

Abstract
Alzheimer’s disease (AD) is the most common cause of dementia worldwide. AD brains display deposits of insoluble amyloid plaques consisting mainly of aggregated amyloid-β (Aβ) peptides, and Aβ oligomers are likely a toxic species in AD pathology. AD patients display altered metal homeostasis, and AD plaques show elevated concentrations of metals such as Cu, Fe, and Zn. Yet, the metal chemistry in AD pathology remains unclear. Ni(II) ions are known to interact with Aβ peptides, but the nature and effects of such interactions are unknown. Here, we use numerous biophysical methods—mainly spectroscopy and imaging techniques—to characterize Aβ/Ni(II) interactions in vitro, for different Aβ variants: Aβ(1–40), Aβ(1–40)(H6A, H13A, H14A), Aβ(4–40), and Aβ(1–42). We show for the first time that Ni(II) ions display specific binding to the N-terminal segment of full-length Aβ monomers. Equimolar amounts of Ni(II) ions retard Aβ aggregation and direct it towards non-structured aggregates. The His6, His13, and His14 residues are implicated as binding ligands, and the Ni(II)·Aβ binding affinity is in the low µM range. The redox-active Ni(II) ions induce formation of dityrosine cross-links via redox chemistry, thereby creating covalent Aβ dimers. In aqueous buffer Ni(II) ions promote formation of beta sheet structure in Aβ monomers, while in a membrane-mimicking environment (SDS micelles) coil–coil helix interactions appear to be induced. For SDS-stabilized Aβ oligomers, Ni(II) ions direct the oligomers towards larger sizes and more diverse (heterogeneous) populations. All of these structural rearrangements may be relevant for the Aβ aggregation processes that are involved in AD brain pathology.
Subject terms: Neuroscience, Metals, Peptides, Protein folding, Biochemistry, Biophysical chemistry, Metals, Neurochemistry, Peptides, Protein folding, Proteins, Biophysics, Biological fluorescence, Protein folding, Neurological disorders
Introduction
Alzheimer’s disease (AD), the leading cause of dementia worldwide, is a progressive, irreversible, and currently incurable chronic neurodegenerative disorder1–3, primarily manifesting as short-term memory loss. Pathological hallmarks of AD include brain atrophy, with extensive brain deposits of amyloid plaques and neurofibrillary Tau tangles occurring years before symptom manifestation3–5. The plaques, which consist mainly of amyloid-β (Aβ) peptides aggregated into insoluble fibrils6, display a characteristic cross-β structure at the core of their constituent fibrils7,8. The plaques are the end-product of an aggregation process involving formation of extra- and intracellular intermediates such as neurotoxic Aβ oligomers9–14. The oligomeric aggregates may spread from neuron to neuron via exosomes15,16. However, the relationship between Aβ aggregation, neurodegenerative mechanisms, cognitive decline, the proposed amyloid cascade hypothesis, and disease progression is not fully understood2,13,14,17.
The 36–43 residues long Aβ peptides found in the plaques are produced by enzymatic cleavage of the membrane-binding amyloid-β precursor protein, APP18. In monomeric form, the Aβ peptides are intrinsically disordered and soluble in water. The central and C-terminal segments are hydrophobic and may interact with membranes or fold into a hairpin conformation that likely is required for aggregation19. The negatively charged N-terminal segment is hydrophilic and readily interacts with metal ions and other cationic molecules20–23.
AD brains typically display altered metal homeostasis17,24,25, and AD plaques accumulate metals such as calcium (Ca), copper (Cu), iron (Fe), and zinc (Zn)26–28. Thus, dysregulated metal chemistry might be part of the AD pathology process29–32. The precursor protein APP is known to bind Cu and Zn ions33, and a possible physiological role of APP (and perhaps its fragments) might be to regulate the Cu(II) and Zn(II) concentrations in the neuronal synaptic clefts, where these ions are released in their free form34 and where Aβ aggregation may be initiated35. Metal ions such as Cu(II), Fe(II), Mn(II), Pb(IV) and Zn(II) have previously been shown to bind to specific Aβ residues and modulate the Aβ aggregation pathways20,29,36–40. Binding of metal ions, and also of other cationic molecules such as polyamines, has furthermore been reported to modulate and sometimes inhibit Aβ toxicity21,22,41. However, it is unclear which possible metal interactions may be relevant for AD pathology, and which exogenous or endogenous metal ions may participate in such interactions30–32.
Nickel (Ni) is a common metal in the industrialized world, where it is used in e.g. stainless steel alloys, Ni–Cd batteries, coins, and jewelry. As a result of low-level exposure, between 10 and 20% of all people have developed some degree of contact allergy towards Ni42,43. It is therefore important to clarify the health effects of long-term Ni exposure, including potential effects on neurodegenerative diseases44. Some studies have demonstrated specific binding between Ni(II) ions and N-terminal Aβ fragments, with possible effects on Aβ structure and toxicity41,45–47. Yet, the interactions between pathologically relevant (i.e., full-length) Aβ peptides and Ni ions are poorly explored.
In this study, we use a range of biophysical spectroscopy and imaging techniques to investigate in vitro interactions between Ni(II) ions and Aβ peptides, with a focus on characterizing binding properties and effects on Aβ structure and aggregation. The different Aβ peptides studied include the pathologically relevant Aβ(1–40), Aβ(4–40), and Aβ(1–42) variants, together with the Aβ(1–40)(H6A, H13A, H14A) mutant. Because the Aβ peptides interact with membranes48, and as membrane-disruption is a possible toxicity mechanism for Aβ oligomers30, the measurements have been carried out in aqueous solution as well as in a membrane-mimetic model consisting of micelles of the SDS (sodium dodecyl sulfate) detergent49. The results are compared to previous studies of the effects of both different chemical environments and metal ion interactions on Aβ peptides37,38,40,48–53.
Biological relevance of Ni and sources of exposure
Even though Ni function is limited in human biology54, it has been suggested as an essential element in humans55, just as it is for many human-associated bacteria56 and possibly all higher plants57. Elevated Ni concentrations are however toxic to plants58. In animals, inadequate Ni amounts have shown adverse effects on nutrient absorption and metabolism59,60. Due to the abundance of Ni in many plant-based foods58,61, Ni deficiency is unlikely to occur in humans, even though only some 10% of ingested Ni is absorbed62. Ni concentrations in potable water vary between 2 and 13 mg/L with a WHO limit of 70 mg/L, which is exceeded in Ni mining regions, where Ni concentrations of 200 mg/L have been found63. Respiratory Ni exposure is related to Ni industries and fossil fuel combustion, which overall are the main global sources of Ni emissions58,62, and to cigarette smoking including e-cigarettes38,64. In addition to Ni–Cd batteries, the main industrial uses of Ni is as a whitening agent in Cu alloys for e.g. coins and jewellery, which may cause allergic contact dermatitis58, and as a provider of corrosion resistance in steel alloys for e.g. surgical tools, biomedical implants, and body piercings. One study of Ni-containing hip prosthetic devices found that Ni blood concentration rose about twofold after metal-on-metal hip arthroplasty65. Ni is also present in some formulations for dental amalgam fillings66.
The human health risks of Ni exposure are well-known and widespread, as Ni use and exposure has gradually grown from human prehistory67 to modern times68. Ni is known to be haematotoxic, immunotoxic, neurotoxic, genotoxic, reproductive toxic, pulmonary toxic, nephrotoxic, hepatotoxic and carcinogenic58,69. Other pathological effects of Ni exposure in the occupational settings are rhinitis, asthma, nasal septum perforation, nasal sinus cancer, and respiratory cancer70. Ni can pass the placental barriers and accumulate in the fetus71. It can also pass the blood–brain barrier, and brain accumulation can result from high levels of exposure54. Yet, most people experience low exposure levels66, and the most common health effect is allergic reactions42,43.
The neurotoxic properties of Ni are well documented, but data on Ni in neurodegenerative disorders are scarce44. A case report of skin tissue Ni concentrations in a patient recovering from ALS after metal chelation described a Ni concentration of 850 µg/kg72. A 44 year old ALS patient died after 9 years of heavy metal exposure in a nickel–cadmium battery factory73. Increased blood Ni concentrations have been detected in multiple sclerosis (MS) patients74, and soil Ni was found to be elevated in a Canadian MS cluster75. Recent data also indicate a possible contribution from Ni in the causation of neurodevelopmental dysfunctional states such as autism76.
As a component of tobacco, cigarette smoke, and air pollution, Ni may contribute to environmental risk factors for AD. Mice exposed to a Ni nanoparticle model of air pollution showed doubled brain levels of Aβ40 and Aβ42 within 24 h, even at a permissible limit of nickel hydroxide exposure according to occupational safety and health standards77. Another study reported higher yet not statistically significantly elevated Ni concentrations in post-mortem brain and ventricular fluid of AD patients (n = 14), compared to healthy controls (n = 15)25. A recent study reported that Ni(II) ions interfered with aggregation of the Tau protein78.
Materials and methods
Samples and preparations
Ni(II) acetate and 2-(N-Morpholino)ethanesulfonic acid hydrate (MES) buffer were purchased from Sigma (Sigma/Merck KGaA, Darmstadt, Germany). The SDS detergent was bought from ICN Biomedicals Inc (USA). Sodium chloride and sodium hydroxide were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Wild-type (wt) Aβ(1–42) peptides, abbreviated as Aβ42, with the primary sequence DAEFR5HDSGY10EVHHQ15KLVFF20AEDVG25SNKGA30IIGLM35VGGVV40IA, were purchased synthetically manufactured from JPT Peptide Technologies (Germany), while recombinantly produced Aβ42 peptides were purchased from rPeptide LLC (USA). Recombinantly produced wild-type (wt) Aβ(1–40) peptides, abbreviated as Aβ40, as well as N-terminal truncated Aβ(4–40) peptides, were purchased as lyophilized powder from AlexoTech AB (Umeå, Sweden). The Aβ40 peptides were either unlabeled, uniformly 15N-labeled, or uniformly 13C,15N-labeled. A recombinantly produced mutant version of Aβ40, where the three histidine residues H6, H13, and H14 have been replaced with alanines, i.e. Aβ(1–40)(H6A, H13A, H14A) was also purchased from AlexoTech AB. This mutant is here abbreviated as Aβ40(NoHis). All Aβ variants were stored at − 80 °C until use, when they were dissolved to monomeric form before the measurements. The Aβ40 and Aβ(4–40) peptides were then dissolved in 10 mM NaOH to 100 µM concentration, and sonicated for 5 min in an ice-bath to dissolve possible pre-formed aggregates. Finally, buffer was added to the peptide solutions. All preparation steps were performed on ice, and the peptide concentrations were determined by weighing the dry powder and/or by NanoDrop measurements of dissolved material.
Preparation of Aβ42 oligomers
Monomeric solutions of Aβ42 peptides were prepared via size exclusion chromatography, according to the following procedure. First, lyophilized Aβ42 powder (1 mg) was dissolved in pure dimethyl sulfoxide (DMSO; 250 µL). A solution of 5 mM NaOH (pH = 12.3) was used to equilibrate a Sephadex G-250 HiTrap desalting column (GE Healthcare, Uppsala), which was then washed with 10–15 mL of 5 mM NaOD (pD = 12.7)79. The Aβ42 solution in DMSO was added to the column, followed by 5 mM NaOD (1.25 mL). Peptide fractions in 5 mM NaOD were then collected on ice at a flow rate of 1 mg/mL. Ten fractions of 1 mL were collected in 1.5 mL low-binding reaction tubes. The Aβ42 concentration in each fraction was measured with a NanoDrop instrument (Eppendorf, Germany) at 280 nm, using a molar extinction coefficient of 1280 M−1 cm−1 for the single tyrosine residue in the peptide80. Liquid nitrogen was used to flash-freeze the fractions, which then were topped with argon gas, and stored at − 80 °C until use. Two well-defined sizes of SDS-stabilized Aβ42 oligomers—named according to the SDS concentration used: AβO0.05%SDS (approximately dodecamers) and AβO0.2%SDS (approximately tetramers)—were prepared using an established protocol81 with the following modifications: the preparations were carried out in D2O without the original dilution step, and at a fourfold lower peptide concentration82. The reaction mixtures, consisting of 100 µM Aβ42 peptide in phosphate buffered saline (PBS) buffer containing either 0.05% SDS or 0.2% SDS, which corresponds to 1.7 mM and 6.9 mM SDS, respectively, were incubated at 37 °C for 24 h together with 0–500 µM of Ni(II) acetate. Liquid nitrogen was used to flash-freeze the prepared oligomer solutions, which then were stored at − 20 °C until further use. When thawed at room temperature for experimental analyses, the oligomers were stable for several days.
NMR spectroscopy measurements of Aβ40 binding to Ni(II) ions
1D and 2D nuclear magnetic resonance (NMR) spectra were recorded on Bruker Avance 500 and 700 MHz spectrometers equipped with cryoprobes. First 42 μM and then 84 μM of Ni(II) acetate was added to 84 μM of monomeric Aβ40 peptides, either 13C,15N-double-labelled or 15N-mono-labelled, in 20 mM sodium phosphate buffer at pH 7.3 or pH 5.6 (90/10 H2O/D2O). During the titrations, 2D 1H,15N-HSQC and 2D 1H,13C-HSQC spectra were recorded at 5 °C. Measurements were also conducted in the presence of 50 mM SDS detergent, at 25 °C. As the critical micelle concentration for SDS is 8.2 mM in water at 25 °C83, micelles have clearly formed under these conditions. SDS micelles are simple membrane models suitable for NMR spectroscopy due to their small size, i.e. on average 62 molecules per micelle49. Thus, there is approximately 0.8 mM of SDS micelles in the sample, i.e., around 10× more micelles than Aβ40 peptides, which means that no micelle should harbour multiple Aβ peptides. All NMR data were processed and evaluated using the Topspin software (v. 3.2), employing already published HSQC crosspeak assignments for Aβ40 in buffer84–86 and in the presence of SDS micelles51.
CD spectroscopy measurements of Ni(II)-induced changes in Aβ secondary structure
Circular dichroism (CD) was carried out in a Chirascan CD spectrometer (Applied Photophysics Ltd., U.K.) using a 2 mm quartz cuvette containing 600 µl of Aβ peptide in 20 mM phosphate buffer, pH 7.3. The studied Aβ variants were Aβ40 (10 µM), Aβ40(NoHis) (10 µM), and Aβ(4–40) (5 µM).
Measurements were conducted either in buffer only or with added SDS micelles. CD spectra were recorded at 25 °C between 190 and 260 nm, using steps of 0.5 nm. After the first recorded spectrum, 50 mM SDS detergent was added to some of the samples. Then, small volumes of Ni(II) acetate (2 mM and 10 mM stock solutions) were titrated to each sample, in steps of 2 µM, 4 µM, 16 µM, 56 µM, 156 µM, 256 µM, and finally 512 µM. All data was processed with an eight points smoothing filter (Savitsky-Golay) using the Chirascan Pro-Data v.4.4.1 software (Applied Photophysics Ltd., U.K.).
Binding affinity of Ni(II)·Aβ complexes
Binding affinities for Ni(II)·Aβ complexes were estimated by fitting NMR and CD titration data, respectively, to Eq. (1) (the Morrison equation87):
| 1 |
This equation assumes a single metal binding site, where [Aβ] is the peptide concentration, [Ni] is the concentration of the titrated Ni(II) ions, I0 is the initial signal intensity, I∞ is the steady-state (saturated) signal intensity at the end of the titration series, and KD is the dissociation constant for the Ni(II)·Aβ complex. As no corrections for buffer conditions are done, the computed dissociation constants should be considered as apparent (KDApp).
For the CD data, a binding curve was generated by plotting the CD intensity at 208 nm vs the Ni(II) concentration. For the NMR data, binding curves were generated by plotting HSQC crosspeak intensities versus Ni(II) concentration. The measured crosspeak intensities were normalized to the intensity of the V40 crosspeak for each step of added Ni(II) ions.
Aβ40 aggregation kinetics monitored via ThT fluorescence measurements
The kinetics of amyloid aggregation over time for Aβ40 peptides together with Ni(II) ions was monitored via measurements with a 96-well plate reader (FLUOstar Omega) of the fluorescence signal of the dye Thioflavin T (ThT), a molecular probe that displays strong fluorescence intensity when bound to amyloid material88,89. The samples contained 10 μΜ Aβ40 peptide, 10 mM sodium phosphate buffer, pH 7.4, 40 μΜ ThT dye, and 0, 1, 2.5, 5, 7.5, 10, 20, or 50 μM of Ni(II) acetate. The peptide concentration was determined using a NanoDrop microvolume spectrophotometer. Measurements were recorded at three-minute intervals for 24 h at 37 °C, with five replicate samples per condition and excitation and emission wavelengths of 440 and 480 nm, respectively. The samples were continuously shaken (orbital mode) between the measurements. To determine the maximum growth rate and the half-time of Aβ aggregation, the resulting ThT fluorescence data were fitted to a sigmoidal curve using Eq. (2)90:
| 2 |
where F0 is the baseline fluorescence intensity, A is the total increase in fluorescence intensity, rmax is the maximum growth rate, and t1/2 is the time when half of the Aβ monomers have aggregated.
Atomic force microscopy images of Aβ40 aggregates
Images of Aβ40 aggregates were recorded with a BioScope Catalyst (Bruker Corp., USA) atomic force microscope (AFM), operating in peak force mode in air and using MSLN and SLN cantilevers (Bruker Corp., USA). The scan rate was 0.51 Hz, with a resolution of 512 Å ~ 512 pixels. Samples of 10 μΜ Aβ40 peptide were incubated for 24 h with either 0, 1, 10, or 50 μΜ Ni(II) acetate, using the same conditions as in the ThT experiments described above. At the end of the procedure, 30 μL samples were diluted in 30 μL Milli-Q water and then applied on freshly prepared mica substrates. After 20 min, the mica substrates were washed three times with Milli-Q water and left to air-dry.
Blue native polyacrylamide gel electrophoresis of Aβ42 oligomers
The Aβ42 oligomer samples prepared with 0–500 µM Ni(II) acetate, as described in the materials section, were analyzed with blue native polyacrylamide gel electrophoresis (BN-PAGE) using the Invitrogen electrophoresis system. First, 4–16% Bis–Tris Novex gels (ThermoFisher Scientific, USA) were loaded with Aβ42 oligomer samples (10 μL) in addition to the Amersham High Molecular Weight Calibration Kit for native electrophoresis (GE Healthcare, USA). The gels were run at 4 °C according to the Invitrogen instructions (ThermoFisher Scientific, USA). Staining was done with the Pierce Silver Staining Kit (ThermoFisher Scientific, USA).
Infrared spectroscopy
Fourier-transformed infrared (FTIR) spectra of the Aβ42 oligomers prepared with 0–500 µM Ni(II) acetate, as described in the materials section, were recorded on a Tensor 37 FTIR spectrometer (Bruker Optics, Germany) operating in transmission mode at room temperature and equipped with a liquid nitrogen-cooled MeCdTe detector and a sample shutter. During the measurements, the instrument was continuously purged with dry air. 8–10 µL of the 80 µM Aβ42 oligomer samples were placed between two flat CaF2 discs, which were separated by a 50 µm plastic spacer that had been covered with vacuum grease at the periphery. The mounted IR cuvette was put in a holder inside the sample chamber, and was then allowed to sit for at least 20 min after the chamber lid was closed, to remove H2O vapor. FTIR spectra were recorded between 1900 and 800 cm−1, at a resolution of 2 cm−1 and with 6 mm aperture. The IR intensity above 2200 cm−1 was blocked with a germanium filter, and that below 1500 cm−1 with a cellulose membrane, to increase the light intensity in the relevant spectral range91. The OPUS 5.5 software was used for analysis and plotting of the spectra. Second derivatives were computed with a smoothing factor of 17.
Fluorescence measurements of Ni(II)-induced formation of dityrosine in Aβ40
A Jobin Yvon Horiba Fluorolog 3 fluorescence spectrometer (Longjumeau, France) was used to record fluorescence emission spectra between 330 and 500 nm (excitation at 315 nm) at room temperature of 10 µM Aβ40 peptide dissolved in 20 mM MES buffer, pH 7.3. The samples were put in a quartz cuvette with 4 mm path length (volume 1 mL). To investigate the effect of Ni(II) ions on dityrosine formation, one sample contained 100 µM Ni(II) acetate. The control sample contained 50 µM of the chelator EDTA, to remove possible free metal ions. Spectra were recorded after 0 and 6 h of incubation, where the sample was kept at room temperature without agitation or other treatment. All experiments were conducted in triplicate, and before the final measurement, 300 µM of EDTA was added to the sample to remove metal ions.
Results
NMR spectroscopy: molecular details of Ni(II) binding to the Aβ40 monomer
High-resolution NMR experiments were conducted to investigate possible residue-specific molecular interactions between Ni(II) ions and monomeric Aβ40 peptides. Figures 1 and 2 show 2D 1H,15N-HSQC spectra for the amide crosspeak region for 84 µM 13C,15N-labeled Aβ40 peptides at either pH 7.3 or pH 5.6, recorded before and after addition of first 42 µM and then 84 µM Ni(II) acetate, i.e. Ni(II):Aβ40 ratios of respectively 1:2 and 1:1. Similar measurements were conducted also for Aβ40 peptides together with SDS micelles (50 mM SDS detergent concentration) at pH 7.3 (Fig. 1C).
Figure 1.

NMR 2D 1H,15N-HSQC-spectra of 84 µM 15N-labeled Aβ40 peptides before (blue peaks) and after addition of first 42 µM (red peaks) and then 84 µM (teal peaks) Ni(II) acetate. Spectra were recorded at 5 °C in 20 mM sodium phosphate buffer at pH 7.3 (A), at pH 5.6 (B), and at pH 7.3 together with 50 mM SDS detergent (C). The peak intensity in the bar charts is given as the ratio between the crosspeak intensity with added Ni(II) ions relative to the intensity before addition of Ni(II) ions, i.e. I/I0.
Figure 2.
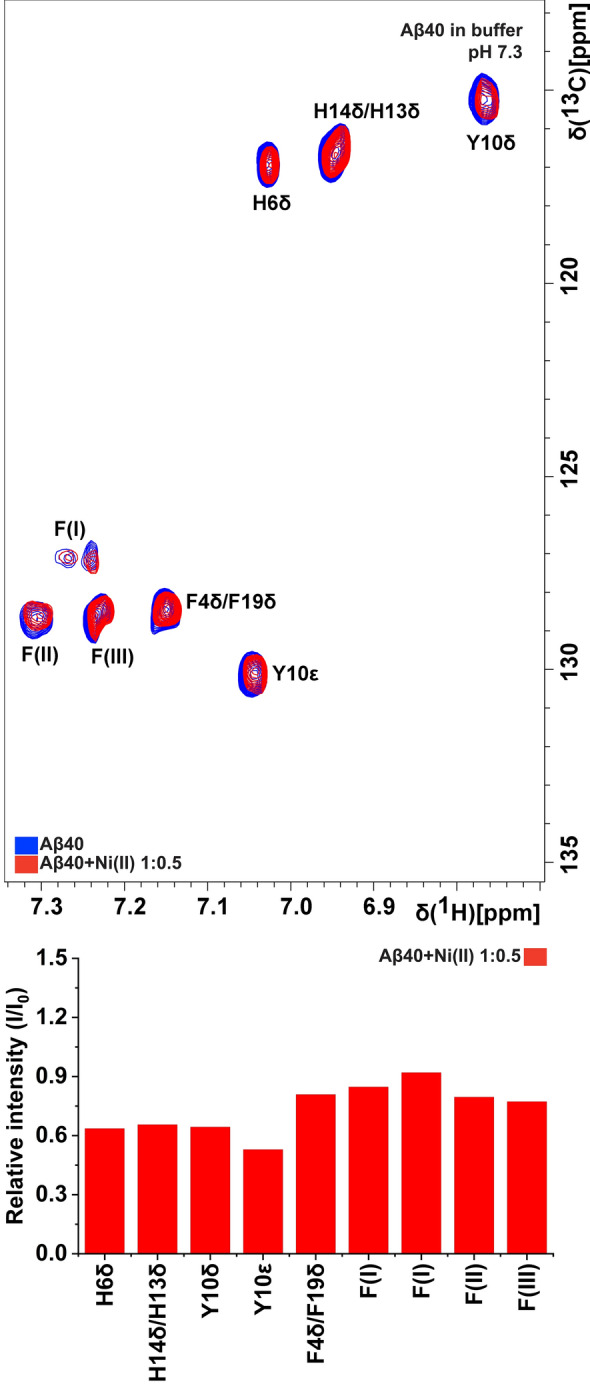
NMR 2D 1H,13C-HSQC spectra of 84 µM 13C,15N-labeled Aβ40 peptides in 20 mM phosphate buffer, pH 7.3, before (blue) and after (red) addition of 42 µM Ni(II) acetate. Some Phe crosspeaks could not be assigned to individual residues, and are instead listed as F(I), F(II), and F(III).
For Aβ40 in aqueous pH 7.3 buffer, addition of Ni(II) ions induces a concentration-dependent loss of amide crosspeak intensity, especially in the N-terminal region (Fig. 1A). This indicates specific binding of Ni(II) ions to N-terminal Aβ40 residues. The specific loss of N-terminal crosspeak intensity is likely caused by intermediate or even slow (on the NMR time-scale) chemical exchange between a free and a bound state of the Aβ40 peptides, similar to the effect induced by Cu(II) and Zn(II) ions40,92, probably together with paramagnetic quenching effects of the Ni(II) ions93. When the Ni(II) ions are added to the sample, no new crosspeaks corresponding to Ni(II)-bound Aβ40 peptides are observed (Fig. 1A). This suggests that no single well-defined Ni(II)·Aβ40 complex exists. Instead, a range of Ni(II)-bound states of the Aβ40 peptides are likely present, probably at different stages of aggregation and oligomerization. Each state is then too weakly populated to create distinct NMR crosspeaks. This is in line with earlier NMR studies reporting that Aβ40 peptides are in a dynamic exchange between NMR-observable monomers and heterogeneous NMR-invisible oligomers (a “dark state”)92,94.
There is also a general loss of crosspeak intensity for all Aβ40 residues, including the central and C-terminal amino acids, upon addition of Ni(II) acetate. This effect is likely caused by a combination of non-specific Ni(II) binding interactions, on an intermediate or slow NMR time-scale, and by the Ni(II) ions promoting aggregation of the Aβ40 peptides into complexes that are too large to be observed with HSQC NMR, or they may simply precipitate out of the solution.
At pH 5.6, the loss of crosspeak intensity after added Ni(II) ions is very uniform, i.e., there is no residue-specific binding (Fig. 1B). The main difference at this lower pH is that histidine residues are protonated, as they have pKa values around 6.8 in short peptides95. Naturally, protonated His residues are less prone to bind cationic metal ions. The loss of residue-specific Ni(II) ion binding at pH 5.6 therefore strongly indicates that histidines are involved in the observed residue-specific Ni(II) ion binding at neutral pH. This is supported by the NMR results at pH 7.3 for the Aβ40 aromatic side chains, where the aromatic rings of the N-terminal residues His6, His13, and His14, together with Tyr10, display a somewhat larger loss of crosspeak signal intensity than the aromatic rings of the Phe4, Phe19, and Phe20 residues (Fig. 2).
Specific loss of crosspeak intensity for N-terminal residues, upon addition of Ni(II) acetate, is observed also for Aβ40 peptides positioned in SDS micelles (Fig. 1C), which constitute a simple model for bio-membranes48,49. The central and C-terminal Aβ regions are known to insert themselves as α-helices into SDS micelles48,51, and thus, the 1H,15N-HSQC spectrum for Aβ40 in SDS micelles corresponds to an α-helical conformation of the Aβ peptide. The N-terminal segment is known to remain unstructured outside the micelle surface, where it is available for interactions with e.g. metal ions96. The C-terminal residues are unaffected by the added Ni(II) ions (Fig. 1C), which shows that they are neither affected by direct Ni(II)-binding, nor by Ni(II)-induced aggregation—most likely Aβ peptides cannot aggregate when bound to SDS micelles, at least when there is on average less than one peptide per micelle. Thus, also in the presence of SDS micelles (Fig. 1C), the NMR spectrum reflects Ni(II) ion binding to monomeric Aβ40 peptides.
CD spectroscopy measurements of Aβ secondary structure
CD spectroscopy was used to investigate possible changes in Aβ secondary structure upon addition of Ni(II) ions, both in aqueous buffer and in a membrane-mimetic environment (i.e., SDS micelles). The three peptide variants Aβ40, Aβ40(NoHis) mutant, and Aβ(4–40) were investigated. In aqueous buffer, the CD spectra for monomers of all three variants display typical random coil signals with minima around 196–198 nm (Fig. 3).
Figure 3.

CD spectra of Ni(II) acetate titrated to Aβ peptides in 20 mM phosphate buffer, pH 7.3, at 25 °C. The titrations were conducted either in the presence of SDS micelles (A–C) or in aqueous buffer only (D–F), for three different peptide variants, i.e. 10 µM Aβ40 (A,D), 10 µM Aβ40(NoHis) (B,E), and 5 µM Aβ(4–40) (C,F). The black spectra show Aβ peptides in buffer only. For samples A–C, 50 mM SDS was then added (red spectra). Next, for all samples, Ni(II) acetate was titrated in steps of 2 µM (orange), 4 µM (yellow), 16 µM (turquoise), 56 µM (green), 156 µM (purple), and finally 256 µM (blue spectra).
Addition of Ni(II) ions to Aβ peptides in aqueous buffer induces concentration-dependent changes in the CD spectra for the monomers of all three peptide variants (Fig. 3D–F). For the Aβ40(NoHis) peptide, these changes correspond to a decrease in intensity, without changing the shape of the spectrum (Fig. 3). This likely means that the Ni(II) ions induce peptide aggregation and precipitation, thereby reducing the effective Aβ concentration in the solution.
For the Aβ40 and Aβ(4–40) variants, addition of Ni(II) ions induces structural transitions between two distinct conformations, as evidenced by the isodichroic points around 210 nm (Fig. 3D,F). Aβ peptides in solution are known to contain some polyproline II (PPII) helix structure, especially at low temperatures50. The loss of signal intensity around 196–198 nm, and the isodichroic points around 210 nm, might be compatible with a conversion of PPII helix into random coil structure50,97,98. However, the difference spectra created by subtracting the CD spectra with no added Ni(II) acetate from those with 256 μM Ni(II) acetate, shown in Supp. Fig. S1, clearly correspond to β-sheet secondary structure99. Formation of β-sheets upon addition of Ni(II) ions is supported also by the IR spectra shown in Supp. Fig. S2. We therefore conclude that Ni(II) ions induce β-sheet structure in Aβ40 and Aβ(4–40) peptides, in aqueous solution at neutral pH.
When SDS micelles were added to the three peptide variants, all of them adopted α-helical secondary structures, producing CD signals with characteristic minima around 208 and 222 nm (Fig. 3A–C). This is consistent with previous studies reporting that Aβ peptides adopt α-helical conformations in membrane-like environments, at least when there is on average less than one peptide per micelle48,49,51,96,100. The Aβ40(NoHis) variant, where the three His residues have been replaced with alanines, displays the strongest α-helical CD signal after addition of SDS, compared to the intensity of the Aβ40(NoHis) random coil signal before adding SDS (Fig. 3). This is reasonable given the strong propensity of alanines to form α-helices101. The Aβ(4–40) peptide also shows a relatively strong α-helical CD signal, compared to the Aβ(4–40) random coil intensity before added SDS. This might be related to a lack of α-helical structure51 in the first three residues of Aβ40, which are missing in the Aβ(4–40) variant.
Addition of Ni(II) ions induces a concentration-dependent loss of CD signal around 208 nm, but not around 222 nm, for the Aβ40 and the Aβ(4–40) peptides, although not for the Aβ40(NoHis) mutant (Fig. 3). As the 222 nm signal intensity remains approximately constant, the observed spectral changes do not correspond to a general loss of α-helicity but may rather be due to a change in helix supercoiling, i.e. when two or more α-helices form coiled coils via hydrophobic interactions102–104. The degree of helix supercoiling is known to be reflected by the [θ222]/[θ208] ratio, where ratios close to 1 reflect large amounts of superhelicity105. During the titrations with Ni(II) acetate the [θ222]/[θ208] ratio increases from 0.70 to 0.89 for Aβ40, and from 0.75 to 1.03 for the Aβ(4–40) variant (Table 1). In both cases this would correspond to a significant increase in superhelicity. Such metal-induced changes of Aβ superhelicity, in a membrane environment, has previously been reported to be induced by Cu(II) ions96. The lack of any Ni(II)-induced structural effects in the Aβ40(NoHis) mutant (Fig. 3; Table 1) appears to support Ni(II) binding to Aβ via the His residues. But it is also possible that the His residues are necessary for forming a coiled-coil Aβ structure.
Table 1.
CD signal intensities at 208 nm and 222 nm for the three Aβ variants Aβ40, Aβ40(NoHis), and Aβ(4–40), as a function of added Ni(II) acetate.
| Wavelength (nm) | 0 µM Ni(II) | 2 µM Ni(II) | 4 µM Ni(II) | 16 µM Ni(II) | 56 µM Ni(II) | 156 µM Ni(II) | 256 µM Ni(II) | |
|---|---|---|---|---|---|---|---|---|
| Aβ40 | 208 | − 11,131 | − 10,808 | − 10,557 | − 9928 | − 9515 | − 9064 | − 9025 |
| 222 | − 7793 | − 7925 | − 8074 | − 8241 | − 8339 | − 8012 | − 8114 | |
| 222/208 | 0.700 | 0.733 | 0.765 | 0.830 | 0.8764 | 0.884 | 0.899 | |
| Aβ40(NoHis) | 208 | − 14,062.7 | − 13,979 | − 14,112.3 | − 13,919.9 | − 13,962.9 | 19,802.9 | 13,725.2 |
| 222 | − 11,184.4 | − 11,100.6 | − 11,249.3 | − 11,138.9 | − 11,172.6 | − 11,149.8 | − 11,039.3 | |
| 222/208 | 0.7953 | 0.7941 | 0.7971 | 0.8002 | 0.8002 | 0.8078 | 0.8043 | |
| Aβ(4–40) | 208 | − 5335.7 | − 5265.9 | − 5258.7 | − 4876.8 | − 4338.2 | − 4235.2 | − 4150.7 |
| 222 | − 3978.0 | − 3989.9 | − 4193.3 | − 4302.0 | − 4205.2 | − 4176.1 | 4268.9 | |
| 222/208 | 0.746 | 0.758 | 0.797 | 0.882 | 0.969 | 0.986 | 1.028 |
Estimates of Aβ·Ni(II) binding affinity
Ni(II)·Aβ binding curves were generated by plotting crosspeak intensities from the NMR titrations (Fig. 1A) and 208 nm intensities from the CD titrations (Table 1) versus Ni(II) concentration (Fig. 4). Fitting Eq. (1) to these curves yields apparent dissociation constants (KD) for the Ni(II)·Aβ complexes.
Figure 4.

Binding curves for Ni(II)·Aβ complexes, derived from the NMR data in Fig. 1A and the CD data in Fig. 3A,C, respectively. The NMR and CD signal intensities are given as the ratio between the intensity with added Ni(II) ions relative to the intensity before addition of Ni(II) ions, i.e. I/I0. Apparent binding affinities were obtained by fitting Eq. (1) to the curves. (A–C) NMR crosspeak intensity vs Ni(II) concentration for three of the crosspeaks/residues in Fig. 1A, i.e. for 84 µM Aβ40 in 20 mM sodium phosphate buffer, pH 7.3 at 5 °C. (D,E) CD intensity at 208 nm vs Ni(II) concentration for the CD data in Fig. 3A,C, i.e. for 10 µM Aβ40 (D) and 5 µM Aβ(4–40) (E) in 20 mM phosphate buffer, pH 7.3 at 25 °C.
For the CD data (Fig. 4D,E), the signal intensities have been normalized to the first value in each titration series, i.e. the signal intensity without added Ni(II) ions. The derived KD values are 7.8 µM for binding to Aβ40, and 17.2 µM for binding to the Aβ(4–40) variant. These KD values should however only be considered as approximations, as there may not be a direct correlation between Ni(II) binding and the structural changes observed in the CD spectra (Fig. 3). Our NMR measurements (Fig. 1C) confirm earlier studies showing that the N-terminal Aβ segment is free to interact with metal ions also when the central and C-terminal Aβ segments are inserted into SDS micelles48,51,96. Yet, earlier studies with Cu(II) ions have shown that the binding affinity for metal ions is reduced when the central Aβ segment is bound to some other entity106, although this effect appears to be minor for SDS micelles96.
For the NMR data, each crosspeak in Fig. 1A generates one binding curve. However, the Ni(II)-induced loss of crosspeak intensity is not only related to concentration-dependent chemical exchange, on an intermediate NMR time-scale, but also to Ni(II)-induced peptide aggregation and paramagnetic quenching of the NMR signal. Even though these effects are somewhat mitigated by normalizing the crosspeak intensity values to the V40 crosspeak intensity, for each titration step (thereby obtaining the relative intensity scale used in Fig. 4A–C), the derived KD values should only be considered as rough approximations. The binding curves shown in Fig. 4A–C correspond to the three Aβ40 crosspeaks that display the strongest apparent binding, with KD values of respectively 5.3 µM, 6.7 µM, and 7.0 µM. These values are very similar to the apparent KD value of 7.8 µM derived for Aβ40 from the CD measurements (Fig. 4D), but they should still only be regarded as lower limits for the true KD value. Thus, we conclude that the binding affinity for the Aβ40·Ni(II) complex is in the low µM range.
Effects of Ni(II) ions on Aβ40 aggregation kinetics
To investigate the influence of Ni(II) ions on Aβ40 aggregation, samples of 10 µM Aβ40 were incubated for 24 h in the absence or presence of increasing concentrations of Ni(II) acetate. The resulting ThT curves are shown in Fig. 5, and the rmax, t1/2, and A parameters obtained from fitting the curves to Eq. (2) are shown in Table 2. For sub-stoichiometric Ni(II):Aβ40 ratios, the Ni(II) ions slow down the Aβ40 aggregation kinetics in a concentration-dependent manner. The aggregation half-time (t1/2) increases with increasing Ni(II) concentrations, i.e. from around 8 h without Ni(II) ions to over 13 h with 10 µM Ni(II) ions. The maximum aggregation rate rmax shows no systematic change with increasing Ni(II) concentration, instead it fluctuates around 0.9 h−1 to 1 h−1 (Table 2). The end-point ThT fluorescence intensities (parameter “A” in Eq. 2) generally decrease with increasing Ni(II) concentrations (Fig. 5), suggesting that less amyloid material (ThT-binding aggregates) is formed when Ni(II) ions are present. Other explanations are however possible, such as binding competition between ThT molecules and Ni(II) ions, or formation of very large Aβ aggregates that may block the transmitted light, or simply precipitate out of the solution. For the samples with high Ni(II) concentrations, i.e. 20 µM and 50 µM, above the stoichiometric Ni(II):Aβ40 ratio, the ThT curves first increase but then decrease back towards the starting value (Fig. 5). Our best explanation for this unusual behaviour is formation of large samples that precipitate, thereby effectively reducing the Aβ40 concentration in the sample. It was not possible to fit Eq. (2) to these data curves.
Figure 5.
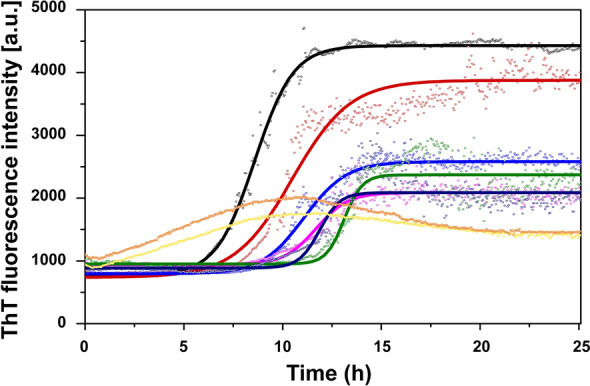
ThT kinetic time curves for aggregation of 10 µM Aβ40, in 10 mM sodium phosphate buffer, pH 7.4, together with different concentrations of Ni(II) acetate: 0 µM (black), 1 µM (red), 2.5 µM (azure blue), 5 µM (magenta), 7.5 µM (green), 10 µM (navy blue), 20 µM (yellow), and 50 µM (orange). The solid lines show curves fitted with Eq. (2) to the ThT data sets.
Table 2.
Parameters rmax, t1/2, and A for aggregation of 10 µM Aβ40 in the presence of different concentrations of Ni(II) acetate.
| Ni(II) | 0 µM | 1 µM | 2.5 µM | 5 µM | 7.5 µM | 10 µM | 20 µM | 50 µM |
|---|---|---|---|---|---|---|---|---|
| rmax [h−1] | 0.96 ± 0.22 | 1.08 ± 0.36 | 0.88 ± 0.34 | 1.0 ± 0.45 | 1.02 ± 0.85 | 0.96 ± 0.58 | n/a | n/a |
| t1/2 [h] | 8.3 ± 1.5 | 10.2 ± 0.3 | 11.8 ± 1.2 | 11.2 ± 0.7 | 12.5 ± 0.5 | 13.5 ± 1.4 | n/a | n/a |
| A | 4115 ± 436 | 2967 ± 238 | 2085 ± 575 | 1449 ± 480 | 1511 ± 132 | 1803 ± 442 | n/a | n/a |
AFM imaging: effects of Ni(II) ions on the morphology of Aβ40 aggregates
To further characterize the influence of Ni(II) ions on Aβ40 fibril morphology, AFM images were recorded on Aβ40 aggregates formed after 24 h incubation in the presence and the absence of Ni(II) (Fig. 6). Without Ni(II) acetate, 10 µM Aβ40 formed typical amyloid fibrils with an apparent height around 4–5 nm (Fig. 2A), which is in line with previously published work on Aβ fibrils formed in vitro107–109. A similar apparent height was observed in the presence of 1 µM Ni(II) ions (Fig. 6B). The presence of 10 µM Ni(II) ions, i.e. a 1:1 Ni(II):Aβ40 ratio, significantly reduces fibril formation: only occasional very short Aβ40 fibril fragments were observed, which display the same height as the fibrils formed by Aβ40 alone (Fig. 6C). In the presence of 50 µM Ni(II) ions no fibrils were present, but instead amorphous clumps of Aβ40 aggregates with variable heights around 13 nm (Fig. 6D). These results are consistent with the concentration-dependent effects of Ni(II) acetate on the Aβ40 aggregation process observed with the ThT measurements (Fig. 5).
Figure 6.

Top row: AFM images of the aggregation products obtained after incubation of 10 µM Aβ40 peptides for 24 h together with either 0, 1, 10, or 50 µM Ni(II) acetate. Bottom row: Representative AFM cross-sections of respectively Aβ40 amyloid fibrils (A,B) and Aβ40 unstructured aggregates (C,D), corresponding to the colored lines shown in the AFM images.
Influence of Ni(II) ions on Aβ42 oligomer formation
PAGE analysis was used to investigate the effect of Ni(II) ions on the formation of Aβ42 oligomers, using a previously published protocol for formation of stable and homogeneous Aβ42 oligomers together with SDS detergent81,82. While most of the current study investigates variants of the Aβ40 peptide, oligomers of Aβ40 are not stable and therefore not suitable model systems. Thus, SEC-purified monomeric solutions of synthetic Aβ42 peptides were mixed with low concentrations of SDS, i.e., below the critical micelle concentration. Incubation of Aβ42 with 0.2% SDS (6.9 mM) leads to formation of mostly tetrameric oligomers (AβO0.2%SDS), while incubation with 0.05% SDS (1.7 mM) produces larger oligomers—predominantly dodecamers (AβO0.05%SDS)81 (Fig. 7, lanes 2 and 6). Figure 7 also shows the effect of different Ni(II) concentrations on oligomer formation. In the absence of Ni(II) ions, the AβO0.05%SDS (Lane 2) and AβO0.2%SDS (Lane 6) oligomers are the dominating species in their respective lanes. With increasing Ni(II) concentration, the band intensity for the major oligomeric structure declines in both cases, while smears towards higher molecular weights appear (Fig. 7, Lanes 3–5 and 7–9). Formation of AβO0.05%SDS is largely disrupted when Ni(II) ions are present at 500 μM concentration (Aβ42:Ni(II) molar ratio of 1:5), while the smear extends over almost the entire length of the lane (Fig. 7, Lane 5). A similar, but less drastic effect is observed for the smaller AβO0.05%SDS (Fig. 7, Lane 9). For both types of SDS-stabilized oligomers, Ni(II) ion concentrations above ~ 100 μM disrupt oligomer formation and more heterogeneous Aβ42 oligomeric solutions containing larger oligomers are produced. A similar smearing effect of Ni(II) ions on the formation of SDS-stabilized Aβ42 oligomers was observed also by SDS-PAGE experiments. These results are shown and discussed in the supplementary information (Fig. S3).
Figure 7.
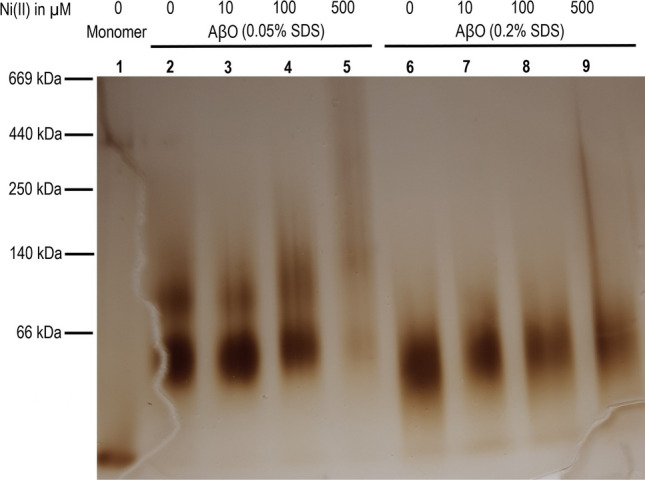
Effects of Ni(II) ions on formation of SDS-stabilized Aβ42 oligomers (AβO0.05%SDS and AβO0.2%SDS) studied by BN-PAGE. Lane 1: Monomers; Lanes 2–5: AβO0.05%SDS oligomers with respectively 0, 10, 100, and 500 µM Ni(II) ions; Lanes 6–9: AβO0.2%SDS oligomers with respectively 0, 10, 100, and 500 µM Ni(II) ions.
FTIR spectroscopy of Aβ42 oligomers formed in the presence of Ni(II) ions
FTIR spectroscopy is a powerful technique for studying the secondary structure of proteins110–116, and can be used to characterize the backbone conformation for different aggregation states of amyloid proteins, including Aβ peptides117–119. Here, the effects of Ni(II) ions on the secondary structures of both AβO0.05%SDS and AβO0.2%SDS oligomers were studied with transmission mode FTIR spectroscopy, using a D2O-based buffer. The results are presented in Fig. 8 as second derivatives of IR absorption spectra, where negative bands indicate the component bands of the absorption spectra. The respective absorbance spectra are shown in Fig. S4 of the Supplementary Information.
Figure 8.

Transmission FTIR data for synthetic AβO0.05%SDS (upper row) and synthetic AβO0.2%SDS (lower row) formed in the presence of different concentrations of Ni(II) ions. The spectra show raw data without normalization. Left: Second derivatives of IR absorbance spectra in the amide I range (1700–1600 cm−1) at zero Ni(II) (red); 1 μM Ni(II) (orange); 10 μM Ni(II)—(green); 100 μM Ni(II) (blue); 500 μM Ni(II) (violet). The black spectrum is for Aβ42 monomers. Right: Dependence on Ni(II) ion concentration for the position (in cm−1) of the main amide I band. For a more clear presentation, the data point at 1 µM was omitted.
For both types of Aβ42 oligomers, two bands are resolved in the amide I region (i.e., 1700–1600 cm−1): a high intensity, low wavenumber band around 1630 cm−1 (the main band for β-sheet structure), and a low intensity, high wavenumber band at 1685 cm−1. This pattern with a split double-band in the amide I region is routinely considered as indicative of the anti-parallel β-sheet structure117,118,120. When Ni(II) acetate is introduced during the Aβ42 oligomer formation reactions, the main band is slightly down-shifted: for AβO0.05%SDS (Fig. 8, upper row) from 1629.2 cm−1 in the absence of Ni(II) acetate to 1628.0 cm−1 at 500 μM of Ni(II) acetate (the highest concentration), and for AβO0.2%SDS (Fig. 8, lower row) from 1630.1 to 1629.6 cm−1 upon addition of Ni(II) acetate up to 500 μM. The downshift is smaller for the oligomers prepared at the higher SDS concentration, indicating that they are less sensitive to Ni(II)-induced effects on the oligomer conformation. The spectral changes observed with Ni(II) are in interesting contrast to the absence of spectral effects upon addition of Li(I) ions121. The downshift is mainly observed in the presence of Ni(II) concentrations between 10 and 100 μM, which agrees with the binding affinities estimated from the CD and NMR results. Detailed analysis of the spectra in Fig. 8 shows that the shifts in band position are associated with a widening of the main β-sheet band on its low wavenumber side, indicating a higher abundance of larger oligomers82. Most of this widening occurs between 10 and 100 μM Ni(II). It further increases between 100 and 500 μM Ni(II), which correlates with the appearance of a high molecular weight smear on the BN-PAGE gel (Fig. 7).
Our previous study on the IR characterization of Aβ42 oligomers has revealed a relationship between oligomer size and position of the main band in the amide I region82. According to these findings, the downshift of the main IR band is associated with an increase in average oligomer size and a concomitant extension of their β-sheet structure. However, the Ni(II)-induced size change is rather modest. The band position of AβO0.2%SDS at the highest Ni(II) ion concentration is still higher than the AβO0.05%SDS band position in the absence of Ni(II) ions, indicating that the oligomers contain less than twelve peptides. Also, the band position of AβO0.05%SDS at 500 µM Ni(II) concentration is considerably higher than that of oligomers formed in the absence of SDS (1623.1 cm−1), which had an average molecular weight of ~ 100 kDa according to Western blotting82. Thus, the dominant β-sheet containing oligomer species of AβO0.05%SDS seem to be smaller than ~ 100 kDa.
The interpretation of the current IR results is in good agreement with the results from the gel electrophoresis experiments, particularly with the BN-PAGE data (Fig. 7). Both IR and PAGE results indicate that addition of Ni(II) ions appears to interfere with the SDS-induced conversion of Aβ42 monomers into homogeneous and stable oligomeric structures, instead favoring formation of larger and more diverse (heterogeneous) oligomer populations.
Ni(II)-induced dityrosine formation
Fluorescence measurements were carried out to investigate if binding of Ni(II) ions could induce formation of covalent dityrosine crosslinks in Aβ peptides, similar to what has been observed for Cu(II) ions122–125. Fluorescence emission spectra for Aβ40 peptides incubated over time with or without Ni(II) acetate are shown in Fig. 9.
Figure 9.
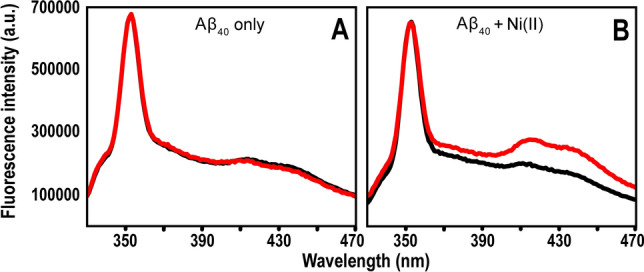
Fluorescence spectra of 10 µM Aβ40 in 20 mM MES buffer, pH 7.3, incubated together with 50 µM EDTA (A) or with 100 µM Ni(II) acetate (B). Black line—0 h; red line—6 h.
The control sample without added Ni(II) ions, which contained 50 µM EDTA to ensure no free metal ions were present, displayed virtually identical spectra before and after 6 h of incubation (Fig. 9A). This is in stark contrast to the sample with 100 µM Ni(II) ions added, where two large new peaks around 410 nm and 435 nm formed during the incubation time (Fig. 9B). The peak around 410 nm is from dityrosine126, while the peak around 435 nm likely is from a different but related system, such as excimers127. These results are not surprising, as nickel is known to be very redox-active. Both the Ni(I)/Ni(II) and Ni(II)/Ni(III) redox pairs could be involved in generating the oxygen radicals required for dityrosine formation128. It should be noted that weak peaks around 410 nm and 435 nm are present in both samples already at time zero (Fig. 9). This shows that some dityrosine cross-links have been generated even before the experiment was initiated, which is consistent with previous reports stating that Aβ peptides can induce oxidative stress on their own, especially in somewhat aggregated states129,130.
Discussion
Nickel is a well-known neurotoxicant, but its role in neurodegenerative diseases remains unclear44. Several studies have investigated the possible effects of transition metals such as Cu and Zn in AD neuropathology, with an emphasis on interactions with the amyloid-forming Aβ peptides29–32. We therefore interpret our current results on Aβ interactions with Ni(II) ions mainly in the light of earlier work on Aβ binding to Cu(II) and Zn(II) ions.
Residue-specific binding of Ni(II) ions to Aβ peptides
Our NMR results show that equimolar amounts of Ni(II) ions display residue-specific binding to the N-terminal segment of the Aβ40 peptide (Fig. 1). This is in line with earlier studies showing that Ni(II) ions can bind N-terminal Aβ fragments45–47. The 2D 1H-13C-HSQC NMR data (Fig. 2) suggest that the three histidine residues His6, His13, and His14 are involved as binding ligands, possibly together with the Tyr10 residue. Previous work has established the metal-binding capacity of the Tyr phenol ring131, and in Aβ peptides the Tyr10 residue seems to be involved in binding to Pb(IV) ions38. The weaker Ni(II)/Aβ40 interactions observed at low pH (Fig. 1) further support the histidines being binding ligands, as these residues become protonated at low pH and therefore less prone to interact with cations92,106,132. The CD spectroscopy measurements also support the histidines being involved as binding ligands: addition of Ni(II) ions induces structural changes in Aβ40 peptides, but not in Aβ40(NoHis) mutant peptides (Fig. 3). This indicates that Ni(II) ions do not bind Aβ peptides when the His residues are absent, which is not surprising, given that Ni(II) ions are known to bind His residues such as those in protein His-tags133. Thus, Ni(II) ions seem to belong to a family of metal ions that coordinate to the Aβ N-terminal segment mainly by the His residues, just like Ag(I), Cu(II), Fe(II), Hg(II), Mn(II), Zn(II), and possibly Pb(IV) ions20,37,38,40,47,52,53,92,134,135. The exact binding coordination could not be determined from our measurements, and it is possible that multiple alternating binding conformations exist, as has been shown for Cu(II) ions136.
According to the Irving-Williams series137, the binding affinities of certain divalent metal ions to peptides and proteins should follow the order Mn(II) < Fe(II) < Co(II) < Ni(II) < Cu(II) > Zn(II). Metal binding affinities are however notoriously difficult to quantify, as they tend to vary both with the experimental conditions (buffer, temperature) and the employed measurement technique138. For example, binding affinities varying by several orders of magnitude have previously been reported for the Aβ·Cu(II) complex, with a consensus value in the low nM region for buffer-corrected affinity138. In our earlier studies, we have reported apparent (not buffer-corrected) KD values around 50–100 µM for Mn(II) ions in phosphate buffer, pH 7.3537, around 1–10 µM for Zn(II) ions in phosphate or Hepes buffer, pH 7.240, and around 0.5 – 2.5 µM for Cu(II) ions in phosphate or Hepes buffer, pH 7.2–pH 7.3540,96,106.
Both the CD and the NMR measurements suggest an affinity in the low µM range for Ni(II) binding to Aβ peptides (Fig. 4), i.e. weaker than Cu(II) ions, stronger than Mn(II) ions, and perhaps somewhat similar to Zn(II) binding affinity, which would be consistent with the Irving-Williams series. As the Ni(II) ions bind to the N-terminal Aβ segment, the binding affinity should be rather the same for Aβ40 and Aβ42 peptides, and also for shorter Aβ versions such as Aβ(1–28) and Aβ(1–16). The CD measurements indicate that the Ni(II) binding affinity to the truncated Aβ(4–40) peptide is similar to, or even somewhat weaker than, the affinity to the full-length Aβ40 peptide (Figs. 3 and 4). This is unexpected, as the Aβ(4–40) peptide has been reported to contain an N-terminal binding motif that supposedly provides very strong binding to Cu(II) and Ni(II) ions139, i.e. possibly fM affinity for Cu(II) ions140. Binding of metal ions to truncated Aβ variants is biologically relevant as such variants, and especially Aβ(4–42), are abundant in amyloid plaques from both healthy and AD brain tissues141–144.
Effects of Ni(II) ions on Aβ structure and aggregation
Similar to e.g. Ag(I), Cu(II), Hg(II), and Zn(II) ions39,40,52,53, Ni(II) ions retard Aβ40 amyloid formation in a concentration-dependent manner by directing the aggregation pathways towards non-fibrillar amorphous aggregates as demonstrated both by ThT fluorescence and AFM imaging (Figs. 5 and 6). Already at a 1:1 Ni(II)/Aβ ratio, Aβ40 fibrillation appears to be completely inhibited. This supports earlier studies reporting that Ni(II) ions can influence protein aggregation145. We have previously shown that low Zn(II) concentrations induce a Zn(II)-bound structure that prevents the Aβ peptides from forming the β-hairpin required for fibrillation19,40. At higher Zn(II) concentrations β-sheet structure was induced92, similar to our current observations with CD spectroscopy that Ni(II) ions induce β-sheet structure in Aβ40 and Aβ(4–40) peptides (Fig. 3D,F). Aβ aggregation is promoted by the direct electrostatic effect of binding cations to the anionic Aβ peptides, thereby reducing repulsion between the Aβ peptides40. Given that Ni(II) and Zn(II) ions have similar charge and binding ligands, they likely affect Aβ aggregation and fibrillation via similar mechanisms.
Although Aβ fibrils, such as those shown in Fig. 6A,B, are the end products of Aβ aggregation, intermediate aggregates known as soluble oligomers are now generally considered to be the main toxic species in AD pathology146,147. The toxic mechanisms are unclear, but may involve membrane disruption30, as some studies have reported that Aβ oligomers can form membrane-spanning “pores” that can induce leakage of e.g. Ca(II) ions148. Interestingly, other studies have reported that this harmful Ca(II) leakage can be inhibited by histidine-associating compounds, such as imidazole, Zn(II), and Ni(II) ions41,149. Because Cu(II), Zn(II), and other divalent ions have been shown to affect the structure, stability, and/or toxicity of Aβ oligomers149–152, it is worth noting that both our FTIR (Figs. 8 and S2) and CD results (Figs. 3 and S1) show that Ni(II) ions induce structural changes in both Aβ oligomers and Aβ monomers. This is in line with earlier studies showing that both SDS-stabilized Aβ42 oligomers81,153,154, and SDS micelle-bound Aβ monomers51,96 contain surface-exposed N-termini which makes it possible for the N-terminal H6, H13, and H14 residues to interact with metal ions. We speculate that Ni(II) ions affect Aβ oligomer structure in similar ways as Cu(II) and Zn(II) ions do, even though the exact mechanisms are not fully understood30. Addition of Ni(II) ions reduces the intensity of the NMR crosspeaks for monomeric Aβ40 peptides in random coil structure (Fig. 1), showing that this conformation becomes less populated. But no new NMR crosspeaks appear (Fig. 1), which shows that the Ni(II) ions do not bind to form a single well-defined Ni(II)·Aβ complex. Instead, a range of heterogeneous β-sheet-containing structures are induced, which most likely exist in different stages of aggregation. They can be observed in CD and FTIR spectra, but not in NMR spectra92,94.
Effects of Ni(II) ions on Aβ dityrosine formation
Ni(II) ions may affect the Aβ aggregation processes also via formation of reactive oxygen species (ROS). Nickel is well known as a redox-active metal that can adopt a wide range of oxidation states, i.e., from − 1 to + 4155. While this can be usefully employed in engineering contexts such as in Ni–Cd batteries, in biological systems it means that Ni can induce oxidative stress, and this may be one of the main mechanisms of Ni toxicity69. Our experiments show that addition of Ni(II) ions initiates formation of Aβ40 dityrosine cross-links (Fig. 9), which is a common ROS effect. Earlier studies have shown that bound redox-active Cu ions can initiate dityrosine formation, both in Aβ and other peptides and proteins96,122–125,152,156,157. It is therefore not surprising that a similar effect is observed for Ni(II) ions, especially as the NMR results indicate that Tyr10 is one of the Ni(II) binding ligands (Fig. 2). As wt Aβ peptides only contain one Tyr residue, i.e. Tyr10, dityrosine formation must involve two Aβ peptides, which then combine into a covalently linked dimer. Such dityrosine-linked Aβ dimers are of biological significance, as they have been found in amyloid plaques in AD brains158. As these plaques contain elevated levels of bound redox-active Cu and Fe ions26,27, it is likely that the Aβ dityrosine links observed in AD patients are generated by metal-induced ROS. Because dimerization is the first step towards peptide aggregation, dityrosine formation in Aβ peptides is clearly a process that influences aggregation. In vitro studies have shown that dityrosine-linked Aβ dimers undergo rapid aggregation into oligomers that are stable, soluble, and neurotoxic159.
Conclusions
We here show for the first time that Ni(II) ions bind to the N-terminal segment of biologically relevant (i.e., full-length) Aβ peptides. The Ni(II) binding affinity is in the low µM range, with the three N-terminal His residues and possibly Tyr10 involved as binding ligands. At equimolar amounts, Ni(II) ions impede Aβ fibrillation by directing the aggregation towards amorphous aggregates. The redox-active Ni(II) ions induce dityrosine cross-links via redox chemistry, thereby creating covalent Aβ dimers. Ni(II) ions induce structural alterations in Aβ monomers, both in aqueous buffer (formation of beta sheets) and in membrane-mimicking SDS micelles (likely formation of coil-coil helix), and affect also Aβ oligomerization. Although Ni(II) binding to Aβ is somewhat weaker than Cu(II) binding, the two metal ions induce similar effects on Aβ structure and aggregation. Exposure to stochiometric amounts of Ni(II) ions induces formation of heterogeneous Aβ oligomers, which can be observed with CD and IR but not NMR spectroscopy. These oligomers, which are in a dynamic equilibrium with Aβ monomers, may be important contributors to AD brain pathology.
Supplementary Information
Acknowledgements
This work was supported by grants from the Swedish Research Council and the Brain Foundation to AG, from the Magnus Bergvall Foundation to SW, AB, and PR, from the Kamprad Research Foundation, the Ulla-Carin Lindquist Foundation for ALS Research, and the Karolinska Institutet IMM strategic grants to PR, from the Swedish Research Council and Umeå University to LMR, and from the Olle Engkvist Foundation and Region Stockholm to AB. We thank Monica Nordberg and Sabrina Sholts for helpful discussions, and Rui Wang and Jing Chi at the office of undergraduate education, Jilin University, for their support to XD.
Author contributions
A.G., E.B., P.R., S.W., and T.S. designed the study. F.V. and A.B carried out and evaluated the IR and PAGE studies. E.B., S.P., and S.W. carried out and evaluated the NMR studies. I.I., J.P., L.A.M.-R., and L.O. carried out and evaluated the AFM studies. E.B., J.J., S.W., T.S., and X.D. carried out and evaluated the CD and fluorescence studies. All authors helped with preparing the manuscript.
Funding
Open access funding provided by Stockholm University.
Data availability
All data generated or analysed during this study are included in this published article (and its supplementary information files). The raw data for the spectroscopy measurements are available from the corresponding author on reasonable request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Elina Berntsson, Email: elina.berntsson@dbb.su.se.
Sebastian K. T. S. Wärmländer, Email: seb@student.su.se
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-023-29901-5.
References
- 1.Prince, M. et al. World Alzheimer Report 2015—The Global Impact of Dementia (Alzheimer’s Disease International (ADI), London, UK, 2015).
- 2.Alzheimer's-Association 2017 Alzheimer's disease facts and figures. Alzheimer's Dementia. 2017;13:325–373. doi: 10.1016/j.jalz.2017.02.001. [DOI] [Google Scholar]
- 3.Fan L, et al. New insights into the pathogenesis of Alzheimer's disease. Front. Neurol. 2019;10:1312. doi: 10.3389/fneur.2019.01312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Masters CL, et al. Alzheimer's disease. Nat. Rev. Dis. Primers. 2015;1:15056. doi: 10.1038/nrdp.2015.56. [DOI] [PubMed] [Google Scholar]
- 5.Irvine GB, El-Agnaf OM, Shankar GM, Walsh DM. Protein aggregation in the brain: The molecular basis for Alzheimer's and Parkinson's diseases. Mol. Med. 2008;14:451–464. doi: 10.2119/2007-00100.Irvine. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Glenner GG, Wong CW. Alzheimer's disease: Initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem. Biophys. Res. Commun. 1984;120:885–890. doi: 10.1016/S0006-291X(84)80190-4. [DOI] [PubMed] [Google Scholar]
- 7.Sunde M, Blake CC. From the globular to the fibrous state: Protein structure and structural conversion in amyloid formation. Q. Rev. Biophys. 1998;31:1–39. doi: 10.1017/S0033583598003400. [DOI] [PubMed] [Google Scholar]
- 8.Eisenberg D, Jucker M. The amyloid state of proteins in human diseases. Cell. 2012;148:1188–1203. doi: 10.1016/j.cell.2012.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Haass C, Selkoe DJ. Soluble protein oligomers in neurodegeneration: Lessons from the Alzheimer's amyloid β-peptide. Nat. Rev. Mol. Cell Biol. 2007;8:101–112. doi: 10.1038/nrm2101. [DOI] [PubMed] [Google Scholar]
- 10.Walsh DM, Selkoe DJ. Aβ oligomers—A decade of discovery. J. Neurochem. 2007;101:1172–1184. doi: 10.1111/j.1471-4159.2006.04426.x. [DOI] [PubMed] [Google Scholar]
- 11.Glabe CG. Structural classification of toxic amyloid oligomers. J. Biol. Chem. 2008;283:29639–29643. doi: 10.1074/jbc.R800016200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sandberg A, et al. Stabilization of neurotoxic Alzheimer amyloid-β oligomers by protein engineering. Proc. Natl. Acad. Sci. USA. 2010;107:15595–15600. doi: 10.1073/pnas.1001740107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Luo J, Wärmländer SK, Gräslund A, Abrahams JP. Cross-interactions between the Alzheimer disease amyloid-β peptide and other amyloid proteins: A further aspect of the amyloid cascade hypothesis. J. Biol. Chem. 2016;291:16485–16493. doi: 10.1074/jbc.R116.714576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Selkoe DJ, Hardy J. The amyloid hypothesis of Alzheimer's disease at 25 years. EMBO Mol. Med. 2016;8:595–608. doi: 10.15252/emmm.201606210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nath S, et al. Spreading of neurodegenerative pathology via neuron-to-neuron transmission of β-amyloid. J. Neurosci. 2012;32:8767–8777. doi: 10.1523/JNEUROSCI.0615-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sardar Sinha M, et al. Alzheimer's disease pathology propagation by exosomes containing toxic amyloid-β oligomers. Acta Neuropathol. 2018;136:41–56. doi: 10.1007/s00401-018-1868-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Aaseth J, et al. Copper, iron, selenium and lipo-glycemic dysmetabolism in Alzheimer's disease. Int. J. Mol. Sci. 2021 doi: 10.3390/ijms22179461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Owen MC, et al. Effects of in vivo conditions on amyloid aggregation. Chem. Soc. Rev. 2019 doi: 10.1039/c8cs00034d. [DOI] [PubMed] [Google Scholar]
- 19.Abelein A, et al. The hairpin conformation of the amyloid β peptide is an important structural motif along the aggregation pathway. J. Biol. Inorg. Chem. 2014;19:623–634. doi: 10.1007/s00775-014-1131-8. [DOI] [PubMed] [Google Scholar]
- 20.Wärmländer S, et al. Biophysical studies of the amyloid β-peptide: Interactions with metal ions and small molecules. ChemBioChem. 2013;14:1692–1704. doi: 10.1002/cbic.201300262. [DOI] [PubMed] [Google Scholar]
- 21.Luo J, et al. Endogenous polyamines reduce the toxicity of soluble Aβ peptide aggregates associated with Alzheimer's disease. Biomacromol. 2014;15:1985–1991. doi: 10.1021/bm401874j. [DOI] [PubMed] [Google Scholar]
- 22.Luo J, et al. Cellular polyamines promote amyloid-beta (Aβ) peptide fibrillation and modulate the aggregation pathways. ACS Chem. Neurosci. 2013;4:454–462. doi: 10.1021/cn300170x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wallin C, Luo J, Jarvet J, Wärmländer S, Gräslund A. The Amyloid-β peptide in amyloid formation processes: Interactions with blood proteins and naturally occurring metal ions. Isr. J. Chem. 2017;57:674–685. doi: 10.1002/ijch.201600105. [DOI] [Google Scholar]
- 24.Duce JA, Bush AI, Adlard PA. Role of amyloid-β–metal interactions in Alzheimer’s disease. Future Neurol. 2011;6:641–659. doi: 10.2217/fnl.11.43. [DOI] [Google Scholar]
- 25.Szabo ST, Harry GJ, Hayden KM, Szabo DT, Birnbaum L. Comparison of metal levels between postmortem brain and ventricular fluid in Alzheimer's disease and nondemented elderly controls. Toxicol. Sci. 2016;150:292–300. doi: 10.1093/toxsci/kfv325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lovell MA, Robertson JD, Teesdale WJ, Campbell JL, Markesbery WR. Copper, iron and zinc in Alzheimer's disease senile plaques. J. Neurol. Sci. 1998;158:47–52. doi: 10.1016/S0022-510X(98)00092-6. [DOI] [PubMed] [Google Scholar]
- 27.Miller LM, et al. Synchrotron-based infrared and X-ray imaging shows focalized accumulation of Cu and Zn co-localized with β-amyloid deposits in Alzheimer's disease. J. Struct. Biol. 2006;155:30–37. doi: 10.1016/j.jsb.2005.09.004. [DOI] [PubMed] [Google Scholar]
- 28.Beauchemin D, Kisilevsky R. A method based on ICP-MS for the analysis of Alzheimer's amyloid plaques. Anal. Chem. 1998;70:1026–1029. doi: 10.1021/ac970783f. [DOI] [PubMed] [Google Scholar]
- 29.Tiiman A, Palumaa P, Tougu V. The missing link in the amyloid cascade of Alzheimer's disease-metal ions. Neurochem. Int. 2013;62:367–378. doi: 10.1016/j.neuint.2013.01.023. [DOI] [PubMed] [Google Scholar]
- 30.Wärmländer SKTS, et al. Metal binding to the amyloid-β peptides in the presence of biomembranes: Potential mechanisms of cell toxicity. J. Biol. Inorganic Chem. 2019;24:1189–1196. doi: 10.1007/s00775-019-01723-9. [DOI] [PubMed] [Google Scholar]
- 31.Huat TJ, et al. Metal toxicity links to Alzheimer's disease and neuroinflammation. J. Mol. Biol. 2019;431:1843–1868. doi: 10.1016/j.jmb.2019.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kim AC, Lim S, Kim YK. Metal ion effects on Aβ and Tau aggregation. Int. J. Mol. Sci. 2018 doi: 10.3390/ijms19010128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wild K, August A, Pietrzik CU, Kins S. Structure and synaptic function of metal binding to the amyloid precursor protein and its proteolytic fragments. Front. Mol. Neurosci. 2017;10:21. doi: 10.3389/fnmol.2017.00021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ayton S, Lei P, Bush AI. Metallostasis in Alzheimer's disease. Free Radic. Biol. Med. 2013;62:76–89. doi: 10.1016/j.freeradbiomed.2012.10.558. [DOI] [PubMed] [Google Scholar]
- 35.Branch T, Barahona M, Dodson CA, Ying L. Kinetic analysis reveals the identity of Aβ-metal complex responsible for the initial aggregation of Aβ in the synapse. ACS Chem. Neurosci. 2017;8:1970–1979. doi: 10.1021/acschemneuro.7b00121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Miller Y, Ma B, Nussinov R. Metal binding sites in amyloid oligomers: Complexes and mechanisms. Coord. Chem. Rev. 2012;256:2245–2252. doi: 10.1016/j.ccr.2011.12.022. [DOI] [Google Scholar]
- 37.Wallin C, et al. Characterization of Mn(II) ion binding to the amyloid-β peptide in Alzheimer's disease. J. Trace Elem. Med. Biol. 2016;38:183–193. doi: 10.1016/j.jtemb.2016.03.009. [DOI] [PubMed] [Google Scholar]
- 38.Wallin C, et al. Alzheimer's disease and cigarette smoke components: Effects of nicotine, PAHs, and Cd(II), Cr(III), Pb(II), Pb(IV) ions on amyloid-β peptide aggregation. Sci. Rep. 2017;7:14423. doi: 10.1038/s41598-017-13759-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Faller P, Hureau C, Berthoumieu O. Role of metal ions in the self-assembly of the Alzheimer's amyloid-β peptide. Inorg. Chem. 2013;52:12193–12206. doi: 10.1021/ic4003059. [DOI] [PubMed] [Google Scholar]
- 40.Danielsson J, Pierattelli R, Banci L, Gräslund A. High-resolution NMR studies of the zinc-binding site of the Alzheimer's amyloid β-peptide. FEBS J. 2007;274:46–59. doi: 10.1111/j.1742-4658.2006.05563.x. [DOI] [PubMed] [Google Scholar]
- 41.Arispe N, Diaz JC, Flora M. Efficiency of histidine-associating compounds for blocking the Alzheimer's Aβ channel activity and cytotoxicity. Biophys. J. 2008;95:4879–4889. doi: 10.1529/biophysj.108.135517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.ATSDR. Agency for Toxic Substances and Disease Registry: Toxicological Profile for Nickel (2005).
- 43.Uter W, Pfahlberg A, Gefeller O, Geier J, Schnuch A. Risk factors for contact allergy to nickel—Results of a multifactorial analysis. Contact Dermatitis. 2003;48:33–38. doi: 10.1034/j.1600-0536.46.s4.29_102.x. [DOI] [PubMed] [Google Scholar]
- 44.Ijomone, O. M. Advances in Neurotoxicology (ed Ashner, M.) 263–284 (Academic Press, 2021).
- 45.Józsa E, et al. Nickel(II) and mixed metal complexes of amyloid-β N-terminus. Dalton Trans. 2010;39:7046–7053. doi: 10.1039/c0dt00189a. [DOI] [PubMed] [Google Scholar]
- 46.Grenács A, Sóvágó I. Copper(II), nickel(II) and zinc(II) complexes of the N-terminal nonapeptide fragment of amyloid-β and its derivatives. J. Inorg. Biochem. 2014;139:49–56. doi: 10.1016/j.jinorgbio.2014.06.001. [DOI] [PubMed] [Google Scholar]
- 47.Lermyte F, et al. Metal ion binding to the amyloid β monomer studied by native top-down FTICR mass spectrometry. J. Am. Soc. Mass Spectrom. 2019 doi: 10.1007/s13361-019-02283-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Österlund N, et al. Amyloid-β peptide interactions with amphiphilic surfactants: Electrostatic and hydrophobic effects. ACS Chem. Neurosci. 2018;9:1680–1692. doi: 10.1021/acschemneuro.8b00065. [DOI] [PubMed] [Google Scholar]
- 49.Österlund N, Luo J, Wärmländer SKTS, Gräslund A. Membrane-mimetic systems for biophysical studies of the amyloid-β peptide. Biochim. Biophys. Acta Proteins Proteom. 1867;492–501:2019. doi: 10.1016/j.bbapap.2018.11.005. [DOI] [PubMed] [Google Scholar]
- 50.Danielsson J, Jarvet J, Damberg P, Gräslund A. The Alzheimer β-peptide shows temperature-dependent transitions between left-handed 3-helix, β-strand and random coil secondary structures. FEBS J. 2005;272:3938–3949. doi: 10.1111/j.1742-4658.2005.04812.x. [DOI] [PubMed] [Google Scholar]
- 51.Jarvet J, Danielsson J, Damberg P, Oleszczuk M, Gräslund A. Positioning of the Alzheimer Aβ(1–40) peptide in SDS micelles using NMR and paramagnetic probes. J. Biomol. NMR. 2007;39:63–72. doi: 10.1007/s10858-007-9176-4. [DOI] [PubMed] [Google Scholar]
- 52.Wallin C, et al. Mercury and Alzheimer's disease: Hg(II) ions display specific binding to the amyloid-β peptide and hinder its fibrillization. Biomolecules. 2019 doi: 10.3390/biom10010044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wallin C, et al. Metal ion coordination delays amyloid-β peptide self-assembly by forming an aggregation-inert complex. J. Biol. Chem. 2020;295:7224–7234. doi: 10.1074/jbc.RA120.012738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Klein, C. & Costa, M. Handbook on the Toxicology of Metals, 4th ed. Volume II: Specific Metals (eds Nordberg, G. F. et al.) (Elsevier, 2015).
- 55.Anke M, Groppel B, Kronemann H, Grun M. Nickel—An essential element. IARC Sci Publ; 1984. pp. 339–365. [PubMed] [Google Scholar]
- 56.Fischer F, et al. Characterization in Helicobacter pylori of a nickel transporter essential for colonization that was acquired during evolution by gastric Helicobacter species. PLoS Pathog. 2016;12:e1006018. doi: 10.1371/journal.ppat.1006018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Eskew DL, Welch RM, Cary EE. Nickel: An essential micronutrient for legumes and possibly all higher plants. Science. 1983;222:621–623. doi: 10.1126/science.222.4624.621. [DOI] [PubMed] [Google Scholar]
- 58.Genchi G, Carocci A, Lauria G, Sinicropi MS, Catalano A. Nickel: Human health and environmental toxicology. Int. J. Environ. Res. Public Health. 2020 doi: 10.3390/ijerph17030679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Nielsen FH, Ollerich DA, Fosmire GJ, Sandstead HH. Nickel deficiency in chicks and rats: Effects on liver morphology, function and polysomal integrity. Adv. Exp. Med. Biol. 1974;48:389–403. doi: 10.1007/978-1-4684-0943-7_18. [DOI] [PubMed] [Google Scholar]
- 60.Stangl GI, Kirchgessner M. Nickel deficiency alters liver lipid metabolism in rats. J. Nutr. 1996;126:2466–2473. doi: 10.1093/jn/126.10.2466. [DOI] [PubMed] [Google Scholar]
- 61.Mania M, Rebeniak M, Postupolski J. Exposure assessment of the population in Poland to the toxic effects of nickel from vegetable and their products. Rocz. Panstw. Zakl. Hig. 2019;70:401–406. doi: 10.32394/rpzh.2019.0095. [DOI] [PubMed] [Google Scholar]
- 62.World Health, O. Air quality guidelines for Europe. WHO Reg Publ Eur Ser, V-X, 1–273 (2000). [PubMed]
- 63.Chain, E. P. o. C. i. t. F. et al. Update of the risk assessment of nickel in food and drinking water. EFSA J. 2020;18:e06268. doi: 10.2903/j.efsa.2020.6268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bernhard D, Rossmann A, Wick G. Metals in cigarette smoke. IUBMB Life. 2005;57:805–809. doi: 10.1080/15216540500459667. [DOI] [PubMed] [Google Scholar]
- 65.Dahlstrand H, Stark A, Anissian L, Hailer NP. Elevated serum concentrations of cobalt, chromium, nickel, and manganese after metal-on-metal alloarthroplasty of the hip: A prospective randomized study. J. Arthroplasty. 2009;24:837–845. doi: 10.1016/j.arth.2008.07.019. [DOI] [PubMed] [Google Scholar]
- 66.Setcos JC, Babaei-Mahani A, Silvio LD, Mjor IA, Wilson NH. The safety of nickel containing dental alloys. Dent. Mater. 2006;22:1163–1168. doi: 10.1016/j.dental.2005.11.033. [DOI] [PubMed] [Google Scholar]
- 67.Cheng CF, Schwitter CM. Nickel in ancient bronzes. Am. J. Archaeol. 1957;61:351–365. doi: 10.2307/500603. [DOI] [Google Scholar]
- 68.Rauch JN, Pacyna JM. Earth's global Ag, Al, Cr, Cu, Fe, Ni, Pb, and Zn cycles. Glob. Biogeochem. Cycles. 2009;23:GB2001. doi: 10.1029/2008GB003376. [DOI] [Google Scholar]
- 69.Das KK, Das SN, Dhundasi SA. Nickel, its adverse health effects & oxidative stress. Indian J. Med. Res. 2008;128:412–425. [PubMed] [Google Scholar]
- 70.Nordberg G, Costa M. Handbook on the Toxicology of Metals. 5. Academic Press; 2022. [Google Scholar]
- 71.Hou YP, et al. The characteristics of placental transfer and tissue concentrations of nickel in late gestational rats and fetuses. Placenta. 2011;32:277–282. doi: 10.1016/j.placenta.2010.12.021. [DOI] [PubMed] [Google Scholar]
- 72.Mangelsdorf I, Walach H, Mutter J. Healing of amyotrophic lateral sclerosis: A case report. Complement. Med. Res. 2017;24:175–181. doi: 10.1159/000477397. [DOI] [PubMed] [Google Scholar]
- 73.Bar-Sela S, Reingold S, Richter ED. Amyotrophic lateral sclerosis in a battery-factory worker exposed to cadmium. Int. J. Occup. Environ. Health. 2001;7:109–112. doi: 10.1179/107735201800339470. [DOI] [PubMed] [Google Scholar]
- 74.Forte G, et al. Quantification of chemical elements in blood of patients affected by multiple sclerosis. Ann. Ist. Super Sanita. 2005;41:213–216. [PubMed] [Google Scholar]
- 75.Irvine DG, Schiefer HB, Hader WJ. Geotoxicology of multiple sclerosis: The Henribourg, Saskatchewan, cluster focus. II. The soil. Sci. Total Environ. 1988;77:175–188. doi: 10.1016/0048-9697(88)90054-x. [DOI] [PubMed] [Google Scholar]
- 76.Frye RE, et al. Early life metal exposure dysregulates cellular bioenergetics in children with regressive autism spectrum disorder. Transl. Psychiatry. 2020;10:223. doi: 10.1038/s41398-020-00905-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Kim SH, et al. Rapid doubling of Alzheimer's amyloid-β40 and 42 levels in brains of mice exposed to a nickel nanoparticle model of air pollution. F1000Res. 2012;1:70. doi: 10.12688/f1000research.1-70.v1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Gorantla NV, Das R, Balaraman E, Chinnathambi S. Transition metal nickel prevents Tau aggregation in Alzheimer's disease. Int. J. Biol. Macromol. 2020;156:1359–1365. doi: 10.1016/j.ijbiomac.2019.11.176. [DOI] [PubMed] [Google Scholar]
- 79.Glasoe PK, Long FA. Use of glass electrodes to measure acidities in deuterium oxide. J. Phys. Chem. 1960;64:88–90. doi: 10.1021/j100830a521. [DOI] [Google Scholar]
- 80.Edelhoch H. Spectroscopic determination of tryptophan and tyrosine in proteins. Biochemistry. 1967;6:1948–1954. doi: 10.1021/bi00859a010. [DOI] [PubMed] [Google Scholar]
- 81.Barghorn S, et al. Globular amyloid β-peptide1-42 oligomer—A homogenous and stable neuropathological protein in Alzheimer’s disease. J. Neurochem. 2005;95:834–847. doi: 10.1111/j.1471-4159.2005.03407.x. [DOI] [PubMed] [Google Scholar]
- 82.Vosough F, Barth A. Characterization of homogeneous and heterogeneous amyloid-β42 oligomer preparations with biochemical methods and infrared spectroscopy reveals a correlation between infrared spectrum and oligomer size. ACS Chem. Neurosci. 2021;12:473–488. doi: 10.1021/acschemneuro.0c00642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Dominguez A, Fernandez A, Gonzalez N, Iglesias E, Motenegro E. Determination of critical micelle concentration of some surfactants by three techniques. J. Chem. Educ. 1997;74:1227–1231. doi: 10.1021/ed074p1227. [DOI] [Google Scholar]
- 84.Danielsson J, Andersson A, Jarvet J, Gräslund A. 15N relaxation study of the amyloid β-peptide: Structural propensities and persistence length. Magn. Reason. Chem. 2006;44:S114–S121. doi: 10.1002/mrc.1814. [DOI] [PubMed] [Google Scholar]
- 85.Roche J, Shen Y, Lee JH, Ying J, Bax A. Monomeric Aβ(1–40) and Aβ (1–42) peptides in solution adopt very similar Ramachandran map distributions that closely resemble random coil. Biochemistry. 2016;55:762–775. doi: 10.1021/acs.biochem.5b01259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Yamaguchi T, Matsuzaki K, Hoshino M. Transient formation of intermediate conformational states of amyloid-β peptide revealed by heteronuclear magnetic resonance spectroscopy. FEBS Lett. 2011;585:1097–1102. doi: 10.1016/j.febslet.2011.03.014. [DOI] [PubMed] [Google Scholar]
- 87.Kuzmic, P. History, variants and usage of the "Morrison equation" in enzyme inhibition kinetics, BioKin Technical Note TN-2015-01 http://www.biokin.com/TN/2015/01 (2015).
- 88.Biancalana M, Koide S. Molecular mechanism of Thioflavin-T binding to amyloid fibrils. Biochem. Biophys. Acta. 1804;1405–1412:2010. doi: 10.1016/j.bbapap.2010.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Gade Malmos K, et al. ThT 101: A primer on the use of thioflavin T to investigate amyloid formation. Amyloid. 2017;24:1–16. doi: 10.1080/13506129.2017.1304905. [DOI] [PubMed] [Google Scholar]
- 90.Hellstrand E, Boland B, Walsh DM, Linse S. Amyloid β-protein aggregation produces highly reproducible kinetic data and occurs by a two-phase process. ACS Chem Neurosci. 2010;1:13–18. doi: 10.1021/cn900015v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Baldassarre M, Barth A. Pushing the detection limit of infrared spectroscopy for structural analysis of dilute protein samples. Analyst. 2014;139:5393–5399. doi: 10.1039/c4an00918e. [DOI] [PubMed] [Google Scholar]
- 92.Ghalebani L, Wahlström A, Danielsson J, Wärmländer SK, Gräslund A. pH-dependence of the specific binding of Cu(II) and Zn(II) ions to the amyloid-β peptide. Biochem. Biophys. Res. Commun. 2012;421:554–560. doi: 10.1016/j.bbrc.2012.04.043. [DOI] [PubMed] [Google Scholar]
- 93.Barnhart JL, Berk RN. Influence of paramagnetic ions and pH on proton NMR relaxation of biologic fluids. Investig. Radiol. 1986;21:132–136. doi: 10.1097/00004424-198602000-00009. [DOI] [PubMed] [Google Scholar]
- 94.Fawzi NL, Ying J, Torchia DA, Clore GM. Kinetics of amyloid β monomer-to-oligomer exchange by NMR relaxation. J. Am. Chem. Soc. 2010;132:9948–9951. doi: 10.1021/ja1048253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Pogostin BH, Malmendal A, Londergan CH, Akerfeldt KS. pKa determination of a histidine residue in a short peptide using Raman spectroscopy. Molecules. 2019 doi: 10.3390/molecules24030405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Tiiman A, et al. Specific binding of Cu(II) ions to amyloid-β peptides bound to aggregation-inhibiting molecules or SDS micelles creates complexes that generate radical oxygen species. J. Alzheimers Dis. 2016;54:971–982. doi: 10.3233/JAD-160427. [DOI] [PubMed] [Google Scholar]
- 97.Gielnik M, et al. The prion protein octarepeat domain forms transient β-sheet structures upon residue-specific Cu(II) and Zn(II) binding. bioRxiv. 2021 doi: 10.1101/2021.12.12.472308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Kjaergaard M, et al. Temperature-dependent structural changes in intrinsically disordered proteins: Formation of alpha-helices or loss of polyproline II? Protein Sci. 2010;19:1555–1564. doi: 10.1002/pro.435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Greenfield N, Fasman GD. Computed circular dichroism spectra for the evaluation of protein conformation. Biochemistry. 1969;8:4108–4116. doi: 10.1021/bi00838a031. [DOI] [PubMed] [Google Scholar]
- 100.Usachev KS, Filippov AV, Khairutdinov BI, Antzutkin ON, Klochkov VV. NMR structure of the Arctic mutation of the Alzheimer’s Aβ(1–40) peptide docked to SDS micelles. J. Mol. Struct. 2014;1076:518–523. doi: 10.1016/j.molstruc.2014.08.030. [DOI] [Google Scholar]
- 101.Lopez-Llano J, Campos LA, Sancho J. Alpha-helix stabilization by alanine relative to glycine: Roles of polar and apolar solvent exposures and of backbone entropy. Proteins. 2006;64:769–778. doi: 10.1002/prot.21041. [DOI] [PubMed] [Google Scholar]
- 102.Lau SY, Taneja AK, Hodges RS. Synthesis of a model protein of defined secondary and quaternary structure. Effect of chain length on the stabilization and formation of two-stranded alpha-helical coiled-coils. J. Biol. Chem. 1984;259:13253–13261. doi: 10.1016/S0021-9258(18)90686-1. [DOI] [PubMed] [Google Scholar]
- 103.Zhou NE, Kay CM, Hodges RS. Synthetic model proteins: The relative contribution of leucine residues at the nonequivalent positions of the 3–4 hydrophobic repeat to the stability of the two-stranded alpha-helical coiled-coil. Biochemistry. 1992;31:5739–5746. doi: 10.1021/bi00140a008. [DOI] [PubMed] [Google Scholar]
- 104.Zhou NE, Kay CM, Hodges RS. Synthetic model proteins. Positional effects of interchain hydrophobic interactions on stability of two-stranded alpha-helical coiled-coils. J. Biol. Chem. 1992;267:2664–2670. doi: 10.1016/S0021-9258(18)45932-7. [DOI] [PubMed] [Google Scholar]
- 105.Barbar E, Nyarko A. NMR characterization of self-association domains promoted by interactions with LC8 hub protein. Comput. Struct. Biotechnol. J. 2014;9:e201402003. doi: 10.5936/csbj.201402003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Lindgren J, et al. Engineered non-fluorescent Affibody molecules facilitate studies of the amyloid-beta (Aβ) peptide in monomeric form: Low pH was found to reduce Aβ/Cu(II) binding affinity. J. Inorg. Biochem. 2013;120:18–23. doi: 10.1016/j.jinorgbio.2012.11.005. [DOI] [PubMed] [Google Scholar]
- 107.Luo J, et al. Inhibiting and reversing amyloid-β peptide (1–40) fibril formation with gramicidin S and engineered analogues. Chemistry. 2013;19:17338–17348. doi: 10.1002/chem.201301535. [DOI] [PubMed] [Google Scholar]
- 108.Luo J, Wärmländer SK, Gräslund A, Abrahams JP. Human lysozyme inhibits the in vitro aggregation of Aβ peptides, which in vivo are associated with Alzheimer's disease. Chem. Commun. 2013;49:6507–6509. doi: 10.1039/c3cc42325e. [DOI] [PubMed] [Google Scholar]
- 109.Luo J, Wärmländer SK, Gräslund A, Abrahams JP. Reciprocal molecular interactions between the Aβ peptide linked to Alzheimer's disease and insulin linked to diabetes mellitus type II. ACS Chem. Neurosci. 2016;7:269–274. doi: 10.1021/acschemneuro.5b00325. [DOI] [PubMed] [Google Scholar]
- 110.Barth A. Infrared spectroscopy of proteins. Biochem. Biophys. Acta. 2007;1767:1073–1101. doi: 10.1016/j.bbabio.2007.06.004. [DOI] [PubMed] [Google Scholar]
- 111.Barth A, Zscherp C. What vibrations tell us about proteins. Q. Rev. Biophys. 2002;35:369–430. doi: 10.1017/s0033583502003815. [DOI] [PubMed] [Google Scholar]
- 112.Yang H, Yang S, Kong J, Dong A, Yu S. Obtaining information about protein secondary structures in aqueous solution using Fourier transform IR spectroscopy. Nat. Protoc. 2015;10:382–396. doi: 10.1038/nprot.2015.024. [DOI] [PubMed] [Google Scholar]
- 113.Goormaghtigh E, Cabiaux V, Ruysschaert JM. Determination of soluble and membrane protein structure by Fourier transform infrared spectroscopy. III. Secondary structures. Subcell Biochem. 1994;23:405–450. doi: 10.1007/978-1-4615-1863-1_10. [DOI] [PubMed] [Google Scholar]
- 114.Jackson M, Mantsch HH. The use and misuse of FTIR spectroscopy in the determination of protein structure. Crit. Rev. Biochem. Mol. Biol. 1995;30:95–120. doi: 10.3109/10409239509085140. [DOI] [PubMed] [Google Scholar]
- 115.Arrondo JL, Muga A, Castresana J, Goni FM. Quantitative studies of the structure of proteins in solution by Fourier-transform infrared spectroscopy. Prog. Biophys. Mol. Biol. 1993;59:23–56. doi: 10.1016/0079-6107(93)90006-6. [DOI] [PubMed] [Google Scholar]
- 116.Fabian, H. & Mäntele, W. Handbook of Vibrational Spectroscopy (eds Chalmers, J. M. & Griffiths, P.) 3399–3426 (Wiley, 2002).
- 117.Sarroukh R, Goormaghtigh E, Ruysschaert JM, Raussens V. ATR-FTIR: A "rejuvenated" tool to investigate amyloid proteins. Biochem. Biophys. Acta. 1828;2328–2338:2013. doi: 10.1016/j.bbamem.2013.04.012. [DOI] [PubMed] [Google Scholar]
- 118.Li H, Lantz R, Du D. Vibrational approach to the dynamics and structure of protein amyloids. Molecules. 2019 doi: 10.3390/molecules24010186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Moran SD, Zanni MT. How to get insight into amyloid structure and formation from infrared spectroscopy. J. Phys. Chem. Lett. 2014;5:1984–1993. doi: 10.1021/jz500794d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Cerf E, et al. Antiparallel β-sheet: A signature structure of the oligomeric amyloid β-peptide. Biochem. J. 2009;421:415–423. doi: 10.1042/BJ20090379. [DOI] [PubMed] [Google Scholar]
- 121.Berntsson E, et al. Lithium ions display weak interaction with amyloid-beta (Aβ) peptides and have minor effects on their aggregation. Acta Biochim. Pol. 2021;68:169–179. doi: 10.18388/abp.2020_5493. [DOI] [PubMed] [Google Scholar]
- 122.Atwood CS, et al. Copper mediates dityrosine cross-linking of Alzheimer's amyloid-β. Biochemistry. 2004;43:560–568. doi: 10.1021/bi0358824. [DOI] [PubMed] [Google Scholar]
- 123.Dong X, et al. Copper ions induce dityrosine-linked dimers in human but not in murine islet amyloid polypeptide (IAPP/amylin) Biochem. Biophys. Res. Commun. 2019;510:520–524. doi: 10.1016/j.bbrc.2019.01.120. [DOI] [PubMed] [Google Scholar]
- 124.Gu M, Bode DC, Viles JH. Copper redox cycling inhibits Aβ fibre formation and promotes fibre fragmentation, while generating a dityrosine Aβ dimer. Sci. Rep. 2018;8:16190. doi: 10.1038/s41598-018-33935-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Vazquez G, et al. Copper, dityrosine cross-links and amyloid-β aggregation. J. Biol. Inorg. Chem. 2019;24:1217–1229. doi: 10.1007/s00775-019-01734-6. [DOI] [PubMed] [Google Scholar]
- 126.Huggins TG, Wells-Knecht MC, Detorie NA, Baynes JW, Thorpe SR. Formation of o-tyrosine and dityrosine in proteins during radiolytic and metal-catalyzed oxidation. J. Biol. Chem. 1993;268:12341–12347. doi: 10.1016/S0021-9258(18)31395-4. [DOI] [PubMed] [Google Scholar]
- 127.Keleti T. The excimer fluorescence of tryptophan, tyrosine and d-glyceraldehyde-3-phosphate dehydrogenase. FEBS Lett. 1970;7:280–282. doi: 10.1016/0014-5793(70)80181-8. [DOI] [PubMed] [Google Scholar]
- 128.Zilbermann I, Maimon E, Cohen H, Meyerstein D. Redox chemistry of nickel complexes in aqueous solutions. Chem. Rev. 2005;105:2609–2625. doi: 10.1021/cr030717f. [DOI] [PubMed] [Google Scholar]
- 129.Harris ME, Hensley K, Butterfield DA, Leedle RA, Carney JM. Direct evidence of oxidative injury produced by the Alzheimer's β-amyloid peptide (1–40) in cultured hippocampal neurons. Exp. Neurol. 1995;131:193–202. doi: 10.1016/0014-4886(95)90041-1. [DOI] [PubMed] [Google Scholar]
- 130.Cenini G, et al. Generation of reactive oxygen species by β amyloid fibrils and oligomers involves different intra/extracellular pathways. Amino Acids. 2010;38:1101–1106. doi: 10.1007/s00726-009-0319-7. [DOI] [PubMed] [Google Scholar]
- 131.Shimazaki Y, Yajima T, Yamauchi O. Properties of the indole ring in metal complexes. A comparison with the phenol ring. J. Inorg. Biochem. 2015;148:105–115. doi: 10.1016/j.jinorgbio.2015.03.001. [DOI] [PubMed] [Google Scholar]
- 132.Zhang S, Casey N, Lee JP. Residual structure in the Alzheimer's disease peptide: Probing the origin of a central hydrophobic cluster. Fold. Des. 1998;3:413–422. doi: 10.1016/S1359-0278(98)00054-6. [DOI] [PubMed] [Google Scholar]
- 133.Knecht S, Ricklin D, Eberle AN, Ernst B. Oligohis-tags: Mechanisms of binding to Ni2+-NTA surfaces. J. Mol. Recognit. 2009;22:270–279. doi: 10.1002/jmr.941. [DOI] [PubMed] [Google Scholar]
- 134.Bousejra-ElGarah F, Bijani C, Coppel Y, Faller P, Hureau C. Iron(II) binding to amyloid-β, the Alzheimer's peptide. Inorg. Chem. 2011;50:9024–9030. doi: 10.1021/ic201233b. [DOI] [PubMed] [Google Scholar]
- 135.Faller P, Hureau C. Bioinorganic chemistry of copper and zinc ions coordinated to amyloid-β peptide. Dalton Trans. 2009 doi: 10.1039/b813398k. [DOI] [PubMed] [Google Scholar]
- 136.Faller P. Copper and zinc binding to amyloid-β: Coordination, dynamics, aggregation, reactivity and metal-ion transfer. ChemBioChem. 2009;10:2837–2845. doi: 10.1002/cbic.200900321. [DOI] [PubMed] [Google Scholar]
- 137.Irving, H. M. N. H. & Williams, R. J. P. The stability of transition-metal complexes. J. Chem. Soc. 3192–3210 (1953).
- 138.Alies B, et al. Cu(II) affinity for the Alzheimer's peptide: Tyrosine fluorescence studies revisited. Anal. Chem. 2013;85:1501–1508. doi: 10.1021/ac302629u. [DOI] [PubMed] [Google Scholar]
- 139.Bar-Or D, Curtis G, Rao N, Bampos N, Lau E. Characterization of the Co(2+) and Ni(2+) binding amino-acid residues of the N-terminus of human albumin. An insight into the mechanism of a new assay for myocardial ischemia. Eur. J. Biochem. 2001;268:42–47. doi: 10.1046/j.1432-1327.2001.01846.x. [DOI] [PubMed] [Google Scholar]
- 140.Mital M, et al. A functional role for Aβ in metal homeostasis? N-truncation and high-affinity copper binding. Angew. Chem. Int. Ed. Engl. 2015;54:10460–10464. doi: 10.1002/anie.201502644. [DOI] [PubMed] [Google Scholar]
- 141.Masters CL, et al. Amyloid plaque core protein in Alzheimer disease and Down syndrome. Proc. Natl. Acad. Sci. USA. 1985;82:4245–4249. doi: 10.1073/pnas.82.12.4245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Masters CL, et al. Neuronal origin of a cerebral amyloid: Neurofibrillary tangles of Alzheimer's disease contain the same protein as the amyloid of plaque cores and blood vessels. EMBO J. 1985;4:2757–2763. doi: 10.1002/j.1460-2075.1985.tb04000.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Lewis H, et al. Quantification of Alzheimer pathology in ageing and dementia: Age-related accumulation of amyloid-β(42) peptide in vascular dementia. Neuropathol. Appl. Neurobiol. 2006;32:103–118. doi: 10.1111/j.1365-2990.2006.00696.x. [DOI] [PubMed] [Google Scholar]
- 144.Portelius E, et al. Mass spectrometric characterization of brain amyloid β isoform signatures in familial and sporadic Alzheimer's disease. Acta Neuropathol. 2010;120:185–193. doi: 10.1007/s00401-010-0690-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Hedberg YS, et al. Synergistic effects of metal-induced aggregation of human serum albumin. Colloids Surf. B Biointerfaces. 2019;173:751–758. doi: 10.1016/j.colsurfb.2018.10.061. [DOI] [PubMed] [Google Scholar]
- 146.Lee SJ, Nam E, Lee HJ, Savelieff MG, Lim MH. Towards an understanding of amyloid-β oligomers: Characterization, toxicity mechanisms, and inhibitors. Chem. Soc. Rev. 2017;46:310–323. doi: 10.1039/c6cs00731g. [DOI] [PubMed] [Google Scholar]
- 147.Sengupta U, Nilson AN, Kayed R. The role of amyloid-β oligomers in toxicity, propagation, and immunotherapy. EBioMedicine. 2016;6:42–49. doi: 10.1016/j.ebiom.2016.03.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Bode DC, Baker MD, Viles JH. Ion channel formation by amyloid-β42 oligomers but not amyloid-β40 in cellular membranes. J. Biol. Chem. 2017;292:1404–1413. doi: 10.1074/jbc.M116.762526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Kim M, Son J, Kim Y. NMR studies of the ion channel-forming human amyloid-β with zinc ion concentrations. Membranes. 2021 doi: 10.3390/membranes11110799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Lee MC, et al. Zinc ion rapidly induces toxic, off-pathway amyloid-β oligomers distinct from amyloid-β derived diffusible ligands in Alzheimer's disease. Sci. Rep. 2018;8:4772. doi: 10.1038/s41598-018-23122-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Garai K, Sengupta P, Sahoo B, Maiti S. Selective destabilization of soluble amyloid β oligomers by divalent metal ions. Biochem. Biophys. Res. Commun. 2006;345:210–215. doi: 10.1016/j.bbrc.2006.04.056. [DOI] [PubMed] [Google Scholar]
- 152.Williams TL, Serpell LC, Urbanc B. Stabilization of native amyloid β-protein oligomers by copper and hydrogen peroxide induced cross-linking of unmodified proteins (CHICUP) Biochem. Biophys. Acta. 1864;249–259:2016. doi: 10.1016/j.bbapap.2015.12.001. [DOI] [PubMed] [Google Scholar]
- 153.Yu L, et al. Structural characterization of a soluble amyloid β-peptide oligomer. Biochemistry. 2009;48:1870–1877. doi: 10.1021/bi802046n. [DOI] [PubMed] [Google Scholar]
- 154.Gu L, Liu C, Guo Z. Structural insights into Aβ42 oligomers using site-directed spin labeling. J. Biol. Chem. 2013;288:18673–18683. doi: 10.1074/jbc.M113.457739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Nag K, Chakravorty A. Monovalent, trivalent and tetravalent nickel. Coord. Chem. Rev. 1980;33:87–147. doi: 10.1016/S0010-8545(00)80405-1. [DOI] [Google Scholar]
- 156.Smith DP, et al. Concentration dependent Cu2+ induced aggregation and dityrosine formation of the Alzheimer's disease amyloid-β peptide. Biochemistry. 2007;46:2881–2891. doi: 10.1021/bi0620961. [DOI] [PubMed] [Google Scholar]
- 157.Wang J, et al. Peptide backbone-copper ring structure: A molecular insight into copper-induced amyloid toxicity. Chin. Phys. B. 2022;31(10):108702. doi: 10.1088/1674-1056/ac8920. [DOI] [Google Scholar]
- 158.Al-Hilaly YK, et al. A central role for dityrosine crosslinking of Amyloid-β in Alzheimer's disease. Acta Neuropathol. Commun. 2013;1:83. doi: 10.1186/2051-5960-1-83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Kok WM, et al. Synthetic dityrosine-linked β-amyloid dimers form stable, soluble, neurotoxic oligomers. Chem. Sci. 2013;4:4449–4454. doi: 10.1039/c3sc22295k. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data generated or analysed during this study are included in this published article (and its supplementary information files). The raw data for the spectroscopy measurements are available from the corresponding author on reasonable request.