

Abstract
Hippocampal (HC) volume has been hypothesized to increase with an increase in food-hoarding specialization in corvids and parids. Recent studies revealed that (i) the HC/hoarding relationship is significant when a difference in HC volume between Eurasian and North American species is controlled for and (ii) the evolutionary association has been acting on a broader phylogenetic context involving avian families outside the Corvidae and Paridae. However, the phylogenetic extent of the continent effect has not been previously addressed. Using data representing 48 avian species, we performed a phylogenetic analysis to test if continental effects are important in a wider evolutionary spectrum. Our results support the observation that Eurasian species have generally larger HC than North American species if variation in food hoarding, which also varied between continents, was held constant. Surprisingly, the relationship between continental distribution and relative HC volume was significant when we included only non-hoarding families in our analysis, indicating that the extent of the continent effect is much broader than originally described. We investigated the potential role of minimal winter temperatures at the northernmost distribution borders in mediating continent effects. The effect of winter temperatures on HC volume was weak and it did not vary consistently along continents. We suggest that the general continental differences in relative HC size are independent of food hoarding and that its determinants should be sought among other ecological factors and life-history traits.
Keywords: Corvidae, food caching, hippocampus, Paridae, phylogeny
1. INTRODUCTION
Cognitive traits and their underlying neural substrates may be shaped by natural selection. Accordingly, specialized food-hoarding birds have long been hypothesized to have larger hippocampal (HC) volumes than non-hoarding species (review in Healy et al. 2005). However, studies investigating the relationship between HC volume and food-hoarding specialization provide conflicting results. For example, Brodin & Lundborg (2003) found no significant support for the hypothesis in either the Corvidae or the Paridae. On the other hand, Lucas et al. (2004) showed that the HC volume of hoarding species was larger than the HC volume of non-hoarders in both families, but only when a previously undescribed continent difference was statistically removed.
In a separate study, Garamszegi & Eens (2004) performed a phylogenetic analysis of 55 bird species from 13 families. With this larger dataset, they showed that the evolutionary change from non-hoarding to specialized hoarding has been accompanied by an evolutionary increase in HC volume. Thus the HC/hoarding relationship appears to be valid in a broad evolutionary context involving distantly related avian families, even outside the Corvidae and Paridae. However, only Lucas et al. (2004) addressed the potential for continent differences. Given the limited taxonomic scope of the Lucas et al. (2004) study, it is still unclear whether the continent effect is restricted to avian families characterized by hoarding specialization or whether the effect is more broadly distributed across other families.
In this paper we test the generality of the ‘continent effect’, relying on the extended dataset of Garamszegi & Eens (2004). We also examine whether the continental differences in HC volume are independent of food-hoarding specialization. We make an effort to identify the role of an ecological factor that may cause continental variation in HC volume. We assess the importance of temperatures that wintering species experience at their northernmost distributions that may affect food hoarding or HC volume. We hypothesize that in cold wintering conditions, animals may hoard more food, retrieve their caches more efficiently and perform better in spatial memory tasks than animals wintering in milder climates (see also Pravosudov & Clayton 2002). Therefore, species that are efficient at exploitation of food resources through specialized food hoarding (or other behavioural traits requiring a larger HC) may be more likely than non-hoarding species to disperse to colder climates. Hence, winter temperatures may shape continental effects, if food hoarding and wintering conditions vary consistently along the continents. If Eurasian species have to adapt to colder winters than North American species, then Eurasian birds are expected to specialize in food hoarding and evolve larger HC volumes than North American birds.
2. MATERIAL AND METHODS
(a) Dataset
Data on relative HC volume were taken from Garamszegi & Eens (2004) and supplemented with an additional species, Poecile carolinensis, from Lucas et al. (2004). We believe that this dataset contains virtually all published data on HC volume. Given the wide oceanic distribution of Oceanodroma leucorrhoa and Fulmarus glacialis, the presence of Pica pica, Sturnus vulgaris and Troglodytes troglodytes in both continents and the absence of Lonchura striata, Taeniopygia guttata and Vidua paradisea in either continent, these species were excluded from the analyses resulting in a list of 48 species of nine avian families. We used the phylogentically adjusted relative HC volumes that are controlled for telencephalon volume (see Garamszegi & Eens 2004). Alternative allometric corrections (e.g. based on body size) give similar results.
We characterized the extent of food hoarding by using the standard three-point scale from Garamszegi & Eens (2004). Species not known to hoard were considered non-hoarders (score of 1), whereas species that store small amounts of food for short time-intervals were categorized as non-specialized hoarders (score of 2). Species that exhibit typical hoarding behaviour by storing thousands of items in the autumn for later recovery were treated as specialized hoarders (score of 3). Since intermediate states are biologically meaningful, food hoarding was treated as a continuous variable. However, food hoarding can be viewed as a categorical variable. Therefore, by collapsing non-specialized and specialized hoarders, food hoarding was also used as a two-state categorical variable.
To obtain wintering temperatures, we first estimated winter distribution ranges from continent-specific field guides (Heinzel et al. 1997; National Geographic 2000). Then, we calculated latitudinal and longitudinal geographical coordinates at the maximal northernmost distribution by the use of a geographical map to the nearest 1 degree. These estimates are significantly repeatable between sources (e.g. Cramp & Simmons 1977–1994; Poole et al. 1993–2002) and between observers (latitude, R = 0.928; longitude, R = 0.975). We preferred using information from field guides because the resolution of distribution maps are similar in continent-specific field guides, whereas the differences in the quality of maps between Cramp & Simmons (1977–1994) and Poole et al. (1993–2002) may raise biases for our specific purposes here. Similar measures of distributions provided biologically meaningful measures (Gaston & Blackburn 1996). Second, based on the geographical coordinates, average January temperatures were estimated to the nearest degree 1 °C by using electronic sources from the RI/LDEO Climate Data Library Web site (http://iridl.ldeo.columbia.edu). Throughout the study, we assumed that when longitudinal effects are also taken into account, the northernmost winter distribution reflects the coldest temperatures that wintering species experience. Given their migratory habits or their southern distribution, quantitative data on the winter distribution for seven species (Ficedula albicollis, Molothrus badius, Molothrus bonariensis, Molothrus rufoaxillaris, Phylloscopus trochilus, Sylvia borin and Urocissa erythrorhyncha) were unavailable.
(b) Phylogenetic methods
To test for the effect of continental distribution as a categorical variable on HC volume, we adopted the ANOVA-like design developed for phylogenetic associations as suggested by Garland et al. (1993). These approaches allow the assessment of a continuous covariate, which we used to investigate the role of the winter temperatures on the continent effect on HC volumes. These phylogenetic models are implemented in Garland’s Pdap package (http://biology.ucr.edu/people/faculty/Garland/PDAP.html), and presented in full detail in Garland et al. (1993) and in the corresponding User’s Guide. Using the software package, we derived the critical F values for the phylogenetically adjusted distribution and compared them with values calculated from a conventional ANOVA (or ANCOVA) analysis of the raw species data. Both the conventional ( p) and the phylogenetically adjusted (pphyl) significance levels are presented. We applied two approaches to control for the potential confounding effect of food hoarding that may vary between continents. First, we used food hoarding as a continuous variable in a phylogenetic analysis of covariance. Second, we excluded food-hoarding families (Corvidae, Paridae, Picidae and Sittidae) and investigated the continental variation in relative HC volume only in avian families with no evidence of food hoarding. When we estimated the correlative evolution of discrete variables, such as continental distribution and food hoarding in a categorical scale, we used Pagel’s discrete model, which applies a continuous-time Markov model to characterize evolutionary changes along each branch of a phylogenetic tree (Pagel 1999). For the evolutionary modelling, we used the phylogenetic hypothesis from Garamszegi & Eens (2004) after removing the excluded species and inserting P. carolinensis, as suggested by Gill et al. (1989).
3. RESULTS
(a) Continent effects
An ANCOVA model, in which we entered food hoarding as a covariate, revealed a significant relationship between relative HC volume and continental distribution (F1,45 = 17.359, p < 0.001, pphyl = 0.022). Similarly, when we used a categorical approach food hoarding showed consistent variation along continents (food hoarding as a continuous variable: F1,46 = 4.949, p = 0.031, pphyl = 0.128; food hoarding as a categorical variable: Fisher’s exact p = 0.041, Discrete modelling, χ2 = 4.734, pphyl = 0.010; figure 1a). Interestingly, non-hoarders and non-specialized hoarders have a similar HC volume in Europe, while the two hoarder categories are similar in relative HC volume and the non-hoarders are different in North America.
Figure 1.
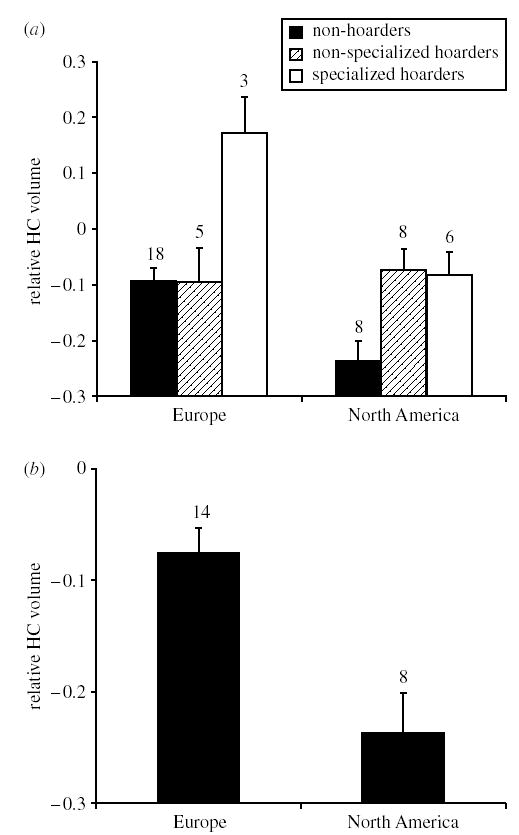
(a) Relative hippocampal (HC) volume (corrected for telecephalon) and food-hoarding specialization in Eurasian and North American birds. (b) Relative HC volume and continental distribution when hoarding families (Corvidae, Paridae, Picidae and Sittidae) have been excluded. Error bars and sample sizes are given.
Our expanded dataset supports the conclusion of Lucas et al. (2004) that Eurasian birds have larger HC volumes than North American birds. However, this trend could be caused by continental differences in HC volumes expressed only in the caching birds. To evaluate the extent of the continent effect, we deleted all families known to hoard and tested for the continent effect in the rest of our dataset. The effect remained significant, even when phylogeny was controlled for (F1,20 = 15.756, p < 0.001, pphyl = 0.039; figure 1b). Thus, the continent effect is broadly distributed across the avifauna of these two continents.
(b) The role of winter temperatures
We found that winter temperatures may be important in explaining variation in HC volume (figure 2), but this effect was non-significant after correcting for phylogenetic relationships (F1,38 = 9.309, p = 0.004, pphyl = 0.359). Similar patterns were found when we controlled for food hoarding by including families not known to hoard (F1,13 = 3.077, p = 0.103, pphyl = 0.155). There are no consistent differences in winter temperatures between continents (F1,39 = 0.565, p = 0.459, pphyl = 0.608).
Figure 2.
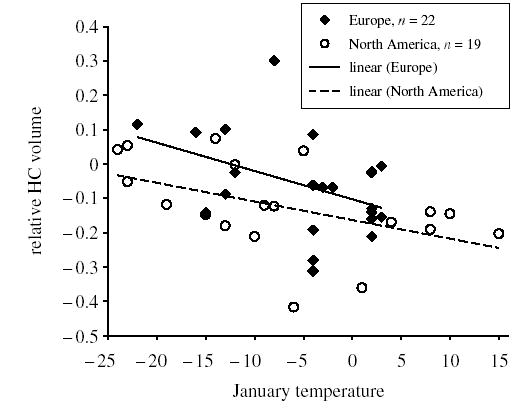
The relationship between January temperatures at the northernmost wintering distribution of species and relative HC volume. Regression lines and sample sizes are given.
4. DISCUSSION
Our phylogenetic analyses indicate that the continent effect on relative HC volume found by Lucas et al. (2004) in corvids and parids is a general phenomenon in birds. It seems that relative HC volume is larger in birds inhabiting the Eurasian subcontinent than in birds from North America. We also found a significant relationship between continental distribution and food hoarding. This result may suggest that differences in food-hoarding specialization between the continents promote differences in relative HC volume. However, when food hoarding is statistically controlled for, the association between continental distribution and relative HC volume is still significant. In addition, the relative volume of the HC is significantly smaller in North America than in Europe when involving typically non-hoarding avian families only (figure 1b). Therefore, continental differences in food-hoarding specialization cannot lead to the phenomenon that Eurasian species have larger HC volumes. This pattern seems to be a general phenomenon among birds and it is not an attribute of food-hoarding families.
Lucas et al. (2004) suggested that systematic methodological differences between different laboratories in how brain measurements were taken may result in differences in HC volume between continents. However, a laboratory effect may only play a minor role because different laboratories are collaborating; analysing data from different laboratories separately gives similar trends for the HC/hoarding relationship (Lucas et al. 2004). In addition, the high between-studies repeatability of relative HC volume also argues against this explanation (Garamszegi & Eens 2004).
Lucas et al. (2004) also proposed that the phylogenetic distribution of birds may be responsible for the observed differences in HC volume between continents. However, the phylogenetic analyses presented here and in Garamszegi & Eens (2004) show that phylogenetic effects are unimportant confounding factors.
As a third possibility, Lucas et al. (2004) put forward the hypothesis that there truly is some continent-wide difference in an ecological factor or a life history trait that mediates differences in the scaling of brains. However, the role of these potential factors remained undetermined. Here we hypothesized that the wintering conditions of species may be different between Europe and North America and that these differences may select for different behavioural adaptations that are ultimately reflected by differences in relative HC volumes. We found a significantly negative relationship between winter temperatures and relative HC volumes at the raw species level, which was non-significant when we controlled for phylogenetic associations. This may indicate that the effect of wintering temperatures on relative HC volume, if it exists at all, is small and, thus, goes undetected in our small sample when controlling for common ancestry. It is also possible that January temperatures at the measured locations do not reflect the coldest temperatures that wintering species experience. However, we conclude that winter temperatures cannot be the primary basis of the continent effects because wintering temperatures did not differ between the continents. We suggest that differences in the degree of food hoarding and relative HC size in Europe and North America should be sought among other ecological factors and life-history traits that are not mirrored by winter temperatures. A comprehensive analysis of the role of biological factors should include a broad range of ecological or life history traits that can be related to HC volume and continental distribution and, thus, potentially shape the continent effect. Ideally, if the mediator variable is found, the continental effects should drop out of significance in the model.
The continent effect on relative HC volume, which is independent of food hoarding and winter temperatures, implies that other behavioural tasks requiring space in this neural substrate also allow species to adapt to environmental factors that vary across continents. Thus, food hoarding seems to be just a part of a complex cognitive adaptation process governed in part by the HC. The evolutionary role played by the HC may be even more important than previously thought.
Acknowledgments
K. Bledsoe, A. Boyko, M. Eens, B. Fanson, C. Jonason, T. Garland, K. Henry, J. Lanier, L. Lohmann, Mark Nolen and R. Van Damme provided useful comments. L.Z.G. was supported by a postdoctoral grant from the FWO-Flanders (Belgium). Part of the work was funded by an NIMH grant to J.R.L., Nicky Clayton and Hubert Schwabl.
References
- Brodin A, Lundborg K. Is hippocampal volume affected by specialization for food hoarding in birds? Proc R Soc B. 2003;270:1555–1563. doi: 10.1098/rspb.2003.2413. (doi:10.1098/rspb. 2003.2413.) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cramp, S. & Simmons, K. E. L. 1977–1994 The birds of the western Palearctic Oxford University Press.
- Garamszegi LZ, Eens M. The evolution of hippocampus volume and brain size in relation to food hoarding in birds. Ecol Lett. 2004;7:1216–1224. [Google Scholar]
- Garland TJ, Dickerman AW, Janis CM, Jones JA. Phylogenetic analysis of covariance by computer simulation. Syst Biol. 1993;42:265–292. [Google Scholar]
- Gaston KJ, Blackburn TM. Global scale macroecology: interactions between population size, geographic range size and body size in the Anseriformes. J Anim Ecol. 1996;65:701–714. [Google Scholar]
- Gill FB, Funk DH, Silverin B. Protein relationship among titmice (Parus) Wilson Bull. 1989;101:182–197. [Google Scholar]
- Healy SD, de Kort SR, Clayton NS. The hippocampus, spatial memory and food hoarding: a puzzle revisited. Trends Ecol Evol. 2005;20:17–22. doi: 10.1016/j.tree.2004.10.006. [DOI] [PubMed] [Google Scholar]
- Heinzel, H., Fitter, R. & Parslow, J. 1997 Birds of Britain and Europe London: Harper Collins Publishers.
- Lucas JR, Brodin A, de Kort SR, Clayton NS. Does hippocampal size correlate with the degree of caching specialization? Proc R Soc B. 2004;271:2423–2429. doi: 10.1098/rspb.2004.2912. (doi:10.1098/rspb.2004.2912.) [DOI] [PMC free article] [PubMed] [Google Scholar]
- National Geographic 2000 Field guide to the birds of North America Washington, DC: National Geographic Society.
- Pagel M. Inferring the historical patterns of biological evolution. Nature. 1999;401:877–884. doi: 10.1038/44766. [DOI] [PubMed] [Google Scholar]
- Poole, A., Stettenheim, P. & Gill, F. 1993–2002 The birds of North America Philadelphia: The American Ornithologists’ Union and The Academy of Natural Sciences of Philadelphia.
- Pravosudov VV, Clayton NS. A test of the adaptive specialization hypothesis: Population differences in caching, memory, and the hippocampus in blackcapped chickadees (Poecile atricapilla) Behav Neurosci. 2002;116:515–522. [PubMed] [Google Scholar]