

Abstract
Background
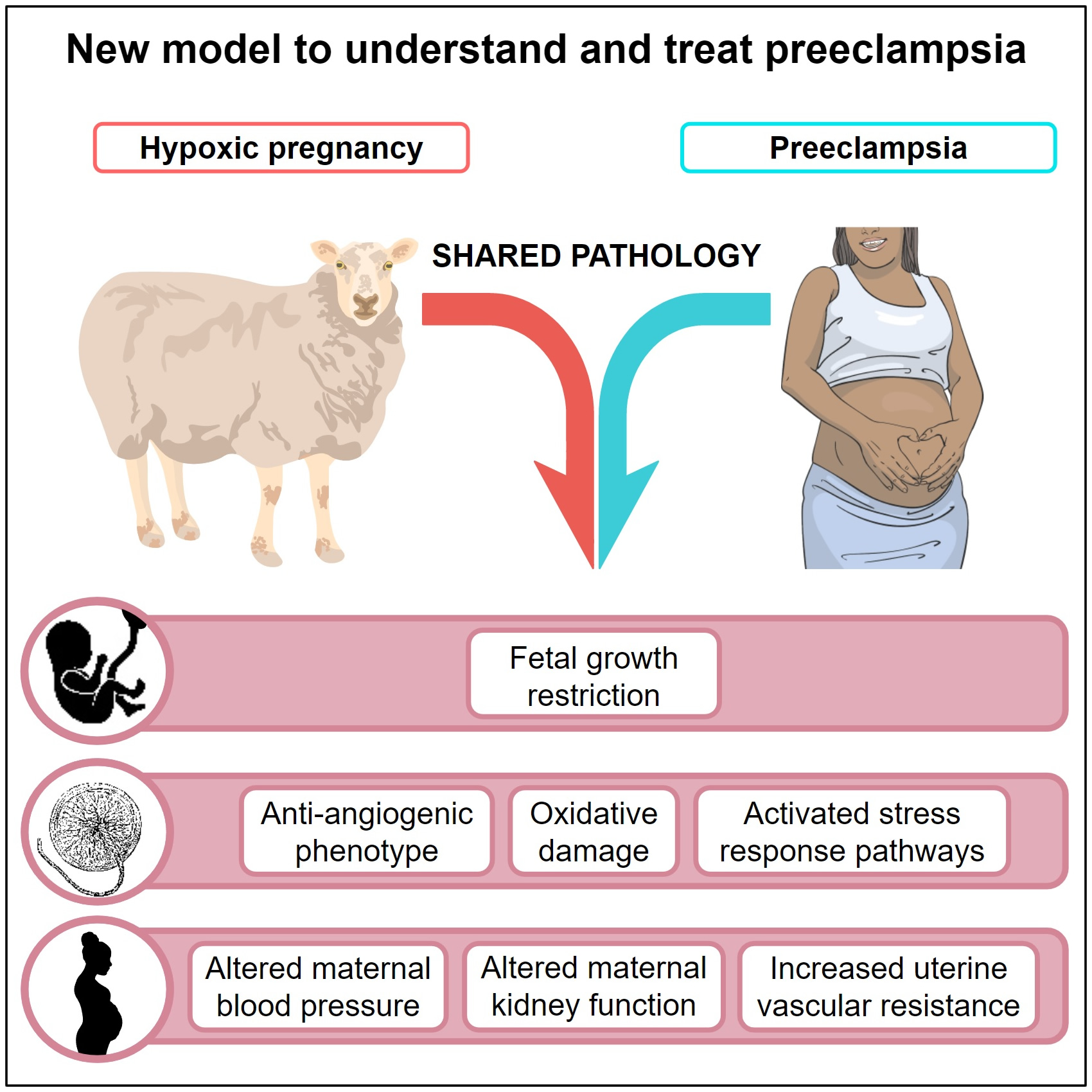
Preeclampsia continues to be a prevalent pregnancy complication and underlying mechanisms remain controversial. A common feature of preeclampsia is utero-placenta hypoxia. In contrast to the impact of hypoxia on the placenta and fetus, comparatively little is known on the maternal physiology.
Methods
We adopted an integrative approach to investigate the inter-relationship between chronic hypoxia during pregnancy with maternal, placental and fetal outcomes, common in preeclampsia. We exploited a novel technique using isobaric hypoxic chambers and in vivo continuous cardiovascular recording technology for measurement of blood pressure in sheep and studied the placental stress in response to hypoxia at cellular and sub-cellular levels.
Results
Chronic hypoxia in ovine pregnancy promoted fetal growth restriction with evidence of fetal brain-sparing, increased placental hypoxia-mediated oxidative damage and activated placental stress response pathways. These changes were linked with dilation of the placental endoplasmic reticulum cisternae and increased placental expression of the antiangiogenic factors sFlt-1 and sEng, combined with a shift towards an angiogenic imbalance in the maternal circulation. Chronic hypoxia further led to an increase in uteroplacental vascular resistance, and the fall in maternal blood pressure with advancing gestation measured in normoxic pregnancy did not occur in hypoxic pregnancy.
Conclusions
Therefore, we show in an ovine model of sea level adverse pregnancy that chronic hypoxia recapitulates physiological and molecular features of preeclampsia in the mother, placenta and offspring.
Keywords: chronic hypoxia, fetal growth restriction, unfolded protein response, placental dysfunction, angiogenic imbalance, oxidative stress
Non-standard abbreviations & acronyms
- ATF6
activating transcription factor 6
- dGA
days gestational age
- ER
endoplasmic reticulum
- ERK
extracellular signal-regulated kinase
- FGR
fetal growth restriction
- GRP78
glucose-related protein 78
- HIF1α
hypoxia-inducible factor 1α
- HSP27
heat shock protein 27
- HSP70
heat shock protein 70
- JNK
C-Jun N-terminal kinase
- PaO2
arterial partial pressure of oxygen
- PDI
protein disulfide isomerase
- PI
pulsatility index
- PlGF
placental growth factor
- sEng
soluble endoglin
- sFlt-1
soluble fms-like tyrosine kinase 1
- UPR
unfolded protein response
- UPRCyt
cytosolic unfolded protein response
- UPRER
endoplasmic reticulum unfolded protein response
- VEGF
vascular endothelial growth factor
Background
Preeclampsia remains a leading cause of perinatal morbidity and mortality, affecting 2-8% of pregnancies worldwide1. Therefore, there is ongoing interest in improving our understanding of the disease. Historically, preeclampsia was thought to develop exclusively in early pregnancy due to failure of spiral artery conversion and reduced uteroplacental perfusion2. However, knowledge has expanded and it is now accepted that preeclampsia encompasses a broader spectrum of disorders, including early- and late-onset preeclampsia3.
Irrespective of aetiology, all forms of preeclampsia resolve after delivery of the placenta, confirming central involvement of the organ. In addition, most forms of preeclampsia present with evidence of impaired uteroplacental perfusion and placental hypoxia2. However, whether placental hypoxia is merely a consequence of the disease or whether it causes the adverse maternal and fetal outcomes is uncertain. In contrast to the impact of hypoxia on the placenta and fetus, comparatively little is known on the maternal physiology. When investigating interactions among mother, placenta and offspring, maternal metabolism, the temporal profile of fetal development, and access to longitudinal physiological measures are three important considerations. Sheep and humans share a similar precocial profile of organ development, and sheep give birth primarily to singleton or twin lambs of similar weight to humans after a relatively long gestation period4. Therefore, the maternal and placental metabolic investment in pregnancy is similar between sheep and humans. In addition, sheep permit longitudinal assessment of uterine blood flow via surgically implanted flow probes as well as serial long-term blood sampling for endocrinology5. In this study, we have tested the hypothesis that placental hypoxia drives phenotypes of preeclampsia by investigating the effects on mother, placenta and fetus of chronic hypoxia during late pregnancy. To achieve this, we developed a preclinical model of improved human translational potential to investigate the symptoms of preeclampsia in late gestation in sheep, independent of maladaptive placental changes in early pregnancy.
Materials & methods
The authors declare that all supporting data are available within the article [and its online supplementary files].
For the purpose of the current study, we exploited recently available novel technology to maintain pregnant sheep under highly controlled isobaric hypoxic conditions, while undergoing wireless recording of maternal cardiovascular function (Figure 1)6–8. Then, we combined measurements in vivo with functional and molecular analyses to determine the inter-relationship between chronic hypoxia and maternal, placental and fetal outcomes. The experimental design was conducted in accordance with the ARRIVE guidelines9.
Figure 1. Isobaric hypoxic chambers and wireless recording CamDAS™ system.

Both panels: A specially designed nitrogen-generating system supplied compressed air and nitrogen to the bespoke isobaric hypoxic chambers housed at The Barcroft Centre, University of Cambridge. Each chamber was equipped with an electronic servo-controlled humidity cool steam injection system to return the appropriate humidity to the inspirate (i). Ambient partial pressures of oxygen and carbon dioxide, humidity, and temperature within each chamber were monitored via sensors (ii). For experimental procedures, each chamber had a double transfer port (iii) to internalise material and a manually operated sliding panel (iv) to bring the ewe into a position, where daily sampling of blood could be achieved through glove compartments (v). Each chamber incorporated a drinking bowl on continuous water supply and a rotating food compartment (vi) for determining food intake. A sealed transfer isolation cart could be attached to a side exit (vii) to couple chambers together for cleaning. Waste could be disposed via a sealable pipe (viii). Panel B: A separate cohort of ewes was instrumented with the CamDAS™ system during surgery, allowing continuous longitudinal monitoring of arterial blood pressure and uterine blood flow. The wireless CamDAS™ system was contained in two parts in a custom-made sheep jacket: the data acquisition box (ix) on one side and a box containing the pressure transducers (x) on the other side. Cables (xi) provided connection between the two boxes and to two battery packs. Measurements made using the CamDAS™ system were transmitted wirelessly via Bluetooth technology (xiii) to a laptop on the outside (xii), on which it was possible to continuously measure and record uterine blood flow and maternal arterial blood pressure during the experimental period (reproduced with permission6,7).
An expanded version of the Materials & Methods is available in the Online Supplement.
Results
Chronic hypoxia causes asymmetric fetal growth restriction
Exposure of pregnant ewes to chronic isobaric hypoxia of 10% inspired oxygen for a month from 105 to 138 days gestational age (dGA; term at 145 dGA) was associated with a 28% reduction in fetal growth, decreasing fetal weight from 3.67±0.17kg in normoxic (N) fetuses to 2.65±0.22kg in hypoxic (H) fetuses at 138 dGA (Figure 2A). There was no change in fetal brain weight in H relative to N pregnancies (N: 47.7±0.9 vs. H: 47.8±1.3g). However, when fetal brain weight was expressed relative to fetal body weight, this ratio was significantly increased in H relative to N fetuses (Figure 2B). In contrast, there was no effect of chronic hypoxia on placental weight (Figure 2C) or on the number or weight distribution of different placentome types (Figure S1). These effects of chronic hypoxia on fetal growth occurred in the absence of changes to maternal food intake (Figure S2).
Figure 2. Hypoxic pregnancy causes asymmetric FGR.

Values are mean ± SEM for fetal weight (A), the ratio of fetal brain to body weight (B) and placental weight (C). Groups are N (○,n=9-13) and H (●,n=7-8). Significant differences (P<0.05) are *N vs. H, Student’s t-test for unpaired data.
Chronic hypoxia leads to activation of the placental unfolded protein response
The levels of hypoxia-inducible factor 1α (HIF1α) were greater in H relative to N placentomes at 138 dGA (Figure 3A). The levels of protein carbonylation were greater in H relative to N placentae (Figure 3B), indicative of oxidative stress. This was associated with an increase in the ratio of the phosphorylated forms of the mitogen activated protein kinases C-jun N-terminal kinase (JNK) and the cell survival extracellular signal-regulated kinase (ERK) compared to total levels of these kinases in H relative to N placentae (Figure 3C and 3D). Oxidative protein damage can trigger activation of unfolded protein response (UPR) pathways in different cellular compartments, including the cytosol and endoplasmic reticulum (ER). The UPR signal activator activating transcription factor 6 (ATF6) was increased in H relative to N placentae (Figure 4A). As part of the endoplasmic reticulum UPR (UPRER), levels of the protein chaperone glucose-related protein 78 (GRP78) and of the protein folding enzyme protein disulfide isomerase (PDI) were higher in H relative to N placentae (Figure 4B). The expression of the cytosolic protein chaperones heat shock protein 27 (HSP27) and heat shock protein 70 (HSP70), part of the cytosolic UPR (UPRCyt), was also greater in H relative to N placentae (Figure 4C). Immunohistochemical analysis showed that ATF6 localised to the nucleus, indicating potential transcriptional activity of ATF6 (Figure 4D). Nuclear staining was more prominent in H compared to N placentae (Figure 4D). Transmission electron microscopy further revealed distended ER morphology in H compared with the ER in N placentae, which displayed a highly defined membrane structure (Figure 4E).
Figure 3. Hypoxic pregnancy activates the placental stress response.

Values are mean ± SEM for the relative ratio of placental levels of HIF1α (A), of protein carbonylation (B), of the ratio of phosphorylated to total stress kinases JNK (C) and ERK (D). Blots for JNK and ERK appear atypical as they were resolved on 14% agarose gels for higher resolution. Groups are N (○,n=9-10) and H (●,n=7). Significant differences (P<0.05) are *N vs. H, Student’s t-test for unpaired data.
Figure 4. Hypoxic pregnancy activates the placental UPR response.

Values are mean ± SEM for the relative ratio of the placental levels of ATF6 (A), GRP78 and PDI (B) and HSP27 and HSP70 (C). Groups are N (○,n=9-10) and H (●,n=7). Significant differences (P<0.05) are *N vs. H, Student’s t-test for unpaired data. In the placenta, ATF6 localises to the nuclei (D), with more prominent nuclear staining in H compared to N placentae. Pictured (D): trophoblast containing binucleate cells (arrows); scale bar = 50μm. Change in trophoblast ER structure was examined by transmission electron microscopy (E). Representative images taken at 5,000x magnification are shown. Arrows indicate the location of ER and stars indicate the location of the nucleus; scale bar = 500nm.
Chronic hypoxia promotes an angiogenic imbalance in the maternal circulation
Placental transcripts encoding the anti-angiogenic factors soluble fms-like tyrosine kinase 1 (sFlt-1) and soluble endoglin (sEng), as well as the ratio of the placental transcripts of sFlt-1 compared to the angiogenic factor vascular endothelial growth factor (VEGF), were increased in H relative to N placentae at 138 dGA, as measured by quantitative reverse transcription PCR (Figure 5A-5C). There were no differences in VEGF and PlGF transcripts, and no differences in the ratio of sFlt-1 compared to PlGF transcripts (Figure S3A-S3C). In N ewes, the concentration of sFlt-1 and the ratios of sFlt-1 to VEGF and to PlGF in plasma did not change in samples taken at baseline and at 138 dGA (Figure 5D-5F). In contrast, in H ewes the concentrations of sFlt-1 and the ratios of sFlt-1 to VEGF and to PlGF were significantly higher at 138 dGA relative to baseline and when compared to values in N ewes at 138 dGA (Figure 5D-5F). Neither N nor H ewes showed changes in sEng, VEGF or PlGF plasma concentrations with increasing gestation and there were no differences between the groups at baseline or 138 dGA (Figure S3D-S3F).
Figure 5. Hypoxic pregnancy changes placental expression of anti-angiogenic factors, increasing the anti- to pro-angiogenic balance in maternal plasma.

Upper panel: Values are mean ± SEM for the relative placental fold change for sFlt-1 (A), sEng (B) and the ratio of sFlt-1 to VEGF (C). Lower panel: Values are mean ± SEM for plasma concentration of sFlt-1 (D) and plasma ratios of sFlt-1 to PlGF (E) and to VEGF (F). Groups are N (○,n=7-9) and H (●,n=7). Significant differences (P<0.05) are *N vs. H or † vs baseline; Student’s t-test for unpaired data or two-way RM-ANOVA where appropriate.
Chronic hypoxia increases uteroplacental vascular resistance and prevents the gestational decrease in maternal arterial blood pressure
At 138 dGA, the uterine artery PI values were greater in H relative to N ewes (Figure 6A). At 138 dGA, independent of treatment, there were significant positive correlations between maternal uterine PI and the maternal plasma sFlt-1 concentration, and between maternal uterine PI and maternal plasma sFlt-1 to PlGF ratio (Figure S4A and S4C). However, there was no correlation between maternal uterine PI and maternal plasma sFlt-1 to VEGF ratio at 138 dGA (Figure S4B). At 138 dGA, plasma creatinine concentrations were slightly higher in H relative to N ewes, which may indicate a reduction in glomerular filtration rate (Figure S5A). However, there was no difference in the urine ratio of albumin to creatinine at 138 dGA (Figure S5B).
Figure 6. Hypoxic pregnancy causes maternal cardiovascular dysfunction.

Values are mean ± SEM for uterine artery PI (A) and the change from baseline in uterine vascular resistance (B) and in arterial blood pressure (C). Groups are N (○,n=5-9), H (●,n=5-7). Significant differences (P<0.05) are *N vs. H, or † vs baseline; Student’s t-test for unpaired data or two-way RM-ANOVA, where appropriate.
Daily changes in maternal arterial blood gas, acid base and metabolic status during exposure to chronic normoxia or chronic hypoxia in the second cohort of animals have been previously reported7. In brief, these data confirm a reduction in maternal arterial partial pressure of oxygen (PaO2) from 105.7±3.7 to 42.0±1.2mmHg and in arterial oxygen saturation 103.5±0.5 to 78.6±5.7% (P<0.05) during exposure to chronic isobaric hypoxia of 10% inspired oxygen (Table S1). Ewes exposed to chronic hypoxia had significantly elevated haematocrit and haemoglobin concentration by the end of exposure relative to baseline and to values in N ewes (Table S1). There was no significant change between groups in maternal arterial pH, partial pressure of arterial carbon dioxide, blood glucose or lactate concentrations (Table S1).
Longitudinal continuous measurement of maternal arterial blood pressure and calculation of uterine vascular resistance via the wireless CamDAS recording revealed the expected fall in both variables with advancing gestation in N ewes but not in H ewes (Figure 6B and 6C). During baseline conditions, prior to hypoxic exposure, between 122-124 dGA, average values for maternal arterial blood pressure (80.4±1.2 vs. 81.2±3.2mmHg) and for uterine vascular resistance (0.29±0.05 vs. 0.20±0.03mmHg.(ml.min-1)-1) were not different between N and H ewes. In N pregnancies, values for maternal arterial blood pressure (80.4±1.2 vs. 73.4±1.1mmHg) and for uterine vascular resistance (0.29±0.05 vs. 0.20±0.06mmHg.(ml.min-1)-1) were significantly lower at 134 dGA compared to earlier in gestation (all P<0.05). In contrast, following chronic hypoxic exposure, values for maternal arterial blood pressure (81.2±3.2 vs. 79.6±4.8mmHg) and for uterine vascular resistance (0.20±0.03 vs. 0.19±0.04mmHg.(ml.min-1)-1) in H pregnancies, were similar at 134 dGA compared to earlier in gestation (P>0.05). Maternal arterial blood pressure was not significantly different between N and H pregnancies at 134 dGA.
Discussion
In the classic two-stage theory of preeclampsia, impaired uteroplacental perfusion promotes placental hypoxia, oxidative stress and ER stress. The subsequent placental release of proinflammatory cytokines, syncytiotrophoblast debris and antiangiogenic factors into the maternal circulation induces the peripheral syndrome10. For example, the angiogenic imbalance caused by the release of sFlt-1 and sEng, which oppose the actions of VEGF and PlGF, promotes global maternal endothelial dysfunction and increased peripheral vascular resistance11. These changes manifest as an increase in the uterine PI, maternal systemic hypertension and impaired renal glomerular filtration rate12. On the fetal side, these changes lead to impaired fetal oxygen and nutrient delivery, yielding asymmetric fetal growth restriction (FGR)13.
Data in the present study show that chronic hypoxia during the last third of pregnancy in sheep stresses the placenta, with upstream adverse effects on the mother and downstream adverse effects on the fetus, akin to those found in preeclampsia. Placentae from the hypoxic cohort showed molecular evidence of hypoxia, increased oxidative stress, activation of the UPR, dilation of ER cisternae, and increased expression of anti-angiogenic factors. Upstream adverse consequences on the ewe included evidence of an angiogenic imbalance in maternal plasma, increased uterine artery PI and a lack of an ontogenic fall in uterine vascular resistance and arterial blood pressure with advancing gestation. Downstream adverse consequences on the hypoxic offspring included FGR with evidence of fetal brain-sparing. Combined, therefore, the data in this study support the hypothesis that chronic hypoxia during the last third of pregnancy in sheep provides a link between placental stress, FGR and maternal cardiovascular dysfunction in adverse pregnancy, as in preeclampsia. However, the differences between this pre-clinical model and preeclampsia are just as informative as the similarities. While the data suggest that some features of preeclampsia can be caused by hypoxia, other features, such as overt maternal hypertension and maternal proteinuria, were not recapitulated. However, ewes undergoing hypoxic pregnancy did not show the significant fall in maternal arterial blood pressure measured in control ewes with advancing gestation. Lack of maternal hypertension may therefore be due to the limited duration of hypoxia towards the end of pregnancy in this ovine model. In contrast, in preeclampsia, the pathophysiology can start during the first trimester.
Chronic hypoxia and asymmetric fetal growth restriction
The level of maternal hypoxia used in this model is clinically relevant. Previous studies from our group have used the hypoxic chambers with pregnant sheep, which were surgically prepared with catheters for daily blood sampling and Transonic flow probes for long-term recording of fetal cardiovascular function7. These studies revealed that the level of maternal hypoxia used in the present study reduced fetal PaO2 in the descending aorta to 12mmHg in a highly controlled manner7. This level of chronic hypoxia equates to that measured by cordocentesis in human growth restricted fetuses in preeclamptic pregnancies14. Our previous studies also revealed that chronic fetal hypoxia promotes a sustained redistribution of blood flow away from the peripheral circulations towards the fetal brain7,15. This is the so-called ‘fetal brain-sparing effect’16 and is responsible for the asymmetric FGR measured in chronically hypoxic fetuses both in humans and animal models15,17. In the present study, the asymmetric FGR resulting from chronic hypoxia during the last third of pregnancy was represented by a smaller fall in the brain relative to the fetal body weight, yielding an increase in the percentage relative brain weight.
Chronic hypoxia and placental stress
Placental hypoxia promotes an increase in placental oxidative stress18. Protein carbonyls are used as biomarkers of reactive oxygen species-mediated protein damage in preeclamptic placentae, and correlate well with the severity of the syndrome19. Accumulation of damaged proteins in the placenta is associated with activation of the kinases ERK and JNK, which mediate several responses to cellular stress 20. As part of the cellular quality control system, the ER ensures protein folding, and is capable of activating a powerful UPR to restore protein homeostasis21. In the present study, both the UPRER and UPRCyt showed increased activation in hypoxic placentae, along with morphological changes in ER structure. We found that both the expression and nuclear translocation of ATF6 was increased, likely mediating the transcriptional activation of UPR target genes in response to ER stress22. In hypoxic placentae the expression of GRP78 and PDI and of HSP27 and HSP70 were increased as part of the UPRER and the UPRCyt, respectively. Transmission electron microscopy further revealed distended ER cristae in hypoxic placentae. Many of these molecular and morphological markers have been reported in placentae from women suffering from preeclampsia23–25.
Chronic hypoxia and maternal adverse effects
Healthy human pregnancy is accompanied by a number of maternal cardiovascular adaptations that help support the growing fetus26. By mid-gestation, there is a fall in utero-placental vascular resistance, which directs perfusion towards the uterine artery, where blood flow is increased from 20-50 ml/min in the non-pregnant state to 450-800 ml/min26. To accommodate this, the uterine artery markedly increases its diameter, driving a fall in uterine vascular resistance and maternal arterial pressure27. Pregnancy at high altitude blunts the rise in uterine blood flow and impairs the fall in maternal arterial blood pressure with advancing gestation in non-indigenous human populations28,29 and sheep30,31. The diminished rise in uterine blood flow in human highland pregnancy is thought to be an important contributor to the enhanced prevalence of preeclampsia and FGR at high altitude32,33. Extensive studies by Zhang and colleagues have shown that gestational hypoxia contributes to the maladaptive uterine haemodynamic phenotype through epigenetic regulation of the large conductance calcium-activated potassium channel30,34.
Data in the present study show that chronic hypoxia during the last third of pregnancy in sheep led to an increase in placental sFlt-1 expression and maternal plasma sFlt-1 concentration. This may be driven by increased placental levels of HIF1α in the hypoxic placenta, which has been previously demonstrated in vitro in placental explants and is supported by raised HIF1α levels in the current study35. In addition, the fall in uterine vascular resistance and maternal blood pressure with advancing gestation monitored using indwelling flow probes and vascular catheters did not occur in hypoxic ewes. Both uterine vascular dysfunction and increased maternal blood pressure have been reported in sheep undergoing high altitude pregnancy34. Therefore, combined, the present study extends previous findings in ovine highland pregnancy34, highlighting the critical role for oxygen deficiency in placental dysfunction and their relationship with maternal cardiovascular changes. Further, there was a significant positive correlation between maternal uterine PI and the maternal plasma concentration of sFlt-1, and between maternal uterine PI and the maternal plasma sFlt-1 to PlGF ratio. The data therefore support that increased expression of antiangiogenic factors in the placenta may contribute to an angiogenic imbalance and endothelial dysfunction in the maternal circulation.
Advances and limitations
Despite great advances in the understanding of preeclampsia, progress in this field has been hampered by many experimental limitations. While advances such as organoid cultures create new and exciting opportunities, in vitro models cannot replicate all in vivo interactions between mother, placenta and offspring. On the other hand, there are no preclinical animal models that spontaneously develop preeclampsia, and those available, in which symptoms are induced, have limitations. The same is true for this ovine model of hypoxic pregnancy. Clearly, there are gross anatomical differences between the human haemochorial and the ovine synepitheliochorial placenta36. However, there are also important similarities. Both sheep and humans have placental counter-current flow of maternal and fetal blood within the placental villous tree, comparable transplacental oxygen gradients and oxygen consumption rates (37 in sheep vs. 34ml.kg-1.min-1 in humans), as well as similar nutrient transporter expression37–41. At the molecular level, the induction of oxidative and ER stress and the activation of the UPR are highly conserved pathways across species21. This is also the case for rodents, in which activation of the placental UPRER has been demonstrated in hypoxic pregnancy34.
There have been many other studies by our and other groups in rodent pregnancy investigating the effects of hypoxic pregnancy on the placental phenotype, FGR and on uterine vascular reactivity42–46. However, few of these studies have had a focus on placental molecular or maternal circulating markers of preeclampsia or investigated associated changes in maternal in vivo cardiovascular function. It is also important to highlight that the murine placenta is functionally divided into distinct zones for endocrine activity and for nutrient transfer47. The labyrinth zone shows high levels of mitochondrial activity, while the junctional zone is less well oxygenated, but prone to ER stress due to its synthetic and secretory activities48. Thus, in the murine placenta, crosstalk between ER and mitochondrial stress is limited, with diverging responses depending on which zone is investigated49,50. Other murine studies using experimental models of preeclampsia by either eNOS knockout or restriction of uteroplacental perfusion also support a link between placental hypoxia with oxidative stress and impaired placental nutrient transport, FGR and abnormal maternal cardiovascular function49,51. This is what we have directly addressed in the present study in an ovine model with increased human translational potential. Nevertheless, extrapolation of these findings to the human clinical condition needs to be viewed with caution.
Perspectives
This work introduces a novel large animal model of isobaric hypoxic pregnancy in sheep that not only promotes fetal growth restriction but also recapitulates many of the physiological and molecular features of preeclampsia in the mother and the placenta. These findings are significant, as any changes occur independent of alterations to placentation in early pregnancy. Therefore, the work offers novel ways of thinking about the syndrome and an established platform to develop interventional therapies.
Supplementary Material
{kind=link}
Novelty and Relevance.
What is new?
A common feature of preeclampsia is placental hypoxia. However, whether placental hypoxia is merely a consequence of the disease or whether it causes the adverse maternal and fetal outcomes is uncertain.
Here, we show using a novel in vivo approach that placental hypoxia drives maternal and fetal phenotypes associated with preeclampsia.
What is relevant?
Pregnancy affected by uteroplacental hypoxia, one of the most common pregnancy complications leading to fetal growth restriction in humans, increases the risk of physiological and molecular markers of preeclampsia.
Clinical and pathophysiological implications?
Chronic hypoxia in ovine pregnancy recapitulates markers of preeclampsia in the mother, placenta and offspring, indicating that placental hypoxia is an initiating factor in the pathoaetiology of preeclampsia.
Therefore, biomarkers of placental hypoxia, oxidative stress and activation of the unfolded protein response must be addressed to guide future clinical management of preeclampsia.
Acknowledgements
W.T. is a PhD student supported by the Centre for Trophoblast Research at the University of Cambridge. We are grateful to the staff of the University of Cambridge Biological Services for helping with the maintenance of the animals at The Barcroft Centre.
Funding sources
This work was supported by The British Heart Foundation (RG/17/8/32924).
Footnotes
Competing interests
The authors declare no competing interests.
Author contributions
Conceptualisation, D.A.G.; Methodology, W.T., B.J.A., K.L.B., O.V.P., Y.N., K.J.B., S.G.F., T.A.G., F.B.P., Q.L., L.Z., J.M., T.C.D., H.W.Y., G.J.B., D.A.G.; Formal Analysis, W.T., B.J.A., K.L.B., O.V.P., Y.N., K.J.B., S.G.F., T.A.G., F.B.P., Q.L., L.Z., J.M., T.C.D., H.W.Y., G.J.B., D.A.G.;.; Writing – Original Draft, W.T., B.J.A., K.L.B., D.A.G.; Writing – Review & Editing, W.T., B.J.A., K.L.B., O.V.P., Y.N., K.J.B., S.G.F., T.A.G., F.B.P., Q.L., L.Z., J.M., T.C.D., H.W.Y., G.J.B.; Visualisation, W.T., D.A.G.; Supervision, W.T., B.J.A., K.L.B., O.V.P., Y.N., K.J.B., S.G.F., T.A.G., F.B.P., Q.L., L.Z., J.M., T.C.D., H.W.Y., G.J.B., D.A.G.; Project Administration, W.T., B.J.A., K.L.B., O.V.P., Y.N., K.J.B., S.G.F., D.A.G.; Funding Acquisition, D.A.G., G.J.B.
Resource availability
Lead contact
Further information and requests for resources and reagents should be directed to and will be fulfilled by the Lead Contact, Professor Dino A. Giussani (dag26@cam.ac.uk).
Materials availability
This study did not generate new unique reagents.
Data and code availability
This study did not generate/analyse any datasets or code
References
- 1.Gynecologists ACoOa. Gestational Hypertension and Preeclampsia: ACOG Practice Bulletin, Number 222. Paper/Poster presented at: Obstetrics & Gynecology. 2020 doi: 10.1097/AOG.0000000000003891. [DOI] [PubMed]
- 2.Steegers EAP, von Dadelszen P, Duvekot JJ, Pijnenborg R. Pre-eclampsia. The Lancet. 2010;376:631–644. doi: 10.1016/S0140-6736(10)60279-6. [DOI] [PubMed] [Google Scholar]
- 3.Burton GJ, Redman CW, Roberts JM, Moffett A. Pre-eclampsia: pathophysiology and clinical implications. BMJ. 2019;366:l2381. doi: 10.1136/bmj.l2381. [DOI] [PubMed] [Google Scholar]
- 4.Barry JS, Anthony RV. The Pregnant Sheep as a Model for Human Pregnancy. Theriogenology. 2008;69:55–67. doi: 10.1016/j.theriogenology.2007.09.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jellyman JK, Gardner DS, Fowden AL, Giussani DA. Effects of dexamethasone on the uterine and umbilical vascular beds during basal and hypoxemic conditions in sheep. American journal of obstetrics and gynecology. 2004;190:825–835. doi: 10.1016/j.ajog.2003.09.046. [DOI] [PubMed] [Google Scholar]
- 6.Brain KL, Allison BJ, Niu Y, Cross CM, et al. Induction of controlled hypoxic pregnancy in large mammalian species. Physiological Reports. 2015;3:e12614. doi: 10.14814/phy2.12614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Allison BJ, Brain KL, Niu Y, Kane AD, et al. Fetal in vivo continuous cardiovascular function during chronic hypoxia. The Journal of Physiology. 2016;594:1247–1264. doi: 10.1113/JP271091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Shaw CJ, Allison BJ, Itani N, Botting KJ, et al. Altered autonomic control of heart rate variability in the chronically hypoxic fetus. The Journal of Physiology. 2018;596:6105–6119. doi: 10.1113/jp275659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kilkenny C, Browne WJ, Cuthill IC, Emerson M, et al. Improving Bioscience Research Reporting: The ARRIVE Guidelines for Reporting Animal Research. PLoS Biology. 2010;8:e1000412. doi: 10.1371/journal.pbio.1000412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Tong W, Giussani DA. Preeclampsia link to gestational hypoxia. Journal of developmental origins of health and disease. 2019;10:322–333. doi: 10.1017/S204017441900014X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Powe CE, Levine RJ, Karumanchi SA. Preeclampsia, a disease of the maternal endothelium: the role of antiangiogenic factors and implications for later cardiovascular disease. Circulation. 2011;123:2856–2869. doi: 10.1161/CIRCULATIONAHA.109.853127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.VanWijk MJ, Kublickiene K, Boer K, VanBavel E. Vascular function in preeclampsia. Cardiovascular research. 2000;47:38–48. doi: 10.1016/S0008-6363(00)00087-0. [DOI] [PubMed] [Google Scholar]
- 13.Poston L. The control of blood flow to the placenta. Experimental Physiology. 1997;82:377–387. doi: 10.1113/expphysiol.1997.sp004033. [DOI] [PubMed] [Google Scholar]
- 14.Nicolaides KH, Rodeck CH, Soothill PW, Campbell S. Ultrasound-guided sampling of umbilical cord and placental blood to assess fetal wellbeing. The Lancet. 1986;327:1065–1067. doi: 10.1016/S0140-6736(86)91333-4. [DOI] [PubMed] [Google Scholar]
- 15.Giussani DA. Breath of Life: Heart Disease Link to Developmental Hypoxia. Circulation. 2021;144:1429–1443. doi: 10.1161/CIRCULATIONAHA.121.054689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Giussani DA. The fetal brain sparing response to hypoxia: physiological mechanisms. The Journal of physiology. 2016;594:1215–1230. doi: 10.1113/JP271099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Swanson AM, David AL. Animal models of fetal growth restriction: Considerations for translational medicine. Placenta. 2015;36:623–630. doi: 10.1016/j.placenta.2015.03.003. [DOI] [PubMed] [Google Scholar]
- 18.Sferruzzi-Perri AN, Higgins JS, Vaughan OR, Murray AJ, et al. Placental mitochondria adapt developmentally and in response to hypoxia to support fetal growth. Proc Natl Acad Sci U S A. 2019;116:1621–1626. doi: 10.1073/pnas.1816056116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zusterzeel PLM, Rütten H, Roelofs HMJ, Peters WHM, et al. Protein Carbonyls in Decidua and Placenta of Pre-eclamptic Women as Markers for Oxidative Stress. Placenta. 2001;22:213–219. doi: 10.1053/plac.2000.0606. [DOI] [PubMed] [Google Scholar]
- 20.Tang C, Liang J, Qian J, Jin L, et al. Opposing role of JNK-p38 kinase and ERK1/2 in hydrogen peroxide-induced oxidative damage of human trophoblast-like JEG-3 cells. International Journal of Clinical and Experimental Pathology. 2014;7:959–968. [PMC free article] [PubMed] [Google Scholar]
- 21.Schröder M, Kaufman RJ. The mammalian unfolded protein response. Annual Review of Biochemistry. 2005;74:739–789. doi: 10.1146/annurev.biochem.73.011303.074134. [DOI] [PubMed] [Google Scholar]
- 22.Adams CJ, Kopp MC, Larburu N, Nowak PR, et al. Structure and Molecular Mechanism of ER Stress Signaling by the Unfolded Protein Response Signal Activator IRE1. Frontiers in Molecular Biosciences. 2019;6 doi: 10.3389/fmolb.2019.00011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Charnock-Jones DS. Placental hypoxia, endoplasmic reticulum stress and maternal endothelial sensitisation by sFLT1 in pre-eclampsia. Journal of Reproductive Immunology. 2016;114:81–85. doi: 10.1016/j.jri.2015.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Yung HW, Cox M, Tissot van Patot M, Burton GJ. Evidence of endoplasmic reticulum stress and protein synthesis inhibition in the placenta of non-native women at high altitude. FASEB J. 2012;26:1970–1981. doi: 10.1096/fj.11-190082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yung HW, Atkinson D, Campion-Smith T, Olovsson M, et al. Differential activation of placental unfolded protein response pathways implies heterogeneity in causation of early- and late-onset pre-eclampsia. The Journal of Pathology. 2014;234:262–276. doi: 10.1002/path.4394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Palmer SK, Zamudio S, Coffin C, Parker S, et al. Quantitative estimation of human uterine artery blood flow and pelvic blood flow redistribution in pregnancy. Obstetrics and gynecology. 1992;80:1000–1006. [PubMed] [Google Scholar]
- 27.Kublickiene KR, Cockell AP, Nisell H, Poston L. Role of nitric oxide in the regulation of vascular tone in pressurized and perfused resistance myometrial arteries from term pregnant women. Am J Obstet Gynecol. 1997;177:1263–1269. doi: 10.1016/s0002-9378(97)70048-6. [DOI] [PubMed] [Google Scholar]
- 28.Palmer SK, Moore LG, Young D, Cregger B, et al. Altered blood pressure course during normal pregnancy and increased preeclampsia at high altitude (3100 meters) in Colorado. Am J Obstet Gynecol. 1999;180:1161–1168. doi: 10.1016/s0002-9378(99)70611-3. [DOI] [PubMed] [Google Scholar]
- 29.Giussani DA, Salinas CE, Villena M, Blanco CE. The role of oxygen in prenatal growth: studies in the chick embryo. The Journal of physiology. 2007;585:911–917. doi: 10.1113/jphysiol.2007.141572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hu X-Q, Chen M, Dasgupta C, Xiao D, et al. Chronic hypoxia upregulates DNA methyltransferase and represses large conductance Ca2+-activated K+ channel function in ovine uterine arteries. Biology of reproduction. 2017;96:424–434. doi: 10.1095/biolreprod.116.145946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ducsay CA, Goyal R, Pearce WJ, Wilson S, et al. Gestational Hypoxia and Developmental Plasticity. Physiological reviews. 2018;98:1241–1334. doi: 10.1152/physrev.00043.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Keyes LE, Armaza JF, Niermeyer S, Vargas E, et al. Intrauterine growth restriction, preeclampsia, and intrauterine mortality at high altitude in Bolivia. Pediatr Res. 2003;54:20–25. doi: 10.1203/01.PDR.0000069846.64389.DC. [DOI] [PubMed] [Google Scholar]
- 33.Julian CG, Galan HL, Wilson MJ, Desilva W, et al. Lower uterine artery blood flow and higher endothelin relative to nitric oxide metabolite levels are associated with reductions in birth weight at high altitude. Am J Physiol Regul Integr Comp Physiol. 2008;295:R906–915. doi: 10.1152/ajpregu.00164.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hu X-Q, Dasgupta C, Xiao D, Huang X, et al. MicroRNA-210 Targets Ten-Eleven Translocation Methylcytosine Dioxygenase 1 and Suppresses Pregnancy-Mediated Adaptation of Large Conductance Ca(2+)-Activated K(+) Channel Expression and Function in Ovine Uterine Arteries. Hypertension (Dallas, Tex 1979) 2017 doi: 10.1161/HYPERTENSIONAHA.117.09864. HYPERTENSIONAHA.117.09864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Iwagaki S, Yokoyama Y, Tang L, Takahashi Y, et al. Augmentation of leptin and hypoxia-inducible factor 1α mRNAs in the pre-eclamptic placenta. Gynecological Endocrinology. 2004;18:263–268. doi: 10.1080/0951359042000196277. [DOI] [PubMed] [Google Scholar]
- 36.Wooding FBP, Burton G. Comparative Placentation: Structures, Functions and Evolution. 2008 [Google Scholar]
- 37.Wilkening RB, Molina RD, Meschia G. Placental oxygen transport in sheep with different hemoglobin types. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 1988;254:R585–R589. doi: 10.1152/ajpregu.1988.254.4.R585. [DOI] [PubMed] [Google Scholar]
- 38.Bonds DR, Crosby LO, Cheek TG, Hä gerdal M, et al. Estimation of human fetal-placental unit metabolic rate by application of the Bohr principle. Journal of developmental physiology. 1986;8:49–54. [PubMed] [Google Scholar]
- 39.Ma Y, Zhu MJ, Uthlaut AB, Nijland MJ, et al. Upregulation of growth signaling and nutrient transporters in cotyledons of early to mid-gestational nutrient restricted ewes. Placenta. 2011;32:255–263. doi: 10.1016/j.placenta.2011.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Regnault TRH, de Vrijer B, Galan HL, Wilkening RB, et al. Umbilical uptakes and transplacental concentration ratios of amino acids in severe fetal growth restriction. Pediatric Research. 2013;73:602. doi: 10.1038/pr.2013.30. [DOI] [PubMed] [Google Scholar]
- 41.Hafez SA, Borowicz P, Reynolds LP, Redmer DA. Maternal and fetal microvasculature in sheep placenta at several stages of gestation. J Anat. 2010;216:292–300. doi: 10.1111/j.1469-7580.2009.01184.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Nuzzo AM, Camm EJ, Sferruzzi-Perri AN, Ashmore TJ, et al. Placental Adaptation to Early-Onset Hypoxic Pregnancy and Mitochondria-Targeted Antioxidant Therapy in a Rodent Model. The American Journal of Pathology. 2018;188:2704–2716. doi: 10.1016/j.ajpath.2018.07.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Skeffington KL, Higgins JS, Mahmoud AD, Evans AM, et al. Hypoxia, AMPK activation and uterine artery vasoreactivity. The Journal of physiology. 2016;594:1357–1369. doi: 10.1113/JP270995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Matheson H, Veerbeek JH, Charnock-Jones S, Burton GJ, et al. Morphological and molecular changes in the murine placenta exposed to normobaric hypoxia throughout pregnancy. The Journal of Physiology. 2016;594:1371–1388. doi: 10.1113/JP271073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Higgins JS, Vaughan OR, Fernandez de Liger E, Fowden AL, et al. Placental phenotype and resource allocation to fetal growth are modified by the timing and degree of hypoxia during mouse pregnancy. The Journal of physiology. 2016;594:1341–1356. doi: 10.1113/JP271057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Camm EJ, Hansell JA, Kane AD, Herrera EA, et al. Partial contributions of developmental hypoxia and undernutrition to prenatal alterations in somatic growth and cardiovascular structure and function. American Journal of Obstetrics and Gynecology. 2010;203:495.e424–495.e434. doi: 10.1016/j.ajog.2010.06.046. [DOI] [PubMed] [Google Scholar]
- 47.Woods L, Perez-Garcia V, Hemberger M. Regulation of Placental Development and Its Impact on Fetal Growth-New Insights From Mouse Models. Front Endocrinol (Lausanne) 2018;9:570. doi: 10.3389/fendo.2018.00570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yung HW, Colleoni F, Dommett E, Cindrova-Davies T, et al. Noncanonical mitochondrial unfolded protein response impairs placental oxidative phosphorylation in early-onset preeclampsia. Proceedings of the National Academy of Sciences. 2019;116:18109–18118. doi: 10.1073/pnas.1907548116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kusinski LC, Stanley JL, Dilworth MR, Hirt CJ, et al. eNOS knockout mouse as a model of fetal growth restriction with an impaired uterine artery function and placental transport phenotype. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 2012;303:R86–R93. doi: 10.1152/ajpregu.00600.2011. [DOI] [PubMed] [Google Scholar]
- 50.Burton GJ, Yung HW, Murray AJ. Mitochondrial – Endoplasmic reticulum interactions in the trophoblast: Stress and senescence. Placenta. 2017;52:146–155. doi: 10.1016/j.placenta.2016.04.001. [DOI] [PubMed] [Google Scholar]
- 51.Amaral LM, Pinheiro LC, Guimaraes DA, Palei ACT, et al. Antihypertensive effects of inducible nitric oxide synthase inhibition in experimental pre-eclampsia. Journal of cellular and molecular medicine. 2013;17:1300–1307. doi: 10.1111/jcmm.12106. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
This study did not generate/analyse any datasets or code