

Abstract
Periplasmic flagella are complex nanomachines responsible for distinctive morphology and motility of spirochetes. Although bacterial flagella have been extensively studied for several decades in model systems Escherichia coli and Salmonella enterica, our understanding of periplasmic flagella in many disease-causing spirochetes remains incomplete. Recent advances, including molecular genetics, biochemistry, structural biology, and cryo-electron tomography, have greatly increased our understanding of structure and function of periplasmic flagella. In this chapter, we summarize some of the recent findings that provide new insights into the structure, assembly and function of periplasmic flagella.
Keywords: molecular machines, bacterial motility, protein secretion, cryo electron microscopy, cryo electron tomography
The flagellum is a major organelle for motility in many bacterial species. It confers locomotion and is often associated with virulence of bacterial pathogens. Flagella from different species share a conserved core, but also exhibit profound variations in flagellar structure, flagellar number and placement (Chen et al., 2011; Zhao et al., 2014), resulting in distinct flagella that appear to be adapted to the specific environments that the bacteria encounter. While many bacteria possess multiple peritrichous flagella such as those found in Escherichia coli and Salmonella enterica, other bacteria such as Vibrio spp. and Pseudomonas aeruginosa normally have a single flagellum at one cell pole (Fig. 1). Spirochetes uniquely assemble flagella that are embedded in periplasmic space between their inner and outer membranes, thus called periplasmic flagella (Charon et al., 2012). Although the flagella of E. coli and Salmonella have been extensively studied for several decades, periplasmic flagella are less understood, despite their profound impact on the distinctive morphology and motility of spirochetes. In this chapter, many aspects of periplasmic flagella will be discussed, with particular focus on their structure and assembly.
Figure 1. Distinctive placement of bacterial flagellum.

(A) Bacteria with flagella distributed all over the cell (e.g. Escherichia coli) are peritrichous. (B) Monotrichous bacteria, such as Vibrio cholera, Pseudomonas aeruginosa, and Caulobacter crescentus have a single flagellum present at one end of the cell. (C). Spirochetes, including species of Borrelia, Treponema, and Leptospira, possess specialized flagella located within the periplasmic space. The rotation of the periplasmic flagella allows this bacterium to swim forward in a corkscrew-like motion.
Spirochetes are a distinctive group of bacteria of significant importance in human health.
Spirochetes cause several major diseases in humans such as Lyme disease (Borrelia burgdorferi), syphilis (Treponema pallidum), leptospirosis (Leptospira interrogans), and periodontitis (Treponema spp.). Lyme disease is the most commonly reported tick-borne illness in the United States, and the incidence is growing rapidly. The number of patients diagnosed with Lyme disease each year in the United States is approximately 300,000. The disease is caused by Borrelia burgdorferi and related organisms, and is transmitted to humans through the bite of infected Ixodes ticks (Radolf et al., 2012). Syphilis is a common sexually transmitted disease in many areas of the world. Leptospirosis is the most common waterborne zoonosis worldwide.
Motility is essential for spirochetes to infect and disseminate in mammalian hosts.
Spirochetal motility is unique, as the entire bacterium is involved in translocation without the involvement of external appendages. The motility is driven by periplasmic flagella, and rotation of the flagella causes a serpentine movement, allowing the organism to very efficiently bore its way through viscous media or tissue (Charon et al., 2012). To complete the host-vector life cycle, B. burgdorferi is able to adapt to divergent host environments and also evade the defense of its mammalian reservoir (Radolf et al., 2012). Several studies provide direct evidence that the unique motility and chemotaxis of B. burgdorferi are essential for the establishment of infection in mammals and the completion of its enzootic cycle (Li et al., 2010; Motaleb et al., 2015; Sultan et al., 2013; Sultan et al., 2015).
Periplasmic flagella are necessary for the flat-wave morphology and distinctive motility of B. burgdorferi.
B. burgdorferi possesses 7–11 periplasmic flagella that are inserted at each cell pole and wrap around the cell cylinder to produce the spirochete’s distinctive flat-wave morphology. Periplasmic flagella are crucial not only for motility but also for the overall shape of B. burgdorferi, as mutant cells lacking flagella are non-motile and exhibit a rod-shaped morphology (Motaleb et al., 2000). Similar to the external flagella found in the model organisms E. coli and S. enterica, periplasmic flagella are composed of the flagellar motor, the hook and the filament. The flagellar motor is a rotary motor that anchors the flagellum to the inner membrane. The motors of the periplasmic flagella are noticeably larger than those of other external flagella (Fig. 2). Importantly, the motor possesses a spirochete-specific “collar” (Fig. 2) (Kudryashev et al., 2009; Liu et al., 2009; Moon et al., 2016; Murphy et al., 2006). The motor can be further divided into two parts: the rotor and the stator. The rotation of the motor is driven by the torque generated by the stator-rotor interaction, utilizing energy generated by the flow of protons through the stator channel. The rotor is composed of the MS-ring, the C-ring, and the rod. The MS-ring is the base of the rotor and it is formed by multiple copies of FliF. The C-ring is located in the cytoplasm and is also known as the switch complex. It consists of the proteins FliG, FliM and FliN and controls the direction of flagellar rotation. The rod serves as a drive shaft and consists of multiple different proteins (FlgB, FlgC, FlgF and FlgG). The hook of the periplasmic flagellum is located in periplasmic space, in contrast to the externally localized hook in E. coli and S. enterica. A recent study indicates that the hook proteins are cross-linked by a covalent bond, an unusual property necessary for transmission of high rotational torque from the motor to the filament (Miller et al., 2016). The filament is the longest component of the periplasmic flagella. Multiple filaments arising from both poles form flat ribbons that wrap around the spirochete cell body in a right-handed fashion (Charon et al., 2009). Flagellar-specific type III secretion system (fT3SS), which is embedded in the flagellar motor, is responsible for the transport and assembly of the protein compoments of the rod, the hook and the filament (Zhao et al., 2013).
Figure 2. Comparison of motor structures from E. coli, Vibrio, H. pylori, and Borrelia.
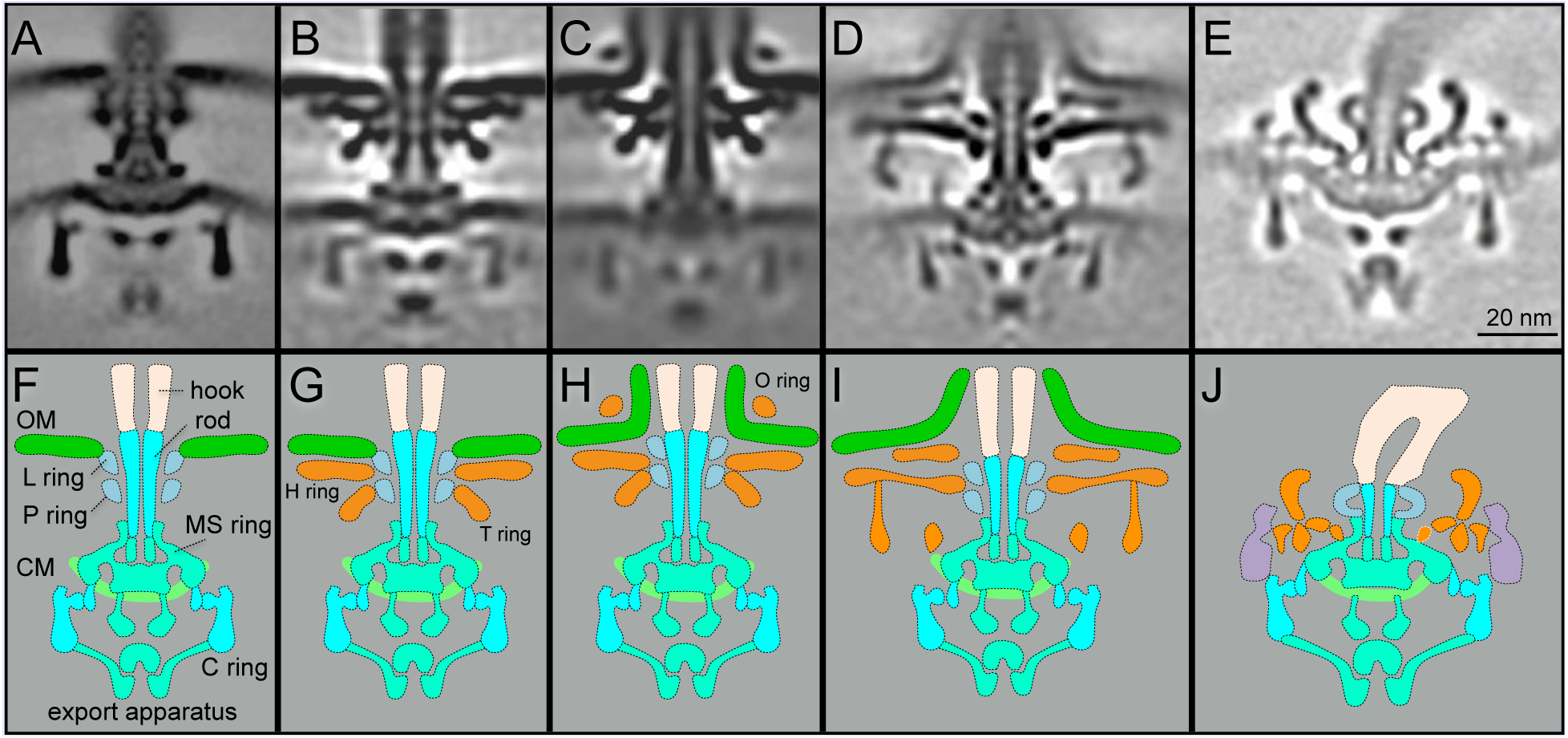
(A) A central section of an E. coli flagellar motor. (B) A central section from a non-sheathed Vibrio flagellar motor. (C) A central section from a sheathed Vibrio flagellar motor. (D) A central section from a sheathed flagellar motor of H. pylori. (E) A central section from Borrelia flagellar motor. (F-J) Schematic models derived from the central sections (A-E), respectively. Adapted from prior publications (Qin et al., 2016; Zhao et al., 2013; Zhu et al., 2017), with permission.
Characterization of the unique periplasmic structure of spirochetal flagella.
The collar is a unique spirochete-specific component that has not been found in other bacterial flagella reported to date (Chen et al., 2011; Zhao et al., 2014). The collar in the B. burgdorferi periplasmic flagellar motor is a large complex with ~71 nm in diameter and ~24 nm in height, presumably composed of many different proteins. However, there is limited information regarding its structure, function and protein components. Recently, a hypothetical membrane protein FlbB was identified as a candidate involved in collar assembly (Moon et al., 2016). In addition, the novel tetratricopeptide repeat protein BB0236 was also proposed to contribute to the collar assembly (Moon et al., 2018). Mutants deficient in either FlbB or BB0236 are non-motile and their periplasmic flagella lack the collar, its associated proteins (including FliL), and the stator (Fig. 3). This finding provides direct evidence that the collar is indeed an important (as well as unique) component of periplasmic flagella (Fig. 2). Although additional unknown proteins are likely involved in the collar assembly, it is evident that the periplasmic collar provides a static framework promoting the recruitment and stable association of stator units, which could in turn facilitate the higher torques to rotate the periplasmic flagellum. The rotation of the flagella within the confinements of the periplasm enables the spirochete to bore its way through complex, viscous environment in vertebrate and tick tissues.
Figure 3. Characterization of the unique features in periplasmic flagella, as examined through mutational analysis.
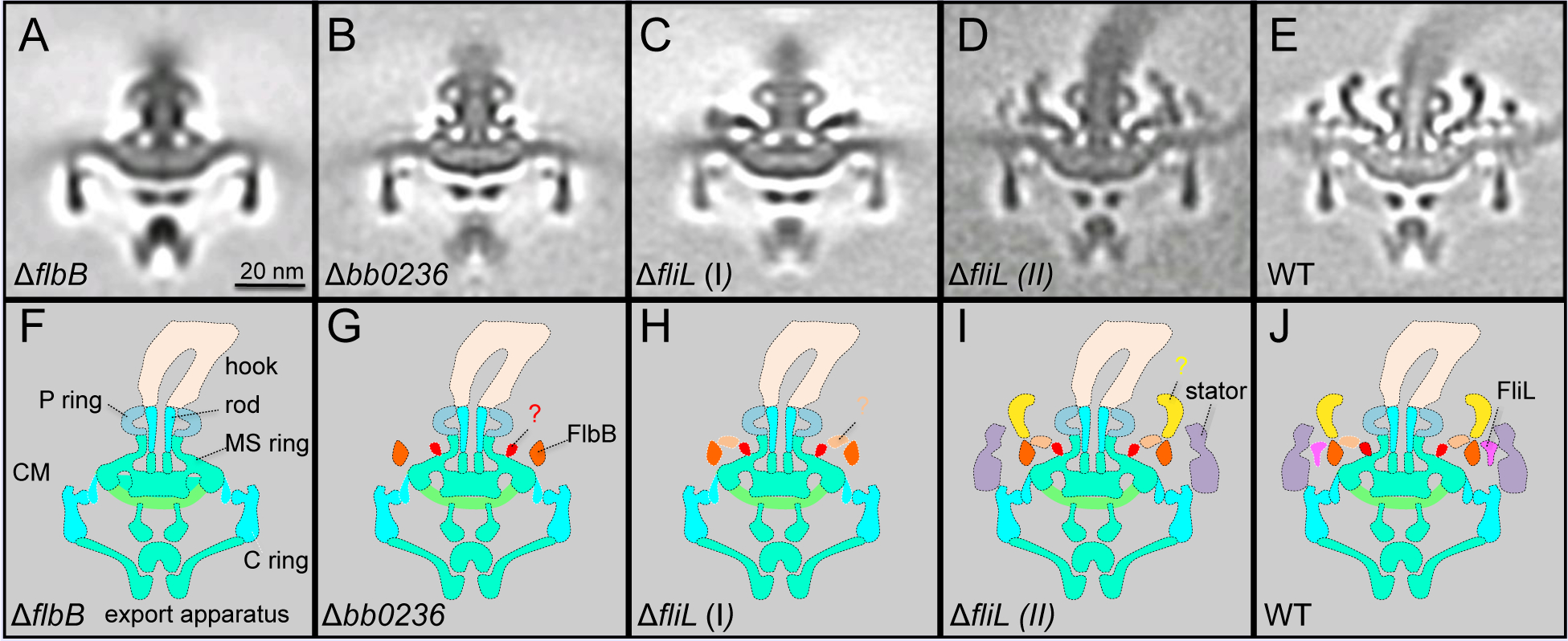
(A) Central section from a mutant lacking FlbB. (B) Central section from a mutant lacking BB0236. (C) Central section from a class average of a mutant lacking FliL. (D) Central section from another class average of a mutant lacking fliL. (E) A central section from WT flagellar motor. (F-J) Schematic models derived from panels (A-E), respectively. Adapted from prior publication (Moon et al., 2018), with permission.
Stator-rotor interaction.
Powered by the electrochemical gradient across the cytoplasmic membrane, the flagellar motor can rotate the filament at high speed. It is believed that the flagellar rotation is mediated by the interaction between the cytoplasmic loop region of MotA, and the C-terminal domain of FliG in the C-ring. However, there is limited structural information on stator-rotor interaction in model systems E. coli and S. enterica, largely because the stator is poorly resolved due to its dynamic nature and low occupancy (Fukuoka et al., 2009; Leake et al., 2006; Paulick et al., 2009). In contrast, the in situ flagellar motor structures of B. burgdorferi and other spirochetes determined by cryo-electron tomography (cryo-ET) reveal more detailed information regarding the stators and their interactions with the C-ring (Chen et al., 2011; Kudryashev et al., 2010; Liu et al., 2010; Liu et al., 2009; Murphy et al., 2006; Raddi et al., 2012). The presence of the collar in spirochetes is likely essential for the better visualization of the stator and its interaction with the rotor, because the collar provides a stable framework to recruit and stabilize the stators.
CheY-P binding and flagellar switching of rotational direction.
The flagellar motor in many bacteria species can rotate in both counter-clockwise (CCW) and clockwise (CW) directions to achieve swimming towards attractants or away from repellents. The rotation direction is controlled by a sophisticated chemotactic system. In the signaling pathway, CheY is phosphorylated by CheA kinase, then the phosphorylated CheY binds to the FliM protein in the C-ring and induces conformational changes that alter the stator-rotation interaction and cause switching (Welch et al., 1993). Studies in E. coli of the correlation between the CW rotation and the intracellular level of the phosphorylated CheY in individual cells indicated that binding and switching are highly cooperative (Cluzel et al., 2000). The switching spreads from one or more nucleation points on the C-ring, a phenomenon referred to as ‘conformational spread’ (Bai et al., 2010). Recent experiments revealed that the flagellar motor can adapt to varied levels of phosphorylated CheY by increasing the content of FliM (Yuan et al., 2012). Additional experiments suggested that it is not CheY-P binding, but rather the direction of motor rotation, that has the largest effect on remodeling of the FliM (Lele et al., 2012). It was suggested that there are ~34 molecules of FliM in a motor with exclusively CW rotation and ~44 molecules in a motor with CCW rotation. These E. coli studies also indicate that motors with even more FliM molecules may exist. It is unclear how the C-ring can accommodate such a large change, and if similar C-ring modifications also occur in spirochetes.
Because periplasmic flagellar motors are located at the two cell poles, it was hypothesized that spirochetal motors rotate asymmetrically at one end relative to the other during a run (Charon and Goldstein, 2002; Li et al., 2002). CheX is the only CheY-P phosphatase identified in the B. burgdorferi genome. A cheX mutant constantly flexes and is not able to run or reverse (Motaleb et al., 2005), while both cheA2 and cheY3 mutant constantly run in one direction (Li et al., 2002; Motaleb et al., 2011). A comparison of the motor structures from two different motions (flex and run) will likely shed new light upon the mechanisms underlying CheY-P binding and the switching of rotational direction.
Flagellar assembly.
The bacterial flagellum is built from the inside out, from proximal to distal structures, in a temporally and spatially regulated fashion. Detailed insights into the flagellar assembly have been well established in E. coli and S. enterica (Chevance and Hughes, 2008; Macnab, 2003). In these organisms, multiple copies of FliF form the MS-ring (Suzuki et al., 2004), which serves as the initial base for flagellar assembly, structural maturation, and function. The MS-ring also serves as a scaffold for the assembly of the C-ring. FliG proteins directly associate with the cytoplasmic face of the MS-ring and form the FliG ring (Lee et al., 2010; Minamino et al., 2011). FliM and FliN proteins form a stable complex with a stoichiometry of 1:4 (Brown et al., 2005; Delalez et al., 2014). The FliM-FliN4 complex will bind to the FliG ring to form the completed C-ring. The export apparatus, which is assembled inside the MS-ring and the C-ring, is responsible to export flagellar axial protein components from the cytoplasm to the distal end of the nascent flagellar apparatus. FliE are likely assembled first and form a junction between the MS-ring and the rod to overcome their symmetry mismatch. Then multiple copies of FlgB,C,F,G form the rod, FlgI form the P-ring and FliH form the L-ring. FlgD cap assembles at the rod tip to support the assembly of the hook. Then the filament-cap (FliD) is formed after the hook assembly to support the assembly of the filament (Zhang et al., 2019). By arresting assembly with a series of genetic mutations, cryo-ET imaging of the motor in B. burgdorferi provided snapshots of the sequential assembly of periplasmic flagella (Zhao et al., 2013).
Flagellar export apparatus and its evolutionarily related injectisome.
The fT3SS consists of five integral membrane proteins (FlhA, FlhB, FliP, FliQ and FliR) and three soluble proteins (FliH, FliI and FliJ), and is located at the center of the cytoplasmic face of the MS-ring. The ATP complex promotes the export process by binding and delivering substrates to the export apparatus (Fraser et al., 2003; Minamino and Imada, 2015). FliI is an ATPase and shows structural similarity with the α and β subunits of the F0F1-ATP synthase (Ibuki et al., 2011); it exhibits its full ATPase activity when it self-assembles into a homo-hexamer (Imada et al., 2007). FliH, FliI, and FliJ coordinately deliver a chaperone-substrate complex to the export gate by binding to the docking platform of the fT3SS for substrate export. FliP, FliQ and FliR form an export gate complex with helical symmetry (Kuhlen et al., 2018).
fT3SSs in different bacterial species are highly conserved. In addition, they are evolutionally related to virulence T3SSs. The evolutionary relationship between the flagellum and the injectisome has garnered significant debate. The latest phylogenomic and comparative analyses of fT3SSs and vT3SSs suggest that the vT3SS derived from a flagellar ancestor. The loss of flagellum-specific genes led to an eventual loss in the motility function, but this system presumably kept the ability to secrete proteins (Abby and Rocha, 2012)
The overall organization of the fT3SS machine in periplasmic flagella shares many similar features as those observed in the vT3SS machine (Hu et al., 2017; Hu et al., 2015; Kawamoto et al., 2013) (Fig 4). However, the ATPase complex of the periplasmic flagella is noticeably different from those observed in the injectisome (Fig 4). There are 23 spokes and one hub in the ATPase complex of the B. burgdorferi periplasmic flagella. Only 6 spokes and one hob were observed in Salmonella injectisome, presumably optimizing for substrate recruitment and export. In contrast, the ATPase complex in the periplasmic flagella not only facilitates substrate recruitment and secretion, but also supports the integrity of the C-ring, which undergoes rotation and switches rotational direction between CW and CCW.
Figure 4. Comparison of the fT3SS from B. burgdorferi and the vT3SS from Salmonella.
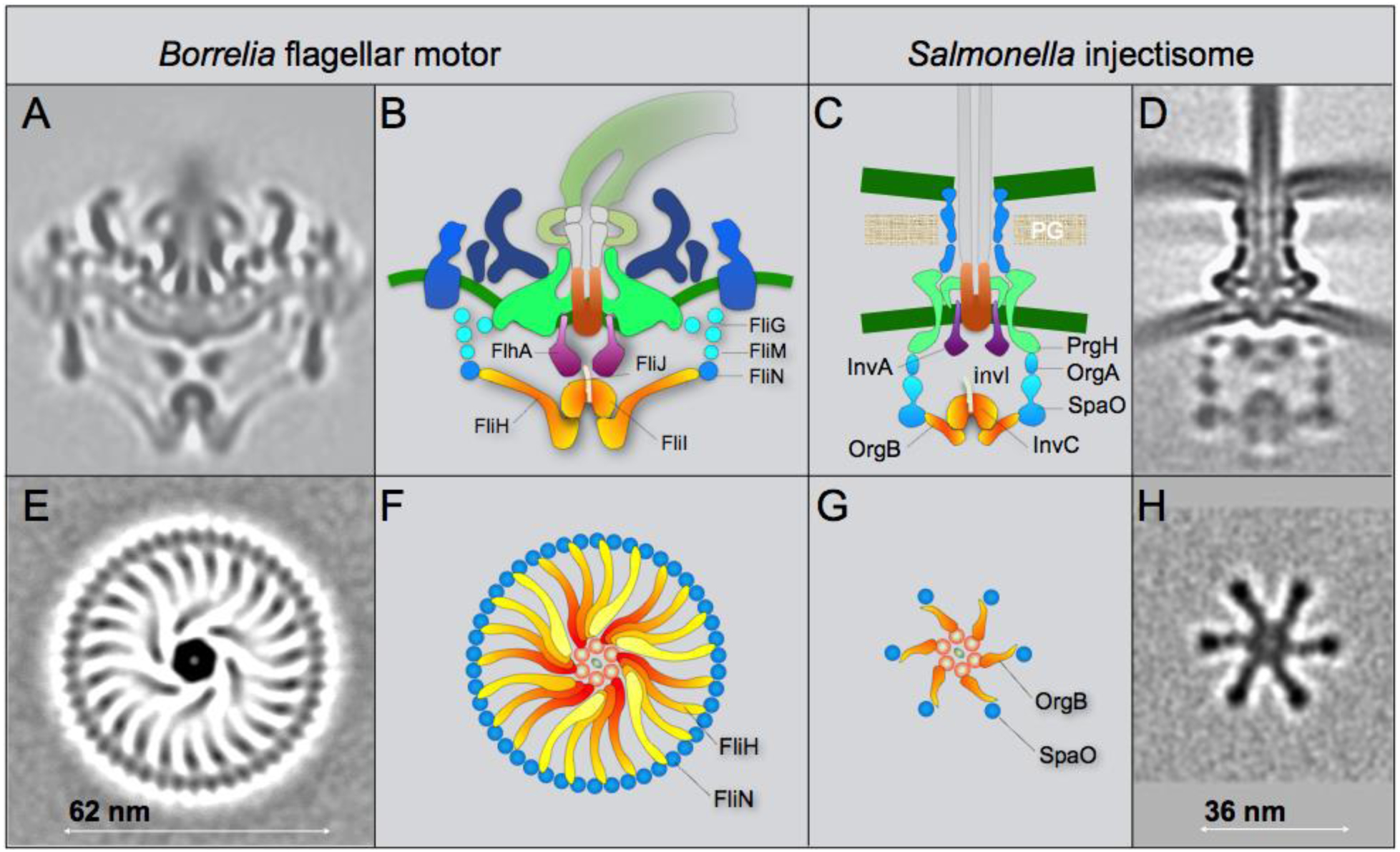
(A) A central section from the B. burgdorferi motor. (B) The fT3SS in the spirochete motor includes the ATPase complex (orange) and the export apparatus (purple) underneath the MS-ring. (C, D) The vT3SS from Salmonella injectisome is modeled in a similar color scheme. The difference between the two T3SSs is striking in a comparison of the cross-sections of their ATPase complexes. Note that the C-ring from the B. burgdorferi motor is a continuous ring with ~46 copies of FliN tetramer. There are 23 visible FliH spokes (E, F). There are six pods in Salmonella injectisome. Only six spokes of the FliH homolog OrgB connect the ATPase complex to the SpaO molecules that compose the pod of the injectisome. Adapted from prior publication (Qin et al., 2018), with permission.
Outlook and perspective.
Although the structure and functions of bacterial flagellum have been studied for several decades, many important questions remain to be addressed. For example, how does the stator couple proton gradient to generate the torque? How does the C-ring change its conformation to generate rotation or switch rotational direction? How does the proton gradient power the expert apparatus and facilitate protein transport across inner membrane? Periplasmic flagella also raise additional questions: how do the periplasmic flagella coordinate their rotation from two cell poles? how does the spirochete specific collar assemble? Given that periplasmic flagella play critical roles in many bacterial pathogens, it will be important to understand not only the conserved structure and function among bacterial flagella, but also the specific features that distinct periplasmic flagella from others. Emerging techniques such as cryo-ET will be increasingly valuable to address many of these fundamental questions in periplasmic flagellum.
Acknowledgements
The work in Liu Laboratory was supported by grants GM107629 and R01AI087946 from National Institutes of Health.
REFERENCE:
- Abby SS, and Rocha EP (2012). The non-flagellar type III secretion system evolved from the bacterial flagellum and diversified into host-cell adapted systems. PLoS genetics 8, e1002983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bai F, Branch RW, Nicolau DV Jr., Pilizota T, Steel BC, Maini PK, and Berry RM (2010). Conformational spread as a mechanism for cooperativity in the bacterial flagellar switch. Science 327, 685–689. [DOI] [PubMed] [Google Scholar]
- Brown PN, Mathews MAA, Joss LA, Hill CP, and Blair DF (2005). Crystal Structure of the Flagellar Rotor Protein FliN from Thermotoga maritima. J Bacteriol 187, 2890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charon NW, Cockburn A, Li C, Liu J, Miller KA, Miller MR, Motaleb MA, and Wolgemuth CW (2012). The unique paradigm of spirochete motility and chemotaxis. Ann Rev Microbiol 66, 349–370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charon NW, and Goldstein SF (2002). Genetics of motility and chemotaxis of a fascinating group of bacteria: the spirochetes. Annu Rev Genet 36, 47–73. [DOI] [PubMed] [Google Scholar]
- Charon NW, Goldstein SF, Marko M, Hsieh C, Gebhardt LL, Motaleb MA, Wolgemuth CW, Limberger RJ, and Rowe N (2009). The flat-ribbon configuration of the periplasmic flagella of Borrelia burgdorferi and its relationship to motility and morphology. J Bacteriol 191, 600–607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen S, Beeby M, Murphy GE, Leadbetter JR, Hendrixson DR, Briegel A, Li Z, Shi J, Tocheva EI, Muller A, et al. (2011). Structural diversity of bacterial flagellar motors. EMBO J 30, 2972–2981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chevance FF, and Hughes KT (2008). Coordinating assembly of a bacterial macromolecular machine. Nat Rev Microbiol 6, 455–465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cluzel P, Surette M, and Leibler S (2000). An ultrasensitive bacterial motor revealed by monitoring signaling proteins in single cells. Science 287, 1652–1655. [DOI] [PubMed] [Google Scholar]
- Delalez NJ, Berry RM, and Armitage JP (2014). Stoichiometry and Turnover of the Bacterial Flagellar Switch Protein FliN. mBio 5, e01216–01214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fraser GM, Gonzalez-Pedrajo B, Tame JR, and Macnab RM (2003). Interactions of FliJ with the Salmonella type III flagellar export apparatus. J Bacteriol 185, 5546–5554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukuoka H, Wada T, Kojima S, Ishijima A, and Homma M (2009). Sodium‐dependent dynamic assembly of membrane complexes in sodium‐driven flagellar motors. Mol Microbiol 71, 825–835. [DOI] [PubMed] [Google Scholar]
- Hu B, Lara-Tejero M, Kong Q, Galan JE, and Liu J (2017). In Situ Molecular Architecture of the Salmonella Type III Secretion Machine. Cell 168, 1065–1074 e1010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu B, Morado DR, Margolin W, Rohde JR, Arizmendi O, Picking WL, Picking WD, and Liu J (2015). Visualization of the type III secretion sorting platform of Shigella flexneri. Proc Natl Acad Sci U S A 112, 1047–1052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibuki T, Imada K, Minamino T, Kato T, Miyata T, and Namba K (2011). Common architecture of the flagellar type III protein export apparatus and F-and V-type ATPases. Nat Struct Mol Biol 18, 277–282. [DOI] [PubMed] [Google Scholar]
- Imada K, Minamino T, Tahara A, and Namba K (2007). Structural similarity between the flagellar type III ATPase FliI and F1-ATPase subunits. Proc Natl Acad Sci U S A 104, 485–490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawamoto A, Morimoto YV, Miyata T, Minamino T, Hughes KT, Kato T, and Namba K (2013). Common and distinct structural features of Salmonella injectisome and flagellar basal body. Sci Rep 3, 3369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kudryashev M, Cyrklaff M, Baumeister W, Simon MM, Wallich R, and Frischknecht F (2009). Comparative cryo-electron tomography of pathogenic Lyme disease spirochetes. Mol Microbiol 71, 1415–1434. [DOI] [PubMed] [Google Scholar]
- Kudryashev M, Cyrklaff M, Wallich R, Baumeister W, and Frischknecht F (2010). Distinct in situ structures of the Borrelia flagellar motor. J Struct Biol 169, 54–61. [DOI] [PubMed] [Google Scholar]
- Kuhlen L, Abrusci P, Johnson S, Gault J, Deme J, Caesar J, Dietsche T, Mebrhatu MT, Ganief T, Macek B, et al. (2018). Structure of the core of the type III secretion system export apparatus. Nat Struct Mol Biol 25, 583–590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leake MC, Chandler JH, Wadhams GH, Bai F, Berry RM, and Armitage JP (2006). Stoichiometry and turnover in single, functioning membrane protein complexes. Nature 443, 355. [DOI] [PubMed] [Google Scholar]
- Lee LK, Ginsburg MA, Crovace C, Donohoe M, and Stock D (2010). Structure of the torque ring of the flagellar motor and the molecular basis for rotational switching. Nature 466, 996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lele PP, Branch RW, Nathan VSJ, and Berg HC (2012). Mechanism for adaptive remodeling of the bacterial flagellar switch. Proc Natl Acad Sci U S A 109, 20018–20022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C, Bakker RG, Motaleb MA, Sartakova ML, Cabello FC, and Charon NW (2002). Asymmetrical flagellar rotation in Borrelia burgdorferi nonchemotactic mutants. Proc Natl Acad Sci U S A 99, 6169–6174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C, Xu H, Zhang K, and Liang FT (2010). Inactivation of a putative flagellar motor switch protein FliG1 prevents Borrelia burgdorferi from swimming in highly viscous media and blocks its infectivity. Mol Microbiol 75, 1563–1576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu J, Howell JK, Bradley SD, Zheng Y, Zhou ZH, and Norris SJ (2010). Cellular architecture of Treponema pallidum: novel flagellum, periplasmic cone, and cell envelope as revealed by cryo electron tomography. J Mol Biol 403, 546–561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu J, Lin T, Botkin DJ, McCrum E, Winkler H, and Norris SJ (2009). Intact flagellar motor of Borrelia burgdorferi revealed by cryo-electron tomography: evidence for stator ring curvature and rotor/C-ring assembly flexion. J Bacteriol 191, 5026–5036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macnab RM (2003). How bacteria assemble flagella. Annu Rev Microbiol 57, 77–100. [DOI] [PubMed] [Google Scholar]
- Miller MR, Miller KA, Bian J, James ME, Zhang S, Lynch MJ, Callery PS, Hettick JM, Cockburn A, Liu J, et al. (2016). Spirochaete flagella hook proteins self-catalyse a lysinoalanine covalent crosslink for motility. Nature Microbiol, 1(10): 16134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minamino T, and Imada K (2015). The bacterial flagellar motor and its structural diversity. Trends Microbiol 23, 267–274. [DOI] [PubMed] [Google Scholar]
- Minamino T, Imada K, Kinoshita M, Nakamura S, Morimoto YV, and Namba K (2011). Structural insight into the rotational switching mechanism of the bacterial flagellar motor. PLoS Biol 9, e1000616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moon KH, Zhao X, Manne A, Wang J, Yu Z, Liu J, and Motaleb MA (2016). Spirochetes flagellar collar protein FlbB has astounding effects in orientation of periplasmic flagella, bacterial shape, motility, and assembly of motors in Borrelia burgdorferi. Mol Microbiol 102, 336–348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moon KH, Zhao X, Xu H, Liu J, and Motaleb MA (2018). A tetratricopeptide repeat domain protein has profound effects on assembly of periplasmic flagella, morphology and motility of the lyme disease spirochete Borrelia burgdorferi. Mol Microbiol 110, 634–647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motaleb MA, Corum L, Bono JL, Elias AF, Rosa P, Samuels DS, and Charon NW (2000). Borrelia burgdorferi periplasmic flagella have both skeletal and motility functions. Proc Natl Acad Sci U S A 97, 10899–10904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motaleb MA, Liu J, and Wooten RM (2015). Spirochetal motility and chemotaxis in the natural enzootic cycle and development of Lyme disease. Curr Opin Microbiol 28, 106–113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motaleb MA, Miller MR, Li C, Bakker RG, Goldstein SF, Silversmith RE, Bourret RB, and Charon NW (2005). CheX is a phosphorylated CheY phosphatase essential for Borrelia burgdorferi chemotaxis. J Bacteriol 187, 7963–7969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motaleb MA, Sultan SZ, Miller MR, Li C, and Charon NW (2011). CheY3 of Borrelia burgdorferi is the key response regulator essential for chemotaxis and forms a long-lived phosphorylated intermediate. J Bacteriol 193, 3332–3341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy GE, Leadbetter JR, and Jensen GJ (2006). In situ structure of the complete Treponema primitia flagellar motor. Nature 442, 1062–1064. [DOI] [PubMed] [Google Scholar]
- Paulick A, Koerdt A, Lassak J, Huntley S, Wilms I, Narberhaus F, and Thormann KM (2009). Two different stator systems drive a single polar flagellum in Shewanella oneidensis MR‐ 1. Mol Microbiol 71, 836–850. [DOI] [PubMed] [Google Scholar]
- Qin Z, Lin WT, Zhu S, Franco AT, and Liu J (2016). Imaging the motility and chemotaxis machineries in Helicobacter pylori by cryo-electron tomography. J Bacteriol 199, e00695–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin Z, Tu J, Lin T, Norris SJ, Li C, Motaleb MA, and Liu J (2018). Cryo-electron tomography of periplasmic flagella in Borrelia burgdorferi reveals a distinct cytoplasmic ATPase complex. PLoS Biol 16, e3000050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raddi G, Morado DR, Yan J, Haake DA, Yang XF, and Liu J (2012). Three-dimensional structures of pathogenic and saprophytic Leptospira species revealed by cryo-electron tomography. J Bacteriol 194, 1299–1306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radolf JD, Caimano MJ, Stevenson B, and Hu LT (2012). Of ticks, mice and men: understanding the dual-host lifestyle of Lyme disease spirochaetes. Nat Rev Microbiol 10, 87–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sultan SZ, Manne A, Stewart PE, Bestor A, Rosa PA, Charon NW, and Motaleb MA (2013). Motility is crucial for the infectious life cycle of Borrelia burgdorferi. Infect Immun 81, 2012–2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sultan SZ, Sekar P, Zhao X, Manne A, Liu J, Wooten RM, and Motaleb MA (2015). Motor rotation is essential for the formation of the periplasmic flagellar ribbon, cellular morphology, and Borrelia burgdorferi persistence within Ixodes scapularis tick and murine hosts. Infect Immun 83, 1765–1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki H, Yonekura K, and Namba K (2004). Structure of the rotor of the bacterial flagellar motor revealed by electron cryomicroscopy and single-particle image analysis. J Mol Biol 337, 105–113. [DOI] [PubMed] [Google Scholar]
- Welch M, Oosawa K, Aizawa S, and Eisenbach M (1993). Phosphorylation-dependent binding of a signal molecule to the flagellar switch of bacteria. Proc Natl Acad Sci U S A 90, 8787–8791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan J, Branch RW, Hosu BG, and Berg HC (2012). Adaptation at the output of the chemotaxis signalling pathway. Nature 484, 233–236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang K, Qin Z, Chang Y, Liu J, Malkowski MG, Shipa S, Li L, Qiu W, Zhang J-R, and Li C (2019). Analysis of a flagellar filament cap mutant reveals that HtrA serine protease degrades unfolded flagellin protein in the periplasm of Borrelia burgdorferi. Mol Microbiol 0(0), 1–19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao X, Norris SJ, and Liu J (2014). Molecular architecture of bacterial flagellar motor in cells. Biochemistry 53, 4323–4333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao X, Zhang K, Boquoi T, Hu B, Motaleb MA, Miller KA, James ME, Charon NW, Manson MD, Norris SJ, et al. (2013). Cryoelectron tomography reveals the sequential assembly of bacterial flagella in Borrelia burgdorferi. Proc Natl Acad Sci U S A 110, 14390–14395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu S, Nishikino T, Hu B, Kojima S, Homma M, and Liu J (2017). Molecular architecture of the sheathed polar flagellum in Vibrio alginolyticus. Proc Natl Acad Sci U S A 114, 10966–10971. [DOI] [PMC free article] [PubMed] [Google Scholar]