

Abstract
The EFSA Panel on Plant Health conducted a pest categorisation of cowpea mosaic virus (CPMV) for the EU territory. The identity of CPMV, a member of the genus Comovirus (family Secoviridae), is established and detection and identification methods are available. The pathogen is not included in the Commission Implementing Regulation (EU) 2019/2072. It has been reported from the Americas, and several countries in Africa and Asia and it is not known to be present in the EU in natural conditions. CPMV is considered a major pathogen of cowpea on which it causes symptoms ranging from mild to severe mosaic, chlorosis and necrosis. The virus has been reported sporadically on some other cultivated species of the family Fabaceae, including soybean and some common bean varieties. CPMV is transmitted by cowpea seeds, with uncertainty on the transmission rate. There is uncertainty on seed transmission by other Fabaceae host species due to lack of information. CPMV is also transmitted by several beetle species, one of which, Diabrotica virgifera virgifera, is present in the EU. Seeds for sowing of cowpea are identified as the major entry pathway. The cultivated area and production of cowpea in the EU territory are mainly limited to local varieties cultivated at a small scale in Mediterranean EU Member States. Should the pest establish in the EU, an impact is expected on cowpea crops at local scale. There is high uncertainty on the potential impact that CPMV would cause on other natural hosts cultivated in the EU due to the lack of information from the areas of CPMV's current distribution. Despite the uncertainty concerning the potential impact on bean and soybean crops in the EU, CPMV satisfies the criteria that are within the remit of EFSA to assess for it to be regarded as a potential Union quarantine pest.
Keywords: Comovirus, CPMV, Diabrotica virgifera virgifera, Fabaceae, pest risk, plant health, quarantine
1. Introduction
1.1. Background and Terms of Reference as provided by the requestor
1.1.1. Background
The new Plant Health Regulation (EU) 2016/2031, on the protective measures against pests of plants, is applying from 14 December 2019. Conditions are laid down in this legislation in order for pests to qualify for listing as Union quarantine pests, protected zone quarantine pests or Union regulated non‐quarantine pests. The lists of the EU regulated pests together with the associated import or internal movement requirements of commodities are included in Commission Implementing Regulation (EU) 2019/2072. Additionally, as stipulated in the Commission Implementing Regulation 2018/2019, certain commodities are provisionally prohibited to enter in the EU (high risk plants, HRP). EFSA is performing the risk assessment of the dossiers submitted by exporting to the EU countries of the HRP commodities, as stipulated in Commission Implementing Regulation 2018/2018. Furthermore, EFSA has evaluated a number of requests from exporting to the EU countries for derogations from specific EU import requirements.
In line with the principles of the new plant health law, the European Commission with the Member States are discussing monthly the reports of the interceptions and the outbreaks of pests notified by the Member States. Notifications of an imminent danger from pests that may fulfil the conditions for inclusion in the list of the Union quarantine pest are included. Furthermore, EFSA has been performing horizon scanning of media and literature.
As a follow‐up of the above‐mentioned activities (reporting of interceptions and outbreaks, HRP, derogation requests and horizon scanning), a number of pests of concern have been identified. EFSA is requested to provide scientific opinions for these pests, in view of their potential inclusion by the risk manager in the lists of Commission Implementing Regulation (EU) 2019/2072 and the inclusion of specific import requirements for relevant host commodities, when deemed necessary by the risk manager.
1.1.2. Terms of reference
EFSA is requested, pursuant to Article 29(1) of Regulation (EC) No 178/2002, to provide scientific opinions in the field of plant health.
EFSA is requested to deliver 53 pest categorisations for the pests listed in Annex 1A, 1B, 1D and 1E (for more details see mandate M‐2021‐00027 on the Open.EFSA portal). Additionally, EFSA is requested to perform pest categorisations for the pests so far not regulated in the EU, identified as pests potentially associated with a commodity in the commodity risk assessments of the HRP dossiers (Annex 1C; for more details see mandate M‐2021‐00027 on the Open.EFSA portal). Such pest categorisations are needed in the case where there are not available risk assessments for the EU.
When the pests of Annex 1A are qualifying as potential Union quarantine pests, EFSA should proceed to phase 2 risk assessment. The opinions should address entry pathways, spread, establishment, impact and include a risk reduction options analysis.
Additionally, EFSA is requested to develop further the quantitative methodology currently followed for risk assessment, in order to have the possibility to deliver an express risk assessment methodology. Such methodological development should take into account the EFSA Plant Health Panel Guidance on quantitative pest risk assessment and the experience obtained during its implementation for the Union candidate priority pests and for the likelihood of pest freedom at entry for the commodity risk assessment of High Risk Plants.
1.2. Interpretation of the Terms of Reference
Cowpea mosaic virus (CPMV) is one of a number of pests listed in Annex 1D to the Terms of Reference (ToR) to be subject to pest categorisation to determine whether it fulfils the criteria of a potential Union quarantine pest (QP) for the area of the EU excluding Ceuta, Melilla and the outermost regions of Member States referred to in Article 355(1) of the Treaty on the Functioning of the European Union (TFEU), other than Madeira and the Azores, and so inform EU decision‐making as to its appropriateness for potential inclusion in the lists of pests of Commission Implementing Regulation (EU) 2019/2072. If a pest fulfils the criteria to be potentially listed as a Union QP, risk reduction options will be identified.
2. Data and methodologies
2.1. Data
2.1.1. Literature search
A literature search on cowpea mosaic virus was conducted at the beginning of the categorisation in the ISI Web of Science bibliographic database, using the scientific name of the pest as search term. Papers relevant for the pest categorisation were reviewed, and further references and information were obtained from experts, as well as from citations within the references and grey literature.
2.1.2. Database search
Pest information, on host(s) and distribution, was retrieved from the EPPO Global Database, the CABI databases and scientific literature databases as referred above in Section 2.1.1.
Data about the import of commodity types that could potentially provide a pathway for the pest to enter the EU and about the area of hosts grown in the EU were obtained from EUROSTAT (Statistical Office of the European Communities).
The Europhyt and TRACES databases were consulted for pest‐specific notifications on interceptions and outbreaks. Europhyt is a web‐based network run by the Directorate General for Health and Food Safety (DG SANTÉ) of the European Commission as a subproject of PHYSAN (Phyto‐Sanitary Controls) specifically concerned with plant health information. TRACES is the European Commission's multilingual online platform for sanitary and phytosanitary certification required for the importation of animals, animal products, food and feed of non‐animal origin and plants into the European Union, and the intra‐EU trade and EU exports of animals and certain animal products. Up until May 2020, the Europhyt database managed notifications of interceptions of plants or plant products that do not comply with EU legislation, as well as notifications of plant pests detected in the territory of the Member States and the phytosanitary measures taken to eradicate or avoid their spread. The recording of interceptions switched from Europhyt to TRACES in May 2020.
GenBank was searched to determine whether it contained any nucleotide sequences for cowpea mosaic virus which could be used as reference material for molecular diagnosis. GenBank® (www.ncbi.nlm.nih.gov/genbank/) is a comprehensive publicly available database that as of August 2019 (release version 227) contained over 6.25 trillion base pairs from over 1.6 billion nucleotide sequences for 450,000 formally described species (Sayers et al., 2020).
2.2. Methodologies
The Panel performed the pest categorisation for cowpea mosaic virus, following guiding principles and steps presented in the EFSA guidance on quantitative pest risk assessment (EFSA PLH Panel et al., 2018), the EFSA guidance on the use of the weight of evidence approach in scientific assessments (EFSA Scientific Committee, 2017) and the International Standards for Phytosanitary Measures No. 11 (FAO, 2013).
The criteria to be considered when categorising a pest as a potential Union QP is given in Regulation (EU) 2016/2031 Article 3 and Annex I, Section 1 of the Regulation. Table 1 presents the Regulation (EU) 2016/2031 pest categorisation criteria on which the Panel bases its conclusions. In judging whether a criterion is met the Panel uses its best professional judgement (EFSA Scientific Committee, 2017) by integrating a range of evidence from a variety of sources (as presented above in Section 2.1) to reach an informed conclusion as to whether or not a criterion is satisfied.
Table 1.
Pest categorisation criteria under evaluation, as derived from Regulation (EU) 2016/2031 on protective measures against pests of plants (the number of the relevant sections of the pest categorisation is shown in brackets in the first column)
| Criterion of pest categorisation | Criterion in Regulation (EU) 2016/2031 regarding Union quarantine pest (Article 3) |
|---|---|
| Identity of the pest (Section 3.1 ) | Is the identity of the pest clearly defined, or has it been shown to produce consistent symptoms and to be transmissible? |
| Absence/presence of the pest in the EU territory (Section 3.2 ) |
Is the pest present in the EU territory? If present, is the pest in a limited part of the EU or is it scarce, irregular, isolated or present infrequently? If so, the pest is considered to be not widely distributed. |
| Pest potential for entry, establishment and spread in the EU territory (Section 3.4 ) | Is the pest able to enter into, become established in, and spread within, the EU territory? If yes, briefly list the pathways for entry and spread. |
| Potential for consequences in the EU territory (Section 3.5 ) | Would the pests' introduction have an economic or environmental impact on the EU territory? |
|
Available measures (Section 3.6 ) |
Are there measures available to prevent pest entry, establishment, spread or impacts? |
| Conclusion of pest categorisation (Section 4 ) | A statement as to whether (1) all criteria assessed by EFSA above for consideration as a potential quarantine pest were met and (2) if not, which one(s) were not met. |
The Panel's conclusions are formulated respecting its remit and particularly with regard to the principle of separation between risk assessment and risk management (EFSA founding regulation (EU) No 178/2002); therefore, instead of determining whether the pest is likely to have an unacceptable impact, deemed to be a risk management decision, the Panel will present a summary of the observed impacts in the areas where the pest occurs, and make a judgement about potential likely impacts in the EU. Whilst the Panel may quote impacts reported from areas where the pest occurs in monetary terms, the Panel will seek to express potential EU impacts in terms of yield and quality losses and not in monetary terms, in agreement with the EFSA guidance on quantitative pest risk assessment (EFSA PLH Panel, 2018). Article 3 (d) of Regulation (EU) 2016/2031 refers to unacceptable social impact as a criterion for QP status. Assessing social impact is outside the remit of the Panel.
3. Pest categorisation
3.1. Identity and biology of the pest
3.1.1. Identity and taxonomy
Is the identity of the pest clearly defined, or has it been shown to produce consistent symptoms and/or to be transmissible?
Yes, the identity of cowpea mosaic virus is clearly defined. The pathogen has been shown to produce consistent symptoms and to be transmissible.
Cowpea mosaic virus (CPMV) is currently classified in the species Cowpea mosaic virus belonging to the genus Comovirus (subfamily Comovirinae, family Secoviridae, order Picornavirales) (ictv.global/taxonomy). The species name (Cowpea mosaic virus) will be changed to Comovirus vignae according to taxonomy proposal 2022.005P.A.v1.Secoviridae_rename (https://ictv.global/files/proposals/pending?fid=7456#block‐teamplus‐page‐title), which has been approved by the International Committee on Taxonomy of Viruses (ICTV) and whose ratification is expected in spring 2023. CPMV icosahedral viral particles, 28 nm in diameter, are composed of two types of coat protein (CP), the large CP (LCP, 41 kDA) and the small CP (SCP, 23.7 kDA) (Lin et al., 1999). Virions separately encapsidate two genomic RNA components, RNA1 and RNA2. Depending on the encapsidated RNA, virus particles may have different buoyant densities in caesium chloride (CsCl) gradients, resulting in two separate components, the bottom (B) and the middle (M). In addition, a third top (T) component may contain empty particles with no detectable RNA. The T component is widely used for biotechnological applications, mainly for the expression of either peptides or polypeptides in plants or as nanoparticles formed by empty virions as tools for human antitumour therapies (Sainsbury et al., 2010; Beatty and Lewis, 2019; Venkataraman and Hefferon, 2021). The bipartite RNA genome possesses a 5’‐VPg and a 3′‐poly(A) tail. RNA1 and RNA2 consist of 5,889 and 3,841 nucleotides (nt), respectively, excluding the poly(A) tail. They do not share any sequence homology, except for the 5′‐ and 3′‐untranslated regions (UTR), and act as messenger RNAs for the expression of large polyproteins, which undergo proteolytic cleavage. RNA1 encodes a 200‐kDa protein (200K), which is subsequently cleaved to give the final products 32K, 58K, VPg, 24K and 87K, with the latter containing the motifs of the RNA‐dependent RNA polymerase (RdRp). RNA2 encodes two polyproteins, 105K and 95K, due to the leaky scanning of the first AUG, which are both cleaved by the RNA1‐encoded 24 K protease to give a 58K or 48K product, respectively, as well as large capsid protein (LCP) and small capsid protein (SCP). The 48K protein is the cell‐to‐cell movement protein, which is part of the tubular structures trespassing the plasmodesmata of CPMV infected cells (Lomonossoff, 2008). LCP and SCP have also a role in long‐distance movement inside the plant. The 58K protein is involved in RNA2 replication. RNA1 is replication competent at the single cell level, however, it cannot move, nor are virions formed (Fuchs et al., 2022).
The genome of CPMV has been completely sequenced (RefSeqs NC_003549, NC_003550). Moreover, the complete coding sequences of at least three other isolates and additional partial sequences are publicly available in GenBank.
The EPPO code1 (Griessinger and Roy, 2015; EPPO, 2019) for this species is: CPMV00 (EPPO, online).
Synonyms: cowpea mosaic comovirus, cowpea yellow mosaic virus, CPMV.
Cowpea severe mosaic virus was considered in the past as a severe strain of CPMV, but since 1982 it has been classified in the different species Cowpea severe mosaic virus (Matthews, 1982). Therefore, uncertainties exist on which of the two species some of the studies dating before the taxonomic revision refer to.
3.1.2. Biology of the pest
Cowpea mosaic disease was first reported in the USA together with its possible transmission by the chrysomelidae beetle Ceratoma trifurcata (Forster) (Smith, 1924). CPMV was initially isolated in 1959 in Nigeria from cowpea (Vigna unguiculata L.). Its natural host range is limited to a few species in the family Fabaceae (Chant, 1959).
CPMV is transmitted by leaf‐feeding beetles (Coleoptera) in the families Chrysomelidae (leaf beetles), and Curculionidae (weevils). The chrysomelid Ceratoma trifurcata (Foster) was the first suspected vector (Smith, 1924). CPMV isolate MP‐5 (Nigeria) was shown to be transmitted by adults of the chrysomelids Ootheca mutabilis Sahlberg and Paraluperodes quaternus (Fairmaire), and the curculionid Nematocerus acerbus (Faust), with O. mutabilis being the most efficient vector (Whitney and Gilmer, 1974). Ceratoma trifurcata, together with some additional chrysmelids (Diabrotica balteata LeConte, D. undecimpunctata howardi Barber, D. virgifera LeConte and Acalymma vittatum (F.)), have been reported as CPMV vectors (Jansen and Staples, 1971; Tolin et al., 2016). The species D. virgifera was splitted in the two subspecies D. virgifera virgifera Le Conte and D. virgifera zeae Krysan & Smith in 1980, after the identification of D. virgifera as a vector of CPMV (Jansen and Staples, 1971). Although experiments by Jansen and Staples (1971) were performed using beetles from Nebraska where D. virgifera zeae is not known to be present (EFSA PLH Panel, 2019), uncertainties exist on whether both subspecies are able to transmit this virus. The ladybird beetle Epilachna varivestis Mulsant (family Coccinellidae) was reported to be a vector of the severe strain of CPMV (Jansen and Staples, 1970). Due to the reallocation of this strain in a new species (see Section 3.1.1), there is uncertainty on the capability of this beetle to transmit CPMV.
In general, chrysomelid adult beetles can become viruliferous in a short time, with apparently no latency period before transmission, although acquisition and transmission efficiency increase with the extension of the feeding time and depend on the virus‐beetle combination (Bakker, 1975; reviewed by Fulton et al., 1987; Tolin et al., 2016). Virus titre in viruliferous beetles decreases over time (Ghabrial and Schultz, 1983). Retention of comoviruses in the insect depends on the beetle feeding activity on the infected plant and could be extended during diapause or in the absence of host plants (Jansen and Staples, 1971; Tolin et al., 2016). Beetle larvae can acquire and transmit comoviruses, with no evidence that the virus can overcome the pupation stage and be retained in the adult (Fulton et al., 1987; Tolin et al., 2016). In the absence of specific data, uncertainty exists on the transmission of CPMV by juvenile beetle stages and retention of transmissibility over extended periods. Based on experimental evidence, among vector species tested O. mutabilis Sahlb. has been reported to lose the infectivity after 48 h (Chant, 1959), while other viruliferous beetles (C. trifurcata, D. balteata, D. undecimpunctata howardi, D. virgifera, A. vittatum) retained the transmission ability of CPMV up to 10 days (Jansen and Staples, 1971). No virus replication occurs in the vector and no transovarial transmission is observed (Lomonossoff, 2008; Tolin et al., 2016).
CPMV transmission has been reported to be facilitated by the nymphs and adults of the thrips Sericothrips occipitalis Hood and Taeniothrips sjostedti (Tryb.) (Thysanoptera: Thripidae) as well as by the nymphs of the grasshopper Catantops spissus spissus Wlk. and the adults of Zonocerus variegatus F. (family Acrididae, order Orthoptera) (Whitney and Gilmer, 1974), which have been reported only in central Africa so far. However, these studies are associated with uncertainties due to reported experimental failures and a reassessment of the role of thrips in the transmission of CPMV based on artificial inoculation tests and ELISA assays concluded that T. sjostedti is unlikely to significantly contribute to the field spread of CPMV in Nigeria (Allen and Van Damme, 1981). Grasshopper faecal pellets were shown to contain infectious CPMV but their relevance in the natural spread of the virus was not assessed (Whitney and Gilmer, 1974). The stability of the virus and its easy mechanical transmission may favour virus spread by human activities.
Cowpea seed transmission (1%–5%) was reported (van Kammen et al., 2001 citing Gilmer et al., 1974). Subsequently, Thottappilly and Rossel (1987) reported that no seed transmission was observed in seedlings from different cowpea breeding lines experimentally inoculated with CPMV and showing clear symptoms of mosaic. Ladipo (1988) recovered an infectious CPMV isolate (originating from Crotalaria juncea) from flower buds, petals, open flowers, seed coat and cotyledons with embryo axis of green seeds and green pods, but not from seed coat of air‐dried seeds or dried pod tissues of two cowpea cultivars (C20‐55 and Ife Brown). Moreover, no symptoms were observed in seedlings germinated from seeds collected from Crotalaria juncea and cowpea plants infected by CPMV. However, due to the limited number of tested seeds, the author did not exclude that transmission at low rate would escape detection in the reported assay (Ladipo, 1988). In addition, negative results of bioassays seem questionable due to the low sensitivity of the detection method. More recently, about 65% virus infection was reported in seedlings emerged from seeds of cowpea plants infected by CPMV‐egn, an isolate reported in Egypt (Younes et al., 2018; Abd El‐Aziz and Younes, 2019; Abdel Aleem et al., 2019). Therefore, there is uncertainty on CPMV seed transmission rate in cowpea. Experimentally, CPMV is mechanically transmissible to several species of the genus Nicotiana (family Solanaceae) and to several members of the families Fabaceae, Chenopodiaceae and Amaranthaceae (Bock, 1971; Ladipo, 1988).
3.1.3. Host range/species affected
Cowpea (Vigna unguiculata L.) is the main natural host of CPMV (Chant, 1959). Natural infections by this virus have also been reported in soybean (Glycine max; Van Kammen et al., 2001 citing Thottappilly and Rossel, 1985; Cho and Lee, 2003) and soybean CPMV isolates from Nigeria are available in DSMZ collection (Germany) (www.dsmz.de, accessed on 09/11/2022). CPMV has been found in some other cultivated plant species belonging to the Fabaceae family, such as red kidney bean and Jamaica bean (Phaseolus vulgaris; Lloyd Thomas, 1981), sunn hemp (Crotalaria juncea; Ladipo, 1988) and lablab bean (Lablab purpureus; Martin et al., 2014). Due to the limited number of reports, which are generally old and not associated with straightforward molecular data, an uncertainty exists on the reported natural host range of CPMV. Other cultivated plant species in the family Fabaceae have been experimentally infected by CPMV (e.g., Cicer arietinum, Cajanus cajan, Phaseolus vulgaris, Vigna radiata, Trifolium pratense) (Bock, 1971). Members of the families Amaranthaceae (i.e., Gomphrena globosa, Lupinus albus), Chenopodiaceae (i.e. Chenopodium quinoa, C. amaranticolor and C. murale), and Solanaceae (i.e. Nicotiana benthamiana, N. debneyi, N. occidentalis, N. repanda and N. sylvestris) were shown to be experimental CPMV hosts (Bock, 1971; Ladipo, 1988). The wild host Phaseolus lathyroides was found to be naturally infected by CPMV and could represent a natural reservoir of the virus (Alconero and Santiago, 1973; Lima and Nelson, 1977). In addition, pigeon pea (Cajanus cajan), which grows as a perennial plant in the coastal areas of Kenya may serve as a virus natural reservoir (Bock, 1971). Other natural hosts may also exist, as supported by the recent identification of CPMV in a pool of weeds studied by metagenomics approaches in Tanzania (Mwaipopo et al., 2021). A detailed list of natural and experimental hosts of CPMV is reported in Appendix A.
3.1.4. Intraspecific diversity
Complete and partial genomic sequences of several CPMV isolates are available in GenBank.
Generally, CPMV isolates from different geographic areas are serologically related. However, although little systematic work has been done in this respect, some differences in the experimental host range have been observed (van Kammen et al., 2001). Phaseolus lunatus was listed as an experimental host for the Surinam CPMV isolate but not for East African isolates (Agrawal, 1964 quoted in van Kammen et al., 2001; Bock, 1971).
3.1.5. Detection and identification of the pest
Are detection and identification methods available for the pest?
Yes, detection and identification methods are available for cowpea mosaic virus.
A range of diagnostic protocols were developed to detect and/or identify CPMV in host plants. Serological methods include DAS‐ELISA, dot‐blot immunoassay and tissue‐blot immunoassays using specific antisera (Lloyd Thomas, 1981; Abd El‐Aziz and Younes, 2019; Abdel Aleem et al., 2019) some of which are also commercially available. These serological methods allowed the identification of CPMV in different plant parts, such as leaves, flowers, pods and seeds (Abd El‐Aziz and Younes, 2019). Monoclonal antibodies were raised and characterised; however, they are not routinely used in virus diagnosis (Porta et al., 1994). Molecular diagnostic protocols based on reverse transcription (RT)‐PCR mainly aim at the amplification of the SCP or RdRP genes with specific primers, which, however, are not used for survey purposes (Abdel Aleem et al., 2019). When available, electron microscopy is used to visualise CPMV virions and cytopathological alterations (Carette et al., 2000; Pouwels et al., 2002; Abdel Aleem et al., 2019). CPMV was recently identified in wild plants in Tanzania using a next‐generation sequencing approach (Mwaipopo et al., 2021).
3.2. Pest distribution
3.2.1. Pest distribution outside the EU
To date, CPMV has been reported in Africa, Asia and America (see Figure 1). A CPMV isolate from Suriname has been partially characterised (Agrawal, 1964). A report of CPMV from Cuba (Kviéala et al., 1970) is considered as inconclusive (Thottappilly and Rossel, 1992) as the identity of the virus was not confirmed by serological or molecular tests. Details on CPMV worldwide distribution are included in Appendix B.
Figure 1.
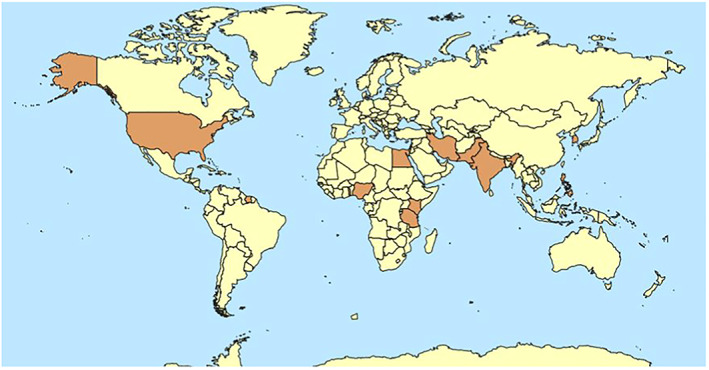
Global distribution of cowpea mosaic virus (CPMV)
3.2.2. Pest distribution in the EU
Is the pest present in the EU territory? If present, is the pest in a limited part of the EU or is it scarce, irregular, isolated or present infrequently? If so, the pest is considered to be not widely distributed.
No, cowpea mosaic virus is not known to be present in the EU territory under natural conditions.
To date, CPMV has not been reported in the EU under natural conditions. However, the virus is used in several EU laboratories.
3.3. Regulatory status
3.3.1. Commission Implementing Regulation 2019/2072
Cowpea mosaic virus is not listed in Annex II of Commission Implementing Regulation (EU) 2019/2072 and implementing acts of Regulation (EU) 2016/2031, or in any emergency plant health legislation.
3.3.2. Hosts or species affected that are prohibited from entering the Union from third countries
None of the natural host plants of cowpea mosaic virus are prohibited from entering the Union from third countries under Commission Implementing Regulation (EU) 2019/2072.
3.3.3. Legislation addressing the organisms that vector CPMV (Commission Implementing Regulation 2019/2072)
Among known vectors of CPMV, Diabrotica undecimpunctata howardi and D. virgifera zeae are QPs included in Annex II, Part A of Commission Implementing Regulation EU 2019/2072, whereas other beetles (O. mutabilis, P. quaternus, N. acerbus, C. trifurcata, D. balteata, D. virgifera virgifera, A. vittatum) are not regulated.
3.4. Entry, establishment and spread in the EU
3.4.1. Entry
Is the pest able to enter into the EU territory? If yes, identify and list the pathways.
Yes, CPMV is able to enter the EU territory through cowpea seeds for sowing, although uncertainty exists on the seed transmission rate. The entry of CPMV into the EU through seeds for sowing of other Fabaceae natural hosts is uncertain because of lack of evidence.
Comment on plants for planting as a pathway.
CPMV hosts are mostly traded as seeds. Therefore, plants for planting other than seeds are unlikely to be a pathway of entry.
The Panel identified cowpea seeds for sowing as the main entry pathway of CPMV in the EU, with uncertainty on seed transmission rate (see Section 3.1.2). Seeds for sowing of other natural hosts have been identified as a potential entry pathway, with uncertainties due to the lack of experimental evidence on seed transmission in these hosts. Green pods and beans as well as dried beans of hosts for feed and food are identified as a minor entry pathway of CPMV: green pods contain immature seeds that are not expected to germinate; although it is unlikely that beans for feed and food will be sown, spillage during unloading operation and transport cannot be excluded.
Although CPMV viruliferous beetles have been reported to retain the transmission capability for up to 10 days (Chant, 1959; Jansen and Staples, 1971), they are not associated with seeds, which is the relevant host traded commodity. Although beetle vectors can be associated with soil during juvenile stages, there is no experimental evidence of CPMV transmission by juvenile stages. Moreover, there is no evidence that the virus can overcome the pupation stage and be retained in the adult of known vectors. Therefore, entry of CPMV through virulent vectors in soil is considered unlikely. In addition, soil not associated with plants is a pathway closed by legislation (Annex VI, 19). Consequently, potential entry pathway associated with the vectors is considered as negligible. Because of the limited virus retention and the low probability of finding the appropriate host, potential entry through hitchhiking of an insect vector is very unlikely.
The Panel notes that CPMV has been used since long in several EU laboratories in absence of quarantine restrictions. Nevertheless, no outbreaks of this virus have been reported in the EU.
CPMV hosts are traded mostly as seeds and therefore, plants for planting other than seeds are unlikely to be a pathway of entry.
There are no prohibitions (Annex VI) or specific requirements (Annex VII) in place regulating CPMV hosts in the EU. Phytosanitary certificate is specifically required for some commodities as indicated in Table 2 where the main potential entry pathway of CPMV is listed.
Table 2.
Potential pathways for cowpea mosaic virus into the EU 27
| Pathways | Life stage | Relevant mitigations (e.g. prohibitions (Annex VI), special requirements (Annex VII) or phytosanitary certificates (Annex XI) within Implementing Regulation 2019/2072) |
|---|---|---|
| Description (e.g. host/intended use/source) | ||
| Seeds for sowing of CPMV hosts | Annex XI (8 and 10) phytosanitary certificates are required for their introduction in the Union territory of Glycine max (seeds for sowing) from all third countries, and of Phaseolus vulgaris (beans for sowing) from third countries other than Switzerland. |
Notifications of interceptions of harmful organisms began to be compiled in Europhyt in May 1994 and in TRACES in May 2020. As of 11 November 2022 and 25 November 2022 there were no records of interception of cowpea mosaic virus in the Europhyt and TRACES databases, respectively. However, since CPMV is not a QP, EU member states have no obligation to notify interceptions of the virus via Europhyt.
3.4.2. Establishment
Is the pest able to become established in the EU territory?
Yes, the virus can potentially establish wherever the hosts are available in the EU.
CPMV could potentially establish in the EU areas where its hosts are present. Natural hosts of CPMV, particularly V. unguiculata, G. max and P. vulgaris are cultivated in the EU. Uncertainty exists on the possibility that some of the experimental hosts may become naturally infected by CPMV. Transfer from an infected plant to other hosts could occur via insect vectors or mechanically. Among the several CPMV insect vectors (Jansen and Staples, 1971; Tolin et al., 2016), the beetle D. virgifera virgifera is present in several EU member states (Figure 2; CABI, 2019) and could contribute to the virus establishment. Whether other vectors may also contribute to the establishment of CPMV in the EU is unknown. Availability of not‐yet identified natural hosts and vectors of CPMV could favour its establishment in the EU.
Figure 2.
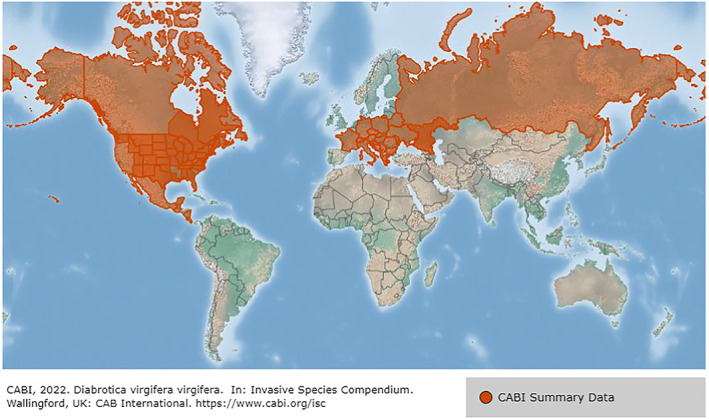
Global distribution map for Diabrotica virgifera virgifera (extracted from the CABI database accessed on 28 October 2022, last updated by CABI on 16 November 2021). Disclaimer: The designations employed and the presentation of material on this map do not imply the expression of any opinion whatsoever on the part of the European Food Safety Authority concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries.
3.4.2.1. EU distribution of main host plants
Cowpea, the major host of CPMV, is cultivated in some EU countries. Details on cowpea production reported in FAOSTAT database in Croatia, Cyprus and Hungary are provided in Table 3. However, cowpea is locally cultivated in several other EU countries, including Italy (Locatelli, 2011), Greece (Lazaridi et al., 2017) and Portugal (Carvalho et al., 2017), where a number of locally adapted populations are conserved at a small scale.
Table 3.
Dried cowpea production (harvested area in ha (production in tonnes)). FAOSTAT database, date of extraction 13 October 2022
| MS/TIME | 2013 | 2014 | 2015 | 2016 | 2017 |
|---|---|---|---|---|---|
| Croatia | 721 (1378) | 678 (1413) | 600 (1346) | 1,534 (3985) | 938 (2347) |
| Cyprus | 210 (235) | 222 (234) | 286 (257) | 153 (196) | 142 (136) |
| Hungary | 1 (1) | 0 | 4 (5) | 4 (7) | 5 (5) |
3.4.2.2. Climatic conditions affecting establishment
Climatic conditions in the EU are not limiting factors for CPMV establishment that rather is dependent on the availability of host plants and vectors in the EU.
3.4.3. Spread
Describe how the pest would be able to spread within the EU territory following establishment.
Following its introduction into the EU, the virus could potentially spread by natural and human‐assisted means.
Comment on plants for planting as a mechanism of spread.
CPMV hosts are traded mostly as seeds. Therefore, plants for planting other than seeds are unlikely to constitute a mechanism of virus spread.
Trade of CPMV infected seeds may spread the virus at long distances. Natural spread in the field is mediated by vectors, one of which (D. virgifera virgifera) is known to be present in the EU territory, namely Austria, Bulgaria, Croatia, Czechia, France, Germany, Greece, Hungary, Italy, Poland, Romania, Slovakia, and Slovenia (CABI, 2019). Although D. virgifera virgifera larvae can only complete development on corn roots, adults of this species, likely flown there from cornfields, have been reported to feed on soybean (Levine et al., 2002), which is a CPMV host (see Section 3.1.3). If the adults of D. virgifera virgifera feed on CPMV infected hosts, they are expected to spread the virus. Whether beetle species already present in the EU may also contribute to virus spread is unknown, as well as there is uncertainty on the role of grasshopper faecal pellets, shown to contain infectious CPMV (Whitney and Gilmer, 1974), or other modes of mechanical transmission in the natural spread of the virus. It is unknown whether wild Fabaceae present in the EU could act as reservoir for CPMV. Following the virus introduction into the EU, both the virus, which is easily mechanically transmissible, and the vectors can be dispersed by human activities. Machinery not appropriately cleaned may carry seeds or viruliferous vectors, thus contributing to the spread of the virus.
3.5. Impacts
Would the pests' introduction have an economic or environmental impact on the EU territory?
Yes, should the virus be introduced in the EU, an economic impact is expected on cowpea production in the EU.
Cowpea plants infected by CPMV show typical mosaic symptoms, which vary in severity, from light green mottle to yellow mosaic. Leaves are distorted, showing significant reduced growth, especially of younger leaves (van Kammen et al., 2001). The number of flowers and pods is also reduced. Premature plant death may follow (Bliss and Robertson, 1971 quoted in Younes et al., 2018). Cowpea cv. California Blackeye shows chlorotic spots with diffuse borders (diam. 1–3 mm) in inoculated primary leaves and a bright yellow or light green mosaic on trifoliolate leaves, moderate distortion, and reduction in size and chlorotic spots on pods (van Kammen et al., 2001). Host plant reactions were described by Hampton et al. (1978) and Ladipo (1988), who reported a CPMV incidence of 1.5% on C. juncea plants showing mosaic, distortion and puckering. In cowpea‐infected fields yield losses occur, ranging from 60% to 100% (Thottappilly and Rossel, 1992). The incidence and impact may depend on the cultivar susceptibility (Thottappilly and Rossel, 1992; van Kammen et al., 2001). In a survey conducted in Nigeria, from 1% to 19% cowpea plants were shown to be infected by CPMV (Shoyinka et al., 1997 quoted in Abdel Aleem et al., 2019). In Egypt, in 2015 and 2016, 6% of 200 cowpea plants showing mosaic, mottling, spots, chlorosis and leaf malformations was reported to be infected by CPMV (Abdel Aleem et al., 2019). CPMV mixed infections with other cowpea infecting viruses were reported in Nigeria (Aliyu et al., 2012). The CPMV impact on cowpea yield can be mitigated by using available resistant or immune cultivars (Beier et al., 1977; van Kammen et al., 2001). Should the virus become established in the EU, an impact can be expected on cowpea production. However, cowpea crops in the EU are mainly restricted to Mediterranean MSs and limited to small‐scale production of locally adapted varieties that contribute to the preservation of cowpea biodiversity. The susceptibility of these local varieties to the virus is not known, generating uncertainty on the magnitude of the impact of CPMV on cowpea in the EU. No outbreaks on bean and soybean crops have been reported in areas where the virus is known to be present. For this reason, the potential impact on other Fabacae hosts species such as bean and soybean crops in the EU is highly uncertain due to lack of information from the areas of CPMV's current distribution.
3.6. Available measures and their limitations
Are there measures available to prevent pest entry, establishment, spread or impacts such that the risk becomes mitigated?
Yes, there are measures available to prevent CPMV entry, establishment, spread and impacts (see Sections 3.3.2, 3.4.1 and 3.6.1).
3.6.1. Identification of potential additional measures
Additional potential risk reduction options and supporting measures are shown in sections 3.6.1.1 and 3.6.1.2.
3.6.1.1. Additional potential risk reduction options
Potential additional control measures are listed in Table 4.
Table 4.
Selected control measures (a full list is available in EFSA PLH Panel et al., 2018) for pest entry/establishment/spread/impact in relation to currently unregulated hosts and pathways. Control measures are measures that have a direct effect on pest abundance
| Control measure/risk reduction option (Blue underline = Zenodo doc, Blue = WIP) | RRO summary | Risk element targeted (entry/establishment/spread/impact) |
|---|---|---|
| Require pest freedom | Use of cowpea seeds for sowing and green pods and beans, and dried beans originating from a country officially free from CPMV or from a pest free area or from a pest free place of production is highly effective. | Entry/spread |
| Crop rotation, associations and density, weed/volunteer control |
Crop rotation, associations and density, weed/volunteer control are used to prevent problems related to pests and are usually applied in various combinations to make the habitat less favourable for pests. The measures deal with (1) allocation of crops to field (over time and space) (multi‐crop, diversity cropping) and (2) to control weeds and volunteers as hosts of pests/vectors. Control of wild hosts and crop rotation may reduce virus inoculum. |
Establishment/spread/impact |
| Use of resistant and tolerant plant species/varieties |
Resistant plants are used to restrict the growth and development of a specified pest and/or the damage they cause when compared to susceptible plant varieties under similar environmental conditions and pest pressure. It is important to distinguish resistant from tolerant species/varieties. Cowpea resistant, tolerant or immune cultivars are available (Beier et al., 1977; van Kammen et al., 2001). |
Entry/establishment/impact |
| Chemical treatments on crops including reproductive material | Chemical treatments against the CPMV vectors are expected to reduce virus spread and disease impact. | Spread/establishment/impact |
| Cleaning and disinfection of facilities, tools and machinery |
The physical and chemical cleaning and disinfection of facilities, tools, machinery, transport means, facilities and other accessories (e.g. boxes, pots, pallets, palox, supports, hand tools). The measures addressed in this information sheet are: washing, sweeping and fumigation. Cleaning measures may remove vector or CPMV‐infected seeds carried over by the machinery. |
Entry/spread |
3.6.1.2. Additional supporting measures
Potential additional supporting measures are listed in Table 5.
Table 5.
Selected supporting measures (a full list is available in EFSA PLH Panel, 2018) in relation to currently unregulated hosts and pathways. Supporting measures are organisational measures or procedures supporting the choice of appropriate risk reduction options that do not directly affect pest abundance
| Supporting measure | Summary | Risk element targeted (entry/establishment/spread/impact) |
|---|---|---|
| Inspection and trapping |
Inspection is defined as the official visual examination of plants, plant products or other regulated articles to determine if pests are present or to determine compliance with phytosanitary regulations (ISPM 5). The effectiveness of sampling and subsequent inspection to detect pests may be enhanced by including trapping and luring techniques. Inspection can be useful to identify early symptoms, when present. |
Entry/establishment/spread |
|
Examination, other than visual, to determine if pests are present using official diagnostic protocols. Diagnostic protocols describe the minimum requirements for reliable diagnosis of regulated pests. Laboratory testing can be performed to assess the possible presence of the pest in its host plants, including seeds. |
Entry | |
| Sampling |
According to ISPM 31, it is usually not feasible to inspect entire consignments, so phytosanitary inspection is performed mainly on samples obtained from a consignment. It is noted that the sampling concepts presented in this standard may also apply to other phytosanitary procedures, notably selection of units for testing. For inspection, testing and/or surveillance purposes the sample may be taken according to a statistically based or a non‐statistical sampling methodology. |
Entry |
| Phytosanitary certificate and plant passport |
An official paper document or its official electronic equivalent, consistent with the model certificates of the IPPC, attesting that a consignment meets phytosanitary import requirements (ISPM 5)
A phytosanitary certification confirming that the seeds for sowing of CPMV host plants are virus‐free could be an effective measure. |
Entry |
| Certification of reproductive material (voluntary/official) |
Plants come from within an approved propagation scheme and are certified pest free (level of infestation) following testing; used to mitigate against pests that are included in a certification scheme. The risk is reduced if seeds for sowing are produced under an approved certification scheme and tested free of the virus. |
Entry/spread |
| Delimitation of Buffer zones |
ISPM 5 defines a buffer zone as “an area surrounding or adjacent to an area officially delimited for phytosanitary purposes in order to minimise the probability of spread of the target pest into or out of the delimited area, and subject to phytosanitary or other control measures, if appropriate” (ISPM 5). The objectives for delimiting a buffer zone can be to prevent spread from the outbreak area and to maintain a pest free production place (PFPP), site (PFPS) or area (PFA). Delimitation of a buffer zone around an outbreak area could potentially prevent the spread of a pest. However, in the case of CPMV, this measure might not be very effective because some of its vectors are polyphagous and move far from the source of infection. |
Spread |
| Surveillance | CPMV is not known to be present in the EU. Surveillance would be an efficient supporting measure. | Spread/establishment |
3.6.1.3. Biological or technical factors limiting the effectiveness of measures
The presence of wild hosts and mechanical transmission by human activities may reduce the efficacy of crop rotation and other measures affecting establishment and spread.
The dispersal potential of vectors could reduce the efficacy of the buffer zone.
4. Uncertainty
Due to the lack of data, there is uncertainty on CPMV seed transmission in Fabaceae natural hosts other than cowpea. Moreover, there is a key uncertainty on the potential impact on bean and soybean crops in the EU due to lack of information on susceptibility of varieties grown in the EU and on outbreaks in these crops in the areas where CPMV is currently present.
5. Conclusions
Despite the uncertainty concerning the potential impact on bean and soybean crops in the EU, CPMV satisfies the criteria that are within the remit of EFSA to assess for it to be regarded as a potential Union QP. Table 6 provides a summary of the PLH Panel conclusions.
Table 6.
The Panel's conclusions on the pest categorisation criteria defined in Regulation (EU) 2016/2031 on protective measures against pests of plants (the number of the relevant sections of the pest categorisation is shown in brackets in the first column)
| Criterion of pest categorisation | Panel's conclusions against criterion in Regulation (EU) 2016/2031 regarding Union quarantine pest | Key uncertainties |
|---|---|---|
| Identity of the pest (Section 3.1 ) | The identity of cowpea mosaic virus is clearly defined. | None |
| Absence/presence of the pest in the EU (Section 3.2 ) | CPMV has not been reported in the EU under natural conditions. However, the virus is used in several EU laboratories. | None |
| Pest potential for entry, establishment and spread in the EU (Section 3.4 ) |
CPMV could enter in the EU with seeds for sowing of CPMV hosts. Potential virus vectors and human activities could contribute to virus establishment and spread. Would the pest enter in the EU it may establish and spread within the EU territory. The Panel notes that CPMV has been used since long in several EU laboratories in absence of quarantine restrictions. Nevertheless, no outbreaks of this virus have been reported in the EU. |
None |
| Potential for consequences in the EU (Section 3.5 ) | Introduction and spread of CPMV could have negative impact on the EU yield and quality production of the cultivated hosts. | Potential impact on bean and soybean crops in the EU |
| Available measures (Section 3.6 ) | Potential control measures are available to mitigate the risk of entry, establishment, spread and impact of CPMV in the EU. | None |
| Conclusion (Section 4 ) | Despite the uncertainty concerning the potential impact on bean and soybean crops in the EU, CPMV satisfies the criteria that are within the remit of EFSA to assess for it to be regarded as a potential Union quarantine pest. | |
| Aspects of assessment to focus on/scenarios to address in future if appropriate: | Information on CPMV seed transmission in Fabaceae hosts other than cowpea, vector transmission and impact on bean and soybean crops | |
Abbreviations
- EPPO
European and Mediterranean Plant Protection Organization
- FAO
Food and Agriculture Organization
- IPPC
International Plant Protection Convention
- ISPM
International Standards for Phytosanitary Measures
- MS
Member State
- PLH
EFSA Panel on Plant Health
- PZ
Protected Zone
- TFEU
Treaty on the Functioning of the European Union
- ToR
Terms of Reference
Glossary
- Containment (of a pest)
Application of phytosanitary measures in and around an infested area to prevent spread of a pest (FAO, 2021)
- Control (of a pest)
Suppression, containment or eradication of a pest population (FAO, 2021)
- Entry (of a pest)
Movement of a pest into an area where it is not yet present, or present but not widely distributed and being officially controlled (FAO, 2021)
- Eradication (of a pest)
Application of phytosanitary measures to eliminate a pest from an area (FAO, 2021)
- Establishment (of a pest)
Perpetuation, for the foreseeable future, of a pest within an area after entry (FAO, 2021)
- Greenhouse
A walk‐in, static, closed place of crop production with a usually translucent outer shell, which allows controlled exchange of material and energy with the surroundings and prevents release of plant protection products (PPPs) into the environment
- Hitchhiker
An organism sheltering or transported accidentally via inanimate pathways including with machinery, shipping containers and vehicles; such organisms are also known as contaminating pests or stowaways (Toy and Newfield, 2010)
- Impact (of a pest)
The impact of the pest on the crop output and quality and on the environment in the occupied spatial units
- Introduction (of a pest)
The entry of a pest resulting in its establishment (FAO, 2021)
- Pathway
Any means that allows the entry or spread of a pest (FAO, 2021)
- Phytosanitary measures
Any legislation, regulation or official procedure having the purpose to prevent the introduction or spread of quarantine pests, or to limit the economic impact of regulated non‐quarantine pests (FAO, 2021)
- Quarantine pest
A pest of potential economic importance to the area endangered thereby and not yet present there, or present but not widely distributed and being officially controlled (FAO, 2021)
- Risk reduction option (RRO)
A measure acting on pest introduction and/or pest spread and/or the magnitude of the biological impact of the pest should the pest be present. A RRO may become a phytosanitary measure, action or procedure according to the decision of the risk manager
- Spread (of a pest)
Expansion of the geographical distribution of a pest within an area (FAO, 2021)
Appendix A – CPMV host plants/species affected
| Host status | Host name | Plant family | Common name | Reference |
|---|---|---|---|---|
| Cultivated hosts | Vigna unguiculata | Fabaceae | Cowpea | Chant (1959) |
| Vigna subterranea | Fabaceae | Bambara groundnut | Thottappilly and Rossel (1987) | |
| Glycine max | Fabaceae | Soybean | Thottappilly and Rossel (1985), Cho and Lee (2003) | |
| Phaseolus vulgaris | Fabaceae | Red kidney bean | Lloyd Thomas (1981) | |
| Jamaica bean | Lloyd Thomas (1981) | |||
| Crotalaria juncea | Fabaceae | Sunn hemp | Ladipo (1988) | |
| Lablab purpureus | Fabaceae | Lablab bean | Martin et al. (2014) | |
| Wild weed hosts | Phaseolus lathyroides | Phasey bean | Lima and Nelson (1977) | |
| Artificial/experimental host | Nicotiana benthamiana | Solanaceae | Lomonossoff (2008) | |
| N. debneyi | Solanaceae | Ladipo (1988) | ||
| N. occidentalis | Solanaceae | Ladipo (1988) | ||
| N. repanda | Solanaceae | Ladipo (1988) | ||
| N. sylvestris | Solanaceae | Ladipo (1988) | ||
| Chenopodium amaranticolor | Chenopodiaceae | Talens and Talens (1979) | ||
| C. quinoa | Chenopodiaceae | Talens and Talens (1979) | ||
| C. murale | Chenopodiaceae | Talens and Talens (1979) | ||
| Cajanus cajan | Fabaceae | Pigeon pea | Bock (1971) | |
| Calapogonium mucunoides | Fabaceae | Calopo | Bock (1971) | |
| Canavalia ensiformis | Fabaceae | Jack bean | Bock (1971) | |
| Cassia occidentalis | Fabaceae | Ladipo (1988) | ||
| Centrosema pubescens | Fabaceae | Butterfly pea | Bock (1971) | |
| Cicer arietinum | Fabaceae | Chickpea | Bock (1971) | |
| Crotolaria intermedia | Fabaceae | Bock (1971) | ||
| C. spectabilis | Fabaceae | Showy rattlepod | Bock (1971) | |
| C. striata | Fabaceae | Bock (1971) | ||
| Desmodium discolor | Fabaceae | Bock (1971) | ||
| D. ovalifolium | Fabaceae | Bock (1971) | ||
| Dolichos biflorus | Fabaceae | Bock (1971) | ||
| Gomphrena globosa | Amaranthaceae | Globe amaranth | Bock (1971) | |
| Lupinus albus | Amaranthaceae | White lupin | Bock (1971) | |
| Mucuna aterrima (syn. Stizolobium deeringianum) | Fabaceae | Bengal velvetbean | Chant (1959), Bock (1971) | |
| Phaseolus vulgaris | Fabaceae | Bean | Talens and Talens (1979) | |
| Phaseolus lunatus | Fabaceae | Lima bean | Agrawal (1964) quoted in van Kammen et al. (2001), Bock (1971) | |
| P. radiatus (syn. Vigna radiata) | Fabaceae | Mung bean | Bock (1971) | |
| P. mungo (syn. Vigna mungo) | Fabaceae | Bock (1971) | ||
| Trifolium pratense | Fabaceae | Red clover | Bock (1971) |
Appendix B – Distribution of cowpea mosaic virus
| Region | Country | Status | Reference |
|---|---|---|---|
| North America | USA | Present | CABI (2019), Martin et al. (2014) |
| South America | Suriname | Agrawal (1964) | |
| Africa | Egypt | Abdel Aleem et al. (2019) | |
| Kenya | Bock (1971) | ||
| Nigeria | Present | CABI (2019) | |
| Togo | Present | CABI (2019) | |
| Tanzania | Mwaipopo et al. (2021) | ||
| Asia | India | Present | CABI (2019) |
| Iran | Ghorbani et al. (2008) | ||
| Philippines | Talens and Talens (1979) | ||
| Pakistan | Present | CABI (2019) | |
| Korea | Cho and Lee (2003) |
Suggested citation: EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard C, Baptista P, Chatzivassiliou E, Gonthier P, Jaques Miret JA, Justesen AF, MacLeod A, Magnusson CS, Milonas P, Navas‐Cortes JA, Parnell S, Potting R, Stefani E, Thulke HH, Van der Werf W, Civera AV, Yuen J, Zappalà L, Streissl F, Carluccio AV, Chiumenti M, Di Serio F, Rubino L and Reignault PL, 2023. Scientific Opinion on the pest categorisation of cowpea mosaic virus. EFSA Journal 2023;21(2):7847, 22 pp. 10.2903/j.efsa.2023.7847
Requestor: European Commission
Question number: EFSA‐Q‐2022‐00392
Panel members: Claude Bragard, Paula Baptista, Elisavet Chatzivassiliou, Francesco Di Serio, Paolo Gonthier, Josep Anton Jaques Miret, Annemarie Fejer Justesen, Alan MacLeod, Christer Sven Magnusson, Panagiotis Milonas, Juan A Navas‐Cortes, Stephen Parnell, Roel Potting, Philippe L Reignault, Emilio Stefani, Hans‐Hermann Thulke, Wopke Van der Werf, Antonio Vicent Civera, Jonathan Yuen and Lucia Zappalà.
Competing interests: In line with EFSA's policy on declarations of interest, Panel member Francesco Di Serio did not participate in the adoption of this scientific output.
Declarations of interest: If you wish to access the declaration of interests of any expert contributing to an EFSA scientific assessment, please contact interestmanagement@efsa.europa.eu.
EFSA may include images or other content for which it does not hold copyright. In such cases, EFSA indicates the copyright holder and users should seek permission to reproduce the content from the original source.
Reproduction of the images listed below is prohibited and permission must be sought directly from the copyright holder: Figure 2: © CABI
Adopted: 26 January 2023
Note
An EPPO code, formerly known as a Bayer code, is a unique identifier linked to the name of a plant or plant pest important in agriculture and plant protection. Codes are based on genus and species names. However, if a scientific name is changed the EPPO code remains the same. This provides a harmonised system to facilitate the management of plant and pest names in computerised databases, as well as data exchange between IT systems (Griessinger and Roy, 2015; EPPO, 2019).
References
- Abd El‐Aziz MH and Younes HA, 2019. Serological and molecular characterization of two seed born cowpea mosaic Comovirus isolates affecting cowpea plants (Vigna unguiculata L.) in northern Egypt. Journal of Plant Science and Phytopathology, 3(3), 86–94. [Google Scholar]
- Abdel Aleem EE, Fattouh FA and Omar SM, 2019. Detection, isolation and characterization of cowpea mosaic virus in Egypt. Journal of Plant Diseases and Protection, 126(6), 599–602. [Google Scholar]
- Agrawal HO, 1964. Identification of cowpea mosaic virus isolates. PhD Thesis, Veenman.
- Alconero R and Santiago A, 1973. Phaseolus lathyroides as a reservoir of cowpea mosaic virus in Puerto Rico. Phytopathology, 63, 120–123. [Google Scholar]
- Aliyu TH, Balogun OS and Kumar L, 2012. Survey of the symptoms and viruses associated with cowpea (Vigna unguiculata L.) in the Agroecological zones of Kwara State, Nigeria. Ethiopian Journal of Environmental Studies and Management, 5(4), 613–619. [Google Scholar]
- Allen DJ and Van Damme P, 1981. On thrips transmission of cowpea (yellow) mosaic virus. Tropical Agriculture, 58, 2. [Google Scholar]
- Bakker W, 1975. Rice yellow mottle virus. lAAB Descr. Plant Viruses No. 149. 4 pp.
- Beatty PH and Lewis JD, 2019. Cowpea mosaic virus nanoparticles for cancer imaging and therapy. Advanced Drug Delivery Reviews, 145, 130–144. [DOI] [PubMed] [Google Scholar]
- Beier H, Siler DJ, Russell ML and Bruening G, 1977. Survey of susceptibility to cowpea mosaic virus among protoplasts and intact plants from Vigna sinensis lines. Phytopathology, 67(7), 917–921. [Google Scholar]
- Bliss FA and Robertson DG, 1971. Genetics of host reaction in Cowpea to Cowpea Yellow Mosaic Virus and Cowpea Mottle Virus 1. Crop Science, 11(2), 258–262. [Google Scholar]
- Bock KR, 1971. Notes on east African plant virus diseases. I. Cowpea mosaic virus. East African Agricultural and Forestry Journal, 37(1), 60–62. [Google Scholar]
- CABI , 2019. Datasheet for Diabrotica virgifera virgifera (western corn rootworm). CABI Crop Protection Compendium. Available online: https://www.cabi.org/cpc/datasheet/18637. Last modified 16th November 2021 [Accessed 28 October 2022].
- Carette JE, Stuiver M, Van Lent J, Wellink J and Van Kammen AB, 2000. Cowpea mosaic virus infection induces a massive proliferation of endoplasmic reticulum but not Golgi membranes and is dependent on de novo membrane synthesis. Journal of virology, 74(14), 6556–6563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carvalho M, Muñoz‐Amatriaín M, Castro I, Lino‐Neto T, Matos M, Egea‐Cortines M, Rosa E, Close T and Carnide V, 2017. Genetic diversity and structure of Iberian Peninsula cowpeas compared to world‐wide cowpea accessions using high density SNP markers. BMC genomics, 18(1), 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chant SR, 1959. Viruses of cowpea, Vigna unguiculata L. (Walp.), in Nigeria. Annals of Applied Biology, 47 3, 565–572. [Google Scholar]
- Cho EK and Lee SH, 2003. Cowpea mosaic virus from Vegetable Soybeans in Korea. The Plant Pathology Journal, 19, 166–170. [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard C, Dehnen-Schmutz K, Di Serio F, Gonthier P, Jacques M-A, Jaques Miret JA, Justesen AF, Magnusson CS, Milonas P, Navas-Cortes JA, Parnell S, Potting R, Reignault PL, Thulke H-H, Van der Werf W, Vicent Civera A, Yuen J, Zappalà L, Czwienczek E and MacLeod A, 2019. Scientific Opinion on the pest categorisation of Diabrotica virgifera zeae . EFSA Journal, 17 11:5858, 29 pp. [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Jeger M, Bragard C, Caffier D, Candresse T, Chatzivassiliou E, Dehnen‐Schmutz K, Gregoire J‐C, Jaques Miret JA, MacLeod A, Navajas Navarro M, Niere B, Parnell S, Potting R, Rafoss T, Rossi V, Urek G, Van Bruggen A, Van Der Werf W, West J, Winter S, Hart A, Schans J, Schrader G, Suffert M, Kertesz V, Kozelska S, Mannino MR, Mosbach‐Schulz O, Pautasso M, Stancanelli G, Tramontini S, Vos S and Gilioli G, 2018. Guidance on quantitative pest risk assessment. EFSA Journal 2018;16(8):5350, 86 pp. 10.2903/j.efsa.2018.5350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA Scientific Committee , Hardy A, Benford D, Halldorsson T, Jeger MJ, Knutsen HK, More S, Naegeli H, Noteborn H, Ockleford C, Ricci A, Rychen G, Schlatter JR, Silano V, Solecki R, Turck D, Benfenati E, Chaudhry QM, Craig P, Frampton G, Greiner M, Hart A, Hogstrand C, Lambre C, Luttik R, Makowski D, Siani A, Wahlstroem H, Aguilera J, Dorne J‐L, Fernandez Dumont A, Hempen M, Valtue~na Martınez S, Martino L, Smeraldi C, Terron A, Georgiadis N and Younes M, 2017. Scientific Opinion on the guidance on the use of the weight of evidence approach in scientific assessments. EFSA Journal 2017;15(8):4971, 69 pp. 10.2903/j.efsa.2017.4971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EPPO , 2019. EPPO codes. Available online: https://www.eppo.int/RESOURCES/eppo_databases/eppo_codes
- FAO (Food and Agriculture Organization of the United Nations) , 2013. ISPM (International Standards for Phytosanitary Measures) 11—Pest Risk Analysis for Quarantine Pests. FAO, Rome, 36 pp. Available online: https://www.ippc.int/sites/default/files/documents/20140512/ispm_11_2013_en_2014-04-30_201405121523-494.65%20KB.pdf [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations) , 2021. International Standards for Phytosanitary Measures. ISPM 5 Glossary of Phytosanitary Terms. FAO, Rome. Available online: https://www.fao.org/3/mc891e/mc891e.pdf [Google Scholar]
- Fuchs M, Hily JM, Petrzik K, Sanfaçon H, Thompson JR, van der Vlugt R and Wetzel T, 2022. ICTV virus taxonomy profile: Secoviridae 2022. Journal of General Virology. 10.1099/jgv.0.001807 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fulton JP, Gergerich RC and Scott HA, 1987. Beetle transmission of plant viruses. Annual review of phytopathology, 25(1), 111–123. [Google Scholar]
- Ghabrial SA and Schultz FJ, 1983. Serological detection of bean pod mottle virus in bean leaf beetles. Phytopathology, 73, 480–483. [Google Scholar]
- Ghorbani S, Shahraein N and Elahinia SA, 2008. Serodiagnosis of cowpea (Vigna unguiculata) viruses in Guilan province, Iran. Iranian Journal of Virology, 2, 30. [Google Scholar]
- Griessinger D and Roy A‐S, 2015. EPPO codes: a brief description. Available online: https://www.eppo.int/media/uploaded_images/RESOURCES/eppo_databases/A4_EPPO_Codes_2018.pdf
- Hampton R, Beczner L, Hagedorn D, Bos L, Inouye T, Barnett O, Musil M and Meiners J, 1978. Host reactions of mechanically transmissible legume viruses of the northern temperate zone. Phytopathology, 68, 989–997. [Google Scholar]
- Jansen WP and Staples R, 1970. Transmission of Cowpea mosaic virus by the Mexican bean beetle. Journal of Economic Entomology, 63(5), 1719–1720. [Google Scholar]
- Jansen WP and Staples R, 1971. Specificity of transmission of cowpea mosaic virus by species within the subfamily Galerucinae, family Chrysomelidae. Journal of Economic Entomology, 64(2), 365–367. [Google Scholar]
- Kviéala BA, Smrz J and Blanco N, 1970. Some properties of Cowpea mosaic virus isolated in Cuba. Phytopathologische Zeitschrift, 69(3), 223–235. [Google Scholar]
- Ladipo JL, 1988. Viruses associated with a mosaic disease of Crotalaria juncea in Nigeria: I. Cowpea mosaic virus. Journal of Phytopathology, 121(1), 8–18. [Google Scholar]
- Lazaridi E, Ntatsi G, Savvas D and Bebeli PJ, 2017. Diversity in cowpea (Vigna unguiculata (L.) Walp.) local populations from Greece. Genetic resources and crop evolution, 64(7),1529–1551. [Google Scholar]
- Levine E, Spencer JL, Isard SA, Onstad DW and Gray ME, 2002. Adaptation of the western corn rootworm to crop rotation: evolution of a new strain in response to a management practice. American Entomologist, 48, 94–107. [Google Scholar]
- Lima JAA and Nelson MR, 1977. Etiology and epidemiology of mosaic of cowpea in Ceara, Brazil. Plant Disease Reporter, 61(10), 864–867. [Google Scholar]
- Lin T, Chen Z, Usha R, Stauffacher CV, Dai JB, Schmidt T and Johnson JE, 1999. The refined crystal structure of cowpea mosaic virus at 2.8 Å resolution. Virology, 265(1), 20–34. [DOI] [PubMed] [Google Scholar]
- Lloyd Thomas OS, 1981. Virus diseases of cultivated plants in Montserrat. International Journal of Pest Management, 27(4), 461–464. [Google Scholar]
- Locatelli A, 2011. Coltivare il fagiolo dall'occhio, legume poco diffuso adatto all'estate. Vita in Campagna, 7–8(2011), 21–23. [Google Scholar]
- Lomonossoff GP, 2008. Cowpea mosaic virus. Encyclopedia of Virology, 2008, 569–574. 10.1016/B978-012374410-4.00562-8 [DOI] [Google Scholar]
- Martin K, Hill JH and Cannon S, 2014. Occurrence and Characterization of Bean common mosaic virus Strain NL1 in Iowa. Plant Disease, 98(11), 1593. [DOI] [PubMed] [Google Scholar]
- Matthews REF, 1982. Classification and nomenclature of viruses. Intervirology, 17(1), 199. [DOI] [PubMed] [Google Scholar]
- Mwaipopo B, Rajamäki ML, Ngowi N, Nchimbi‐Msolla S, Njau PJ, Valkonen JP and Mbanzibwa DR, 2021. Next‐generation sequencing‐based detection of common bean viruses in wild plants from Tanzania and their mechanical transmission to common bean plants. Plant disease, 105(9), 2541–2550. [DOI] [PubMed] [Google Scholar]
- Porta C, Wang G, Cheng H, Chen Z, Baker TS and Johnson JE, 1994. Direct imaging of interactions between an icosahedral virus and conjugate Fab fragments by cryoelectron microscopy and X‐ray crystallography. Virology, 204(2), 777–788. [DOI] [PubMed] [Google Scholar]
- Pouwels J, Carette JE, Van Lent J and Wellink J, 2002. Cowpea mosaic virus: effects on host cell processes. Molecular Plant Pathology, 3(6), 411–418. [DOI] [PubMed] [Google Scholar]
- Sainsbury F, Cañizares MC and Lomonossoff GP, 2010. Cowpea mosaic virus: the plant virus‐based biotechnology workhorse. [DOI] [PubMed]
- Sayers EW, Cavanaugh M, Clark K, Ostell J, Pruitt KD and Karsch‐Mizrachi I, 2020. Genbank. Nucleic Acids Research, 48, Database issue. 10.1093/nar/gkz956 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shoyinka SA, Thottappilly G, Adebayo GG and Anno‐Nyako FO, 1997. Survey on cowpea virus incidence and distribution in Nigeria. International Journal of Pest Management, 43(2), 127–132. [Google Scholar]
- Smith CE, 1924. Transmission of cowpea mosaic by the bean leaf‐beetle. Science, 60(1551), 268. [DOI] [PubMed] [Google Scholar]
- Talens LT and Talens AD, 1979. Cowpea mosaic virus: I. Host range and symptomatology. Philippine. Journal of Crop Science, 4(1),27–36. [Google Scholar]
- Thottappilly G and Rossel HW, 1985. Cowpea research, production and utilization (Singh SR and Rachie KO, eds.). John Wiley & Sons, Chichester. [Google Scholar]
- Thottappilly G and Rossel HW, 1987. Seed transmission of cowpea (yellow) mosaic virus unlikely in cowpea. Tropical Grain Legume Bulletin no. 34, 1987 (IITA).
- Thottappilly G and Rossel HW, 1992. Virus diseases of cowpea in tropical Africa. International Journal of Pest Management, 38(4), 337–348. [Google Scholar]
- Tolin SA, Langham MA and Gergerich RC, 2016. Beetle transmission: A unique alliance of virus, vector, and host. In: Brown JK (ed.). Vector‐Mediated Transmission of Plant Pathogens. The American Phytopathological Society, Saint‐Paul, MN, USA. pp. 131–146. [Google Scholar]
- Toy SJ and Newfield MJ, 2010. The accidental introduction of invasive animals as hitchhikers through inanimate pathways: a New Zealand perspective. Revue scientifique et technique (International Office of Epizootics), 29(1), 123–133. [DOI] [PubMed] [Google Scholar]
- van Kammen A, van Lent J and Wellink J, 2001. Cowpea mosaic virus. AAB Description of Plant Viruses. Available online: https://dpvweb.net/dpv/showdpv/?dpvno=378
- Venkataraman S and Hefferon K, 2021. Application of plant viruses in biotechnology, medicine, and human health. Viruses, 13(9), 1697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitney WK and Gilmer RM, 1974. Insect vectors of cowpea mosaic virus in Nigeria. Annals of Applied Biology, 77(1), 17–21. [DOI] [PubMed] [Google Scholar]
- Younes HA, Aseel DG and El‐Aziz ME, 2018. Identification and purification of cowpea mosaic virus Comovirus isolated from infected cowpea (Vigna unguiculata L.) in northern Egypt. International Journal of Agricultural and Environmental research, 4, 206–212. [Google Scholar]