

ABSTRACT
Listeria monocytogenes is a foodborne pathogen that can tolerate a variety of extreme environments. In particular, its acid resistance (AR) capability is considered one of the key factors threating food safety. Here, we employed a microbial functional genomic technology termed transposon sequencing (Tn-seq), leading to the identification of two genes involved in cell wall peptidoglycan biosynthesis (murF) and phosphate transport (lmo2248) that play key roles in lactic acid resistance (LAR) of L. monocytogenes. Deletion of lmo2248 significantly impaired the ability of LAR in L. monocytogenes, demonstrating the accuracy of the Tn-seq results. Transcriptome analysis revealed that 31.7% of the L. monocytogenes genes on the genome were differentially expressed under lactic acid (LA) treatment, in which genes involved in phosphate transport were influenced most significantly. These findings shed light on the LAR mechanisms of L. monocytogenes, which may contribute to the development of novel strategies against foodborne pathogens.
IMPORTANCE Listeria monocytogenes is a Gram-positive foodborne pathogen with high lethality and strong stress resistance, and its strong acid tolerance leads to many foodborne illnesses occurring in low-pH foods. Lactic acid is a generally recognized as safe (GRAS) food additive approved for use by the FDA. However, the genetic determinants of lactic acid resistance in L. monocytogenes have not been fully identified. In this study, the lactic acid resistance determinants of L. monocytogenes were comprehensively identified by Tn-seq on a genome-wide scale. Two genes, murF (cell wall peptidoglycan biosynthesis) and lmo2248 (phosphate transport), were identified to play an important role in the lactic acid resistance. Moreover, genome-wide transcriptomic analysis showed that phosphotransferase system (PTS)-related genes play a key role at the transcriptional level. These findings contribute to a better understanding of the lactic acid resistance mechanism of L. monocytogenes and may provide unique targets for the development of other novel antimicrobial agents.
KEYWORDS: lactic acid, Listeria monocytogenes, RNA-seq, Tn-seq
INTRODUCTION
Listeria monocytogenes is a common Gram-positive, facultative, anaerobic bacterium and one of the most fatal foodborne pathogens worldwide (1). It is mainly transmitted by food and widely found in meat, eggs, poultry, seafood, dairy products, and vegetables (2). In immunocompromised individuals, even low levels of food contamination (approximately 102 to 104 bacteria) can cause symptoms such as bacterial sepsis, meningitis, and abortion to pregnancy, with mortality rates as high as 20 to 30% (3). Foodborne diseases caused by L. monocytogenes infection have remained a major problem in the food industry over the past decade (4, 5).
In recent years, the rapid emergence of antibiotic-resistant bacteria has gradually reduced the effectiveness of antibiotics (6). Therefore, considerable effort has been devoted to finding some efficient natural antimicrobials (7, 8). Organic acids (lactic acid, malic acid, citric acid, propionic acid, acetic acid, etc.) are widely used as preservatives and antimicrobial agents in meat products, dairy products, and other foods because of their advantages of low price, simple processing, and significant bacteriostatic effect (9). As a representative organic acid, lactic acid has been widely reported to exhibit broad-spectrum antibacterial activity against Gram-negative and Gram-positive pathogens. Wang et al. (10) found that 0.5% lactic acid could completely inhibit the growth of Salmonella enterica serovar Enteritidis, Escherichia coli, and L. monocytogenes, while causing a large amount of protein leakage. Moreover, both the FDA and the European Food Safety Authority have approved the addition of lactic acid (LA) for the food industry as an antibacterial agent (11, 12). However, many bacteria have been reported to have the ability to adapt to acid environments and survive under low-pH conditions. For instance, Yu et al. (13) found that E. coli O157 and O26 could survive for more than 12 h after exposure to pH 3.5. Mani-López et al. found (9, 14) that Salmonella is particularly capable of adapting to acidic environments and surviving under intense pH conditions and that its inducible acid tolerance response (ATR) is critical for its low pH tolerance. As a result, many foodborne illness outbreaks occur in low-pH foods (yogurt, juice, cheese, etc.). Hence, it is important to study the acid resistance (AR) mechanisms of foodborne pathogens. Previous studies have shown that the adaptive ATR mechanism of L. monocytogenes is closely related to the LisRK 2-component regulatory system, the SOS response, components of the σB regulon, changes in membrane fluidity, the F0F1-ATPase proton pump, the glutamate decarboxylase (GAD) system, and the arginine deiminase (ADI) system (15, 16). Moreover, Begley et al. and Madeo et al. also found that genes btlA and thiT contribute to the acid tolerance of L. monocytogenes (17, 18). However, many LA resistance (LAR)-related genes have not yet been fully identified, so we need to further explore the LAR mechanism of L. monocytogenes from a genome-wide level.
Transposon sequencing (Tn-seq) is a powerful method for the study of bacterial functional genomics, which combines transposon mutagenesis with high-throughput sequencing (19). It can directly link phenotype to genotype in a genome-wide scale (20, 21). Tn-seq technology facilities comprehensive identification of conditionally essential genes of foodborne pathogens in particular growth environments. For instance, Zhang et al. (22) have identified 37 genes that are essential for the growth of Enterococcus faecium in human serum. Jayeola et al. (23) have revealed the essential genes that contribute to the growth of Cronobacter sakazakii and Salmonella in low-moisture foods (LMFs). In addition, Tn-seq also plays an important role in the identification of antibiotic resistance genes in many clinically important pathogens (including the ESKAPE [E. faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter] species) (24–26). In this study, Tn-seq and transcriptome sequencing (RNA-seq) were employed to investigate the lactic acid resistance (LAR) mechanisms in L. monocytogenes, which may lead to a deeper understanding of the LAR mechanism of L. monocytogenes in response to extreme pH and shed light on the development of novel antibacterial drugs.
RESULTS
Antibacterial effect of LA against L. monocytogenes.
As shown in Fig. 1, LA exhibited a significant inhibitory effect on the growth of L. monocytogenes and showed a significant dose-dependent manner. When the concentration of LA reached 4 mg/mL (pH 4.68), the growth of L. monocytogenes was completely inhibited. At 12 h, the addition of LA (1 mg/mL [pH 6.52] and 2 mg/mL [pH 5.92]) inhibited the growth of L. monocytogenes by 15% and 36%, respectively, compared with the control group. The result indicated that high doses of LA (≥4 mg/mL) were effective inhibitors for controlling L. monocytogenes, while L. monocytogenes showed significant acid resistance (AR) to low doses of LA (≤2 mg/mL).
FIG 1.
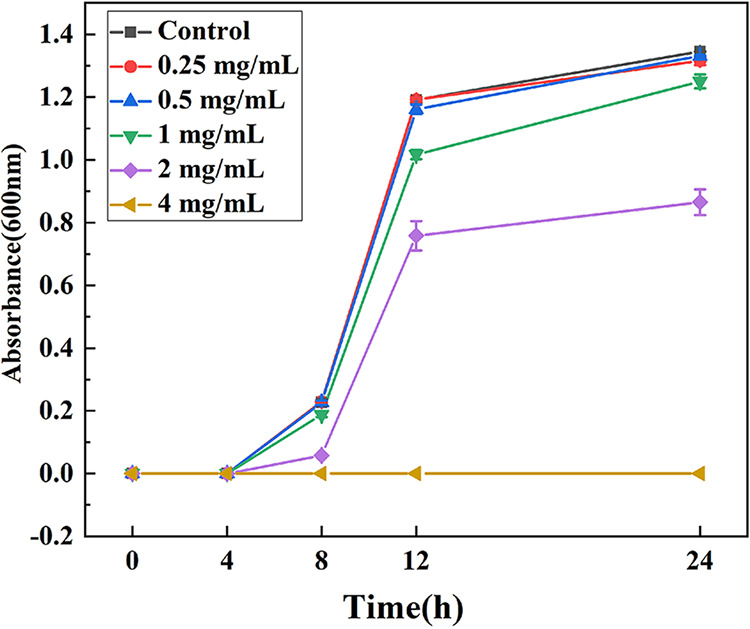
Growth curves of L. monocytogenes under different concentrations of LA treatment. Data are presented as the mean ± standard deviation (n = 3).
Identification of genetic determinants involved in LAR in L. monocytogenes by Tn-seq.
To identify the genes required for LAR in L. monocytogenes, Tn-seq was performed to determine which mutants were selectively lost during culturing in the presence of LA. Though 10 genes (fold change [FC], >2 and P < 0.05) were identified to be involved in LAR, only 2 genes (murF and lmo2248) had strict statistical significance (Benjamini-Hochberg corrected P value (BH) < 0.05) (Fig. 2 and Table 1). A single gene, murF (locus tag lmo0856), encoding a UDP-N-acetylmuramoylalanyl-d-glutamyl-2,6-diaminopimelate-d-alanyl-d-alanyl ligase, was identified by Tn-seq with the highest fold change (FC = 13.5). Another significant gene, lmo2248 (FC, 4.4), was a putative phosphate transport regulator.
FIG 2.

Tn-seq analysis to identify L. monocytogenes genes involved in LA resistance. (A) Schematic depiction of the Tn-seq. (B) Identification of genes involved in LAR by Tn-seq analysis. Different bubbles represent different genes, and bubble size corresponds to different statistical analysis values (–log[P-values]). The larger the bubble size, the stronger the significance. The x axis represents the location of the gene in the chromosome, and the y axis represents the fold change value (LA treatment versus control). Genes with significant changes (P < 0.05) are represented by colored bubbles; other genes are represented in gray. Two genes with a BH of <0.05 are shown in yellow and red.
TABLE 1.
L. monocytogenes genes involved in lactic acid resistance as determined by Tn-seq analysis
| Synonym | Gene name | Annotation | Fold changea | P value | BH valueb |
|---|---|---|---|---|---|
| lmo0856 | murF | UDP-N-acetylmuramoylalanyl-d-glutamyl-2,6-diamino pimelate-d-alanyl-d-alanyl ligase | 13.5 | 3.6E-02 | 6.6E-03 |
| lmo2248 | hypothetical protein | 4.4 | 1.3E-03 | 6.6E-03 | |
| lmo1222 | pheT | Phenylalanyl-tRNA synthetase subunit beta | 4.1 | 3.3E-02 | NS |
| lmo1078 | UDP-glucose pyrophosphorylase | 3.7 | 4.2E-02 | NS | |
| lmo2718 | cydA | Cytochrome d ubiquinol oxidase subunit I | 3.6 | 1.7E-02 | NS |
| lmo1811 | ATP-dependent DNA helicase RecG | 3.5 | 1.3E-03 | NS | |
| lmo2267 | ATP-dependent deoxyribonuclease subunit A | 3.5 | 1.3E-03 | NS | |
| lmo0885 | acpS | 4′-Phosphopantetheinyl transferase | 2.1 | 1.5E-02 | NS |
| lmo1553 | hemL | Glutamate-1-semialdehyde aminotransferase | 2.1 | 1.9E-02 | NS |
| lmo1580 | Hypothetical protein | 2.1 | 2.1E-03 | NS |
Indicates the fold change derived from the ratio of the unselected control library to the lactic acid competitively selected library.
NS, not significant (BH value of >0.05).
Effect of LA on growth of L. monocytogenes wild type and Δlmo2248 mutant.
In order to validate the results of the Tn-seq data and to determine the role of the identified genes in LA resistance, we constructed a markerless deletion mutant, Δlmo2248, in L. monocytogenes. Since murF is an essential gene (27), we selected the second-ranked gene, lmo2248, for further validation and functional studies. In the absence of LA, the growth of the wild-type strain was not significantly different from that of the mutant Δlmo2248 and the complemented strain Δlmo2248+lmo2248 (Fig. 3A). However, when the WT and Δlmo2248 were grown in brain heart infusion (BHI) with 1 mg/mL LA, the growth rate of the Δlmo2248 mutant decreased dramatically compared to that of the WT (Fig. 3B). Moreover, it can be visualized in Fig. 3C that the deletion strain of lmo2248 was almost completely unable to grow on agar plates spiked with LA (2 mg/mL). The complemented strain Δlmo2248+lmo2248 could partially restore the LA resistance to the wild-type level (Fig. 3B and C). These findings suggest that gene lmo2248 indeed contributes to LA resistance in L. monocytogenes.
FIG 3.

The effect of targeted mutation of lmo2248 on growth and morphology of L. monocytogenes in the presence of LA. (A and B) Growth curve of wild-type L. monocytogenes (WT), Δlmo2248, and Δlmo2248+lmo2248 in the absence (A) and presence (B) (1 mg/mL) of LA. (C) Growth of wild-type (WT), Δlmo2248, and Δlmo2248+lmo2248 strains on agar plates. (D and E) SEM (D) and TEM (E) images of L. monocytogenes and Δlmo2248. The WT and Δlmo2248 were both treated with LA (4 mg/mL) for 2 h.
Morphological observation of L. monocytogenes and Δlmo2248.
Scanning electron microscopy (SEM) and transmission electron microscopy (TEM) were applied to observe the effect of LA on the cell ultrastructure of L. monocytogenes (WT) and Δlmo2248. As shown in Fig. 3D, under treatment with the same concentration of LA, obvious shrinkage and damage (red arrows) appeared on the cell surface of the Δlmo2248 mutant, while no significant changes appeared in the wild type. Similarly, in the internal morphological changes of the cells (Fig. 3E), the intracellular substances (nucleic acids and proteins) of the Δlmo2248 mutant were leaked in large quantities, and the structure of the cell membrane was severely damaged. Overall, these findings suggest that the Δlmo2248 mutant is more sensitive to LA-mediated bactericidal effects than the wild type. Therefore, the accuracy of the Tn-seq data was further validated, indicating that the lmo2248 gene is indeed critical for LA resistance in L. monocytogenes.
Transcriptomic analysis of L. monocytogenes during growth in rich medium and in LA.
Transcriptomic analysis was performed to analyze differential gene expression of L. monocytogenes under LA treatment. Compared with the control, 944 differentially expressed genes (DEGs) were identified in the LA-treated group, including 413 upregulated genes and 531 downregulated genes (Fig. 4A). Among these DEGs, lmo2248 was also significantly upregulated (FC, 2.65; P < 0.01) in transcriptomic analysis. As can be seen from the heat map, the transcriptome profiles of the two groups were significantly different (Fig. 4B).
FIG 4.

Transcriptomic analysis of L. monocytogenes during growth in rich medium and in LA. (A) Volcano plot of the DEGs. (B) Heat map of DEGs. (C) Z-score bubble plot for GO enrichment analysis. The ordinate is –log10(P value), the abscissa is the up-down normalization value (the ratio of the difference between the number of differentially upregulated genes and the number of differentially downregulated genes to the total differential genes), and the size of the bubble represents the target gene enriched by the current GO term number; the yellow line represents the threshold of a P value of 0.05; the right side is the list of the top 20 terms of the P value, and different colors represent different ontologies.
Gene Ontology (GO) enrichment analysis and KEGG pathway analysis were used to further analyze the function of DEGs. The most significant top 20 GO terms are shown in Fig. 4C. Upregulated genes were concentrated in the cellular component, while downregulated genes were concentrated in the biological process and molecular function categories. For biological process, the most notable subcategory is “phosphoenolpyruvate-dependent sugar phosphotransferase system,” and for cellular component and molecular function, “ribosome” and “protein-N(PI)-phosphohistidine-sugar phosphotransferase activity” were the most significant subcategories, respectively.
KEGG pathway analysis of DEGs in L. monocytogenes was conducted in the LA treatment group compared to the control group. “Ribosome,” “arginine biosynthesis,” and “alanine, aspartate and glutamate metabolism” were the three most significantly upregulated pathways (Fig. 5A). The most significantly downregulated pathways included “phosphotransferase system (PTS),” “fructose and mannose metabolism,” and “starch and sucrose metabolism” (Fig. 5B). In addition, the associations between different pathways were also shown in Fig. 5C. More interestingly, phosphate-related genes (Table 2) were significantly enriched in both GO and KEGG analyses, indicating that they play a crucial role in LA resistance.
FIG 5.

KEGG pathway analysis of differentially expressed genes (DEGs) in L. monocytogenes under LA treatment. (A) Top 20 enriched KEGG pathways for upregulated genes. (B) Top 20 enriched KEGG pathways for downregulated genes. (C) KEGG enrichment network map. Different nodes represent different KEGG pathways, the size of the node represents the number of genes enriched in the pathway after KEGG enrichment analysis, and the gradient color of the node represents the P value of KEGG enrichment analysis. A solid line indicates that there is a connection between a pathway and a pathway or a pathway and a gene, and an isolated node in the figure indicates that the pathway is not directly related to other pathways in the figure. (D) KEGG pathway diagram for the phosphotransferase system (PTS); the downregulated genes are shown in green.
TABLE 2.
Genes related to phosphotransferase system (PTS)a
| Synonym | Gene name | Annotation | Fold change | Padj |
|---|---|---|---|---|
| lmo0021 | lmo0021 | PTS fructose transporter subunit IIA | 0.2244 | 2.05E-07 |
| lmo0023 | lmo0023 | PTS fructose transporter subunit IIC | 0.0331 | 6.73E-13 |
| lmo0027 | lmo0027 | PTS beta-glucoside transporter subunit IIABC | 0.0224 | 9.12E-287 |
| lmo0034 | lmo0034 | PTS cellobiose transporter subunit IIC | 0.2085 | 2.15E-05 |
| lmo0298 | lmo0298 | PTS beta-glucoside transporter subunit IIC | 0.0113 | 2.36E-40 |
| lmo0299 | lmo0299 | PTS beta-glucoside transporter subunit IIB | 0.0042 | 7.70E-10 |
| lmo0301 | lmo0301 | PTS beta-glucoside transporter subunit IIA | 0.0549 | 2.20E-06 |
| lmo0357 | lmo0357 | PTS sugar transporter subunit IIA | 0.3374 | 1.68E-03 |
| lmo0358 | lmo0358 | PTS fructose transporter subunit IIBC | 0.2495 | 2.06E-04 |
| lmo0374 | lmo0374 | PTS beta-glucoside transporter subunit IIB | 0.2696 | 4.37E-03 |
| lmo0399 | lmo0399 | PTS fructose transporter subunit IIB | 0.1191 | 4.36E-10 |
| lmo0400 | lmo0400 | PTS fructose transporter subunit IIC | 0.0261 | 8.89E-14 |
| lmo0426 | lmo0426 | PTS fructose transporter subunit IIA | 0.2280 | 8.76E-07 |
| lmo0427 | lmo0427 | PTS fructose transporter subunit IIB | 0.2581 | 1.04E-03 |
| lmo0428 | lmo0428 | PTS fructose transporter subunit IIC | 0.2212 | 5.17E-26 |
| lmo0503 | lmo0503 | PTS fructose transporter subunit IIA | 0.0411 | 9.05E-28 |
| lmo0507 | lmo0507 | PTS galactitol transporter subunit IIB | 0.1156 | 7.72E-09 |
| lmo0508 | lmo0508 | PTS galactitol transporter subunit IIC | 0.1089 | 2.51E-42 |
| lmo0542 | lmo0542 | PTS sorbitol transporter subunit IIA | 0.1163 | 2.30E-02 |
| lmo0543 | lmo0543 | PTS sorbitol transporter subunit IIBC | 0.1233 | 5.02E-05 |
| lmo0544 | lmo0544 | PTS sorbitol transporter subunit IIC | 0.0485 | 9.23E-05 |
| lmo0738 | lmo0738 | PTS beta-glucoside transporter subunit IIABC | 0.0009 | 0.00E + 00 |
| lmo0781 | lmo0781 | PTS mannose transporter subunit IID | 0.4964 | 3.58E-05 |
| lmo0874 | lmo0874 | PTS sugar transporter subunit IIA | 0.1111 | 3.00E-02 |
| lmo0875 | lmo0875 | PTS beta-glucoside transporter subunit IIB | 0.0355 | 2.07E-04 |
| lmo0876 | lmo0876 | PTS sugar transporter subunit IIC | 0.2438 | 3.55E-04 |
| lmo0914 | lmo0914 | PTS sugar transporter subunit IIB | 0.1141 | 5.70E-65 |
| lmo0915 | lmo0915 | PTS sugar transporter subunit IIC | 0.0134 | 1.32E-270 |
| lmo0916 | lmo0916 | PTS sugar transporter subunit IIA | 0.0051 | 4.39E-70 |
| lmo1971 | ulaA | PTS ascorbate transporter subunit IIC | 0.0581 | 2.09E-07 |
| lmo1997 | lmo1997 | PTS mannose transporter subunit IIA | 0.1123 | 5.88E-14 |
| lmo2000 | lmo2000 | PTS mannose transporter subunit IID | 0.0514 | 9.43E-34 |
| lmo2001 | lmo2001 | PTS mannose transporter subunit IIC | 0.0472 | 1.58E-40 |
| lmo2002 | lmo2002 | PTS mannose transporter subunit IIB | 0.1204 | 1.07E-17 |
| lmo2096 | lmo2096 | PTS galacticol transporter subunit IIC | 0.1197 | 4.27E-09 |
| lmo2098 | lmo2098 | PTS galacticol transporter subunit IIA | 0.1191 | 1.33E-03 |
| lmo2135 | lmo2135 | PTS fructose transporter subunit IIC | 0.2246 | 1.88E-04 |
| lmo2136 | lmo2136 | PTS fructose transporter subunit IIB | 0.1801 | 8.24E-03 |
| lmo2137 | lmo2137 | PTS fructose transporter subunit IIA | 0.0825 | 1.19E-04 |
| lmo2373 | lmo2373 | PTS beta-glucoside transporter subunit IIB | 0.4827 | 5.21E-25 |
| lmo2649 | ulaA | PTS system ascorbate transporter subunit IIC | 0.0017 | 0.00E + 00 |
| lmo2650 | lmo2650 | MFS transporter | 0.0101 | 6.86E-156 |
| lmo2651 | lmo2651 | PTS mannitol transporter subunit IIA | 0.0615 | 2.08E-143 |
| lmo2665 | lmo2665 | PTS galacticol transporter subunit IIC | 0.0033 | 0.00E + 00 |
| lmo2666 | lmo2666 | PTS galacticol transporter subunit IIB | 0.0026 | 0.00E + 00 |
| lmo2667 | lmo2667 | PTS galacticol transporter subunit IIA | 0.0024 | 0.00E + 00 |
| lmo2685 | lmo2685 | PTS cellobiose transporter subunit IIA | 0.0828 | 6.78E-50 |
| lmo2708 | lmo2708 | PTS cellobiose transporter subunit IIC | 0.4097 | 3.16E-10 |
| lmo2733 | lmo2733 | PTS fructose transporter subunit IIABC | 0.4917 | 9.23E-05 |
| lmo2762 | lmo2762 | PTS cellobiose transporter subunit IIB | 0.0644 | 3.57E-06 |
| lmo2763 | lmo2763 | PTS cellobiose transporter subunit IIC | 0.1640 | 1.13E-16 |
| lmo2765 | lmo2765 | PTS cellobiose transporter subunit IIA | 0.3373 | 4.58E-02 |
| lmo2772 | lmo2772 | PTS beta-glucoside transporter subunit IIABC | 0.0834 | 1.54E-13 |
| lmo2782 | lmo2782 | PTS cellobiose transporter subunit IIB | 0.1655 | 4.48E-02 |
| lmo2787 | bvrB | beta-Glucoside-specific phosphotransferase enzyme II ABC component | 0.2198 | 3.85E-04 |
| lmo2797 | lmo2797 | PTS mannitol transporter subunit IIA | 0.0452 | 8.74E-29 |
| lmo2799 | lmo2799 | PTS mannitol transporter subunit IIBC | 0.0043 | 5.16E-23 |
Padj, adjusted P value.
Validation of RNA-seq data by qPCR.
The accuracy of the transcriptome data was verified by real-time quantitative PCR (qPCR). As shown in Fig. 6, the RNA-seq and qPCR data of the LA-treated group and the control group were highly similar, which demonstrated the reliability and stability of the transcriptome analysis results.
FIG 6.
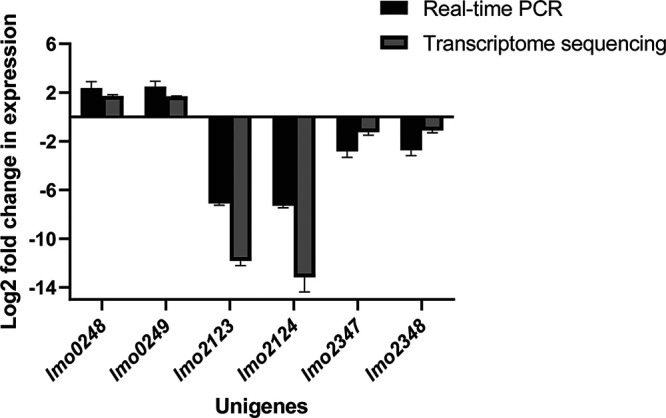
Real-time qPCR (RT-qPCR) validation of RNA-seq experiments. Gene drm was used as the housekeeping control. Each group was performed with 3 biological replicates (n = 3).
DISCUSSION
The addition of lactic acid can significantly inhibit the growth of L. monocytogenes, so it is widely used in the prevention and control of L. monocytogenes in the food industry (28, 29). However, the application and development of LA are often limited due to the strong acid resistance of L. monocytogenes. Therefore, it is very important to explore the lactic acid resistance (LAR) mechanism of L. monocytogenes.
The lack of suitable genetic tools has long been a bottleneck in the study of L. monocytogenes acid resistance mechanisms. In this study, we employed Tn-seq (19), a bacterial functional genomics research method that combines genome-wide transposon mutagenesis with high-throughput sequencing, to explore the LAR mechanism of L. monocytogenes.
Two novel LAR genes (murF and lmo2248) were identified through this genome-wide technology. Gene murF is mainly involved in the synthesis of cell wall peptidoglycan by catalyzing the final step in the synthesis of UDP-N-acetylmuramyl pentapeptide (the precursor of murein) (30). In S. aureus, the murF gene not only plays an important role in peptidoglycan biosynthesis, but may also be involved in the control of cell division (27). Gene lmo2248 was annotated as a hypothetical protein; however, based on previous studies (31), it may be a putative regulator of phosphate transport. In addition, lmo2249, a downstream gene of lmo2248, was identified to encode a low-affinity inorganic phosphate transporter mainly responsible for phosphate transport (32). Hence, the gene lmo2248 may promote LAR in L. monocytogenes by playing a role in phosphate transport. In addition, the significant upregulation of lmo2248 in the transcriptomic analysis indicated that it also plays an important role in the LAR of L. monocytogenes at the transcriptional level.
The significant differences in growth and killing curves of markerless deletion mutant Δlmo2248 and the wild type in the presence of LA strongly illustrate the accuracy of the Tn-seq data and the importance of the identified genes in LAR. In addition, their morphologically significant changes under the same concentration of LA treatment also further confirmed the importance of the genes identified in this study for LAR.
Our transcriptomic analysis of L. monocytogenes grown in LA revealed pervasive changes in gene expression compared to growth in rich media. In the GO enrichment analysis, “phosphoenolpyruvate-dependent sugar phosphotransferase system,” “ribosome,” and “protein-N(PI)-phosphohistidine-sugar phosphotransferase activity” were the most significantly enriched subcategories in the biological process, cellular component, and molecular function categories, respectively. PTS mainly phosphorylates various sugars and their derivatives through the phosphorylation cascade and then transports them into the cell (33). It not only participates in carbon and nitrogen central metabolism, regulates iron and potassium homeostasis, and regulates the virulence of certain pathogens, but also mediates stress responses (34). Interestingly, in the KEGG pathway analysis, PTS-related pathways (Fig. 5D) were also significantly enriched. Notably, gene lmo2248, identified in Tn-seq experiments, was also shown to be closely related to phosphate transport, indicating that phosphate transport-related genes play an important role in the LAR of L. monocytogenes. The results also suggested that ribosome-related genes were significantly upregulated under LA treatment, which was consistent with previous findings (12). Ribosome-related pathways were also found to be significantly enriched under other processing conditions, such as high hydrostatic pressure (35), suggesting that it may be a general stress response.
Conclusion. In this study, a genome-wide screening of genetic determinants LAR in L. monocytogenes was performed by using Tn-seq, which led to the comprehensive identification of genes that contribute to LAR, including murF and lmo2248. Moreover, transcriptional analysis revealed that phosphate transport systems were significantly regulated under LA stress conditions, which may play a crucial role in the LAR of L. monocytogenes. These findings provide new insights into the molecular mechanisms of LAR in L. monocytogenes and may contribute to the development of novel strategies against L. monocytogenes and other foodborne pathogens.
MATERIALS AND METHODS
Transposon mutant library construction and evaluation.
The high-density transposon mutant library (containing 63,666 unique insertion mutants) was constructed from previous studies in our laboratory (36, 37). Briefly, the temperature-sensitive plasmid pGPA2 was first electroporated into competent cells of L. monocytogenes. Then, the plasmid-containing strains were grown overnight at 30°C in BHI supplemented with chloramphenicol, and 200 μL of this culture was then added to BHI containing gentamicin (25 μg/mL) and nisin (25 ng/mL) for overnight growth. Next, the cultures were subjected to two consecutive passages in BHI at 37°C to obtain the mutant library. The randomness and coverage of the transposon library were assessed by PCR footprinting and reverse PCR as previously described (38).
Bacterial strains, mutants, and complement construction.
L. monocytogenes EGD-e (ATCC BAA-679, serovar 1/2a) was used throughout this study. The markerless mutant Δlmo2248 and the complemented strain Δlmo2248+lmo2248 were deposited in our laboratory. Markerless gene deletion mutant Δlmo2248 was created via the Cre-lox recombination system as previously described (38). The primers are listed in Table S1 in the supplemental material. Briefly, the upstream and downstream homology arms of lmo2248 and the gentamicin maker were PCR amplified with the primers in Table S1. Then, the three DNA fragments were seamlessly cloned (NovoRec plus one-step PCR cloning kit) into the pWS3 vector cleaved with SmaI to obtain the plasmid pWS3-lmo2248 using EC1000 as the cloning host. The recombinant plasmid was electroporated into L. monocytogenes competent cells, and the gentamicin-resistant transformants were cultured at the appropriate temperature and supplemented with the appropriate antibiotics to obtain marked mutants. Next, the plasmid pWS3-erm-cre was electroporated into the marked mutant to obtain the markerless mutants. Plasmids for the in trans complementation of the lmo2248 mutant were produced by PCR amplification of the gene using the primers listed in Table S1. The target gene fragment was then ligated to the downstream region of the PnisA promoter of pMSP3535. Next, the recombinant plasmid was introduced into Δlmo2248 competent cells and verified by PCR. Unless otherwise mentioned, L. monocytogenes and Δlmo2248 were grown in brain heart infusion (BHI) broth (Hopebio) or agar at 37°C.
Antibacterial activity assessment.
To evaluate the antibacterial activity of different concentrations of lactic acid on L. monocytogenes, 50 mL of BHI broth with different concentrations of lactic acid (Aladdin, Shanghai, China) (0 [pH 7.03], 0.25 [pH = 6.91], 0.5 [pH = 6.78], 1 [pH = 6.52], 2 [pH = 5.92], and 4 mg/mL [pH = 4.68]) were prepared. Then, the logarithmic phase L. monocytogenes was harvested and washed three times (4,000 rpm, 2 min) with phosphate-buffered saline (PBS) and diluted to obtain a final inoculum of 106 CFU/mL as the seed. Next, 1 mL of L. monocytogenes (1.0 × 106 CFU/mL) seed solution was inoculated into the shake flasks (50 mL BHI broth with different concentrations of lactic acid), and all the flasks were incubated on an orbital shaker (150 rpm at 37°C). Bacterial concentrations were monitored by measuring the optical density at 600 nm (OD600) using a UV spectrophotometer (Thermo, USA) after 4 h, 8 h, 12 h, and 24.
Tn-seq analysis of conditionally essential genes of L. monocytogenes under lactic acid treatment.
Tn-seq, a high-throughput tool for the functional genomic study of foodborne pathogens, has previously been reported in detail (21). In this study, we used this technique to perform a genome-wide identification of conditionally essential genes of L. monocytogenes under lactic acid treatment. The procedure was similar to that described previously (36). In brief, aliquots containing approximately 107 CFU from the L. monocytogenes mutant pool were used to inoculate 100 mL of BHI broth or BHI broth supplemented with 1 mg/mL lactic acid (Aladdin, Shanghai, China). After incubation at 37°C for 16 h, 1 mL culture was used to extract the genomic DNA using the genomic DNA extraction kit (NUOWEIZAN, Nanjing, China). Next, further processing steps (including fragmentation, adapter connection, PCR amplification, etc.) were carried out as previously described (38). Then, high-throughput sequencing was performed on an Illumina HiSeq PE150 instrument (Personalbio, Shanghai, China). This experiment was performed in triplicate.
Bioinformatics analysis of Tn-seq data.
Tn-seq data analysis was conducted in a previous study (22). After Illumina sequencing, the raw reads were split based on barcodes using Fastp (39), and 16-bp nucleotide fragments corresponding to each reading of the L. monocytogenes sequence were mapped to the L. monocytogenes genome using Bowtie 2 (40). Then, Integrative Genomics Viewer (IGV) was used to sort and count the results of the alignment. Next, the read counts of each gene were normalized using the following formula: RPTAM (reads per TA sites per million input reads) = (number of reads mapped to a gene × 106)/(total mapped input reads in the sample × number of TA sites in this gene). Cyber-T (41) (http://cybert.ics.uci.edu/) was used for statistical analysis of RPTAM values between different groups. Genes exhibiting a BH value (Benjamini-Hochberg corrected P value) of <0.05 were determined to be statistically significant.
Determination of growth curves of the wild type and Δlmo2248.
A microplate reader (BioTek ELX808IU) was used to determine the effects of lactic acid on bacterial growth. The wild type and the mutant (Δlmo2248) were grown overnight in BHI. Bacterial cells were inoculated at an initial OD630 of 0.005 into 200 μL BHI and BHI with 1 mg/mL of lactic acid, respectively. The cultures were incubated in the microplate reader at 37°C, and the absorbance at 630 nm (A630) was recorded every hour for 22 h. Each experiment was carried out in triplicate.
Overnight cultures of L. monocytogenes, Δlmo2248, and Δlmo2248+ lmo2248 were diluted 100-fold and inoculated into the fresh BHI broth. Next, 2 mL of the cell cultures (exponential phase) were spun down and resuspended in PBS to an OD600 of 1. Then the bacterial solution was serially diluted in a 96-well plate, and 10 μL was added to the agar plate supplemented with LA (2 mg/mL).
SEM and TEM.
Bacterial cells (L. monocytogenes and Δlmo2248) were cultured in BHI to the logarithmic growth stage (OD600, 0.8) and then treated with 4 mg/mL lactic acid at 37°C for 2 h. Next, the samples were fixed overnight at 4°C in 2.5% glutaraldehyde solution. After a series of pretreatments as previously described (35), the samples were sent to the bio-ultrastructure analysis Lab of Zhejiang University for scanning electron microscopy (GeminiSEM 300, Oberkochen, Germany) and transmission electron microscopy (Hitachi H-7650, Tokyo, Japan) observations.
Transcriptome profiling.
L. monocytogenes was incubated in BHI broth and BHI broth supplemented with 2 mg/mL lactic acid for 16 h. Bacterial cells were centrifuged at 4,000 rpm for 5 min at room temperature and snap-frozen in liquid nitrogen. RNA extraction and quantification were performed as described in previous studies (1, 35). Then, the amplified and purified RNA-seq libraries were sequenced on the NovaSeq 6000 platform (Illumina, San Diego, CA, USA) and 150-bp paired-end reads were generated. Differentially expressed gene (DEG) analysis and enrichment analysis (Gene Ontology [GO] and Kyoto Encyclopedia of Genes and Genomes [KEGG] enrichment analyses) were performed according to the method described previously (35).
Validation of transcriptome results by qPCR analysis.
The cDNA was synthesized according to the instructions for the PrimeScript RT reagent kit with gDNA Eraser (TaKaRa, Beijing, China). Then, qPCR was conducted using the TB Green premix Ex Taq (Tli RNaseH Plus) kit (Thermo Fisher Scientific, Waltham, MA, USA) and the Applied Biosystems QuantStudio 3 instrument (Thermo Fisher Scientific). Transcript levels, relative to drm, of the assayed genes were calculated using QuantStudio Design & Analysis Software 1.3.1 (Thermo Fisher Scientific). Data analysis was performed using the 2−ΔΔCT method. Each group contained 3 biological replicates.
Statistical analysis.
All analyses of significance were performed using one-way analysis of variance (ANOVA) and Duncan’s multiple-range test (DMRT). In all analyses, only when P was <0.05, were data considered statistically significant. All tests were conducted in triplicate.
Data availability.
The raw RNA-seq data are available at the NCBI Sequence Read Archive (SRA) under BioProject accession no. PRJNA862260, and the Tn-seq data are available at NCBI SRA under BioProject accession no. PRJNA862615.
Supplementary Material
ACKNOWLEDGMENTS
This work was supported by the Natural Science Foundation of Shandong Province, China (ZR2019ZD21), the Taishan Scholars Program of Shandong Province, China (ts20190955), the Project of Shandong Province Higher Educational Outstanding Youth Innovation Team (2019KJF011), and the National Key Research and Development Program of China (2019YFE0103900).
We declare that we have no conflict of interest.
Footnotes
Supplemental material is available online only.
Contributor Information
Jianrui Niu, Email: niujianrui@lyu.edu.cn.
Xinglin Zhang, Email: zhangxinglin@lyu.edu.cn.
Luxin Wang, University of California Davis.
Augustine Agyekum, University of Georgia.
REFERENCES
- 1.Liu X, Shu Q, Chen Q, Pang X, Wu Y, Zhou W, Wu Y, Niu J, Zhang X. 2020. Antibacterial efficacy and mechanism of mannosylerythritol lipids-A on Listeria monocytogenes. Molecules 25:4857. doi: 10.3390/molecules25204857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zhu Q, Gooneratne R, Hussain MA. 2017. Listeria monocytogenes in fresh produce: outbreaks, prevalence and contamination levels. Foods 6:21. doi: 10.3390/foods6030021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Radoshevich L, Cossart P. 2018. Listeria monocytogenes: towards a complete picture of its physiology and pathogenesis. Nat Rev Microbiol 16:32–46. doi: 10.1038/nrmicro.2017.126. [DOI] [PubMed] [Google Scholar]
- 4.European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC). 2018. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J 16:e05500. doi: 10.2903/j.efsa.2018.5500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Boelaert F, Stoicescu A, Amore G, Messens W, Hempen M, Rizzi V, Antoniou S-E, Baldinelli F, Dorbek-Kolin E, Van der Stede Y, Niskanen T, Haussig J, Kaczmarek M, Dias JG, Barco L, Mancin M, Mantovani C, Sardella A, Antonelli P, Leati M, Lettini AA, Losasso C, Morabito S, Scavia G, Knijn A, Tozzoli R, Iacoponi F, Moro O, D'Errico ML, Gattuso A, Suffredini E, Di Bartolo I, Delibato E, Anniballi F, Ianiro G, Altieri I, Morales MAG, Casulli A, Caccio S, Danan C, Felix B, European Food Safety A, European Ctr Dis P . 2021. The European Union One Health 2019 Zoonoses Report. EFSA J 19:6406. doi: 10.2903/j.efsa.2021.6406. [DOI] [Google Scholar]
- 6.Capita R, Alonso-Calleja C. 2013. Antibiotic-resistant bacteria: a challenge for the food industry. Crit Rev Food Sci Nutr 53:11–48. doi: 10.1080/10408398.2010.519837. [DOI] [PubMed] [Google Scholar]
- 7.Gyawali R, Ibrahim SA. 2014. Natural products as antimicrobial agents. Food Control 46:412–429. doi: 10.1016/j.foodcont.2014.05.047. [DOI] [Google Scholar]
- 8.Chibane LB, Degraeve P, Ferhout H, Bouajila J, Oulahal N. 2019. Plant antimicrobial polyphenols as potential natural food preservatives. J Sci Food Agric 99:1457–1474. doi: 10.1002/jsfa.9357. [DOI] [PubMed] [Google Scholar]
- 9.Mani-López E, García HS, López-Malo A. 2012. Organic acids as antimicrobials to control Salmonella in meat and poultry products. Food Res International 45:713–721. doi: 10.1016/j.foodres.2011.04.043. [DOI] [Google Scholar]
- 10.Wang C, Chang T, Yang H, Cui M. 2015. Antibacterial mechanism of lactic acid on physiological and morphological properties of Salmonella Enteritidis, Escherichia coli and Listeria monocytogenes. Food Control 47:231–236. doi: 10.1016/j.foodcont.2014.06.034. [DOI] [Google Scholar]
- 11.EFSA Panel on Biological Hazards (BIOHAZ). 2011. Scientific opinion on the evaluation of the safety and efficacy of lactic acid for the removal of microbial surface contamination of beef carcasses, cuts and trimmings. EFSA J 9:2317. doi: 10.2903/j.efsa.2011.2317. [DOI] [Google Scholar]
- 12.Ning Y, Fu Y, Hou L, Ma M, Wang Z, Li X, Jia Y. 2021. iTRAQ-based quantitative proteomic analysis of synergistic antibacterial mechanism of phenyllactic acid and lactic acid against Bacillus cereus. Food Res Int 139:109562. doi: 10.1016/j.foodres.2020.109562. [DOI] [PubMed] [Google Scholar]
- 13.Yu L, Zhang S, Xu Y, Mi X, Xing T, Li J, Zhang L, Gao F, Jiang Y. 2021. Acid resistance of E. coli O157:H7 and O26:H11 exposure to lactic acid revealed by transcriptomic analysis. LWT 136:110352. doi: 10.1016/j.lwt.2020.110352. [DOI] [Google Scholar]
- 14.Foster JW. 1995. Low pH adaptation and the acid tolerance response of salmonella Typhimurium. Crit Rev Microbiol 21:215–237. doi: 10.3109/10408419509113541. [DOI] [PubMed] [Google Scholar]
- 15.Ryan S, Hill C, Gahan CGM. 2008. Acid stress responses in Listeria monocytogenes. Adv Appl Microbiol 65:67–91. doi: 10.1016/S0065-2164(08)00603-5. [DOI] [PubMed] [Google Scholar]
- 16.Smith JL, Liu Y, Paoli GC. 2013. How does Listeria monocytogenes combat acid conditions? Can J Microbiol 59:141–152. doi: 10.1139/cjm-2012-0392. [DOI] [PubMed] [Google Scholar]
- 17.Begley M, Hill C, Gahan CGM. 2003. Identification and disruption of btlA, a locus involved in bile tolerance and general stress resistance in Listeria monocytogenes. FEMS Microbiol Lett 218:31–38. doi: 10.1111/j.1574-6968.2003.tb11494.x. [DOI] [PubMed] [Google Scholar]
- 18.Madeo M, O’Riordan N, Fuchs TM, Utratna M, Karatzas KAG, O’Byrne CP. 2012. Thiamine plays a critical role in the acid tolerance of Listeria monocytogenes. FEMS Microbiol Lett 326:137–143. doi: 10.1111/j.1574-6968.2011.02442.x. [DOI] [PubMed] [Google Scholar]
- 19.van Opijnen T, Bodi KL, Camilli A. 2009. Tn-seq: high-throughput parallel sequencing for fitness and genetic interaction studies in microorganisms. Nat Methods 6:767–772. doi: 10.1038/nmeth.1377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cain AK, Barquist L, Goodman AL, Paulsen IT, Parkhill J, van Opijnen T. 2020. A decade of advances in transposon-insertion sequencing. Nat Rev Genet 21:526–540. doi: 10.1038/s41576-020-0244-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Liu X, Liu G, Wu Y, Pang X, Wu Y, Niu J, Chen Q, Zhang X. 2021. Transposon sequencing: a powerful tool for the functional genomic study of food-borne pathogens. Trends Food Sci Technol 118:679–687. doi: 10.1016/j.tifs.2021.06.032. [DOI] [Google Scholar]
- 22.Zhang X, de Maat V, Prieto AMG, Prajsnar TK, Bayjanov JR, de Been M, Rogers MRC, Bonten MJM, Mesnage S, Willems RJL, van Schaik W. 2017. RNA-seq and Tn-seq reveal fitness determinants of vancomycin-resistant Enterococcus faecium during growth in human serum. BMC Genomics 18:893. doi: 10.1186/s12864-017-4299-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jayeola V, McClelland M, Porwollik S, Chu W, Farber J, Kathariou S. 2020. Identification of novel genes mediating survival of Salmonella on low-moisture foods via transposon sequencing analysis. Front Microbiol 11:726. doi: 10.3389/fmicb.2020.00726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jana B, Cain AK, Doerrler WT, Boinett CJ, Fookes MC, Parkhill J, Guardabassi L. 2017. The secondary resistome of multidrug-resistant Klebsiella pneumoniae. Sci Rep 7:42483. doi: 10.1038/srep42483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Boinett CJ, Cain AK, Hawkey J, Do Hoang NT, Khanh NNT, Thanh DP, Dordel J, Campbell JI, Lan NPH, Mayho M, Langridge GC, Hadfield J, Chau NVV, Thwaites GE, Parkhill J, Thomson NR, Holt KE, Baker S. 2019. Clinical and laboratory-induced colistin-resistance mechanisms in Acinetobacter baumannii. Microb Genom 5:e000246. doi: 10.1099/mgen.0.000246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Coe KA, Lee W, Stone MC, Komazin-Meredith G, Meredith TC, Grad YH, Walker S. 2019. Multi-strain Tn-Seq reveals common daptomycin resistance determinants in Staphylococcus aureus. PLoS Pathog 15:e1007862. doi: 10.1371/journal.ppat.1007862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sobral RG, Ludovice AM, de Lencastre H, Tomasz A. 2006. Role of murF in cell wall biosynthesis: isolation and characterization of a murF conditional mutant of Staphylococcus aureus. J Bacteriol 188:2543–2553. doi: 10.1128/JB.188.7.2543-2553.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Byelashov OA, Daskalov H, Geornaras I, Kendall PA, Belk KE, Scanga JA, Smith GC, Sofos JN. 2010. Reduction of Listeria monocytogenes on frankfurters treated with lactic acid solutions of various temperatures. Food Microbiol 27:783–790. doi: 10.1016/j.fm.2010.04.003. [DOI] [PubMed] [Google Scholar]
- 29.Gónzalez-Fandos E, Martinez-Laorden A, Perez-Arnedo I. 2021. Efficacy of combinations of lactic acid and potassium sorbate against Listeria monocytogenes in chicken stored under modified atmospheres. Food Microbiol 93:103596. doi: 10.1016/j.fm.2020.103596. [DOI] [PubMed] [Google Scholar]
- 30.Egan AJF, Errington J, Vollmer W. 2020. Regulation of peptidoglycan synthesis and remodelling. Nat Rev Microbiol 18:446–460. doi: 10.1038/s41579-020-0366-3. [DOI] [PubMed] [Google Scholar]
- 31.Schultze T, Hilker R, Mannala GK, Gentil K, Weigel M, Farmani N, Windhorst AC, Goesmann A, Chakraborty T, Hain T. 2015. A detailed view of the intracellular transcriptome of Listeria monocytogenes in murine macrophages using RNA-seq. Front Microbiol 6:1199. doi: 10.3389/fmicb.2015.01199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Schauer K, Geginat G, Liang C, Goebel W, Dandekar T, Fuchs TM. 2010. Deciphering the intracellular metabolism of Listeria monocytogenes by mutant screening and modelling. BMC Genomics 11:573. doi: 10.1186/1471-2164-11-573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Deutscher J, Francke C, Postma PW. 2006. How phosphotransferase system-related protein phosphorylation regulates carbohydrate metabolism in bacteria. Microbiol Mol Biol Rev 70:939–1031. doi: 10.1128/MMBR.00024-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Saier MH, Jr. 2015. The bacterial phosphotransferase system: new frontiers 50 years after its discovery. J Mol Microbiol Biotechnol 25:73–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Liu X, Zhang L, Pang X, Wu Y, Wu Y, Shu Q, Chen Q, Zhang X. 2022. Synergistic antibacterial effect and mechanism of high hydrostatic pressure and mannosylerythritol lipid-A on Listeria monocytogenes. Food Control 135:108797. doi: 10.1016/j.foodcont.2021.108797. [DOI] [Google Scholar]
- 36.Zhang X, Bierschenk D, Top J, Anastasiou I, Bonten MJM, Willems RJL, van Schaik W. 2013. Functional genomic analysis of bile salt resistance in Enterococcus faecium. BMC Genomics 14:299. doi: 10.1186/1471-2164-14-299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wu Y, Pang X, Liu X, Wu Y, Zhang X. 2022. Functional genomics identified novel genes involved in growth at low temperatures in Listeria monocytogenes. Microbiol Spectr 10:e0071022. doi: 10.1128/spectrum.00710-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zhang X, Paganelli FL, Bierschenk D, Kuipers A, Bonten MJM, Willems RJL, van Schaik W. 2012. Genome-wide identification of ampicillin resistance determinants in Enterococcus faecium. PLoS Genet 8:e1002804. doi: 10.1371/journal.pgen.1002804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chen S, Zhou Y, Chen Y, Gu J. 2018. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34:i884–i890. doi: 10.1093/bioinformatics/bty560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Langmead B, Salzberg SL. 2012. Fast gapped-read alignment with Bowtie 2. Nat Methods 9:357–359. doi: 10.1038/nmeth.1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kayala MA, Baldi P. 2012. Cyber-T web server: differential analysis of high-throughput data. Nucleic Acids Res 40:W553–W559. doi: 10.1093/nar/gks420. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. Download spectrum.02750-22-s0001.pdf, PDF file, 0.05 MB (55.5KB, pdf)
Data Availability Statement
The raw RNA-seq data are available at the NCBI Sequence Read Archive (SRA) under BioProject accession no. PRJNA862260, and the Tn-seq data are available at NCBI SRA under BioProject accession no. PRJNA862615.