

Abstract
Suicide attempts (SA) among African Americans have increased at a greater rate than any other racial/ethnic group. Research in European ancestry populations has indicated that SA are genetically influenced; however, less is known about the genetic contributors that underpin SA among African Americans. We examined whether genetic propensity for depression and risky behaviors (assessed via polygenic risk scores; PRS) independently and jointly are associated with SA among urban, African Americans and whether sex differences exist in these relations. Participants (N = 1,157, 45.0% male) were originally recruited as part of two first grade universal school-based prevention trials. Participants reported in adolescence and young adulthood on whether they ever attempted suicide in their life. Depression and risky behaviors PRS were created based on large-scale genome-wide association studies conducted by Howard et al. (2019) and Karlson Línner et al. (2019), respectively. There was a significant interaction between the risky behavior PRS and depression PRS such that the combination of high risky behavior polygenic risk and low/moderate polygenic risk for depression was associated with greater risk for lifetime SA among the whole sample and African American males specifically. In addition, the risky behavior PRS was significantly positively associated with lifetime SA among African American males. These findings provide preliminary evidence regarding the importance of examining risky behavior and depression polygenic risk in relation to SA among African Americans, though replication of our findings in other African American samples is needed.
Keywords: African Americans, depression, polygenic risk scores, risky behavior, suicide attempt
1 |. INTRODUCTION
Suicide attempt is a robust risk factor for suicide death, with some studies indicating that close to 9% of individuals who attempt suicide will eventually die by suicide (Bostwick & Pankratz, 2000) and other work indicating that over 80% of deaths by suicide occur within a year of the first suicide attempt (Bostwick, Pabbati, Geske, & McKean, 2016). The epidemiology of suicide and suicide attempts in the United States has shifted over time and an emerging risk group is urban, African American young people (Price & Khubchandani, 2019). Estimates of lifetime suicide attempts among African American youth have ranged from 4.1% to 11% (Nock et al., 2013; Tomek et al., 2015). From 1991 to 2017, suicide attempts increased by over 70% among African American male and female adolescents and attempts requiring medical attention increased by 122% among African American males (Lindsey, Sheftall, Xiao, & Joe, 2019). Given the dramatic increase in suicide attempts among African Americans, an examination into individual-specific risk factors that may explain this increase, such as genetic predispositions, is warranted.
Most research that has examined the impact of genetic variation on suicide attempt has been limited to exclusively European American samples, with African Americans largely underrepresented in this research. The lack of inclusion of diverse ancestral populations, such as African Americans, in genomics research poses a significant problem given differences in linkage disequilibrium (LD), allele frequencies, and environmental/contextual experiences across ancestral populations. Each of these differences may reduce the generalizability of findings observed in European ancestry samples to other ancestral groups (Marnetto et al., 2020; Martin et al., 2017) such as African Americans. For example, several studies have revealed a net attenuation in the variance accounted for in a number of phenotypes (e.g., educational attainment; Rabinowitz et al., 2019) when the ancestry of genetic discovery samples does not reflect the population under study (Martin et al., 2019). The current study aims to examine whether polygenic risk scores for depression and risky behaviors derived from European ancestry discovery samples (Howard et al., 2019; Karlson Línner et al., 2019) independently and jointly are associated with lifetime suicide attempt in an urban, African American sample. This study is a first step in determining whether conclusions drawn from genomics research on suicidal behaviors among European ancestral populations are applicable to urban, African Americans.
1.1 |. Genetics and suicide attempt
Genetic variation is one factor that may influence susceptibility to attempting suicide. For example, a number of studies have indicated that suicidal behavior is moderately heritable (h2 = .30–.50) (Baldessarini & Hennen, 2004; Pedersen & Fiske, 2010; Voracek & Loibl, 2007), with twin and family studies documenting a higher concordance rate for suicide attempt among monozygotic (MZ) (concordance rate range = 0.12–0.47) compared with dizygotic (DZ) twins (concordance rate range = 0.00–0.25) (Pedersen & Fiske, 2010). Other work has indicated that offspring of probands who have attempted suicide are close to five times more likely to attempt suicide themselves (Brent et al., 2015; Lutz, Mechawar, & Turecki, 2017).
Within the last several years, a number of genome-wide association studies (GWAS) have been conducted to identify genetic liabilities associated with suicidal behaviors (Docherty et al., 2020). Based on discovery results from GWAS, polygenic risk scores (PRS) can be generated which are weighted summed scores that often include hundreds of thousands of single nucleotide polymorphisms (SNPs) of very small effect size that represent an individual’s cumulative genetic risk for a specific phenotype (Maher, 2015). Ideally, the use of a PRS approach in relation to suicide attempt may help reveal key etiological factors related to suicide attempts, improve the accuracy of suicidal risk models, and inform more tailored suicide prevention approaches.
1.2 |. Depression genetics and suicide attempt
Polygenic risk for depressive symptoms may play a significant role in suicide attempt manifestation (Lim et al., 2020; Mullins et al., 2014). Research examining the genetic relations between depression and suicide attempt has revealed positive genetic correlations between suicide attempt, major depression, and depressive symptoms (r range = .40–.50) (Ruderfer et al., 2020). Most of the work that has examined whether depressive symptoms PRS are associated with suicide attempt has been conducted in predominantly European ancestry samples. For example, using a twin sample of European ancestry, depression PRS were significantly associated with suicide attempt with a small amount of variance accounted for (R2 range = 0.12%–0.30%) (Maciejewski et al., 2017; Mullins et al., 2014). Other work has indicated that depression PRS were associated with suicide attempt among individuals of European ancestry that met diagnostic criteria for psychiatric disorders (R2 range = 0.25%–0.70%) (Mullins et al., 2014).
To our knowledge, only one study has examined links between genetic liability for depression and phenotypic suicide attempt among African Americans. In particular, Levey et al. (2019) observed that higher levels of depression PRS were tied to greater lethality of suicide attempt (R2 = 0.70%) with a small fraction of the variance explained in a predominantly African American cohort that met diagnostic criteria for substance use disorders. Based on findings from Levey et al. (2019) and work conducted in European American samples (Mullins et al., 2014), it is possible that higher depression polygenic risk may similarly exacerbate risk for attempting suicide in a community sample of urban African Americans, although whether this is the case remains unknown.
1.3 |. Risky behavior genetics and suicide attempt
In addition to the potential association between polygenic risk for depression and phenotypic suicide attempt, genetic propensity for engaging in risky behaviors may also influence liability for attempting suicide; however, there is a dearth of research in this area. Many suicide attempts, particularly those carried out by adolescents and young adults, are impulsive, relatively unplanned events, making impulsivity and risk taking potentially important factors to consider in relation to suicide attempts in this population (Bridge, Goldstein, & Brent, 2006; McHugh et al., 2019). At the phenotypic level, higher levels of impulsivity may reflect difficulties in inhibiting responses, preferences for immediate versus delayed rewards, self-regulation deficits, and may manifest in higher levels of risk taking behaviors specifically (Gvion, Levi-Belz, Hadlaczky, & Apter, 2015). A robust literature base has indicated that greater engagement in risky behavior has been strongly tied to suicide attempts in young people (McHugh et al., 2019). For example, one study found that adolescents who engaged in impulsive, risky behaviors, including greater drug use, drinking, risky sexual behavior, or/and violence, reported more frequent suicide attempts (Thullen, Taliaferro, & Muehlenkamp, 2016). Other work has shown that higher levels of sexual risk taking, tobacco/alcohol use, illicit drug use, and delinquent acts among adolescents were positively associated with suicide attempt when each of these variables was examined in separate models (Ammerman, Steinberg, & McCloskey, 2018).
There is also some evidence that genetic liability for engaging in risky behaviors may be associated with increased risk for suicide attempt. Like suicide behaviors, individual variation in risky behavior engagement is heritable and has distinct genetic underpinnings (Nicolaou & Shane, 2020). Indeed, a recent GWAS conducted in European ancestry samples identified numerous genetic loci implicated in four risky behaviors, including number of sexual partners, drinks per week, lifetime history of smoking, and automobile speeding propensity (Karlson Línner et al., 2019). In terms of the genetic association between risky behaviors and suicide attempt, research has observed moderate positive genetic correlations between risk tolerance (i.e., one’s preference for taking risks) and suicide attempts (rg = 0.33) (Colbert et al., 2021). Similarly, using a twin design, other research has indicated that close to 40% of the genetic variation in suicidal behaviors was shared with impulsive-aggressive behaviors (Orri et al., 2020). To our knowledge, no research has examined whether genetic propensity for engaging in risky behaviors is associated with suicide attempt, something the current study sought to examine.
1.4 |. Sex differences
It is likely that sex differences exist in the association between depression and risky behavior polygenic risk and phenotypic suicide attempt. For example, a number of studies have indicated differences in the base rates of suicide attempt based on sex (Lewinsohn, Rohde, Seeley, & Baldwin, 2001). Genetic risk for suicidal behavior has also been observed to differ depending on participant sex, with research demonstrating higher heritability estimates (h2 = .74) for self-reported thoughts of harm and suicide among females compared with males (h2 = .45) (Althoff et al., 2012). Similarly, twin studies have indicated greater differences in the concordance rates of suicide between MZ and DZ twins among females (MZ concordance rate = 11%, DZ concordance rate = 0%) compared with males (MZ concordance rate = 3%, DZ concordance rate = 2%), suggesting greater genetic effects among women (Pedersen & Fiske, 2010).
1.5 |. Current study
In the current study, we examine whether depression and risky behavior genetic predispositions (assessed via PRS) are associated independently with reported lifetime suicide attempt from adolescence into young adulthood among African Americans. In addition, we examine whether the relations between risky behavior genetic propensity and lifetime suicide attempt differ based on levels of depression polygenic risk and vice versa, consistent with research indicating that the combination of phenotypic depression and risky behavior influences suicide attempt risk (Hart et al., 2017). Determining whether depression and risky behavior polygenic risk scores (derived from European ancestry discovery samples) are associated with suicide attempt may inform etiological models of suicidal behaviors and eventually result in more effective screening for and prevention of suicide attempts among African American adolescents and young adults.
2 |. METHOD
2.1 |. Participants
The study’s analytic sample was drawn from three cohorts of participants in a series of randomized controlled trials of elementary school-based universal preventive interventions targeting early aggression and academic achievement. The trials were carried out within a single urban school district in a Mid-Atlantic region of the United States when children were in first grade. In terms of inclusion criteria, children had to attend one of the participating schools, be in first grade, and be in a mainstream as opposed to a self-contained special education classroom. The interventions employed in the first two cohorts (first trial) are detailed in Dolan et al. (1993) whereas Ialongo et al. (1999) describes the interventions employed in the third cohort (second trial). Although the targets of the interventions were the same in the first and second trials, the nature of the interventions varied. Across all three cohorts, the interventions were administered universally, or classroom-wide, and participants were followed from first grade to young adulthood (~age 30). The trials and follow-up studies were approved by a University Institutional Review Board. Participants provided written informed consent upon reaching age 18, whereas prior to age 18, parents provided written consent and participants gave verbal assent.
Three-thousand one-hundred and ten first graders were available for recruitment across the three cohorts, of which 2,194 were African American. Of the 2,194 African Americans, 1,157 comprised the analytic sample for this study, given that they had successful genome-wide assays and were asked at least once in a follow-up interview in adolescence or young adulthood whether they had ever attempted suicide in their lifetime. Participant demographic information for the analytic sample is outlined in Table 1. Details regarding DNA collection and processing can be found in the Appnedix S1.
TABLE 1.
Sample characteristics (N = 1,157)
| Characteristic | n |
|---|---|
| Sex | |
| Male | 527 |
| Female | 630 |
| Free/reduced-priced luncha | |
| Yes | 223 |
| No | 881 |
| Intervention | |
| Yes | 600 |
| No | 557 |
| Cohort identification | |
| Cohort 1 | 361 |
| Cohort 2 | 315 |
| Cohort 3 | 481 |
| Suicide attempt | |
| Whole sample | |
| Yes | 178 |
| No | 979 |
| Male | |
| Yes | 67 |
| No | 460 |
| Female | |
| Yes | 111 |
| No | 519 |
Free/reduced priced lunch was assessed when participants were in first grade.
With respect to differences in first grade demographic characteristics (sex and free/reduced priced lunch status [a proxy for family income]) between the analytic sample (i.e., 1,157 participants in cohorts 1, 2, and 3) and those who did not provide DNA and/or were not interviewed in adolescence or young adulthood, there was a greater proportion of females in the analytic sample (analytic sample, 55%, vs. non-analytic sample, 45%, p < .001). In addition, although there was a greater percentage of those assigned to one of the two intervention conditions in the analytic sample as compared with the control condition, the difference was not statistically significant at the p ≤ .05 level (analytic sample, 52% vs. non-analytic sample, 48%).
2.2 |. Measures
2.2.1 |. Lifetime suicide attempt
Participants were interviewed periodically in adolescence and young adulthood and were asked if they had ever attempted suicide in their lifetime. We created a binary variable that reflected ever attempting suicide in one’s lifetime (0 = no lifetime attempt of suicide 1; 1 = lifetime attempt of suicide).
2.2.2 |. Risky behavior GWAS discovery sample
We generated a risky behavior PRS based on a GWAS (N = 315,894) conducted by Karlson Línner et al. (2019). Using data drawn from the UK Biobank, an adult European ancestry sample, Karlson Línner et al. (2019) conducted a principal component analysis (PCA) involving four risky behaviors, namely drinks per week, automobile speeding propensity, ever been a smoker, and number of sexual partners. The GWAS was conducted on the first PC which accounted for the most variability in the four risky behaviors; summary statistics derived from this GWAS served as the basis from which the risky behavior PRS was generated in the present sample.
2.2.3 |. Depression GWAS discovery sample
Depression PRS were created based on the largest GWAS meta-analysis of depression to date. This meta-analysis GWAS included 807,553 individuals (246,363 cases and 561,190 controls) of European ancestry across several samples (Howard et al., 2019). Summary statistics for the UK Biobank and Psychiatric Genomics Consortium (PGC) cohorts were obtained from the online repository (https://datashare.is.ed.ac.uk/handle/10283/3203). Summary statistics for the 23andMe cohort were obtained under an agreement with 23andMe that protects the privacy of the 23andMe participants (see http://research.23andme.com/collaborate/#dataset-access for more information).
2.2.4 |. Coronary artery disease GWAS discovery sample
To demonstrate specificity of results, we created a “control” PRS based on a GWAS conducted on coronary artery disease (van der Harst & Verweij, 2018). This GWAS included 34,541 cases with coronary artery disease and 261,984 controls drawn from UK Biobank. Results from these analyses are presented in the Appnedix S1.
2.3 |. Polygenic risk scores generation
PRS were generated using LDpred, a recently developed and commonly implemented method that obtains posterior estimates of joint effect sizes of the SNPs by incorporating LD patterns and assumes a spike and slab prior on the effect sizes (Vilhjalmsson et al., 2015). To ensure matched ancestry between the LD reference panel and the GWAS, we used the hg38 conversions genotype data from 1,000 Genomes Phase 3 Reference Panel that includes 498 unrelated European ancestry individuals. As suggested by Vilhjalmsson et al. (2015), we selected the 1,111,236 HapMap3 SNPs with minor allele frequency (MAF) greater than 0.01 that were also available in the 1,000 Genomes Phase 3 Reference Panel and the genotype data of the target participants. We set the LD radius (i.e., the number of SNPs for which we adjusted LD on each side of a given SNP) to 400 (approximately the total number of SNPs divided by 3,000) as suggested by Vilhjalmsson et al. (2015), and considered pf = 1, 0.3, 0.1, 0.03, 0.01, 0.003, 0.001, 0.0003, and 0.0001 as the candidates for the fraction of causal variants.
Our cohort was split into two groups: a test dataset of European American ancestry (N = 337; 62 of whom reported a lifetime suicide attempt), which was used to select the causal SNP fraction parameter pf, and a validation dataset (i.e., our target sample), which included only the African American participants comprising the analytic sample in the present study (N = 1,157; see Table 1 for lifetime suicide attempt frequency in our analytic sample). To tune the PRS threshold pf, we calculated various PRS for the individuals in the test dataset under different values of pf, and selected the value of pf that led to the highest squared correlation (r2) between the corresponding PRS and the phenotype. The proportion of causal SNPs for each PRS that was most predictive of lifetime suicide attempt in the test dataset is as follows: risky behavior PRS at pf = 0.01, depression PRS at pf = 0.0001, and coronary artery disease PRS at pf = 0.03. We examined the performance of these PRS thresholds in the validation dataset (i.e., our African American target analytic sample).
2.4 |. Population stratification
To reduce population stratification, we used principal component analysis to create the population stratification control variables in PLINK 2.0 (Chang et al., 2015). The first 10 genetic ancestry principal components (PCs) were included as covariates in our analyses in an attempt to control for population stratification.
3 |. STATISTICAL ANALYSES
The primary analyses were conducted in R version 4.0.2 (R Core Team, 2020). Logistic regression models were conducted in the whole sample to examine (a) the main effect of the covariates (intervention status, free/reduced priced lunch status and the top 10 genetic ancestry PCs), (b) the main effects of the risky behavior PRS and depression PRS adjusted for the covariates referenced above, and (c) the interaction between the risky behavior PRS and depression PRS in relation to lifetime suicide attempt controlling for the main effects of the risky behavior PRS, depression PRS, and covariates referenced above. To further characterize the relationship between phenotypic lifetime suicide attempt and risky behavior PRS and depression PRS levels, we plotted the percentage of individuals who endorsed a lifetime suicide attempt across deciles of the polygenic risk scores.
Given sex differences in heritability of suicide attempt observed among males and females (Althoff et al., 2012; Pedersen & Fiske, 2010), we conducted interaction analyses involving participant sex and the PRS. A model including two, 2-way interactions (i.e., participant sex × risky behavior PRS and participant sex × depression PRS interaction) was conducted; this model controlled for the covariates referenced above and the main effects of the risky behavior PRS and depression PRS. An additional model was run including an interaction between participant sex, the risky behavior PRS, and depression PRS, which adjusted for the covariates, main effects of both PRS, as well as three, two-way interactions (i.e., sex × risky behavior PRS; sex × depression PRS; risky behavior PRS × depression PRS). Given the possibility we may have been underpowered to detect sex differences, we also conducted a series of analyses stratified by participant sex to further examine whether different results patterns emerged based on participant sex. Across models, we report area under the ROC curve (AUC) which is a metric ranging from 0.5 (poor separation) to 1 (perfect separation) and indicates the probability that the model will correctly categorize individuals (Streiner & Cairney, 2007).
In the presence of a significant interaction, the probability of a lifetime suicide attempt at different values of the depression and risky behavior PRS were plotted, holding all other covariates constant at their sample means. Model coefficients were extracted and multiplied by the sample mean for covariates and by a range of values from −3 to 3 in 0.1 increments for the PRS variables; these values were summed to obtain logit values for lifetime suicide attempt and then converted to the probability scale and plotted. Three lines were plotted including (a) one corresponding to possible depression PRS values while holding the risky behavior PRS constant at 0 (the sample mean after standardization), (b) one corresponding to possible risky behavior PRS values while holding the depression PRS constant at 0, and (c) one corresponding to simultaneous changes in both the depression and risky behavior PRS. For significant interactions, confidence and prediction intervals were plotted to aid in interpretation of the interaction effects.
4 |. RESULTS
One hundred and seventy-eight individuals (15.4% of the sample) reported a lifetime suicide attempt by their young adult years. Approximately 13% of African American males (n = 67) reported attempting suicide in their lifetime, whereas close to 18% of African American females endorsed attempting suicide in their lifetime (n = 111). Results from the primary analyses are presented below.
4.1 |. Whole sample
The initial covariates-only model had an AUC of 0.59 (95% CI: 0.55, 0.64). There was a main effect of participant sex such that males were less likely to attempt suicide relative to females (aOR = 0.67, 95% CI: 0.48, 0.95, p = .024). The second model added main effects for the risky behavior PRS and depression PRS to the model which increased the AUC to 0.60 (95% CI: 0.55, 0.65). Main effects of the risky behavior PRS (aOR = 1.06, 95% CI: 0.88, 1.28, p = .556) and depression PRS (aOR = 1.11, 95% CI: 0.94, 1.32, p = .232) with suicide attempt were not significant (Table 2). Distributions are shown in Figure 1 for descriptive purposes only. A greater percentage of individuals with a lifetime history of suicide attempt were in the top decile of depression polygenic risk. With regards to risky behavior polygenic risk, the percentage of individuals with a lifetime history of suicide attempt was higher among individuals in the lowest decile of risky behavior PRS, as well as higher deciles of risky behavior PRS (Figure 1).
TABLE 2.
Summary of associations between the risky behavior PRS and depression PRS with suicide attempt among the whole sample
| aOR (95% CI) | p-value | |
|---|---|---|
| PRS main effects | ||
| Risky behavior PRS | 1.06 (0.88, 1.28) | .556 |
| Depression PRS | 1.11 (0.94, 1.32) | .232 |
| PRS interaction | ||
| Risky behavior PRS × depression PRS | 0.85 (0.72, 0.99) | .034 |
| 2-way sex × single PRS interactions | ||
| Sex × risky behavior PRS | 1.39 (0.98, 1.97) | .063 |
| Sex × depression PRS | 0.87 (0.62, 1.24) | .444 |
| 3-way sex × PRS interaction | ||
| Sex × risky behavior PRS × depression PRS | 0.78 (0.54, 1.10) | .159 |
FIGURE 1.
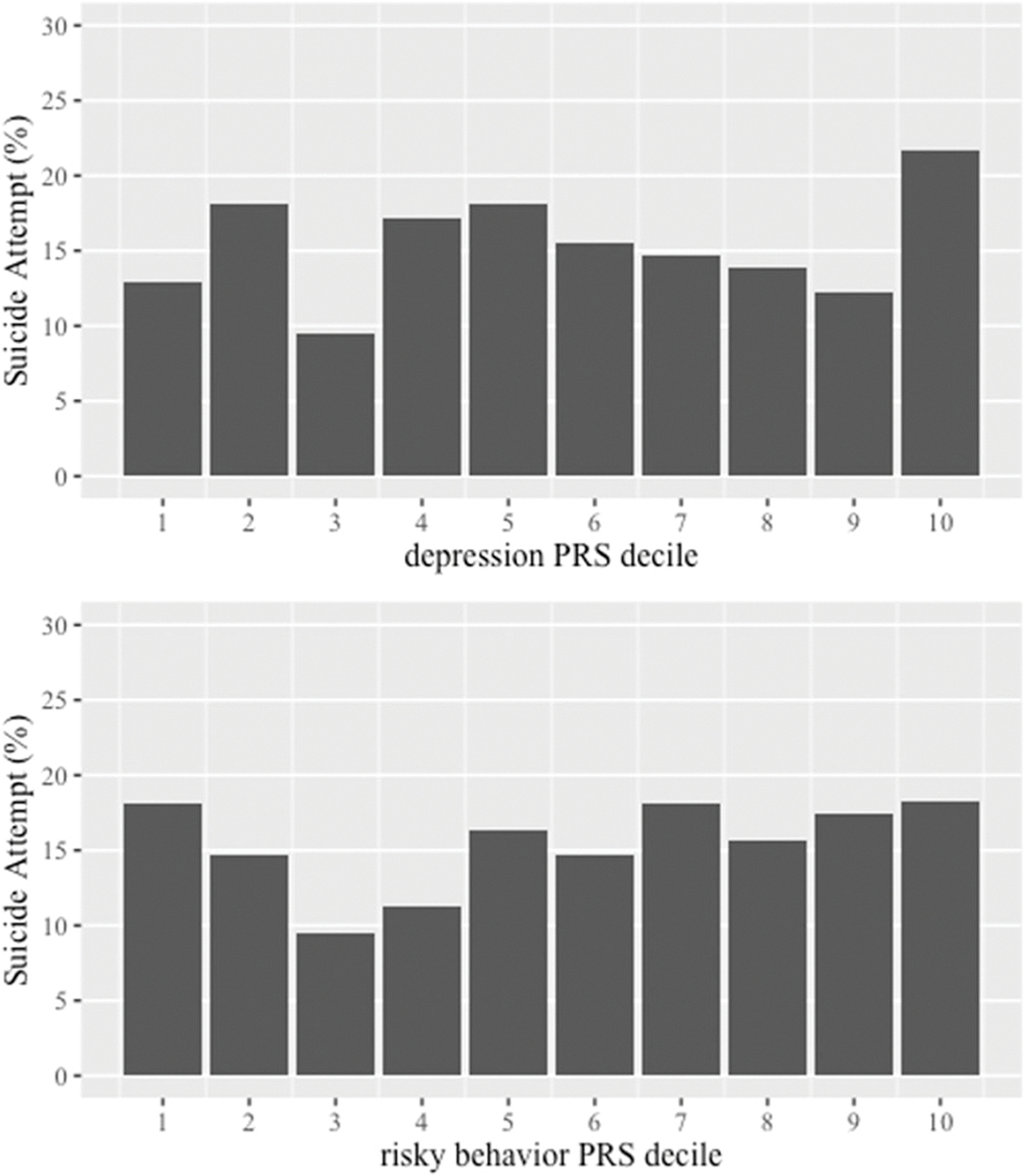
Prevalence of suicide attempt based on deciles of the risky behavior PRS and depression PRS among the whole sample
The addition of the risky behavior × depression PRS interaction term to the model increased the AUC to 0.61 (95 CI: 0.57, 0.66). There was a statistically significant interaction between the risky behavior PRS and depression PRS in relation to lifetime suicide attempt (p = .034, Table 2). As illustrated in Figure 2, when depression PRS was low (−1), the aOR associated with risky behavior PRS was 1.31 (95% CI: 1.04, 1.64); when depression PRS was average (0), the aOR was 1.11 (95% CI: 0.93, 1.31); and when depression PRS was high (1), the aOR was 0.94 (95% CI: 0.74, 1.18). These findings are further supported in Figure 3a,b which display the confidence intervals and prediction intervals for this interaction.
FIGURE 2.
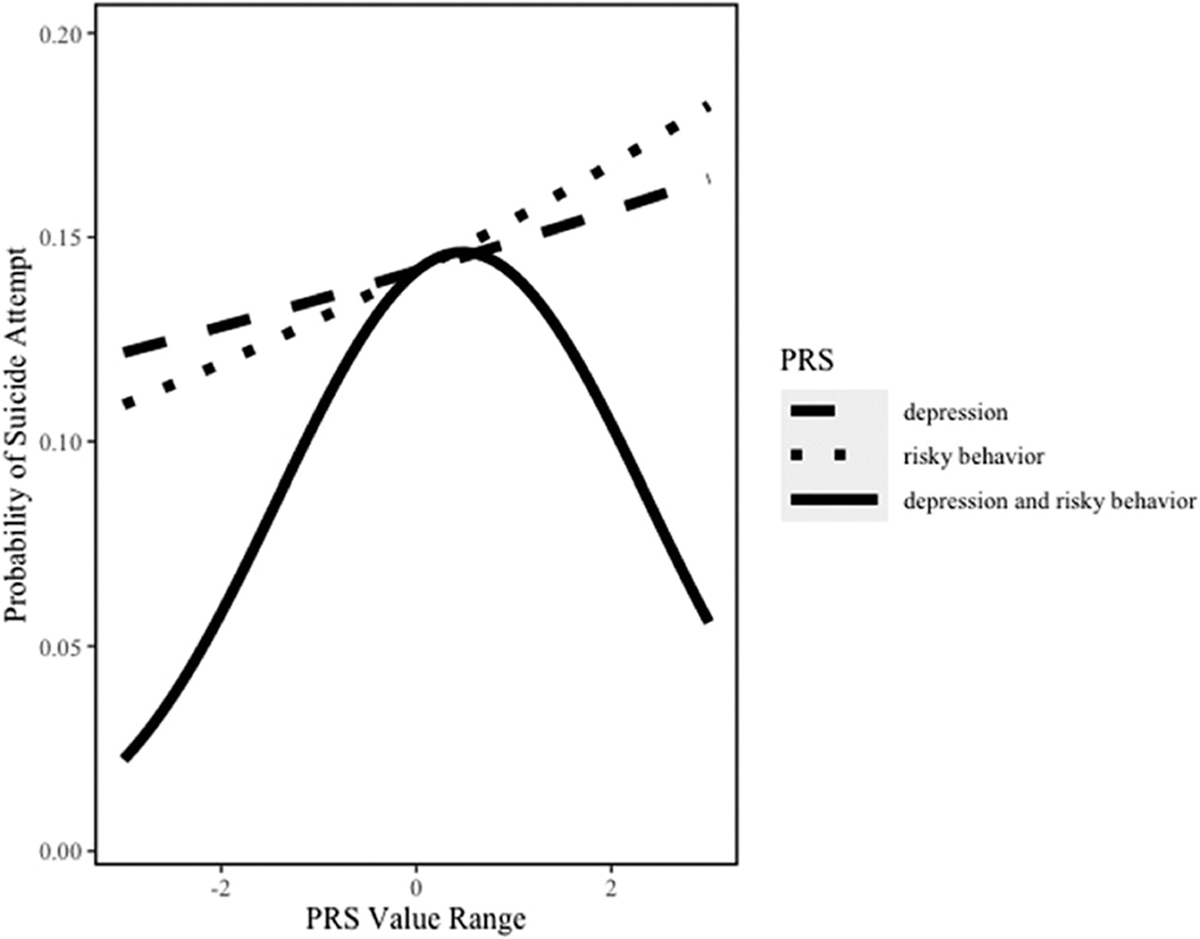
Relation between risky behavior PRS and depression PRS in relation to suicide attempt among the whole sample. *All lines plot the model estimated probability of suicide attempt while varying the indicated PRS—either depression only, risky behavior only, or both simultaneously—and holding all other covariates constant at their sample means
FIGURE 3.
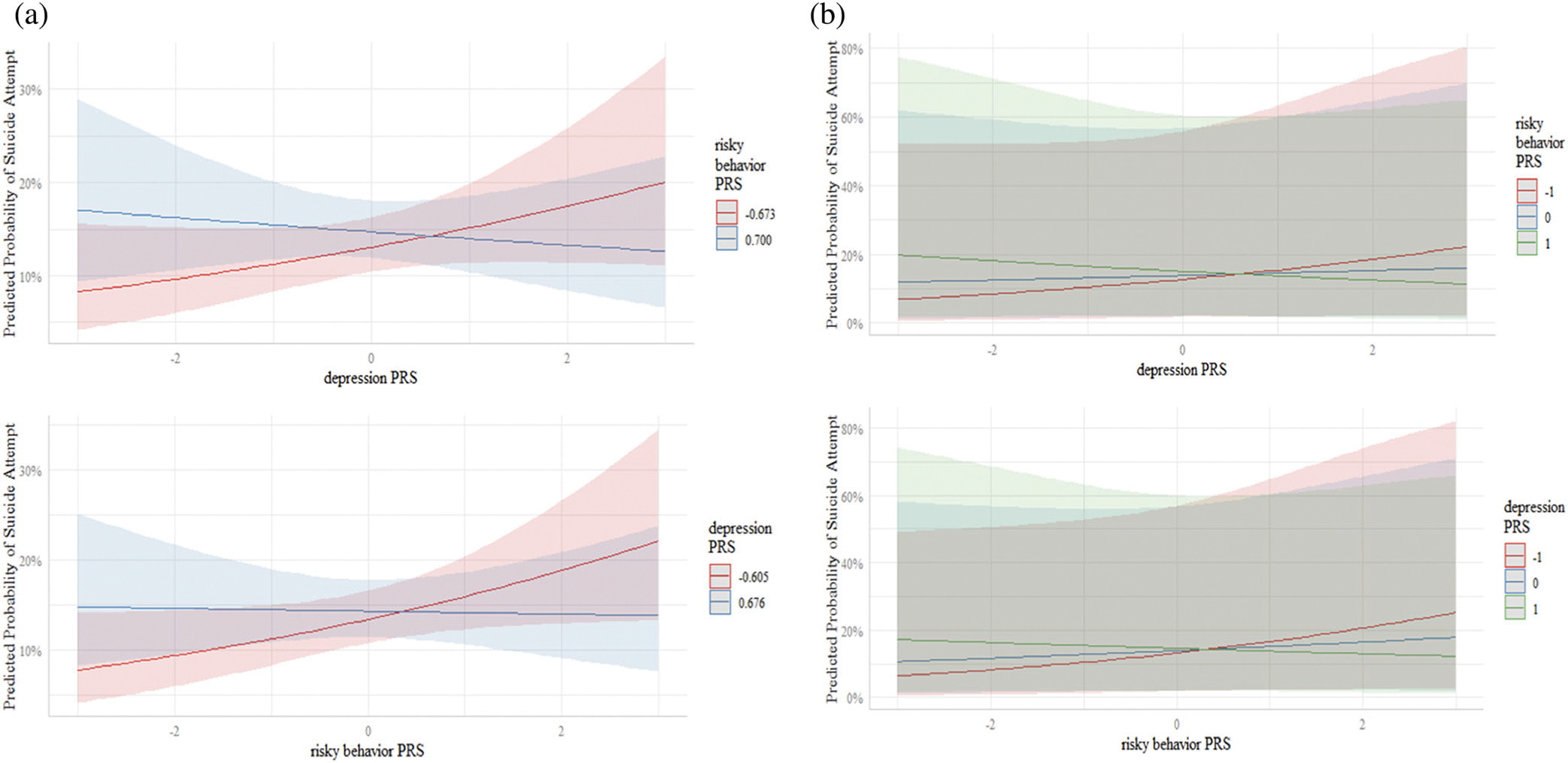
(a) Predicted probability of suicide attempt (with 95% confidence intervals) as a function of depression PRS with risky behavior PRS fixed at the 25th and 75th quartiles, and vice versa. (b) Predicted probability of suicide attempt (with 95% prediction intervals) as a function of depression PRS with risky behavior PRS fixed at 1 SD below the mean, the sample mean, and 1 SD above the mean, and vice versa
Next, two-way interactions involving participant sex and the risky behavior PRS and depression PRS were added to the model. There was a marginally significant interaction between participant sex and the risky behavior PRS (aOR = 1.39, 95% CI: 0.98, 1.97, p = .063) such that males with a higher risky behavior PRS were more likely to report a lifetime suicide attempt relative to females (Table 2). There was no evidence of a statistically significant interaction between participant sex and the depression PRS in relation to lifetime suicide attempt (aOR = 0.87, 95% CI: 0.62, 1.24, p = .444). The addition of the sex and PRS interactions did not result in an AUC improvement.
Finally, the three-way interaction between participant sex, the risky behavior PRS, and depression PRS was not statistically significant (p = .159). The addition of this interaction increased the AUC to 0.62 (95% CI: 0.57, 0.67).
4.2 |. Analyses stratified by sex
4.2.1 |. African American males
The model that only included the covariates had an AUC of 0.64 (95 CI: 0.57, 0.71). The AUC of the model increased to 0.66 (95% CI: 0.59, 0.72) after including the risky behavior PRS and depression PRS. As shown in Figure 4, a greater percentage of males who attempted suicide in their lifetime was in the top decile of risky behavior polygenic risk, whereas the percentage of males who attempted suicide in their lifetime was relatively similar regardless of their depression PRS levels. The risky behavior PRS was significantly associated with suicide attempt such that males with a higher risky behavior PRS were more likely to report a lifetime suicide attempt (aOR = 1.43, 95% CI: 1.07, 1.91, p = .016; Table 3). There was no statistically significant evidence that the depression PRS was associated with a lifetime suicide attempt (aOR = 0.94, 95% CI: 0.68, 1.30, p = .728).
FIGURE 4.
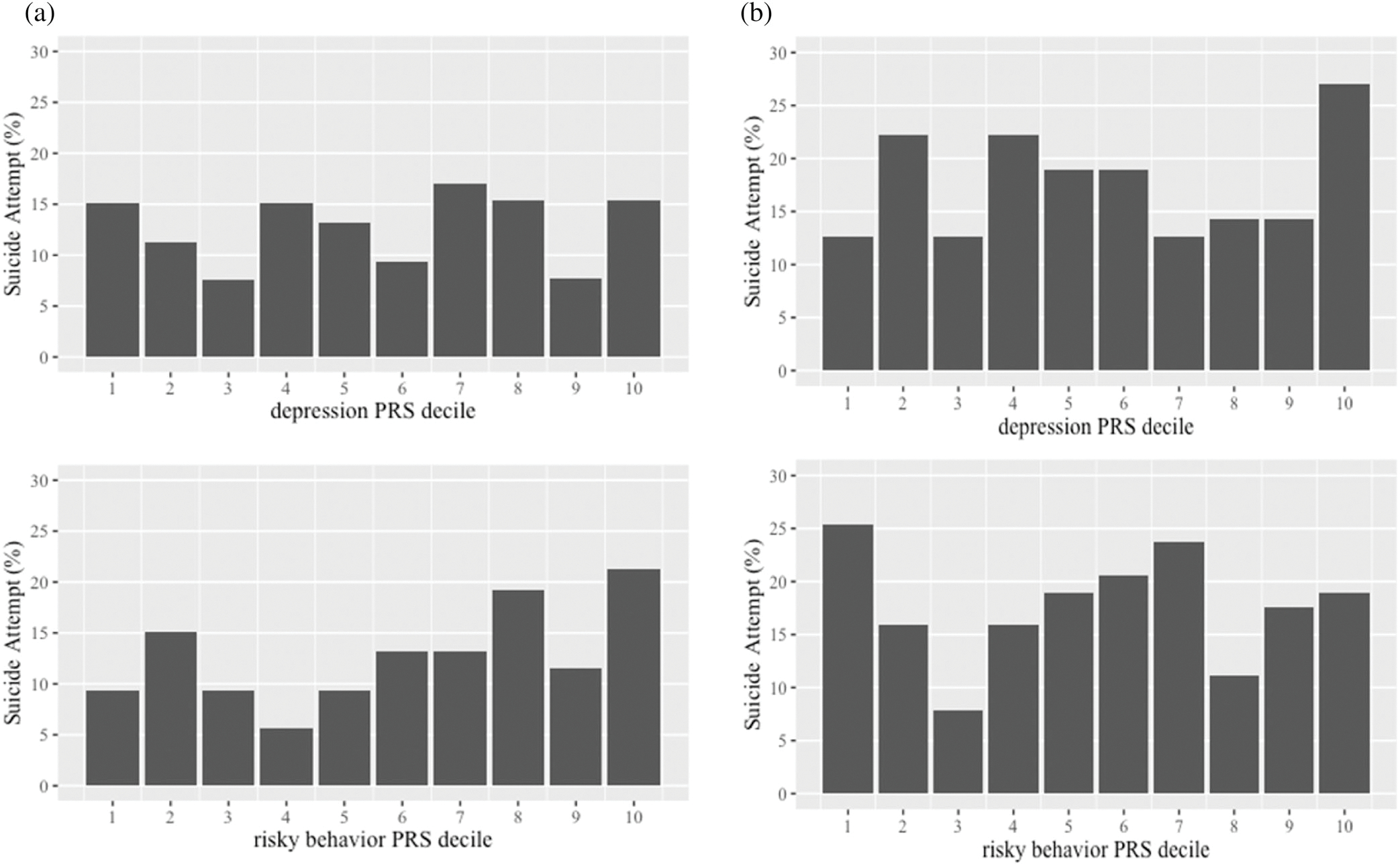
(a) Prevalence of suicide attempt based on deciles of the risky behavior PRS and depression PRS among African American males, and (b) African American females
TABLE 3.
Stratified analyses involving associations between the risky behavior PRS and depression PRS with suicide attempt among males and females
| aOR (95% CI) | p-value | |
|---|---|---|
| Males | ||
| PRS main effects | ||
| Risky behavior PRS | 1.43 (1.07, 1.91) | .016 |
| Depression PRS | 0.94 (0.68, 1.30) | .728 |
| PRS interaction | ||
| Risky behavior PRS × depression PRS | 0.70 (0.51, 0.95) | .023 |
| Females | ||
| PRS main effects | ||
| Risky behavior PRS | 0.98 (0.79, 1.23) | .883 |
| Depression PRS | 1.15 (0.90, 1.47) | .251 |
| PRS interaction | ||
| Risky behavior PRS × depression PRS | 0.93 (0.77, 1.14) | .478 |
The AUC of the model with the addition of the risky behavior PRS × depression PRS interaction term was 0.68 (95% CI: 0.61, 0.74). There was evidence of a statistically significant interaction between the risky behavior PRS and the depression PRS (p = .023, Table 3). As shown in Figure 5, the risky behavior PRS was associated with an increased risk of suicide attempt when the depression PRS was low to moderate; however, this association disappeared and appeared to invert at higher levels of depression PRS. When depression PRS was low (−1), the aOR associated with risky behavior PRS was 1.98 (95% CI: 1.30, 2.99); when depression PRS was average (0), the aOR was 1.38 (95% CI: 1.03, 1.86); and when depression PRS was high (1), the aOR was 0.97 (95% CI: 0.63, 1.49). These findings are further supported in Figure 6a,b which display the confidence intervals and prediction intervals of this interaction.
FIGURE 5.
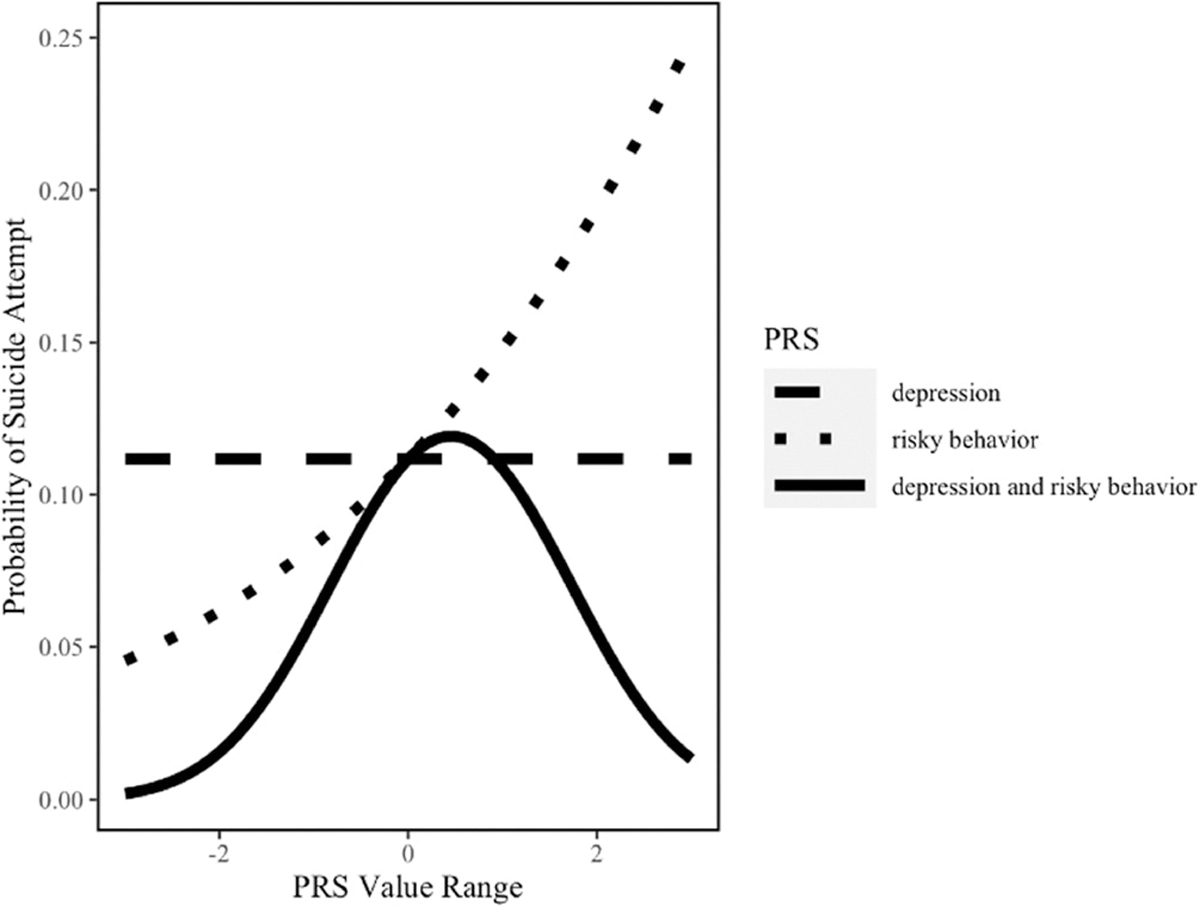
Relation between depression PRS and risky behavior PRS in relation to suicide attempt among African American males based on stratified analyses. *All lines plot the model estimated probability of suicide attempt while varying the indicated PRS—either depression only, risky behavior only, or both simultaneously—and holding all other covariates constant at their sample means
FIGURE 6.
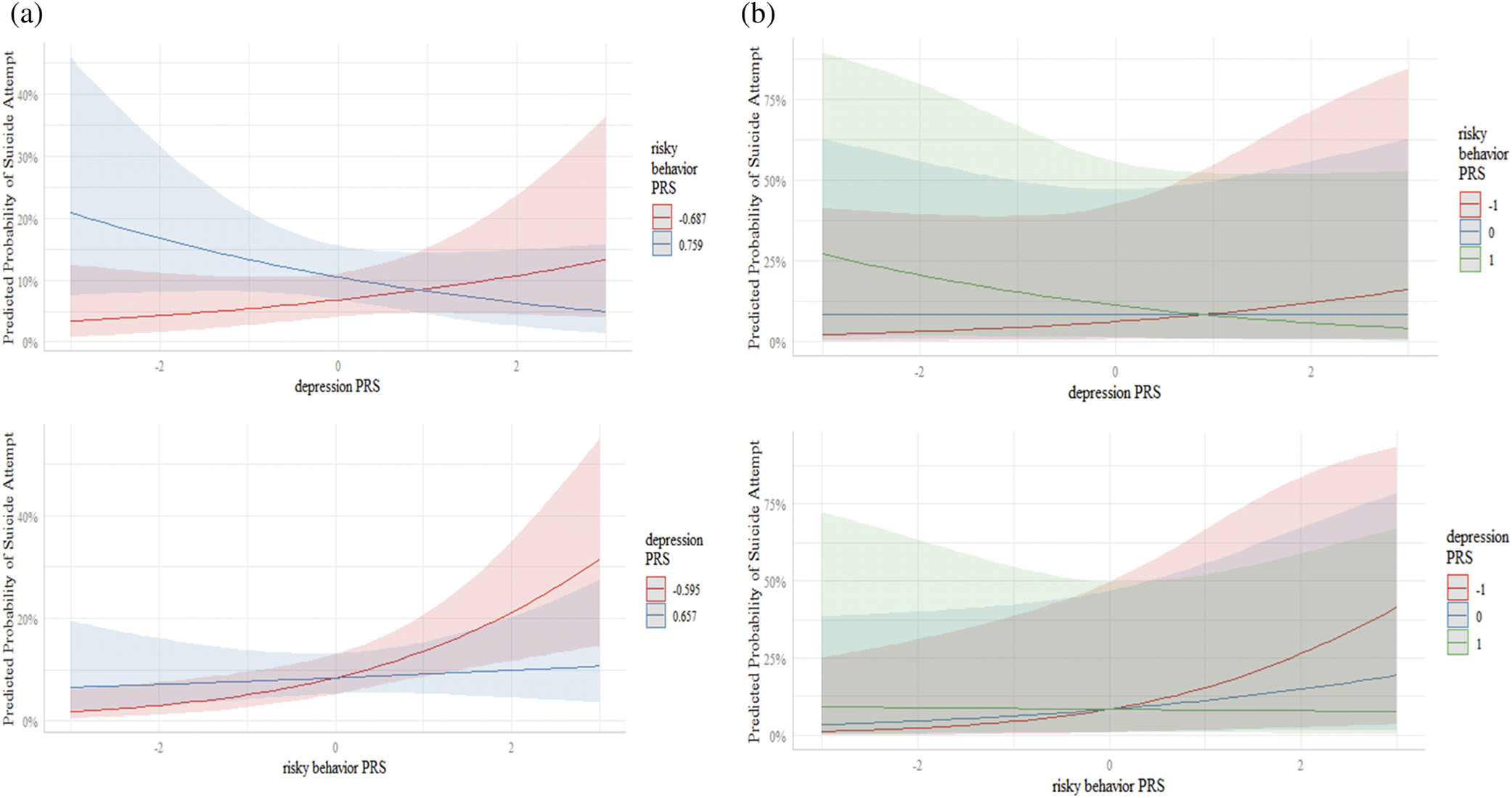
(a) Predicted probability of suicide attempt (with 95% confidence intervals) as a function of depression PRS with risky behavior PRS fixed at the 25th and 75th quartiles, and vice versa. (b) Predicted probability of suicide attempt (with 95% prediction intervals) as a function of depression PRS with risky behavior PRS fixed at 1 SD below the mean, the sample mean, and 1 SD above the mean, and vice versa
4.2.2 |. African American females
The model that only included the covariates had an AUC of 0.61 (95% CI: 0.55, 0.67). The model had an increased AUC of 0.62 (95% CI: 0.56, 0.68) after further including the risky behavior PRS and depression PRS. As shown in Figure 4b, a greater percentage of females who attempted suicide was in the lowest decile of risky behavior polygenic risk; the percentage of females who attempted suicide was greatest in the highest decile of depression PRS. As shown in Table 3, there was no statistically significant evidence that the (a) risky behavior PRS (aOR = 0.98, 95% CI: 0.79, 1.21, p = .883), (b) depression PRS (aOR = 1.15, 95% CI: 0.90, 1.47, p = .251), or (c) interaction between the risky behavior PRS and depression PRS was associated with lifetime suicide attempt (aOR = 0.93, 95% CI: 0.77, 1.14, p = .478); the AUC for the model involving the interaction term was 0.62 (95% CI: 0.57, 0.68).
5 |. DISCUSSION
Substantial progress in genomic discoveries has resulted in the uncovering of genetic loci implicated in suicide attempt and related phenotypes (Docherty et al., 2020; Mullins et al., 2014). However, African Americans have been vastly underrepresented in this work with genetic advances mostly benefiting individuals of European ancestry (Sirugo, Williams, & Tishkoff, 2019). This fact, along with the increasing rate of suicidal behaviors among young African Americans (Lindsey et al., 2019), prompted us to investigate whether genetic propensity for depressive symptoms and risky behavior independently and jointly were associated with a lifetime report of a suicide attempt in an urban, African American young adult sample.
There was a significant interaction between the depression PRS and risky behavior PRS in relation to a lifetime suicide attempt among the whole sample and among males (observed in stratified analyses). Notably, the inclusion of the interaction term in the analytic models involving the whole sample and males improved prediction, as evidenced by a higher AUC, compared with examining the risky behavior PRS and depression PRS in isolation, highlighting the importance of considering polygenic risk for depression and risky behavior together. These findings are consistent with some work indicating that at low levels of phenotypic depression, engagement in risky behaviors (i.e., more frequent alcohol use) was associated with augmented risk for suicide attempt (McManama O’Brien, Becker, Spirito, Simon, & Prinstein, 2014). Individuals with lower depression polygenic risk and higher risky behavior polygenic risk may manifest lower levels of phenotypic depressive symptoms (e.g., sadness, anhedonia, and lethargy); as such, these individuals may possess the motivation to engage in risk taking behaviors that confer risk for attempting suicide. The current findings may also capture a distinct subgroup of individuals, impulsive suicide attempters, that has been previously identified in the literature (Brown, Overholser, Spirito, & Fritz, 1991; Simon et al., 2001). Previous work has shown that impulsive suicide attempters often attempt suicide within a relatively short period of time of experiencing an internal or external stressor without having made a plan and generally report lower levels of depressive symptoms (Simon et al., 2001; Wyder & De Leo, 2007). Future work should consider investigating pathways through which the combination of low to moderate depression polygenic risk and higher risky behavior polygenic risk amplify risk for attempting suicide among African American young adults generally and males specifically.
We found statistically significant evidence that the risky behavior PRS was associated with a lifetime suicide attempt among African American males (in stratified analyses), but not African American females. These findings are consistent with the literature suggesting that greater behavioral engagement in risky behaviors (e.g., heavier substance use, greater number of sexual partners) is associated with heightened risk for attempting suicide (Ammerman et al., 2018; King et al., 2001; Thullen et al., 2016). There are a number of explanations regarding the fact that greater genetic propensity for risky behavior exacerbated risk for suicide attempt among males, but not females, including differences in heritability of complex traits and environmental exposures. In terms of the former, research has shown that as individuals age from adolescence to young adulthood, heritability of risk taking increases by over 55% among males, but not females (Anokhin, Golosheykin, Grant, & Heath, 2012). In terms of the latter, males and females are typically socialized differently with males often encouraged to be more autonomous and it is more socially sanctioned for males to engage in risk taking practices during adolescence and young adulthood (Pawlowski & Atwal, 2008; Shulman, Harden, Chein, & Steinberg, 2015). In these contexts, it is possible that greater genetic propensity for risky behavior serves as a diathesis among males that has downstream consequences for adverse health outcomes, such as attempting suicide (Mann, Waternaux, Haas, & Malone, 1999). For example, individuals with heightened genetic propensity to engage in risky behaviors may be more likely to act impulsively with little reflection on the consequences of their behavior, which may exacerbate risk for attempting suicide (Turecki & Brent, 2016).
It is notable that there was no statistically significant evidence that the depression PRS was associated with a lifetime suicide attempt. There are several explanations to account for these null findings. It is possible that individuals with higher depression PRS manifest higher levels of amotivation or fatigue that may impede the carrying out of a suicide attempt, in line with some research indicating that recovery from major depression is associated with increased risk for suicide attempt (Mittal, Brown, & Shorter, 2009). Another potential explanation might be the nature of the sample studied; indeed, the current study sample is a low-income, urban population who experience numerous stressors including exposure to stressful life events, violence, and racial discrimination, which have been shown to independently predict suicide behavior manifestation (Kaslow et al., 2005). The diathesis-stress model suggests that outcomes like depression and suicidal behavior are the result of the intersection of internal vulnerabilities and external stressors, but in the presence of extremely high stressors, even people with minimal vulnerabilities may become depressed. Stated another way, genetic vulnerability to depression may not be a strong predictor of phenotypic depression in a population that universally experiences high levels of environmental stressors, and thus, the association between the depression PRS and suicidal behavior may become attenuated. Last, previous studies have indicated that depression PRS often account for a very small percentage of the variance in suicide attempt (<1%) (Maciejewski et al., 2017; Mullins et al., 2014), which coupled with the ancestry mismatch between the target and discovery samples, may have resulted in statistically null main associations between the depression PRS and suicide attempt that we observed.
Findings should be interpreted in the context of some limitations. Although the current study is one of very few to examine genetic correlates of suicide attempt in an African American sample, we did not consider contextual influences that may affect suicidal behavior manifestation. Of course, genetics alone is not sufficient in explaining phenomena as complex as suicide attempts and should be considered in concert with a host of other risk factors such as family history, stressful live events, exposure to racial and community trauma, and so on (Zalsman, 2010). Another limitation is that the depression and risky behavior PRS were constructed from summary statistics derived from European ancestry GWAS in the absence of African American GWAS. It is well established that allele frequencies can differ across ancestral groups due to differences in population demographic history (i.e., migration, isolation) (De La Vega & Bustamante, 2018; Marnetto et al., 2020). Consequently, genetic loci implicated in a given phenotype in one population may not be relevant to another population particularly when genetic divergence between populations increases, which may reduce the portability and predictive utility of PRS to other ancestral groups (Martin et al., 2019). This strategy can also suffer from low predictive power due to heterogeneity in LD structure and effect size distribution of genetic variants across ancestries (Martin et al., 2019). For these reasons, caution should be taken regarding the interpretation of our results and the implications of our work for suicide prevention efforts. Moreover, the inclusion of more diverse ancestral populations in GWAS, and work such as that presented here that examines the transferability of PRS derived from primarily European ancestry samples, is crucial to ensuring equity in the application of precision medicine initiatives.
Advances in statistical and genomic methodologies offer the promise of creating more accurate evidence-based risk prediction models of suicide and may ideally help clinicians in the identification of subgroups of individuals at heightened risk for attempting suicide. However, before this happens, substantial research is needed to determine how to best integrate genomic information into clinical care, and efforts aimed at prioritizing the inclusion of diverse populations in genomics research are surely needed.
Supplementary Material
ACKNOWLEDGEMENTS
The authors would like to acknowledge support from the following NIDA grants (R01DA044184-02S1; DA11796 MH57005; DA 04392; DA 09897) and NIMH grants (P50 MH 38725; R01 MH 42968).
Footnotes
CONFLICT OF INTEREST
The authors report no conflicts of interest.
SUPPORTING INFORMATION
Additional supporting information may be found online in the Supporting Information section at the end of this article.
DATA AVAILABILITY STATEMENT
Data are not publicly available, but are available upon request. Persons interested in accessing the data could contact Dr. Nicholas Ialongo (nialong1@jhmi.edu).
REFERENCES
- Althoff RR, Hudziak JJ, Willemsen G, Hudziak V, Bartels M, & Boomsma DI (2012). Genetic and environmental contributions to self-reported thoughts of self-harm and suicide. American Journal of Medical Genetics. Part B, Neuropsychiatric Genetics, 159B(1), 120–127. 10.1002/ajmg.b.32010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ammerman BA, Steinberg L, & McCloskey MS (2018). Risk-taking behavior and suicidality: The unique role of adolescent drug use. Journal of Clinical Child and Adolescent Psychology, 47(1), 131–141. 10.1080/15374416.2016.1220313 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anokhin AP, Golosheykin S, Grant J, & Heath AC (2012). Heritability of risk-taking in adolescence: A longitudinal twin study. Twin Research and Human Genetics, 12(4), 366–371. 10.1375/twin.12.4.366 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baldessarini RJ, & Hennen J (2004). Genetics of suicide: An overview. Harvard Review of Psychiatry, 12(1), 1–13. 10.1080/10673220490425915 [DOI] [PubMed] [Google Scholar]
- Bostwick JM, Pabbati C, Geske JR, & McKean AJ (2016). Suicide attempt as a risk factor for completed suicide: Even more lethal than we knew. The American Journal of Psychiatry, 173(11), 1094–1100. 10.1176/appi.ajp.2016.15070854 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bostwick JM, & Pankratz VS (2000). Affective disorders and suicide risk: A reexamination. The American Journal of Psychiatry, 157(12), 1925–1932. [DOI] [PubMed] [Google Scholar]
- Brent DA, Melhem NM, Oquendo M, Burke A, Birmaher B, Stanley B, … Mann JJ (2015). Familial pathways to early-onset suicide attempt: A 5.6-year prospective study. JAMA Psychiatry, 72(2), 160–168. 10.1001/jamapsychiatry.2014.2141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bridge JA, Goldstein TR, & Brent DA (2006). Adolescent suicide and suicidal behavior. Journal of Child Psychology and Psychiatry, 47, 372–394. 10.1111/j.1469-7610.2006.01615.x [DOI] [PubMed] [Google Scholar]
- Brown LK, Overholser J, Spirito A, & Fritz GK (1991). The correlates of planning in adolescent suicide attempts. Journal of the American Academy of Child & Adolescent Psychiatry, 30(1), 95–99. 10.1097/00004583-199101000-00014 [DOI] [PubMed] [Google Scholar]
- Chang CC, Chow CC, Tellier LC, Vattikuti S, Purcell SM, & Lee JJ (2015). Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience, 4, 7. 10.1186/s13742-015-0047-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colbert SMC, Hatoum AS, Shabalin A, Coon H, Nelson EC, Agrawal A, … Johnson EC (2021). Exploring the genetic overlap of suicide-related behaviors and substance use disorders. medRxiv. 10.1101/2021.04.05.21254944 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De La Vega FM, & Bustamante CD (2018). Polygenic risk scores: A biased prediction? Genome Medicine, 10(1), 100. 10.1186/s13073-018-0610-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Docherty AR, Shabalin AA, DiBlasi E, Monson E, Mullins N, Adkins DE, … Coon H (2020). Genome-wide association study of suicide death and polygenic prediction of clinical antecedents. The American Journal of Psychiatry, 177(10), 917–927. 10.1176/appi.ajp.2020.19101025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dolan LJ, Kellam SG, Brown CH, Werthamer-Larsson L, Rebow GW, Mayer LS, … Turkkan JS (1993). The short-term impact of two classroom-based preventive interventions on aggressive and shy behaviors and poor achievement. Journal of Applied Developmental Psychology, 14, 317–345. [Google Scholar]
- Gvion Y, Levi-Belz Y, Hadlaczky G, & Apter A (2015). On the role of impulsivity and decision-making in suicidal behavior. World Journal of Psychiatry, 5(3), 255–259. 10.5498/wjp.v5.i3.255 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hart SR, Van Eck K, Ballard ED, Musci RJ, Newcomer A, & Wilcox HC (2017). Subtypes of suicide attempters based on longitudinal childhood profiles of co-occurring depressive, anxious and aggressive behavior symptoms. Psychiatry Research, 257, 150–155. 10.1016/j.psychres.2017.07.032 [DOI] [PubMed] [Google Scholar]
- Howard DM, Adams MJ, Clarke TK, Hafferty JD, Gibson J, Shirali M, … McIntosh AM (2019). Genome-wide meta-analysis of depression identifies 102 independent variants and highlights the importance of the prefrontal brain regions. Nature Neuroscience, 22(3), 343–352. 10.1038/s41593-018-0326-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ialongo NS, Werthamer L, Kellam SG, Brown CH, Wang S, & Lin Y (1999). Proximal impact of two first-grade preventive interventions on the early risk behaviors for later substance abuse, depression, and antisocial behavior. American Journal of Community Psychology, 27 (5), 599–641. [DOI] [PubMed] [Google Scholar]
- Karlson Línner R, Biroli P, Kong E, Meddens SFW, Wedow R, Fontana MA, … Beauchamp JP (2019). Genome-wide association analyses of risk tolerance and risky behaviors in over 1 million individuals identify hundreds of loci and shared genetic influences. Nature Genetics, 51(2), 245–257. 10.1038/s41588-018-0309-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaslow NJ, Sherry A, Bethea K, Wyckoff S, Compton MT, Grall MB, Scholl L, Price AW, Kellerman A, Thompson N, and Parker R (2005). Social risk and protective factors for suicide attempts in low income African American men and women. Suicide and Life-Threatening Behavior, 35(4), 400–412. [DOI] [PubMed] [Google Scholar]
- King RA, Schwab-Stone M, Flisher AJ, Greenwald S, Kramer RA, Goodman SH, … Gould MS (2001). Psychosocial and risk behavior correlates of youth suicide attempts and suicidal ideation. Journal of the American Academy of Child and Adolescent Psychiatry, 40(7), 837–846. 10.1097/00004583-200107000-00019 [DOI] [PubMed] [Google Scholar]
- Levey DF, Polimanti R, Cheng Z, Zhou H, Nunez YZ, Jain S, … Gelernter J (2019). Genetic associations with suicide attempt severity and genetic overlap with major depression. Translational Psychiatry, 9 (1), 22. 10.1038/s41398-018-0340-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewinsohn P, Rohde P, Seeley JR, & Baldwin CL (2001). Gender differences in suicide attempts from adolescence to young adulthood. Journal of the American Academy of Child and Adolescent Psychiatry, 40(4), 427–434. 10.1097/00004583-200104000-00011 [DOI] [PubMed] [Google Scholar]
- Lim KX, Rijsdijk F, Hagenaars SP, Socrates A, Choi SW, Coleman JRI, … Pingault JB (2020). Studying individual risk factors for self-harm in the UKbiobank: A polygenic scoring and Mendelian randomisation study. PLoS Medicine, 17(6), e1003137. 10.1371/journal.pmed.1003137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindsey MA, Sheftall AH, Xiao Y, & Joe S (2019). Trends of suicidal behaviors among high school students in the United States: 1991–2017. Pediatrics, 144(5), e20191187. 10.1542/peds.2019-1187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lutz PE, Mechawar N, & Turecki G (2017). Neuropathology of suicide: Recent findings and future directions. Molecular Psychiatry, 22(10), 1395–1412. 10.1038/mp.2017.141 [DOI] [PubMed] [Google Scholar]
- Maciejewski DF, Renteria ME, Abdellaoui A, Medland SE, Few LR, Gordon SD, … Verweij KJ (2017). The association of genetic predisposition to depressive symptoms with non-suicidal and suicidal self-injuries. Behavior Genetics, 47(1), 3–10. 10.1007/s10519-016-9809-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maher BS (2015). Polygenic scores in epidemiology: Risk prediction, etiology, and clinical utility. Current Epidemiology Reports, 2(4), 239–244. 10.1007/s40471-015-0055-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mann JJ, Waternaux C, Haas GL, & Malone KM (1999). Toward a clinical model of suicidal behavior in psychiatric patients. American Journal of Psychiatry, 156(2), 181–189. [DOI] [PubMed] [Google Scholar]
- Marnetto D, Parna K, Lall K, Molinaro L, Montinaro F, Haller T, … Pagani L (2020). Ancestry deconvolution and partial polygenic score can improve susceptibility predictions in recently admixed individuals. Nature Communications, 11(1), 1628. 10.1038/s41467-020-15464-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin AR, Gignoux CR, Walters RK, Wojcik GL, Neale BM, Gravel S, … Kenny EE (2017). Human demographic history impacts genetic risk prediction across diverse populations. American Journal of Human Genetics, 100(4), 635–649. 10.1016/j.ajhg.2017.03.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin AR, Kanai M, Kamatani Y, Okada Y, Neale BM, & Daly MJ (2019). Clinical use of current polygenic risk scores may exacerbate health disparities. Nature Genetics, 51(4), 584–591. 10.1038/s41588-019-0379-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- McHugh CM, Chun Lee RS, Hermens DF, Corderoy A, Large M, & Hickie IB (2019). Impulsivity in the self-harm and suicidal behavior of young people: A systematic review and meta-analysis. Journal of Psychiatric Research, 116, 51–60. 10.1016/j.jpsychires.2019.05.012 [DOI] [PubMed] [Google Scholar]
- McManama O’Brien KH, Becker SJ, Spirito A, Simon V, & Prinstein MJ (2014). Differentiating adolescent suicide attempters from ideators: Examining the interaction between depression severity and alcohol use. Suicide & Life-Threatening Behavior, 44(1), 23–33. 10.1111/sltb.12050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mittal V, Brown WA, & Shorter E (2009). Are patients with depression at heightened risk of suicide as they begin to recover? Psychiatric Services, 60(3), 384–386. 10.1176/appi.ps.60.3.38410.1176/ps.2009.60.3.384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mullins N, Perroud N, Uher R, Butler AW, Cohen-Woods S, Rivera M, … Lewis CM (2014). Genetic relationships between suicide attempts, suicidal ideation and major psychiatric disorders: A genome-wide association and polygenic scoring study. American Journal of Medical Genetics. Part B, Neuropsychiatric Genetics, 165B(5), 428–437. 10.1002/ajmg.b.32247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicolaou N, & Shane S (2020). Common genetic effects on risk-taking preferences and choices. Journal of Risk and Uncertainty, 59(3), 261–279. 10.1007/s11166-019-09316-2 [DOI] [Google Scholar]
- Nock MK, Green JG, Hwang I, McLaughlin KA, Sampson NA, Zaslavsky AM, & Kessler RC (2013). Prevalence, correlates, and treatment of lifetime suicidal behavior among adolescents: Results from the national comorbidity survey replication adolescent supplement. JAMA Psychiatry, 70(3), 300–310. 10.1001/2013.jamapsychiatry.55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orri M, Seguin JR, Castellanos-Ryan N, Tremblay RE, Cote SM, Turecki G, & Geoffroy MC (2020). A genetically informed study on the association of cannabis, alcohol, and tobacco smoking with suicide attempt. Molecular Psychiatry. 10.1038/s41380-020-0785-6 [DOI] [PubMed] [Google Scholar]
- Pawlowski B, & Atwal R (2008). Sex differences in everyday risk-taking behavior in humans. Evolutionary Psychology, 6(1), 29–42. [Google Scholar]
- Pedersen NL, & Fiske A (2010). Genetic influences on suicide and nonfatal suicidal behavior: Twin study findings. European Psychiatry, 25(5), 264–267. 10.1016/j.eurpsy.2009.12.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price JH, & Khubchandani J (2019). The changing characteristics of African-American adolescent suicides, 2001–2017. Journal of Community Health, 44(4), 756–763. 10.1007/s10900-019-00678-x [DOI] [PubMed] [Google Scholar]
- R Core Team. (2020). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Retrieved from. https://www.R-project.org/ [Google Scholar]
- Rabinowitz JA, Kuo SI, Felder W, Musci RJ, Bettencourt A, Benke K, … Ialongo NS (2019). Associations between an educational attainment polygenic score with educational attainment in an African American sample. Genes, Brain, and Behavior, 18(5), e12558. 10.1111/gbb.12558 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruderfer DM, Walsh C, Aguirre MW, Tanigawa Y, Ribeiro JD, Franklin JC, & Rivas MA (2020). Significant shared heritability underlies suicide attempt and clinically predicted probability of attempting suicide. Molecular Psychiatry, 25(10), 2422–2430. 10.1038/s41380-018-0326-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shulman EP, Harden KP, Chein JM, & Steinberg L (2015). Sex differences in the developmental trajectories of impulse control and sensation-seeking from early adolescence to early adulthood. Journal of Youth and Adolescence, 44(1), 1–17. 10.1007/s10964-014-0116-9 [DOI] [PubMed] [Google Scholar]
- Simon TR, Swann AC, Powell KE, Potter LB, Kresnow M-J, & O’Carroll PW (2001). Characteristics of impulsive suicide attempts and attempters. Suicide & Life-Threatening Behavior, 32, 49–59. 10.1521/suli.32.1.5.49.24212 [DOI] [PubMed] [Google Scholar]
- Sirugo G, Williams SM, & Tishkoff SA (2019). The missing diversity in human genetic studies. Cell, 177(1), 26–31. 10.1016/j.cell.2019.02.048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Streiner DL, & Cairney J (2007). What’s under the ROC? An introduction to receiver operating characteristics curves. The Canadian Journal of Psychiatry, 52(2), 121–128. [DOI] [PubMed] [Google Scholar]
- Thullen MJ, Taliaferro LA, & Muehlenkamp JJ (2016). Suicide ideation and attempts among adolescents engaged in risk behaviors: A latent class analysis. Journal of Research on Adolescence, 26(3), 587–594. 10.1111/jora.12199 [DOI] [PubMed] [Google Scholar]
- Tomek S, Hooper LM, Church WT 2nd, Bolland KA, Bolland JM, & Wilcox K. (2015). Relations among suicidality, recent/frequent alcohol use, and gender in a black american adolescent sample: A longitudinal investigation. Journal of Clinical Psychology, 71 (6), 544–560. 10.1002/jclp.22169 [DOI] [PubMed] [Google Scholar]
- Turecki G, & Brent DA (2016). Suicide and suicidal behaviour. The Lancet, 387(10024), 1227–1239. 10.1016/s0140-6736(15)00234-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Harst P, & Verweij N (2018). Identification of 64 novel genetic loci provides an expanded view on the genetic architecture of coronary artery disease. Circulation Research, 122(3), 433–443. 10.1161/CIRCRESAHA.117.312086 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vilhjalmsson BJ, Yang J, Finucane HK, Gusev A, Lindstrom S, Ripke S, … Price AL (2015). Modeling linkage disequilibrium increases accuracy of polygenic risk scores. American Journal of Human Genetics, 97(4), 576–592. 10.1016/j.ajhg.2015.09.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voracek M, & Loibl LM (2007). Genetics of suicide: A systematic review of twin studies. Wiener Klinische Wochenschrift, 119(15–16), 463–475. 10.1007/s00508-007-0823-2 [DOI] [PubMed] [Google Scholar]
- Wyder M, & De Leo D (2007). Behind impulsive suicide attempts: Indications from a community study. Journal of Affective Disorders, 104(1–3), 167–173. 10.1016/j.jad.2007.02.015 [DOI] [PubMed] [Google Scholar]
- Zalsman G (2010). Timing is critical: Gene, environment and timing interactions in genetics of suicide in children and adolescents. European Psychiatry, 25(5), 284–286. 10.1016/j.eurpsy.2010.01.007 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data are not publicly available, but are available upon request. Persons interested in accessing the data could contact Dr. Nicholas Ialongo (nialong1@jhmi.edu).