

Abstract
Rationale & Objective.
The nasal passages harbor both commensal and pathogenic bacteria. In this study, we sought to characterize the anterior nasal microbiota in PD patients using 16S rRNA gene sequencing.
Study Design.
Cross-sectional.
Setting & Participants.
We recruited 32 PD patients, 37 kidney transplant (KTx) recipients, 22 living donor/healthy control (HC) participants and collected anterior nasal swabs at a single point in time.
Predictors.
We performed 16S rRNA gene sequencing of the V4-V5 hypervariable region to determine the nasal microbiota.
Outcomes.
Nasal microbiota profiles were determined at the genus level as well as the amplicon sequencing variant level.
Analytical Approach.
We compared nasal abundance of common genera among the 3 groups using Wilcoxon rank sum testing with Benjamini-Hochberg adjustment. DESeq2 was also utilized to compare the groups at the ASV levels.
Results.
In the entire cohort, the most abundant genera in the nasal microbiota included: Staphylococcus, Corynebacterium, Streptococcus, and Anaerococcus. Correlational analyses revealed a significant inverse relationship between the nasal abundance of Staphylococcus and that of Corynebacterium. PD patients have a higher nasal abundance of Streptococcus than KTx recipients and HC participants. PD patients have a more diverse representation of Staphylococcus and Streptococcus than KTx recipients and HC participants. PD patients who concurrently have or who developed future Staphylococcus peritonitis had a numerically higher nasal abundance of Staphylococcus than PD patients who did not develop Staphylococcus peritonitis.
Limitations.
16S RNA gene sequencing provides taxonomic information to the genus level.
Conclusions.
We find a distinct nasal microbiota signature in PD patients compared to KTx recipients and HC participants. Given the potential relationship between the nasal pathogenic bacteria and infectious complications, further studies are needed to define the nasal microbiota associated with these infectious complications and to conduct studies on the manipulation of the nasal microbiota to prevent such complications.
INTRODUCTION
The anterior nasal microbiota is at the interface between the external environment and the nasal passages and contains a combination of commensal and pathogenic bacteria. The most common genera defined in healthy individuals in the Human Microbiome Project are Staphylococcus, Corynebacterium, Propionibacterium, and Moraxella (1). Subsequent studies on the nasal microbiota have revealed microbiota dysbiosis in diseased states such as chronic rhinosinusitis (2) and have linked the nasal microbiota to infectious complications after elective surgical procedures (3).
Peritoneal dialysis (PD) patients undergo dialysis through PD catheter through their abdomen. Despite being taught sterile technique, PD patients experience both exit site infections around the catheter and infectious peritonitis. Prior work has established that pathogenic bacteria in the nasal passages may be associated with infectious complications in PD patients. Luzar et al. reported that Staphylococcus aureus nasal colonization was associated with exit site infections in a cohort of 140 PD patients (4). Other studies have found that persistent nasal colonization with S. aureus was also associated with peritonitis (5, 6). Decolonization with mupirocin has been suggested to prevent infections and the MUPIROCIN Study Group found that nasal mupirocin prevented S. aureus exit site infection (7). Despite these data, International Society of Peritoneal Dialysis (ISPD) guidelines do not support the routine use of nasal mupirocin (8).
Because no study to date has comprehensively evaluated the anterior nasal microbiota in PD patients, we performed a pilot study to evaluate the anterior nasal microbiota using 16S rRNA gene sequencing of the V4-V5 hypervariable region in PD patients, in kidney transplant recipients, and healthy controls.
METHODS
Study Cohort Recruitment and Nasal Swab Specimen Collection
From August 2021 to January 2022, we recruited patients receiving peritoneal dialysis (PD), kidney transplant (KTx) recipients, and living donor/healthy control (HC) participants for anterior nasal swab specimen collection. All kidney transplant recipients and living donor candidates were recruited from the clinic. Most PD patients were recruited in the PD clinic; several were recruited during hospitalization. The Weill Cornell Institutional Review Board approved this protocol (IRB # 1604017181) and all participants provided written informed consent.
Anterior nasal swab specimens were collected once from each participant using the Human Microbiome Project protocol. A Copan Eswab (Copan Diagnostics, Murietta, CA, USA) was inserted into the anterior part of one nostril of the participant and turned twice and was then inserted into the anterior part of the other nostril and turned twice. The Copan Eswab was then placed into 1 mL of liquid Amies provided by the Copan Eswab technology and immediately stored on ice or 4°C. Aliquots of 300 uL were created in 2 mL cryovial and stored at −80°C within 12 hours.
16S rRNA gene sequencing of the V4-V5 hypervariable region
A single aliquot of approximately 285 μL was deposited into a Qiagen PowerBead glass 0.1 mm tube. Using a Promega Maxwell RSC PureFood GMO and Authentication Kit (AS1600), 1mL of CTAB buffer & 20 μL of RNAse A Solution was added to the PowerBead tube containing the sample. The sample/buffer was mixed for 10 seconds on a Vortex Genie2 and then incubated at 95°C for 5 minutes on an Eppendorf ThermoMixer F2.0, shaking at 1500 rpm. The tube was removed and clipped to a horizontal microtube attachment on a Vortex Genie2 (SI-H524) and vortexed at high-speed for 20 minutes. The sample was removed from the Vortex and centrifuged on an Eppendorf Centrifuge 5430R at 40°C, 12700 rpm for 10 minutes. Upon completion, the sample was centrifuged again for an additional 10 minutes to eliminate foam. The tube was then added to a Promega MaxPrep Liquid Handler tube rack. The Liquid Handler instrument was loaded with proteinase K tubes, lysis buffer, elution buffer, 1000mL tips, 50mL tips, 96-sample deep-well plate, and Promega Maxwell RSC 48 plunger tips. The Promega MaxPrep Liquid Handler instrument was programed to use 300 μL of sample and transfer all sample lysate into Promega Maxwell RSC 48 extraction cartridge for DNA extraction. Upon completion, the extraction cartridge was loaded into Promega Maxwell RSC 48 for DNA extraction & elution. DNA was eluted in 100 μL and transferred to a standard 96-well plate. DNA was quantified using Quant-iT dsDNA High Sensitivity Assay Kit using Promega GloMax plate reader on a microplate (655087). 16S rRNA library generation followed the protocol from the Earth Microbiome Project.
Amplicon libraries were washed using Beckman Coulter AMPure XP magnetic beads. Library quality & size verification was performed using PerkinElmer LabChip GXII instrument with DNA 1K Reagent Kit (CLS760673). Library concentrations were quantified using Quant-iT dsDNA High Sensitivity Assay Kit using Promega GloMax plate reader on a microplate (655087). Library molarity was calculated based on library peak size & concentration. Libraries were normalized to 2nM using the PerkinElmer Zephyr G3 NGS Workstation (133750) and pooled together using the same volume across all normalized libraries into a 1.5mL Eppendorf DNA tube (022431021). Sequencing was performed on an Illumina MiSeq instrument at loading concentration of 7 pM with 15% PhiX, paired-end 250 using MiSeq Reagent Kit v2, 500-cycles (MS-102-2003).
Bioinformatics Pipeline
Demultiplexed raw reads were processed using the Nextflow (9) nf-core (10) ampliseq pipeline (11), version 2.2.0, with the following parameters: -profile singularity --input SampleSheet.tsv -- FW_primer GTGYCAGCMGCCGCGGTAA --RV_primer CCGYCAATTYMTTTRAGTTT -- metadata Metadata.tsv --outdir results --dada_ref_taxonomy silva --ignore_empty_input_files -- ignore_failed_trimming --min_frequency 10 --retain_untrimmed --trunclenf 240 --trunclenr 160. Specifically, reads were trimmed with cutadapt (12), PhiX and quality filtering, read pair merging, and amplicon sequence variant resolution was performed with DADA2 (13). Subsequent taxonomic assignment was also performed with DADA2, using the Silva reference database (14), version 138. Sequences that were assigned the families, Chloroplast and Mitochondria, were removed from downstream analyses.
Biostatistical Analyses
The distribution of categorial variables were compared using Fisher’s exact tests. The distribution of continuous variables were compared using Wilcoxon rank sum tests and to account for the comparison of multiple taxa, adjusted p values were calculated using Benjamini-Hochberg adjustment for multiple comparisons. DESeq2 was utilized to detect differences at the ASV between the groups using Benjamini-Hochberg adjustment. Comparison of correlations using a correlational matrix was adjusted for multiple comparisons using the Bonferonni method. All statistical tests were performed using R 4.1.3 in RStudio.
Data Availability
Sequencing data that support the findings of this study will be made available in the database of Genotypes and Phenotypes (dbGaP) phs002251.v1.p1 after peer-reviewed acceptance. Local institutional review board approval will be needed to access the data.
RESULTS
Characteristics of the Study Cohort and Nasal Microbial Sequencing
The microbiota in the anterior nares was performing using 16S rRNA gene sequencing of the V4-V5 hypervariable region in 32 PD patients, 37 KTx recipients, 22 HC participants, and 3 negative controls. A total of 1,116,291 reads with assigned taxonomy was obtained in the cohort of 91 participants with a median of 12,713 assigned reads with an interquartile range of 7,132 and 16,018 assigned reads. The number of assigned reads in the 3 negative controls were 146, 308, and 529, below the number in the cohort of participants.
Table 1 shows the demographics of the participants. In general, the PD patients were older than the HC participants and similar in age to the KTx recipients. More than 50% of PD patients performed automated peritoneal dialysis and 15% had current Staphylococcus peritonitis or developed future Staphylococcus peritonitis within 10 to 12 months from the nasal specimen collection (last follow up). Approximately a third of KTx recipients received deceased donor transplantation and 32% were on trimethoprim/sulfamethoxazole (TMP-SMX) prophylaxis.
TABLE 1. Demographics of the Cohort.
Categorical variables are represented by the number followed by the percentage in parentheses. Continuous variables are represented by the median followed by the interquartile range in parentheses.
| Characteristic | PD Cohort (n=32) median abundance |
KTx Cohort (n = 37) median abundance |
HC Cohort (n = 22) median abundance |
|---|---|---|---|
| Age, years | 63 (51, 73) | 59 (52, 67) | 51 (33, 61) |
| Female Sex | 20 (63%) | 17 (46%) | 13 (59%) |
| Ethnicity | |||
| Hispanic | 3 (9%) | 2 (5%) | 3 (14%) |
| Non-Hispanic | 27 (84%) | 33 (89%) | 16 (73%) |
| Declined | 2 (6%) | 2 (5%) | 3 (14%) |
| Race | |||
| Asian | 4 (11%) | 4 (11%) | 1 (5%) |
| Black | 13 (41%) | 10 (27%) | 5 (23%) |
| White | 11 (34%) | 20 (54%) | 11 (50%) |
| Other | 3 (9%) | 2 (5%) | 1 (5%) |
| Declined | 1 (3%) | 1 (3%) | 4 (18%) |
| History of Hypertension | 26 (81%) | 36 (97%) | 2 (9%) |
| History of Diabetes Mellitus | 9 (28%) | 12 (32%) | 0 (0%) |
| Years on PD | 1.2 (0.6, 2.4) | ||
| Automated Peritoneal Dialysis | 21 (66%) | ||
| Concurrent or Develops Future | |||
| Staphylococcus Peritonitis | 6 (19%) | ||
| Decreased Donor Transplantation | 14 (38%) | ||
| Days Post Transplantation | 364 (51, 1637) | ||
| History of Prior Transplantation | 4 (11%) | ||
| Maintenance Immunosuppression | |||
| Tacro/mycophenolic mofetil | 19 (51%) | ||
| Tacro/mycophenolic mofetil/pred | 14 (38%) | ||
| Tacro/mycophenolic acid | 1 (3%) | ||
| Tacro/mycophenolic acid/pred | 3 (8%) | ||
| Trimethoprim/Sulfamethoxazole PPx | 12 (32%) |
Anterior Nasal Microbial Diversity Differs Across the Study Cohort
Microbial diversity among the study participants was measured at the ASV level using the Shannon diversity, an index that evaluates the richness and evenness in a community, as well as Chao1, an index that estimates the total number of ASVs in the specimens. Fig. 1A and B show box and whisker plots of these diversity indices and reveal that PD patients had a significantly higher Shannon diversity index and Chao1 diversity index than KTx recipients (P<0.05, Wilcoxon rank sum test) but similar to the HC participants (P>0.05).
FIGURE 1. Distinct Differences in Nasal Microbial Diversity among the Study Cohort.

Panel A shows box and whisker plots of Chao1 index, the estimated number of amplicon sequence variants, in the anterior nasal specimens from the peritoneal dialysis (PD) cohort, the kidney transplant cohort (Ktx), and the living donor /healthy control (HC) cohort. The Chao1 index is on the y axis and the study group is on the x-axis. P value shown was calculated by Wilcoxon rank sum test. Panel B shows box and whisker plots of Shannon diversity index, a measure of evenness and richness, in the anterior nasal specimens from the 3 cohorts. The Shannon diversity index is on the y axis and the study group is on the x-axis. P values shown were calculated by Wilcoxon rank sum test.
Staphylococcus Abundance Negatively Correlates with Corynebacterium Abundance in Anterior Nasal Specimens
We further evaluated the anterior nasal microbiota among the study cohort at the genus level. At the genus level, the top abundant genera (>1% mean abundance across the cohort) included Staphylococcus, Corynebacterium, Anaerococcus, Streptococcus, unspecified Neisseriaceae, Moraxella, Cutibacterium, Peptoniphilus, and Finegoldia (Fig. 2A). We performed a correlational matrix analysis among each of the genera (Fig. 2B). The relative abundance of Staphylococcus was inversely correlated with that of Corynebacterium (Pearson r = −0.66, adjusted P value < 0.10, Benjamini-Hochberg adjustment). The relative abundances of Peptoniphilus was positively associated with that of Anaerococcus (r=0.52, adjusted P value < 0.10) and of Finegoldia (r=0.27, adjusted P value < 0.10). The relative abundance of Finegoldia was positively associated with that of Anaerococcus (r= 0.60, adjusted P value < 0.10). The relative abundance of Unspecified Neisseriaceae was positively associated with that of Cutibacterium (r= 0.31, adjusted P value < 0.10).
FIGURE 2. Significant Correlations among the Most Common Genera in the Study Cohort.
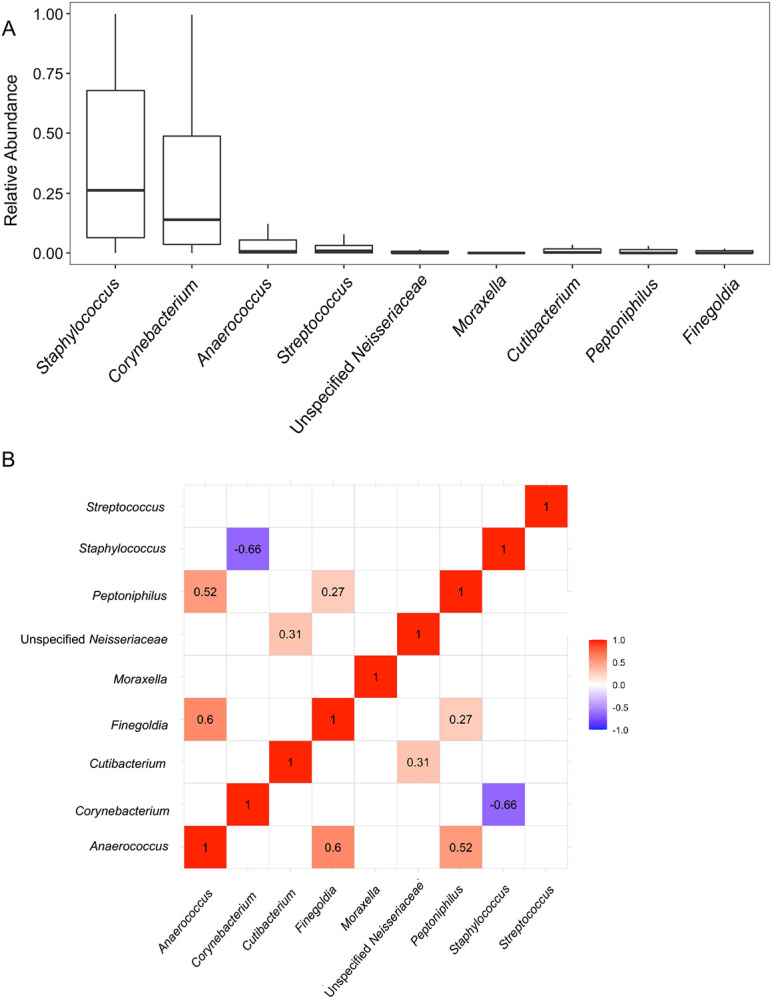
Panel A shows the most common genera in the anterior nasal microbiota (>1% mean relative abundance in the cohort). Box and whisker plots are represented to show the variation in the relative abundance of the genus (y-axis) with the genus on the x-axis. Panel B shows a correlational matrix between the nasal abundance of the most common genera using Pearson’s r correlations with Benjamini-Hochberg adjustment for multiple hypotheses. The numbers shown are Pearson’s r correlations that had an adjusted P value < 0.10. The color shows the strength of the correlation with red showing a positive correlation between two genera and blue showing a negative correlation between two genera.
Distinct Anterior Nasal Microbiota Define PD Patients and Kidney Transplant Recipients
The individual profiles of the top genera in anterior nasal microbiota are shown in the PD patients, KTx recipients, and HC participants (Fig. 3). Fig. 4 shows box and whisker plots of the top 9 taxa among the PD patients, the KTx recipients, and the HC participants and Table 2 shows the comparisons among the groups using Wilcoxon rank sum testing with Benjamini-Hochberg adjustment. PD patients had a distinctly higher relative abundance of Streptococcus than KTx recipients or HC participants (Adjusted P value < 0.10, Wilcoxon rank sum test, Benjamini-Hochberg adjustment) (Fig. 4D). PD patients also had lower abundance of unspecified Neisseriaceae, Moraxella, Cutibacterium, and Peptoniphilus than HC participants (Fig. 4G) (Adjusted P value < 0.10). Kidney transplant recipients had a lower abundance of Moraxella than HC participants (Adjusted P value < 0.10). Other than Streptococcus, KTx recipients had similar abundance of the top genera compared to PD patients (Adjusted P value > 0.10).
FIGURE 3. Individual Microbiota Profiles by the Study Cohort.

The relative abundance of microbiota is on the y axis and individual nasal specimens are on the x-axis. The relative abundance of each genus is represented by color. The top panel represents anterior nasal microbiota profiles from the 32 peritoneal dialysis (PD) patients, the middle panel represents the anterior nasal microbiota profiles from the 37 kidney transplant (KTx) patients, and the bottom panel represents the anterior nasal microbiota profiles from the 22 potential living donor / healthy control (HC) participants. The right panel represents the microbiota from 3 negative controls.
FIGURE 4. Distinct microbial differences among the cohort at the genus level.

Box and whisker plots are represented with the relative abundance of individual genera on the y axis and the group on the x axis. PD, peritoneal dialysis cohort (n=32). KTx, kidney transplant cohort (n=37). HC, living donor / healthy control cohort (n=22). P values were calculated using Wilcoxon rank sum testing with Benjamini-Hochberg adjustment for multiple hypothesis. ** Adjusted P value < 0.05 * Adjusted P value < 0.10. Panel A, Staphylococcus analysis. Panel B, Corynebacterium analysis. Panel C, Anaerococcus analysis. Panel D, Streptococcus analysis. Panel E, Unspecified Neisseriaceae analysis. Panel F, Moraxella analysis. Panel G, Cutibacterium analysis. Panel H, Peptoniphilus analysis. Panel I. Finegoldia analysis.
TABLE 2. Comparison of the Nasal Abundance Among the 3 Cohorts at the Genus Level.
The median abundance of the most common genera are shown for the peritoneal dialysis (PD) cohort, the kidney transplant cohort (KTx), and the living donor /healthy control (HC) cohort. P values shown were calculated using Wilcoxon rank sum test between groups. Adjusted P value (Adj P Value) were calculated using Benjamini-Hochberg adjustment. P values with NA were unable to be calculated because the abundances were 0 in both groups.
| Genus | PD Cohort (n=32) median abundance |
KTx Cohort (n = 37) median abundance |
HC Cohort (n = 22) median abundance |
PD vs. HC P value |
PD vs. HC Adj P Value |
PD vs. KTx P value |
PD vs. KTx Adj P Value |
KTx vs. HC P value |
KTx vs. HC Adj P Value |
|---|---|---|---|---|---|---|---|---|---|
| Anaerococcus | 0.020 | 0.004 | 0.014 | 0.999 | 0.999 | 0.238 | 0.722 | 0.303 | 0.389 |
| Corynebacterium | 0.098 | 0.200 | 0.127 | 0.509 | 0.573 | 0.271 | 0.722 | 0.701 | 0.701 |
| Cutibacterium | 0.000 | 0.000 | 0.009 | 0.016 | 0.049 | 0.942 | 0.942 | 0.029 | 0.133 |
| Finegoldia | 0.000 | 0.000 | 0.001 | 0.098 | 0.147 | 0.660 | 0.880 | 0.178 | 0.320 |
| Moraxella | 0.000 | 0.000 | 0.000 | 0.014 | 0.049 | NA | NA | 0.008 | 0.074 |
| Unspecified | |||||||||
| Neisseriaceae | 0.000 | 0.000 | 0.003 | 0.034 | 0.062 | 0.510 | 0.817 | 0.105 | 0.237 |
| Peptoniphilus | 0.000 | 0.000 | 0.013 | 0.022 | 0.049 | 0.382 | 0.763 | 0.046 | 0.139 |
| Staphylococcus | 0.249 | 0.403 | 0.257 | 0.406 | 0.522 | 0.891 | 0.942 | 0.285 | 0.389 |
| Streptococcus | 0.027 | 0.005 | 0.003 | 0.001 | 0.008 | 0.003 | 0.027 | 0.584 | 0.657 |
In order to gain further insight, we evaluated the taxa at the ASV level. We performed pairwise DESeq2 between the groups to identify ASVs that were consistently different among the groups. Fig. 5 shows the significant log2 fold abundance changes between the groups and SI Tables 1 to 3 reveal the changes in the nasal abundances of the groups. Both PD patients and KTx recipients had significantly higher nasal abundances of Staphylococcus ASV #1 and Corynebacterium ASV #1 and lower abundance of Anaerococcus ASV #1 than HC participants (Adjusted p value < 0.10, Benjamini-Hochberg adjustment). PD patients had higher nasal abundance of Staphylococcus ASV #2, Abiotrophia ASV #1, and Porphyromonas ASV #1 than KTx recipients or HC participants (Adjusted p value < 0.10).
FIGURE 5. Differential abundance analyses among the cohort at the amplicon sequence variant level.

Differential abundance analyses were performed on the anterior nasal microbiota between the groups using DESeq2 with Benjamini-Hochberg adjustment for multiple hypothesis testing. On the y axis is the individual amplicon sequence variant with genus shown and on the x axis is the fold difference in abundance. PD, peritoneal dialysis cohort. KTx, kidney transplant cohort. HC, living donor / healthy control cohort. The fold difference directionality is represented above the graph. Panel A represents differential abundance analyses between the HC Group and the PD Group. Panel B represents differential abundance analyses between the KTx Group and the PD Group. Panel C represents differential abundance analyses between the HC Group and the KTx Group.
PD Patients Have a More Diverse Representation of Staphylococcus and Streptococcus than KTx Recipients and HC Participants
To further understand why PD patients have higher microbial diversity, we evaluated the diversity of ASVs in the most common genera: Staphylococcus, Corynebacterium, and Streptococcus. There were 61 different Staphylococcus ASVs identified in the whole cohort. PD patients had a significantly higher number of Staphylococcus ASVs per specimen than KTx patients (P=0.03, Wilcoxon rank sum test) and HC participants (P=0.04). There were 95 different Corynebacterium ASVs identified in the whole cohort. PD patients, KTx patients, and HC participants had similar number of Corynebacterium ASVs per specimen (P>0.10). There were 46 different Streptococcus ASVs identified in the whole cohort. PD patients had a significantly higher number of Streptococcus ASVs per specimen than KTx patients (P=0.04) and HC participants (P=0.05).
Clinical Factors, Outcomes, and the Nasal Microbiota
We next evaluated the relationship among the nasal microbiota, clinical factors, and outcomes in the cohort. There were no significant differences in the nasal abundances of the most common genera based upon age greater than or equal to 65 years old (SI Table 4). The relative abundance of Peptoniphilus was significantly higher in male patients than in female patients (adjusted P value < 0.10) (SI Table 5). Twelve of the kidney transplant recipients were on TMP-SMX prophylaxis for Pneumocystic jirovecii prophylaxis and 35 were not. There were no significant differences in the nasal abundance of the most common genera between the kidney transplant recipients on TMP-SMX and those who were not (SI Table 6). In the PD cohort, 6 PD patients concurrently had Staphylococcus peritonitis or developed future Staphylococcus peritonitis within 10 to 12 months (last follow up) (Staph Peritonitis Group) and 26 PD patients did not (No Staph Peritonitis Group). The nasal abundance of Staphylococcus was higher in the Staph Peritonitis Group than in the No Staph Peritonitis Group but the difference was not statistically significant (median abundance 52% vs. 24%, respectively, adjusted P value 0.73). There were no significant differences in the nasal abundance of the other most common genera between the Staph Peritonitis Group and the No Staph Peritonitis Group (SI Table 7).
DISCUSSION
This study aimed to describe the anterior nasal microbiota across different groups of patients with kidney disease. We detect a distinct microbial signature in the anterior nares of PD patients compared to KTx recipients and HC participants.
Many of the most common genera in the kidney cohort overlap with those reported in healthy individuals and include Staphylococccus, Corynebacterium, Finegoldia, and Cutibacterium (15, 16). However, there were some distinct differences among the groups. PD patients had a higher nasal abundance of Streptococcus than HC participants or KTx recipients. Interestingly, having a higher nasal abundance of Streptococcus has been associated with respiratory infections such as bronchiolitis in infants (17). While the most common type of infectious peritonitis is Staphylococcus in origin, Streptococcus peritonitis also occurs in PD patients. Our study was not able to directly address whether PD patients with nasal abundance of Streptococcus is associated with Streptococcus peritonitis and/or respiratory viral infections, but such a link would provide the groundwork for novel approaches to manipulate the nasal microbiota to prevent such complications.
In our analysis, we noticed a higher nasal microbial diversity in the PD patients compared to KTx recipients and HC participants. Further analysis showed that part of this increased microbial diversity may be due to a more diverse representation of Staphylococcus and Streptococcus in PD patients (Fig. 6). This may have interesting implications as prior data suggests that PD patients with Staphylococcus aureus colonization had a higher incidence of exit site infections (Ref). In our study, we did find an increased nasal abundance of Staphylococcus in PD patients who had a history of Staphylococcus peritonitis and/or developed Staphylococcus peritonitis. While the association was not significant, it could be due to the low number of PD patients in the PD Staph peritonitis as our study was not powered to detect the differences. At the 16S rRNA level, we were not able to determine Staphylococcus at the species level and this limitation prevented us to assess this association in more detail.
FIGURE 6. Diverse representation of the most common genera in the study cohort.

Each set of graphs represents the number of amplicon sequence variant (ASV) from a particular genus by group. PD, peritoneal dialysis group (n=32). KTx, kidney transplant group (n=37). HC, living donor / healthy control group (n=22). The top graph presents box and whisker plots of the number of ASVs per specimen by group. P value was calculated using Wilcoxon rank sum test: ** P value < 0.05 * P value < 0.10. The bottom 3 graphs represent box and whisker plots of the relative abundance of individual ASVs from a particular genus. The relative abundance is on the y axis with the color representing individual ASVs and anterior nasal specimens are on the x axis with the second top graph representing the PD Cohort, the second bottom graph representing the KTx Cohort, and the bottom graph representing the HC Cohort. Panel A shows the diversity of Staphylococcus ASVs in the study cohort. Panel B shows the diversity of Corynebacterium ASVs in the study cohort. Panel C shows the diversity of Streptococcus ASVs in the study cohort.
Our data also highlight a strong inverse association between the nasal abundance of Staphylococcus and that of Corynebacterium. Interesting mechanistic studies have shown a complicated relationship between these two taxa, which represent the most common taxa in the nasal microbiota. One study found that Corynebacterium species can secrete antimicrobial peptides against Staphylococcus aureus (18). Another study has shown that Corynebacterium species can decrease the virulence of S. aureus (19). Taken together, our data are consistent with the inverse relationship and suggest potential novel approaches to manipulate the nasal microbiota. For example, since Staphylococcus peritonitis is much more common than Corynebacterium peritonitis, establishing a Corynebacterium dominant nasal microbiota may be preventative of Staphylococcus in the nasal passages and possibly decrease the risk for Staphylococcus exit site infection and/or peritonitis.
A surprising result is that we did not find an association between the TMP/SMX and nasal microbiota differences. TMP/SMX has broad coverage against gram positive cocci including Staphylococcus species. There are few studies which have investigated the role of oral antibiotics on the nasal microbiota and it is possible that intra-nasal antibiotics rather than oral antibiotics may more efficiently impact the nasal microbiota. While our study is limited by the population size and the cross-sectional nature, our study raises this possibility.
There are several limitations to our study. As mentioned prior, we are unable to assess species level identification via 16S rRNA gene sequencing of the V4-V5 hypervariable region. Future studies using whole gene 16S rRNA gene sequencing or metagenomic sequencing may provide better resolution on the intricate intra-species competition between the microbiota, particularly between Staphylococcus species. and Corynebacterium species. Given the low biomass of the nasal microbiota, environmental contamination and/or contamination through the DNA processing steps could artificially introduce microbiota in our specimens. However, we did sequence negative controls (Fig. 3) and the most abundant microbiota identified were not the most common nasal microbiota flora previously reported, suggesting that the nasal microbiota identified in our cohort was present in higher quantities and distinct. The cross-sectional nature of our study provides a snapshot of the microbiota across different groups of patients with kidney disease but does not provide longitudinal changes. Such a longitudinal study may provide more insight into the relationship between the microbiota and clinical factors and outcomes in the populations.
In conclusion, we provide the first description of a distinct nasal microbiota signature in PD patients compared to KTx recipients and HC participants. We find a higher abundance of Streptococcus and a more diverse representation of Staphylococcus and Streptococcus in PD patients. Given the potential relationship between the nasal bacteria and infectious complications in PD patients, further studies are needed to define the nasal microbiota associated with these infectious complications and to conduct studies on the manipulation of the nasal microbiota to prevent such complications.
Supplementary Material
Acknowledgments.
We thank Dr. Manikkam Suthanthiran for his overall guidance in this project.
Funding.
This study was supported in part by R21AI164093 (JRL, IDW).
Footnotes
Financial Disclosure.
DMD, JRL, and IDV hold patent US-2020-0048713-A1 titled “Methods of Detecting Cell-Free DNA in Biological Samples” licensed to Eurofins Viracor. LFW has received or receives research support from Accelerate Diagnostics, Inc., bioMérieux, Inc., Hardy Diagnostics, and Roche Molecular Systems, Inc., and consulting fees from Roche Molecular Systems, Inc., Shionogi, Inc., and Talis Biomedical. MJS reports research support from Merck, Biomérieux, and SNIPRBiome, compensation for consulting from Shionogi, and compensation for participation in a Data and Safety Monitoring Board from AbbVie. CK is a consultant for Rebiotix/Ferring and on the Scientific Advisory Board of Seres Therapeutics. DMD serves as a consultant for CareDx and is a site clinical investigator for studies sponsored by AlloVir and CSL Behring. JS is the Co-Chair of the American Society of Nephrology Workgroup on COVID-19 and other Emerging Threats and of the American Society of Nephrology Emergency Partnership Initiative and has received consulting fees from Alkahest, Bayer, Honeywell, Kaneka and St. Gobain. IDV is a member of the Scientific Advisory Board of Karius Inc., Kanvas Biosciences and GenDX. VS reports receiving speakers’ fees from Baxter Healthcare. JRL received research support under an investigator-initiated research grant from BioFire Diagnostics, LLC. JRL receives speakers’ fees from Astellas.
REFERENCES
- 1.Human Microbiome Project C. Structure, function and diversity of the healthy human microbiome. Nature. 2012;486(7402):207–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chen F, Gao W, Yu C, Li J, Yu F, Xia M, et al. Age-Associated Changes of Nasal Bacterial Microbiome in Patients With Chronic Rhinosinusitis. Front Cell Infect Microbiol. 2022;12:786481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hsiao CJ, Paulson JN, Singh S, Mongodin EF, Carroll KC, Fraser CM, et al. Nasal Microbiota and Infectious Complications After Elective Surgical Procedures. JAMA Netw Open. 2021;4(4):e218386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Luzar MA, Coles GA, Faller B, Slingeneyer A, Dah GD, Briat C, et al. Staphylococcus aureus nasal carriage and infection in patients on continuous ambulatory peritoneal dialysis. N Engl J Med. 1990;322(8):505–9. [DOI] [PubMed] [Google Scholar]
- 5.Nouwen JL, Fieren MW, Snijders S, Verbrugh HA, and van Belkum A. Persistent (not intermittent) nasal carriage of Staphylococcus aureus is the determinant of CPD-related infections. Kidney Int. 2005;67(3):1084–92. [DOI] [PubMed] [Google Scholar]
- 6.Ong LM, Ch'ng CC, Wee HC, Supramaniam P, Zainal H, Goh BL, et al. Risk of Peritoneal Dialysis-Related Peritonitis in a Multi-Racial Asian Population. Perit Dial Int. 2017;37(1):35–43. [DOI] [PubMed] [Google Scholar]
- 7.Nasal mupirocin prevents Staphylococcus aureus exit-site infection during peritoneal dialysis. Mupirocin Study Group. J Am Soc Nephrol. 1996;7(11):2403–8. [DOI] [PubMed] [Google Scholar]
- 8.Li PK, Szeto CC, Piraino B, de Arteaga J, Fan S, Figueiredo AE, et al. ISPD Peritonitis Recommendations: 2016 Update on Prevention and Treatment. Perit Dial Int. 2016;36(5):481–508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Di Tommaso P, Chatzou M, Floden EW, Barja PP, Palumbo E, and Notredame C. Nextflow enables reproducible computational workflows. Nat Biotechnol. 2017;35(4):316–9. [DOI] [PubMed] [Google Scholar]
- 10.Ewels PA, Peltzer A, Fillinger S, Patel H, Alneberg J, Wilm A, et al. The nf-core framework for community-curated bioinformatics pipelines. Nat Biotechnol. 2020;38(3):276–8. [DOI] [PubMed] [Google Scholar]
- 11.Straub D, Blackwell N, Langarica-Fuentes A, Peltzer A, Nahnsen S, and Kleindienst S. Interpretations of Environmental Microbial Community Studies Are Biased by the Selected 16S rRNA (Gene) Amplicon Sequencing Pipeline. Front Microbiol. 2020;11:550420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Martin M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnetjournal. 2011;17: 10.14806/ej.17.1.200. [DOI] [Google Scholar]
- 13.Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJ, and Holmes SP. DADA2: High-resolution sample inference from Illumina amplicon data. Nat Methods. 2016;13(7):581–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 2013;41(Database issue):D590–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Oh J, Conlan S, Polley EC, Segre JA, and Kong HH. Shifts in human skin and nares microbiota of healthy children and adults. Genome Med. 2012;4(10):77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Pereira PAB, Aho VTE, Paulin L, Pekkonen E, Auvinen P, and Scheperjans F. Oral and nasal microbiota in Parkinson's disease. Parkinsonism Relat Disord. 2017;38:61–7. [DOI] [PubMed] [Google Scholar]
- 17.Hasegawa K, Linnemann RW, Mansbach JM, Ajami NJ, Espinola JA, Petrosino JF, et al. Nasal Airway Microbiota Profile and Severe Bronchiolitis in Infants: A Case-control Study. Pediatr Infect Dis J. 2017;36(11):1044–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Menberu MA, Liu S, Cooksley C, Hayes AJ, Psaltis AJ, Wormald PJ, et al. Corynebacterium accolens Has Antimicrobial Activity against Staphylococcus aureus and Methicillin-Resistant S. aureus Pathogens Isolated from the Sinonasal Niche of Chronic Rhinosinusitis Patients. Pathogens. 2021;10(2). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ramsey MM, Freire MO, Gabrilska RA, Rumbaugh KP, and Lemon KP. Staphylococcus aureus Shifts toward Commensalism in Response to Corynebacterium Species. Front Microbiol. 2016;7:1230. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Sequencing data that support the findings of this study will be made available in the database of Genotypes and Phenotypes (dbGaP) phs002251.v1.p1 after peer-reviewed acceptance. Local institutional review board approval will be needed to access the data.