

Abstract
Triterpenoids possess potent biological activities, but their polycyclic skeletons are challenging to synthesize. The skeletal diversity of triterpenoids in plants is generated by oxidosqualene cyclases based on epoxide-triggered cationic rearrangement cascades. Normally, triterpenoid skeletons then remain unaltered during subsequent tailoring steps. In contrast, the highly modified triterpenoids found in Sapindales plants imply the existence of post-cyclization skeletal rearrangement enzymes that have not yet been found. We report here a biosynthetic pathway in Sapindales plants for the modification of already cyclized tirucallane triterpenoids, controlling the pathway bifurcation between different plant triterpenoid classes. Using a combination of bioinformatics, heterologous expression in plants and chemical analyses, we identified a cytochrome P450 monooxygenase and two isomerases which harness the epoxidation-rearrangement biosynthetic logic of triterpene cyclizations for modifying the tirucallane scaffold. The two isomerases share the same epoxide substrate made by the cytochrome P450 monooxygenase CYP88A154, but generate two different rearrangement products, one containing a cyclopropane ring. Our findings reveal a process for skeletal rearrangements of triterpenoids in nature that expands their scaffold diversity after the initial cyclization. In addition, the enzymes described here are crucial for the biotechnological production of limonoid, quassinoid, apoprotolimonoid, and glabretane triterpenoids.
Introduction
Triterpenoids are of great interest to natural product chemists, organic chemists, and medicinal chemists alike due to their complex structures and a wide array of bioactivities.1,2 The regio- and stereoselective synthesis or modification of such polycyclic and often densely functionalized molecules remains an outstanding challenge and severely hinders drug development of such compounds.3−5 As an alternative to synthesis, many organisms, particularly plants, possess elaborate biochemical machinery to produce diverse triterpenoids with high selectivity. The biosynthesis of plant triterpenoids is generally divided into two phases (Figure 1A): (1) First, the underlying carbon skeleton is generated by an oxidosqualene cyclase (OSC), based on epoxide-mediated rearrangements; (2) then, tailoring enzymes such as cytochrome P450 monooxygenases (P450s) and glycosyltransferases (GTs) introduce specific functionalizations and decorations, e.g., oxidations or glycosylations, but leave the carbon skeleton unaltered.6−11 An enzymatic way to modify the skeletons of already functionalized triterpenoids would be highly desirable to rapidly expand their chemical space. So far, however, no enzyme is known that can achieve such skeletal rearrangements of already cyclized triterpenoid scaffolds.9
Figure 1.

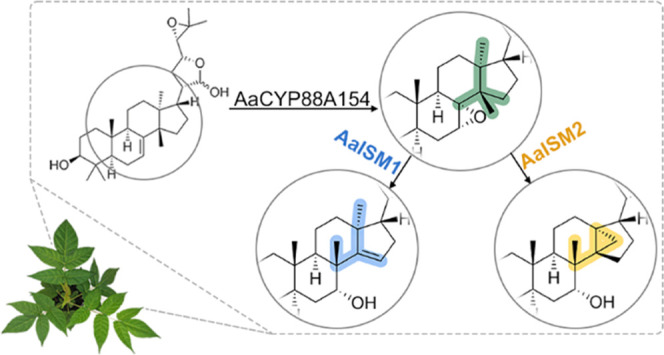
Uncharacterized skeletal rearrangements in plant triterpenoid biosynthesis. (A) Biosynthetic proposals for triterpenoids from Sapindales plants imply enzymatic modification of the already cyclized skeleton, which contrasts canonical triterpenoid biosynthesis. In comparison to the preceding protolimonoid melianol (8), the methyl group at C-14 must migrate to C-8, partially involving cyclopropane formation. The enzymes and intermediates involved in these post-cyclization skeletal rearrangements have remained unknown so far. (B and C) Globally invasive plant tree of heaven (Ailanthus altissima, family Simaroubaceae, order Sapindales) was used as a model system in this study. (B) Typical growth of A. altissima in an urban environment. (C) Eight-week-old A. altissima seedling grown in the laboratory. Abbreviations: OSC, oxidosqualene cyclase; P450: cytochrome P450 monooxygenase, GT: glycosyltransferase, GlcA: glucuronic acid.
Plants of the order Sapindales are known for their rich diversity of structurally complex triterpenoids. The two best-known groups are limonoids and quassinoids, which include many industrially and ecologically relevant members (Figure 1A).12−14 The limonoid azadirachtin (1) is a potent insecticide and key active principle of neem oil.14,15 The allelopathic quassinoid ailanthone (2) plays a crucial role for the ecological success of the globally invasive tree of heaven (A. altissima), as it occurs in root exudates and helps to outgrow surrounding plants.16−18 In addition to limonoids and quassinoids, structurally simpler triterpenoids also occur in Sapindales plants, e.g., brujavanone L (3) (belonging to apoprotolimonoids),19 and cyclopropane-containing compounds like glabretal (4) (named glabretanes herein).20 Based on increasing structural complexity, protolimonoids are considered to be precursors of other triterpenoid classes in Sapindales plants.21,22 Protolimonoid biosynthesis requires three steps starting from the common triterpenoid precursor 2,3-oxidosqualene (5).21,23 First, 2,3-oxidosqualene (5) is converted by an oxidosqualene cyclase (OSC) into tirucalla-7,24-dien-3β-ol (6), followed by multiple oxidations carried out by two cytochrome P450 monooxygenases (CYP450s), which sequentially oxidize 6 to dihydroniloticin (7) and then to the protolimonoid melianol (8), which is considered to be a key intermediate in these metabolic pathways (Figure 1A).21,23
Remarkably, the carbon skeletons of protolimonoids exhibit distinct differences from other Sapindales triterpenoid classes, namely, the positioning of a methyl group at either C-14 or C-8 and the presence or absence of a cyclopropane ring. In contrast to the canonical triterpenoid biosynthesis paradigm (Figure 1A), this suggests that there are yet unknown enzymes in Sapindales plants that modify the protolimonoid skeleton after the initial cyclization, leading to pathway bifurcation between the apoprotolimonoids/limonoids/quassinoids groups and the cyclopropane-containing glabretanes. Here, we use a combination of co-expression analysis via self-organizing maps, transient co-expression in the plant host Nicotiana benthamiana, and NMR-based structure elucidation to unravel the enzymatic steps and intermediates of this metabolic branchpoint. Three enzymes – a cytochrome P450 monooxygenase and two homologous isomerases evolved from sterol metabolism – are responsible for the skeletal rearrangements and pathway branching en route to biologically active triterpenoids of the limonoid, quassinoid, apoprotolimonoid, and glabretane classes and form the basis for future biotechnological approaches.
Results and Discussion
Gene Candidate Selection with Self-Organizing Maps
The accessibility of the tree of heaven (A. altissima) as an invasive plant makes it an ideal model system for studying the biosynthetic pathways of Sapindales triterpenoids (Figure 1B,C). Elucidation of biosynthetic pathways in plants is challenging compared to microbes, as biosynthetic genes are typically not physically clustered. Many recent examples demonstrate that co-expression analysis is a helpful tool to discover novel biosynthetic genes.24−27 During our recent discovery of the genes required for melianol (8) biosynthesis in A. altissima based on de novo transcriptome sequencing,23 we observed that these genes were highly and exclusively expressed in roots. We therefore expected that further, yet unknown genes involved in triterpenoid biosynthesis may share a similar root-specific expression profile. We therefore searched our A. altissima expression data from previous work23 for gene candidates co-expressed with the first three genes in the pathway. To facilitate visual analysis of the underlying multidimensional data set, we employed self-organizing map (SOM) analysis (Figure 2), which arranges large numbers of transcripts into clusters based on their expression profile.28 This process successfully grouped the two previously identified cytochrome P450 monooxygenase (CYP450) genes (AaCYP71CD4 and AaCYP71BQ17)23 into a single cluster with predominant average expression in roots. This cluster was of high quality, i.e., representing a homogeneous group of 695 contigs that was well separated from neighboring clusters (Figures 2 and S1). The oxidosqualene cyclase gene AaOSC2 was found in a neighboring cluster that represented transcripts with high expression in both stem bark and roots. Due to the order of biosynthetic steps, we hypothesized that further pathway genes should have an expression profile more similar to the P450 genes than to the OSC gene and therefore decided to focus on the first cluster for detailed analysis.
Figure 2.
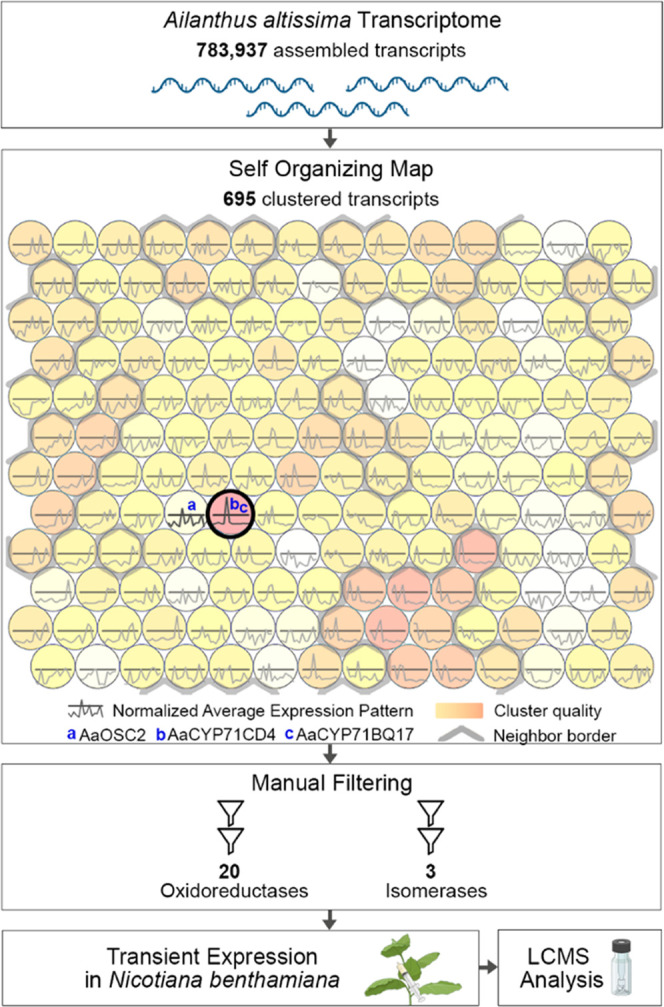
Selection of triterpenoid biosynthesis gene candidates by self-organizing map (SOM) analysis. Candidate genes co-expressed with the two pathway genes AaCYP71CD4 and AaCYP71BQ17 were obtained by self-organizing map analysis from multidimensional RNA-Seq expression data of A. altissima transcripts. Both CYP genes were grouped in the same cluster with the highest quality that contained 695 transcripts. The 695 contigs were manually filtered (Figure S2), leading to 20 oxidoreductase and 3 isomerase gene candidates, which were functionally evaluated by transient expression in N. benthamiana. See the main text and Supporting Information for further details. The figure was created with BioRender.com.
It was proposed earlier that the rearrangement of the protolimonoid skeleton could be triggered by epoxidation of the C-7,8 double bond.21,22,29 To search for the corresponding enzymes, we filtered the 695 contigs obtained by self-organizing map analysis for suitable annotations and manually excluded contigs of insufficient length (<1 kb) and low expression in the root (<50 transcripts per million) (Figure S2). This resulted in a list of 20 oxidoreductase and 3 isomerase gene candidates that were selected for functional screening.
Epoxidation of Melianol (8) by AaCYP88A154
To test if one of these 20 oxidoreductase candidates uses melianol (8) as a substrate, we co-expressed the gene candidates with the other pathway genes AaOSC2, AaCYP71CD4, and AaCYP71BQ17 as well as mevalonate pathway genes to boost levels of the precursor 2,3-oxidosqualene (5) in the plant host N. benthamiana. N. benthamiana is a popular tool for elucidating plant biosynthetic pathways, thanks to the capacity to rapidly co-express multiple gene candidates without the need for multiple selection markers.27,30,31 Eighteen of the 20 oxidase gene candidates were successfully cloned into Agrobacterium tumefaciens and co-infiltrated with AaOSC2, AaCYP71CD4, and AaCYP71BQ17 into N. benthamiana. Crude extracts of the co-expressing plants were then analyzed by LC-MS to look for consumption of melianol (8) and the production of new compounds. Gratifyingly, co-expression of one candidate gene (AaCYP88A154), encoding a cytochrome P450 monooxygenase, showed a clear decrease of melianol (8) compared to controls and other samples (Figure 3), whereas all other candidates showed no or inconsistent activity on melianol (8) or its precursors 6 or 7. Two major new peaks (compound 9 at 8.0 min, compound 10 at 6.8 min) and a minor peak at 5.5 min were observed. The new peaks showed putative molecular ions of m/z 511 ([M + Na]+). In comparison to melianol (m/z 495 for [M + Na]+), this implied incorporation of an additional oxygen atom. We suspected that one of the new products might be the previously postulated epoxide of melianol (8), while the others could be rearrangement, degradation, or shunt products. To support this hypothesis, we treated melianol (8) with meta-chloroperoxybenzoic acid (m-CPBA), a common epoxidation reagent.32 Indeed, analysis of the reaction profile showed the complete disappearance of 8 and the same three peaks as judged by retention times and mass spectra (Figure 3A). To understand the reaction course, we attempted to purify the two major products from large-scale expression in N. benthamiana. This was aggravated by the fact that all products were highly sensitive to traces of acid, e.g., from silica, formic acid, or CDCl3 (Figures S3, S4, S12). For 9, switching from CDCl3 to C6D6 as an NMR solvent proved critical to prevent degradation during measurements. We succeeded to purify 9 and 10 as mixtures of lactol epimers and fully elucidate their structures by NMR spectroscopy (Figure 3B). Compound 9 was structurally similar to melianol (8), but lacked signals for the C-7,8 double bond. Instead, two new carbon signals at 63.22 and 55.05 ppm for the major epimer suggested the presence of an epoxide at this position. Compound 10 still contained two olefinic carbons, but at C-14,15 instead of C-7,8, and in addition, a hydroxy group could be identified at C-7. By detailed two-dimensional (2D) NMR analysis, we also identified substructures of two major degradation products present in our NMR sample of 10 formed by opening of the C-24/25 epoxide in the side chain (Table S5). Both 9 and 10 are new natural products that we named 7,8-epoxymelianol and isomeliandiol, respectively. Even though isomeliandiol (10) has not been found in nature before, ca. 120 so called apoprotolimonoids with the same carbon skeleton have been isolated from Simaroubaceae, Meliaceae, and Rutaceae plants (Table S7). The presence of an oxygen atom at C-7 and the shift of the methyl group from C-14 to C-8 is also a hallmark feature of mature quassinoids and limonoids.22,33 Close homologues of AaCYP88A154 exist in the limonoid-producing plants Citrus sinensis (Cs7g22820.1, 85% AA identity) and Citrus grandis (CgUng000240.1, 87% AA identity). Hence, in support of earlier biosynthetic proposals,22,29 we conclude that the epoxidase AaCYP88A154 catalyzes a central step in the biosynthetic pathway of apoprotolimonoids, quassinoids, and limonoids, and that 7,8-epoxymelianol (9) and isomeliandiol (10) are true biosynthetic intermediates that have so far been overlooked due to their instability and rapid conversion.
Figure 3.
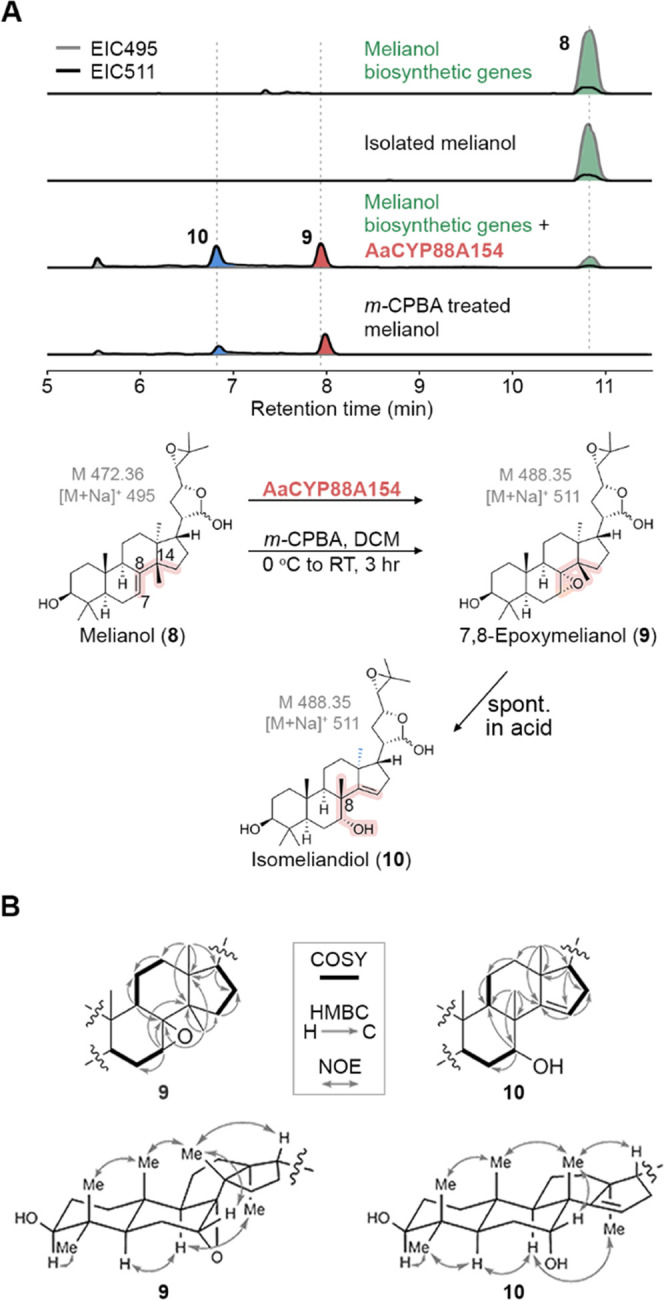
Discovery of the key intermediate 7,8-epoxymelianol (9). (A) Epoxidation of melianol by AaCYP88A154 or chemically using meta-chloroperoxybenzoic acid (m-CPBA) generates the previously unknown biosynthetic intermediates 7,8-epoxymelianol (9) and isomeliandiol (10). Melianol was generated in situ by co-expression of melianol biosynthetic genes (AaOSC2, AaCYP71CD4, and AaCYP71BQ17) and mevalonate pathway genes in N. benthamiana. (B) Key COSY, HMBC, and NOE correlations for the structure elucidation of 9 and 10.
Isomerases ISM1 and ISM2 Catalyze Skeletal Rearrangements
Given the high reactivity of 7,8-epoxymelianol (9), we speculated that additional enzymes might direct its further conversion in planta. The involvement of nonoxidative enzymes such as isomerases for this step was also proposed previously by Hodgson et al.21 We therefore next focused on the three isomerase candidates from our self-organizing map analysis (Figures 2 and S2). The isomerase gene candidates were again cloned into a transient expression vector and co-expressed with the other pathway genes AaOSC2, AaCYP71CD4, AaCYP71BQ17, and AaCYP88A154 in N. benthamiana. Strikingly, in the presence of candidate ISM1, a strong shift toward isomeliandiol (10) as the major product occurred (Figure 4A). To our bigger surprise, the presence of candidate ISM2 also led to almost complete disappearance of epoxide 9, but a new product peak (compound 11) at 6.6 min was observed. The last candidate did not show any changes to the metabolic profile compared to controls. Like for 9 and 10, the mass spectrum of 11 showed a molecular ion of m/z 511, suggesting it to be a previously not observed isomer. We isolated 11 from large-scale co-expression in N. benthamiana. Strikingly, NMR analysis of 11 clearly showed the presence of a cyclopropane, as judged by a CH2 group with unusual high field shifts (δC = 13.9 ppm, δH = 0.65/0.45 ppm) (Figure 4B).34 Full structure elucidation of 11 (Figure 4C) indicated a novel natural product structurally related to glabretal, a triterpenoid previously isolated from Guarea glabra (Meliaceae),20 one of ca. 110 natural products with the same carbon skeleton found in the families Meliaceae, Rutaceae, and Simaroubaceae for which we suggest the name glabretanes (Table S8). Hence, we named 11 protoglabretal. AaCYP88A154, ISM1, and ISM2 were also heterologously produced in baker’s yeast (Saccharomyces cerevisiae) and used for in vitro assays with microsomes, which showed the same enzymatic activity observed in N. benthamiana (Figure S30A/B). ISM2 accepted only 7,8-epoxymelianol (9) as a substrate, but not isomeliandiol (10) (Figure S30C). Taken together, our findings show that the two isomerases ISM1 and ISM2 control the skeletal rearrangement cascade of 7,8-epoxymelianol (9), leading to two different triterpenoid skeletons. While ISM1 merely channels the spontaneous reaction toward isomeliandiol (10), ISM2 generates a product that is not observed when the rearrangement occurs spontaneously. Even though both ISM1 and ISM2 genes have highly similar expression profiles (Figure S1B), co-expression of both genes in N. benthamiana demonstrated that isomeliandiol (10) and protoglabretal (11) can be formed in parallel in planta (Figure 4A). We therefore conclude that ISM1 and ISM2 are central gatekeepers in plant triterpenoid metabolism, controlling the formation of the apoprotolimonoid and glabretane structural subclasses.
Figure 4.

Homologous isomerases AaISM1 and AaISM2 channel the rearrangement of 7,8-epoxymelianol (9) toward isomeliandiol (10) and protoglabretal (11), respectively. (A) LCMS profiles of AaISM1 and AaISM2 genes co-expressed with genes for the biosynthesis of 7,8-epoxymelianol (9) in N. benthamiana. The product distribution based on relative peak areas of compounds 9, 10, 11 and the minor side product at 5.5 min compared to their total peak area is additionally shown as a bar plot for three biological replicates each. (B) Structures of the new natural products isomeliandiol (10) and protoglabretal (11) together with chemical shifts at C-18 (highlighted). (C) Key COSY, HMBC, and NOE correlations for 11. (D) Maximum-likelihood phylogenetic tree of sterol isomerases. The scale bar indicates the phylogenetic distance. Bootstrap values for 1000 replicates are shown. Homologues of AaISM1 and AaISM2 are found in Meliaceae (Toona sinensis (IDs start with Maker)) and Rutaceae (C. sinensis; C. grandis) plants, but not in plants from other Sapindales families (Acer yangbiense, Sapindaceae; Mangifera indica, Anacardiaceae), matching the distribution of quassinoids, limonoids, apoprotolimonoids, and glabretanes in these families. For further details, see main text and Supporting Information.
ISM1 and ISM2 Diverged from General Sterol Biogenesis
The amino acid identity between ISM1 and ISM2 is 50%. Both are related to C-8,7 sterol isomerases (8,7SI) from primary metabolism, which catalyze the key isomerization of the Δ8 double bond to Δ7 in sterol biosynthesis in all eukaryotes (Figure S31).35−39 The most well-known C-8,7 sterol isomerase from plants is At8,7SI (encoded by HYDRA1) from Arabidopsis thaliana.39,40 The amino acid identity of At8,7SI compared to ISM1 and ISM2 is 45 and 53%, respectively. We generated a phylogenetic tree with other C-8,7 sterol isomerases from primary metabolism as well as putative homologues of ISM1 and ISM2 found in publicly available genome data of the limonoid-producing plants C. sinensis (sweet orange), C. grandis (pomelo),41,42 and T. sinensis.43 This analysis suggested that homologues of AaISM1 and AaISM2 are conserved in Rutaceae and Meliaceae but not in other plants, matching the occurrence of quassinoid, limonoid, apoprotolimonoid, and glabretane triterpenoids and thus supporting the proposed key roles of ISM1 and ISM2 for triterpenoid metabolism (Figure 4D). Our phylogenetic analysis suggests that both ISM1 and ISM2 evolved by duplication of a C-8,7 sterol isomerase gene from primary metabolism. Such gene duplication and neofunctionalization events are known as key drivers for the evolution of plant specialized metabolism.44−46
Only very few other examples of isomerases in plant specialized metabolism are known, and none performs a skeletal rearrangement or affects more than three adjacent atoms (Figure S32). Most of these examples play a role in nonterpenoid metabolic pathways, namely, chalcone isomerase in flavonoid metabolism,47,48 the BAHD acyltransferase COSY in coumarin biosynthesis,49 and neopinone isomerase in opiate production.50 Two isomerases are known from plant terpenoid metabolism but only catalyze double bond shifts: In withanolide biosynthesis, an isomerase evolved from a reductase performs Δ24(28) to a Δ24(25) double bond isomerization.51 A similar double bond shift was reported for Δ5-3-ketosteroid isomerase in the context of cardenolide biosynthesis.52 In contrast to these known isomerases, ISM1 and ISM2 modify the underlying scaffolds of their substrates. Mechanistically, we propose that the preceding epoxidation enables a cationic rearrangement cascade that involves multiple carbon atoms in spatial proximity, typical for terpenoid cyclizations and rearrangements (Figure 5).53−55 While several ways for enzymatic cyclopropanation are already known,56−59 ISM2 represents a new way how a cyclopropane can be installed onto an existing triterpenoid skeleton. The fact that the two related enzymes ISM1 and ISM2 generate different rearrangement products from the same epoxide substrate also suggests that protein engineering is highly promising to harness these isomerases for even further skeletal modifications in the future. Recent data additionally supports the role of these enzymes in limonoid biosynthesis.60
Figure 5.

Proposed mechanism for post-cyclization skeletal rearrangements of melianol (8) by a cationic rearrangement cascade. Triggered by C-7,8 epoxidation via the cytochrome P450 monooxygenase AaCYP88A154, the two isomerases AaISM1 and AaISM2 generate the previously unknown triterpenoids isomeliandiol (10) and protoglabretal (11) by directing the fate of carbon cation intermediates. Thereby, ISM1 and ISM2 control the branching between the apoprotolimonoid/limonoid/quassinoid and glabretane classes of triterpenoids, respectively.
Conclusions
Taken together, we discovered three novel enzymes in plant triterpenoid metabolism, a cytochrome P450 and two isomerases, that perform skeletal rearrangements of triterpenoids in Sapindales plants at a metabolic branchpoint. The two isomerases AaISM1 and AaISM2 share the same substrate, 7,8-epoxymelianol (9), formed by the cytochrome P450 monooxygenase AaCYP88A154, but generate two different rearrangement products isomeliandiol (10) and protoglabretal (11), representing different classes of triterpenoids. Traditionally, the skeletal diversity of triterpenoids was considered to be solely derived from the initial cyclization. Our discovery here now shows how triterpenoid skeletal diversity can be expanded after the cyclization phase in nature by exploiting the same epoxide-rearrangement biosynthetic logic that is also employed for the initial cyclization. Our findings pave the way for developing new strategies to modify already cyclized triterpenoid skeletons, which would greatly facilitate the speed by which already functionalized triterpenoids can be generated for medicinal chemistry and other applications. Last, the biosynthetic genes described herein will also be crucial to develop biotechnological tools for the production of industrially relevant triterpenoids like the insecticidal limonoid azadirachtin (1) and many other triterpenoids belonging to the apoprotolimonoid, limonoid, quassinoid, or glabretane classes.
Acknowledgments
The authors thank Prof. Dr. David Nelson (Department of Molecular Science, University of Tennessee, Memphis, USA) and the P450 nomenclature committee for naming AaCYP88A154, Yvonne Leye and Miriam Fent for excellent horticultural support, Katja Körner for excellent technical support, and Johanna Wolf and Yue Sun for assistance with the transient expression screening. The authors thank Prof. Dr. Christian Hertweck (Leibniz Institute for Natural Product Research and Infection Biology, HKI, Jena, Germany) and Prof. Dr. Sarah O’Connor (Max Planck Institute for Chemical Ecology, Jena, Germany) for helpful discussions.
Supporting Information Available
The Supporting Information is available free of charge at https://pubs.acs.org/doi/10.1021/jacs.2c10838.
Experimental details, materials, primer sequences, NMR data and spectra for all isolated compounds (PDF)
Author Contributions
§ L.C. and S.L. contributed equally to this work. The manuscript was written through contributions of all authors. All authors have given approval to the final version of the manuscript.
The authors gratefully acknowledge financial support by the Fonds der Chemischen Industrie, the Emmy Noether program of the Deutsche Forschungsgemeinschaft (FR 3720/3-1), and the SMART BIOTECS alliance between the Technische Universität Braunschweig and the Leibniz Universität Hannover, supported by the Ministry of Science and Culture (MWK) of Lower Saxony. The authors also thank the DFG for the provision of NMR equipment (INST 187/686-1). In addition, this work was supported by the LUH compute cluster, which is funded by the Leibniz Universität Hannover, the Lower Saxony Ministry of Science and Culture (MWK), and the German Research Association (DFG).
The authors declare no competing financial interest.
Supplementary Material
References
- Hill R. A.; Connolly J. D. Triterpenoids. Nat. Prod. Rep. 2020, 37, 962–998. 10.1039/C9NP00067D. [DOI] [PubMed] [Google Scholar]
- Zhou M.; Zhang R.-H.; Wang M.; Xu G.-B.; Liao S.-G. Prodrugs of Triterpenoids and Their Derivatives. Eur. J. Med. Chem. 2017, 131, 222–236. 10.1016/j.ejmech.2017.03.005. [DOI] [PubMed] [Google Scholar]
- Yamashita S.; Naruko A.; Nakazawa Y.; Zhao L.; Hayashi Y.; Hirama M. Total Synthesis of Limonin. Angew. Chem., Int. Ed. 2015, 54, 8538–8541. 10.1002/anie.201503794. [DOI] [PubMed] [Google Scholar]
- Furiassi L.; Tonogai E. J.; Hergenrother P. J. Limonin as a Starting Point for the Construction of Compounds with High Scaffold Diversity. Angew. Chem., Int. Ed. 2021, 60, 16119–16128. 10.1002/anie.202104228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pazur E. J.; Wipf P. Recent Syntheses and Biological Profiling of Quassinoids. Org. Biomol. Chem. 2022, 20, 3870–3889. 10.1039/D2OB00490A. [DOI] [PubMed] [Google Scholar]
- Thimmappa R.; Geisler K.; Louveau T.; O’Maille P.; Osbourn A. Triterpene Biosynthesis in Plants. Annu. Rev. Plant Biol. 2014, 65, 225–257. 10.1146/annurev-arplant-050312-120229. [DOI] [PubMed] [Google Scholar]
- Ghosh S. Triterpene Structural Diversification by Plant Cytochrome P450 Enzymes. Front. Plant Sci. 2017, 8, 1886 10.3389/fpls.2017.01886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawai S.; Saito K. Triterpenoid Biosynthesis and Engineering in Plants. Front. Plant Sci. 2011, 2, 25 10.3389/fpls.2011.00025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almeida A.; Dong L.; Appendino G.; Bak S. Plant Triterpenoids with Bond-Missing Skeletons: Biogenesis, Distribution and Bioactivity. Nat. Prod. Rep. 2020, 37, 1207–1228. 10.1039/C9NP00030E. [DOI] [PubMed] [Google Scholar]
- Hu D.; Gao H.; Yao X.. 1.18 - Biosynthesis of Triterpenoid Natural Products. In Comprehensive Natural Products III; Liu H.-W. Ben.; Begley T. P., Eds.; Elsevier: Oxford, 2020; pp 577–612 10.1016/B978-0-12-409547-2.14678-5. [DOI] [Google Scholar]
- Cárdenas P. D.; Almeida A.; Bak S. Evolution of Structural Diversity of Triterpenoids. Front. Plant Sci. 2019, 10, 1523 10.3389/fpls.2019.01523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Houël E.; Stien D.; Bourdy G.; Deharo E.. Quassinoids: Anticancer and Antimalarial Activities. In Natural Products; Ramawat K. G.; Mérillon J.-M., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2013; pp 3775–3802 10.1007/978-3-642-22144-6_161. [DOI] [Google Scholar]
- Fiaschetti G.; A Grotzer M.; Shalaby T.; Castelletti D.; Arcaro A. Quassinoids: From Traditional Drugs to New Cancer Therapeutics. Curr. Med. Chem. 2011, 18, 316–328. 10.2174/092986711794839205. [DOI] [PubMed] [Google Scholar]
- Tan Q.-G.; Luo X.-D. Meliaceous Limonoids: Chemistry and Biological Activities. Chem. Rev. 2011, 111, 7437–7522. 10.1021/cr9004023. [DOI] [PubMed] [Google Scholar]
- Fan S.; Zhang C.; Luo T.; Wang J.; Tang Y.; Chen Z.; Yu L. Limonin: A Review of Its Pharmacology, Toxicity, and Pharmacokinetics. Molecules 2019, 24, 3679. 10.3390/molecules24203679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heisey R. M. Identification of an Allelopathic Compound from Ailanthus Altissima (Simaroubaceae) and Characterization of Its Herbicidal Activity. Am. J. Bot. 1996, 83, 192–200. 10.1002/j.1537-2197.1996.tb12697.x. [DOI] [Google Scholar]
- Alves I. A. B. S.; Miranda H. M.; Soares L. A. L.; Randau K. P. Simaroubaceae Family: Botany, Chemical Composition and Biological Activities. Rev. Bras. Farmacogn. 2014, 24, 481–501. 10.1016/j.bjp.2014.07.021. [DOI] [Google Scholar]
- Sladonja B.; Sušek M.; Guillermic J. Review on Invasive Tree of Heaven (Ailanthus Altissima (Mill.) Swingle) Conflicting Values: Assessment of Its Ecosystem Services and Potential Biological Threat. Environ. Manage. 2015, 56, 1009–1034. 10.1007/s00267-015-0546-5. [DOI] [PubMed] [Google Scholar]
- Dong S.-H.; Liu J.; Ge Y.-Z.; Dong L.; Xu C.-H.; Ding J.; Yue J.-M. Chemical Constituents from Brucea Javanica. Phytochemistry 2013, 85, 175–184. 10.1016/j.phytochem.2012.08.018. [DOI] [PubMed] [Google Scholar]
- Ferguson G.; Gunn P. A.; Marsh W. C.; McCrindle R.; Restivo R.; Connolly J. D.; Fulke J. W. B.; Henderson M. S. Triterpenoids from Guarea Glabra (Meliaceae): A New Skeletal Class Identified by Chemical, Spectroscopic, and X-Ray Evidence. J. Chem. Soc. Chem. Commun. 1973, 159–160. 10.1039/C39730000159. [DOI] [Google Scholar]
- Hodgson H.; Peña R. D. L.; Stephenson M. J.; Thimmappa R.; Vincent J. L.; Sattely E. S.; Osbourn A. Identification of Key Enzymes Responsible for Protolimonoid Biosynthesis in Plants: Opening the Door to Azadirachtin Production. Proc. Natl. Acad. Sci. U.S.A. 2019, 116, 17096–17104. 10.1073/pnas.1906083116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das M. F.; Da Silva G. F.; Gottlieb O. R. Evolution of Quassinoids and Limonoids in the Rutales. Biochem. Syst. Ecol. 1987, 15, 85–103. 10.1016/0305-1978(87)90086-X. [DOI] [Google Scholar]
- Chuang L.; Liu S.; Biedermann D.; Franke J. Identification of Early Quassinoid Biosynthesis in the Invasive Tree of Heaven (Ailanthus Altissima) Confirms Evolutionary Origin from Protolimonoids. Front. Plant Sci. 2022, 13, 958138 10.3389/fpls.2022.958138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caputi L.; Franke J.; Farrow S. C.; Chung K.; Payne R. M. E.; Nguyen T.-D.; Dang T.-T. T.; Carqueijeiro I. S. T.; Koudounas K.; Bernonville T. D.; de Ameyaw B.; Jones D. M.; Vieira I. J. C.; Courdavault V.; O’Connor S. E. Missing Enzymes in the Biosynthesis of the Anticancer Drug Vinblastine in Madagascar Periwinkle. Science 2018, 360, 1235–1239. 10.1126/science.aat4100. [DOI] [PubMed] [Google Scholar]
- Dang T.-T. T.; Franke J.; Carqueijeiro I. S. T.; Langley C.; Courdavault V.; O’Connor S. E. Sarpagan Bridge Enzyme Has Substrate-Controlled Cyclization and Aromatization Modes. Nat. Chem. Biol. 2018, 14, 760–763. 10.1038/s41589-018-0078-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nett R. S.; Dho Y.; Low Y.-Y.; Sattely E. S. A Metabolic Regulon Reveals Early and Late Acting Enzymes in Neuroactive Lycopodium Alkaloid Biosynthesis. Proc. Natl. Acad. Sci. U.S.A. 2021, 118, e2102949118 10.1073/pnas.2102949118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobowitz J. R.; Weng J.-K. Exploring Uncharted Territories of Plant Specialized Metabolism in the Postgenomic Era. Annu. Rev. Plant Biol. 2020, 71, 631–658. 10.1146/annurev-arplant-081519-035634. [DOI] [PubMed] [Google Scholar]
- Payne R. M. E.; Xu D.; Foureau E.; Teto Carqueijeiro M. I. S.; Oudin A.; de Bernonville T. D.; Novak V.; Burow M.; Olsen C.-E.; Jones D. M.; Tatsis E. C.; Pendle A.; Halkier B. A.; Geu-Flores F.; Courdavault V.; Nour-Eldin H. H.; O’Connor S. E. An NPF Transporter Exports a Central Monoterpene Indole Alkaloid Intermediate from the Vacuole. Nat. Plants 2017, 3, 16208 10.1038/nplants.2016.208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polonsky J.Quassinoid Bitter Principles. In Fortschritte Der Chemie Organischer Naturstoffe: Progress in the Chemistry of Organic Natural Products; Cormier M. J.; Flasch H.; Franck B.; Hori K.; Jaenicke L.; Keller-Schierlein W.; Locksley H. D.; Müller D. G.; Polonsky J.; Tschesche R.; Wampler J. E.; Wulff G.; Grisebach H.; Kirby G. W.; Herz W., Eds.; Fortschritte der Chemie organischer Naturstoffe Progress in the Chemistry of Organic Natural Products; Springer: Vienna, 1973; pp 101–150 10.1007/978-3-7091-7102-8_3. [DOI] [PubMed] [Google Scholar]
- Chuang L.; Franke J.. Rapid Combinatorial Coexpression of Biosynthetic Genes by Transient Expression in the Plant Host Nicotiana Benthamiana. In Engineering Natural Product Biosynthesis: Methods and Protocols; Skellam E., Ed.; Methods in Molecular Biology; Springer US: New York, NY, 2022; pp 395–420 10.1007/978-1-0716-2273-5_20. [DOI] [PubMed] [Google Scholar]
- Bally J.; Jung H.; Mortimer C.; Naim F.; Philips J. G.; Hellens R.; Bombarely A.; Goodin M. M.; Waterhouse P. M. The Rise and Rise of Nicotiana Benthamiana: A Plant for All Reasons. Annu. Rev. Phytopathol. 2018, 56, 405–426. 10.1146/annurev-phyto-080417-050141. [DOI] [PubMed] [Google Scholar]
- Hussain H.; Al-Harrasi A.; R Green I.; Ahmed I.; Abbas G.; Ur Rehman N. Meta -Chloroperbenzoic Acid (m CPBA): A Versatile Reagent in Organic Synthesis. RSC Adv. 2014, 4, 12882–12917. 10.1039/C3RA45702H. [DOI] [Google Scholar]
- Seigler D. S.Limonoids, Quassinoids, and Related Compounds. In Plant Secondary Metabolism; Springer: Boston, MA, 1998; pp 473–485 10.1007/978-1-4615-4913-0_25. [DOI] [Google Scholar]
- Trottmann F.; Franke J.; Richter I.; Ishida K.; Cyrulies M.; Dahse H.-M.; Regestein L.; Hertweck C. Cyclopropanol Warhead in Malleicyprol Confers Virulence of Human- and Animal-Pathogenic Burkholderia Species. Angew. Chem., Int. Ed. 2019, 58, 14129–14133. 10.1002/anie.201907324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Long T.; Hassan A.; Thompson B. M.; McDonald J. G.; Wang J.; Li X. Structural Basis for Human Sterol Isomerase in Cholesterol Biosynthesis and Multidrug Recognition. Nat. Commun. 2019, 10, 2452 10.1038/s41467-019-10279-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benveniste P. Biosynthesis and Accumulation of Sterols. Annu. Rev. Plant Biol. 2004, 55, 429–457. 10.1146/annurev.arplant.55.031903.141616. [DOI] [PubMed] [Google Scholar]
- Desmond E.; Gribaldo S. Phylogenomics of Sterol Synthesis: Insights into the Origin, Evolution, and Diversity of a Key Eukaryotic Feature. Genome Biol. Evol. 2009, 1, 364–381. 10.1093/gbe/evp036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang T.; Yuan D.; Xie J.; Lei Y.; Li J.; Fang G.; Tian L.; Liu J.; Cui Y.; Zhang M.; Xiao Y.; Xu Y.; Zhang J.; Zhu M.; Zhan S.; Li S. Evolution of the Cholesterol Biosynthesis Pathway in Animals. Mol. Biol. Evol. 2019, 36, 2548–2556. 10.1093/molbev/msz167. [DOI] [PubMed] [Google Scholar]
- Grebenok R. J.; Ohnmeiss T. E.; Yamamoto A.; Huntley E. D.; Galbraith D. W.; Della Penna D. Isolation and Characterization of an Arabidopsis Thaliana C-8,7 Sterol Isomerase: Functional and Structural Similarities to Mammalian C-8,7 Sterol Isomerase/Emopamil-Binding Protein. Plant Mol. Biol. 1998, 38, 807–815. 10.1023/A:1006028623875. [DOI] [PubMed] [Google Scholar]
- Souter M.; Topping J.; Pullen M.; Friml J.; Palme K.; Hackett R.; Grierson D.; Lindsey K. Hydra Mutants of Arabidopsis Are Defective in Sterol Profiles and Auxin and Ethylene Signaling. Plant Cell 2002, 14, 1017–1031. 10.1105/tpc.001248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu Q.; Chen L.-L.; Ruan X.; Chen D.; Zhu A.; Chen C.; Bertrand D.; Jiao W.-B.; Hao B.-H.; Lyon M. P.; Chen J.; Gao S.; Xing F.; Lan H.; Chang J.-W.; Ge X.; Lei Y.; Hu Q.; Miao Y.; Wang L.; Xiao S.; Biswas M. K.; Zeng W.; Guo F.; Cao H.; Yang X.; Xu X.-W.; Cheng Y.-J.; Xu J.; Liu J.-H.; Luo O. J.; Tang Z.; Guo W.-W.; Kuang H.; Zhang H.-Y.; Roose M. L.; Nagarajan N.; Deng X.-X.; Ruan Y. The Draft Genome of Sweet Orange (Citrus Sinensis). Nat. Genet. 2013, 45, 59–66. 10.1038/ng.2472. [DOI] [PubMed] [Google Scholar]
- Wu G. A.; Prochnik S.; Jenkins J.; Salse J.; Hellsten U.; Murat F.; Perrier X.; Ruiz M.; Scalabrin S.; Terol J.; Takita M. A.; Labadie K.; Poulain J.; Couloux A.; Jabbari K.; Cattonaro F.; Del Fabbro C.; Pinosio S.; Zuccolo A.; Chapman J.; Grimwood J.; Tadeo F. R.; Estornell L. H.; Muñoz-Sanz J. V.; Ibanez V.; Herrero-Ortega A.; Aleza P.; Pérez-Pérez J.; Ramón D.; Brunel D.; Luro F.; Chen C.; Farmerie W. G.; Desany B.; Kodira C.; Mohiuddin M.; Harkins T.; Fredrikson K.; Burns P.; Lomsadze A.; Borodovsky M.; Reforgiato G.; Freitas-Astúa J.; Quetier F.; Navarro L.; Roose M.; Wincker P.; Schmutz J.; Morgante M.; Machado M. A.; Talon M.; Jaillon O.; Ollitrault P.; Gmitter F.; Rokhsar D. Sequencing of Diverse Mandarin, Pummelo and Orange Genomes Reveals Complex History of Admixture during Citrus Domestication. Nat. Biotechnol. 2014, 32, 656–662. 10.1038/nbt.2906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ji Y.-T.; Xiu Z.; Chen C.-H.; Wang Y.; Yang J.-X.; Sui J.-J.; Jiang S.-J.; Wang P.; Yue S.-Y.; Zhang Q.-Q.; Jin J.; Wang G.-S.; Wei Q.-Q.; Wei B.; Wang J.; Zhang H.-L.; Zhang Q.-Y.; Liu J.; Liu C.-J.; Jian J.-B.; Qu C.-Q. Long Read Sequencing of Toona Sinensis (A. Juss) Roem: A Chromosome-Level Reference Genome for the Family Meliaceae. Mol. Ecol. Resour. 2021, 21, 1243–1255. 10.1111/1755-0998.13318. [DOI] [PubMed] [Google Scholar]
- Sonawane P. D.; Jozwiak A.; Panda S.; Aharoni A. ‘Hijacking’ Core Metabolism: A New Panache for the Evolution of Steroidal Glycoalkaloids Structural Diversity. Curr. Opin. Plant Biol. 2020, 55, 118–128. 10.1016/j.pbi.2020.03.008. [DOI] [PubMed] [Google Scholar]
- Jozwiak A.; Sonawane P. D.; Panda S.; Garagounis C.; Papadopoulou K. K.; Abebie B.; Massalha H.; Almekias-Siegl E.; Scherf T.; Aharoni A. Plant Terpenoid Metabolism Co-Opts a Component of the Cell Wall Biosynthesis Machinery. Nat. Chem. Biol. 2020, 16, 740–748. 10.1038/s41589-020-0541-x. [DOI] [PubMed] [Google Scholar]
- Tohge T.; Fernie A. R. Co-Regulation of Clustered and Neo-Functionalized Genes in Plant-Specialized Metabolism. Plants 2020, 9, 622. 10.3390/plants9050622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin Y.-c.; Zhang X.; Gao Z.; Hu T.; Liu Y. The Research Progress of Chalcone Isomerase (CHI) in Plants. Mol. Biotechnol. 2019, 61, 32–52. 10.1007/s12033-018-0130-3. [DOI] [PubMed] [Google Scholar]
- Jez J. M.; Bowman M. E.; Dixon R. A.; Noel J. P. Structure and Mechanism of the Evolutionarily Unique Plant Enzyme Chalcone Isomerase. Nat. Struct. Biol. 2000, 7, 786–791. 10.1038/79025. [DOI] [PubMed] [Google Scholar]
- Vanholme R.; Sundin L.; Seetso K. C.; Kim H.; Liu X.; Li J.; Meester B. D.; Hoengenaert L.; Goeminne G.; Morreel K.; Haustraete J.; Tsai H.-H.; Schmidt W.; Vanholme B.; Ralph J.; Boerjan W. COSY Catalyses Trans – Cis Isomerization and Lactonization in the Biosynthesis of Coumarins. Nat. Plants 2019, 5, 1066–1075. 10.1038/s41477-019-0510-0. [DOI] [PubMed] [Google Scholar]
- Dastmalchi M.; Chen X.; Hagel J. M.; Chang L.; Chen R.; Ramasamy S.; Yeaman S.; Facchini P. J. Neopinone Isomerase Is Involved in Codeine and Morphine Biosynthesis in Opium Poppy. Nat. Chem. Biol. 2019, 15, 384–390. 10.1038/s41589-019-0247-0. [DOI] [PubMed] [Google Scholar]
- Knoch E.; Sugawara S.; Mori T.; Poulsen C.; Fukushima A.; Harholt J.; Fujimoto Y.; Umemoto N.; Saito K. Third DWF1 Paralog in Solanaceae, Sterol Δ24-Isomerase, Branches Withanolide Biosynthesis from the General Phytosterol Pathway. Proc. Natl. Acad. Sci. U.S.A. 2018, 115, E8096–E8103. 10.1073/pnas.1807482115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meitinger N.; Geiger D.; Augusto T. W.; Maia de Pádua R.; Kreis W. Purification of Δ5-3-Ketosteroid Isomerase from Digitalis Lanata. Phytochemistry 2015, 109, 6–13. 10.1016/j.phytochem.2014.10.025. [DOI] [PubMed] [Google Scholar]
- Christianson D. W. Structural and Chemical Biology of Terpenoid Cyclases. Chem. Rev. 2017, 117, 11570–11648. 10.1021/acs.chemrev.7b00287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickschat J. S. Bacterial Diterpene Biosynthesis. Angew. Chem., Int. Ed. 2019, 58, 15964–15976. 10.1002/anie.201905312. [DOI] [PubMed] [Google Scholar]
- Dickschat J. S. Bacterial Terpene Cyclases. Nat. Prod. Rep. 2016, 33, 87–110. 10.1039/C5NP00102A. [DOI] [PubMed] [Google Scholar]
- Ma S.; Mandalapu D.; Wang S.; Zhang Q. Biosynthesis of Cyclopropane in Natural Products. Nat. Prod. Rep. 2022, 39, 926–945. 10.1039/D1NP00065A. [DOI] [PubMed] [Google Scholar]
- Thibodeaux C. J.; Chang W.; Liu H. Enzymatic Chemistry of Cyclopropane, Epoxide, and Aziridine Biosynthesis. Chem. Rev. 2012, 112, 1681–1709. 10.1021/cr200073d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wessjohann L. A.; Brandt W.; Thiemann T. Biosynthesis and Metabolism of Cyclopropane Rings in Natural Compounds. Chem. Rev. 2003, 103, 1625–1648. 10.1021/cr0100188. [DOI] [PubMed] [Google Scholar]
- Trottmann F.; Ishida K.; Ishida-Ito M.; Kries H.; Groll M.; Hertweck C. Pathogenic Bacteria Remodel Central Metabolic Enzyme to Build a Cyclopropanol Warhead. Nat. Chem. 2022, 14, 884–890. 10.1038/s41557-022-01005-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De La Peña R.; Hodgson H.; Liu J. C.-T.; Stephenson M. J.; Martin A. C.; Owen C.; Harkess A.; Leebens-Mack J.; Jimenez L. E.; Osbourn A.; Sattely E. S. Complex Scaffold Remodeling in Plant Triterpene Biosynthesis. Science 2023, 379, 361–368. 10.1126/science.adf1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.