Calcium channel blockers | Combination CCBs with cisplatni can inhibit the viability and proliferation of OCSCs |
| Lee [124] | 2020 | Calcium channel blockers | Poziotinib with a CCB can effectively inhibit OCSC survival and function |
Table 3.
Therapeutics targeting the OCSCs
| Agent | Target | Mechanism of action | Study phase |
|---|---|---|---|
| DAPT | Notch pathway | GSIs | In vitro |
| WFA | Notch pathway | Notch1 inhibitor | Both in vitro and in vivo |
| Eugenol | Notch pathway | Notch-Hes1 inhibitor | Both in vitro and in vivo |
| TF3 | Wnt/β-Catenin | β-Catenin down-regulator | In vitro |
| Ginsenoside-Rb1 metabolite compound K | Wnt/β-Catenin | β-Catenin down-regulator | Both in vitro and in vivo |
| Calcitriol | Wnt/β-Catenin | VDR down-regulator | Clinical trial phase 1 |
| N-t-boc-Daidzein | PI3K/Akt/mTOR | Akt/mTOR down-regulator | In vitro |
| DFOG | PI3K/Akt/mTOR | FOXM1 inhibitor | In vitro |
| EriB | NF-κB | Nuclear p54 inhibitor | In vitro |
| 3PO | NF-κB | PFKFB3 inhibitor | In vitro |
| PFK158 | NF-κB | PFKFB3 inhibitor | Clinical trial phase 1 |
| VP | Hippo | YAP inhibitor | Both in vitro and in vivo |
| dCD133KDEL | CD133 | CD133 monoclonal antibody | Both in vitro and in vivo |
| ATRA | ALDH | ALDH1 inhibitor | Both in vitro and in vivo |
| DDB2 | ALDH | ALDH1a1 inhibitor | Both in vitro and in vivo |
| Metformin | multiple | CD44 down-regulator | Clinical trial phase 2 |
| Salinomycin | multiple | Apoptotic proteins promoter | Both in vitro and in vivo |
| CCBs | AKT/ERK pathway | Apoptotic proteins promoter | In vitro |
-
2.
OCSCs-related signaling pathways
A range of signaling pathways were shown to regulate the maintenance, self-replication, differentiation and drug resistance properties of OCSCs. Identification of signaling pathways regulating OCSCs is the key for eradicati–ng OCSCs and in turn harnessing drug resistance and relapse of tumors. We have summarized the related studies as follows and showed the pathways in Figs. 3, 4, 5, 6 and 7.
Fig. 3.

Notch signaling pathway in OCSCs
Fig. 4.

Wnt/β-catenin signaling pathway in OCSCs
Fig. 5.

PI3K/AKT/mTOR signaling pathway in OCSCs
Fig. 6.
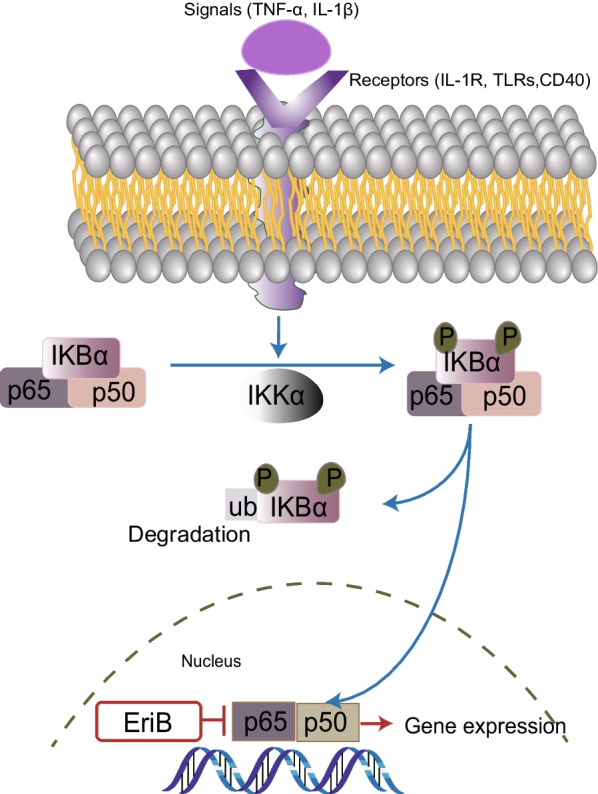
NF-κB signaling pathway in OCSCs
Fig. 7.
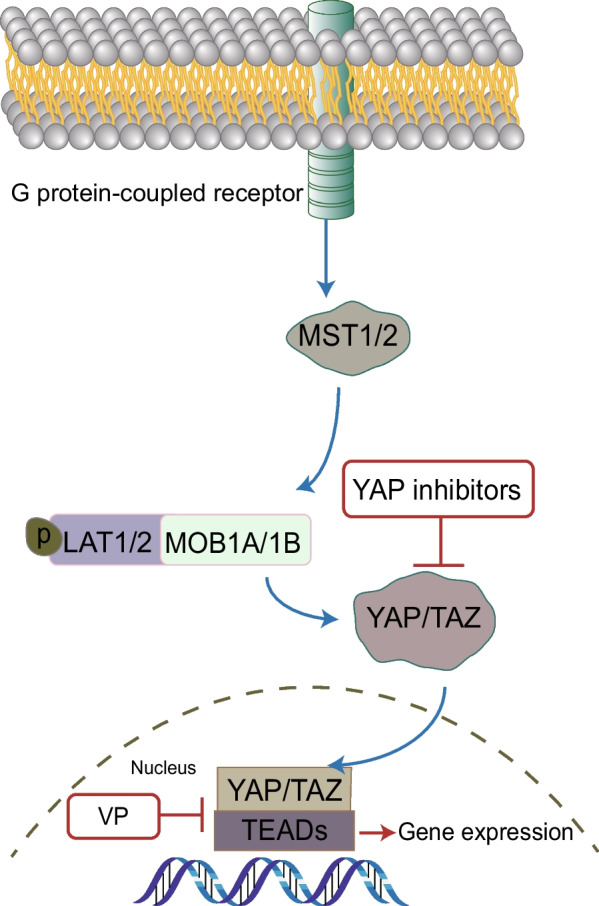
Hippo signaling pathway in OCSCs
Notch signaling pathway
The Notch signaling pathway is one of the vital pathways that constitute the CSC signaling network and is responsible for CSC maintenance, proliferation and differentiation [31]. There are four Notch receptors (Notch 1–4). In particular, the Notch3 signaling pathway is important for OCSC maintenance and tumor resistance to platinum. Notch3 is overexpressed in more than 20% of ovarian serous adenocarcinomas and is related to aggressive subtypes [32]. Notch is a large transmembrane protein that contains an EGF-like extracellular domain that is involved in ligand binding, a transmembrane domain, and an intracellular, nuclear localization domain. After the binding of ligand proteins and Notch receptors, the Notch intracellular domain (NICD) is released by γ-secretase, following the nuclear translocation of NICD to induce transcriptional activation of Notch target genes [33].
There are several Notch pathway inhibitors with different targets and mechanisms of action. γ-Secretase inhibitors (GSIs) are the most classical agents targeting Notch signaling and block the second proteolytic cleavage of Notch receptors and the Notch intracellular domain [34]. Li et al. claimed that inhibition of Notch signaling with a γ-secretase inhibitor markedly inhibits OCSCs from self-renewing and proliferating, significantly downregulates the expression of OCSC-specific surface markers, and reduces the protein and mRNA expression of Oct4 and Sox2 in OCSCs [35]. In addition to GSIs, some other compounds that inhibit Notch, such as withaferin A (WFA), have been proven to suppress tumor growth and eliminate CSCs [36]. WFA is a bioactive compound isolated from the plant Withania somnifera, and when used in combination with cisplatin, it can prevent the increase of Notch 1, Hes1 and Hey1 expression, which presents a more efficacious therapy for ovarian cancer [36]. Hyeok demonstrated that Galectin-3 maintains OCSCs by activating the Notch1 intracellular domain (NICD1) and leads to an increase in OCSCs and resistance to cisplatin and paclitaxel-induced cell death [37]. Syed et al. proved that adding eugenol to cisplatin-treated ovarian cancer cells synergistically inhibits the Notch-Hes1 pathway and downregulates drug resistance ABC transporter genes, resulting in the inhibition of OCSC growth and elimination of resistant OCSCs [38].
Wnt/β-catenin signaling pathway
The wingless/integrated (Wnt) signaling pathway is divided into the β-catenin-dependent pathway and noncanonical Wnt pathway. It plays a significant role in cell proliferation, differentiation, migration, and organ development in the period of embryogenesis [39]. Wnt signaling requires a complex combining frizzled (Fz) and lipoprotein receptor-related protein (LRP)-Wnt-induced Fz-LRP6 complex, and complex phosphorylation is a key event in receptor activation, resulting in stabilization of β-catenin. β-Catenin and transcriptional coactivators such as T cell factor (TCF) form complexes that contribute to the transcription of Wnt-responsive genes, inducing cellular responses [40]. Wnt/β-catenin has also been associated with the progression and development of cancer, and it has been identified as one of the most vital pathways involved in CSC maintenance [41]. Accumulated β-catenin activates β-catenin signaling and results in excessive stem cell renewal and proliferation in colorectal cancer [40]. It has been claimed that the Wnt/β-catenin signaling pathway is activated in OCSCs, and targeted inhibition of β-catenin potently sensitizes cells to cisplatin and decreases OCSC sphere formation [42].
Chau et al. claimed that c-kit is a key regulator of OCSC survival and proliferation. Studies have shown that the activation of Wnt/β-catenin and ATP-binding cassette G2 downstream of c-kit lead to the acquisition and maintenance of chemoresistance and the tumor-initiating capacity of ovarian cancer cells, indicating that Wnt/β-catenin combined with c-kit could be significant for improving OC patient prognoses [43]. Several chemical compounds have been proven to exhibit inhibitory effects against OCSCs by suppressing the Wnt/β-catenin signaling pathway. Theaflavin-3,3′-digallate (TF3), a polyphenolic compound extracted from black tea, exhibits anticancer properties by downregulating the protein levels of β-catenin, LEF-1, c-Myc and cyclin D1 in OCSCs. Additionally, the overexpression of β-catenin attenuated the inhibitory effect of TF3 on the viability and tumor sphere formation capacity of OCSCs [44]. Ginsenoside-Rb1, a natural saponin isolated from the rhizome of Panax quinquefolius and notoginseng, and its metabolite compound K could effectively suppress OCSC self-renewal by downregulating β-catenin/T cell factor-dependent transcription and the expression of its target genes ATP-binding cassette G2 and P-glycoprotein, which is associated with the Wnt/β-catenin pathway [45]. Calcitriol, an active metabolite compound of vitamin D, could deplete the OCSC population and impede the growth of xenograft tumors by inhibiting the Wnt/β-catenin pathway via the VDR pathway [46].
PI3K/AKT/mTOR signaling pathway
The phosphoinositide 3-kinase (PI3K)/AKT/mammalian target of rapamycin (mTOR) pathway links the response to growth-related hormone receptor interactions to downstream targets, affecting cell proliferation, survival, and apoptosis [47]. The PI3K/AKT/mTOR pathway, found in approximately 70% of ovarian cancers, is associated with aggressive phenotypes, chemoresistance and poor prognosis [48]. PI3K activates AKT/mTOR/p70S6K1 signaling to regulate the G1 cell cycle and apoptosis in ovarian cancer, and inhibition of the PI3K/AKT/mTOR pathway has been found to disrupt ovarian cancer cell proliferation and trigger cell death [49]. Although it has been widely accepted that the PI3K/AKT/mTOR pathway is an attractive therapeutic target for OC treatment, many drugs have not advanced into late-phase clinical studies for several reasons [48].
Deng et al. claimed that the acquisition of EMT and enhancement of CSC marker expression in chemoresistant epithelial ovarian cancer cells were associated with the activation of PI3K/AKT/mTOR signaling [50]. Similar to the two pathways above, several emerging compounds have the potential to be used as novel chemotherapeutic drugs for OC by targeting PI3K/AKT/mTOR signaling. N-t-boc-Daidzein is an isoflavone Daidzein that is capable of inducing apoptosis in EOC cells, including OCSCs, which may be accomplished by promoting the degradation of AKT and inhibiting the mTOR pathway [51]. A newly synthesized genistein derivative, DFOG, has been proven to exert anticancer activity by targeting OCSCs and to reduce OCSC oncogenicity in vitro through inhibitory effects on the PI3K/AKT/mTOR and/or ERK and/or NF-κB pathways [52]. ST6GALNAC1 is responsible for encoding a sialyltransferase that acts as a catalyst for the synthesis of cancer-related sialyl-Tn antigen, which is related to metastasis and poor prognosis in patients with cancer. Wang et al. claimed that ST6GALNAC1 promoted cell proliferation, migration and invasion abilities by activating the PI3K/AKT pathway in OCSCs, providing a novel approach to the study of drug resistance and may contribute to discovering new therapies curing OC [53].
Other signaling pathways
The Nuclear factor-κ light chain enhancer of activated B cells (NF-κB) is a group of transcription factors involved widely in inflammation, cell growth and apoptosis, including RelA (p65), c-Rel, RelB, p50 (NF-κB 1) and p52 (NF-κB 2) [54]. The NF-κB pathway can be divided into canonical and non-canonical pathway, which may be crucial to the maintenance of the proliferative cell population and CSC populations in tumors, respectively [55]. It has been reported that CD44 + cells promote the stem characteristics of OCSCs by up-regulating the expression of RelA, RelB and IKKα and adjusting the nuclear activation of p50/RelA dimer [56]. The compound Eriocalyxin B (EriB), an analogue of oridonin, has been proved to inhibit NF-κB activity by decreasing the levels of nuclear p65 in the OCSCs, accompanied with the suppression of cytokine production and eventually promoting OCSCs death [57]. 6-phosphofructo-2-kinase/fructose-2,3-biphosphatase 3 (PFKFB3) is vital to the first controlled step in glycolysis which is critical for CSCs. Jiang et al. claimed that PFKFB3 upregulated in CSC-enriched ovarian cancer cells and prompted clonogenicity, sphere-formation in ovarian cancer cells. Furthermore, the over-expression of PFKFB3 has been proved to up-regulate NF-κB activation. The compound 3PO has been proved to inhibit the expression of PFKB3; however, it has not been applied clinically on account of its poor water solubility. Another PFKFB3 inhibitor, PFK158 has been reported to impede tumorigenesis and tumor-initiation in vivo and in vitro, also the first PFKFB3 inhibitor being approved by the U.S. Food and Drug Administrator (FDA) to clinical trial in patients with tumors including ovarian cancer [58].
The Hippo signaling pathway, consisted of a broad range of proteins, is a highly evolutionarily conserved regulator of biological processes, including differentiation, cellular proliferation and cell growth [59]. The core of the Hippo pathway comprises a core kinase cassette that consists of protein kinase MST1/2 and tumor suppressor LSTSA/2. The major targets of the Hippo pathway are Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ), whose activity controlled by multiple upstream branches [60]. While plenty of researches show that the activation of YAP and TAZ can induce CSC properties in various malignancies, they can be prevented from interacting with transcriptional enhancer factor TEA domain family (TEADs) for suppressing the transcription of target genes. Xia et al. firstly claimed the self-renewal and chemoresistance properties of OCSCs depend on YAP and TEADS. AKT phosphorylation has been detected in OCSCs followed by YAP/TEADs silenced, indicating that the Hippo pathway may interact with other signaling pathways [61]. Verteporfin (VP), a light-activated drug used in photodynamic therapy for the choroidal neovascular membranes, has been reported as a newly identified YAP inhibitor. Feng et al. reported that VP treatment can prevent the accumulation of YAP in nucleus and the transcriptional coactivator activity of YAP, resulting in the suppression of OC progression [62].
-
3.
Cell surface markers in OCSCs
It is now clear that in the same type of tumor, there are different populations of CSCs defined by one specific marker or different combinations of markers. Identifying and functionally characterizing OCSCs could support the development of effective therapies. Herein, we will discuss several reliable OSCS biomarkers and their relationships to OC chemoresistance and targeting potential for therapies.
CD44
CD44, a widely expressed surface transmembrane glycoprotein and the main surface receptor with a role in cell–cell interaction, adhesion and migration, is found in many solid tumors, including OC [63]. CD44 was first identified and cloned in 1989 and represents a polymorphic group of surface proteins that range from 80 to 200 kDa in size [64–66]. It was originally identified as a receptor for hyaluronan or hyaluronic acid and later a receptor `to several other ligands, also a marker for cancer stem cells of several types. Several studies have demonstrated the association of CD44 with poor prognosis in EOC patients [67, 68]. More importantly, overexpression of CD44 in OC has been shown to be strongly related to the occurrence of metastasis and disease relapse [69]. Among these studies, the connection between CD44 and CSC maintenance has been emphasized, and CD44 + ovarian cancer cell subpopulations have been proven to initiate tumorigenesis and promote disease recurrence by recapitulating the original tumor [70].
Casagrande et al. demonstrated that OCSCs were characterized as CD44+/MyD88 + cells showing cytokine and chemokine production, high capacity for repair, resistance to conventional chemotherapies, capacity to form spheroids in suspension, and most importantly, ability to recapitulate the original tumor in vivo. In that study, CD44 + OCSCs were capable of expressing high levels of the gene encoding the tight junction protein claudin-4, which was proven to be a high-affinity natural receptor for Clostridium perfringens enterotoxin (CPE). Animals with chemotherapy-resistant CD44 + OCSC tumors can gain cure or long-term survival through repeated i.p. injections of CPE. This study supports that CPE-based therapy targeting CD44 + OCSCs may have great potential for chemotherapy-resistant OC patients [71]. MiR-199a was found to specifically regulate CD44, and the overexpression of miR-199a led to reduced expression of CD44, resulting in the inhibition of proliferation, migration and invasion of OCSCs [72].
CD133
CD133, also known as Prominin-1, is a 97 kDa pentaspan membrane glycoprotein originally found on neuroepithelial stem cells in mice and later in human tissues [73]. The physiologic function of CD133 in normal biology and the progression of cancer remains elusive, and most studies focus on its use as a cell surface marker for the detection of somatic stem cells and CSCs [74]. CD133 alone or in combination with other markers has widely been used to identify stem cells from different kinds of cancers, including brain cancer, prostate cancer and OC. Among these studies, CD133-expressing CSCs show self-renewal potential and the ability to regenerate a histologically similar tumor mass [17, 75, 76]. Although there is no adequate knowledge about the molecular underpinnings of CD133 in cancer, a large range of current studies demonstrate that CD133 plays an important role in predicting overall survival, disease-free survival, and progression-free survival in several kinds of solid cancers [77]. Herein, a spectrum of therapies targeting or relating to CD133 have been developed.
A monoclonal antibody (mAb) against the CD133 fusion protein dCD133KDEL has been found to be effective in targeting CD133 + tumor-initiating cells, which are also CSCs, resulting in a decrease in both OVCAR5 cells and ovarian cancer tumor growth. This novel deimmunized toxin could arrest the proliferation of OC cells in vitro and in vivo, and it can serve as a novel treatment regimen for targeting CD133 + OCSCs [78]. Due to the complex molecular structure of CD133, the effect of CD133 antibody-targeted therapy is still limited. Long et al. utilized the Cre/LoxP regulation system to selectively augment tBid, a potent suicide gene, and introduced tBid to CD133 + OCSCs, significantly inducing cell apoptosis and inhibiting CD133 + OCSC growth both in vitro and in vivo. In addition, Cre/LoxP system-mediated tBid overexpression augmented the cytotoxic effect of cisplatin in CD133 + OCSCs, representing a future clinical approach for preventing OC metastasis and recurrence [79]. Regarding the direct interaction between CSCs and chemokines produced by inflammatory cells, some studies have explored the role of chemokines and cytokines, such as interleukin (IL)-17 and IL-23, in a subpopulation of OCSCs. Xiang et al. demonstrated that IL-17 could promote self-renewal in vitro and enhance the tumorigenic potential of CD133 + OCSCs in xenograft mice through the NF-κB and p38 MAPK signaling pathways [80]. Similarly, Wang et al. indicated that IL-23 and its receptor are both mainly expressed in OCSCs and that IL-23 could promote its self-renewal ability via activation of the STAT3 and NF-κB pathways [81].
ALDH
Aldehyde dehydrogenase (ALDH) is a family of ubiquitous enzymes located in nearly all mammalian tissues [82]. Several studies have proven that ALDH plays a role in catalyzing the oxidation of aldehydes, subsequently contributing to cellular homeostasis related to the function of stem cells, such as self-renewal capability and stress-resistant properties [83]. ALDH1 is one of the ALDH superfamily of enzymes that converts retinol to retinoic acid and functions as a modulator of cell proliferation, cellular detoxification and stem cell differentiation [84, 85]. It has been proven to be a useful marker for CSCs and is widely used to isolate CSCs in various malignancies, including OC [86, 87]. In OC, ALDH1A1 has been proven to be associated with chemoresistance and may be a potential therapeutic molecular target [88]. However, the mechanisms and signaling pathways of the biological effects of ALDH1 in OCSCs remain unclear, and there remains some controversy about the correlation between ALDH1A1 and OC patient prognosis, which deserves further exploration [89, 90].
Kim et al. indicated that high ALDH1A1, the main isoenzyme of ALDH1, led to NRF2 activation through the p62-associated pathway in ALDH1-high OCSCs, resulting in CSC properties such as drug resistance, colony formation, tumor growth and high stemness marker expression. More importantly, all-trans retinoic acid (ATRA) was proven to have the ability to suppress ALDH1 expression, subsequently suppressing CSC properties in ALDH1-high cancer cells by inhibiting NRF2 activation, which suggests the molecular basis of the ATRA effect in CSCs [91]. Similarly, Young et al. claimed that ATRA can target ALDH1 + OCSCs and suppress sphere formation ability, cell migration and invasion and tumorigenesis via ALDH1/FoxM1/Notch1 signaling [92]. In addition to ATRA, DNA damage-binding protein 2 (DDB2) was proven to suppress non-CSC-to-CSC conversion through repression of ALDH1A1 transcription, limit the CSC subpopulation, and finally halt tumor growth in OC [93]. Kakar et al. claimed that WFA alone or in combination with cisplatin significantly inhibited the spheroid formation of isolated ALDH1 + OCSCs in vitro and reduced its expression in tumors collected from mice bearing orthotopic OC by suppressing the expression of securing, an “oncogene,” which suggested the mechanisms of their antitumor effects [94]. According to several studies [23, 76, 95], ALDH combined with CD133 may be the best supported OCSC marker. Choi et al. found that bone morphologenetic protein 2 (BMP2) could act as a feedback mechanism by promoting the expansion of ALDH + CD133 + OCSCs and restricting the growth of progenitors, which supported BMP2 as a therapeutic target in OC [96].
-
4.
Drugs targeting OCSCs
A new method to find novel therapies for diseases or target molecules for drugs with already existing indications is drug repositioning or repurposing, which may drastically save time and other resources compared with traditional drug development. There are also some well-known drugs targeting OCSCs, showing a potential way to treat OC.
Metformin
Metformin, a biguanide derivative, is considered a first-line antidiabetic drug for managing type 2 diabetes. A range of studies have indicated that metformin may improve lipid metabolism and weight loss, reduce cardiovascular incidence, inhibit brain function, slow cognitive decline, and reduce the risk of dementia [97–99]. More importantly, several studies have proven that metformin has antitumor effects and improves chemotherapy sensitivity [100, 101]. Epidemiological studies have indicated that in many types of cancer, the cancer-related mortality rate of patients who take metformin is significantly lower, including pancreatic, breast, liver and endometrial cancers [102]. In OC, studies have proven that metformin has anti-proliferative and pro-apoptotic effects on cancer in vitro and in vivo. It has also been demonstrated that a major mechanism for metformin’s ability to inhibit OC growth is metformin impacting OCSC growth.
Shank et al. reported that metformin was active against primary human OCSCs in vitro and that metformin therapy alone slowed the growth of OCSCs in vivo [103]. Based on the theory that CSCs and tumor vasculature may be mutually dependent on each other, they demonstrated that the antiangiogenic effects of metformin are due to the actions of metformin on CSCs [103, 104]. Interestingly, they also mentioned earlier studies indicating that metformin acted primarily on tumors with p53 mutations, while 95% OC carries p53 mutations [105]. This study indicated the mechanisms of metformin’s antitumor effect and provided evidence of its potential significance for OC patients. Zhang et al. showed that low concentrations of metformin selectively inhibited CD44 + CD117 + OCSCs via downregulation of epithelial mesenchymal transformation [106]. They also illustrated that metformin inhibited tumor sphere formation, decreased SKOV3 and primary OC growth, and most importantly, enhanced the anticancer effect of cisplatin, which may be effective in preventing OC recurrence and improving patients’ long-term survival.
Salinomycin
Salinomycin (Sal), a monocarboxylic polyether antibiotic naturally isolated from Streptomyces albus and initially used as an agricultural antimicrobial agent [107], acts as an ionophore and promotes the transfer of cations across biological membranes via an exchange diffusion mechanism [107]. Studies have proven that Sal selectively kills CSCs in some types of cancer, such as leukemia, breast cancer, colorectal cancer, lung cancer, and gastric cancer, but the underlying mechanisms have not been well elucidated [107–112]. It has been well accepted that paclitaxel combined with Sal can produce strong antitumor effects for the eradication of breast cancer and CSCs, which provides inspiration for the application of Sal in OC [113]. It has been demonstrated that Sal inhibits growth and induces apoptosis in the cisplatin-resistant human OC cell line C13 in vitro and exhibits significant efficacy in vivo, suggesting that Sal is a promising antitumor agent in cisplatin-resistant OC therapy. There are no adequate studies about the activity against OCSCs, which demands deep exploration.
SOX2 has been recognized as a cancer stem cell-associated gene, and the overexpression of SOX2 is related to cell proliferation, cell migration, resistance to cisplatin treatment and tumorigenicity of ovarian cancer cells [114]. Lee et al. demonstrated that paclitaxel combined with Sal silenced SOX2 expression and increased apoptosis of OCSCs, while the underlying mechanism required further investigation [115]. In addition to combining Sal with paclitaxel, Sal can also be utilized with other CSC markers to improve therapeutic efficiency. Mi et al. showed that Sal exerted significantly increased cytotoxic effects in CD133 + OC cells and reduced the CSC percentage in OC cells [116]. Due to the poor water solubility of Sal, Sal-loaded antibody-conjugated nanoparticles have been developed for their delivery to CSCs. They utilized CD133-SAL-NPs to improve the targeting of Sal toward OCSCs, resulting in an increased cytotoxic effect, which represented a promising approach for the treatment of OC.
Calcium channel blockers (CCBs)
Calcium is a significant signal transduction element involved in many different cell processes, such as proliferation, differentiation, growth, cell death and apoptosis [117]. In tumor cells, it is well known that calcium channels play important roles in tumorigenesis and tumor progression by controlling gene expression, DNA synthesis, cell cycle progression, apoptosis, proliferation and migration [118–120]. The importance of calcium channels in tumorigenesis and tumor progression shows the possibility of targeting calcium channels during tumorigenesis. Studies proved that in glioblastoma stem cells, calcium channels and signaling pathways were enriched, which elicited vital cell functions in response to extracellular cues [121]. Through a screen of 72 ion channel blockers, there were 10 drugs acting on calcium-related channels among the 12 drugs capable of reducing glioblastoma stem cells [122]. In OC, the calcium channels involved in promoting cancer behaviors are mostly voltage-gated calcium channels, nonvoltage-activated calcium channels and intracellular calcium channels, such as the IP3R and Orai families.
Lee et al. established a high-throughput screening system to identify agents that inhibit the sphere-forming property and proliferation of CSCs and screened approximately 1000 compounds [123]. They found that four calcium channel blockers had anticancer effects against OCSCs, mediated via L- or T-type calcium channels. These four CCBs induced OCSC apoptosis via inhibition of AKT and ERK signaling; more importantly, the combination treatment with CCBs and cisplatin enhanced drug sensitivity in a CSC-enriched epithelial OC population. Similarly, CCBs combined with poziotnib, a panepithermal growth factor receptor inhibitor, could inhibit the expression of stem cell markers, especially CD133, NANOG, and KLF4, and could suppress the phosphorylation of STAT5, AKT and ERK, a process involved in the self-renewal ability of OCSCs [124].
-
5.
Challenges and future
Chemoresistance remains one of the major challenges for OC patients. There are various molecular mechanisms involved in chemoresistance, such as decreasing drug concentrations by drug efflux pumps, degrading drugs by detoxifying enzymes, which are related to the inherent ability of cancer cells to survive chemotherapy. The acquired chemoresistance ability of cancer cells is related to the mutation and expression of genes, which is developed through natural selection of changes to gain survival advantages [125]. p53 is known as tumor-suppressed gene and barrier to CSCs formation [126]. It is known that most of HGSOC harbor mutant p53 status, which enhances metastasis and chemoresistance. Accumulated data suggest that the chemoresistant property of CSCs interweave with mutant p53 status [127]. Ramraj et al. developed an p53-targeted strategy, mainly aimed at OC cells that harbor mutant p53, for maintenance therapy of HGSOC [128]. The contribution and mechanisms of OCSCs inducing cancer chemoresistance and recurrence need further discussion, which may effectively eradicate OC. Another great challenge is the isolation of OCSCs. Although the most common method to isolate OCSCs is to select for cells by the expression of markers, already discussed above, it is quite reliable. It may be more reliable to classify OCSC populations by transcriptional profiling considering the inherent heterogeneity of OC cells.
Conclusion
CSCs are associated with the entire process of ovarian cancer development, including initiation, metastatic progression, therapeutic resistance or disease recurrence. To overcome cancer metastasis, new strategies are still needed in current therapy. Despite our increasing knowledge of the traits of OCSCs, there remains much that we do not know. As we delve more into tumor biology and the mechanisms regulating OCSCs, there is a greater understanding of the strategies that may help to find reliable strategies for OC therapy. Our review has shown that there are different therapies to eliminate OCSCs in different ways. Findings from such a review could assist researchers in exploring targeted therapy against OCSCs, which will improve patient survival and decrease tumor relapse among OC patients.
Acknowledgements
Not applicable.
Abbreviations
- OC
Ovarian cancer
- CSCs
Cancer stem cells
- OCSCs
Ovarian cancer stem cells
- NICD
The Notch intracellular domain
- GSIs
γ-Secretase inhibitors
- WFA
Withaferin A
- NICD1
The Notch1 intracellular domain
- Wnt
Wingless/integrated
- Fz
Frizzled
- LRP
Lipoprotein receptor-related protein
- TCF
T cell factor
- TF3
Theaflavin-3,3′-digallate
- PI3K
Phosphoinositide 3-kinase
- mTOR
Mammalian target of rapamycin
- CPE
Clostridium perfringens Enterotoxin
- mAb
Monoclonal antibody
- ALDH
Aldehyde dehydrogenase
- ATRA
All-trans retinoic acid
- DDB2
DNA damage-binding protein 2
- BMP2
Bone morphologenetic protein 2
- Sal
Salinomycin
- CCBs
Calcium channel blockers
Author contributions
The manuscript was written by HM. ZC and TT provided critical revision of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2020. CA Cancer J Clin. 2020;70(1):7–30. doi: 10.3322/caac.21590. [DOI] [PubMed] [Google Scholar]
- 2.Kurman RJ, Carcangiu ML, Herrington CS, et al. WHO classification of tumours of the female reproductive organs. Lyon: WHO Press; 2014. [Google Scholar]
- 3.Hatano Y, Hatano K, Tamada M, Morishige KI, Tomita H, Yanai H, Hara A. A comprehensive review of ovarian serous carcinoma. Adv Anat Pathol. 2019;26(5):329–339. doi: 10.1097/PAP.0000000000000243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kurman RJ, Shih IM. The dualistic model of ovarian carcinogenesis: revisited, revised, and expanded. Am J Pathol. 2016;186(4):733–747. doi: 10.1016/j.ajpath.2015.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kim A, Ueda Y, Naka T, Enomoto T. Therapeutic strategies in epithelial ovarian cancer. J Exp Clin Cancer Res. 2012;31(1):14. doi: 10.1186/1756-9966-31-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Siddiqui MK, Tyczynski J, Pahwa A, Fernandes AW. Objective response rate is a possible surrogate endpoint for survival in patients with advanced, recurrent ovarian cancer. Gynecol Oncol. 2017;146(1):44–51. doi: 10.1016/j.ygyno.2017.03.515. [DOI] [PubMed] [Google Scholar]
- 7.Bonadio RC, Estevez-Diz MDP. Perspectives on PARP inhibitor combinations for ovarian cancer. Front Oncol. 2021;11:754524. doi: 10.3389/fonc.2021.754524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Xu Q, Li Z. Update on poly ADP-ribose polymerase inhibitors in ovarian cancer with non-BRCA mutations. Front Pharmacol. 2021;12:743073. doi: 10.3389/fphar.2021.743073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Franzese E, Centonze S, Diana A, Carlino F, Guerrera LP, Di Napoli M, De Vita F, Pignata S, Ciardiello F, Orditura M. PARP inhibitors in ovarian cancer. Cancer Treat Rev. 2019;73:1–9. doi: 10.1016/j.ctrv.2018.12.002. [DOI] [PubMed] [Google Scholar]
- 10.Yang C, Xia BR, Zhang ZC, Zhang YJ, Lou G, Jin WL. Immunotherapy for ovarian cancer: adjuvant, combination, and neoadjuvant. Front Immunol. 2020;11:577869. doi: 10.3389/fimmu.2020.577869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zamarin D. Novel therapeutics: response and resistance in ovarian cancer. Int J Gynecol Cancer. 2019;29(Suppl 2):s16–s21. doi: 10.1136/ijgc-2019-000456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kossaï M, Leary A, Scoazec JY, Genestie C. Ovarian cancer: a heterogeneous disease. Pathobiology. 2018;85(1–2):41–49. doi: 10.1159/000479006. [DOI] [PubMed] [Google Scholar]
- 13.Terraneo N, Jacob F, Dubrovska A, Grünberg J. Novel therapeutic strategies for ovarian cancer stem cells. Front Oncol. 2020;10:319. doi: 10.3389/fonc.2020.00319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jordan CT, Guzman ML, Noble M. Cancer stem cells. N Engl J Med. 2006;355(12):1253–1261. doi: 10.1056/NEJMra061808. [DOI] [PubMed] [Google Scholar]
- 15.Bonnet D, Dick JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med. 1997;3(7):730–737. doi: 10.1038/nm0797-730. [DOI] [PubMed] [Google Scholar]
- 16.Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci USA. 2003;100(7):3983–8. 10.1073/pnas.0530291100. Erratum in: Proc Natl Acad Sci U S A. 2003;100(11):6890. [DOI] [PMC free article] [PubMed]
- 17.Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T, Henkelman RM, Cusimano MD, Dirks PB. Identification of human brain tumour initiating cells. Nature. 2004;432(7015):396–401. doi: 10.1038/nature03128. [DOI] [PubMed] [Google Scholar]
- 18.Collins AT, Maitland NJ. Prostate cancer stem cells. Eur J Cancer. 2006;42(9):1213–1218. doi: 10.1016/j.ejca.2006.01.037. [DOI] [PubMed] [Google Scholar]
- 19.Ricci-Vitiani L, Lombardi DG, Pilozzi E, Biffoni M, Todaro M, Peschle C, De Maria R. Identification and expansion of human colon-cancer-initiating cells. Nature. 2007;445(7123):111–115. doi: 10.1038/nature05384. [DOI] [PubMed] [Google Scholar]
- 20.Bapat SA, Mali AM, Koppikar CB, Kurrey NK. Stem and progenitor-like cells contribute to the aggressive behavior of human epithelial ovarian cancer. Cancer Res. 2005;65(8):3025–3029. doi: 10.1158/0008-5472.CAN-04-3931. [DOI] [PubMed] [Google Scholar]
- 21.Zhang S, Balch C, Chan MW, Lai HC, Matei D, Schilder JM, Yan PS, Huang TH, Nephew KP. Identification and characterization of ovarian cancer-initiating cells from primary human tumors. Cancer Res. 2008;68(11):4311–4320. doi: 10.1158/0008-5472.CAN-08-0364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gao MQ, Choi YP, Kang S, Youn JH, Cho NH. CD24+ cells from hierarchically organized ovarian cancer are enriched in cancer stem cells. Oncogene. 2010;29(18):2672–2680. doi: 10.1038/onc.2010.35. [DOI] [PubMed] [Google Scholar]
- 23.Curley MD, Therrien VA, Cummings CL, Sergent PA, Koulouris CR, Friel AM, Roberts DJ, Seiden MV, Scadden DT, Rueda BR, Foster R. CD133 expression defines a tumor initiating cell population in primary human ovarian cancer. Stem Cells. 2009;27(12):2875–2883. doi: 10.1002/stem.236. [DOI] [PubMed] [Google Scholar]
- 24.Korkaya H, Liu S, Wicha MS. Regulation of cancer stem cells by cytokine networks: attacking cancer’s inflammatory roots. Clin Cancer Res. 2011;17(19):6125–6129. doi: 10.1158/1078-0432.CCR-10-2743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Todaro M, Alea MP, Di Stefano AB, Cammareri P, Vermeulen L, Iovino F, Tripodo C, Russo A, Gulotta G, Medema JP, Stassi G. Colon cancer stem cells dictate tumor growth and resist cell death by production of interleukin-4. Cell Stem Cell. 2007;1(4):389–402. doi: 10.1016/j.stem.2007.08.001. [DOI] [PubMed] [Google Scholar]
- 26.Kim CF, Dirks PB. Cancer and stem cell biology: how tightly intertwined? Cell Stem Cell. 2008;3(2):147–150. doi: 10.1016/j.stem.2008.07.012. [DOI] [PubMed] [Google Scholar]
- 27.Ahmed N, Abubaker K, Findlay J, Quinn M. Cancerous ovarian stem cells: obscure targets for therapy but relevant to chemoresistance. J Cell Biochem. 2013;114(1):21–34. doi: 10.1002/jcb.24317. [DOI] [PubMed] [Google Scholar]
- 28.Soo RA, Anderson BO, Cho BC, Yang CH, Liao M, Lim WT, Goldstraw P, Mok TS, Asian Oncology Summit First-line systemic treatment of advanced stage non-small-cell lung cancer in Asia: consensus statement from the Asian Oncology Summit 2009. Lancet Oncol. 2009;10(11):1102–1110. doi: 10.1016/S1470-2045(09)70238-4. [DOI] [PubMed] [Google Scholar]
- 29.Arksey H, O’Malley L. Scoping studies: towards a methodological framework. Int J Soc Res Methodol. 2005;8(1):19–32. doi: 10.1080/1364557032000119616. [DOI] [Google Scholar]
- 30.Levac D, Colquhoun H, O’Brien KK. Scoping studies: advancing the methodology. Implement Sci. 2010;5:69. doi: 10.1186/1748-5908-5-69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Clara JA, Monge C, Yang Y, Takebe N. Targeting signalling pathways and the immune microenvironment of cancer stem cells—a clinical update. Nat Rev Clin Oncol. 2020;17(4):204–232. doi: 10.1038/s41571-019-0293-2. [DOI] [PubMed] [Google Scholar]
- 32.McAuliffe SM, Morgan SL, Wyant GA, Tran LT, Muto KW, Chen YS, Chin KT, Partridge JC, Poole BB, Cheng KH, Daggett J, Jr, Cullen K, Kantoff E, Hasselbatt K, Berkowitz J, Muto MG, Berkowitz RS, Aster JC, Matulonis UA, Dinulescu DM. Targeting Notch, a key pathway for ovarian cancer stem cells, sensitizes tumors to platinum therapy. Proc Natl Acad Sci USA. 2012;109(43):E2939–E2948. doi: 10.1073/pnas.1206400109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Choi JH, Park JT, Davidson B, Morin PJ, Shih IM, Wang TL. Jagged-1 and Notch3 juxtacrine loop regulates ovarian tumor growth and adhesion. Cancer Res. 2008;68(14):5716–5723. doi: 10.1158/0008-5472.CAN-08-0001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch signaling: cell fate control and signal integration in development. Science. 1999;284(5415):770–776. doi: 10.1126/science.284.5415.770. [DOI] [PubMed] [Google Scholar]
- 35.Jiang LY, Zhang XL, Du P, Zheng JH. γ-Secretase inhibitor, DAPT inhibits self-renewal and stemness maintenance of ovarian cancer stem-like cells in vitro. Chin J Cancer Res. 2011;23(2):140–146. doi: 10.1007/s11670-011-0140-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kakar SS, Ratajczak MZ, Powell KS, Moghadamfalahi M, Miller DM, Batra SK, Singh SK. Withaferin a alone and in combination with cisplatin suppresses growth and metastasis of ovarian cancer by targeting putative cancer stem cells. PLoS ONE. 2014;9(9):e107596. doi: 10.1371/journal.pone.0107596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kang HG, Kim DH, Kim SJ, Cho Y, Jung J, Jang W, Chun KH. Galectin-3 supports stemness in ovarian cancer stem cells by activation of the Notch1 intracellular domain. Oncotarget. 2016;7(42):68229–68241. doi: 10.18632/oncotarget.11920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Islam SS, Aboussekhra A. Sequential combination of cisplatin with eugenol targets ovarian cancer stem cells through the Notch-Hes1 signalling pathway. J Exp Clin Cancer Res. 2019;38(1):382. doi: 10.1186/s13046-019-1360-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Clevers H. Wnt/beta-catenin signaling in development and disease. Cell. 2006;127(3):469–480. doi: 10.1016/j.cell.2006.10.018. [DOI] [PubMed] [Google Scholar]
- 40.MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell. 2009;17(1):9–26. doi: 10.1016/j.devcel.2009.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wang Y, Krivtsov AV, Sinha AU, North TE, Goessling W, Feng Z, Zon LI, Armstrong SA. The Wnt/beta-catenin pathway is required for the development of leukemia stem cells in AML. Science. 2010;327(5973):1650–1653. doi: 10.1126/science.1186624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Nagaraj AB, Joseph P, Kovalenko O, Singh S, Armstrong A, Redline R, Resnick K, Zanotti K, Waggoner S, DiFeo A. Critical role of Wnt/β-catenin signaling in driving epithelial ovarian cancer platinum resistance. Oncotarget. 2015;6(27):23720–23734. doi: 10.18632/oncotarget.4690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chau WK, Ip CK, Mak ASC, et al. c-Kit mediates chemoresistance and tumor-initiating capacity of ovarian cancer cells through activation of Wnt/β-catenin-ATP-binding cassette G2 signaling. Oncogene. 2013;32:2767–2781. doi: 10.1038/onc.2012.290. [DOI] [PubMed] [Google Scholar]
- 44.Pan H, Kim E, Rankin GO, Rojanasakul Y, Tu Y, Chen YC. Theaflavin-3, 3′-digallate inhibits ovarian cancer stem cells by suppressing Wnt/β-catenin signaling pathway. J Funct Foods. 2018;50:1–7. doi: 10.1016/j.jff.2018.09.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Deng S, Wong CKC, Lai HC, Wong AST. Ginsenoside-Rb1 targets chemotherapy-resistant ovarian cancer stem cells via simultaneous inhibition of Wnt/β-catenin signaling and epithelial-to-mesenchymal transition. Oncotarget. 2017;8(16):25897–25914. doi: 10.18632/oncotarget.13071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Srivastava AK, Rizvi A, Cui T, Han C, Banerjee A, Naseem I, Zheng Y, Wani AA, Wang QE. Depleting ovarian cancer stem cells with calcitriol. Oncotarget. 2018;9(18):14481–14491. doi: 10.18632/oncotarget.24520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.McAuliffe PF, Meric-Bernstam F, Mills GB, Gonzalez-Angulo AM. Deciphering the role of PI3K/Akt/mTOR pathway in breast cancer biology and pathogenesis. Clin Breast Cancer. 2010;10(Suppl 3):S59–65. doi: 10.3816/CBC.2010.s.013. [DOI] [PubMed] [Google Scholar]
- 48.Ediriweera MK, Tennekoon KH, Samarakoon SR. Role of the PI3K/AKT/mTOR signaling pathway in ovarian cancer: Biological and therapeutic significance. Semin Cancer Biol. 2019;59:147–160. doi: 10.1016/j.semcancer.2019.05.012. [DOI] [PubMed] [Google Scholar]
- 49.Gao N, Flynn DC, Zhang Z, Zhong XS, Walker V, Liu KJ, Shi X, Jiang BH. G1 cell cycle progression and the expression of G1 cyclins are regulated by PI3K/AKT/mTOR/p70S6K1 signaling in human ovarian cancer cells. Am J Physiol Cell Physiol. 2004;287(2):C281–C291. doi: 10.1152/ajpcell.00422.2003. [DOI] [PubMed] [Google Scholar]
- 50.Deng J, Bai X, Feng X, Ni J, Beretov J, Graham P, Li Y. Inhibition of PI3K/Akt/mTOR signaling pathway alleviates ovarian cancer chemoresistance through reversing epithelial-mesenchymal transition and decreasing cancer stem cell marker expression. BMC Cancer. 2019;19(1):618. doi: 10.1186/s12885-019-5824-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Green JM, Alvero AB, Kohen F, Mor G. 7-(O)-Carboxymethyl daidzein conjugated to N-t-Boc-hexylenediamine: a novel compound capable of inducing cell death in epithelial ovarian cancer stem cells. Cancer Biol Ther. 2009;8(18):1747–1753. doi: 10.4161/cbt.8.18.9285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ning Y, Xu M, Cao X, Chen X, Luo X. Inactivation of AKT, ERK and NF-κB by genistein derivative, 7-difluoromethoxyl-5,4′-di-n-octylygenistein, reduces ovarian carcinoma oncogenicity. Oncol Rep. 2017;38(2):949–958. doi: 10.3892/or.2017.5709. [DOI] [PubMed] [Google Scholar]
- 53.Wang WY, Cao YX, Zhou X, Wei B, Zhan L, Sun SY. Stimulative role of ST6GALNAC1 in proliferation, migration and invasion of ovarian cancer stem cells via the Akt signaling pathway. Cancer Cell Int. 2019;19:86. doi: 10.1186/s12935-019-0780-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Oeckinghaus A, Ghosh S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb Perspect Biol. 2009;1(4):a000034. doi: 10.1101/cshperspect.a000034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Harrington BS, Annunziata CM. NF-κB signaling in ovarian cancer. Cancers (Basel) 2019;11(8):1182. doi: 10.3390/cancers11081182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Gonzalez-Torres C, Gaytan-Cervantes J, Vazquez-Santillan K, Mandujano-Tinoco EA, Ceballos-Cancino G, Garcia-Venzor A, Zampedri C, Sanchez-Maldonado P, Mojica-Espinosa R, Jimenez-Hernandez LE, Maldonado V. NF-κB participates in the stem cell phenotype of ovarian cancer cells. Arch Med Res. 2017;48(4):343–351. doi: 10.1016/j.arcmed.2017.08.001. [DOI] [PubMed] [Google Scholar]
- 57.Leizer AL, Alvero AB, Fu HH, Holmberg JC, Cheng YC, Silasi DA, Rutherford T, Mor G. Regulation of inflammation by the NF-κB pathway in ovarian cancer stem cells. Am J Reprod Immunol. 2011;65(4):438–447. doi: 10.1111/j.1600-0897.2010.00914.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Jiang YX, Siu MKY, Wang JJ, Leung THY, Chan DW, Cheung ANY, Ngan HYS, Chan KKL. PFKFB3 regulates chemoresistance, metastasis and stemness via IAP proteins and the NF-κB signaling pathway in ovarian cancer. Front Oncol. 2022;12:748403. doi: 10.3389/fonc.2022.748403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Harvey KF, Zhang X, Thomas DM. The Hippo pathway and human cancer. Nat Rev Cancer. 2013;13(4):246–257. doi: 10.1038/nrc3458. [DOI] [PubMed] [Google Scholar]
- 60.Song Y, Pan S, Li K, Chen X, Wang ZP, Zhu X. Insight into the role of multiple signaling pathways in regulating cancer stem cells of gynecologic cancers. Semin Cancer Biol. 2022;85:219–233. doi: 10.1016/j.semcancer.2021.06.001. [DOI] [PubMed] [Google Scholar]
- 61.Xia Y, Zhang YL, Yu C, Chang T, Fan HY. YAP/TEAD co-activator regulated pluripotency and chemoresistance in ovarian cancer initiated cells. PLoS ONE. 2014;9(11):e109575. doi: 10.1371/journal.pone.0109575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Feng J, Gou J, Jia J, Yi T, Cui T, Li Z. Verteporfin, a suppressor of YAP-TEAD complex, presents promising antitumor properties on ovarian cancer. Onco Targets Ther. 2016;9:5371–5381. doi: 10.2147/OTT.S109979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Bartakova A, Michalova K, Presl J, Vlasak P, Kostun J, Bouda J. CD44 as a cancer stem cell marker and its prognostic value in patients with ovarian carcinoma. J Obstet Gynaecol. 2018;38(1):110–114. doi: 10.1080/01443615.2017.1336753. [DOI] [PubMed] [Google Scholar]
- 64.Chen C, Zhao S, Karnad A, Freeman JW. The biology and role of CD44 in cancer progression: therapeutic implications. J Hematol Oncol. 2018;11(1):64. doi: 10.1186/s13045-018-0605-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Basakran NS. CD44 as a potential diagnostic tumor marker. Saudi Med J. 2015;36(3):273–279. doi: 10.15537/smj.2015.3.9622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Senbanjo LT, Chellaiah MA. CD44: a multifunctional cell surface adhesion receptor is a regulator of progression and metastasis of cancer cells. Front Cell Dev Biol. 2017;5:18. doi: 10.3389/fcell.2017.00018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Cho EY, Choi Y, Chae SW, Sohn JH, Ahn GH. Immunohistochemical study of the expression of adhesion molecules in ovarian serous neoplasms. Pathol Int. 2006;56(2):62–70. doi: 10.1111/j.1440-1827.2006.01925.x. [DOI] [PubMed] [Google Scholar]
- 68.Sacks JD, Barbolina MV. Expression and function of CD44 in epithelial ovarian carcinoma. Biomolecules. 2015;5(4):3051–3066. doi: 10.3390/biom5043051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Chen H, Hao J, Wang L, Li Y. Coexpression of invasive markers (uPA, CD44) and multiple drug-resistance proteins (MDR1, MRP2) is correlated with epithelial ovarian cancer progression. Br J Cancer. 2009;101(3):432–440. doi: 10.1038/sj.bjc.6605185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Alvero AB, Chen R, Fu HH, Montagna M, Schwartz PE, Rutherford T, Silasi DA, Steffensen KD, Waldstrom M, Visintin I, Mor G. Molecular phenotyping of human ovarian cancer stem cells unravels the mechanisms for repair and chemoresistance. Cell Cycle. 2009;8(1):158–166. doi: 10.4161/cc.8.1.7533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Casagrande F, Cocco E, Bellone S, Richter CE, Bellone M, Todeschini P, Siegel E, Varughese J, Arin-Silasi D, Azodi M, Rutherford TJ, Pecorelli S, Schwartz PE, Santin AD. Eradication of chemotherapy-resistant CD44+ human ovarian cancer stem cells in mice by intraperitoneal administration of Clostridium perfringens enterotoxin. Cancer. 2011;117(24):5519–5528. doi: 10.1002/cncr.26215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Cheng W, Liu T, Wan X, Gao Y, Wang H. MicroRNA-199a targets CD44 to suppress the tumorigenicity and multidrug resistance of ovarian cancer-initiating cells. FEBS J. 2012;279(11):2047–2059. doi: 10.1111/j.1742-4658.2012.08589.x. [DOI] [PubMed] [Google Scholar]
- 73.Taïeb N, Maresca M, Guo XJ, Garmy N, Fantini J, Yahi N. The first extracellular domain of the tumour stem cell marker CD133 contains an antigenic ganglioside-binding motif. Cancer Lett. 2009;278(2):164–173. doi: 10.1016/j.canlet.2009.01.013. [DOI] [PubMed] [Google Scholar]
- 74.Glumac PM, LeBeau AM. The role of CD133 in cancer: a concise review. Clin Transl Med. 2018;7(1):18. doi: 10.1186/s40169-018-0198-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Collins AT, Berry PA, Hyde C, Stower MJ, Maitland NJ. Prospective identification of tumorigenic prostate cancer stem cells. Cancer Res. 2005;65(23):10946–10951. doi: 10.1158/0008-5472.CAN-05-2018. [DOI] [PubMed] [Google Scholar]
- 76.Kryczek I, Liu S, Roh M, Vatan L, Szeliga W, Wei S, Banerjee M, Mao Y, Kotarski J, Wicha MS, Liu R, Zou W. Expression of aldehyde dehydrogenase and CD133 defines ovarian cancer stem cells. Int J Cancer. 2012;130(1):29–39. doi: 10.1002/ijc.25967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Grosse-Gehling P, Fargeas CA, Dittfeld C, Garbe Y, Alison MR, Corbeil D, Kunz-Schughart LA. CD133 as a biomarker for putative cancer stem cells in solid tumours: limitations, problems and challenges. J Pathol. 2013;229(3):355–378. doi: 10.1002/path.4086. [DOI] [PubMed] [Google Scholar]
- 78.Skubitz AP, Taras EP, Boylan KL, Waldron NN, Oh S, Panoskaltsis-Mortari A, Vallera DA. Targeting CD133 in an in vivo ovarian cancer model reduces ovarian cancer progression. Gynecol Oncol. 2013;130(3):579–587. doi: 10.1016/j.ygyno.2013.05.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Long Q, Yang R, Lu W, Zhu W, Zhou J, Zheng C, Zhou D, Yu L, Wu J. Adenovirus-mediated truncated Bid overexpression induced by the Cre/LoxP system promotes the cell apoptosis of CD133+ ovarian cancer stem cells. Oncol Rep. 2017;37(1):155–162. doi: 10.3892/or.2016.5263. [DOI] [PubMed] [Google Scholar]
- 80.Xiang T, Long H, He L, Han X, Lin K, Liang Z, Zhuo W, Xie R, Zhu B. Interleukin-17 produced by tumor microenvironment promotes self-renewal of CD133+ cancer stem-like cells in ovarian cancer. Oncogene. 2015;34(2):165–176. doi: 10.1038/onc.2013.537. [DOI] [PubMed] [Google Scholar]
- 81.Wang D, Xiang T, Zhao Z, Lin K, Yin P, Jiang L, Liang Z, Zhu B. Autocrine interleukin-23 promotes self-renewal of CD133+ ovarian cancer stem-like cells. Oncotarget. 2016;7(46):76006–76020. doi: 10.18632/oncotarget.12579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Lindahl R. Aldehyde dehydrogenases and their role in carcinogenesis. Crit Rev Biochem Mol Biol. 1992;27(4–5):283–335. doi: 10.3109/10409239209082565. [DOI] [PubMed] [Google Scholar]
- 83.Xia Y, Wei X, Gong H, Ni Y. Aldehyde dehydrogenase serves as a biomarker for worse survival profiles in ovarian cancer patients: an updated meta-analysis. BMC Womens Health. 2018;18(1):199. doi: 10.1186/s12905-018-0686-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Carpentino JE, Hynes MJ, Appelman HD, Zheng T, Steindler DA, Scott EW, Huang EH. Aldehyde dehydrogenase-expressing colon stem cells contribute to tumorigenesis in the transition from colitis to cancer. Cancer Res. 2009;69(20):8208–8215. doi: 10.1158/0008-5472.CAN-09-1132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Sun S, Wang Z. ALDH high adenoid cystic carcinoma cells display cancer stem cell properties and are responsible for mediating metastasis. Biochem Biophys Res Commun. 2010;396(4):843–848. doi: 10.1016/j.bbrc.2010.04.170. [DOI] [PubMed] [Google Scholar]
- 86.Deng S, Yang X, Lassus H, Liang S, Kaur S, Ye Q, Li C, Wang LP, Roby KF, Orsulic S, Connolly DC, Zhang Y, Montone K, Bützow R, Coukos G, Zhang L. Distinct expression levels and patterns of stem cell marker, aldehyde dehydrogenase isoform 1 (ALDH1), in human epithelial cancers. PLoS ONE. 2010;5(4):e10277. doi: 10.1371/journal.pone.0010277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.McLean K, Gong Y, Choi Y, Deng N, Yang K, Bai S, Cabrera L, Keller E, McCauley L, Cho KR, Buckanovich RJ. Human ovarian carcinoma-associated mesenchymal stem cells regulate cancer stem cells and tumorigenesis via altered BMP production. J Clin Investig. 2011;121(8):3206–3219. doi: 10.1172/JCI45273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Januchowski R, Wojtowicz K, Sterzyſska K, Sosiſska P, Andrzejewska M, Zawierucha P, Nowicki M, Zabel M. Inhibition of ALDH1A1 activity decreases expression of drug transporters and reduces chemotherapy resistance in ovarian cancer cell lines. Int J Biochem Cell Biol. 2016;78:248–259. doi: 10.1016/j.biocel.2016.07.017. [DOI] [PubMed] [Google Scholar]
- 89.Liebscher CA, Prinzler J, Sinn BV, Budczies J, Denkert C, Noske A, Sehouli J, Braicu EI, Dietel M, Darb-Esfahani S. Aldehyde dehydrogenase 1/epidermal growth factor receptor coexpression is characteristic of a highly aggressive, poor-prognosis subgroup of high-grade serous ovarian carcinoma. Hum Pathol. 2013;44(8):1465–1471. doi: 10.1016/j.humpath.2012.12.016. [DOI] [PubMed] [Google Scholar]
- 90.Chang B, Liu G, Xue F, Rosen DG, Xiao L, Wang X, Liu J. ALDH1 expression correlates with favorable prognosis in ovarian cancers. Mod Pathol. 2009;22(6):817–823. doi: 10.1038/modpathol.2009.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Kim D, Choi BH, Ryoo IG, Kwak MK. High NRF2 level mediates cancer stem cell-like properties of aldehyde dehydrogenase (ALDH)-high ovarian cancer cells: inhibitory role of all-trans retinoic acid in ALDH/NRF2 signaling. Cell Death Dis. 2018;9(9):896. doi: 10.1038/s41419-018-0903-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Young MJ, Wu YH, Chiu WT, Weng TY, Huang YF, Chou CY. All-trans retinoic acid downregulates ALDH1-mediated stemness and inhibits tumour formation in ovarian cancer cells. Carcinogenesis. 2015;36(4):498–507. doi: 10.1093/carcin/bgv018. [DOI] [PubMed] [Google Scholar]
- 93.Cui T, Srivastava AK, Han C, Wu D, Wani N, Liu L, Gao Z, Qu M, Zou N, Zhang X, Yi P, Yu J, Bell EH, Yang SM, Maloney DJ, Zheng Y, Wani AA, Wang QE. DDB2 represses ovarian cancer cell dedifferentiation by suppressing ALDH1A1. Cell Death Dis. 2018;9(5):561. doi: 10.1038/s41419-018-0585-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Kakar SS, Parte S, Carter K, Joshua IG, Worth C, Rameshwar P, Ratajczak MZ. Withaferin A (WFA) inhibits tumor growth and metastasis by targeting ovarian cancer stem cells. Oncotarget. 2017;8(43):74494–505. 10.18632/oncotarget.20170. Erratum in: Oncotarget. 2020; 11(32):3103–4. [DOI] [PMC free article] [PubMed]
- 95.Landen CN, Jr, Goodman B, Katre AA, Steg AD, Nick AM, Stone RL, Miller LD, Mejia PV, Jennings NB, Gershenson DM, Bast RC, Jr, Coleman RL, Lopez-Berestein G, Sood AK. Targeting aldehyde dehydrogenase cancer stem cells in ovarian cancer. Mol Cancer Ther. 2010;9(12):3186–3199. doi: 10.1158/1535-7163.MCT-10-0563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Choi YJ, Ingram PN, Yang K, Coffman L, Iyengar M, Bai S, Thomas DG, Yoon E, Buckanovich RJ. Identifying an ovarian cancer cell hierarchy regulated by bone morphogenetic protein 2. Proc Natl Acad Sci USA. 2015;112(50):E6882–E6888. doi: 10.1073/pnas.1507899112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Day EA, Ford RJ, Smith BK, Mohammadi-Shemirani P, Morrow MR, Gutgesell RM, Lu R, Raphenya AR, Kabiri M, McArthur AG, McInnes N, Hess S, Paré G, Gerstein HC, Steinberg GR. Metformin-induced increases in GDF15 are important for suppressing appetite and promoting weight loss. Nat Metab. 2019;1(12):1202–1208. doi: 10.1038/s42255-019-0146-4. [DOI] [PubMed] [Google Scholar]
- 98.Bharath LP, Agrawal M, McCambridge G, Nicholas DA, Hasturk H, Liu J, Jiang K, Liu R, Guo Z, Deeney J, Apovian CM, Snyder-Cappione J, Hawk GS, Fleeman RM, Pihl RMF, Thompson K, Belkina AC, Cui L, Proctor EA, Kern PA, Nikolajczyk BS. Metformin enhances autophagy and normalizes mitochondrial function to alleviate aging-associated inflammation. Cell Metab. 2020;32(1):44–55. doi: 10.1016/j.cmet.2020.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Samaras K, Makkar S, Crawford JD, Kochan NA, Wen W, Draper B, Trollor JN, Brodaty H, Sachdev PS. Metformin Use is associated with slowed cognitive decline and reduced incident dementia in older adults with type 2 diabetes: the Sydney memory and ageing study. Diabetes Care. 2020;43(11):2691–2701. doi: 10.2337/dc20-0892. [DOI] [PubMed] [Google Scholar]
- 100.Chan AT. Metformin for cancer prevention: a reason for optimism. Lancet Oncol. 2016;17(4):407–409. doi: 10.1016/S1470-2045(16)00006-1. [DOI] [PubMed] [Google Scholar]
- 101.Wheaton WW, Weinberg SE, Hamanaka RB, Soberanes S, Sullivan LB, Anso E, Glasauer A, Dufour E, Mutlu GM, Budigner GS, Chandel NS. Metformin inhibits mitochondrial complex I of cancer cells to reduce tumorigenesis. Elife. 2014;3:e02242. doi: 10.7554/eLife.02242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Franciosi M, Lucisano G, Lapice E, Strippoli GF, Pellegrini F, Nicolucci A. Metformin therapy and risk of cancer in patients with type 2 diabetes: systematic review. PLoS ONE. 2013;8(8):e71583. doi: 10.1371/journal.pone.0071583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Shank JJ, Yang K, Ghannam J, Cabrera L, Johnston CJ, Reynolds RK, Buckanovich RJ. Metformin targets ovarian cancer stem cells in vitro and in vivo. Gynecol Oncol. 2012;127(2):390–397. doi: 10.1016/j.ygyno.2012.07.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Calabrese C, Poppleton H, Kocak M, Hogg TL, Fuller C, Hamner B, Oh EY, Gaber MW, Finklestein D, Allen M, Frank A, Bayazitov IT, Zakharenko SS, Gajjar A, Davidoff A, Gilbertson RJ. A perivascular niche for brain tumor stem cells. Cancer Cell. 2007;11(1):69–82. doi: 10.1016/j.ccr.2006.11.020. [DOI] [PubMed] [Google Scholar]
- 105.Buzzai M, Jones RG, Amaravadi RK, Lum JJ, DeBerardinis RJ, Zhao F, Viollet B, Thompson CB. Systemic treatment with the antidiabetic drug metformin selectively impairs p53-deficient tumor cell growth. Cancer Res. 2007;67(14):6745–6752. doi: 10.1158/0008-5472.CAN-06-4447. [DOI] [PubMed] [Google Scholar]
- 106.Zhang R, Zhang P, Wang H, Hou D, Li W, Xiao G, Li C. Inhibitory effects of metformin at low concentration on epithelial-mesenchymal transition of CD44(+)CD117(+) ovarian cancer stem cells. Stem Cell Res Ther. 2015;6:262. doi: 10.1186/s13287-015-0249-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Markowska A, Sajdak S, Huczyński A, Rehlis S, Markowska J. Ovarian cancer stem cells: a target for oncological therapy. Adv Clin Exp Med. 2018;27(7):1017–1020. doi: 10.17219/acem/73999. [DOI] [PubMed] [Google Scholar]
- 108.Fuchs D, Daniel V, Sadeghi M, Opelz G, Naujokat C. Salinomycin overcomes ABC transporter-mediated multidrug and apoptosis resistance in human leukemia stem cell-like KG-1a cells. Biochem Biophys Res Commun. 2010;394(4):1098–1104. doi: 10.1016/j.bbrc.2010.03.138. [DOI] [PubMed] [Google Scholar]
- 109.Dong TT, Zhou HM, Wang LL, Feng B, Lv B, Zheng MH. Salinomycin selectively targets ‘CD133+’ cell subpopulations and decreases malignant traits in colorectal cancer lines. Ann Surg Oncol. 2011;18(6):1797–1804. doi: 10.1245/s10434-011-1561-2. [DOI] [PubMed] [Google Scholar]
- 110.Wang Y. Effects of salinomycin on cancer stem cell in human lung adenocarcinoma A549 cells. Med Chem. 2011;7(2):106–111. doi: 10.2174/157340611794859307. [DOI] [PubMed] [Google Scholar]
- 111.Zhi QM, Chen XH, Ji J, Zhang JN, Li JF, Cai Q, Liu BY, Gu QL, Zhu ZG, Yu YY. Salinomycin can effectively kill ALDH(high) stem-like cells on gastric cancer. Biomed Pharmacother. 2011;65(7):509–515. doi: 10.1016/j.biopha.2011.06.006. [DOI] [PubMed] [Google Scholar]
- 112.Oak PS, Kopp F, Thakur C, Ellwart JW, Rapp UR, Ullrich A, Wagner E, Knyazev P, Roidl A. Combinatorial treatment of mammospheres with trastuzumab and salinomycin efficiently targets HER2-positive cancer cells and cancer stem cells. Int J Cancer. 2012;131(12):2808–2819. doi: 10.1002/ijc.27595. [DOI] [PubMed] [Google Scholar]
- 113.Zhang Y, Zhang H, Wang X, Wang J, Zhang X, Zhang Q. The eradication of breast cancer and cancer stem cells using octreotide modified paclitaxel active targeting micelles and salinomycin passive targeting micelles. Biomaterials. 2012;33(2):679–691. doi: 10.1016/j.biomaterials.2011.09.072. [DOI] [PubMed] [Google Scholar]
- 114.Wen Y, Hou Y, Huang Z, Cai J, Wang Z. SOX2 is required to maintain cancer stem cells in ovarian cancer. Cancer Sci. 2017;108(4):719–731. doi: 10.1111/cas.13186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Lee HG, Shin SJ, Chung HW, Kwon SH, Cha SD, Lee JE, Cho CH. Salinomycin reduces stemness and induces apoptosis on human ovarian cancer stem cell. J Gynecol Oncol. 2017;28(2):e14. doi: 10.3802/jgo.2017.28.e14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Mi Y, Huang Y, Deng J. The enhanced delivery of salinomycin to CD133+ ovarian cancer stem cells through CD133 antibody conjugation with poly(lactic-co-glycolic acid)-poly(ethylene glycol) nanoparticles. Oncol Lett. 2018;15(5):6611–6621. doi: 10.3892/ol.2018.8140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Berridge MJ, Bootman MD, Roderick HL. Calcium signalling: dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol. 2003;4(7):517–529. doi: 10.1038/nrm1155. [DOI] [PubMed] [Google Scholar]
- 118.Becchetti A, Arcangeli A. Integrins and ion channels in cell migration: implications for neuronal development, wound healing and metastatic spread. Adv Exp Med Biol. 2010;674:107–123. doi: 10.1007/978-1-4419-6066-5_10. [DOI] [PubMed] [Google Scholar]
- 119.Burgoyne RD. Neuronal calcium sensor proteins: generating diversity in neuronal Ca2+ signalling. Nat Rev Neurosci. 2007;8(3):182–193. doi: 10.1038/nrn2093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Capiod T. The need for calcium channels in cell proliferation. Recent Pat Anticancer Drug Discov. 2013;8(1):4–17. doi: 10.2174/15748928130102. [DOI] [PubMed] [Google Scholar]
- 121.Wee S, Niklasson M, Marinescu VD, Segerman A, Schmidt L, Hermansson A, Dirks P, Forsberg-Nilsson K, Westermark B, Uhrbom L, Linnarsson S, Nelander S, Andäng M. Selective calcium sensitivity in immature glioma cancer stem cells. PLoS ONE. 2014;9(12):e115698. doi: 10.1371/journal.pone.0115698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Niklasson M, Maddalo G, Sramkova Z, Mutlu E, Wee S, Sekyrova P, Schmidt L, Fritz N, Dehnisch I, Kyriatzis G, Krafcikova M, Carson BB, Feenstra JM, Marinescu VD, Segerman A, Haraldsson M, Gustavsson AL, Hammarström LG, Jenmalm Jensen A, Uhrbom L, Altelaar AF, Linnarsson S, Uhlén P, Trantirek L, Vincent CT, Nelander S, Enger PØ, Andäng M. Membrane-depolarizing channel blockers induce selective glioma cell death by impairing nutrient transport and unfolded protein/amino acid responses. Cancer Res. 2017;77(7):1741–1752. doi: 10.1158/0008-5472.CAN-16-2274. [DOI] [PubMed] [Google Scholar]
- 123.Lee H, Kim JW, Kim DK, Choi DK, Lee S, Yu JH, Kwon OB, Lee J, Lee DS, Kim JH, Min SH. Calcium channels as novel therapeutic targets for ovarian cancer stem cells. Int J Mol Sci. 2020;21(7):2327. doi: 10.3390/ijms21072327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Lee H, Kim JW, Lee DS, Min SH. Combined poziotinib with manidipine treatment suppresses ovarian cancer stem-cell proliferation and stemness. Int J Mol Sci. 2020;21(19):7379. doi: 10.3390/ijms21197379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Cornelison R, Llaneza DC, Landen CN. Emerging therapeutics to overcome chemoresistance in epithelial ovarian cancer: a mini-review. Int J Mol Sci. 2017;18(10):2171. doi: 10.3390/ijms18102171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Aloni-Grinstein R, Shetzer Y, Kaufman T, Rotter V. p53: the barrier to cancer stem cell formation. FEBS Lett. 2014;588(16):2580–2589. doi: 10.1016/j.febslet.2014.02.011. [DOI] [PubMed] [Google Scholar]
- 127.Escoll M, Gargini R, Cuadrado A, Anton IM, Wandosell F. Mutant p53 oncogenic functions in cancer stem cells are regulated by WIP through YAP/TAZ. Oncogene. 2017;36(25):3515–3527. doi: 10.1038/onc.2016.518. [DOI] [PubMed] [Google Scholar]
- 128.Solomon H, Dinowitz N, Pateras IS, Cooks T, Shetzer Y, Molchadsky A, Charni M, Rabani S, Koifman G, Tarcic O, Porat Z, Kogan-Sakin I, Goldfinger N, Oren M, Harris CC, Gorgoulis VG, Rotter V. Mutant p53 gain of function underlies high expression levels of colorectal cancer stem cells markers. Oncogene. 2018;37(12):1669–1684. doi: 10.1038/s41388-017-0060-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Seo EJ, Kim DK, Jang IH, Choi EJ, Shin SH, Lee SI, Kwon SM, Kim KH, Suh DS, Kim JH. Hypoxia-NOTCH1-SOX2 signaling is important for maintaining cancer stem cells in ovarian cancer. Oncotarget. 2016;7(34):55624–55638. doi: 10.18632/oncotarget.10954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Zhu W, Niu J, He M, Zhang L, Lv X, Liu F, Jiang L, Zhang J, Yu Z, Zhao L, Bi J, Yan Y, Wei Q, Huo H, Fan Y, Chen Y, Ding J, Wei M. SNORD89 promotes stemness phenotype of ovarian cancer cells by regulating Notch1-c-Myc pathway. J Transl Med. 2019;17(1):259. doi: 10.1186/s12967-019-2005-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Zhang L, Ma R, Gao M, Zhao Y, Lv X, Zhu W, Han L, Su P, Fan Y, Yan Y, Zhao L, Ma H, Wei M, He M. SNORA72 activates the Notch1/c-Myc pathway to promote stemness transformation of ovarian cancer cells. Front Cell Dev Biol. 2020;8:583087. doi: 10.3389/fcell.2020.583087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Mariya T, Hirohashi Y, Torigoe T, Tabuchi Y, Asano T, Saijo H, Kuroda T, Yasuda K, Mizuuchi M, Saito T, Sato N. Matrix metalloproteinase-10 regulates stemness of ovarian cancer stem-like cells by activation of canonical Wnt signaling and can be a target of chemotherapy-resistant ovarian cancer. Oncotarget. 2016;7(18):26806–26822. doi: 10.18632/oncotarget.8645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Chen MW, Yang ST, Chien MH, Hua KT, Wu CJ, Hsiao SM, Lin H, Hsiao M, Su JL, Wei LH. The STAT3-miRNA-92-Wnt signaling pathway regulates spheroid formation and malignant progression in ovarian cancer. Cancer Res. 2017;77(8):1955–1967. doi: 10.1158/0008-5472.CAN-16-1115. [DOI] [PubMed] [Google Scholar]
- 134.Wu G, Liu A, Zhu J, Lei F, Wu S, Zhang X, Ye L, Cao L, He S. MiR-1207 overexpression promotes cancer stem cell-like traits in ovarian cancer by activating the Wnt/β-catenin signaling pathway. Oncotarget. 2015;6(30):28882–28894. doi: 10.18632/oncotarget.4921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Wu D, Wang J, Cai Y, Ren M, Zhang Y, Shi F, Zhao F, He X, Pan M, Yan C, Dou J. Effect of targeted ovarian cancer immunotherapy using ovarian cancer stem cell vaccine. J Ovarian Res. 2015;8:68. doi: 10.1186/s13048-015-0196-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.