

Abstract
Chronic wounds require extensive healing time and place patients at risk for both infection and amputation. Recently, a fresh hypothermically-stored amniotic membrane (HSAM) was developed and has subsequently shown promise in its ability to effectively heal chronic wounds. The purpose of this study is to investigate the mechanisms of action that contribute to wound healing responses observed with HSAM. A proteomic analysis was conducted on HSAM, measuring 25 growth factors specific to wound healing within the grafts. The rate of release of these cytokines from HSAMs were also measured. To model the effect of these cytokines and their role in wound healing, proliferation and migration assays with human fibroblasts and keratinocytes were conducted, along with tube formation assays measuring angiogenesis using media conditioned from HSAM. Additionally, the cell-matrix interactions between fibroblasts and HSAM were investigated. Conditioned media from HSAM significantly increased both fibroblast and keratinocyte proliferation and migration and induced more robust tube formation in angiogenesis assays. Fibroblasts cultured on HSAMs were found to migrate into and deposit matrix molecules within the HSAM graft. These collective results suggest HSAM positively affects various critical pathways in chronic wound healing; lending further support to promising qualitative results seen clinically and providing further validation for ongoing clinical trials.
Keywords: hypothermically-stored amniotic membrane (HSAM), chronic wounds, chronic wound healing, amnion, regenerative healing
Introduction
Diabetic foot ulcers (DFUs) are a major co-morbidity of diabetes, often resulting from poorly controlled hyperglycemia. There are 29.1 million Type II diabetics in the United States (9.3% of the total population) (1), and with DFUs at a lifetime incidence rate of 15%−25% (2), this complication accounts for healthcare costs in excess of 9–13 billion dollars annually (3,4). In many cases, DFUs lead to amputations which are associated with mortality rates between 30%−50% at 1 year (5). DFUs are not only difficult to close, owing to the complex diabetic wound environment, but they are also expensive to treat, with the cost of treating a DFU is estimated to run in excess of $18,000 per case (6). Improving healing rates and tissue quality of regenerated tissue in patients with these chronic wounds would not only improve patient quality of life but also reduce the overall burden to the healthcare system.
Under normal conditions, wound healing proceeds through four specifically timed phases: hemostasis, inflammation, repair, and remodeling (7). Chronic wounds develop when the healing process becomes stalled, most often in the inflammatory stage, and fails to progress though the subsequent phases of healing (8). Chronic wounds are characterized by prolonged expression of high levels of pro-inflammatory cytokines including tissue necrosis factor alpha (TNF-α), interleukin-6 (IL-6) and interleukin- 1 beta (IL-1β). This imbalance in inflammatory cytokines then leads to a prolonged recruitment period for macrophages, neutrophils and mast cells at the wound site. These cells, in turn, produce high levels of reactive oxidative species (ROS) and proteases.
Of the proteases produced within a chronic wound, matrix metalloproteases (MMPs) are of interest due to their multifunctional role. MMPs are an integral part of the normal wound healing process, serving to function in several capacities including: eliminating damaged proteins, destroying the provisional extracellular matrix (ECM), and facilitating cell migration and tissue remodeling (9). However, in the chronic wound environment, the influx of cells and the increase in pro-inflammatory cytokines leads to elevated and prolonged expression of MMPs and down regulated expression of MMP inhibitors (tissue inhibitors of matrix metalloproteinases, TIMPs). Many studies have shown that levels of MMPs are higher in the exudates of chronic wounds than in those of acute wounds, pointing to a potential mechanism underlying the chronic nature of DFUs (10). In a study by Muller et.al. (9) evaluating MMPs longitudinally in diabetic foot ulcers, it was found that despite similar initial levels of MMPs, good healers were characterized by a spike in MMP-1 at 2 weeks and a marked decrease in MMPs by week 4 (compared to no significant changes in MMP levels over time in poor healers). These studies point to the careful regulation of these proteases as an essential component to the successful healing of chronic wounds, since increased levels of proteases collectively cause an excessive break down of ECM, growth factors and growth factor specific receptors (8,11).
A key consequence of the destruction of growth factors and their receptors is a drop in the mitogenic activity of cells and a decrease in the recruitment of additional cells to the wound bed (12,13). Fibroblasts, which are known to be the building blocks of dermal tissue, are one example of cells effected by the chronic wound environment (14). Fibroblasts from diabetic patients have been shown to have a decreased responsiveness to growth factors (15), resulting in a lower proliferative response and deficiency in reorganizing the ECM. The chronic wound environment also detrimentally affects keratinocytes. Comparisons of keratinocytes obtained from healthy skin and DFUs have indicated that there are significant changes in many of the signaling pathways associated with apoptosis, migration and proliferation (16). Keratinocytes analyzed along the margins of DFUs have been found to be hyper-proliferative and lacking in their ability to migrate and completely differentiate (17–19).
The cellular processes involved in effective wound healing require a significant amount of oxygen, and this oxygen is received through normal blood flow. In many chronic wounds, the oxygen levels needed to proceed with efficient wound healing are unavailable, due to underlying ischemic conditions. Ischemic conditions are often a consequence of diabetic vascular complications which can affect the blood flow to the wound site (20), and cause deficiencies in blood vessel formation in peripheral tissues (21). Another consequence of the proteolytic environment within chronic wounds is the inhibition of angiogenesis. This occurs through the destruction of ECM proteins and cell adhesion molecules necessary for blood vessel formation, and the destruction of angiogenic growth factors (22), consequently effecting the delivery of oxygen to the wound microenvironment, impeding normal healing. (23).
Clinically, there are many types of advanced dressings and biologic products used to treat difficult to heal diabetic foot ulcers. One category that has grown in utilization recently is cellular tissue products, specifically those derived from placental tissues. These membranes in their native configuration serve to surround and protect the fetus; placental derived grafts consist of one or more layers of amnion and/or chorion. With consent from the mother, these membranes are collected post-delivery of the full-term, healthy baby. These placental-derived allografts are especially suited for chronic wound healing, because they are known to naturally address many of the contributing factors of chronic wound development, including: suppressing dysregulated/uncontrolled inflammatory responses, increasing levels of MMP inhibitors in the wound environment, stimulating proliferation and migration of important cell types, and promoting angiogenesis (24–26). Current processing technologies focus on dehydration or cryopreservation of placental derived membranes for clinical use; the goal of this paper is to evaluate wound healing responses elicited using a novel hypothermically stored amniotic membrane (HSAM).
Methods
HSAM (Affinity ®, NuTech Medical ® Birmingham, AL) is a hypothermically stored allograft composed of amniotic membrane derived from human placenta. Placentas were donated with informed consent after planned cesarean sections, and all processing were completed in accordance with the Food and Drug Administration’s (FDA) Good Tissue Practices and American Association of Tissue Banks standards. All donors were tested to be free of infectious diseases, including human immunodeficiency virus, human T-lymphotropic virus I/II, hepatitis B and C, and syphilis. HSAM is aseptically processed and stored in a proprietary hypothermic storage solution using the Allofresh™ process.
In order to determine the growth factor content present within HSAMs, multiple 1cm2 samples from nine donors were assessed for a variety of cytokines and growth factors by utilizing a quantitative multiplex enzyme-linked immunosorbent assay (ELISA) proteomics microarray (RayBiotech, Inc., Norcross, GA). Growth factors evaluated in this study are thought to be relevant to wound healing and have previously been identified within placental derived tissue (25,26). For the purposes of this evaluation cytokines have been categorized into general functional areas (detailed in Table 1).
Table 1.
Relevant Cytokine Categories for Wound Healing
| Angiogenic |
| Acidic fibroblast growth factor (aFGF) |
| Basic fibroblast growth factor (bFGF) |
| Vascular endothelial growth factor (VEGF) |
| Endocrine gland-derived vascular endothelial growth factor (EG-VEGF) |
| Vascular endothelial growth factor D (VEGF-D) |
| Platelet-derived growth factor BB (PDGF-BB) |
| Angiopoietin (ANG) |
| Thrombospondin 1 (TSP-1) |
| Angiopoietin-2 (ANG-2) |
| Placental growth factor (PlGF) |
| Angiopoietin-like 4 (APL4) |
| Regenerative |
| Epidermal growth factor (EGF) |
| Hepatocyte growth factor (HGF) |
| Galectin-7 (GAL) |
| Insulin-like Growth Factor-1 (IGF-I) |
| Insulin-like Growth Factor-2 (IGF-II) |
| Transforming growth factor beta 1 (TGF-β1) |
| Transforming growth factor beta 3 (TGF-β3) |
| Insulin-like growth factor-binding protein 1 (IGFBP-1) |
| Insulin-like growth factor-binding protein 5 (IGFBP-5) |
| Anti-Inflammatory |
| Transforming growth factor alpha (TGF-α) |
| Tissue Inhibitor of Metalloproteinase 1 (TIMP-1) |
| Tissue Inhibitor of Metalloproteinase 2 (TIMP-2) |
| Interleukin 1 receptor antagonist (IL-1ra) |
| Interleukin 10 (IL-10) |
For the proteomics array, HSAM grafts were selected from 9 donors, unless otherwise specified. Grafts were washed in saline and then homogenized using a Retsch cryomill (Verder Scientific, Inc., Newtown, PA). After cryomilling, the tissue was incubated overnight in total protein extraction buffer with a protease inhibitor cocktail (EMD Millipore, Billerica, MA) at 4°C with agitation. Following overnight incubation, the supernatant was removed and loaded into the microarray chambers per the manufacturer’s instructions. The slides were imaged using a GenePix 4000B microarray scanner (Molecular Devices, Sunnyvale, CA), and scanned images were imported and analyzed using GenePix Pro 7 software (Molecular Devices, Sunnyvale, CA).
To investigate the release of growth factors from HSAM grafts over time, samples were cut to 1.5 × 1.5 cm2 sections and placed in a 48 well plate in a serum free media. The HSAM samples were cultured under standard conditions for 7 days with gentle rocking. At 4, 12, 24, 48, 72, 96, 120, and 168 hours the supernatant was collected, sterile filtered, and stored at −80°C. At the end of the experiment, cultured HSAM grafts were collected and also stored at −80°C. Culture media and HSAM grafts were then evaluated using a proteomics array, using methods described above.
To evaluate the effect of HSAM on cell proliferation, commercially available adult human fibroblasts or keratinocytes (Lonza, Walkersville, MD) were cultured in the presence of assay media conditioned with HSAM. Conditioned media (CM) was obtained by incubating HSAM in assay media for 5 days at 4°C at a concentration of 1 cm2 HSAM membrane per milliliter (mL) of assay media. Following incubation, assay media was filter sterilized prior to use. Media was stored at 4°C for up to 14 days. Cells were then seeded into 48 well plates at a concentration of 3,300 and 10,000 cells per well for fibroblasts and keratinocytes, respectively. CM was added to assay media at concentrations of 50%, 25%, and 10% (v/v) at 1, 4, 7, and 10 days of culture immediately following evaluation with AlamarBlue. Specifically, AlamarBlue assays were conducted according to manufacturer’s instructions (Invitrogen, Carlsbad, CA) by incubating cells in 350 μL of a 10% AlamarBlue working solution in Dulbecco’s Modified Eagle Medium with 2.5% FBS for 4 hours under standard culture conditions. Following incubation, fluorescence was measured in a plate reader from 100μL samples using a fluorescence excitation wavelength of 540–570 nm and fluorescence emission wavelength of 580–610 nm.
Cell migration assays were conducted with adult human dermal fibroblasts, keratinocytes, and HUVECs using a standard Boyden chamber assay (27). Three lots of fibroblasts were used; one from ZenBio (ZenBio, Inc., Research Triangle Park, NC) and two from Lonza (Lonza, Walkersville, MD). HUVECs were purchased from Lonza (Lonza, Walkersville, MD) and Life Technologies (Life Technologies, Carlsbad, CA). Keratinocytes were purchased from Lonza (Lonza, Walkersville, MD). Prior to the start of the assay, cells at less than or equal to 80% confluent, were serum-starved overnight. Reservoirs were loaded with either assay media alone (negative control), assay media + 10% FBS (positive control), or CM from HSAM at concentrations of 50%, 25%, and 10% (v/v). Prior to adding cells to the migration chambers, both the cell inserts and reservoirs were coated with 5 μg/ml Fibronectin (ThermoFisher, Waltham, MA) overnight at room temperature to promote initial cell attachment. Inserts and reservoirs were then washed three times with PBS. Next, cells were trypsinized and added to the top of the inserts at a concentration of 10,000 (for fibroblasts) or 20,000 (for HUVECs and keratinocytes) cells per insert and then incubated for 24 hours to allow for migration. Non-migrating cells remaining at the top of the cell inserts were removed with a cotton tip applicator, and cells which had migrated to the bottom of the insert were fixed with a 4% solution of neutral buffered formalin in PBS for 10 minutes prior to staining with a crystal violet solution. Images of the inserts were taken with an inverted microscope (Nikon Eclipse Ti, Tokyo, Japan), and representative images were used to count the number of migrated cells.
Tube formation assays were conducted using the angiogenesis starter kit (Life Technologies, Carlsbad, CA) per the manufacturer’s instructions. Two lots of HUVECs (passages 3–5) were used in these experiments (Life Technologies, Carlsbad, CA) (28,29). HUVECs were expanded in Medium 200 with large vessel endothelial supplement (Life Technologies, Carlsbad, CA) and were passaged at least once after thawing and prior to beginning the tube formation assay. Once cells had reached 80% confluency, HUVECs were seeded into 48-well plates with a reduced growth factor basement membrane matrix (Geltrex, Life Technologies, Carlsbad, CA) at a concentration of 25,000 cells/cm2. Cells were then cultured overnight with either CM from HSAM or assay media alone (EBM). Conditioned media was obtained as described in detail above. After overnight culture, cells were then fixed in methanol for 1 minute, rinsed thrice in distilled water and imaged via phase-contrast with an inverted microscope (Nikon Eclipse Ti, Tokyo, Japan). Images were then imported into ImageJ (NIH, Bethesda, MD) and average tube length was quantified.
For all quantitative mechanistic assays (proliferation, migration and angiogenesis), statistical analysis was conducted using a one-way ANOVA with a post-hoc Tukey test where p<0.05 was considered significant. Throughout * denotes p<0.05, ** denotes p< 0.01, *** denotes p<0.001 and **** denotes p< 0.0001. For qualitative imaging, representative images of all groups were taken and presented.
To qualitatively analyze how fibroblasts interacted with HSAM, 50,000 human dermal fibroblasts were seeded onto HSAM grafts (2.5cm x 2.5 cm). Grafts were then cultured in 6 well plates for two weeks under standard culture conditions. Post 2-week culture, grafts were fixed in 4% neutral buffered formalin for 24 hours. Following paraffin embedding, serial sections 5μm thick were cut from the tissue blocks and floated onto charged glass slides (Super-Frost Plus, Fisher Scientific, Pittsburgh, PA) and dried overnight at 60° C. Sections were stained for hemotoxylin and eosin (H&E), Masson’s trichrome, Verhoeff’s stain, and Alcian blue. All sections for immunohistochemistry were deparaffinized and hydrated using graded concentrations of ethanol to deionized water. For fibronectin immunohistochemistry, antigen retrieval was performed using a solution of Proteinase K (P6556, 1:50 dilution in TE-CaCl2 buffer pH8) for 15 min at 37°C. For Collagen I and 3 immunohistochemistry, tissues sections were subjected to antigen retrieval using a 0.01 M Tris-1mM EDTA buffer (pH 9) at 70°C for 20 min. Following antigen retrieval, all sections were washed gently in deionized water, then transferred to a 0.05 M Tris-based solution in 0.15M NaCl with 0.1% v/v Triton-X-100, pH 7.6 (TBST). Endogenous peroxidase was blocked with 3% hydrogen peroxide for 20 min. To reduce further nonspecific background staining, slides were incubated with 3% normal goat serum for 30 min (Sigma-Aldrich, St. Louis, MO) at room temperature. All slides then were incubated at 4° C overnight with various antibodies (Fibronectin, PA5–29578, Thermo Fisher, 1:100; Collagen 1, ab34710, Abcam, 1;100; Collagen 3, ab7778, Abcam, 1:100). Negative controls were produced by eliminating the primary antibodies from the diluents. After washing with TBST, sections were then incubated with the matching secondary antibodies conjugated with horseradish peroxidase. Diaminobenzidine (DAB; Scy Tek Laboratories, Logan, UT) was used as the chromagen and hematoxylin (Richard-Allen Scientific, Kalamazoo, MI) as the counterstain. All stained samples were visualized using an inverted Microscope (Nikon Eclipse Ti, Tokyo, Japan Nikon, Melvile, NY) with a 40x objective.
Results
Proteomic microarrays confirmed physiologically relevant levels of numerous growth factors and cytokines important for wound healing (Table 1). These growth factors were broken down using general categories to describe their activity, including: angiogenic, regenerative and anti-inflammatory growth factors (Figure 1. average ± standard deviation, n=9 unless otherwise noted). There is some sample-to-sample variability observed in the levels of growth factors and cytokines present in the tissue. This level of variability is expected and has been observed in previous studies of dehydrated human amnion-chorion membranes (dHACM) (25,30) and of cryopreserved human amniotic membrane (cHAM) (31–33). Many of the same growth factors are found to be present in cHAM, dHACM, and HSAM. However, differences have been noted which may be contributed to by several factors including: degradation of growth factors or cytokines occurring as a result of tissue processing (dehydration, lyophilization, and sterilization), the layers included in the product (amnion vs. amnion and chorion), as well as study-related factors such as protein extraction methods, methods for evaluation of growth factors, and sample size. The most abundant growth factors present in the HSAM were TSP-1, APL4, and IGF-1 in nearly all of the samples that were tested, suggesting that while specific levels of growth factors in the tissues may vary to some degree, relative concentrations of these most abundant factors are consistent.
Figure 1.

Multiplex ELISA proteomics microarray measuring growth factors and cytokines relating to mechanisms of wound healing including: a) angiogenic growth factors, (b) regenerative growth factors, and c) anti-inflammatory growth factors. For all growth factors and cytokines, results are shown as average ± standard deviation (n=9, except where marked with º n=4)
Subsequently, we were interested in evaluating the time course for release of growth factors from HSAMs over a 7-day time course. All growth factors analyzed in Figure 1 were included in this analysis and the total growth factor and cytokine release over 7 days was approximately 70.6 ± 19.9% of total measured cytokines (Figure 2A). Key growth factors including bFGF, HGF, IL-1ra, TGF-β3, and TIMP-1/2 were evaluated and graphed individually (Figure 2B-F). After 7 days, 91.7±5.6% (n=3) of the bFGF had been released; bFGF is widely known to accelerate wound healing, primarily though it mitogenic effects on cells (34) (Figure 2B). HGF is particularly important to the wound healing environment due to its role in stimulating proliferation and cell motility in cells of epithelial origin (35,36); in our study 89.9±4.5% (n=3) of HGF was released from HSAM after 7 days (Figure 2C). IL-1ra is a natural inhibitor of pro-inflammatory IL-1B and is thought to also play a role in re-epithelialization. Of interest, delayed healing has been observed in IL-1ra deficient mice as evidenced by problems with collagen deposition and vascularization (37). Over the course of 7 days, approximately 92.2±9.9% (n=3) IL-1ra was released from HSAM (Figure 2D). After 7 days, approximately 97.8±3.7% (n=3) of TGF-β3 in HSAM had been released (Figure 2E). TGF-β3 is thought to assists in wound healing by reducing fibrosis and scarring at the wound site (38,39).
Figure 2.

Release of growth factors from HSAM: a) cumulative release curve for all 25 growth factors listed in Table 1, b) basic fibroblast growth factor (bFGF), c) hepatocyte growth factor (HGF), d) interleukin1 receptor antagonist (IL-1ra), e) transforming growth factor beta 3 (TGF-β3), and f) tissue inhibitor of metalloproteinase 1 & 2 (TIMP-1/2). Cytokine concentrations are shown as average ± standard deviation for all time points, and total % average ± standard deviation released at 7 days is inset for all release curves (n=3).
Another important component of the wound healing process is appropriate regulation of protease activity. Overexpression of MMPs is thought to be one of the causative factors for chronic wounds; therefore inhibitors of MMPs (TIMPs) and their availability are of particular interest (40). We found that the majority of TIMP-2 (93.6±4.3%, n=3) was released from the membrane by day 7; we also found that TIMP-1 was released from the membrane, however only 10.7±5.4% (n=3) of the total quantity of TIMP-1 was released by day 7. The low release rate of TIMP-1 compared to TIMP-2 may be, in part, explained by the interaction of TIMP-1 with the β1 integrin (41). Because of its interaction with the ECM, the release profile of TIMP-1 in particular is likely much different in vivo, due to interactions of the graft with the chronic wound environment and subsequent break down of the ECM.
In addition to studying the growth factor/cytokine profile and their respective release characteristics, we also investigated the effects released cytokines have on cell types important to wound healing. Specifically, we investigated the effects of cytokines released from HSAM on the behavior of human dermal fibroblasts, human keratinocytes, and human endothelial cells. Fibroblast migration and proliferation is thought to be an indicator of healing in wounds (7,42,43). In Figure 3 A&B, it was found that fibroblasts proliferate more quickly in media conditioned from HSAM than in assay media alone, which represents the baseline control. Quantitatively, it was found that 50%, 25%, and 10% conditioned media (CM) significantly increased fibroblast proliferation at 7, 10 and 14 days, although there were no measurable differences between the CM groups of different concentrations. Qualitative images were taken at 14 days, and these images confirmed HSAM CM groups had higher density of cells present than the assay media group (Figure 3B).
Figure 3.
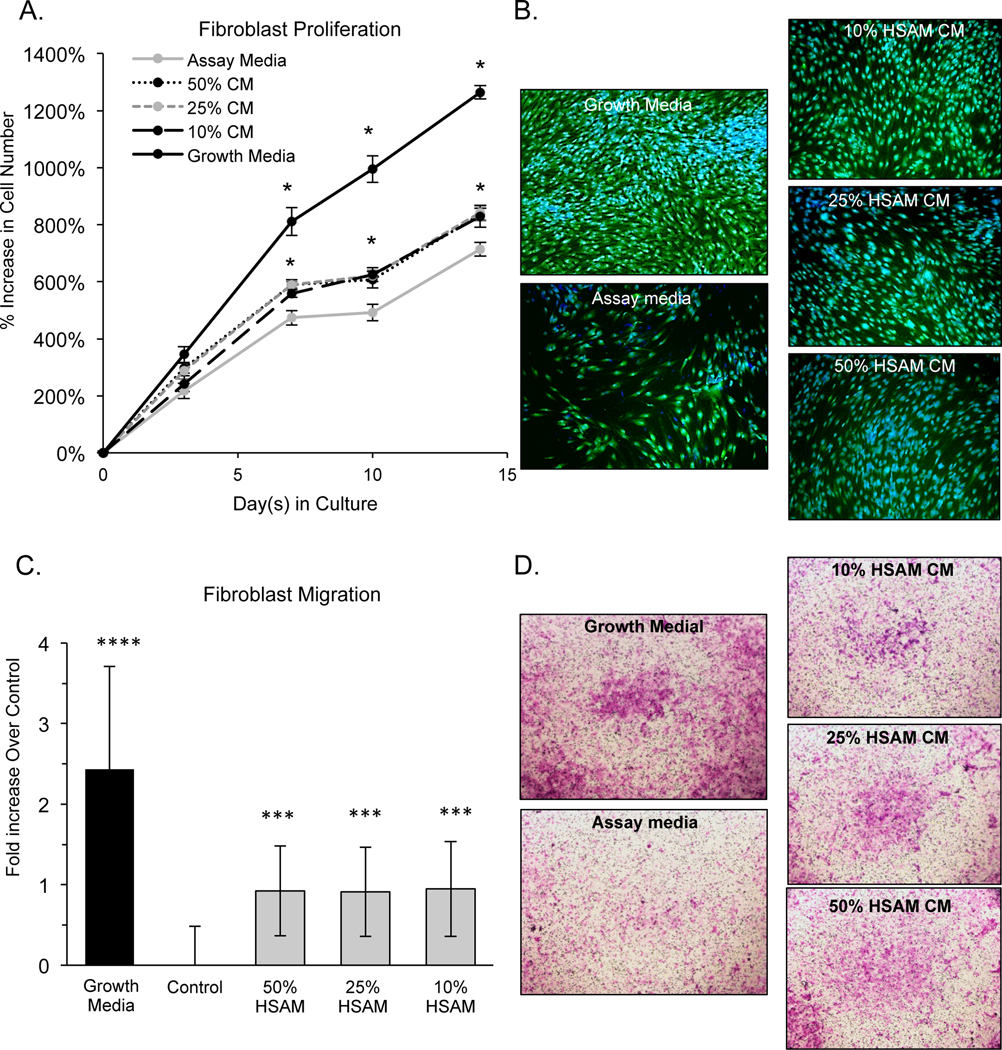
Evaluation of HSAM effects on fibroblasts: a) quantitative proliferation assay in assay media alone or HSAM conditioned media in assay media (with 50%, 25%, and 10% conditioned media by volume), b) qualitative analysis of proliferation assay with CFDA (green) and Dapi (blue) staining at 14 days, c) quantitative analysis of fibroblast migration over 24 hours in growth media, assay media, and 50%, 25% and 10% HSAM conditioned media by volume, and d) confirmatory images of cell migration with cells stained with crystal violet. For quantitative assays significance compared to assay media is denoted by: * (p<0.05), **(p<0.01), *** (p<0.001), and **** (p<0.0001).
Additionally, migration of fibroblasts through a microporous membrane was evaluated. Fibroblast migration assays showed that the presence of HSAM CM resulted in a significant increase in migration relative to assay media alone. Interestingly, the dose of CM (50%, 25%, or 10%) did not affect the increase in fibroblast migration with all groups showing between 0.91 and 0.95-fold increase over assay media alone. Crystal violet stained cells qualitatively confirmed greater migration in groups with HSAM CM than in assay media alone (Figure 3D).
Human epidermal keratinocytes (HEKs) are essential to wound closure; in the normal healing process HEKs proliferate and migrate into the wound and form an epithelial layer (44–46). To model this in vitro, we assessed the effect HSAM CM would have on HEK proliferation and migration. (Figure 4). After 7 days, all CM groups evaluated (10%, 25%, and 50% HSAM CM) resulted significantly increased cell proliferation relative to assay media alone. Migration of keratinocytes is considered an important step in wound healing as they play an integral role in re-epithelialization of the wound bed (47). To model this effect in vitro, HEK migration was evaluated using a Boyden chamber assay (Figure 4B). It was found that 50% and 25% HSAM CM stimulated significantly increased cell migration compared to assay media, 340% and 232%, respectively. Qualitatively, these changes were confirmed with crystal violet staining of cells attached to the Boyden chamber membrane (Figure 4C).
Figure 4.

Evaluation of HSAM effects on human epidermal keratinocytes (HEK). a) Quantitative analysis of proliferation of HEKs cultured under various conditions including growth media, assay media, and HSAM conditioned media (50%, 25%, and 10%). Migration of HEKs in response to growth media, assay media, and 50%, 25%, and 10% HSAM CM evaluated quantitatively (b) and qualitatively with crystal violet staining (c). For quantitative assays significance compared to assay media is denoted by: * (p<0.05), ** (p<0.01), *** (p<0.001), and **** (p<0.0001).
Angiogenesis, or the formation of new blood vessels, is essential for wound healing (48). However, this process is often disrupted in chronic wounds. In this study, HSAM CM or assay media was applied to a monolayer of HUVECs and formation of tubules was assessed (Figure 5). HSAM CM at concentrations of 50% and 25% significantly increased average tubule length by 104% and 95%, respectively. 10% and 1% HSAM CM showed no appreciable increases in tubule length. Representative phase-contrast images are shown in Figure 5B for each of the groups analyzed, with qualitative increases in tube formation shown in higher concentrations of HSAM CM.
Figure 5.

Impact of HSAM on angiogenesis as measured by tube formation:.a) Tube formation was evaluated after culture of HUVECs overnight with or without CM from HSAM (50%, 25%, 10% and 1%), and b) representative phase-contrast images of tube formation were taken. Image J was used to quantify tube length and data is presented as average ± standard deviation. Significance is denoted by * (p<0.05).
To model the interaction of fibroblasts with HSAM, we cultured fibroblasts on HSAM for up to 2 weeks and subsequently evaluated the tissue using H&E, Masson’s trichrome, Verhoeff’s stain, and Alcian blue (Figure 6). Immunohistochemistry was also conducted for collagen I, III, and fibronectin (Figure 7). Images show a series of histological results for HSAM with (Figures 6B, 6D, 6F, 6H, 7B, 7D, and 7F) and without (Figures 6A, 6C, 6E, 6G, 7A, 7C, and 7E) fibroblasts seeded for up to 2 weeks. Based on the H&E and Masson’s trichrome images, it is clear that fibroblasts readily infiltrate and proliferate within the HSAM grafts. Masson’s trichrome and Verheoff’s staining showed deposition of collagen and elastin into the HSAM by fibroblasts. While HSAM contains a high quantity of glycosaminoglycans, alcian blue images do not indicate deposition of additional glycosaminoglycans by fibroblasts. HSAM was found to contain high amounts of collagen I throughout stromal layer, collagen III specifically localized to the basement membrane layer and the spongy layer, and Fibronectin within the basement membrane layer and around cells. Fibroblasts cultured on HSAM deposited additional collagen III and Fibronectin throughout the HSAM. While we also expected to see fibroblasts producing collagen I, the high levels of native collagen I found throughout the HSAM make it difficult to detect an additional protein deposition.
Figure 6.

Qualitative evaluation of fibroblast interaction with HSAM was completed using staining. HSAM was cultured with or without fibroblasts for two weeks, and samples were processed and stained with Hematoxylan and Eosin (H&E), Masson’s Trichrome, Alcian Blue, or Verhoeff’s Stainor immunohistochemical stains. Representative images were taken of H&E(a&b), collagen IMasson’s Trichrome (c&d), collagen IIAlcian Blue (e&f), and fibronectin Verhoeff’s stain (g&h). Black arrows point to fibronectin deposited throughout the matrix.
Figure 7.

Qualitative evaluation of fibroblast interaction with HSAM was completed using immunohistochemical stains. Representative images were taken of collagen I (a&b), collagen II (c&d), and fibronectin (e&f). Black arrows point to fibronectin deposited throughout the matrix.
Discussion
The use of placental based tissues is proposed to aid in healing through various mechanisms including: reduction in inflammation, enhancement of cell migration into the wound environment, stimulation of cell proliferation, increased production of ECM, and improved angiogenesis (25,49–51). A significant part of the function of these tissues is attributed to the growth factor content released from the membranes. Selected growth factors and cytokines of interest are shown in Table 1; while this list is not comprehensive of all molecules found in placental-based tissues, these molecules were selected due to the important role they play in wound healing. Some growth factors and cytokine concentrations are lower than in singular recombinant growth factor therapies used for chronic wounds, but it is important to note that there may be a substantial benefit in the sustained delivery of multiple cytokines with different roles/signaling processes in the healing wound (52,53).
The management of inflammation is essential to resolving chronic wounds. Amniotic membranes and the cells within these membranes have been shown to elicit an anti-inflammatory response. Cell derived from amniotic membranes have been shown to down-regulate the production of certain inflammatory cytokines including TNF-α, CXCL10, CXCL9, and CCL5 (54,55). The suppression of TNF-α is of particular interest since TNF-α has been found to be elevated in non-healing DFUs (56). Amniotic membranes have been shown to suppress the production of TGF-β1, IL-8, and granulocyte macrophage colony-stimulating factor by fibroblasts which are known to be stimulated under inflammatory conditions (57). Another relevant cytokine know to trigger inflammatory responses within chronic wounds, IL-1β, was found to be significantly reduced in gingival crevicular fluid following the application of amniotic membranes with bone grafting for periodontal pockets in a 30 patient clinical study (58). In a mouse bleomycin injury model, amnion derived epithelial cells have been shown to limit macrophage infiltration into the lungs and to also drive a majority of pulmonary macrophages to the anti-inflammatory M2 phenotype (59). The shift to M2 polarization in response to amnion derived cells is relevant to DFUs since M2 and M2-like macrophages induce anti-inflammatory, regulatory, and reparative functions that lead to wound closure (60–62). While the exact mechanisms responsible for these anti-inflammatory effects remains unclear, potential sources include release of interleukins from the membrane including both IL-10 and IL-1ra, which were measured in HSAM. IL-10 is known to counteract the effects of several pro-inflammatory cytokines including IL-6, IL-1, IL-8 and TNF-α (63), and IL-1ra is a potent inhibitor of IL-1, shown to suppress the inflammatory response triggered by IL-1 (64). While not measured in this study, we suspect the presence of heavy chain hyaluronic acid (65), prostaglandin E2 (66), and macrophage migration-inhibitory factor (67,68), may also play a role in the anti-inflammatory effects observed with HSAM.
An essential part of resolving chronic wounds is the reduction of proteases within the wound environment to prevent the destruction of growth factors and ECM proteins necessary for wound healing. MMP-9 has been found to be elevated in non-healing DFUs (56), and the addition of inhibitors to MMPs (TIMPs) to the chronic wound environment has been shown to be beneficial to wound healing (9,25,69). Amniotic membranes have been previously been shown to inhibit MMP activity (70,71), and here we have demonstrated the presence and the release curves of TIMP-1 and TIMP-2 within HSAM. In addition to TIMP-1 and 2, possible mechanisms behind the protease inhibition observed with amniotic membranes include contributions by type-1 plasminogen activator inhibitor and thrombospondin-1 (24).
The destruction of essential growth factors and their receptors is another consequence of excessive protease levels within chronic wounds. This consequently leads to cellular dysregulation especially with fibroblasts and keratinocytes. For fibroblasts, a marked decrease in cellular migration, proliferation, and ECM deposition has been observed (72,73). Here we have demonstrated that CM from HSAM significantly improves both fibroblast proliferation and migration. Fibroblasts were also shown to migrate into and deposit ECM proteins within HSAMs. The likely cause of this proliferative and migratory response of fibroblasts to CM is from the release of FGF, TGF-β1, TFG-β3, and IGF-1 from the HSAM, which have been shown to lead to the activation and migration of fibroblasts and stimulate their proliferation (42). Keratinocytes along the borders of chronic wounds have also been found to demonstrate abnormal behavior including failure to differentiate and migrate (17). Here we have demonstrated that HSAM contains several growth factors know to stimulate keratinocyte migration and proliferation including EGF, PDGF-BB, and TGFβ−1. More importantly we have shown that growth factors/cytokines found within CM from HSAM are active and can promote both proliferation and migration of keratinocytes. In a recent study by Zhao et al., human amniotic epithelial cells were found to improve keratinocyte migration and proliferation. This increase in keratinocyte migration and proliferation was found to be primarily the result of EGF and KGF produced by these cells which, in turn, activated the ERK, JNK, and AKT pathways in keratinocytes (74). Fibroblasts and keratinocytes are known to interact in the wound environment (35,75). Suppression of fibroblast proliferation and migration in chronic wounds have been shown to result in changed cytokine and protease release profiles (76,77), and these changes result in deleterious impacts on keratinocyte proliferation and migration (78). We expect the combined effects of HSAM CM on both fibroblasts and keratinocytes to promote significant improvements clinically in chronic wound healing.
Prolonged hypoxia in the wound environment can prevent wounds from healing normally; therefore, angiogenesis is critical to restore oxygen and nutrients to the injured tissue (7). Because many patients with chronic wounds, especially DFUs, have underlying vascular disease, lack of effective angiogenesis can be particularly problematic in healing these patients. In addition to these underlying issues, there are several anti-angiogenic factors present within chronic wounds including inflammatory factors, proteases, and cell death mediators which deter pro-angiogenic processes (22). We expect HSAM may mitigate both the prolonged pro-inflammatory response and high protease content within the wound to create an environment more favorable to angiogenesis. HSAM contains numerous angiogenic growth factors including FGF, VEGF, PDGF-BB, ANG, PLGF, APL4, and TSP1. The results from in vitro tube formation assays reflected the activity of high levels of angiogenic growth factors; these results support the hypothesis that the application of HSAMs to chronic wounds may promote angiogenesis resulting in improved wound healing and closure.
Placental-derived membranes have a long history in wound care (79,80); however most current commercially available membranes are processed in such a way that they do not maintain native viable cell populations. While these membranes have shown positive effects on healing in chronic wound care (51,81), the absence of a viable cell population prevents these grafts from dynamically responding to the chronic wound environment (82). Numerous studies have shown that the various cell types populating the amniotic membrane, which are present within HSAM, produce factors which act to reduce inflammation, improve cell proliferation and migration, and improve angiogenesis (54,55,59,66,67). Recent clinical data suggests that cell containing amniotic grafts may be more effective in treating chronic wounds compared to non-viable placental-derived alternatives (83). Future studies will focus on how the cell populations within HSAM respond to chronic wound environments through in vitro models in the context of inflammation and in vivo studies utilizing an ischemic animal model which more closely mimics the chronic wound environment (84).
Conclusion
Hypothermically-stored amniotic membranes offer a new treatment alternative to promote healing in chronic wounds; these membranes deliver a native extracellular matrix, growth factors and cytokines, and a viable cell content. In this study, we have focused on the overall activity of the grafts primarily by using media conditioned by the grafts to study mechanisms of action. We have reported concentrations of numerous growth factors and cytokines known to contribute to wound healing and detailed their released from HSAM over 7 days. Furthermore, we have shown that HSAM CM significantly increases fibroblast and keratinocyte proliferation and migration, suggesting its potential to improve healing in chronic wounds where the normal activity of these cells is known to be suppressed. Finally, this study shows that HSAM promotes increased tube formation in HUVECs suggesting a potential mechanism for HSAM to promote and improve angiogenesis. In sum, we believe hypothermically-stored amniotic grafts are a compelling option for promoting pro-healing activities spanning various functions including: reducing inflammation and protease activity as well as promoting angiogenesis and cellular responses.
Acknowledgements
The authors would like to acknowledge the support of the University of Alabama at Birmingham’s Pathology Core Research lab, in particular Dr. Dezhi Wang, for their assistance with all histology and immunohistochemistry. This study was supported and funded by NuTech Medical, Birmingham AL. JPM, JBV, KCM are employees of NuTech Medical.
Abbreviations:
- aFGF
Acidic fibroblast growth factor
- ANG
Angiopoietin
- ANG-2
Angiopoietin-2
- APL4
Angiopoietin-like 4
- bFGF
Basic fibroblast growth factor
- CM
Conditioned media
- cHAM
cryopreserved human amniotic membrane
- DFU
diabetic foot ulcers
- dHACM
dehydrated human amnion-chorion membrane
- ECM
extracellular matrix
- ELISA
enzyme-linked immunosorbent assay
- EG-VEGF
Endocrine gland-derived vascular endothelial growth factor
- EGF
Epidermal growth factor
- GAL
Galectin-7
- HEK
Human epidermal keratinocytes
- HGF
Hepatocyte growth factor
- HSAM
hypothermically stored amniotic membrane
- IL-1β
interleukin- 1 beta
- IL-6
interleukin-6
- IGF-1
Insulin-like Growth Factor-
- IGF-II
Insulin-like Growth Factor-2
- IGFBP-1
Insulin-like growth factor-binding protein 1
- IGFBP-5
Insulin-like growth factor-binding protein 5
- IL-1ra
Interleukin 1 receptor antagonist
- IL-10
Interleukin 10
- MMPs
matrix metalloproteases
- PIGF
Placental growth factor
- PDGF-BB
Platelet-derived growth factor BB
- TBST
Tris-based solution
- TSP-1
Thrombospondin 1
- TIMPs
tissue inhibitors of matrix metalloproteinases
- TNF-α
tissue necrosis factor alpha
- TGF-β1
Transforming growth factor beta 1
- TGF-β3
Transforming growth factor beta 3
- VEGF
Vascular endothelial growth factor
- VEGF-D
Vascular endothelial growth factor D
References
- 1.Centers for Disease Control and Prevention. National Diabetes Statistics Report: Estimates of Diabetes and Its Burden in the United States. Atlanta, GA; 2014. [Google Scholar]
- 2.Boulton AJM, Vileikyte L, Ragnarson-Tennvall G, Apelqvist J. The global burden of diabetic foot disease. Lancet (London, England) [Internet]. 2005. November 12;366(9498):1719–24. [DOI] [PubMed] [Google Scholar]
- 3.Brantley JN, Verla TD. Use of Placental Membranes for the Treatment of Chronic Diabetic Foot Ulcers. Adv wound care [Internet]. 2015. September 1;4(9):545–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Rice JB, Desai U, Cummings AKG, Birnbaum HG, Skornicki M, Parsons NB. Burden of diabetic foot ulcers for medicare and private insurers. Diabetes Care [Internet]. 2014;37(3):651–8. [DOI] [PubMed] [Google Scholar]
- 5.Fortington LV, Geertzen JHB, van Netten JJ, Postema K, Rommers GM, Dijkstra PU. Short and long term mortality rates after a lower limb amputation. Eur J Vasc Endovasc Surg [Internet]. Elsevier Ltd; 2013. July;46(1):124–31. [DOI] [PubMed] [Google Scholar]
- 6.Rice JB, Desai U, Ristovska L, Cummings AKG, Birnbaum HG, Skornicki M, et al. Economic outcomes among Medicare patients receiving bioengineered cellular technologies for treatment of diabetic foot ulcers. J Med Econ [Internet]. Informa UK Ltd.; 2015;18(8):586–95. [DOI] [PubMed] [Google Scholar]
- 7.Falanga V. Wound healing and its impairment in the diabetic foot. Lancet (London, England) [Internet]. 2005. November 12;366(9498):1736–43. [DOI] [PubMed] [Google Scholar]
- 8.McCarty SM, Percival SL. Proteases and Delayed Wound Healing. Adv wound care [Internet]. 2013. October;2(8):438–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Muller M, Trocme C, Lardy B, Morel F, Halimi S, Benhamou PY. Matrix metalloproteinases and diabetic foot ulcers: the ratio of MMP-1 to TIMP-1 is a predictor of wound healing. Diabet Med [Internet]. 2008. April;25(4):419–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lobmann R, Schultz G, Lehnert H. Proteases and the diabetic foot syndrome: mechanisms and therapeutic implications. Diabetes Care [Internet]. 2005. February;28(2):461–71. [DOI] [PubMed] [Google Scholar]
- 11.McCarty SM, Cochrane CA, Clegg PD, Percival SL. The role of endogenous and exogenous enzymes in chronic wounds: a focus on the implications of aberrant levels of both host and bacterial proteases in wound healing. Wound Repair Regen [Internet]. 2012. March;20(2):125–36. [DOI] [PubMed] [Google Scholar]
- 12.Grazul-Bilska AT, Johnson ML, Bilski JJ, Redmer DA, Reynolds LP, Abdullah A, et al. Wound healing: the role of growth factors. Drugs Today (Barc) [Internet]. 2003. October;39(10):787–800. [DOI] [PubMed] [Google Scholar]
- 13.Nwomeh BC, Yager DR, Cohen IK. Physiology of the chronic wound. Clin Plast Surg [Internet]. 1998. July 29;25(3):341–56. [PubMed] [Google Scholar]
- 14.Bainbridge P. Wound healing and the role of fibroblasts. J Wound Care [Internet]. 2013. August;22(8):407–8, 410–2. [DOI] [PubMed] [Google Scholar]
- 15.Loot MAM, Kenter SB, Au FL, van Galen WJM, Middelkoop E, Bos JD, et al. Fibroblasts derived from chronic diabetic ulcers differ in their response to stimulation with EGF, IGF-I, bFGF and PDGF-AB compared to controls. Eur J Cell Biol [Internet]. 2002. March;81(3):153–60. [DOI] [PubMed] [Google Scholar]
- 16.Hoke GD, Ramos C, Hoke NN, Crossland MC, Shawler LG, Boykin JV. Atypical Diabetic Foot Ulcer Keratinocyte Protein Signaling Correlates with Impaired Wound Healing. J Diabetes Res [Internet]. 2016;2016:1586927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Usui ML, Mansbridge JN, Carter WG, Fujita M, Olerud JE. Keratinocyte Migration, Proliferation, and Differentiation in Chronic Ulcers From Patients With Diabetes and Normal Wounds. J Histochem Cytochem [Internet]. 2008;56(7):687–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Brem H, Tomic-Canic M. Cellular and molecular basis of wound healing in diabetes. J Clin Invest [Internet]. 2007. May;117(5):1219–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Stojadinovic O, Brem H, Vouthounis C, Lee B, Fallon J, Stallcup M, et al. Molecular pathogenesis of chronic wounds: the role of beta-catenin and c-myc in the inhibition of epithelialization and wound healing. Am J Pathol [Internet]. 2005. July;167(1):59–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mustoe TA, O’Shaughnessy K, Kloeters O. Chronic wound pathogenesis and current treatment strategies: a unifying hypothesis. Plast Reconstr Surg [Internet]. 2006. June;117(7 Suppl):35S–41S. [DOI] [PubMed] [Google Scholar]
- 21.Martin A, Komada MR, Sane DC. Abnormal angiogenesis in diabetes mellitus. Med Res Rev [Internet]. 2003. March;23(2):117–45. [DOI] [PubMed] [Google Scholar]
- 22.Krisp C, Jacobsen F, McKay MJ, Molloy MP, Steinstraesser L, Wolters DA. Proteome analysis reveals antiangiogenic environments in chronic wounds of diabetes mellitus type 2 patients. Proteomics [Internet]. 2013. September;13(17):2670–81. [DOI] [PubMed] [Google Scholar]
- 23.Bao P, Kodra A, Tomic-Canic M, Golinko MS, Ehrlich HP, Brem H. The role of vascular endothelial growth factor in wound healing. J Surg Res [Internet]. 2009. May 15;153(2):347–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Litwiniuk M, Bikowska B, Niderla-Bielińska J, Jóźwiak J, Kamiński A, Skopiński P, et al. Potential role of metalloproteinase inhibitors from radiation‑sterilized amnion dressings in the healing of venous leg ulcers. Mol Med Rep [Internet]. 2012. October;6(4):723–8. [DOI] [PubMed] [Google Scholar]
- 25.Koob TJ, Rennert R, Zabek N, Massee M, Lim JJ, Temenoff JS, et al. Biological properties of dehydrated human amnion/chorion composite graft: implications for chronic wound healing. Int Wound J [Internet]. 2013. October;10(5):493–500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Steed DL, Trumpower C, Duffy D, Smith C, Marshall V, Rupp R, et al. Amnion-derived cellular cytokine solution: a physiological combination of cytokines for wound healing. Eplasty [Internet]. 2008. April 7;8(e18):e18. [PMC free article] [PubMed] [Google Scholar]
- 27.Chen H-C. Boyden chamber assay. Methods Mol Biol [Internet]. 2005;294(294):15–22. [DOI] [PubMed] [Google Scholar]
- 28.Arnaoutova I, Kleinman HK. In vitro angiogenesis: endothelial cell tube formation on gelled basement membrane extract. Nat Protoc [Internet]. 2010. April;5(4):628–35. [DOI] [PubMed] [Google Scholar]
- 29.DeCicco-Skinner KL, Henry GH, Cataisson C, Tabib T, Gwilliam JC, Watson NJ, et al. Endothelial cell tube formation assay for the in vitro study of angiogenesis. J Vis Exp [Internet]. 2014. September 1;91(91):e51312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Koob TJ, Lim JJ, Zabek N, Massee M. Cytokines in single layer amnion allografts compared to multilayer amnion/chorion allografts for wound healing. J Biomed Mater Res B Appl Biomater [Internet]. 2015. July [cited 2016 Jun 7];103(5):1133–40. [DOI] [PubMed] [Google Scholar]
- 31.Kruse FE, Joussen AM, Rohrschneider K, You L, Sinn B, Baumann J, et al. Cryopreserved human amniotic membrane for ocular surface reconstruction. Graefes Arch Clin Exp Ophthalmol [Internet]. 2000. January;238(1):68–75. [DOI] [PubMed] [Google Scholar]
- 32.Niknejad H, Peirovi H, Jorjani M, Ahmadiani A, Ghanavi J, Seifalian AM. Properties of the amniotic membrane for potential use in tissue engineering. Eur Cells Mater. 2008;15:88–99. [DOI] [PubMed] [Google Scholar]
- 33.Hao Y, Ma DH-K, Hwang DG, Kim WS, Zhang F. Identification of antiangiogenic and antiinflammatory proteins in human amniotic membrane. Cornea [Internet]. 2000. May;19(3):348–52. [DOI] [PubMed] [Google Scholar]
- 34.Akita S, Akino K, Hirano A. Basic Fibroblast Growth Factor in Scarless Wound Healing. Adv wound care [Internet]. 2013. March;2(2):44–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Werner S, Grose R. Regulation of wound healing by growth factors and cytokines. Physiol Rev. American Physiological Society; 2003. July;83(3):835–70. [DOI] [PubMed] [Google Scholar]
- 36.Matsumoto K, Hashimoto K, Yoshikawa K, Nakamura T. Marked stimulation of growth and motility of human keratinocytes by hepatocyte growth factor. Exp Cell Res [Internet]. 1991. September;196(1):114–20. [DOI] [PubMed] [Google Scholar]
- 37.Ishida Y, Kondo T, Kimura A, Matsushima K, Mukaida N. Absence of IL-1 receptor antagonist impaired wound healing along with aberrant NF-kappaB activation and a reciprocal suppression of TGF-beta signal pathway. J Immunol [Internet]. 2006. May 1;176(9):5598–606. [DOI] [PubMed] [Google Scholar]
- 38.Barrientos S, Stojadinovic O, Golinko MS, Brem H, Tomic-Canic M. Growth factors and cytokines in wound healing. Wound Repair Regen [Internet]. 2008;16(5):585–601. [DOI] [PubMed] [Google Scholar]
- 39.O’Kane S, Ferguson MW. Transforming growth factor beta s and wound healing. Int J Biochem Cell Biol [Internet]. 1997. January;29(1):63–78. [DOI] [PubMed] [Google Scholar]
- 40.Schreml S, Szeimies R-M, Prantl L, Landthaler M, Babilas P. Wound healing in the 21st century. J Am Acad Dermatol [Internet]. 2010. November;63(5):866–81. [DOI] [PubMed] [Google Scholar]
- 41.Ries C. Cytokine functions of TIMP-1. Cell Mol Life Sci [Internet]. 2014. February;71(4):659–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Martin P. Wound healing--aiming for perfect skin regeneration. Science [Internet]. 1997. April 4;276(5309):75–81. [DOI] [PubMed] [Google Scholar]
- 43.Diegelmann RF, Evans MC. Wound healing: an overview of acute, fibrotic and delayed healing. Front Biosci [Internet]. 2004. January 1;9:283–9. [DOI] [PubMed] [Google Scholar]
- 44.Pastar I, Stojadinovic O, Tomic-Canic M. Role of keratinocytes in healing of chronic wounds. Surg Technol Int [Internet]. 2008. January [cited 2016 May 24];17:105–12. [PubMed] [Google Scholar]
- 45.Li J, Chen J, Kirsner R. Pathophysiology of acute wound healing. Clin Dermatol [Internet]. 2007;25(1):9–18. [DOI] [PubMed] [Google Scholar]
- 46.Shirakata Y, Kimura R, Nanba D, Iwamoto R, Tokumaru S, Morimoto C, et al. Heparin-binding EGF-like growth factor accelerates keratinocyte migration and skin wound healing. J Cell Sci [Internet]. 2005. June 1;118(Pt 11):2363–70. [DOI] [PubMed] [Google Scholar]
- 47.Freedberg IM, Tomic-Canic M, Komine M, Blumenberg M. Keratins and the keratinocyte activation cycle. J Invest Dermatol [Internet]. 2001. May;116(5):633–40. [DOI] [PubMed] [Google Scholar]
- 48.Tonnesen MG, Feng X, Clark RA. Angiogenesis in wound healing. J Investig Dermatol Symp Proc [Internet]. 2000. December [cited 2016 May 24];5(1):40–6. [DOI] [PubMed] [Google Scholar]
- 49.Massee M, Chinn K, Lim JJ, Godwin L, Young CS, Koob TJ. Type I and II Diabetic Adipose-Derived Stem Cells Respond In Vitro to Dehydrated Human Amnion/Chorion Membrane Allograft Treatment by Increasing Proliferation, Migration, and Altering Cytokine Secretion. Adv wound care [Internet]. 2016. February 1 [cited 2016 Jun 2];5(2):43–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Perepelkin NMJ, Hayward K, Mokoena T, Bentley MJ, Ross-Rodriguez LU, Marquez-Curtis L, et al. Cryopreserved amniotic membrane as transplant allograft: viability and post-transplant outcome. Cell Tissue Bank [Internet]. 2016. March [cited 2016 Jun 2];17(1):39–50. [DOI] [PubMed] [Google Scholar]
- 51.Zelen CM, Serena TE, Denoziere G, Fetterolf DE. A prospective randomised comparative parallel study of amniotic membrane wound graft in the management of diabetic foot ulcers. Int Wound J. 2013;10(5):502–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Pachuau L. Recent developments in novel drug delivery systems for wound healing. Expert Opin Drug Deliv [Internet]. 2015;12(12):1895–909. [DOI] [PubMed] [Google Scholar]
- 53.Ashraf A, Lee PHU, Kim K, Zaporojan V, Bonassar L, Valentini R, et al. Effect of sustained-release PDGF and TGF-beta on cyclophosphamide-induced impaired wound healing. Plast Reconstr Surg [Internet]. 2009. October;124(4):1118–24. [DOI] [PubMed] [Google Scholar]
- 54.Magatti M, De Munari S, Vertua E, Nassauto C, Albertini A, Wengler GS, et al. Amniotic mesenchymal tissue cells inhibit dendritic cell differentiation of peripheral blood and amnion resident monocytes. Cell Transplant [Internet]. 2009;18(8):899–914. [DOI] [PubMed] [Google Scholar]
- 55.Magatti M, Caruso M, De Munari S, Vertua E, De D, Manuelpillai U, et al. Human Amniotic Membrane-Derived Mesenchymal and Epithelial Cells Exert Different Effects on Monocyte-Derived Dendritic Cell Differentiation and Function. Cell Transplant [Internet]. 2015;24(9):1733–52. [DOI] [PubMed] [Google Scholar]
- 56.Dinh T, Tecilazich F, Kafanas A, Doupis J, Gnardellis C, Leal E, et al. Mechanisms involved in the development and healing of diabetic foot ulceration. Diabetes [Internet]. 2012. November;61(11):2937–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Solomon A, Wajngarten M, Alviano F, Anteby I, Elchalal U, Pe’er J, et al. Suppression of inflammatory and fibrotic responses in allergic inflammation by the amniotic membrane stromal matrix. Clin Exp Allergy [Internet]. 2005. July;35(7):941–8. [DOI] [PubMed] [Google Scholar]
- 58.Kumar A, Chandra RV, Reddy AA, Reddy BH, Reddy C, Naveen A. Evaluation of clinical, antiinflammatory and antiinfective properties of amniotic membrane used for guided tissue regeneration: A randomized controlled trial. Dent Res J (Isfahan) [Internet]. 2015;12(2):127–35. [PMC free article] [PubMed] [Google Scholar]
- 59.Tan JL, Chan ST, Wallace EM, Lim R. Human amnion epithelial cells mediate lung repair by directly modulating macrophage recruitment and polarization. Cell Transplant [Internet]. 2014. March;23(3):319–28. [DOI] [PubMed] [Google Scholar]
- 60.Mantovani A, Biswas SK, Galdiero MR, Sica A, Locati M. Macrophage plasticity and polarization in tissue repair and remodelling. J Pathol [Internet]. 2013. January;229(2):176–85. [DOI] [PubMed] [Google Scholar]
- 61.Murray PJ, Wynn TA. Protective and pathogenic functions of macrophage subsets. Nat Rev Immunol [Internet]. 2011. October 14;11(11):723–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.MacLeod AS, Mansbridge JN. The Innate Immune System in Acute and Chronic Wounds. Adv wound care [Internet]. 2016. February 1;5(2):65–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Pestka S, Krause CD, Sarkar D, Walter MR, Shi Y, Fisher PB. Interleukin-10 and related cytokines and receptors. Annu Rev Immunol [Internet]. 2004;22(1):929–79. [DOI] [PubMed] [Google Scholar]
- 64.Dinarello CA, Wolff SM. The role of interleukin-1 in disease. N Engl J Med [Internet]. 1993. January 14;328(2):106–13. [DOI] [PubMed] [Google Scholar]
- 65.He H, Li W, Tseng DY, Zhang S, Chen S-Y, Day AJ, et al. Biochemical characterization and function of complexes formed by hyaluronan and the heavy chains of inter-alpha-inhibitor (HC*HA) purified from extracts of human amniotic membrane. J Biol Chem [Internet]. 2009. July 24;284(30):20136–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Rossi D, Pianta S, Magatti M, Sedlmayr P, Parolini O. Characterization of the conditioned medium from amniotic membrane cells: prostaglandins as key effectors of its immunomodulatory activity. PLoS One [Internet]. 2012;7(10):e46956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Li H, Niederkorn JY, Neelam S, Mayhew E, Word RA, McCulley JP, et al. Immunosuppressive factors secreted by human amniotic epithelial cells. Invest Ophthalmol Vis Sci [Internet]. 2005. March;46(3):900–7. [DOI] [PubMed] [Google Scholar]
- 68.Apte RS, Sinha D, Mayhew E, Wistow GJ, Niederkorn JY. Cutting edge: role of macrophage migration inhibitory factor in inhibiting NK cell activity and preserving immune privilege. J Immunol [Internet]. 1998. June 15;160(12):5693–6. [PubMed] [Google Scholar]
- 69.Cook H, Davies KJ, Harding KG, Thomas DW. Defective extracellular matrix reorganization by chronic wound fibroblasts is associated with alterations in TIMP-1, TIMP-2, and MMP-2 activity. J Invest Dermatol [Internet]. 2000. August;115(2):225–33. [DOI] [PubMed] [Google Scholar]
- 70.Kim JS, Kim JC, Na BK, Jeong JM, Song CY. Amniotic membrane patching promotes healing and inhibits proteinase activity on wound healing following acute corneal alkali burn. Exp Eye Res [Internet]. 2000. March;70(3):329–37. [DOI] [PubMed] [Google Scholar]
- 71.Litwiniuk M, Grzela T. Amniotic membrane: new concepts for an old dressing. Wound Repair Regen [Internet]. 2014;22(4):451–6. [DOI] [PubMed] [Google Scholar]
- 72.Bucalo B, Eaglstein WH, Falanga V. Inhibition of cell proliferation by chronic wound fluid. Wound Repair Regen. 1993;1(3):181–6. [DOI] [PubMed] [Google Scholar]
- 73.Widgerow AD. Chronic wounds - is cellular “reception” at fault? Examining integrins and intracellular signalling. Int Wound J [Internet]. 2013. April;10(2):185–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zhao B, Liu JQ, Zheng Z, Zhang J, Wang SY, Han SC, et al. Human amniotic epithelial stem cells promote wound healing by facilitating migration and proliferation of keratinocytes via ERK, JNK and AKT signaling pathways. Cell Tissue Res. 2016;365(1):85–99. [DOI] [PubMed] [Google Scholar]
- 75.Werner S, Krieg T, Smola H. Keratinocyte–Fibroblast Interactions in Wound Healing Epithelial–mesenchymal interactions in tissue repair. J Invest Dermatol. 2007;127:998–1008. [DOI] [PubMed] [Google Scholar]
- 76.Harding KG, Moore K, Phillips TJ. Wound chronicity and fibroblast senescence - implications for treatment. Int Wound J [Internet]. 2005. December;2(4):364–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Campisi J. The role of cellular senescence in skin aging. J Investig dermatology Symp Proc [Internet]. Elsevier Masson SAS; 1998. August;3(1):1–5. [PubMed] [Google Scholar]
- 78.Widgerow AD. Chronic wounds - is cellular “reception” at fault? Examining integrins and intracellular signalling. Int Wound J. 2013. April;10(2):185–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Gruss JS, Jirsch DW. Human amniotic membrane: a versatile wound dressing. Can Med Assoc J [Internet]. 1978. May 20;118(10):1237–46. [PMC free article] [PubMed] [Google Scholar]
- 80.Sawhney CP. Amniotic membrane as a biological dressing in the management of burns. Burns [Internet]. 1989. October;15(5):339–42. [DOI] [PubMed] [Google Scholar]
- 81.Zelen CM, Serena TE, Snyder RJ. A prospective, randomised comparative study of weekly versus biweekly application of dehydrated human amnion/chorion membrane allograft in the management of diabetic foot ulcers. Int Wound J. 2014;11(2):122–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Duan-Arnold Y, Gyurdieva A, Johnson A, Uveges TE, Jacobstein DA, Danilkovitch A. Retention of Endogenous Viable Cells Enhances the Anti-Inflammatory Activity of Cryopreserved Amnion. Adv wound care [Internet]. 2015. September 1;4(9):523–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Johnson E, Marshall J, Michael G. A Comparative Outcomes Analysis Evaluating Clinical Effectiveness in Two Different Human Placental Membrane Products for Wound Management. Wound Repair Regen [Internet]. 2016. December 20. [DOI] [PubMed] [Google Scholar]
- 84.Roy S, Biswas S, Khanna S, Gordillo G, Bergdall V, Green J, et al. Characterization of a preclinical model of chronic ischemic wound. Physiol Genomics [Internet]. 2009. May 13;37(3):211–24. [DOI] [PMC free article] [PubMed] [Google Scholar]