

Abstract
This study investigates the effects of soybean meal fermented by Enterococcus faecium as a replacement for soybean meal on growth performance, apparent total tract digestibility, blood profile and gut microbiota of weaned pigs. Eighty piglets (weaned at 21 days) [(Landrace × Yorkshire) × Duroc] with average body weight of 6.52 ± 0.59 kg) were selected and assigned to 4 treatments/4 replicate pens (3 barrows and 2 gilts). The four diets (SBM, 3, 6 and 9% FSBM) were formulated using fermented soybean meal to replace 0, 3, 6 and 9% of soybean meal, respectively. The trial lasted for 42 days phase 1, 2 and 3. Result showed that supplemental FSBM increased (P < 0.05) the body weight gain (BWG) of piglets at day 7, 21 and 42 and ADG at days 1–7, 8–21, 22–42 and 1–42, and ADFI at days 8–21, 22–42 and 1–42 and G: F at days 1–7, 8–21 and 1–42, and crude protein, dry matter, and gross energy digestibility at day 42, and lowered (P < 0.05) diarrhea at days 1–21 and 22–42. The concentration of glucose levels, WBC, RBC, and lymphocytes were increased while, concentration of BUN level in the serum was lowered in the FSBM treatment compared to the SBM group (P < 0.05). Microbiota sequencing found that FSBM supplementation increased the microbial Shannon, Simpsons and Chao indexs, (P < 0.05) and the abundances of the phylum Firmicutes, and genera prevotella, Lactobacillus, Lachnospiraceae and Lachnoclostridium (P < 0.05), lower in the abundances of the phylum bacteroidetes, Proteobacteria, genera Escherichia-Shigella, Clostridium sensu stricto1, Bacteroides and Parabacteroides (P < 0.05). Overall, FSBM replacing SBM improved the growth performance, apparent total tract digestibility, and blood profiles; perhaps via altering the faecal microbiota and its metabolites in weaned pigs. The present study provides theoretical support for applying FSBM at 6–9% to promote immune characteristics and regulate intestinal health in weaning piglets.
Subject terms: Microbiology, Gastroenterology
Introduction
In the swine husbandry, weaning produce gut system dysfunctions and results from inconstant impairment of the gut barrier function, oxidative stress and absorption as a result of poor growth, diarrhoea and other diseases1,2. Weaning pigs are immediately required to undergo a change from high digestible sow’s milk to solid diets as well as complex protein3,4. Therefore, it is important to explore potential protein sources that should be added to diets to alleviate the weaning stress of piglets. The high cost, finite supply, and unstable variation of sources animal protein had become major reasons for limiting its supplement in diets on weaning pigs. Soybean meal (SBM) is an important protein feed ingredient in livestock diet, but variety of antinutritional factors (ANF) as well as β-conglycinin, glycinin and trypsin inhibitor, which would lead to immune responses, digestive disorders and negative effects on animal health3,5.
Probiotics are alternatives to in-feed antibiotics because they are successful in improving livestock production, efficiency and welfare6. Enterococcus faecium is a gram-positive gamma-hemolytic or non-hemolytic bacterium in the genus Enterococcus is a kind of facultative anaerobic lactobacillus that can colonize the gastrointestinal tract of humans and animals7. Enterococcus faecium bacterium feeds have shown positive effects in piglets intestinal microbiota, increasing immunoreactivity while reducing diarrhea8. The administration of living microbial preparations is one part of probiotic treatments. While, the another effective pathway is microbial fermentation of feed. Cheng et al.9 and Li et al.10 reported that bacterial fermentation could reduce content by ANFs and improving on nutritional quality and nutrient bioavailability. Fermented soybean meal (FSBM), a manufactured product mixed with solid SBM, liquid phases and the vaccinating the mixture with E. faecium11 could improve protein quality and reduce on ANFs levels of SBM with solid state fermentation12. It has been reported that FSBM has partial or total replacement of SBM to improve the growth performance and apparent tract total digestibility (ATTD) of crude protein and gross energy immune and antioxidant capacity in weaning pigs2. Jeong et al.13 found that compared with SBM, FSBM had improve the growth performance and the apparent ileal digestibility of crude protein, dry matter, and gross energy and amino acids in weaning piglets. Moreover, compared with SBM, FSBM showed greater concentration of crude protein and amino acid and reduce trypsin inhibitor and ANFs14.
The ideal situation would be to select feed probiotics that would improve soybean meal quality and enhance resistance to weaning stress by boosting the gut microbiota. Using Enterococcus faecium (E. faecium) toproduce a fermented soybean meal (FSBM) in weaning pigs has not been stated earlier and a probiotic-fermented SBM might show some different effects on piglets. The objective of the current study was to compare the effects of dietary SBM and FSBM on growth performance, ATTD, blood profile and gut microbiota of weaning pigs.
Materials and methods
Animal care
The animal care and experimental procedures described in this experiment were conducted according to the Animal Welfare Committee guidelines and had the approval of the Ethics committee of Animal Resource and Science College of Dankook University (DK-2-1936, Cheonan, South Korea). And the experiments were performed in accordance with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines (https://arriveguidelines.org).
Preparation of fermented soybean meal (FSBM)
The Enterococcus faecium SLB130 were grown in de Man, Rogosa and Sharpe (MRS) medium at 37 °C. Fermentation was initiated by soaking SBM in distilled water to a achieve 30% moisture content. Water-soaked SBM and inoculated with a 10% of Enterococcus faecium SLB130 to achieve 106 cfu/g in SBM. SBM mixtures were anaerobically solid-state fermented at 37 °C for 48 h using a previously published protocol15. Finally, the fermented SBM was dried at 50–60 °C to a moisture concentration of 10% and then ground in a hammer mill. Final fermented SBM consisted of 5.8 × 107/g. Crude protein, KOH protein solubility of FSBM was determined by official methods of analysis16. Glycinin, β-conglycinin and trypsin inhibitor in FSBM were tested using a commercial kit (Feed Up Co., Ltd., Republic of Korea) (Table 1).
Table 1.
Compositions of soybean meal (SBM) and fermented soybean meal (FSBM).
| Item | SBM | FSBM |
|---|---|---|
| Crude protein (%)* | 33.13 ± 0.43b | 39.78 ± 0.26a |
| Metabolizable energy**, kcal/kg | 3732 | 3720 |
| Crude fiber, % | 4.00 ± 96 | 4.08 ± 56 |
| Crude Ash, % | 5.56 ± 0.29 | 5.84 ± 0.084 |
| KOH protein solubility (%) | 86.69 ± 0.51a | 75.45 ± 1.83b |
| TCA soluble protein (%) | 2.21 ± 0.06b | 11.06 ± 0.08a |
| Glycinin (mg/g)* | 140.22 ± 0.08a | 28.88 ± 1.33b |
| β-conglycinin (mg/g)* | 113.42 ± 1.49a | 36.13 ± 0.29b |
| Trypsin inhibitor (mg/g)* | 11.16 ± 0.4a | 0.33 ± 0.02b |
| Stachyose (%) | 4.57 ± 0.057a | 0.18 ± 0.03b |
| Raffinose (%) | 2.81 ± 0.16a | 0.54 ± 0.03b |
*On a dry matter basis; a,b, Means within rows with different letters differed significantly (P < 0.05). The comparison was conducted in a horizontal manner.
**Metabolizable energy of SBM, FSBM is measured.
Experimental design, animals and feeding method
Eighty [(Landrace × Yorkshire) × Duroc] crossed weaned piglets (21 of age; 6.52 ± 0.59 kg) were randomly selected and allocated into 4 diets with 4 pens replicates according to the average initial body weight and sex, and each pen has 5 piglets (3 barrows and 2 gilts). Four diets were formulated using FSBM to replace 0, 3, 6 and 9% of SBM, respectively. The trial lasted for 42 days (phase 1 (days 1–7), phase 2 (8–21) and phase 3 (days 22–42). Basal diets were formulated to meet the NRC (2012) requirements (Tables 2, 3 and 4). Throughout the trial, all pigs had free access to feed and water, and the room temperature was maintained at 24–26 °C with 60–70% humidity, respectively.
Table 2.
Composition of weaning pig diets (as fed-basis).
| Item | Dietary treatmentsa Phase 1 (1–7) | |||
|---|---|---|---|---|
| FSBM0 | FSBM3 | FSBM6 | FSBM9 | |
| Ingredients (%) | ||||
| Corn | 39.29 | 39.89 | 39.67 | 39.45 |
| Soybean meal | 22.18 | 20.06 | 17.64 | 15.22 |
| FSBM | – | 3.00 | 6.00 | 9.00 |
| Spray-dried plasma protein | 6.00 | 6.00 | 6.00 | 6.00 |
| Tallow | 0.78 | 1.36 | 1.02 | 0.69 |
| Coconut powder (60% fat) | 2 | – | – | – |
| Lactose | 13.46 | 13.46 | 13.46 | 13.46 |
| Sugar | 2.00 | 2.00 | 2.00 | 2.00 |
| Whey protein | 11.00 | 11.00 | 11.00 | 11.00 |
| Monodicalcium phosphate | 1.40 | 1.38 | 1.38 | 1.38 |
| Limestone | 0.94 | 0.94 | 0.94 | 0.94 |
| Salt | 0.10 | 0.10 | 0.10 | 0.10 |
| Methionine (99%) | 0.19 | 0.18 | 0.18 | 0.17 |
| Lysine | 0.23 | 0.20 | 0.18 | 0.16 |
| Mineral mixtureb | 0.20 | 0.20 | 0.20 | 0.20 |
| Vitamin mixturec | 0.20 | 0.20 | 0.20 | 0.20 |
| Choline (25%) | 0.03 | 0.03 | 0.03 | 0.03 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 |
| Calculated chemical composition | ||||
| Crude protein, % | 20.00 | 20.00 | 20.00 | 20.00 |
| Metabolizable energy, kcal/kg | 3400 | 3400 | 3400 | 3400 |
| Calcium, % | 0.80 | 0.80 | 0.80 | 0.80 |
| Available phosphorus, % | 0.70 | 0.70 | 0.70 | 0.70 |
| Lysine, % | 1.40 | 1.40 | 1.40 | 1.40 |
| Methionine, % | 0.45 | 0.45 | 0.45 | 0.45 |
| FAT, % | 3.82 | 3.72 | 3.95 | 4.2 |
| Lactose, % | 20.00 | 20.00 | 20.00 | 20.00 |
aFSBM0, FSBM3, FSBM6 and FSBM9 diets were made by substituting fermented soybean meal for 0, 3, 6 and 9% soybean meal.
bProvided per kg diet: Fe, 100 mg as ferrous sulfate; Cu, 17 mg as copper sulfate; Mn, 17 mg as manganese oxide; I, 0.5 mg as potassium iodide; and Se, 0.3 mg as sodium selenite.
cProvided per kilograms of diet: vitamin A, 10,800 IU; vitamin D3, 4000 IU; vitamin E, 40 IU; vitamin K3, 4 mg; vitamin B1, 6 mg; vitamin B2, 12 mg; vitamin B6, 6 mg; vitamin B12, 0.05 mg; biotin, 0.2 mg; folic acid, 2 mg; niacin, 50 mg; D-calcium pantothenate, 25 mg.
Table 3.
Composition of weaning pig diets (as fed-basis).
| Item | Dietary treatmentsa Phase 2 (8–21) | |||
|---|---|---|---|---|
| FSBM0 | FSBM3 | FSBM6 | FSBM9 | |
| Ingredients (%) | ||||
| Corn | 54.75 | 55.34 | 55.1 | 54.91 |
| Soybean meal | 27.42 | 25.30 | 22.88 | 20.42 |
| FSBM | – | 3.00 | 6.00 | 9.00 |
| Spray-dried plasma protein | 1.50 | 1.50 | 1.50 | 1.50 |
| Tallow | 2.73 | 3.31 | 2.99 | 2.65 |
| Coconut powder (60% fat) | 2.00 | – | – | – |
| Lactose | 3.89 | 3.89 | 3.89 | 3.89 |
| Sugar | 2.00 | 2.00 | 2.00 | 2.00 |
| Whey protein | 2.00 | 2.00 | 2.00 | 2.00 |
| Monodicalcium phosphate | 1.58 | 1.56 | 1.56 | 1.56 |
| Limestone | 0.98 | 0.99 | 0.99 | 0.99 |
| Salt | 0.10 | 0.10 | 0.10 | 0.10 |
| Methionine (99%) | 0.17 | 0.16 | 0.16 | 0.16 |
| Lysine | 0.45 | 0.42 | 0.4 | 0.39 |
| Mineral mixtureb | 0.20 | 0.20 | 0.20 | 0.20 |
| Vitamin mixturec | 0.20 | 0.20 | 0.20 | 0.20 |
| Choline (25%) | 0.03 | 0.03 | 0.03 | 0.03 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 |
| Calculated chemical composition | ||||
| Crude protein, % | 19.50 | 19.50 | 19.50 | 19.50 |
| Metabolizable energy, kcal/kg | 3400 | 3400 | 3400 | 3400 |
| Calcium, % | 0.80 | 0.80 | 0.80 | 0.80 |
| Available phosphorus, % | 0.70 | 0.70 | 0.70 | 0.70 |
| Lysine, % | 1.40 | 1.40 | 1.40 | 1.40 |
| Methionine, % | 0.45 | 0.45 | 0.45 | 0.45 |
| FAT, % | 6.28 | 6.19 | 6.44 | 6.67 |
| Lactose, % | 4.98 | 4.98 | 4.98 | 4.98 |
aFSBM0, FSBM3, FSBM6 and FSBM9 diets were made by substituting fermented soybean meal for 0, 3, 6 and 9% soybean meal.
bProvided per kg diet: Fe, 100 mg as ferrous sulfate; Cu, 17 mg as copper sulfate; Mn, 17 mg as manganese oxide; I, 0.5 mg as potassium iodide; and Se, 0.3 mg as sodium selenite.
cProvided per kilograms of diet: vitamin A, 10,800 IU; vitamin D3, 4000 IU; vitamin E, 40 IU; vitamin K3, 4 mg; vitamin B1, 6 mg; vitamin B2, 12 mg; vitamin B6, 6 mg; vitamin B12, 0.05 mg; biotin, 0.2 mg; folic acid, 2 mg; niacin, 50 mg; D-calcium pantothenate, 25 mg.
Table 4.
Composition of weaning pig diets (as fed-basis).
| Item | Dietary treatmentsa Phase 3 (22–42) | |||
|---|---|---|---|---|
| FSBM0 | FSBM3 | FSBM6 | FSBM9 | |
| Ingredients (%) | ||||
| Corn | 59.12 | 59.7 | 59.47 | 59.24 |
| Soybean meal | 28.32 | 26.18 | 23.76 | 21.34 |
| FSBM | – | 3.00 | 6.00 | 9.00 |
| Tallow | 2.00 | 2.60 | 2.27 | 1.94 |
| Coconut powder (60% fat) | 2.00 | – | – | – |
| Lactose | 2.69 | 2.69 | 2.69 | 2.69 |
| Sugar | 2.00 | 2.00 | 2.00 | 2.00 |
| Monodicalcium phosphate | 1.64 | 1.64 | 1.64 | 1.64 |
| Limestone | 0.98 | 0.98 | 0.98 | 0.98 |
| Salt | 0.10 | 0.10 | 0.10 | 0.10 |
| Methionine (99%) | 0.17 | 0.16 | 0.16 | 0.16 |
| Lysine | 0.55 | 0.52 | 0.50 | 0.48 |
| Mineral mixtureb | 0.20 | 0.20 | 0.20 | 0.20 |
| Vitamin mixturec | 0.20 | 0.20 | 0.20 | 0.20 |
| Choline (25%) | 0.03 | 0.03 | 0.03 | 0.03 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 |
| Calculated chemical composition | ||||
| Crude protein, % | 19.00 | 19.00 | 19.00 | 19.00 |
| Metabolizable energy, kcal/kg | 3350 | 3350 | 3350 | 3350 |
| Calcium, % | 0.80 | 0.80 | 0.80 | 0.80 |
| Available phosphorus, % | 0.70 | 0.70 | 0.70 | 0.70 |
| Lysine, % | 1.40 | 1.40 | 1.40 | 1.40 |
| Methionine, % | 0.45 | 0.45 | 0.45 | 0.45 |
| FAT, % | 5.72 | 5.64 | 5.88 | 6.12 |
| Lactose, % | 2.50 | 2.50 | 2.50 | 2.50 |
aFSBM0, FSBM3, FSBM6 and FSBM9 diets were made by substituting fermented soybean meal for 0, 3, 6 and 9% soybean meal.
bProvided per kg diet: Fe, 100 mg as ferrous sulfate; Cu, 17 mg as copper sulfate; Mn, 17 mg as manganese oxide; I, 0.5 mg as potassium iodide; and Se, 0.3 mg as sodium selenite.
cProvided per kilograms of diet: vitamin A, 10,800 IU; vitamin D3, 4000 IU; vitamin E, 40 IU; vitamin K3, 4 mg; vitamin B1, 6 mg; vitamin B2, 12 mg; vitamin B6, 6 mg; vitamin B12, 0.05 mg; biotin, 0.2 mg; folic acid, 2 mg; niacin, 50 mg; D-calcium pantothenate, 25 mg.
Growth performance
Each piglet was weighed on days 0, 7, 21 and 42 and feed consumption was also recorded on a pen basis to determine average daily gain (ADG), average daily feed intake (ADFI) and gain to feed ratio (G:F = ADG/ADFI).
Diarrhea score
The piglets’ anuses were checked one by one at 09:00 and 17:00 daily during the experiment to observe and recorded any fecal contamination and redness method by Ma et al.17. The number of piglets with diarrhea per treatment was counted at the days 1–21 and 22–42 and the diarrhea rate was calculated with the following formulation:
Apparent total tract digestibility
To determine dry matter, crude protein, and gross energy digestibility, chromium oxide was added to the diet as an indigestible marker at 2 g/kg of the diet for7 day prior to fecal collection. Fecal samples were collected from 8 pigs randomly selected per treatment via rectal massage, and the sample was stored in a freezer at − 20 °C and was dried in a 65 °C for 72 h and the feed and fecal samples were grounded to passed through 1-mm sieve for the measurement of dry matter, crude protein, and gross energy of FSBM, SBM, diets and feces samples were determined following to the Association of Official Analytical Chemists18 procedures.
Blood profile
At 06:00 on days 21 and 42, two pigs for each replicate was randomly selected to collect blood from the jugular vein and subsequently centrifuged at 3,000 × g for 15 min at 4 °C to obtain the serum sample and was kept at − 80 °C until analysis by Muniyappan et al.19 The concentrations of lymphocyte counts, Red blood cells (RBC), White blood cell (WBC), Blood urea nitrogen (BUN), and glucose in serum were measured with an automatic biochemical analyzer (Model 7020, Hitachi, Tokyo, Japan). The serum creatinine concentration was determined using anAstra-8 analyzer (Beckman Instruments, Inc., Brea, CA, US).
Characteristics of microbial population in feces
On day 42, fresh fecal samples were collected from six pigs for microbiota analysis. Genomic DNA of fecal samples was extracted by using a DNA Kit (Omega Bio-tek, Norcross, GA, USA), according to manufacturer’s instructions. The quantity and quality of extracted genomic DNA were checked using a UV spectrophotometer (Mecasys, Daejeon, Korea). Amplification and sequencing of the V5–V6 hypervariable region of the 16S rRNA gene was performed using an Illumina MiSeq platform (Illumina, San Diego, CA, USA). We performed alpha-diversity and taxonomic analyses of the raw paired-end sequences using EZBioCloud pipeline20. Then the samples were grouped into microbiome taxonomic profile sets for further analyses. Relative abundance cut-offs at the phylum and genus levels were set to 0.1%. Charts depicting the results from the alpha-diversity and taxonomic analyses were generated using GraphPad Prism software version 9.0 (GraphPad, San Diego, California, USA).
Statistical analysis
All results are presented as mean ± standard deviation (SD). GraphPad 9.0 was used for figures. Statistical analyses were performed using IBM SPSS 20.0 (SPSS Inc., Chicago, IL, USA) and the differences between treatments were compared with one-way ANOVA followed by Dunnett’s multiple comparison procedure. For all tests, P < 0.05 was considered as significant.
Results
Growth performance
As shown the Fig. 1, the BW of piglets fed dietary FSBM was higher than SBM. The 3% and 6% FSBM (P < 0.05) during days 7, 21 and 42, 9% FSBM (P < 0.05) during days 7 and 22, (P < 0.01) during day 42 all showed significant increases. Compared with SBM treatments, piglets fed 3, 6 and 9% FSBM (P < 0.05) on the higher ADG, ADFI and G:F during days 1–7, 8–21, 22–42 and 1–42. Importantly, the diarrhea rate lower in FSBM treatments (1–21, 22–42). (P < 0.05).
Figure 1.

Effect of fermented soybean meal (FSBM) on growth performance in piglets. (A) Body weight (BW). (B) Average daily gain (ADG). (C) Average daily feed intake (ADFI). (D) Gain-to-feed ratio (G:F). Data are presented as means ± SD. *P < 0.05, **P < 0.01. SBM, soybean meal; FSBM, fermented soybean meal.
Diarrhea score
Piglets fed FSBM diets had significantly lower (P < 0.05) the diarrhea score on days 1–21 and 22–42 (Fig. 2).
Figure 2.
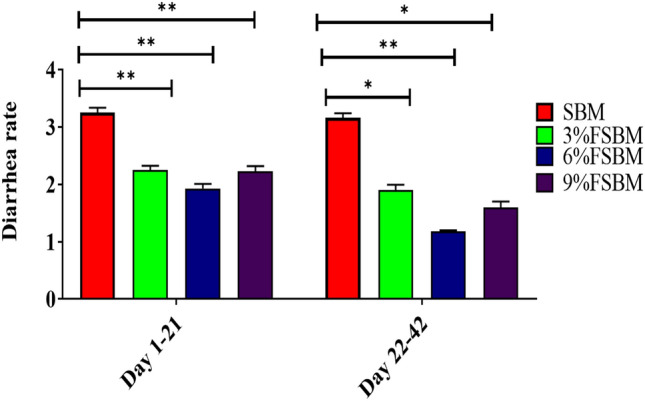
Effect of fermented soybean meal (FSBM) on diarrhea rate in piglets. Data are presented as means ± SD. *P < 0.05, **P < 0.01. SBM, soybean meal; FSBM, fermented soybean meal.
Apparent total tract digestibility
Dietary supplementation with 3, 6 and 9% FSBM increased (P < 0.05) the ATTD of dry matter, crude protein, and gross energy compared to the SBM (Fig. 3). Higher (P < 0.05) dry matter was found in the piglets dietary supplemented with 6% FSBM than 3 and 9% FSBM. However, 9% FSBM diet showed elevated crude protein levels than piglets fed 3 and 6% FSBM.
Figure 3.

Effect of fermented soybean meal (FSBM) on apparent total tract digestibility in piglets. (A) Dry matter. (B) Crude protein. (C) Gross energy. *P < 0.05, **P < 0.01. SBM, soybean meal; FSBM, fermented soybean meal.
Blood profile
As illustrated in Fig. 4, piglets diets FSBM diets had increased (P < 0.05) the glucose levels, WBC, RBC, and lymphocytes and lowered (P < 0.05) the BUN level on days day 21 and 42. High (P < 0.05) glucose, RBC, WBC, Lymphocyte, and lower BUN compared with SBM was observed (Fig. 3) in serum gathered from days 21 and 42 piglets supplemented with the 6% FSBM. BUN was lower in the latter two group than in 3 and 6% FSBM. The Creatinine was not influenced in the serum by feeding 3, 6 and 9%FSBM.
Figure 4.

Effect of fermented soybean meal (FSBM) on blood profiles in piglets. (A) Glucose levels. (B) White blood cell (WBC). (C) Red blood cell (RBC). (D) Lymphocyte. (E) Blood urea nitrogen (BUN). (F) Creatinine. *P < 0.05, **P < 0.01. SBM, soybean meal; FSBM, fermented soybean meal.
Effects of FSBM on fecal microbial composition
The OTUs Venn analysis identified 21, 28, 46 and 69 unique OTUs among FSBM and SBM treatments, respectively (Fig. 5A). Figure 4B illustrates the results of alpha diversity analysis of which the Shannon, Simpsons and Chao indices were improved (P < 0.05) on FSBM compared to the SBM (Fig. 5B).
Figure 5.

Fecal microbial richness and diversity. (A) Venn diagrams of dietary treatments at the OTUs levels. (B) Microbial richness estimates (Observed species, Chao, and Ace) and diversity indices (Shannon and Simpson). *P < 0.05, **P < 0.01. SBM, soybean meal; FSBM, fermented soybean meal.
As shown in Fig. 6, the phylum levels analysis illustrated that the dietary supplementation of FSBM higher in the abundance of Firmicutes (P < 0.05) and lower the abundance of Bacteroidetes and Proteobacteria (P < 0.05). At the genus level, FSBM group increased Lactobacillus, prevotella, Lachnospiraceae and Lachnoclostridium (P < 0.05) in fecal microbiota (P < 0.05). Compared with SBM treatment, FSBM treatments lower in the abundance of Escherichia-Shigella, Clostridium sensu stricto1, Bacteroides and Parabacteroides (P < 0.05) (Fig. 7).
Figure 6.

Effects of dietary FSBM on fecal microbial composition at the phylum level. *P < 0.05, **P < 0.01, and ***P < 0.001. SBM, soybean meal; FSBM, fermented soybean meal.
Figure 7.

Effects of dietary FSBM on fecal microbial composition at the genus level. *P < 0.05, **P < 0.01, and ***P < 0.001. SBM, soybean meal; FSBM, fermented soybean meal.
Discussion
Growth performance and diarrhea score
Sources of plant protein like FSBM have been extensively used in the diets of weaning pigs to enhance growth and immune status2,4. Earlier studies, illustrated that dietary FSBM supplementation increased the growth performance on weaning pigs21–23 and broilers9. Zhu et al.24 reported a significantly in the increased growth performance in weaning piglets due to dietary FSBM supplementation. In current study, the BW, ADG, and ADFI were improved in weaning pigs supplemented with FSBM treatments. Trypsin inhibitor content and antigenic protein content was reduced to 0.33 and 20%, respectively. Stachyose and raffinose were completely absent in FSBM. Pigs fed with FSBM could improve nutrient digestibility that may explain an enhanced ADG in weaning pigs.
Diarrhea score
The positive effect of SBM substitution by FSBM on diarrhea reduction may be attributed to the degradation of the main antigenic soybean proteins such as α and α′ subunits of β-conglycinin and acidic subunits of glycinin and some antinutritional factors such as trypsin inhibitors25,26. These proteins are partially digested by proteases secreted by microorganisms responsible for the fermentation process27,28. Similarly, Jeong et al.11 showed that degradation of most antigenic proteins (glycinin and β-conglycinin) and protease inhibitors in SBM fermented by Enterococcus Faecium improved intestinal morphology and digestive enzyme activities in weaned pigs. Fermentation of soybeans also degrades its proteins and carbohydrates to the extent of low molecular weight and water-soluble compounds that facilitate their digestibility29. The combination of a better nutritional status and a reduced immunological challenge when pigs are fed FSBM, or FSB may help to prevent diarrhea after weaning. Previous study, we demonstrated that dietary FSBM inclusion reduce the diarrhea in weaning pigs24,30. In the current study, piglets fed FSBM decreased the diarrhea in weaning pigs. In addition, it has been proposed that some microorganisms present in fermented products can inhibit intestinal colonization of pathogens that causes diarrhea in pigs31. The reduction of diarrhea is of great importance in pig production as it decreases the predisposition of these animals to Escherichia coli infection and improves feed efficiency, especially in weaning pigs32.
Apparent total tract digestibility
In the current study, supplementation of FSBM significantly increased digestibility of crude protein, dry matter, and gross energy which was consistent with Yan and Kim33 and Muniyappan et al.15 who reported that FSBM inclusion increased apparent total tract of crude protein, dry matter, and gross energy in weaning pigs. The beneficial effects of FSBM on nutrient digestibility may be attributed to the effects of FSBM on gastrointestinal development and the concomitant increase in digestive enzyme secretion4. Similarly, Min et al.34 found that dietary FSBM did not have significant effect on the digestibility of dry matter, crude protein, and gross energy in weaning pigs. In contrast, the negative effects of crude protein, dry matter, and gross energy on nutrient digestibility were also observed in previous studies13,35. Similarly, Hossain et al.36 and Lan and Kim37 found that dietary inclusion of FSMB did not have a significant effect on the digestibility of crude protein, dry matter, and gross energy in weaning pigs. The inconsistent results may be related to many factors such as FSBM source, additional levels, and composition of the basal diet, with effects being more pronounced when moderate amounts of FSBM are added to SBM basal diets21,34.
Blood profile
The BUN concentration and producing are affected by way of protein catabolism, and its concentrating is negatively associated with digestibility of proteins and amino acids38. Creatinine exists a natural waste result arises from the muscles and is eliminated from the body through kidney. Pigs fed FSBM showed decreased diarrhea score and the glucose levels improved which was in accordance with earlier studies24,39. Intriguingly, serum blood urea nitrogen reduced in FSBM treatments compared with the control. This shown that the fermenting procedure change nitrogen distribution inside the feed40. The levels of white blood cells, lymphocytes, were also increased, in the current studies. Lymphocyte growth exists as a major phase during the immune reaction in an animal and a proliferative reaction is a specific antigen41. Gizzarelli et al.42 and Wang et al.43 reported that weaning pig immunity is lower on β-conglycinin is not adequate deactivated during fermentation. Our results found a higher immune resistance on piglets this directly corresponded with a decrease in glycinin (80.29%) and β-conglycinin (69.43%) on FSBM. Therefore, overall growth performance and blood profiles were in accordance with each other.
Effects of FSBM on fecal microbial composition
Diets offer available substrates inclusive of protein carbohydrates for the gut microbiota and affect microbial structure and metabolism might encourage the performance and gut health of weaned pigs2. Generally, dietary nutrients are absorbed and digested in the foregut, later undigested food residues and endogenous are fermented by gut microbiota in the hindgut44,45. Fermentation of carbohydrates is enhanced to colonic cells due to the production of SCFA2,45, during fermentation of indigestible protein manufacture potentially metabolite toxicity inclusive of Ammonia, amines, phenols, and indoles that have harmful effects on intestinal health44. Alpha diversity can be utilized as an indicator of the functional resilience of ecological diversity of the intestinal microbiota, including species richness indices (Observed species, Chao, and Ace), and species diversity (Shannon and Simpson)46. In the current study, dietary inclusion with FSBM significantly increased the Shannon, Simpson, and Chao1 richness, which was in accordance with our previous findings in pigs that dietary inclusion of FSBM significant difference the Shannon, Simpson47. Bacteroidetes, Firmicutes, Actinobacteria, Proteobacteria, and Tenericutes were the most pre-dominant bacteria phylum in the piglets48. In the current study, FSBM group shapes gut microbiota in weaning pigs, including lower in the abundances of the phylum Bacteroidetes, and Proteobacteria and the genera Escherichia-Shigella, Clostridium sensu stricto1, Bacteroides and Parabacteroides, and a higher in the abundances of the phylum Firmicutes and the genera Lactobacillus, prevotella, Lachnospiraceae and Lachnoclostridium. Bacteroidetes, Proteobacteria and Firmicutes as three major communities, are essential to growth performance and energy metabolism homeostasis49,50. Shin et al.51 found a lower in the abundance of the phylum Proteobacteria in the gut of healthy humans. Therefore, Litvak et al.52 reported that increase in Proteobacteria abundance have been associated with in humans with gut inflammation, colorectal cancer, irritable bowel syndrome and metabolic syndrome and could be bacterial signature of gut dysbiosis51. The abundance of the phylum Proteobacteria contains many possible opportunistic pathogens, as well as Campylobacter spp, Klebsiella spp,Escherichia, and Salmonella and its rise could be shown as a potential indicator of gut diseases. The Firmicutes abundance have been evidence to be positive relationship with energy and active transport, facilitated diffusion, endocytosis and passive diffusion, whereas improve in fecal Proteobacteria and Bacteroidetes is associated with inferior nutrient digestibility2. Therefore, higher abundances of the phylum Firmicutes along with lower abundances of the phylum Proteobacteria and Bacteroidetes might promote nutrient digestibility in weaning pigs. Eren et al.53 reported that the herbivores have an increase abundance of Lachnospiraceae than carnivores in animal. Vacca et al.54 reported that reduce abundance of Lachnospiraceae, while multiple sclerosis and ulcerative colitis patients. All participants of Lachnospiraceae are anaerobic, hemoorganotrophic and fermentative and could degrade non-starch polysaccharides and butyrate and acetic acid. Butyrate provide the major energy source for intestinal epithelial cell growth, increased intestinal protection mediated epithelial cells and favoring the suppression of inhibits inflammatory responses55,56. Stanley et al.57 showed that Lachnospiraceae is associated with improve growth performance in animals. The abundances of the genera Lachnoclostridium—butyric-acid-producing microbes that have been associated in the mitigation on intestinal inflammation- were better in the FSBM groups10. The Lactobacillus as a possibility probiotic, possess the opposition to pathogen, anti– inflammatory, antioxidant capacity, and capability to higher of fecal microbiota58,59. Zhu et al.48 reported an improved Lactobacilli and totality anaerobic bacteria counts in the gut microbiota weaning piglets due to dietary FSBM supplementation. In the current study, inclusion of FSBM treatments an increased in the abundances of the genera Lactobacillus were significantly increased RBC, WBC, Lymphocyte, glucose levels and decreased BUN. Lactobacillus is familiar to have a positive influence on the GIT, growth performance, and nutrient digestibility in pigs and regularly used as probiotics in livestock production60,61. Yan et al.62 reported that Lactobacillus could commonly increase growth performance and the GIT of animals by defense the intestine from pathogens and encourage efficient nutrient and energy extraction by the host. The genus Prevotella is saccharolytic and produce succinic and acetic acids as ending fermentation products63. Prevotella specialized in degrading fiber diet, which had also been correspondent with could improve intestinal immune and decrease diarrhea64. Wu et al.65 reported evidence indicated a close relationship among Prevotella and long period of time carbohydrates diets or carbohydrates from fiber-rich diets. It was also reported that a higher abundance of Prevotellaceae dominated in fecal microbiota of healthy piglets when compared to post-weaning diarrheic piglets 64. Feng et al.47 and Forsyth et al.66 reported a higher Prevotella abundance, the higher the mucin composite content, but bacterial toxins would reduce gut penetrability and the sensitivity of the region-specific gut to systemic exposure. Accordingly, higher in the Prevotella abundances in FSBM it could be helpful in improve on growth performance in weaning piglets. The relative abundance of Clostridium sensu stricto1 and bacteoides showed a positively correlated with the frequency of diarrhea. Clostridium is the main cause of diarrhoea in humans and is responsible for community-acquired out breaks67. Harlow et al.68 reported that Clostridium perfringens, Clostridium difficile and Salmonella spp. are the most common microbes associated with diarrhea. In our study, lower in the abundances of the genera Clostridium sensu stricto1 in the FSBM group, and was positively correlated with decreased diarrhoea. Another result of FSBM groups were a decrease Escherichia-Shigella, and lower Escherichia-Shigella was main participant of the decreased in Proteobacteria abundance compared to the SBM treatments. The genera Escherichia–Shigella comprises an opportunistically pathogenic bacterium. Sousa et al.69 and Gong et al.70 reported the genera Escherichia–Shigella could destroy the gut structure and must be pro-inflammatory actions through multiple pathways and like the secretion of virulence factors, consequent in the improved danger of infection and diarrhea in the host. Parabacteroides and Bacteroides, which occur at the initial phase of the lifetime, have been informed to generate gamma amino butyric acid, closely related to growth71,72. A higher abundance of the genera Bacteroides is commonly used in the events of colorectal cancer, functional gastrointestinal disorders and ulcerative colitis73–75. Therefore, the biological results found that appropriate inclusion of FSBM in diets can inhibit the gut pathogens (such as Escherichia–Shigella, Bacteroides and Parabacteroides) and enhance beneficial bacteria (such as Prevotella and Lactobacillus) and further enhance the immunity and health status of weaning piglets.
Conclusions
In conclusion, FSBM replacing SBM improved the growth performance, apparent total tract digestibility, blood profiles, possibly via altering gut microbiota profile in weaned piglets. The present study provides theoretical support for applying FSBM at 6 to 9% promote blood profiles and regulate intestinal health in weaning piglets.
Acknowledgements
This research was supported by Basic Science Research Capacity Enhancement Project through Korea Basic Science Institute (National research Facilities and Equipment Center) grant funded by the Ministry of Education (Grant No. 2019R1A6C1010033). The Department of Animal Resource & Science was supported through the Research-Focused Department Promotion & Interdisciplinary Convergence Research Projects as a part of the University Innovation Support Program for Dankook University in 2022.
Author contributions
M.M., S.S., J.H.P., K.H. and I.H.K.: Conceptualization and designed the trials. M.M.: writing – original draft preparation, performed the animal trials, M.M., S.S., I.H.K., J.H.P. and K.H.: Software, Methodology, Formal analysis, Writing – review and editing, I.H.K.: Supervision. All authors contributed to the article and approved the submitted version.
Data availability
All data generated in this study are included in the published article. The datasets generated during the current study are available from the corresponding author on demand upon reasonable request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Madesh Muniyappan and Sureshkumar Shanmugam.
Contributor Information
Kyudong Han, Email: jim97@dankook.ac.kr.
In Ho Kim, Email: inhokim@dankook.ac.kr.
References
- 1.Hu R, He Z, Liu M, Tan J, Zhang H, Hou DX, Wu S. Dietary protocatechuic acid ameliorates inflammation and up-regulates intestinal tight junction proteins by modulating gut microbiota in LPS-challenged piglets. J. Anim. Sci. Biotechnol. 2020;11:1–12. doi: 10.1186/s40104-020-00492-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zhang L, Piao X. Different dietary protein sources influence growth performance, antioxidant capacity, immunity, fecal microbiota and metabolites in weaned piglets. Anim. Nutr. 2022;8:71–81. doi: 10.1016/j.aninu.2021.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ma XK, Shang QH, Wang QQ, Hu JX, Piao XS. Comparative effects of enzymolytic soybean meal and antibiotics in diets on growth performance, antioxidant capacity, immunity, and intestinal barrier function in weaned pigs. Anim. Feed Sci. Technol. 2019;248:47–58. doi: 10.1016/j.anifeedsci.2018.12.003. [DOI] [Google Scholar]
- 4.Ma X, Shang Q, Hu J, Liu H, Brøkner C, Piao X. Effects of replacing soybean meal, soy protein concentrate, fermented soybean meal or fish meal with enzyme-treated soybean meal on growth performance, nutrient digestibility, antioxidant capacity, immunity and intestinal morphology in weaned pigs. Livest. Sci. 2019;225:39–46. doi: 10.1016/j.livsci.2019.04.016. [DOI] [Google Scholar]
- 5.Gu X, Li Z, Wang J, Chen J, Jiang Q, Liu N, Ma X. Fermented cottonseed meal as a partial replacement for soybean meal could improve the growth performance, immunity and antioxidant properties, and nutrient digestibility by altering the gut microbiota profile of weaned piglets. Front. Microbiol. 2021;12:100. doi: 10.3389/fmicb.2021.734389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wang Y, Wu Y, Wang B, Cao X, Fu A, Li Y, Li W. Effects of probiotic Bacillus as a substitute for antibiotics on antioxidant capacity and intestinal autophagy of piglets. AMB Express. 2017;7:1–11. doi: 10.1186/s13568-017-0353-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sureshkumar S, Park JH, Kim IH. Effects of Enterococcus faecium SLB 130 probiotic on the performance of weaning pigs. Vet. Med. 2022;67:562–568. doi: 10.17221/41/2022-VETMED. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Scharek-Tedin L, Kreuzer-Redmer S, Twardziok SO, Siepert B, Klopfleisch R, Tedin K, Pieper R. Probiotic treatment decreases the number of CD14-expressing cells in porcine milk which correlates with several intestinal immune parameters in the piglets. Front. Immunol. 2015;6:108. doi: 10.3389/fimmu.2015.00108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cheng YH, Hsiao FSH, Wen CM, Wu CY, Dybus A, Yu YH. Mixed fermentation of soybean meal by protease and probiotics and its effects on the growth performance and immune response in broilers. J. Appl. Anim. Res. 2019;47:339–348. doi: 10.1080/09712119.2019.1637344. [DOI] [Google Scholar]
- 10.Li Y, Guo B, Wu Z, Wang W, Li C, Liu G, Cai H. Effects of fermented soybean meal supplementation on the growth performance and cecal microbiota community of broiler chickens. Animals. 2020;10:1098. doi: 10.3390/ani10061098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jeong JS, Kim IH. Comparative efficacy of up to 50% partial fish meal replacement with fermented soybean meal or enzymatically prepared soybean meal on growth performance, nutrient digestibility and fecal microflora in weaned pigs. Anim. Sci. J. 2015;86:624–633. doi: 10.1111/asj.12335. [DOI] [PubMed] [Google Scholar]
- 12.Song YS, Pérez VG, Pettigrew JE, Martinez-Villaluenga C, de Mejia EG. Fermentation of soybean meal and its inclusion in diets for newly weaned pigs reduced diarrhea and measures of immunoreactivity in the plasma. Anim. Feed Sci. Technol. 2010;159:41–49. doi: 10.1016/j.anifeedsci.2010.04.011. [DOI] [Google Scholar]
- 13.Jeong JS, Park JW, Lee SI, Kim IH. Apparent ileal digestibility of nutrients and amino acids in soybean meal, fish meal, spray-dried plasma protein and fermented soybean meal to weaned pigs. Anim. Sci. J. 2016;87:697–702. doi: 10.1111/asj.12483. [DOI] [PubMed] [Google Scholar]
- 14.Cervantes-Pahm SK, Stein HH. Ileal digestibility of amino acids in conventional, fermented, and enzyme-treated soybean meal and in soy protein isolate, fish meal, and casein fed to weanling pigs. J. Anim. Sci. 2010;88:2674–2683. doi: 10.2527/jas.2009-2677. [DOI] [PubMed] [Google Scholar]
- 15.Muniyappan M, Lee Y, Kim IH. Comparative efficacy of soybean meal vs fermented soybean meal on ileal digestibility and urine contents in weaned pigs. Livest. Sci. 2022;263:105016. doi: 10.1016/j.livsci.2022.105016. [DOI] [Google Scholar]
- 16.Weiner ML, Ferguson HE, Thorsrud BA, Nelson KG, Blakemore WR, Zeigler B, Mahadevan B. An infant formula toxicity and toxicokinetic feeding study on carrageenan in preweaning piglets with special attention to the immune system and gastrointestinal tract. Food Chem. Toxicol. 2015;77:120–131. doi: 10.1016/j.fct.2014.12.022. [DOI] [PubMed] [Google Scholar]
- 17.Ma J, Long S, Wang J, Gao J, Piao X. Microencapsulated essential oils combined with organic acids improves immune antioxidant capacity and intestinal barrier function as well as modulates the hindgut microbial community in piglets. J. Anim. Sci. Biotechnol. 2022;13:1–17. doi: 10.1186/s40104-021-00670-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.AOAC International. Official Methods of Analysis of AOAC. International, 16th edn. Association of Official Analytical Chemists, Washington, DC, USA (2000).
- 19.Muniyappan M, Palanisamy T, Kim IH. Effect of microencapsulated organic acids on growth performance, nutrient digestibility, blood profile, fecal gas emission, fecal microbial, and meat-carcass grade quality of growing-finishing pigs. Livest. Sci. 2021;252:104658. doi: 10.1016/j.livsci.2021.104658. [DOI] [Google Scholar]
- 20.Yoon SH, Ha SM, Kwon S, Lim J, Kim Y, Seo H, Chun J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017;67(5):1613. doi: 10.1099/ijsem.0.001755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kim SW, Van Heugten E, Ji F, Lee CH, Mateo RD. Fermented soybean meal as a vegetable protein source for nursery pigs: I. Effects on growth performance of nursery pigs. J. Anim. Sci. 2010;88:214–224. doi: 10.2527/jas.2009-1993. [DOI] [PubMed] [Google Scholar]
- 22.Jones CK, DeRouchey JM, Nelssen JL, Tokach MD, Dritz SS, Goodband RD. Effects of fermented soybean meal and specialty animal protein sources on nursery pig performance. J. Anim. Sci. 2010;88:1725–1732. doi: 10.2527/jas.2009-2110. [DOI] [PubMed] [Google Scholar]
- 23.Zhang HY, Yi JQ, Piao XS, Li PF, Zeng ZK, Wang D, Han X. The metabolizable energy value, standardized ileal digestibility of amino acids in soybean meal, soy protein concentrate and fermented soybean meal, and the application of these products in early-weaned piglets. Asian Australas. J. Anim. Sci. 2013;26:691. doi: 10.5713/ajas.2012.12429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhu J, Gao M, Zhang R, Sun Z, Wang C, Yang F, Hao Z. Effects of soybean meal fermented by L. plantarum, B. subtilis and S. cerevisieae on growth, immune function and intestinal morphology in weaned piglets. Microb. Cell Factories. 2017;16:1–10. doi: 10.1186/s12934-017-0809-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hong KJ, Lee CH, Kim SW. Aspergillus oryzae GB-107 fermentation improves nutritional quality of food soybeans and feed soybean meals. J. Med. Food. 2004;7:430–435. doi: 10.1089/jmf.2004.7.430. [DOI] [PubMed] [Google Scholar]
- 26.Min BJ, Hong JW, Kwon OS, Lee WB, Kim YC, Kim IH, Kim JH. The effect of feeding processed soy protein on the growth performance and apparent ileal digestibility in weanling pigs. Asian Australas. J. Anim. Sci. 2004;17:1271–1276. doi: 10.5713/ajas.2004.1271. [DOI] [Google Scholar]
- 27.Frias J, Song YS, Martínez-Villaluenga C, De Mejia EG, Vidal-Valverde C. Immunoreactivity and amino acid content of fermented soybean products. J. Agric. Food Chem. 2008;56:99–105. doi: 10.1021/jf072177j. [DOI] [PubMed] [Google Scholar]
- 28.Feng J, Liu X, Xu ZR, Lu YP, Liu YY. The effect of Aspergillus oryzae fermented soybean meal on growth performance, digestibility of dietary components and activities of intestinal enzymes in weaned piglets. Anim. Feed Sci. Technol. 2007;134:295–303. doi: 10.1016/j.anifeedsci.2006.10.004. [DOI] [Google Scholar]
- 29.Kiers JL, Rombouts FM, Nout MJR. In vitro digestibility of Bacillus fermented soya bean. Int. J. Food Microbiol. 2000;60:163–169. doi: 10.1016/S0168-1605(00)00308-1. [DOI] [PubMed] [Google Scholar]
- 30.Kim YG, Lohakare JD, Yun JH, Heo S, Chae BJ. Effect of feeding levels of microbial fermented soy protein on the growth performance, nutrient digestibility and intestinal morphology in weaned piglets. Asian Australas. J. Anim. Sci. 2007;20:399–404. doi: 10.5713/ajas.2007.399. [DOI] [Google Scholar]
- 31.Kiers JL, Meijer JC, Nout MJR, Rombouts FM, Nabuurs MJA, Van der Meulen J. Effect of fermented soya beans on diarrhoea and feed efficiency in weaned piglets. J. Appl. Microbiol. 2003;95:545–552. doi: 10.1046/j.1365-2672.2003.02011.x. [DOI] [PubMed] [Google Scholar]
- 32.Pluske JR, Pethick DW, Hopwood DE, Hampson DJ. Nutritional influences on some major enteric bacterial diseases of pig. Nutr. Res. Rev. 2002;15:333–371. doi: 10.1079/NRR200242. [DOI] [PubMed] [Google Scholar]
- 33.Yan L, Kim IH. Effect of probiotics supplementation in diets with different nutrient densities on growth performance, nutrient digestibility, blood characteristics, faecal microbial population and faecal noxious gas content in growing pigs. J. Appl. Anim. Res. 2013;41:23–28. doi: 10.1080/09712119.2012.739092. [DOI] [Google Scholar]
- 34.Min BJ, Cho JH, Chen YJ, Kim HJ, Yoo JS, Lee CY, Kim IH. Effects of fermented soy protein on growth performance and blood protein contents in nursery pigs. Asian Australas. J. Anim. Sci. 2009;22(7):1038–1042. doi: 10.5713/ajas.2009.80240. [DOI] [Google Scholar]
- 35.Ding Z, Chang KH, Kim I. Effects of fermented soybean meal on growth performance, nutrients digestibility, blood profile and fecal microflora in weaning pigs. Korean J. Agric. Sci. 2020;47(1):1–10. doi: 10.7744/kjoas.20190062. [DOI] [Google Scholar]
- 36.Hossain MM, Begum M, Park JH, Lee SJ, Jang KH, Hong YH, Kim IH. Apparent total tract digestibility and ileal digestibility of dry matter, nitrogen, energy and amino acids in conventional, Bacillus subtilis-fermented and enzyme-treated soybean meal fed to weanling pigs. Vet. Med. 2016;61(12):669–680. doi: 10.17221/389/2014-VETMED. [DOI] [Google Scholar]
- 37.Lan R, Kim I. Enterococcus faecium supplementation in sows during gestation and lactation improves the performance of sucking piglets. Vet. Med. Sci. 2020;6:92–99. doi: 10.1002/vms3.215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Cai Y. Impact of nutritional and environmental factors on plasma urea and amino acid concentrations in pigs. Doctoral dissertation, Iowa State University (1992).
- 39.Shi C, He J, Wang J, Yu J, Yu B, Mao X, Chen D. Effects of Aspergillus niger fermented rapeseed meal on nutrient digestibility, growth performance and serum parameters in growing pigs. Anim. Sci. J. 2016;8:557–563. doi: 10.1111/asj.12457. [DOI] [PubMed] [Google Scholar]
- 40.Cho JH, Min BJ, Chen YJ, Yoo JS, Wang Q, Kim JD, Kim IH. Evaluation of FSP (fermented soy protein) to replace soybean meal in weaned pigs: Growth performance, blood urea nitrogen and total protein concentrations in serum and nutrient digestibility. Asian Australas. J. Anim. Sci. 2007;20:1874–1879. doi: 10.5713/ajas.2007.1874. [DOI] [Google Scholar]
- 41.Sadeghi G, Pourreza J. Serum proteins and some biochemical parameters in broiler chickens fed with raw and treated bitter vetch (Vicia ervilia) seeds. Pak. J. Biol. Sci. 2007;10:977–981. doi: 10.3923/pjbs.2007.977.981. [DOI] [PubMed] [Google Scholar]
- 42.Gizzarelli F, Corinti S, Barletta B, Iacovacci P, Brunetto B, Butteroni C, Tinghino R. Evaluation of allergenicity of genetically modified soybean protein extract in a murine model of oral allergen-specific sensitization. Clin. Exp. Allergy. 2006;36:238–248. doi: 10.1111/j.1365-2222.2005.02415.x. [DOI] [PubMed] [Google Scholar]
- 43.Wang X, Geng F, Wu J, Kou Y, Xu S, Sun Z, Luo Y. Effects of β-conglycinin on growth performance, immunoglobulins and intestinal mucosal morphology in piglets. Arch. Anim. Nutr. 2014;68:186–195. doi: 10.1080/1745039X.2014.919733. [DOI] [PubMed] [Google Scholar]
- 44.Li R, Chang L, Hou G, Song Z, Fan Z, He X, Hou DX. Colonic microbiota and metabolites response to different dietary protein sources in a piglet model. Front. Nutr. 2019;6:151. doi: 10.3389/fnut.2019.00151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Li R, Hou G, Jiang X, Song Z, Fan Z, Hou DX, He X. Different dietary protein sources in low protein diets regulate colonic microbiota and barrier function in a piglet model. Food. Funct. 2019;10:6417–6428. doi: 10.1039/C9FO01154D. [DOI] [PubMed] [Google Scholar]
- 46.Ijaz MU, Ahmed MI, Zou X, Hussain M, Zhang M, Zhao F, Li C. Beef, casein, and soy proteins differentially affect lipid metabolism, triglycerides accumulation and gut microbiota of high-fat diet-fed C57BL/6J mice. Front. Microbiol. 2018;9:2200. doi: 10.3389/fmicb.2018.02200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Feng H, Qu H, Liu Y, Shi Y, Wu S, Bao W. Effect of fermented soybean meal supplementation on some growth performance, blood chemical parameters, and fecal microflora of finishing pigs. R. Bras. Zootec. 2020 doi: 10.37496/rbz4920190096. [DOI] [Google Scholar]
- 48.Zhu JJ, Gao MX, Song XJ, Zhao L, Li YW, Hao ZH. Changes in bacterial diversity and composition in the faeces and colon of weaned piglets after feeding fermented soybean meal. J. Med. Microbiol. 2018;67:1181–1190. doi: 10.1099/jmm.0.000766. [DOI] [PubMed] [Google Scholar]
- 49.Singh KM, Shah T, Deshpande S, Jakhesara SJ, Koringa PG, Rank DN, Joshi CG. High through put 16S rRNA gene-based pyrosequencing analysis of the fecal microbiota of high FCR and low FCR broiler growers. Mol. Biol. Rep. 2012;39:10595–10602. doi: 10.1007/s11033-012-1947-7. [DOI] [PubMed] [Google Scholar]
- 50.Bai M, Liu H, Wang S, Shu Q, Xu K, Zhou J, Liu ZA. Dietary Moutan cortex Radicis improves serum antioxidant capacity and intestinal immunity and alters colonic microbiota in weaned piglets. Front. Nutr. 2021;8:266. doi: 10.3389/fnut.2021.679129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Shin NR, Whon TW, Bae JW. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015;33:496–503. doi: 10.1016/j.tibtech.2015.06.011. [DOI] [PubMed] [Google Scholar]
- 52.Litvak Y, Byndloss MX, Tsolis RM, Bäumler AJ. Dysbiotic Proteobacteria expansion: A microbial signature of epithelial dysfunction. Curr. Opin. Microbiol. 2017;39:1–6. doi: 10.1016/j.mib.2017.07.003. [DOI] [PubMed] [Google Scholar]
- 53.Eren AM, Sogin ML, Morrison HG, Vineis JH, Fisher JC, Newton RJ, McLellan SL. A single genus in the gut microbiome reflects host preference and specificity. ISME J. 2015;9:90–100. doi: 10.1038/ismej.2014.97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Vacca M, Celano G, Calabrese FM, Portincasa P, Gobbetti M, De Angelis M. The controversial role of human gut lachnospiraceae. Microorganisms. 2020;8:573. doi: 10.3390/microorganisms8040573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Yang P, Zhao J. Variations on gut health and energy metabolism in pigs and humans by intake of different dietary fibers. Food Sci. Nutr. 2021;9:4639–4654. doi: 10.1002/fsn3.2421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Gelmez E, Jeron A, Bruder D. Negative elongation factor: A key factor in the maintenance of intestinal epithelial barrier integrity. Cell Mol. Immunol. 2022 doi: 10.1038/s41423-021-00817-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Stanley D, Hughes RJ, Geier MS, Moore RJ. Bacteria within the gastrointestinal tract microbiota correlated with improved growth and feed conversion: Challenges presented for the identification of performance enhancing probiotic bacteria. Front. Microbiol. 2016;7:187. doi: 10.3389/fmicb.2016.00187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.He T, Zhu YH, Yu J, Xia B, Liu X, Yang GY, Wang JF. Lactobacillus johnsonii L531 reduces pathogen load and helps maintain short-chain fatty acid levels in the intestines of pigs challenged with Salmonella enterica Infantis. Vet. Microbiol. 2019;230:187–194. doi: 10.1016/j.vetmic.2019.02.003. [DOI] [PubMed] [Google Scholar]
- 59.Yang A, Zuo L, Cheng Y, Wu Z, Li X, Tong P, Chen H. Degradation of major allergens and allergenicity reduction of soybean meal through solid-state fermentation with microorganisms. Food. Funct. 2018;9:1899–1909. doi: 10.1039/C7FO01824J. [DOI] [PubMed] [Google Scholar]
- 60.Sato Y, Kuroki Y, Oka K, Takahashi M, Rao S, Sukegawa S, Fujimura T. Effects of dietary supplementation with enterococcus faecium and clostridium butyricum, either alone or in combination, on growth and fecal microbiota composition of post-weaning pigs at a commercial farm. Front. Vet. Sci. 2019;6:26. doi: 10.3389/fvets.2019.00026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.He T, Zheng Y, Piao X, Long S. Determination of the available energy, standardized ileal digestibility of amino acids of fermented corn germ meal replacing soybean meal in growing pig diets. Anim. Nutr. 2022;9:259–268. doi: 10.1016/j.aninu.2021.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yan W, Sun C, Yuan J, Yang N. Gut metagenomic analysis reveals prominent roles of Lactobacillus and cecal microbiota in chicken feed efficiency. Sci. Rep. 2017;7:1–11. doi: 10.1038/srep45308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Yu T, Wang Y, Chen S, Hu M, Wang Z, Wu G, Zheng C. Low-molecular-weight chitosan supplementation increases the population of Prevotella in the cecal contents of weanling pigs. Front. Microbial. 2017;8:2182. doi: 10.3389/fmicb.2017.02182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Dou S, Gadonna-Widehem P, Rome V, Hamoudi D, Rhazi L, Lakhal L, Abdennebi-Najar L. Characterisation of early-life fecal microbiota in susceptible and healthy pigs to post-weaning diarrhoea. PLoS ONE. 2017;12:e0169851. doi: 10.1371/journal.pone.0169851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wu GD, Chen J, Hoffmann C, Bittinger K, Chen YY, Keilbaugh SA, Lewis JD. Linking long-term dietary patterns with gut microbial enterotypes. Science. 2011;334:105–108. doi: 10.1126/science.1208344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Forsyth CB, Shannon KM, Kordower JH, Voigt RM, Shaikh M, Jaglin JA, Keshavarzian A. Increased intestinal permeability correlates with sigmoid mucosa alpha-synuclein staining and endotoxin exposure markers in early Parkinson’s disease. PLoS ONE. 2011;6:e28032. doi: 10.1371/journal.pone.0028032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Alcalá L, Reigadas E, Marín M, Martín A, Catalán P, Bouza E. Impact of clinical awareness and diagnostic tests on the underdiagnosis of Clostridium difficile infection. Eur. J. Clin. Microbiol. Infect. Dis. 2015;34:1515–1525. doi: 10.1007/s10096-015-2380-3. [DOI] [PubMed] [Google Scholar]
- 68.Harlow BE, Lawrence LM, Flythe MD. Diarrhea-associated pathogens, lactobacilli and cellulolytic bacteria in equine feces: Responses to antibiotic challenge. Vet. Microbiol. 2013;166:225–232. doi: 10.1016/j.vetmic.2013.05.003. [DOI] [PubMed] [Google Scholar]
- 69.Sousa MAB, Mendes EN, Apolônio ACM, Farias LDM, Magalhaes PP. Bacteriocin production by Shigella sonnei isolated from faeces of children with acute diarrhoea. APMIS. 2010;118:125–135. doi: 10.1111/j.1600-0463.2009.02570.x. [DOI] [PubMed] [Google Scholar]
- 70.Gong Y, Yang H, Wang X, Xia W, Lv W, Xiao Y, Zou X. Early intervention with cecal fermentation broth regulates the colonization and development of gut microbiota in broiler chickens. Front. Microbiol. 2019 doi: 10.3389/fmicb.2019.01422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Yan H, Wei W, Hu L, Zhang Y, Zhang H, Liu J. Reduced feeding frequency improves feed efficiency associated with altered fecal microbiota and bile acid composition in pigs. Front. Microbiol. 2021 doi: 10.3389/fmicb.2021.761210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Strandwitz P, Kim KH, Terekhova D, Liu JK, Sharma A, Levering J, Lewis K. GABA-modulating bacteria of the human gut microbiota. Nat. Microbiol. 2019;4:396–403. doi: 10.1038/s41564-018-0307-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Hills RD, Jr, Pontefract BA, Mishcon HR, Black CA, Sutton SC, Theberge CR. Gut microbiome: Profound implications for diet and disease. Nutrients. 2019;11:1613. doi: 10.3390/nu11071613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zhai H, Adeola O, Liu J. Phosphorus nutrition of growing pigs. Anim. Nutr. 2022;9:127–137. doi: 10.1016/j.aninu.2022.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Liu JB, Xue PC, Cao SC, Liu J, Chen L, Zhang HF. Effects of dietary phosphorus concentration and body weight on postileal phosphorus digestion in pigs. Anim. Feed Sci. Technol. 2018;242:86–94. doi: 10.1016/j.anifeedsci.2018.06.003. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data generated in this study are included in the published article. The datasets generated during the current study are available from the corresponding author on demand upon reasonable request.