

Abstract
Translation of mRNA is a fundamental process in gene expression and control of translation is an important method by which protein synthesis is regulated in cells. The primary hallmark of eukaryotic mRNAs is their 5′ cap, whose molecular contacts to the eukaryotic translation initiation factor eIF4E govern initiation of translation. Here we report 5′ cap analogues with photo-cleavable groups (FlashCaps) that prohibit binding to eIF4E and resist cleavage by decapping enzymes. These compounds are compatible with the general and efficient production of mRNAs by in vitro transcription. In FlashCap-mRNAs, the single photocaging group abrogates translation in vitro and in mammalian cells without increasing immunogenicity. Irradiation restores the native cap, triggering efficient translation. FlashCaps overcome the problem of remaining sequence or structure changes in mRNA after irradiation, which limited previous designs. Together, these results demonstrate that FlashCaps offer a route to regulate the expression of any given mRNA and to dose mRNA-therapeutics with spatio-temporal control.
Messenger RNAs (mRNAs) have recently entered the public stage as most versatile medical modalities. Prominent examples are the mRNA-based vaccines by Moderna and BioNTech/Pfizer coding for the spike proteins to protect against infection by SARS-Cov21. The mRNA technology is not limited to vaccination, and can also greatly improve, for example, therapy for autoimmune diseases or personalised cancer treatment2. Translation of mRNA into proteins is one of the fundamental and highly conserved processes in the cell and works for endogenous and exogenous transcripts (Fig. 1a). Its regulation is essential in cell differentiation, cell proliferation and localized translation3,4 but also relevant for pathologies5. In mRNA therapy, however, at present, one cannot control when and where mRNA has an effect, i.e. when and where it is translated into proteins, which then have a pharmacological effect.
Figure 1. The 5′ cap is a hallmark of eukaryotic mRNAs governing translation initiation.

a) Key steps in translation initiation. The eukaryotic translation initiation factor eIF4E binds directly to the 5’ cap. The heterotrimeric eIF4F complex assembles on the 5′ cap leading to binding of the 40S ribosomal subunit, assembly of the eukaryotic 80S ribosome and translation initiation. b) Eukaryotic mRNA featuring cap 0 structure with recognition site for eIF4E, the site used for chemical modification in this study and the first transcribed nucleotide. c) Structure of eIF4E highlighting molecular interactions for cap 0 recognition. d) Concept of FlashCaps for light-induced translation. A single photo-cleavable group (red triangle) at the cap 0 impairs binding to eIF4E. FlashCaps are compatible with routine protocols for transcription and transfection. Following light-induced deprotection, the native mRNA with a 5’ cap 0 is released and translated. Abbreviations: UTR: untranslated region; PABP: poly(A) binding protein; ORF: open reading frame.
A hallmark of eukaryotic mRNAs is their 5′ cap, which in its simplest form (cap 0) links an N7-methylated guanosine to the first transcribed nucleotide via a 5′-5′ triphosphate bridge (Fig. 1b). Higher order cap structures contain additional methyl groups6. The 5′ cap plays a key role in translation initiation as the N7-methylated guanosine is essential for recognition by the translation initiation factor eIF4E (Fig. 1c). Importantly, the molecular contacts with the cap are sequence-independent, i.e. identical for all mRNAs7. The 5′ cap is also crucial for many mRNA processing and quality control steps, and protects eukaryotic mRNAs from degradation by exonucleases3,5,8. Dedicated decapping enzymes (Dcp1-2, DcpS) are required for mRNA turnover and homeostasis9,10. Together with the poly(A) tail at the 3′ end, the 5′ cap forms an mRNA ‘closed-loop’, facilitated by interactions between the cap-binding eIFs and the poly(A)-binding protein (PABP). The ‘closed loop’ promotes recruitment of the small ribosomal subunit (40S) and the complex enters the next initiation stages leading to formation of the 80S ribosome and translation (Fig. 1a)5. RNA without 5′ cap is barely translated and highly immunogenic11–13. Therefore, production of mRNAs for biological studies and therapeutic applications routinely involves in vitro transcription in the presence of synthetic cap analogues to obtain 5′ capped mRNAs14–16 (Fig. 1d).
In nature, the initiation phase of translation is the target of multiple types of regulatory intervention, enabling confinement of gene expression to a certain time span and cell region, e.g. in neurons, or multicellular organism5,17. The ability to control translation by external triggers – especially by light – would greatly enrich our ability to dissect cellular processes at the molecular level with high spatio-temporal precision. The directed release of mRNAs for translation at a certain time and destination would also provide an avenue to control the pharmacokinetics of mRNAs therapeutics. In this context, it would be important to avoid a drastic increase of immunogenicity.
However, methods to control gene expression externally at the mRNA level are scarce. Natural mechanisms triggering mRNA translation by light are still unknown and only one example of integrating photo-sensitive units in translation was reported to date18. Chemical approaches to directly photocage RNAs provide control of several RNA-regulated processes involving short regulatory RNAs, such as siRNAs, miRNAs, morpholinos and aptamers19–23. The chemical or chemo-enzymatic synthesis of long mRNAs, however, suffers from low yields24. Moreover, the installation of multiple modifications in the mRNA does not necessarily impede the ribosome25,26. Previous approaches towards controlling mRNA translation by light required tags27, multiple photocaging groups28 or photoswitches29–32– including photoswitches at the 5ʹ cap – that left the RNA altered. Remaining chemical modifications in the mRNA might affect the properties of the mRNA as shown for natural modifications33,34. Additional sequence elements may alter mRNA interactions, potentially disrupting regulatory processes of mRNA turnover35.
Here we report optochemical control of mRNA translation in eukaryotic cells. Our approach is based on a synthetic cap analogue (FlashCap) that efficiently interferes with the initiation stage of translation. Irradiation of FlashCap-mRNAs liberates an unaltered cap 0-mRNA molecule accessible for translation into hundreds of protein copies (Fig. 1a,d). This concept capitalizes on a single photocaging group at a defined position to leverage strong effects on translation of ~1000 nt long mRNAs. It is generally applicable, as synthetic 5′ cap analogues are routinely used in the production of mRNAs by in vitro transcription for research and therapeutic purposes. FlashCaps are therefore an efficient and readily applicable solution to make mRNA studies controllable by light, without requiring new production steps and without introducing artefacts into measurements.
Results
To achieve a strong effect on translation, we analysed the molecular interactions between the 5′ cap and the translation initiation factor eIF4E (Fig. 1c) as well as previous work on the effect of cap modifications on binding16,29,36–38. We anticipated that the installation of a sterically demanding residue, such as a photo-cleavable group, at the N2 position of the guanosine should interfere with direct hydrogen bonding to E103 required for proper positioning of the 5′ cap (Fig. 1c). At the same time, photo-deprotection should rapidly reconstitute the natural cap 0 and initiate translation. We therefore developed a synthesis route to 5′ caps with photo-cleavable groups at the N2 position of the cap guanosine (Fig. 2). To promote cap 0 release, we connected the photo-cleavable group via a self-immolative carbamate linkage. Photo-cleavage releases CO2 driving the deprotection reaction. Starting from guanosine (3), we first protected the three hydroxyl groups using trimethylsilyl (TMS) chloride. In a one-pot reaction, we then converted the free amino group of the guanosine to isocyanate, which was directly reacted with the ortho-nitrobenzyl (ONB) alcohol (4c) as the photo-cleavable group or the red-shifted derivatives 3,4-dimethoxy-2-nitrobenzyl (DMNB), 6-nitropiperonyl (NP), or 6-nitropiperonyl-methyl (NPM) alcohol (4a-d, Supplementary Fig. 22). During workup in THF with aqueous ammonia, the TMS groups were removed to obtain the photocaged guanosines (5a-d). The photocaged guanosines were then monophosphorylated at the 5′-OH to give 6a-b, methylated to 7a-b and coupled to guanosine 5′-diphosphate imidazolide prepared from GDP as previously described39.
Figure 2. A general strategy for synthesizing cap analogues with photo-cleavable groups (FlashCaps) for triggering translation by light.

The self-immolative carbamate linkage ensures efficient light-mediated release of cap 0. For details see the extended data (Extended Data Fig. 1).
We measured the absorption spectra of the synthesized guanosines with photo-cleavable groups at the N2 position (5a-d). ONB guanosine (5c) showed only low absorbance above 300 nm, DMNB guanosine (5a) showed an absorption maximum at 350 nm, NP-(5d) and NPM (5b) guanosine were slightly red-shifted with a maximum at 360 nm (Fig. 3a), in line with literature on the respective photo-cleavable groups20,40,41.
Figure 3. Characterization of FlashCaps.

a) Absorption spectra of indicated guanosines and cap analogues. b) Time- and wavelength-dependent photo-deprotection for indicated caged guanosines. c) Stability of FlashCaps in cell lysate at 37 °C in comparison to cap 0. d-e) Affinity measurements of FlashCaps before and after irradiation with eIF4E (Cy5-labelled) using microscale thermophoresis (MST). The average of three independent measurements is shown. f) Stability of FlashCaps against enzymatic degradation by DcpS with or without irradiation at 15 min (dashed line) (365 nm, 30 s). g) Stability of irradiatedFlashCaps against enzymatic degradation by DcpS (365 nm, 30 s) in comparison to cap 0. Data of n=3 independent experiments are shown as mean values +/- SD.
To choose the most suitable photo-cleavable group for biological applications, we irradiated 5a-c in aqueous solution at neutral pH and analysed their decrease as well as formation of the native guanosine using HPLC and LC-MS (Fig. 3b, Extendet Data Fig. 1 Supplementary Fig.3, 8, 10). Time-dependent analyses revealed that at 365 nm (LED, 140 mW/cm2), short irradiation (5-15 s) was sufficient to remove the photo-cleavable group in 10 μL of a 500 μM solution of 5a, b, d and release the free guanosine (Fig. 3b, Extended Data Fig. 1). At 405 nm, the NP, NPM and DMNB groups were efficiently removed after 60 s, more efficiently than the ONB group (Figure 3b, Extended Data Fig. 1). At 420 nm, the NPM group was completely removed after 120 s (Figure 3b, Extended Data Fig. 1). We therefore chose DMNB and NPM groups for further studies and synthesized the respective cap 0 analogs. The resulting FlashCaps contain the DMNB (1) or the NPM (2) group at the N2 position connected via a carbamate functionality (Fig. 2). Their absorption spectra above 300 nm and their uncaging kinetics were similar to the respective photocaged guanosines (5a,b) (Fig. 3a, Extended Data Fig. 1) and formation of cap 0 was confirmed (Supplementary Fig. 7-10). We also assessed the biological stability of the carbamate linkage by incubating cap 0 (0) or FlashCaps (1, 2) in cell lysate followed by HPLC analysis (Fig. 3c, Supplementary Fig. 4). FlashCaps exhibited high stabilities over 30 h, similar to the cap 0, suggesting that the carbamate linkage is not the primary point of degradation in lysate.
Next, we evaluated how the photo-cleavable groups affect interaction of the 5′ cap with eIF4E. Binding measurements of FlashCaps and Cy5-labelled eIF4E using microscale thermophoresis (MST) did not result in a binding curve in case of the photocaged caps (1, 2) (Fig. 3d,e). Under identical conditions, a Kd value of 0.3 μM was determined for cap 0 (Supplementary Fig. 5), in line with the literature36,42. Importantly, after light-induced removal of the photocaging groups from 1 or 2, the characteristic binding curve and a Kd value in a similar range to cap 0 was obtained (Fig. 3d,e), indicating efficient formation of cap 0 (Supplementary Table 2).
We also investigated how the photo-cleavable groups affected interactions with cap-modifying enzymes. DcpS is a pyrophosphatase hydrolysing the cap structure to m7GMP and GDP in eukaryotic cells (Fig. 3f)8. Similar to the results with eIF4E, DcpS (H277N) – a binding but non-cleaving variant of the decapping enzyme – interacted with cap 0 but not with FlashCaps (Supplementary Fig. 6). However, if FlashCaps were briefly irradiated before the assay was performed, the Kd value of DcpS variant was in the same range as for cap 0 (Supplementary Fig. 6), indicating light-induced liberation of functional cap 0.
We also tested whether the photo-cleavable groups would affect the enzymatic degradation of cap structures (Fig. 3f,g). Catalytically active DcpS-WT rapidly cleaved cap 0 into m7GMP and GDP, resulting in >50 % degradation within 15 min (Fig. 3g)8. In contrast, the FlashCaps 1 and 2 remained almost completely intact during that time (~98 % undigested cap), demonstrating that the photo-cleavable groups abrogate enzymatic cleavage of FlashCaps (Fig. 3f). As expected, the DcpS-mediated cleavage of FlashCaps was triggered in situ by irradiation with light (365 nm, 30 s), confirming that light-mediated release of the photo-cleavable group renders the reconstituted cap 0 readily available to enzymatic conversion (Fig. 3f).
Taken together, these data demonstrate that FlashCaps efficiently impede the interaction with cap-binding proteins and cap-degrading enzymes and that irradiation by light releases fully functional cap 0 that is readily recognized by cap-binding partners in vitro.
Next, we were interested whether FlashCaps are suitable for the preparation of long mRNAs containing a photocaged 5ʹ cap (FlashCap-mRNAs) using standard molecular biology methods. In vitro transcription (IVT) using phage T7 RNA polymerase and synthetic cap analogues is routinely used to produce capped mRNAs for biological studies43 and therapeutic applications44. The cap analogue is incorporated as the first G by transcriptional priming, yielding capped and uncapped RNA. The latter can be removed by enzymatic treatment with polyphosphatase and XRN116. Comparative evaluation of IVT with FlashCaps or cap 0 revealed that all tested 5′ caps yielded intact mRNAs (Fig. 4a). The yield and capping efficiency in the presence of 1 or 2 were slightly lower but in the same range as for cap 0, according to our analysis of four different mRNAs (Fig. 4b). These data show that transcriptional priming with FlashCaps is efficient and that 1 and 2 can be routinely used for IVT with T7 polymerase to produce long FlashCap-mRNAs with yields comparable to cap 0.
Figure 4. Characterisation of cap-caged mRNA.

a) PAGE (Polyacrylamide gel electrophoresis) analysis of mRNA from in vitro transcription (IVT) with commercial caps (0, AG) or FlashCaps (1, 2). Shown is one representative gel from n=3 independent replicates. b) Yield and capping efficiency of indicated mRNAs from IVT (50 µL) in the presence of FlashCaps. c) Stability of differently capped (0, 1, 2) mRNA against Dcp1/2 before/after irradiation. Samples were either incubated with enzyme (Dcp1/2) or without (none). Shown is one representative gel from n=3 independent replicates. d) In vitro translation of FlashCap-FLuc-mRNA before/after irradiation. Data of n=3 independent experiments are shown as mean values +/- SD. Statistical significance was determined by two-tailed students t-test. Significance levels were defined as p<0.05:*, p<0.01:**, p<0.001:***. P value for 2(+) versus 2(-) is 9.08 × 10−4. P value for 1(+) versus 1(-) is 2.3 × 10−5. e) Same as d) but with modified nucleotides (m1Ψ, m5C). Data of n=3 independent experiments are shown as mean values +/- SD. Statistical significance was determined by two-tailed students t-test. Significance levels were defined as p<0.05:*, p<0.01:**, p<0.001:***. P value for 2(+) versus 2(-) is 2.69 × 10−4. P value for 1(+) versus 1(-) is 4.3 × 10−5. Abbreviations: nt: nucleotides; irr: irradiated (365 nm, 30 s), M = Marker, AG: ApppG cap, FLuc: Firefly Luciferase, RLuc: Renilla Luciferase, GLuc: Gaussia Luciferase, eGFP: enhanced green fluorescent protein.
We then probed the interaction of long FlashCap-mRNAs with cap-binding proteins or cap-modifying enzymes. In the major mRNA turnover pathway, the decapping enzyme Dcp1/2 cleaves mRNA to release the 5′ monophosphorylated mRNA, which is degraded by the exoribonuclease XRN19,10. We tested this cap-dependent decay in vitro by treating cap 0-mRNA and FlashCap-mRNA with Dcp1/2 followed by XRN1 digestion (Fig. 4c). This treatment completely degraded cap 0-mRNA, whereas FlashCap-mRNAs with 1 or 2 remained intact (Fig. 4c). When FlashCap-mRNAs were irradiated prior to the enzymatic treatment, they became susceptible to enzymatic degradation, indicating light-dependent release of the free cap 0, which is recognized by Dcp1/2. In control reactions, which were irradiated but not treated with the enzymes, mRNAs with cap 0 or FlashCaps remained intact, confirming that irradiation alone does not degrade long mRNAs (Fig. 4c).
As FlashCaps abrogate eIF4E binding, which is the rate limiting step for translation initiation, we were curious how FlashCap-mRNA would impact translation. We therefore tested in vitro translation (IVTL) of luciferase-mRNAs with cap 0 and FlashCaps using rabbit reticulocyte lysate (RRL). To our delight, the translation of FlashCap-mRNAs was drastically reduced (Fig. 4d), in line with results from our eIF4E-binding studies (Fig. 3d, e). FlashCap-RLuc-mRNAs with 1 or 2 exhibited only 2–4% of luciferase activity relative to cap 0-mRNA (Fig. 4d). However, if the FlashCap-mRNAs were irradiated, translation was increased by 15-20-fold, reaching 41±2% (1) (SD, n = 3) or 59±11% (2) (SD, n = 3) relative to the native cap 0. Under the same irradiation conditions, the IVTL of cap 0-mRNAs was only slightly reduced (to 91±5%; SD, n = 3) and the mRNAs stayed intact (Fig. 4a,d).
Taken together, these data demonstrate that FlashCap-mRNAs are translationally muted and efficiently activated by brief irradiation with light. The released mRNAs are intact, functional and contain a 5ʹ cap 0, but no sequence changes or remaining chemical modifications.
Next, we investigated the translation of FlashCap-mRNAs in cultured mammalian cells using Gaussia luciferase (GLuc) or enhanced green fluorescent protein (eGFP) as secreted or intracellular reporter. Luciferase activity for HeLa cells transfected with FlashCap-mRNAs or controls was normalisednormalised to cap-dependent translation of cap 0-mRNA. The cap-dependent translation of FlashCap-mRNAs with 1 or 2 was reduced to 6±1% (SEM, n = 3) and 2±2% (SEM, n = 3), respectively (Fig. 5a). Half of the cell samples were briefly irradiated 6 h after transfection. Irradiation strikingly increased the luciferase signal of cells transfected with FlashCap-mRNA, resulting in 72±8% (SEM, n = 3) in the case of 1 and 54±4% (SEM, n = 3) in the case of 2. This corresponds to a remarkable 12–27-fold irradiation-dependent increase in translation. A 32-fold increase was observed, when FlashCap-mRNA with 2 was irradiated before transfection. The irradiation itself only slightly decreased the translation (77±7%; SEM, n = 3) in HeLa cells, as shown by controls with cap0-mRNA. Of note, the absolute amount of cap-dependent translation triggered by light almost reaches the level of irradiated cells transfected with control mRNA (72±8%, SEM, n = 3), supporting the notion that intracellular uncaging is efficient and fully functional mRNA is generated (Fig. 5a). Taken together, these data demonstrate that irradiation efficiently releases cap 0-mRNA and triggers translation in living cells transfected with FlashCap-mRNAs, without compromising cell viability and mRNA integrity (Fig. 3a, Supplementary Fig.13).
Figure 5. Light-induced translation in cells.

a) Relative Luciferase activity from HeLa cells transfected with differently capped GLuc-mRNAs. The mRNA was capped with the indicated cap analogue. Data of n=3 independent experiments are shown as mean values +/- SEM. Statistical significance was determined by two-tailed students t-test. Significance levels were defined as p<0.05:*, p<0.01:**, p<0.001:***. P value for 1(+ irr. cells, - irr. mRNA) versus 1(- irr. cells, - irr. mRNA) is 1.26 × 10−3. P value for 2(+ irr. cells, - irr. mRNA) versus 2(- irr. cells, - irr. mRNA) is 3.53 × 10−4. P value for 2(- irr. cells, + irr. mRNA) versus 2(- irr. cells, - irr. mRNA) is 1.52 × 10−4. b) Confocal laser scanning microscopy images of HeLa cells co-transfected with differently capped eGFP-mRNAs and cap 0-mScarlet-I mRNA. mRNAs contain m5C and m1Ψ. AG: ApppG-capped mRNA represents cap-independent translation. (0): m7GpppG capped eGFP-mRNA. 2: NPM cap-eGFP mRNA, either non-irradiated, irradiated in cells or irradiated before transfection (irradiated mRNA). The top row shows the eGFP fluorescence, the bottom row the mScarlet-I fluorescence. Scale bars are 50 µm. For all images background substraction was performed with ImageJ (30 pixels). Shown is one representative set of n=3 independent experiments. c) Flow cytometry of HeLa cells transfected with differently capped mRNAs (with cap analogues 0, 1, 2 or ApppG (AG)). Untransfected cells are set as gate for eGFP negative cells. Irradiation is indicated by LED icon. Shown is one representative measurement of n=3 independent experiments.
In current mRNA-based therapeutics, modified nucleosides are widely used to increase translation33 and reduce immunogenicity34,45. To test whether FlashCaps are compatible with such modifications, we produced FlashCap-mRNAs containing 5-methylcytosine (m5C) and N1-methyl-pseudouridine (m1Ψ). As expected, these internal RNA modifications increased the amount of protein produced in all cases (Supplementary Fig. 14)33. Normalised to control-mRNAs containing the same modifications, the light-dependent turn-on effect of FlashCap-mRNAs remained in the same range both in vitro (Fig. 4e) and in cells (Supplementary Fig. 15). Light-induced translation of FlashCap-mRNAs was also achieved in HEK293T cells demonstrating their functionality in different human cell lines (Extended Data Fig. 2, Supplementary Fig. 16).
To assess the effect of FlashCaps and light on translation for a different mRNA and using a different assay, we co-transfected HeLa cells with differently capped eGFP-mRNAs and cap 0-mScarlet-I-mRNAs as internal reference. Imaging by confocal microscopy revealed that a green fluorescent signal was barely detectable when FlashCap-eGFP-mRNA with 2 was used (Fig. 5b). Control cells transfected with cap0-eGFP-mRNA showed bright fluorescence under the same conditions. However, if the cells transfected with FlashCap-mRNA were irradiated, strong green fluorescence was visible, comparable to cells transfected with cap 0-mRNA (Fig. 5b). Similarly, irradiation of FlashCap-mRNA before transfection strongly increased the fluorescence. Quantification of the microscopy images confirmed a significant increase, supporting the data obtained by the luminescence assay (Supplementary Fig. 19). Furthermore, we tested a transcript coding for Rheb, a GTP-binding protein that is ubiquitously expressed in humans. Western Blot confirmed that FlashCap-Rheb-mRNA was muted, but efficiently translated upon irradiation (Extended Data Fig. 4), indicating that FlashCaps are compatible with biologically relevant mRNAs.
To analyze the effect of irradiation on translation also on the single-cell level, we performed flow cytometry of HeLa cells transfected with differently capped eGFP-mRNAs (Fig. 5c). Direct comparison revealed a marked increase in eGFP-positive cells, when FlashCap-mRNA-containing cells had been irradiated. FlashCap-mRNAs with 1 or 2 then led to 36.0 % or 41.4 %, eGFP-positive-cells. These values are close to the 48.6% observed for the positive control (cap 0-mRNA) (Fig. 5c). Without irradiation, FlashCap-mRNAs led to a significantly lower fraction of eGFP-positive cells (18-19%), albeit higher than the negative control (4%). This can be attributed to partial uncaging during this long experiment, as the same FlashCap-eGFP-mRNA shows no significant background in CLSM images (Fig 5b), nor in Western blots (Extended Data Fig. 4). The histograms unambiguously show that the eGFP intensity of the irradiated samples is much higher compared to non-irradiated samples. Taken together, the flow cytometry data show on a single-cell level that the eGFP fluorescence intensity is increased for FlashCap-mRNAs in response to irradiation (Fig. 5c). The data independently confirm the findings from luminescence, Western blot, and microscopy analysis, showing that irradiation of FlashCap-mRNAs highly increases translation of a variety of reporter mRNAs.
A key feature of light-triggered processes is the exquisite and facile spatio-temporal control. Using a confocal laser scanning microscope, we tested whether brief irradiation of a predefined circle with a diameter of 120 μm using the 405 nm laser would activate translation in a subset of cells. Indeed, we observed that cells transfected with FlashCap-eGFP-mRNA containing 2 developed green fluorescence exclusively in the circled area (Fig. 6a). These data show that FlashCap-mRNAs enable control of translation in a subset of cells. Spatial control of translation in the μm-scale can be readily achieved using a commercial CLSM setup.
Figure 6. Spatio-temporal control of translation, stability, and immune response of FlashCap-mRNAs in cells.

a) Irradiation of circled area in a confocal laser scanning microscope and analysis of fluorescence of HeLa cells transfected with FlashCap 2-eGFP-mRNA. Nuclei are stained by DAPI (blue). Scale bars are 100 µm or 50 µm (inset). Individual colour channels were adjusted. Shown is one representative image from n=3 independent experiments. The microscopy images were hyperstacked and the background subtracted (30 pixels) with ImageJ. b) Stability of FlashCap-mRNAs. RTqPCR data showing the relative RLuc mRNA level at 4 h or 10 h post transfection in HeLa cells. The 4 h time point is used for normalization and was set as 100 %. Data of n=3 independent experiments are shown as mean values +/- SEM. c) Immune response of FlashCap-mRNAs. FLuc activity of four different HEK-NF-ĸB cell lines (Null, TLR3, TLR7, TLR8) transfected with differently capped RLuc-mRNAs (either cap 0 or FlashCap1 or 2). TLR3, TLR7 and TLR8 indicates the overexpression of the respective Toll-Like-Receptor (TLR) in that cell line. Data are normalized to the cap 0-mRNA without irradiation. Data of n=4 independent experiments are shown as mean values +/- SEM.
mRNA therapeutics have recently gained enormous interest. For the use of modified mRNAs in vivo/in humans it is important to estimate the effects on the stability of the mRNA as well as on the elicited immune response. Previous studies reported that untranslated mRNAs are subjected to degradation as part of a quality control mechanism46. To assess whether translationally muted FlashCap-mRNAs are prone to degradation, we determined the stability of mRNAs in cells in comparison to cap 0-mRNAs (Fig. 6b). Using RTqPCR, we compared the amount of differently capped mRNAs at 4 h and 10 h after transfection. We observed similar levels of remaining mRNA 10 h after transfection, suggesting that the half-life of mRNAs is not affected by the photo-cleavable group at the 5′ cap (Fig. 6b, Supplementary Fig. 21).
To assess the effect of FlashCaps on immune response, we used reporter HEK-NF-ĸB cell lines overexpressing an NF-ĸB driven Firefly Luciferase and different Toll-like receptors (TLR).47 The control cell line (Null) has no TLR overexpressed and provides a measure for the activation of endogenously expressed pathogen recognition receptors. The FlashCap-mRNAs did not exhibit a significant increase in response to TLR3, TLR7, TLR8 nor to the control cell line in comparison to cap 0-mRNA (Fig. 6c). This was observed both for the unirradiated and the irradiated form. These data suggest that the application and activation of FlashCap-mRNAs can be expected to elicit an immune response similar to cap 0-mRNAs and may thus prove suitable for application in therapeutic mRNAs.
Conclusions
With the approval of mRNAs as therapeutic modality, the number of studies on mRNA aiming to improve the technology and addressing other diseases can be expected to rise, both in the field of basic research as well as in pre-clinical and clinical studies. However, to date, no strategy exists to efficiently time the expression of the administered mRNA, nor to control the delivery and uptake into certain tissues without alterations remaining in the mRNA. Even in cell culture the administration/liberation of exogenous mRNA currently cannot be efficiently controlled in space and time.
We developed a technique to control the translation of any given mRNA by light. FlashCaps are 5’ cap analogues containing a single photocaging group connected via a self-immolative carbamate linkage, leading to fast and efficient liberation of the natural cap 0 structure as demonstrated by multiple assays in vitro. FlashCaps are compatible with common molecular biology techniques. They are simply added instead of the synthetic 5’ cap analogue to the in vitro transcription to make any mRNA of interest with efficiencies similar to the cap 0-mRNA. The resulting FlashCap-mRNAs are (1) translationally muted in vitro and in cells, (2) contain only a single photo-cleavable group, (3) release native cap 0-mRNA, (4) do not require changes in sequence or permanent chemical alterations and (5) are not immunogenic. We demonstrate the functionality of FlashCap-mRNAs in two different cell lines and for light-activated translation into both intracellular (eGFP, RLuc, Rheb) and secreted (GLuc) proteins. The irradiation conditions required to release cap 0-mRNA are compatible with cell viability and translation, and the photo-cleavable groups have even proven compatible with animal models in previous studies.48,49 An up to 32-fold light-induced increase of translation was observed in HeLa cells. We also confirmed that translationally muted FlashCap-mRNAs are not preferentially degraded and do not elicit an increased immune response compared to cap 0-mRNAs. FlashCaps are therefore a highly efficient and readilyapplicable solution to make mRNA studies controllable by light, without requiring new production steps and without introducing permanent artefacts.
Methods
Absorbance spectra analysis
The analysis of the absorbance properties of the photocaged guanosines was performed using a quartz cuvette (Hellma) together with the FP–8500 Fluorescence Spectrometer (Jasco). The respective guanosines were dissolved in water at a final concentration of 100 μM. For the absorbance measurement 20 μL of the solution were further diluted in water to give a final volume of 100 μL (20 μM), which were transferred into the cuvette followed by the absorbance measurement. The values were normalised to the highest measured value of each measurement.
Irradiation of samples
LEDs (LED Engin) were used to irradiate mRNA samples, guanosines, cells and cap analogues. The UV–A–LED (λmax = 365 nm) and the blue light LEDs (λmax = 405 nm, λmax = 420 nm) were operated with 5 V and 600 mA input power, the respective output power is shown in Supplementary Figure 1.
Irradiation was performed in a custom-made LED setup at 23 °C. The samples were irradiated in a PCR-tube or a cell culture dish (Supplementary Fig. 2) unless stated otherwise. Samples were irradiated at 365 nm (142 mW/cm2) for 30 seconds, 405 nm (142 mW/cm2) for 60 seconds or 420 nm (52 mW/cm2) for 120 seconds, unless otherwise noted.
Guanosine and dinucleotide irradiation studies
The respective guanosines or cap analogues were dissolved in ddH2O if possible (if needed, organic solvents were added to increase solubility) to give a solution with a final concentration of 500 μM. The solution (10 μL) was transferred into a PCR-tube and irradiated as described above. Subsequently, the solution was analysed by HPLC.
HPLC analysis
HPLC analysis and purification of cap analogues were performed on an Agilent1260 Infinity HPLC equipped with a diode array detector (DAD) (190–640 nm) using a Nucleodur®C18 Pyramid reversed–phase column (5 μm, 125 x 4mm) from Macherey–Nagel. Elution was done at a flow rate of 1 mL/min applying a linear gradient from buffer A (50 mM ammonium acetate, pH=6.0) to buffer B (1:1 buffer A : acetonitrile). If other conditions were used, this is described in the respective section.
MST measurements
Microscale thermophoresis (MST) measurements were performed on a Monolith NT.115 series instrument (NanoTemper). Prior to thermophoresis measurements proteins were labeled by incubation with Cy5-NHS (Lumiprobe) for 30 min at room temperature. Unreacted dye was separated from the protein using PD SpinTrap™ G-25 gel filtration columns (GE Healthcare) according to the manufacturer’s protocol. Serial dilutions of the cap analogues (starting from 200 μM of cap analog) in MST reaction buffer (20 mM HEPES, 50 mMKCl, 0.2 mM EDTA, 0.01% Triton-X, 700 μM mercaptoethanol, 0.01% Tween-20, pH = 8) were prepared and mixed with an equal volume of the labeled protein (~50 nM). The mixture was filled into premium coated capillaries (4 μL) and directly measured. MST power was set to 30–40%, LED power was set to 20% Red (Exc.: 625nm, Em.: 680 nm). Thermophoresis measurements were performed with the following settings: fluorescence before (5 s), MST on (30 s), fluorescence after (3 s). The capillaries were measured three times in direct succession as technical replicates. MST data were normalised to baseline differences and Kd values were calculated using nonlinear regression assuming a Hill coefficient of 1.0 (GraphPad Prism). MST is known to produce occasional outliers. This was handled as follows: 16 data points were measured per binding curve and at least 12 data point were used for each fit.
yDcpS hydrolysis assay
The hydrolytic activity of yDcpS (New England Biolabs) was assayed using the following experimental conditions: 50 mM Tris HCl containing 1 mM Mg(Ac)2, 30 mM (NH4)2SO4 and 1 mM DTT (final pH 8.0) at 37 °C. Together with the respective cap analogue, an internal standard (either adenine monophosphate or 4,5,7-trihydroxy-3-phenyl-coumarin) with a final concentration of 200 mM was added. At last, 20 U of yDcpS were added. The hydrolysis process was started by incubation at 37 °C. At 0, 5, 10, 15, 30, 45, 60 min of the hydrolysis, 10 μL aliquots of the reaction mixture were withdrawn and the reaction was stopped by heat inactivation of the enzyme (10 min at 90 °C). The samples were then subjected to analytical HPLC and analysed at 260 nm. Hydrolysis products were identified by comparison of their retention times with those of reference standards.
Expression and purification of MTAN, LuxS, hTgs, hDcpS and eIF4E
The enzymes MTAN, LuxS, hDcpS H277N, eIF4E and hTgs were produced and purified as previously described.16,36,50,51
In vitro transcription
The DNA template required for the in vitro transcription was synthesized by PCR, in which the DNA sequence coding for eGFP, Firefly luciferase (FLuc), Gaussia luciferase (GLuc), Renilla luciferase (RLuc) were amplified from pMRNA–vectors containing the respective sequence. After purification (NucleoSpin Gel and PCR Clean up, Macherey–Nagel), the resulting linear dsDNA was used as template (200 ng). The run-off template is an alternative to the PCR–DNA template that was used for GLuc, mScarlet-I and eGFP mRNAs used in cell studies and fluorescence microscopy. Plasmid DNA (3 μg) was incubated with 1x FastDigest buffer (Thermo Fisher) and 3 μL PacI FastDigest enzyme for 10 min at 37 °C, followed by inactivation at 65 °C for 10 min. Subsequently, the ends were dephosphorylated by adding 3 μL FastAP and incubation at 37 °C for 15 min and inactivation at 65 °C for 5 min. The product was purified using the NucleoSpin® Gel and PCR Clean–up kit (Macherey–Nagel). The concentration was measured at 260 nm with the Tecan Infinite© M1000 PRO. The resulting linear dsDNA was used as template (400 ng). The in vitro transcription was performed with T7 polymerase (Thermo Scientific) in transcription buffer (40 mMTris/HCl, 25 mM NaCl, 8 mM MgCl2, 2 mM spermidine(HCl)3) by adding either a A/C/UTP (0.5 mM) mix or A/m5C/m1ΨTP mix (0.5 mM), GTP (0.25 mM), the respective cap analogue (1 mM), T7 RNA polymerase (50 U) (Thermo Scientific) and pyrophosphatase (0.1 U) (Thermo Scientific) for 4 h at 37°C. After the reaction, the DNA template was digested in presence of 2 U DNase I for 1 h at 37°C and then mRNAs were purified using the RNA Clean & Concentrator™–5 Kit (Zymo Research). To digest non-capped RNAs, 10 U of the RNA 5’–polyphosphatase (Epicentre) as well as the supplied reaction buffer were added to purified mRNAs. After an incubation period of 30 min at 37 °C, 0.5 U of the 5′ – 3′ exoribonuclease XRN1 (NEB) and MgCl2 (5 mM) were added. The reaction mixture was incubated for 60 min at 37 °C. Subsequently, capped mRNAs were purified using the RNA Clean & Concentrator™ –5 Kit (Zymo Research).
In vitro Luminescence assay
For in vitro translation the Retic Lysate IVT™ kit (Invitrogen), a eukaryotic cell–free protein expression system, was used. In a total volume of 15 μL and containing 40 ng of the (as indicated capped) FLuc–mRNA, 50 μM L-methionine and 150 mM potassium acetate. Samples were mixed with 8.5 μL of the reticulocyte lysate and incubated for 90 min at 30 °C. Afterwards, 2 μL of the respective translation mix were further used in a luminescence assay. The translation efficiencies of the differently capped FLuc–mRNAs were measured using a luciferase assay based on the Beetle–juice Luciferase Assay Firefly (pjk). Luciferase activity was determined after adding 50 μL freshly prepared substrate solution to the translation mixture. Luminescence was assessed using a Tecan infinite® M1000 PRO microplate reader with an integration time of 3 s. Differently capped mRNAs were used. ApppG–capped mRNA represents cap-independent translation and was subtracted as background from the other samples. All values were normalised to m7GpppG capped mRNA.
Mammalian cell culture
HeLa cells (Merck) were cultured in MEM Earle’s media (PAN) supplemented with l-glutamine (2 mM, PAN), non-essential amino acids (1%, PAN), penicillin and streptomycin (1%, PAN), and fetal calf serum (FCS, 10%, PAN) under standard conditions (5% CO2, 37 °C). HEK 293T cells (DSMZ) were cultured in DMEM (PAN) supplemented with L-glutamine (2 mM, PAN), penicillin and streptomycin (1%, PAN), and fetal calf serum (FCS, 10%, PAN) under standard conditions (5% CO2, 37 °C).
HEK-NF-κB cells (from TRON in Mainz, Germany) were cultured under standard conditions (5 % CO2, 37 °C) in Dulbecco´s Modified Eagle´s medium (DMEM) supplemented with FCS (10 %), HEPES Puffer (1 %), l-glutamine (1 %), non-essential amino acids (1 %) and sodium pyruvate (1 %). For selection, the following antibiotics were added to the culture of the HEK-NF-κB-Null, HEK-NF-κB-TLR7 and HEK-NF-κB-TLR8 cell line: Blasticidin (10 μg/mL), Zeocin (100 μg/mL) and Geneticin (G418) (250 μg/mL). The HEK-NF-ĸB-TLR3 cell line was cultured in the absence of Geneticin. All of the cell lines overexpress an NF-κB driven Firefly luciferase, which allows the detection of NF-κB production in a luminescence assay and can be used as an indicator for the induction of an immune response.47 Additionally, the cell lines HEK-NF-κB-TLR3, HEK-NF-κB-TLR7 and HEK-NF-κB-TLR8 overexpress the respective Toll-like receptor (TLR).
Stability assay of 5′ caps in cell lysate
For preparation of HeLa cell lysate, HeLa cells were cultured as mentioned above. 24 h prior to cell lysis, 3 × 106 cells were seeded on a Petri dish (90 mm). The cells were harvested and pelleted by centrifugation. The cell pellets were stored at –80 °C. For cell lysis, the medium was removed and the cells were washed with 1× PBS, then lysed with CelLytic™ M reagent (1.5 mL, Sigma Aldrich) according to the manufacturer's instructions and stored at –80 °C. The lysis mixture was centrifuged (11,000 rpm, 3 min, 4 °C) and the supernatant was used for the cell lysate stability assay. To the cell lysate the respective cap analogue (500 μM) and 4,5,7-trihydroxy-3-phenyl-coumarin (100 μM) as internal standard were added and incubated for different periods of time (0 h, 0.5 h, 1 h, 5 h, 18 h, 30 h) at 37 °C. The samples were analysed by HPLC.
MTT Assay
HeLa cells (Merck) were cultured as mentioned above. One day prior to transfection cells were seeded in a 96-well plate (30,000 cells/well) and cultured in minimal essential medium (MEM) with antibiotics. The cells were transfected with mRNA (100 ng) in Opti-MEM (10 μL) using Lipofectamine™ MessengerMAX™ Transfection Reagent (0.3 μL) in Opti-MEM (9.7 μL). The cells were incubated with the mRNA/ Lipofectamine™ MessengerMAX™ mixture for 6 hours at 37 °C in a total volume of 100 μL. The samples were irradiated under the indicated conditions. Subsequently, the cell media with the transfection agent was replaced by fresh media and the cells were incubated overnight at 37 °C in media. 24 h post transfection the MTT solution (16.5 mg MTT in 3.3 mL PBS) was added to the 96-well plate (12.5 μL/well). After 4 h of incubation at 37°C the supernatant was removed and 0.04 M HCl in isopropanol was added to the wells. After incubation for 1.5 h at room temperature, 100 μL of the supernatant was placed in a new 96-well plate and absorption at 550 nm was measured using the Tecan Infinite© M1000 PRO plate reader.
In-cell luminescence assay
HeLa or HEK293T cells were cultured as mentioned above. One day prior to transfection, cells were seeded in a 96-well plate (30,000 cells/well) and cultured in minimal essential medium (MEM) with antibiotics. The cell were transfected with mRNA (100 ng) in Opti-MEM (10 μL) using Lipofectamine™ MessengerMAX™ Transfection Reagent (0.3 μL) in Opti-MEM (9.6 μL). The cells were incubated with the mRNA/Lipofectamine™ MessengerMAX™ mixture for 6 h at 37 °C in a total volume of 100 μL. The samples were irradiated with 365 nm for 30 seconds if not stated otherwise. Subsequently, the cell media with the transfection agent was replaced with fresh one and the cells were incubated overnight at 37 °C in media. At 24 h post transfection the supernatant was collected. To perform the luminescence measurement, the Gaussia-Juice Luciferase Assay-Kit (PJK GmbH) was used. The supernatant of the previously prepared samples was transferred to a 96-well plate (5 μL supernatant per well). Afterwards, 50 μL of a reaction mixture (PJK Reconstruction buffer and Coelenterazine) were added to the wells and the luminescence activity was measured using a Tecan Infinite© M1000 PRO plate reader. The activity in relative light units (RLU) was determined with an integration time of 3 s. Differently capped mRNAs were used. ApppG-capped mRNA represents cap-independent translation and was subtracted as background from the other samples. All values were normalised to m7GpppG capped mRNA.
Confocal laser scanning microscopy (CLSM)
For microscopic imaging, HeLa cells were cultured as mentioned above. One day before transfection, 2×105 cells were seeded on glass coverslips in a 12-well plate in 1 mL medium (indicated in cell culture section). Cells were transfected using 1.5 μL Lipofectamine™ MessengerMAX™ (Invitrogen) in Opti-MEM (48.5 μL) and eGFP-mRNA (containing m5C and m1Ψ) (1μg) in Opti-MEM (50 μL). In case of mScarlet-I/eGFP cotransfection a total amount of 1 μg mRNA (eGFP 800 ng, mScarlet-I 200 ng) was used. For confocal microscopy the runoff plasmid of pRNA2-(A)128 (Addgene) with eGFP or mScarlet-I was used as template for in vitro transcription. 24 h post transfection cells were fixed with 300 μL/well 4 % paraformaldehyde in PBS for 10 min at rt. After washing, the nuclei were stained with DAPI (1:10 in PBS). After washing with PBS and water the coverslips were mounted on microscopy slides using Aqua-Poly/mount (Polysciences). A Leica TCS SP8 confocal laser scanning microscope (CLSM) was used to image fixed cells with a 63x water immersion objective lens (HC PL APO 63x/1.20 W CORR UVIS CS2). Images were captured at a green channel for eGFP fluorescence (λex = 488 nm, λem = 492–558 nm), a red channel for mScarlet-I fluorescence (λex = 568 nm, λem = 583–693 nm), a blue channel for DAPI (λex = 358 nm, λem = 443–510 nm) and at the differential interference correlation (DIC) channel. The objectives used in this study were HC PL APO 63x/1.20 W CORR UVIS CS2 and HC PL FLUOSTAR 10x/0.30 Ph1. Lasers: Diode laser 405 nm (405 nm; 8.3 mW) (Laser power in focus plane with 10x objective). Detectors: Photomultiplier (Hamamatsu R 9624) HyD Detector. For all respective microscopy images hyperstacking and background substraction was performed with ImageJ (30 pixels).
RNA isolation and reverse transcription with quantitative real-time PCR (RTqPCR)
For RTqPCR, HeLa cells (Merck), were cultured as described above. One day prior to transfection, 2×105 cells were seeded in media (1 mL) in a 12-well plate. Cells were transfected using 1.5 μL Lipofectamine™ MessengerMAX™ (Invitrogen) in Opti-MEM (48.5 μL). 1 μg RLuc-mRNA in Opti-MEM (50 μL) was prepared. The cells were incubated with the mRNA/ Lipofectamine™ MessengerMAX™ mixture for 4 h at 37 °C in a total volume of 1 mL. Subsequently, the cell medium with the transfection agent was replaced with fresh medium. The cells were harvested 4 h or 10 h post transfection by adding 500 μL lysis buffer (10 mM Tris HCl (pH 8), 150 mM NaCl, 0.5 mM EDTA, 0.1 % NP40). The RNA was isolated from the cell lysate via phenol/chloroform extraction. The isolated total RNA was incubated with DNase I (2 U) in DNase reaction buffer (1x) in a total volume of 20 μL for 30 min at 37 °C to digest remaining DNA. Addition of EDTA (final concentration 5 mM) and incubation for 2 min at 65 °C was used to inactivate the enzymes. For reverse transcription, 1 × RT buffer, dNTPs (final concentration 0.5 mM) with random hexamer primer (5 μM) and Maxima H Minus Reverse Transcriptase (25 U) were mixed for 10 min at 25 °C followed by 30 min at 50 °C and finally 5 min at 85 °C. The resulting cDNA was diluted 1:3 in ddH2O and 3 μL of the diluted cDNA were added in a 96-well qPCR plate. 17 μL of mastermix, containing forward primer (0.5 μM), reverse primer (0.5 μM) and 1 × iTaq Universal SYBR® Green Supermix (Bio-Rad), were added to the provided cDNA in the 96-well plate (Supplementary Table 1). The following PCR program was conducted: (a) initial denaturation (95 °C for 3 min), (b) denaturation (95 °C for 5 s), (c) elongation (55 °C for 30 s), (d) plate read, (e) 39 × cycle (b-d), (f) melt curve (60 °C—95 °C, 0.5 °C/4 s), (g) plate read. Quantitative real-time PCR measurements were performed on a Bio-Rad CFX96TM Real-Time System with a C1000TM Touch Thermal Cycler. Data analysis was performed with the CFX Manager 3.1 (Bio-Rad).
Flow cytometry
For flow cytometry, HeLa cells were cultured as mentioned above. One day prior to transfection, 2×105 cells were seeded in a 12-well plate in 1 mL media. Cells were transfected using 1.5 μL Lipofectamine™ MessengerMAX™ (Invitrogen) in Opti-MEM (48.5 μL) and 1 μg eGFP mRNA in Opti-MEM (50 μL). The cells were incubated with the mRNA/ Lipofectamine™ MessengerMAX™ mixture for 4 h at 37 °C in a total volume of 1 mL. The samples were irradiated with 365 nm for 30 sec. Subsequently, the cell medium with the transfection agent was replaced with fresh medium and the cells were incubated overnight at 37 °C. At 24 h post transfection the cells were harvested with trypsin/EDTA and washed with PBS. The cell suspension was filtered through a 40 μm filter to avoid cell clumps. The eGFP signal was measured with a flow cytometer (Beckman Coulter Cytomics FC 500). During Flow Cytometry 10,000 cells (total cell count) were measured per sample. Analysis was performed with the CxP Analysis Software.
Detection of immunogenicity in HEK-NF-ĸB cells
HEK-NF-ĸB cells were cultured as mentioned above. One day before transfection, 1.5×105 cells were seeded in media (500 μL) in a 24-well plate. Cells were transfected using Metafectene® Pro (2 μL) (Biontex) in PBS (28 μL) and (non-irradiated or irradiated) RLuc-mRNA (500 ng) in PBS (30 μL). 20 h post transfection the cells were harvested and washed with PBS. The pellets were resuspended in 50 μL PBS and used for the luminescence assay. The luminescence measurement was performed using the Beetle-Juice Luciferase assay Firefly Kit (pjk). The reagents were prepared as suggested by the manufacturer. The 50 μL of cell-suspension were mixed with 50 μL of 2× Lysis Juice. After incubation for 15 min at 37 °C (450 U/min), 20 μL of the cell lysate were transferred to a 96-well plate (in duplicates). Then 50 μL of the freshly prepared Firefly reaction mixture was injected to the well with an acquisition time of 3000 ms. The samples were normalised to the m7GpppG capped-mRNA.
Western Blots
HEK293T and HeLa cells were cultured as mentioned above. One day prior to transfection, 2×105 cells were seeded in a 12-well plate in 1 mL medium. Cells were transfected using 1.5 μL Lipofectamine™ MessengerMAX™ (Invitrogen) in Opti-MEM (48.5 μL) and 1 μg eGFP mRNA in Opti-MEM (50 μL). 4 h post transfection cells were irradiated (142 mW/cm2, 365 nm, 30 s) and transfection medium was replaced with fresh medium. 24 h post transfection the cells were harvested and washed with PBS. The cells were lysed with CelLytic M (Sigma-Aldrich). To determine the protein concentration of the cell lysate, a Bradford assay was performed using BSA calibration standards and a dilution of cell lysate (1:25). 50 μL of sample were incubated (10 min, RT, exclusion of light) with 1× Roti®-Quant (Roth) staining solution (200 μL) and then the extinction at 595 nm was determined. 40 μg of proteins were separated via tris-glycine-PAGE (12 % PAA gel, 120 V, 1.5 h, RT). The proteins were transferred onto a nitrocellulose membrane Roti®-NC (Roth) in a semi-dry transfer buffer with 90 mA for 75 min at RT. To validate protein transfer, a Ponceau S (0.5 % PonceauS + 1 % glacial acetic acid) stain was performed. The membrane was cut into two appropriate pieces for subsequent antibody treatment and washed with 1× PBS + 0.01 % Tween (PBST). Blocking of the membrane was performed in blocking buffer (3 % BSA in PBS) for 1 h at RT, followed by incubation with the respective primary antibodies – anti-eGFP mouse monoclonal antibody (Santa Cruz Biotechnology) and anti-Nucleolin mouse monoclonal antibody (Thermo Fisher Scientific) – overnight at 4°C and three times washing with PBST for 5 min at RT. The membrane pieces were incubated with a HRP-conjugated secondary antibody (polyclonal rabbit anti-mouse immunoglobulins/HRP; Dako Diagnostica GmbH) for 1 h at RT and then washed three times with PBST. For chemiluminescence detection the EZ-ECL Chemiluminescence detection kit (Biological Industries) was used and results were analysed with a Chemo Star Advanced Fluorescence & ECL Imager (Intas).
Decapping assay
The RNA was prepared as mentioned above. 1 μg of capped eGFP-mRNA was mixed with mRNA Decapping Enzyme Reaction Buffer (NEB) (final concentration 1x) in a total volume of 19.7 μL. The mixture was irradiated (except for the control without irradiation). Then either 0.3 μL of mRNA Decapping Enzyme (NEB) or 0.3 μL H2O as negative control were added. After incubation for 30 min. at 37°C, 1 μL XRN1 and 2.5 μL MgCl2 were added to each Eppendorf tube (to the controls as well) and incubated for 1h at 37°C. 2 μL of each sample were loaded on a 7.5 % PAA gel. RiboRuler Low Range (Thermo Fisher) was used as marker.
Remethylation assay
A solution of LuxS (5 μM), MTAN (5 μM), the corresponding cap analogue (400 μM), SAM (6 mM) and hTgs (20 μM) in buffer (5 mM Tris-HCl, 10 mM MgCl2, 5 mM KCl (pH = 8.0)) was incubated at 37 °C. At 0, 5, 15, 30, 60 min of the methylation reaction, 10 μL aliquots of the reaction mixture were withdrawn and the reaction was stopped by heat inactivation of the enzyme (10 min at 90 °C). The samples were then analysed by HPLC monitoring the absorbance at 260 nm. Methylation products were assigned by comparison of their retention times with those of reference standards.
Statistical analysis
For the statistical analysis of the luminescence data an unpaired, parametric, two-tailed t-test was used. When compared with the m7GpppG mRNA an additional Welsh Correction was used p<0.05:*, p<0.01:**, p<0.001:***.
Extended Data
Extended Data Fig. 1. Irradiation studies of photocaged guanosines and FlashCaps.

Photo-cleavage reaction by the example of 5c (a).Illustration of the decrease of caged guanosine (G) after irradiation with 365 nm (b), 405 nm (c) or 420 nm (d)and the decrease of FlashCaps after irradiation with 365 nm (e), 405 nm (f) or 420 nm (g)for variousperiods of time. The samples (500 µM) were analysed via HPLC after irradiation via LED (365 nm (142 mW/cm2), 405 nm (142 mW/cm2); 420 nm (60 mW/cm2)). The percentage of uncaged guanosine or FlashCapwas calculated by integration of the resulting peaks. Data points and error bars denote mean values ± standard deviation of the mean for n=3 independent replicates.
Extended Data Fig. 2. In-cell translation assay showing luciferase activity of FlashCap-GLuc mRNAs normalized to m7GpppG-mRNA.

The samples were either irradiated before transfection (++), in cells (+) or left untreated (-). a, Measured and normalized luminescence values, which were obtained from the cell media of HeLa cells that were previously transfected with differently capped GLuc-mRNA (containing m5C and m1Ψ) either irradiated or not irradiated in a 96 well plate. The P value for 1(+) versus 1(-) is 4.66 × 10−4. The P value for 2(+) versus 2(-) is 5.70 × 10−5. The P value for 2(++) versus 2(-) is 1.16 × 10−5. b, Measured and normalized luminescence values, which were obtained from the cell media of HEK293T cells that were previously transfected with differently capped GLuc-mRNA and either irradiated or not irradiated in a 96 well plate. The P value for 1(+) versus 1(-) is 3.57 × 10−3. The P value for 2(+) versus 2(-) is 1.7 × 10−4. The P value for 2(++) versus 2(-) is 4.3 × 10−4. c, Measured and normalized luminescence values, which were obtained from the cell media of HEK293T cells that were previously transfected with differently capped GLuc mRNA (containing m5C and m1Ψ) and either irradiated or not irradiated in a 96 well plate. The P value for 1(+) versus 1(-) is 9.05 × 10−4. The P value for 2(+) versus 2(-) is 9.32 × 10−4. The P value for 2(++) versus 2(-) is 2.09 × 10−4. Statistical significance was determined by two-tailed t-test. Data and error bars represent average and standard error of the mean of three independent (n=3) cell experiments. Significance-levels weredefinesas p<0.05:*, p<0.01:**, p<0.001:***.
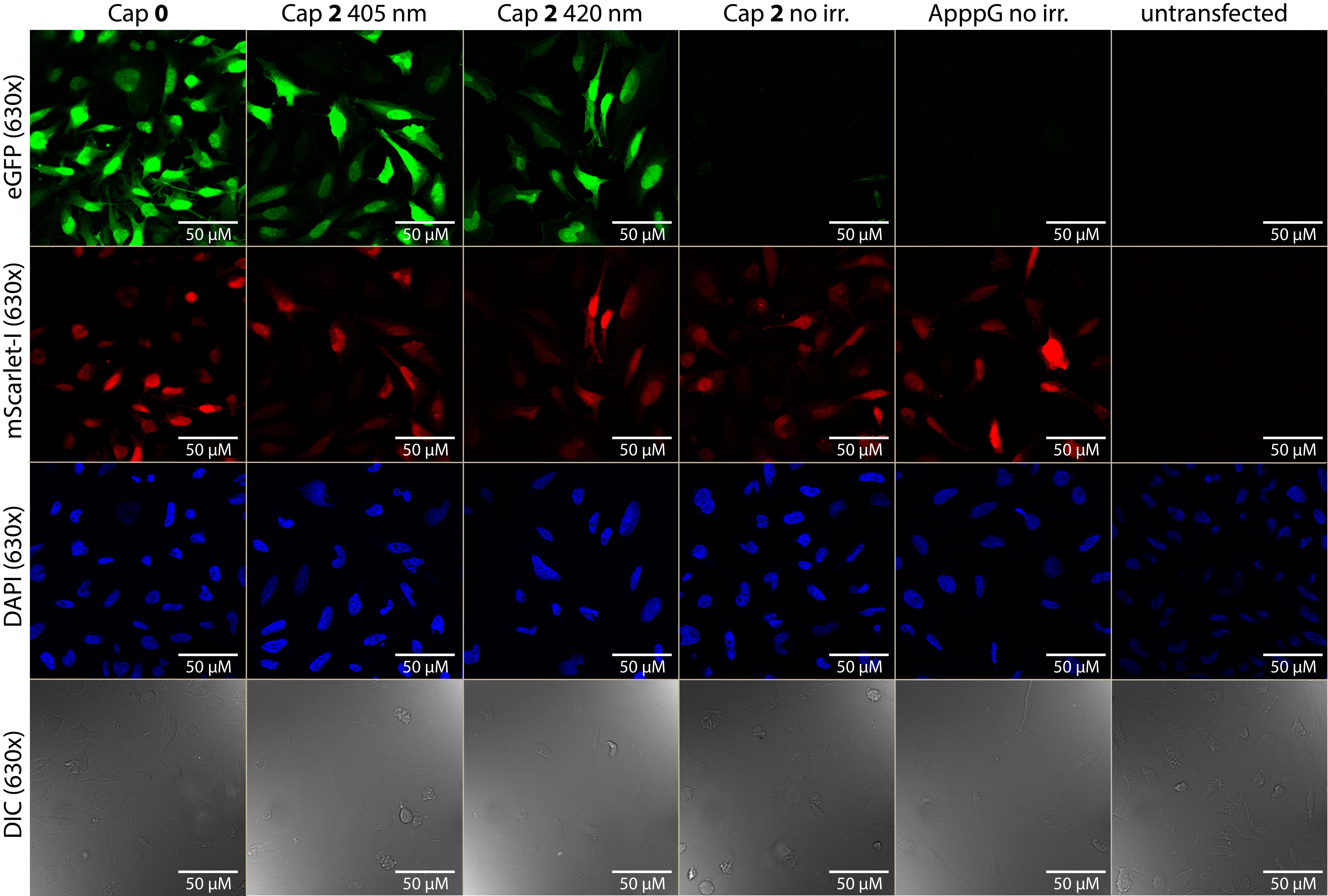
Extended Data Fig. 3. Representative 630x magnification confocal microscopy images of irradiated (405 nm, 420 nm) and non-irradiated HeLa cells transfected with eGFP- and mScarlet-I-mRNA with DAPI staining.

HeLa cells were transfected with differently capped eGFP-mRNA containing m5C and m1Ψ and m7GpppG-capped mScarlet-I-mRNA containing m5C and m1Ψ. Untransfected cells served as control. ApppG-capped mRNA represents cap-independent translation. The m7GpppG cappedeGFP-mRNA (0) served as positive control. The NPM-(2) caged eGFP-mRNA was either not irradiated or irradiated in cells (405 nm, 60 s/420 nm 180 s). The top two rows show the 630x magnification (63x objective) of the red channel (mScarlet-I) or the green channel (eGFP) while the bottom two show the DAPI staining and DIC channel. Shown is one representative experiment of three independent experiments (n=3).
Extended Data Fig. 4. Western Blots of eGFP and Rheb_eGFP.

The eGFP, Rheb_eGFP and Nucleolin protein levels of HEK293T and HeLa cell samples transfected with either ApppG-, m7GpppG- or NPM- capped mRNA were analyzed via Western blotting at 24 h post transfection. Irradiation of transfected cells was performed 4 h post transfection at 365 nm for 30 sec. As marker, the prestainedPageRuler (ThermoFisher) was used and as primary antibodies anti-nucleolin-antibody or anti-eGFP-antibody were used, respectively. Additionally, a HRP secondary antibody was used. Shown is one representative gel from n=3 independent experiments.
Supplementary Material
{kind=link}
Acknowledgements
This project has received funding from the European Research Council (ERC) under the European Union's Horizon 2020 research and innovation programme (grant agreement No 772280; A.R.). We gratefully acknowledge funding by the DFG (RE2796/7-1; A.R.). We thank TRON (Translational Oncology at the University Medical Center of the Johannes Gutenberg University Mainz) for providing us with the HEK-NF-ĸB cell lines and Prof. Dr. D. Kümmel (Westfälische Wilhelms Universität Münster) for providing us the human Rheb plasmid DNA. The authors thank A.-M. Dörner, H. Schepers, N. Kück, A. Bollu, M. Dittmar for excellent technical and experimental assistance (all Westfälische Wilhelms Universität Münster). The NMR and mass measurements were supported by Dr. K. Bergander, the NMR- and MS-facility of the Organic Chemistry Institute at the Westfälische Wilhelms Universität Münster.
Footnotes
Author Contributions Statement
N.K. designed and performed the biochemical experiments. M.v.D. designed and performed the biomolecular and cell experiments. F.W. and P.S. designed and performed the chemical syntheses. S.H. performed cell experiments. A.R. conceived and supervised the project. N.K., F.W., M.v.D., P.S. and A.R. evaluated the data and discussed the results. N.K., F.W., M.v.D., P.S. and A.R. wrote the manuscript.
Competing Interests Statement
P.S., N.K., F.W. and A.R. are the inventors on a European patent application (EP 1 184 349.5, pending) of the University of Münster covering the synthesis and use of photocleavable 5'-cap analogs as well as RNA molecules comprising photocleavable 5'-cap analogs.
Data Availability
The data generated or analysed during this study are included in this article and its supplementary information files. Source Data for Figs. 3–6 and Extended Data Figures 1-4 are provided with the paper. Protein structures and models used for the figures are available under the accession codes: 1EJ1.
Code Availability
No custom software was used in this study. Software used for data collection: Leica Application Suite X (3.5.6.21594), CxP Analysis Software, CFX Manager Software V 3.1 (Bio-Rad), Qualitative Navigator B.08.00, Agilent ChemStation for LC 3D systems Rev. B.04.03, Agilent OpenLab CDS ChemStation Edition Rev. C.01.10. Software used for data analysis: Origin (2016b), Origin (2021b), Origin (2019b), GraphPad Prism 7, ImageJ (Version 20160205), MestReNova 14, LasX (Version), Excel 2016,, PerkinElmer ChemDraw 20.1.
References
- 1.Zhang C, Maruggi G, Shan H, Li J. Advances in mRNA Vaccines for Infectious Diseases. Frontiers in Immunology. 2019;10:594. doi: 10.3389/fimmu.2019.00594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sahin U, Kariko K, Tureci O. mRNA-based therapeutics--developing a new class of drugs. Nature Reviews Drug Discovery. 2014;13:759–780. doi: 10.1038/nrd4278. [DOI] [PubMed] [Google Scholar]
- 3.Besse F, Ephrussi A. Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat Struct Mol Biol. 2008;9:971–980. doi: 10.1038/nrm2548. [DOI] [PubMed] [Google Scholar]
- 4.Jansen RP, Niessing D, Baumann S, Feldbrugge M. mRNA transport meets membrane traffic. Trends Genet. 2014;30:408–417. doi: 10.1016/j.tig.2014.07.002. [DOI] [PubMed] [Google Scholar]
- 5.Shirokikh NE, Preiss T. Translation initiation by cap-dependent ribosome recruitment: Recent insights and open questions. Wires Rna. 2018;9:e1473. doi: 10.1002/wrna.1473. [DOI] [PubMed] [Google Scholar]
- 6.Mikkola S, Salomaki S, Zhang Z, Maki E, Lonnberg H. Preparation and properties of mRNA 5 '-cap structure. Curr Org Chem. 2005;9:999–1022. [Google Scholar]
- 7.von der Haar T, Gross JD, Wagner G, McCarthy JEG. The mRNA cap-binding protein eIF4E in post-transcriptional gene expression. Nat Struct Mol Biol. 2004;11:503–511. doi: 10.1038/nsmb779. [DOI] [PubMed] [Google Scholar]
- 8.Liu HD, Rodgers ND, Jiao X, Kiledjian M. The scavenger mRNA decapping enzyme DcpS is a member of the HIT family of pyrophosphatases. Embo J. 2002;21:4699–4708. doi: 10.1093/emboj/cdf448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Charenton C, et al. Structure of the active form of Dcp1-Dcp2 decapping enzyme bound to m(7)GDP and its Edc3 activator. Nat Struct Mol Biol. 2016;23:982–986. doi: 10.1038/nsmb.3300. [DOI] [PubMed] [Google Scholar]
- 10.Deshmukh MV, et al. mRNA decapping is promoted by an RNA-binding channel in Dcp2. Mol Cell. 2008;29:324–336. doi: 10.1016/j.molcel.2007.11.027. [DOI] [PubMed] [Google Scholar]
- 11.Goubau D, et al. Antiviral immunity via RIG-I-mediated recognition of RNA bearing 5'-diphosphates. Nature. 2014;514:372–375. doi: 10.1038/nature13590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Nallagatla SR, et al. 5'-triphosphate-dependent activation of PKR by RNAs with short stem-loops. Science. 2007;318:1455–1458. doi: 10.1126/science.1147347. [DOI] [PubMed] [Google Scholar]
- 13.De Gregorio E, Preiss T, Hentze MW. Translational activation of uncapped mRNAs by the central part of human eIF4G is 5' end-dependent. RNA. 1998;4:828–836. doi: 10.1017/s1355838298980372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wojtczak BA, et al. 5 '-Phosphorothiolate Dinucleotide Cap Analogues: Reagents for Messenger RNA Modification and Potent Small-Molecular Inhibitors of Decapping Enzymes. J Am Chem Soc. 2018;140:5987–5999. doi: 10.1021/jacs.8b02597. [DOI] [PubMed] [Google Scholar]
- 15.Kauffman KJ, et al. Optimization of Lipid Nanoparticle Formulations for mRNA Delivery in Vivo with Fractional Factorial and Definitive Screening Designs. Nano Letters. 2015;15:7300–7306. doi: 10.1021/acs.nanolett.5b02497. [DOI] [PubMed] [Google Scholar]
- 16.Holstein JM, Anhauser L, Rentmeister A. Modifying the 5'-Cap for Click Reactions of Eukaryotic mRNA and To Tune Translation Efficiency in Living Cells. Angewandte Chemie, International Edition in English. 2016;55:10899–10903. doi: 10.1002/anie.201604107. [DOI] [PubMed] [Google Scholar]
- 17.Gebauer F, Hentze MW. Molecular mechanisms of translational control. Nat Struct Mol Biol. 2004;5:827–835. doi: 10.1038/nrm1488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Weber AM, et al. A blue light receptor that mediates RNA binding and translational regulation. Nat Chem Biol. 2019;15:1085–1092. doi: 10.1038/s41589-019-0346-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Govan JM, et al. Optochemical control of RNA interference in mammalian cells. Nucleic Acids Research. 2013;41:10518–10528. doi: 10.1093/nar/gkt806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dhamodharan V, Nomura Y, Dwidar M, Yokobayashi Y. Optochemical control of gene expression by photocaged guanine and riboswitches. Chemical Communications (Cambridge, United Kingdom) 2018;54:6181–6183. doi: 10.1039/c8cc02290a. [DOI] [PubMed] [Google Scholar]
- 21.Shestopalov IA, Sinha S, Chen JK. Light-controlled gene silencing in zebrafish embryos. Nat Chem Biol. 2007;3:650–651. doi: 10.1038/nchembio.2007.30. [DOI] [PubMed] [Google Scholar]
- 22.Bardhan A, Deiters A, Ettensohn CA. Conditional gene knockdowns in sea urchins using caged morpholinos. Dev Biol. 2021;475:21–29. doi: 10.1016/j.ydbio.2021.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tang XJ, Maegawa S, Weinberg ES, Dmochowski IJ. Regulating gene expression in zebrafish embryos using light-activated, negatively charged peptide nucleic acids. J Am Chem Soc. 2007;129:11000. doi: 10.1021/ja073723s. [DOI] [PubMed] [Google Scholar]
- 24.Keyhani S, Goldau T, Bluemler A, Heckel A, Schwalbe H. Chemo-Enzymatic Synthesis of Position-Specifically Modified RNA for Biophysical Studies including Light Control and NMR Spectroscopy. Angew Chem Int Edit. 2018;57:12017–12021. doi: 10.1002/anie.201807125. [DOI] [PubMed] [Google Scholar]
- 25.Wilkie GS, Davis I. Drosophila wingless and pair-rule transcripts localize apically by dyneinmediated transport of RNA particles. Cell. 2001;105:209–219. doi: 10.1016/s0092-8674(01)00312-9. [DOI] [PubMed] [Google Scholar]
- 26.Hoernes TP, et al. Translation of non-standard codon nucleotides reveals minimal requirements for codon-anticodon interactions. Nature Communications. 2018;9:4865. doi: 10.1038/s41467-018-07321-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhang D, Jin S, Piao X, Devaraj NK. Multiplexed Photoactivation of mRNA with Single-Cell Resolution. ACS Chemical Biology. 2020;15:1773–1779. doi: 10.1021/acschembio.0c00205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ando H, Furuta T, Tsien RY, Okamoto H. Photo-mediated gene activation using caged RNA/DNA in zebrafish embryos. Nat Genet. 2001;28:317–325. doi: 10.1038/ng583. [DOI] [PubMed] [Google Scholar]
- 29.Ogasawara S. Duration Control of Protein Expression in Vivo by Light-Mediated Reversible Activation of Translation. ACS Chemical Biology. 2017;12:351–356. doi: 10.1021/acschembio.6b00684. [DOI] [PubMed] [Google Scholar]
- 30.Rotstan KA, et al. Regulation of mRNA translation by a photoriboswitch. eLife. 2020;9:e51737. doi: 10.7554/eLife.51737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhang D, Zhou CY, Busby KN, Alexander SC, Devaraj NK. Light-Activated Control of Translation by Enzymatic Covalent mRNA Labeling. Angew Chem Int Edit. 2018;57:2822–2826. doi: 10.1002/anie.201710917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ogasawara S. Control of cellular function by reversible photoregulation of translation. Chembiochem. 2014;15:2652–2655. doi: 10.1002/cbic.201402495. [DOI] [PubMed] [Google Scholar]
- 33.Mauger DM, et al. mRNA structure regulates protein expression through changes in functional half-life. P Natl Acad Sci USA. 2019;116:24075–24083. doi: 10.1073/pnas.1908052116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kariko K, Buckstein M, Ni H, Weissman D. Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity. 2005;23:165–175. doi: 10.1016/j.immuni.2005.06.008. [DOI] [PubMed] [Google Scholar]
- 35.Garcia JF, Parker R. MS2 coat proteins bound to yeast mRNAs block 5' to 3' degradation and trap mRNA decay products: implications for the localization of mRNAs by MS2-MCP system. RNA. 2015;21:1393–1395. doi: 10.1261/rna.051797.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Anhauser L, et al. A Benzophenone-Based Photocaging Strategy for the N7 Position of Guanosine. Angewandte Chemie, International Edition in English. 2020;59:3161–3165. doi: 10.1002/anie.201914573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Rydzik AM, et al. Synthetic dinucleotide mRNA cap analogs with tetraphosphate 5',5' bridge containing methylenebis(phosphonate) modification. Organic & Biomolecular Chemistry. 2009;7:4763–4776. doi: 10.1039/b911347a. [DOI] [PubMed] [Google Scholar]
- 38.Grudzien-Nogalska E, et al. Synthetic mRNAs with superior translation and stability properties. Methods Molecular Biology. 2013;969:55–72. doi: 10.1007/978-1-62703-260-5_4. [DOI] [PubMed] [Google Scholar]
- 39.Jemielity J, et al. Novel "anti-reverse" cap analogs with superior translational properties. RNA. 2003;9:1108–1122. doi: 10.1261/rna.5430403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Aujard I, et al. o-nitrobenzyl photolabile protecting groups with red-shifted absorption: syntheses and uncaging cross-sections for one- and two-photon excitation. Chemistry. 2006;12:6865–6879. doi: 10.1002/chem.200501393. [DOI] [PubMed] [Google Scholar]
- 41.Bohacova S, et al. Protected 5-(hydroxymethyl) uracil nucleotides bearing visible-light photocleavable groups as building blocks for polymerase synthesis of photocaged DNA. Organic & Biomolecular Chemistry. 2018;16:1527–1535. doi: 10.1039/c8ob00160j. [DOI] [PubMed] [Google Scholar]
- 42.Liu W, et al. Structural basis for nematode eIF4E binding an m(2,2,7)G-Cap and its implications for translation initiation. Nucleic Acids Research. 2011;39:8820–8832. doi: 10.1093/nar/gkr650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Anhauser L, Huwel S, Zobel T, Rentmeister A. Multiple covalent fluorescence labeling of eukaryotic mRNA at the poly(A) tail enhances translation and can be performed in living cells. Nucleic Acids Research. 2019;47:e42. doi: 10.1093/nar/gkz084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kormann MS, et al. Expression of therapeutic proteins after delivery of chemically modified mRNA in mice. Nature biotechnology. 2011;29:154–157. doi: 10.1038/nbt.1733. [DOI] [PubMed] [Google Scholar]
- 45.Kariko K, et al. Incorporation of Pseudouridine Into mRNA Yields Superior Nonimmunogenic Vector With Increased Translational Capacity and Biological Stability. Mol Ther. 2008;16:1833–1840. doi: 10.1038/mt.2008.200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Jia L, et al. Decoding mRNA translatability and stability from the 5' UTR. Nat Struct Mol Biol. 2020;27:814–821. doi: 10.1038/s41594-020-0465-x. [DOI] [PubMed] [Google Scholar]
- 47.van Dülmen M, Muthmann N, Rentmeister A. Chemo-Enzymatic Modification of the 5' Cap Maintains Translation and Increases Immunogenic Properties of mRNA. Angewandte Chemie, International Edition in English. 2021;60:13280–13286. doi: 10.1002/anie.202100352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lucas T, et al. Light-inducible antimiR-92a as a therapeutic strategy to promote skin repair in healing-impaired diabetic mice. Nature Communications. 2017;8 doi: 10.1038/ncomms15162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Chen C, et al. Dextran-Conjugated Caged siRNA Nanoparticles for Photochemical Regulation of RNAi-Induced Gene Silencing in Cells and Mice. Bioconjug Chem. 2019;30:1459–1465. doi: 10.1021/acs.bioconjchem.9b00204. [DOI] [PubMed] [Google Scholar]
- 50.Muthmann N, et al. Combining Chemical Synthesis and Enzymatic Methylation to Access Short RNAs with Various 5' Caps. ChemBioChem. 2019;20:1693–1700. doi: 10.1002/cbic.201900037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Holstein JM, Stummer D, Rentmeister A. Enzymatic modification of 5'-capped RNA with a 4-vinylbenzyl group provides a platform for photoclick and inverse electron-demand Diels-Alder reaction. Chem Sci. 2015;6:1362–1369. doi: 10.1039/c4sc03182b. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data generated or analysed during this study are included in this article and its supplementary information files. Source Data for Figs. 3–6 and Extended Data Figures 1-4 are provided with the paper. Protein structures and models used for the figures are available under the accession codes: 1EJ1.
No custom software was used in this study. Software used for data collection: Leica Application Suite X (3.5.6.21594), CxP Analysis Software, CFX Manager Software V 3.1 (Bio-Rad), Qualitative Navigator B.08.00, Agilent ChemStation for LC 3D systems Rev. B.04.03, Agilent OpenLab CDS ChemStation Edition Rev. C.01.10. Software used for data analysis: Origin (2016b), Origin (2021b), Origin (2019b), GraphPad Prism 7, ImageJ (Version 20160205), MestReNova 14, LasX (Version), Excel 2016,, PerkinElmer ChemDraw 20.1.