

Abstract
Objective
Findings from animal studies indicate that the early gut bacteriome is a potential mechanism linking maternal prenatal stress with health trajectories in offspring. However, clinical studies are scarce and the associations of maternal psychological profiles with the early infant faecal bacteriome is unknown. This study aimed to investigate the associations of prenatal stressors and distress with early infant faecal bacterial profiles in a South African birth cohort study.
Methods
Associations between prenatal symptoms of depression, distress, intimate partner violence (IPV) and posttraumatic stress-disorder (PTSD) and faecal bacterial profiles were evaluated in meconium and subsequent stool specimens from 84 mothers and 101 infants at birth, and longitudinally from a subset of 69 and 36 infants at 4–12 and 20–28 weeks of age, respectively in a South African birth cohort study.
Results
Infants born to mothers that were exposed to high levels of IPV had significantly higher proportions of Citrobacter and three unclassified genera, all of which belonging to the family Enterobacteriaceae detected at birth. Proportions of these Enterobacteriaceae remained significantly increased over time (birth to 20-28 weeks of life) in infants born to mothers with high levels of IPV exposure compared to infants from mothers with no/low IPV exposure. Infants born to mothers exposed to IPV also had higher proportions of the genus Weissella at 4-12 weeks compared to infants from mothers with no/low IPV exposure. Faecal specimens from mothers exposed to IPV had higher proportions of the family Lactobacillaceae and lower proportions of Peptostreptococcaceae at birth. Maternal psychological distress was associated with decreased proportions of the family Veillonellaceae in infants at 20-28 weeks and a slower decline in Gammaproteobacteria over time. No changes in beta diversity were apparent for maternal or infant faecal bacterial profiles in relation to any of the prenatal measures for psychological adversities.
Conclusion
IPV during pregnancy is associated with altered bacterial profiles in infant and maternal faecal bacteria. These findings may provide insights in the involvement of the gut bacteria linking maternal psychological adversity and the maturing infant brain.
Keywords: Microbiota, Meconium, Domestic violence, Intimate partner violence, Maternal antenatal stress
Introduction
The prevalence of maternal prenatal stressors and distress are particularly high in rural South Africa. Previous studies reported that 30–40% of pregnant mothers have depression (Vythilingum et al., 2012, Rochat et al., 2006, Hartley et al., 2011), 11–19% have posttraumatic stress disorder (PTSD) (van Heyningen et al., 2017, Koen et al., 2016) and 21–32% experienced intimate partner violence (IPV) within the past year (Koen et al., 2016, Bernstein et al., 2016). Psychological stressors and distress during pregnancy may have a range of negative consequences on the development and health of the foetus and infant, including premature delivery, impaired cognitive development, poor infant growth and dysregulated behaviour of infants (Stein et al., 2014). The biological mechanisms responsible for the intergenerational transmission of the effects of maternal psychological stress during pregnancy are, however, still unclear. Studies in mice (Gur et al., 2017) and monkeys (Bailey et al., 2004) have found that exposure to psychological stressors during pregnancy alters the profile of the maternal faecal microbiota, which is then transferred to the offspring. There are, however, few human studies. A Dutch human study (Zijlmans et al., 2015) found that a higher self-reported stress during pregnancy was associated with temporal differences in infants’ faecal bacterial profiles, starting 7 days after delivery. Furthermore, infant faecal bacterial profiles from mothers reporting higher stress levels were characterized by higher relative abundances of Proteobacteria (including the Enterobacteriaceae Escherichia, Serratia and Enterobacter) and a lower relative abundances of lactic acid bacteria and Bifidobacteria (Zijlmans et al., 2015). A recent study examined the associations of maternal prenatal anxiety, depression and stress during pregnancy with changes to the microbiome in meconium of 75 new-borns (Hu et al., 2019). Higher measures of pregnancy-related anxiety were associated with lower abundances of an unidentified genus in the family Enterobacteriaceae (Hu et al., 2019). However, the association of other types of psychological stressors and distress during pregnancy with the temporal changes in the infant faecal microbiome has not yet been studied.
The acquired faecal microbiota may contribute to child neurodevelopment and health trajectories (Jašarević et al., 2015, O’Mahony et al., 2017, Sordillo et al., 2019). A delayed or altered establishment of a mature intestinal microbiota in childhood (termed microbiota immaturity) has been associated with various diseases, including diarrhoea, malnutrition, atopic conditions, inflammatory bowel disease, obesity, diabetes, neurological conditions and impaired neurodevelopment (Arrieta et al., 2014, Carlson et al., 2018). Studies in mice demonstrated that the gut microbiome may function as mechanistic link between maternal prenatal stress and reduction in social behavioural and neurobiological changes, i.e., neuroinflammation, in the offspring (Gur et al., 2017, Gur et al., 2019). The early establishment of the gut bacterial environment may have long-lasting effects on health and brain functioning. In this respect, a study in germ-free mice showed that colonization of the gut directly after birth was essential for the development of the hypothalamic-pituitary-adrenocortical (HPA) axis (Sudo et al., 2004). A study in humans also showed that changes in gut bacteria at one year of age were associated with with lower cognition, including visual reception and expressive language skills in infants at two years of age (Carlson et al., 2018). However, evidence for the association between maternal prenatal psychological adversities with changes to the infant gut microbiome in utero, i.e., meconium, is still limited.
Aims of the study
The primary aim of this study was to determine the association between different profiles of maternal prenatal stressors and distress (including IPV, psychological distress, depression, and PTSD) experienced by South African mothers and the development of infants’ faecal microbiome early in life. The secondary aim included the investigation of the relationship of these prenatal psychological adversities with maternal faecal bacterial profiles at delivery.
Materials and methods
Study participants
Mother-infant dyads were evaluated in a sub-sample from the Drakenstein Child Health Study (DCHS). The DCHS is multidisciplinary birth cohort study investigating determinants of child health over time. Further details of participant recruitment, data collection and the setting of the DCHS has been provided previously (Zar et al., 2015, Stein et al., 2015). Pregnant women were recruited from two low socioeconomic communities, TC Newman and Mbekweni in a peri-urban area outside Cape Town, South Africa. Enrolment of pregnant women took place at 20-28 weeks of gestation at routine antenatal visits to health care facilities. Mothers provided informed, written consent for enrolment of their infants at the time of delivery and follow up until five years of age. Inclusion criteria for this sub-study were longitudinal collection of faecal specimens and availability of maternal prenatal psychological measurements. Exclusion criteria for this sub-study were maternal age < 18 years, residence outside of the Drakenstein sub-district, and intention to move out of the region within 2 years of giving birth.
This study and parent study [the Drakenstein Child Health Study (DCHS)] both received ethical approval from the Faculty of Health Sciences, Human Research Ethics Committee (HREC) of the University of Cape Town (401/2009 and 742/2013, respectively) (Zar et al., 2015).
Measurements of maternal prenatal psychological stress and distress
Locally validated and reliable psychological measures of stress and distress were performed in pregnant mothers when enrolled at the antenatal clinic as previously described (Stein et al., 2015, Zar et al., 2015, Koen et al., 2016). The measures for maternal prenatal stressors and distress in this sub-study were evaluated at a gestational age of 27.4 ± 4.2 weeks by field workers in the absence of the partner/husband. On-site female fieldworkers administered a battery of self-report measures described below. Female fieldworkers were selected based on criteria previously found to affect women’s willingness to divulge intimate information, including measures of IPV exposure (Jansen et al., 2004). All female fieldworkers had at least a Grade 12 certificate and had prior experience in psychiatric/psychological research. They were fluent both in English and Afrikaans or isiXhosa and were therefore able to administer questionnaires in the participants’ preferred language. Further, fieldworkers received extensive in-service training on all aspects of Good Clinical Practice (GCP) ((WHO), 2002).
The self-reporting questionnaire 20-item (SRQ-20) (Harding et al., 1980), a WHO-endorsed measure of symptoms of psychological distress, with good face validity and reliability, has been used widely in international studies and in South African settings (Rumble et al., 1996, Mari and Williams, 1986). Symptoms of depression were measured by the Beck Depression Inventory (BDI-II), a widely-used and reliable measure of depressive symptoms (Beck et al., 1988, Beck et al., 1996). The modified PTSD Symptom Scale (MPSS) is a 17-item self-report scale with excellent internal consistency, high test-retest reliability and concurrent validity to diagnose PTSD consistent with DSM-IV criteria as defined by DSM-IV (Falsetti et al., 1993, Foa and Tolin, 2000). The validity of the MPSS was established in a South African student sample with a Cronbach’s alpha of 0.92, in relation to a diagnosis by psychiatrists based on clinical interviews for PTSD, for which kappa was found to be 0.68 (McGowan and Kagee, 2013).The IPV Questionnaire used in this study was adapted from the WHO multi-country study (Jewkes, 2002) and the Women’s Health Study in Zimbabwe (Shamu et al., 2011). The IPV questionnaire assess lifetime and recent (past-year) exposure to emotional, physical and sexual IPV. Each category of violence was assessed across multiple items measuring the frequency of a number of specified violent acts. Each item is scored using a frequency scale from 1 (“never”) to 4 (“many times”). Bivariable analyses were performed on scoring guidelines based on prior work in South Africa (Dunkle et al., 2004). Psychological assessments of IPV were previously validated and optimized in local studies of maternal mental health for their use in the South African setting (Stein et al., 2009).
Faecal bacteria analyses
Details of the study methods related to the analyses of faecal bacteria have previously been reported (Claassen-Weitz et al., 2018). In short, faecal specimens were collected using sterile spatulas and faecal screw-cap containers. Study staff collected faecal specimens from mothers and infants prior to hospital discharge or during visits to clinics. All specimens collected at hospital or visits to clinics were immediately stored at −20 °C. Mothers were instructed to collect and immediately store infant faecal specimens at −20 °C in the event where faecal specimens were not passed at hospital or during scheduled visits to clinics. All home collections were transported to clinics using ice-boxes and were delivered to study staff within 24 hours of collection. Transport of faecal specimens between the study site and laboratory was performed under controlled conditions using ice boxes. Upon arrival at the laboratory, faecal specimens were stored at −80 °C until further processing. Faecal specimens included in this study were selected based on the availability of faecal specimens from mother-infant dyads at birth and at follow-up visits. Specimens were selected to include the maximum number of longitudinal sample sets at the time of study.
Nucleic acid was extracted from each of the faecal specimens using approximately 50mg of starting material as described previously (Claassen-Weitz et al., 2018, Claassen et al., 2013). A detailed description of the amplicon library preparation, sequencing and bio-informatics steps was published previously, including access to the raw sequence files supporting the findings of this manuscript (Claassen-Weitz et al., 2018). Briefly, amplicon library preparation was performed by amplifying the V4 hypervariable region of the 16S ribosomal ribonucleic acid (rRNA) gene (Claassen-Weitz et al., 2018). Sequencing was carried out using the Illumina® MiSeq™ platform and the MiSeq Reagent Kit v3, 600 cycles (Illumina, CA, USA) (Claassen-Weitz et al., 2018). Quality filtering steps of raw sequences, removal of potential contaminants, de-replication of sequences occurring more than twice, clustering of sequences into operational taxonomic units (OTUs), removal of chimeras and taxonomic assignment was performed using an in-house bio-informatics pipeline incorporating various software tools (Claassen-Weitz et al., 2018).
Covariates
Data on maternal demographics (residential area, education) and health (HIV status, smoking) were obtained at enrolment. Maternal body mass index (BMI) was determined at 6–10 weeks postpartum. Data for mode of delivery, gestational age, birth weight and length, infant sex, antibiotic use and household members were obtained at the time of delivery at Paarl Hospital, where all births took place. Information on feeding practices was obtained at infant follow-up visits at 4-12, and 20-28 weeks of age (Zar et al., 2015). The covariates included in this study were chosen based on their potential effect on faecal bacterial profiles as shown previously (Claassen-Weitz et al., 2018). Variables that were associated with prenatal psychological measures; or bacterial taxa at phylum, class, order, family or genus-level at each of the time points under study; or diversity indices at each of the time points under study were included as covariates and referenced in the results section. Measures for other covariates, for example, maternal diet at birth, that may affect the associations investigated in this study, were not assessed in this study.
Statistical analyses
R software version 3.1.1 (R_Development_Core_Team, 2014) together with RStudio software version 0.98.50751 was used for all statistical analyses as well as graphical representations of the data. Count data (Anders and Huber, 2010) was transformed to compositional data by calculating the relative abundance of each OTU per specimen (McMurdie and Holmes, 2014, Fernandes et al., 2014). Alpha-diversities were determined using the Shannon diversity (H′) index (Shannon, 1948, Morgan and Huttenhower, 2012) using the vegan R package (Oksanen et al., 2013). Beta-diversities were computed based on the “w” metric as decribed in Koleff et al. (Koleff et al., 2003) with the vegan R package and represented in a SMACOF multidimensional scaling map (I and PJF, 2005). Maternal prenatal psychological measures in this study included symptoms of depression, psychological distress, measures for IPV and PTSD. Measures for psychological distress (SRQ-20) and symptoms of depression (BDI-II) were used as continuous variables. Dichotomized measures for IPV were used and PTSD was categorized according to “trauma-exposed PTSD”, “suspected PTSD” and “no exposure to PTSD”. A multivariate approach was followed, testing simultaneously which covariates significantly influenced the microbiome composition by performing PERMANOVA (Anderson, 2001) on the Bray-Curtis dissimilarity matrix (calculated using the [vegdist] function from the R package vegan) (Morgan and Huttenhower, 2012, Faith et al., 1987, Bray and Curtis, 1957, Clarke and Warwick, 2001). For those covariates identified as significant, generalized linear models (GLMs) with a log link function and a Quasi-Poisson family allowing for overdispersion were used to evaluate the association of prenatal psychological measures with each taxon and alpha-diversity at each of the time points under study (Claassen-Weitz et al., 2018). Similarly, generalized linear mixed models (GLMMs) were used to investigate covariate effects across time points using a subset of 36 infants with complete longitudinal data sets. Hypothesis testing was performed at a 5% significance level and since the GLM and GLMM models were fitted for one taxon at a time, multiple testing necessitated controlling the false discovery rate with the Benjamini-Hochberg correction (Benjamini and Hochberg, 1995).
Results
Study characteristics
Faecal specimens of 90 mothers, 107 infants at birth, 72 infants at 4–12 weeks and 36 infants at 20–28 weeks were collected and successfully sequenced (Claassen-Weitz et al., 2018). Maternal prenatal psychological measures were not collected from six of the mothers. These mothers and their infants (six infants at birth and three infants at 4–12 weeks) were excluded from this study. Associations between prenatal distress measures and infant faecal bacterial profiles were therefore investigated from 101 infant meconium specimens, 69 faecal specimens collected at 4-12 weeks and 36 faecal specimens collected at 20-28 weeks. Associations between prenatal distress measures and maternal faecal bacterial profiles were analysed from 84 maternal faecal specimens collected at the time of delivery. A median of 5465 [interquartile range (IQR): 3159-9877)] post-filtered reads per specimen were obtained following the removal of potential contaminant reads. Infant meconium specimens produced the highest number of post-filtered reads (median: 10002; IQR: 5065–14830), followed by infant faecal specimens collected at 4–12 weeks (median: 6407; IQR: 4069–9042) and 20–28 weeks (median: 5636; IQR: 4228–7084) (Claassen-Weitz et al., 2018). Maternal faecal specimens sampled at birth produced the lowest number of post-filtered reads (median: 3155; IQR: 2104–4355) (Claassen-Weitz et al., 2018). All specimen types produced sufficient sequencing depth for calculating Shannon diversity indices in our study (Supplementary Fig. 1).
Demographic and clinical data of the mothers and infants are reported in Table 1. The median (IQR) measures for psychological distress (SRQ-20) and for symptoms of depression (BDI) from mothers with available data (n = 101) were 4.0 (1-7) and 13 (7-12), respectively. Trauma-associated PTSD was diagnosed among 3% (3/101) of mothers. A total of 45% (45/101) of the mothers were exposed to any form of IPV, of which 32% (34/101) experienced emotional abuse, 29% (31/101) physical abuse and 13% (14/101) sexual abuse. Furthermore, 30% (32/101) of the mothers were recently exposed to IPV (during the past 12 months), and 45 % (45/101) had lifetime IPV exposure.
Table 1:
Demographic and clinical characteristics of mothers and infants included in the study
| Characteristics | Mothers at delivery (n=84) | Infants at birth (n=101) | Infants at 4–12 weeks (n=69) | Infants at 20–28 weeks (n=36) |
|---|---|---|---|---|
| Age at which specimens were collected, median (IQR) | 25.3 (21.9 – 31.24) years | 12.4 (4.2 – 24.6) hours | 7.8 (6.9 – 9.3) weeks | 24.1 (23.9 – 26.4) weeks |
| Residential area: | ||||
| - Mbekweni, n (%) | 37 (44.0 %) | 54 (53.5) % | 39 (56.5 %) | 20 (55.6 %) |
| - TC Newman n (%) | 47 (56.0 %) | 47 (46.5) % | 30 (43.5 %) | 16 (44.4 %) |
| Maternal education: | ||||
| - Primary level, n (%) | 8 (9.5 %) | 11 (10.9 %) | 8 (11.6 %) | 7 (19.4 %) |
| - Secondary level, n (%) | 70 (83.4 %) | 82 (81.2 %) | 57 (82.6 %) | 26 (72.2 %) |
| - Tertiary level, n (%) | 6 (7.1 %) | 8 (7.9 %) | 4 (5.8 %) | 3 (8.3 %) |
| Maternal prenatal distress | ||||
| -SRQ-20, median (IQR) | 4.0 (2-6) | 4.0 (1-7) | 5 (1-8) | 4.5 (1.3-8) |
| -BDI, median (IQR) | 10 (6-18) | 13 (7-22) | 13 (7-23) | 13 (7-22.5) |
| -PTSD | ||||
| -Trauma-exposed, n (%) | 3 (3.6 %) | 3 (3.0 %) | 2 (2.9 %) | 0 (0%) |
| -Suspected PTSD, n (%) | 0 (0 %) | 1 (1.0 %) | 1 (1.4 %) | 1 (2.8 %) |
| -No exposure, n (%) | 81 (96.4 %) | 97 (96.0 %) | 66 (95.7 %) | 35 (97. 2 %) |
| -IPV | ||||
| -IPV emotional, n (%) | 29 (34.5 %) | 34 (33.7 %) | 24 (34.8 %) | 13 (36.1 %) |
| -IPV physical, n (%) | 26 (31.0 %) | 31 (30.7 %) | 23 (33.3 %) | 10 (27.8%) |
| -IPV sexual, n (%) | 10 (11.9 %) | 14 (13.9 %) | 11 (15.9 %) | 7 (19.4 %) |
| -IPV any, n (%) | 36 (42.9 %) | 45 (44.6 %) | 32 (46.4%) | 18 (50. 0 %) |
| -IPV any recent, n (%) | 27 (32.1 %) | 32 (31.7 %) | 25 (36.2 %) | 15 (41.7 %) |
| -IPV any lifetime, n (%) | 36 (42.9 %) | 45 (44.6 %) | 32 (46.4 %) | 18 (50.0 %) |
| Maternal HIV status: | ||||
| - HIV infected, n (%) | 18 (21.4 %) | 26 (25.7 %) | 19 (27.5 %) | 10 (27.8 %) |
| Maternal BMI at 6-10 weeks postnatal, median (IQR) | 25.4 (23.2 - 29.2)* | 25.1 (23.0 - 28.8)* | 25.8 (22.9 - 29.7)* | 25.1 (22.4 - 30.4)* |
| Maternal smoking status:# | ||||
| - Active smoker (cotinine levels ≥ 500), n (%) | 28 (33.3 %) | 28 (27.7 %) | 21 (30.4 %) | 13 (36.1 %) |
| - Passive smoker (cotinine levels > 10 < 500), n (%) | 36 (42.9 %) | 36 (35.6 %) | 19 (27.5 %) | 9 (25.0 %) |
| - Non-smoker (cotinine levels ≤ 10), n (%) | 20 (23.8 %) | 20 (19.8 %) | 12 (17.4 %) | 6 (16.7 %) |
| Mode of delivery: | ||||
| - Vaginal delivery, n (%) | 69 (82.1 %) | 81 (80.2 %) | 57 (82.6 %) | 29 (80.6 %) |
| - Elective caesarean-section delivery, n (%) | 3 (3.6 %) | 5 (5.0 %) | 4 (5.8 %) | 3 (8.3 %) |
| - Emergency caesarean-section delivery, n (%) | 12 (14.3 %) | 15 (14.9 %) | 8 (11.6 %) | 4 (11.1 %) |
| Gestational age (weeks), median (IQR) | 39 (37.3 - 40.0) | 39.0 (38.0 - 40.0) | 39.0 (38.0 - 40.0) | 39.0 (38.3 - 40.0) |
| Birth weight (kg), median (IQR) | 3.0 (2.7 - 3.3) | 3.0 (2.7 - 3.4) | 3.1 (2.7 - 3.5) | 3.0 (2.6 - 3.4) |
| Birth length (cm), median (IQR) | 50.5 (48.0- 52.8) | 50 (48.0 - 53.0) | 51 (48.5 - 53.0) | 50.5 (49 - 53) |
| Sex: | ||||
| - Male, n (%) | 36 (42.9 %) | 43 (42.6 %) | 30 (43.5 %) | 18 (50 %) |
| Milk feeding: | ||||
| - Exclusive breastfeeding, n (%) | 71 (84.5 %)§‡ | 81 (80.2 %)§‡ | 32 (46.4 %)§φ | 6 (16.7 %)§⤉ |
| - Exclusive formula feeding, n (%) | 9 (10.7 %)§‡ | 16 (15.8 %)§‡ | 11 (15.9 %)§φ | 7 (19.4 %)§⤉ |
| - Mixed feeding⫯, n (%) | - | - | 25 (36.2 %)§φ | 23 (63.9 %)§⤉ |
| Antibiotic use, n (%) | 9 (10.7 %) | 15 (14.9 %) | 9 (13.0 %) | 3 (8.3 %) |
| Household members, median (IQR) | 5 (3 - 7) | 4 (3 - 6) | 4 (3 - 6) | 4 (3 - 6) |
Number of observations recorded: Mothers at delivery: 84; Infants at birth: 84; Infants at 4-12 weeks: 52; Infants at 20-28 weeks: 28.
Number of observations recorded: Mothers at delivery: 52; Infants at birth: 65; Infants at 4-12 weeks: 51; Infants at 20-28 weeks: 27.
Number of observations recorded: Mothers at birth: 86; Infants at birth: 103; Infants at 4-12 weeks: 71; Infants at 20-28 weeks: 36.
Feeding recorded prior to discharge from hospital.
Feeding recorded at 4-12 weeks.
Feeding recorded at 20-28 weeks.
breast milk, formula milk and/or solids. IQR: Interquartile range. SRQ-20: psychological distress. BDI: Beck depression inventory. PTSD: Post-traumatic stress disorder. IPV: Intimate partner violence. BMI: Body mass index.
Association of maternal prenatal psychological measures with infant faecal bacterial profiles
Infants born to mothers with exposure to lifetime IPV had higher proportions of unclassified genera within the family Enterobacteriaceae [OTU 101 (p < 0.01); OTU 615 (p = 0.02); OTU 616 (p = 0.01)] measured from meconium (Fig. 1 A–C) (covariates significantly associated with prenatal psychological measures or bacterial taxa that were included in the statistical model: maternal BMI and breastfeeding).
Figure 1.
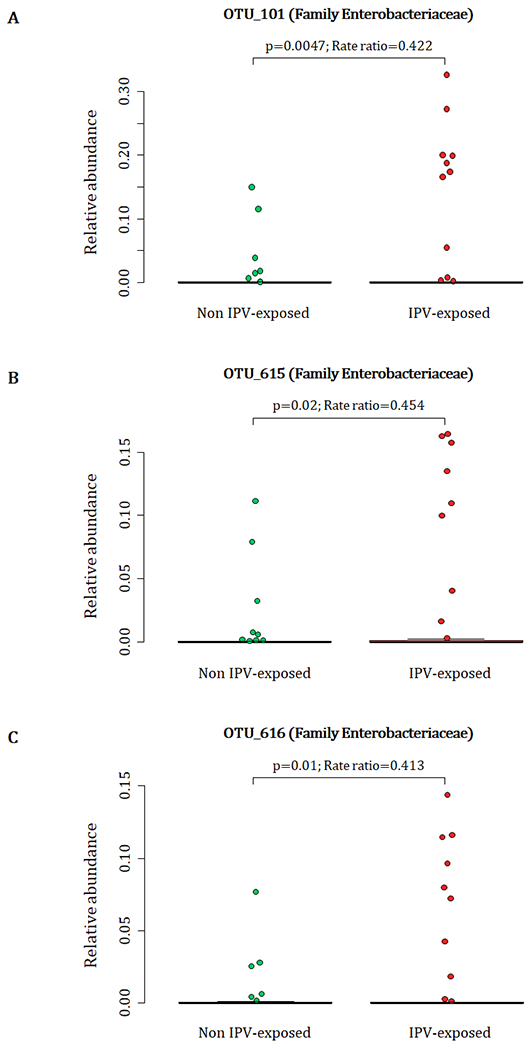
Relationship between maternal lifetime exposure to IPV and the infant meconium bacterial profiles. The number of participants represented in this figure were infants from which we detected the respective OTUs at proportions > 0. A) Differences in proportions of OTU 101 detected from 88 infant meconium specimens; B) differences in proportions of OTU_615 detected from 73 infant meconium specimens; and C) differences in proportions of OTU_616 detected from 71 infant meconium specimens at proportions > 0. IPV=intimate partner violence.
A total of 88 participants at birth had OTU 101, 73 infants had OTU 615 and 71 infants had OTU 616 detected at proportions > 0 from their meconium (Fig. 1 A–C). At 4-12 weeks of age, infants born to mothers with exposure to lifetime IPV had higher proportions of the genus Weissella (Phylum Firmicutes; Class Bacilli; Order Lactobacillales; Family Leuconostocaceae) (p < 0.001) when compared to infants born to mothers who were not exposed to lifetime IPV (Fig. 2) (covariates significantly associated with prenatal psychological measures, bacterial taxa or diversity that were included in the statistical model: area and maternal HIV status). A total of 44 infants at 4-12 weeks of age had Weissella detected at proportions > 0 from their faecal specimens (Fig. 2). This association was not found to be significant at birth or at 20-28 weeks (Supplementary figure 2).
Figure 2.
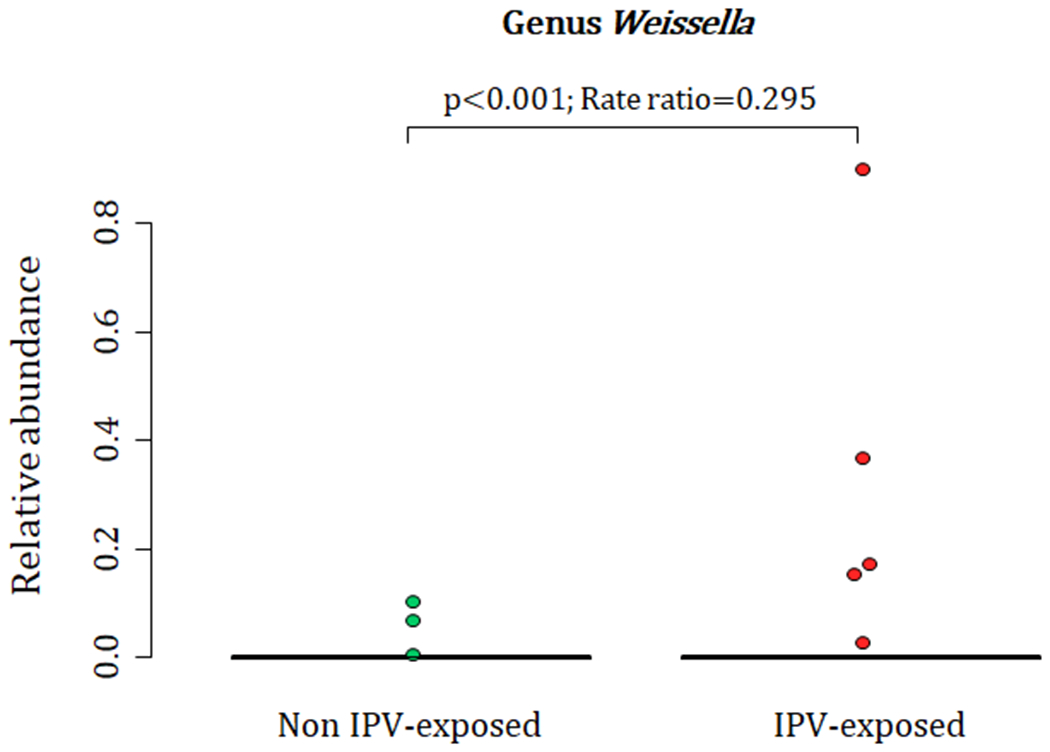
Relationship between maternal lifetime exposure to IPV and the infant faecal bacteria at 4-12 weeks. The figure presents differences in proportions of genus Weissella that was detected from 44 infant faecal specimens at proportions > 0. IPV=intimate partner violence.
At 20-28 weeks, increased measures of maternal psychological distress (SRQ-20) were significantly associated with lower proportions of the family Veillonellaceae (Phylum Firmicutes, Class Negativicutes (p = 0.003), Order Selenomondales (p = 0.003)) (p = 0.01) (covariates significantly associated with psychological measures, bacterial taxa or diversity which were included in the statistical model: area, maternal HIV status, maternal education and gender of infant) (Fig. 3 A–C). This association was only significant at 20-28 weeks (Supplementary figure 3).
Figure 3.
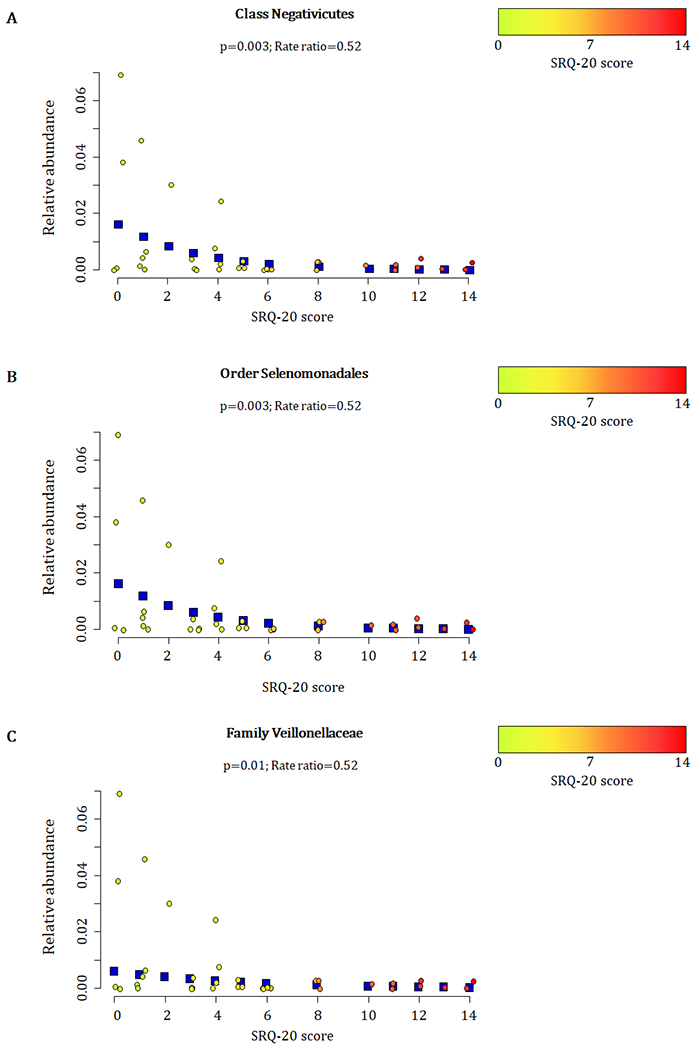
Relationship between maternal prenatal psychological distress (SRQ-20) and infant faecal bacterial profiles at 20-28 weeks. Proportions of the A) class Negativicutes, B) order Selenomonadales, and C) family Veillonellaceae observed from faecal specimens collected at 20-28 weeks were inversely correlated with maternal psychological distress (SRQ-20) (p ≤ 0.01). Individual maternal SRQ-20 scores for the 36 infants investigated at 20-28 weeks are represented by circles using a gradient of colours ranging from green (low SRQ-20 scores) to red (high SRQ-20 scores). Blue squares represent the non-parametric regression smoother used to show the trend in the observations.
When measuring the effect of maternal prenatal stressors and distress on n=36 infant faecal bacterial profiles over time, we found that infants from mothers with exposure to lifetime IPV had increased proportions of taxa within the family Enterobacteriaceae [genus Citrobacter (p = 0.002) (Fig. 4 A) and unclassified genera OTU 101 (p = 0.006) (Fig. 4 B), OTU 615 (p=0.003) (Fig. 4 C) and OTU 616 (p = 0.003) (Fig. 4 D)] at birth through to 20-28 weeks when compared to infants from mothers with no/low IPV exposure (covariates significantly associated with psychological measures or bacterial taxa that were included in the statistical model: gender, area and maternal HIV status). No interaction for age and bacterial proportions with reference to the four genera within the family Enterobacteriaceae from IPV exposed nor unexposed infants was found, and longitudinal analyses were therefore performed for results presented in Figs. 4 A–D. A significant interaction effect between age and IPV exposure was found for the family Streptococcaceae (p = 0.046) and analysis was therefore stratified according to the age groups under investigation (Fig. 4 E). Higher proportions of the Streptococcaceae family was found at birth in infants born to mothers with IPV versus infants from IPV unexposed mothers when compared to these infants at 4-12 weeks (p=0.038) and 20-28 weeks (p=0.018) of age (Fig. 4 E). A significant interaction effect (p = 0.003) was found between age and SRQ for the class Gammaproteobacteria as shown in Fig. 4 F. Overall, lower proportions of the class Gammaproteobacteria was observed at age groups 4-12 weeks and 20-28 weeks compared to the proportions detected at birth (Fig. 4 F). This decrease was more pronounced in infants born to mothers with low prenatal psychological distress at 4-12 weeks (p = 0.007), and a similar non-significant trend was observed at 20-28 weeks (p= 0.089) of age, compared to these infants at birth. (covariates significantly associated with psychological measures or bacterial taxa that were included in the statistical model: gender, area and maternal HIV status) (Fig. 4 F).
Figure 4.
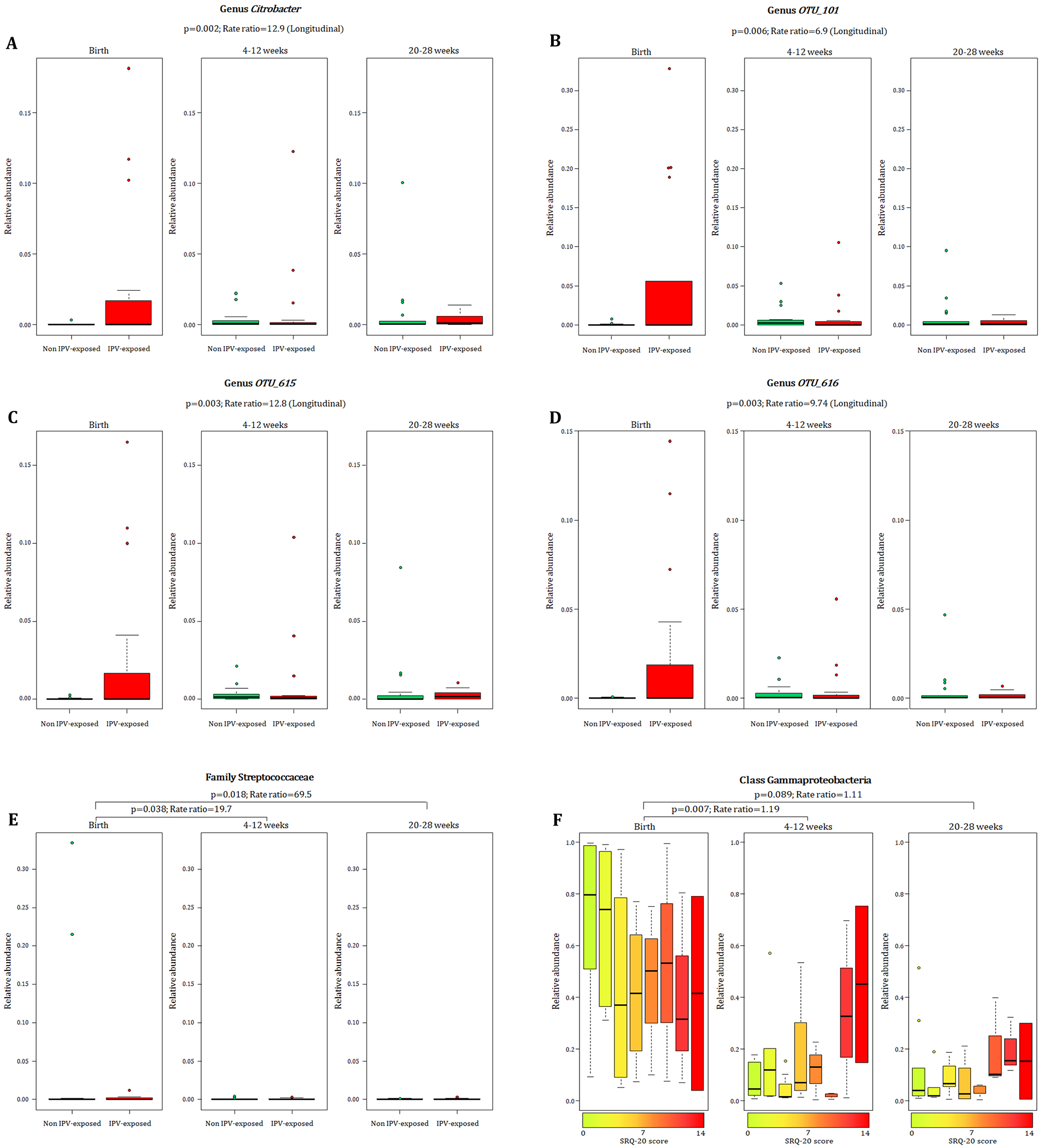
Relationship between maternal exposure to lifetime IPV, psychological distress and temporal changes with infant faecal bacterial profiles. Longitudinal changes to the proportions of A) Genus Citrobacter and unclassified genera within the family Enterobacteriaceae B-D) [OTU_101 (p = 0.006); OTU_615 (p = 0.003); OTU_616 (p = 0.003)] in the infants born to mothers exposed to lifetime IPV compared to infants from mothers exposed to no/low lifetime IPV. E) Differential decrease in proportions of the family Streptococcaceae from birth to ages 4-12 weeks (p = 0.038) and from birth to 20-28 weeks (p = 0.018) with a more pronounced decline in Streptococcaceae for infants from mothers with no/low lifetime IPV exposure compared to infants from mothers exposed to lifetime IPV. F) Association of continuous measures of maternal prenatal psychological distress (SRQ-20) with faecal Gammaproteobacteria in infants at birth compared to the infants at ages 4-12 weeks (p = 0.007) and 20-28 weeks (p = 0.089). Individual maternal SRQ-20 scores for the 36 infants with faecal specimens collected at all three timepoints are represented by a gradient of colours ranging from green (low SRQ-20 scores) to red (high SRQ-20 scores). IPV = intimate partner violence.
No significant associations with infant faecal bacterial profiles were found when assessing symptoms of depression or PTSD. Supplementary figures 4–8 show different measures for maternal prenatal stressors and distress with the infant microbiota at genus level measured over time. No significant associations were found between any maternal prenatal psychological measures and infant faecal bacterial diversity indices (data not shown).
Prenatal psychological measures and maternal faecal bacterial profiles at delivery
Maternal exposure to recent IPV (past year) was associated with higher proportions of the family Lactobacillaceae (p = 0.03) (phylum Firmicutes, class Bacilli, order Lactobacillales) and lower proportions of the family Peptostreptococcaceae (p = 0.04) (phylum Firmicutes, class Clostridia, order Clostridiales) in maternal faecal specimens at delivery (covariates significantly associated with psychological measures or bacterial taxa that were included in the statistical model: mode of delivery) (Fig 5. A and B). None of the prenatal psychological measures were associated with maternal faecal bacterial diversity indices (data not shown).
Figure 5.
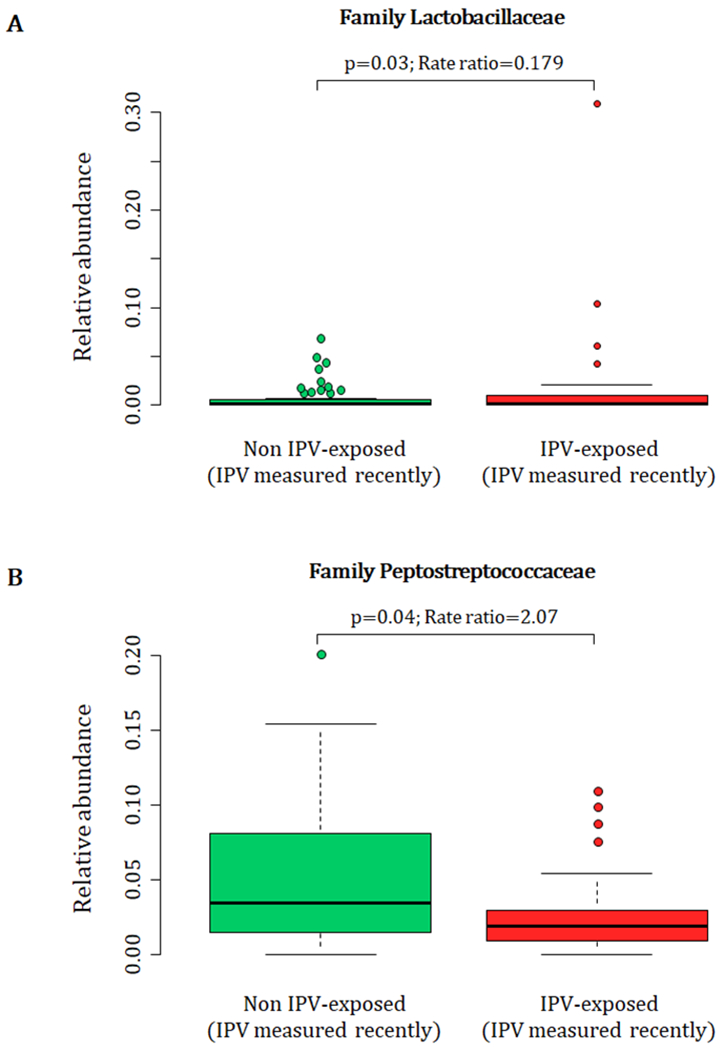
Relationship between maternal exposure to recent IPV (past year) during pregnancy and maternal faecal bacterial profiles at delivery. IPV = intimate partner violence.
Beta diversity and measures for maternal psychological adversities
No clustering patterns were apparent for maternal or infant faecal bacterial profiles in relation to any of the prenatal measures for psychological adversities based on the “w’ metric for beta diversities (results not shown).
Discussion
This study characterized the association of maternal prenatal stressors and distress with early life infant faecal and maternal bacterial profiles. Our results demonstrate that maternal exposure to lifetime IPV and psychological distress were significantly associated with early infant faecal bacterial profiles.
Until recently, the foetus has been considered to exist in a sterile environment with initial microbial colonization of the newborn taking place during birth (D’Argenio, 2018). Recent studies have reported the detection of microbes from amniotic fluid, placenta and meconium, suggesting that microbial colonization may begin in utero (Willyard, 2018, Stinson et al., 2019). In addition, a number of studies have reported bacterial communities from meconium specimens (Seferovic et al., 2018). Dysbiosis of faecal bacterial profiles early in life has been associated with a number of neonatal diseases, including asthma, allergic diseases, and obesity (D’Argenio, 2018), which further emphasises the importance of in utero colonization on the health of newborns.
With cross-sectional analysis we detected higher abundances of unclassified taxa (OTU 101, OTU 615 and OTU 616) within the family Enterobacteriaceae from meconium specimens of infants born to mothers exposed to IPV. Analysis of longitudinal data showed that these findings persisted over the first six months after birth in this study, as higher proportions of these unclassified Enterobacteriaceae taxa, as well as the genus Citrobacter (also a member of the family Enterobacteriaceae), were detected from infants born to IPV exposed mothers as compared to infants from IPV unexposed mothers. Our findings in relation to the family Enterobacteriaceae is of interest, given previous findings. A higher cumulative score of self-reported maternal stress during pregnancy has previously been associated with increased relative abundances of Proteobacteria in infant faecal specimens over time (Zijlmans et al., 2015). The family Enterobacteriaceae (within the phylum Proteobacteria) has also been associated with depression in an adult population (Jiang et al., 2015), and leakage of lipopolysaccharide from Enterobacteriaceae into the circulation has been associated with major depression. Increased faecal Enterobacteriaceae in infants at 3 to 6 months of age has also been associated with impaired follow-up fine motor skills in the children at 3 years (Sordillo et al., 2019). Our findings on the other hand differ from a study by Hu and colleagues that showed an association of higher scores for pregnancy-related anxiety with lower abundances of an unclassified genus in the family Enterobacteriaceae from meconium of 75 infants (Hu et al., 2019). This discrepancy may be due differences of the psychological measures and possibly the larger proportion of mothers (27% vs. 11%) that used antibiotics prior to delivery compared to our study.
We also found that infants born to mothers with IPV exposure had higher proportions of the genus Weissella (phylum Firmicutes) at 4-12 weeks of age. The relative abundance of Weissella increased in all infants with age. However, this increase occurred earlier for infants born to mothers with IPV exposure and subsequently reached the same level for all infants at 20-28 weeks of age. To the best of our knowledge, no prior associations have been reported between psychological stressors or distress and the genus Weissella. Furthermore, premature delivery has been associated with distinct infant faecal bacterial profiles, including higher abundances of Weissella (Arboleya et al., 2012). However, the infants assessed in this study sample had a median gestational age of 39 weeks (IQR: 38-40) (Claassen-Weitz et al., 2018). Moreover, there were no significant age differences (p > 0.5) between infants from mothers that were IPV-exposed vs. non IPV-exposed at any of the time points at which specimens were collected.
Our results further show that proportions of Gammaproteobacteria detected from stool decreased at 4-12 weeks and 20-28 weeks compared to profiles detected from infants at birth. This decrease was less pronounced at 4-12 weeks and 20-28 weeks after birth for infants born to mothers with a higher ratio of psychological distress (SRQ-20). Our results therefore indicate that infants from mothers with higher measures of psychological distress (SRQ-20) had a slower decrease in Gammaproteobacteria during the first six months after birth. The class Gammaproteobacteria (and family Enterobacteriaceae in particular) are the most abundant faecal bacteria in the first month of life, subsequently substituted by anaerobes (Bokulich et al., 2016). Slower reduction rates of these bacteria over time may be an indication of impaired maturation of the infant’s gut bacteriome (Bokulich et al., 2016). Furthermore, higher measures of maternal prenatal psychological distress were associated with lower relative abundance of Veillonellaceae (phylum Firmicutes) in infant faecal specimens at 20-28 weeks. The median relative abundance of Veillonellaceae increased with age overall amongst infants in this study. However, at the age of 4-12 weeks, this abundance was relatively lower (not significant) in infants from mothers with higher scores for psychological distress and more pronounced and significantly lower in these infants at the age of 20-28 weeks. Veillonellaceae are lactate utilizing bacteria that are abundant in infant faecal specimens up to 6 months and subsequently decrease during the transition to toddler and then remaining at stable levels throughout healthy adulthood (Pannaraj et al., 2017, Drell et al., 2017, Laursen et al., 2016). Decreased abundances of Veillonellaceae have been found in children with autism (Kang et al., 2013), as well as in patients with depression (Jiang et al., 2015). A recent Belgian population cohort showed that the genus Dialister (within the family Veillonellaceae) was significantly deceased in depression, a finding that was further validated in a large Dutch cohort (Valles-Colomer et al., 2019). These findings suggest a consistent association between decreased abundance of Veillonellaceae and neurodevelopmental impairment or psychological distress.
Overall, our results illustrate that the associations of different psychological profiles during pregnancy on faecal bacteria may vary. Exposure to lifetime IPV in particular, was associated with differences in the infant and maternal faecal bacteria. IPV is a traumatic stressor that often occurs over prolonged and repeated episodes (Thompson et al., 2006) and IPV during pregnancy often persists postpartum (Charles and Perreira, 2007, Groves et al., 2015, Barnett et al., 2018). It should be noted that other effects of IPV which were not controlled for in this study may contribute to its association with changes to infant faecal bacterial composition. For example, mothers who have experienced IPV are more likely to have weakened bonding with their infant (Zeitlin et al., 1999, Barnett et al., 2018), which may in turn affect the infant faecal bacterial composition. Supporting evidence from animal work showed that male rat pups that were stressed by maternal separation had elevated faecal levels of Enterobacteria and Bacteroides during adulthood (Garcia-Rodenas et al., 2006). Another animal study found that maternal separation of pups resulted in a reduced ratio of Firmicutes to Bacteroides in the adult gut of these female rats (Pusceddu et al., 2015). The effect of IPV on child feeding may also contribute to faecal bacterial changes. Studies have showed that maternal IPV can contribute to infant malnutrition in low to middle income countries (Rico et al., 2011, Misch and Yount, 2014, Barnett et al., 2018). Malnutrition has been associated with marked changes in infants’ faecal bacterial composition (Kane et al., 2015). Therefore, other measures that accompany IPV post-partum may play a role in the development of infant faecal bacterial composition.
Several limitations to our study deserve emphasis. First, there were missing faecal specimens over time, due mainly to the inability to collect stool at the scheduled study visits (stool was only collected if the infant was able to produce a stool sample during the study visit). Therefore, our results should be interpreted with caution, especially at the 20-28 weeks interval. Second, given the limited number of mothers with PTSD, our analysis has insufficient statistical power to draw definitive conclusions about the possible association between PTSD and an altered faecal bacteriome. Third, the cross-sectional analysis for the individual time points does not allow for definitive conclusions about their longitudinal changes in the faecal bacterial profiles. Further work is needed to understand whether IPV is a proxy for other key variables that may affect the associations found with the infants’ faecal gut bacteria.
In summary, maternal prenatal psychological stressors and distress, particularly IPV during pregnancy, is associated with early changes in infant faecal bacterial profiles in a semi-rural South African area. Further studies are needed to verify our findings, and to determine wheteher these changes to the gut bacteriome due to maternal prenatal psychological stressors and distress are associated with neurodevelopmental impairments in children over time. In addition, it would be of interest to examine if preventative strategies to reduce the risk of prenatal psychological stressors and distress can prevent longitudinal changes to the infant’s gut bacteria and associated health risks. Such work may provide further insights into understanding relevant mechanisms and identifying appropriate targets, with the ultimate aim of preventing the negative effects of intergenerational transmission of psychopathology on the developing brain.
Supplementary Material
Significant outcomes
This is the first study to report an association between maternal prenatal stressors and distress with an altered bacterial profile in their infants’ meconium and longitudinal stool specimens.
Maternal prenatal exposure to intimate partner violence (IPV) is associated with differences in faecal bacterial profiles of mothers at delivery (Lactobacillaceae and Peptostreptococcaceae) and of their infants at birth (family Enterobacteriaceae) and 4-12 weeks of age (genus Weissella). These infants had increased proportions of taxa within the family Enterobacteriaceae (Citrobacter; OTU 101; OTU 615; OTU 616) during the first six months after birth compared to infants from mothers with no/low IPV exposure.
Higher psychological distress during pregnancy is associated with lower infant faecal bacterial profiles of the family Veillonellaceae at 20-28 weeks.
Limitations
Missing faecal specimens over time.
Limited number of mothers with posttraumatic stress disorder.
Acknowledgements
We would like to thank the families and their children that participated in this study. We thank the study staff, administrative staff and clinical staff of the Western Cape Government Health Department at TC Newman and Mbekweni clinics, and Paarl Hospital for their support of the study. Petrus Naudé, Shantelle Claassen-Weitz, Mamadou Kaba, Heather Zar, Mark Nicol and Dan Stein contributed to the concept and design of the study. Shantelle Claassen-Weitz, Sugnet Gardner-Lubbe, Gerrit Botha and Mamadou Kaba contributed to the acquisition and analysis of data. All mentioned authors contributed to the interpretation of the results presented in this manuscript. Petrus Naudé and Shantelle Claassen-Weitz drafted the manuscript. The critical revisions by Sugnet Gardner-Lubbe, Gerrit Botha, Mamadou Kaba, Heather Zar, Mark Nicol and Dan Stein were essential for the intellectual content of this manuscript. All listed authors provided final approval of the version to be published.
Financial support
The Drakenstein Child Health Study, is funded by Bill and Melinda Gates Foundation (OPP1017641; OPP1017579). HJZ and DJS are supported by the South African Medical Research Council. This study was also supported by an H3Africa U01 award from the National Institutes of Health of the USA to MPN and HJZ (1U01AI110466-01A1). MK is a recipient of Carnegie Corporation of New York (USA) early-career fellowship, Wellcome Trust Training Fellowship, United Kingdom (102429/Z/13/Z) and CTN International fellowship (Canada). PJWN is supported by the National Alliance for Research on Schizophrenia and Depression Young Investigator Grant (No. 25199) and Scott-Gentle Foundation.
Footnotes
Statement of interest
None
Ethical standards
The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional committees on human experimentation and with the Helsinki Declaration of 1975, as revised in 2008.
References
- Anders S and Huber W (2010) Differential expression analysis for sequence count data. Genome Biology 11, R106–R106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecology 26, 32–46. [Google Scholar]
- Arboleya S, Binetti A, Salazar N, Fernández N, Solís G, Hernández-Barranco A, Margolles A, de los Reyes-Gavilán CG and Gueimonde M (2012) Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiology Ecology 79, 763–772. [DOI] [PubMed] [Google Scholar]
- Arrieta MC, Stiemsma LT, Amenyogbe N, Brown EM and Finlay B (2014) The intestinal microbiome in early life: health and disease. Frontiers in Immunology 5, 427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey MT, Lubach GR and Coe CL (2004) Prenatal stress alters bacterial colonization of the gut in infant monkeys. Journal of Pediatric Gastroenterology and Nutrition 38, 414–421. [DOI] [PubMed] [Google Scholar]
- Barnett W, Halligan S, Heron J, Fraser A, Koen N, Zar HJ, Donald KA and Stein DJ (2018) Maltreatment in childhood and intimate partner violence: A latent class growth analysis in a South African pregnancy cohort. Child Abuse and Neglect 86, 336–348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beck AT, Steer RA and Brown GK (1996) Manual for the Beck Depression Inventory-II. Psychological Corporation; San Antonio. [Google Scholar]
- Beck AT, Steer RA and Carbin MG (1988) Psychometric properties of the Beck Depression Inventory: Twenty-five years of evaluation. Clinical Psychology Review 8, 77–100. [Google Scholar]
- Benjamini Y and Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society, Series B (Methodology) 57, 289–300. [Google Scholar]
- Bernstein M, Phillips T, Zerbe A, McIntyre JA, Brittain K, Petro G, Abrams EJ and Myer L (2016) Intimate partner violence experienced by HIV-infected pregnant women in South Africa: a cross-sectional study. BMJ Open 6, e011999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bokulich NA, Chung J, Battaglia T, Henderson N, Jay M, Li H, A DL, Wu F, Perez-Perez GI, Chen Y, Schweizer W, Zheng X, Contreras M, Dominguez-Bello MG and Blaser MJ (2016) Antibiotics, birth mode, and diet shape microbiome maturation during early life. Science Translational Medicine 8, 343ra82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bray JR and Curtis JT (1957) An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecological Monographs 27, 325–349. [Google Scholar]
- Carlson AL, Xia K, Azcarate-Peril MA, Goldman BD, Ahn M, Styner MA, Thompson AL, Geng X, Gilmore JH and Knickmeyer RC (2018) Infant Gut Microbiome Associated With Cognitive Development. Biol Psychiatry 83, 148–159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charles P and Perreira KM (2007) Intimate Partner Violence During Pregnancy and 1-Year Post-Partum. Journal of Family Violence 22, 609–619. [Google Scholar]
- Claassen-Weitz S, Gardner-Lubbe S, Nicol P, Botha G, Mounaud S, Shankar J, Nierman WC, Mulder N, Budree S, Zar HJ, Nicol MP and Kaba M (2018) HIV-exposure, early life feeding practices and delivery mode impacts on faecal bacterial profiles in a South African birth cohort. Scientific Reports 8, 5078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Claassen S, du Toit E, Kaba M, Moodley C, Zar HJ and Nicol MP (2013) A comparison of the efficiency of five different commercial DNA extraction kits for extraction of DNA from faecal samples. Journal of Microbiological Methods 94, 103–110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke RK and Warwick RM (2001) Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. Primer-E Ltd: Plymouth, UK. [Google Scholar]
- D’Argenio V (2018) The Prenatal Microbiome: A New Player for Human Health. High Throughput 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drell T, Štšepetova J, Simm J, Rull K, Aleksejeva A, Antson A, Tillmann V, Metsis M, Sepp E, Salumets A and Mändar R (2017) The Influence of Different Maternal Microbial Communities on the Development of Infant Gut and Oral Microbiota. Scientific reports 7, 9940–9940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunkle KL, Jewkes RK, Brown HC, Gray GE, McIntryre JA and Harlow SD (2004) Gender-based violence, relationship power, and risk of HIV infection in women attending antenatal clinics in South Africa. Lancet 363, 1415–21. [DOI] [PubMed] [Google Scholar]
- Faith DP, Minchin PR and Belbin L (1987) Compositional dissimilarity as a robust measure of ecological distance. Vegetatio 69, 57–68. [Google Scholar]
- Falsetti SA, Resnick HS, Resick PA and Kilpatrick D (1993) The Modified PTSD Symptom Scale: A brief self-report measure of posttraumatic stress disorder. The Behavioral Therapist 16, 161–162. [Google Scholar]
- Fernandes AD, Reid JNS, Macklaim JM, McMurrough TA, Edgell DR and Gloor GB (2014) Unifying the analysis of high-throughput sequencing datasets: characterizing RNA-seq, 16S rRNA gene sequencing and selective growth experiments by compositional data analysis. Microbiome 2, 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foa EB and Tolin DF (2000) Comparison of the PTSD Symptom Scale–Interview Version and the Clinician-Administered PTSD Scale. Journal of Traumatic Stress 13, 181–191. [DOI] [PubMed] [Google Scholar]
- Garcia-Rodenas CL, Bergonzelli GE, Nutten S, Schumann A, Cherbut C, Turini M, Ornstein K, Rochat F and Corthesy-Theulaz I (2006) Nutritional approach to restore impaired intestinal barrier function and growth after neonatal stress in rats. Journal of Pediatric Gastroenterology and Nutrition 43, 16–24. [DOI] [PubMed] [Google Scholar]
- Groves AK, Moodley D, McNaughton-Reyes L, Martin SL, Foshee V and Maman S (2015) Prevalence, rates and correlates of intimate partner violence among South African women during pregnancy and the postpartum period. Maternal and child health journal 19, 487–495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gur TL, Palkar AV, Rajasekera T, Allen J, Niraula A, Godbout J and Bailey MT (2019) Prenatal stress disrupts social behavior, cortical neurobiology and commensal microbes in adult male offspring. Behavioural Brain Research 359, 886–894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gur TL, Shay L, Palkar AV, Fisher S, Varaljay VA, Dowd S and Bailey MT (2017) Prenatal stress affects placental cytokines and neurotrophins, commensal microbes, and anxiety-like behavior in adult female offspring. Brain, Behavior, and Immunity 64, 50–58. [DOI] [PubMed] [Google Scholar]
- Harding TW, de Arango MV, Baltazar J, Climent CE, Ibrahim HH, Ladrido-Ignacio L, Murthy RS and Wig NN (1980) Mental disorders in primary health care: a study of their frequency and diagnosis in four developing countries. Psychological Medicine 10, 231–241. [DOI] [PubMed] [Google Scholar]
- Hartley M, Tomlinson M, Greco E, Comulada WS, Stewart J, le Roux I, Mbewu N andRotheram-Borus MJ (2011) Depressed mood in pregnancy: Prevalence and correlates in two Cape Town peri-urban settlements. Reproductive Health 8, 9–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu J, Ly J, Zhang W, Huang Y, Glover V, Peter I, Hurd YL and Nomura Y (2019) Microbiota of newborn meconium is associated with maternal anxiety experienced during pregnancy. Developmental Psychobiology 61, 640–649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borg I and Groenen PJF (2005) Modern multidimensional scaling: Theory and applications, Springer Science & Business Media. [Google Scholar]
- Jansen HAFM, Watts, Ellsberg M, Heise L and García-Moreno (2004) Interviewer Training in the WHO Multi-Country Study on Women’s Health and Domestic Violence. Violence Against Women 10, 831–849. [Google Scholar]
- Jašarević E, Rodgers AB and Bale TL (2015) A novel role for maternal stress and microbial transmission in early life programming and neurodevelopment. Neurobiology of Stress 1, 81–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jewkes R (2002) Intimate partner violence: causes and prevention. Lancet 359, 1423–1429. [DOI] [PubMed] [Google Scholar]
- Jiang H, Ling Z, Zhang Y, Mao H, Ma Z, Yin Y, Wang W, Tang W, Tan Z, Shi J, Li L and Ruan B (2015) Altered fecal microbiota composition in patients with major depressive disorder. Brain, Behavior, and Immunity 48, 186–194. [DOI] [PubMed] [Google Scholar]
- Kane AV, Dinh DM and Ward HD (2015) Childhood malnutrition and the intestinal microbiome. Pediatric research 77, 256–262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang D-W, Park JG, Ilhan ZE, Wallstrom G, LaBaer J, Adams JB and Krajmalnik-Brown R (2013) Reduced Incidence of Prevotella and Other Fermenters in Intestinal Microflora of Autistic Children. PLOS ONE 8, e68322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koen N, Brittain K, Donald KA, Barnett W, Koopowitz S, Maré K, Zar HJ and Stein DJ (2016) Psychological trauma and posttraumatic stress disorder: risk factors and associations with birth outcomes in the Drakenstein Child Health Study. European Journal of Psychotraumatology 7, 10.3402/ejpt.v7.28720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koleff P, Gaston KJ and Lennon JJ (2003) Measuring beta diversity for presence–absence data. Journal of Animal Ecology 72, 367–382. [Google Scholar]
- Laursen MF, Andersen LBB, Michaelsen KF, Mølgaard C, Trolle E, Bahl MI and Licht TR (2016) Infant Gut Microbiota Development Is Driven by Transition to Family Foods Independent of Maternal Obesity. mSphere 1, e00069–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mari JJ and Williams P (1986) A validity study of a psychiatric screening questionnaire (SRQ-20) in primary care in the city of Sao Paulo. The British Journal of Psychiatry 148, 23–26. [DOI] [PubMed] [Google Scholar]
- McGowan TC and Kagee A (2013) Exposure to traumatic events and symptoms of post-traumatic stress among South African university students. South African Journal of Psychology 43, 327–339. [Google Scholar]
- McMurdie PJ and Holmes S (2014) Waste Not, Want Not: Why Rarefying Microbiome Data Is Inadmissible. PLOS Computational Biology 10, e1003531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Misch ES and Yount KM (2014) Intimate partner violence and breastfeeding in Africa. Matern Child Health J 18, 688–697. [DOI] [PubMed] [Google Scholar]
- Morgan XC and Huttenhower C (2012) Chapter 12: Human microbiome analysis. PLoS Comput Biol 8, e1002808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Mahony SM, Clarke G, Dinan TG and Cryan JF (2017) Early-life adversity and brain development: Is the microbiome a missing piece of the puzzle? Neuroscience 342, 37–54. [DOI] [PubMed] [Google Scholar]
- Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’hara R, Simpson GL, Solymos P, Stevens MHH and Wagner H (2013) Package ‘vegan’. Community ecology package, version 2. [Google Scholar]
- Pannaraj PS, Li F, Cerini C, Bender JM, Yang S, Rollie A, Adisetiyo H, Zabih S, Lincez PJ, Bittinger K, Bailey A, Bushman FD, Sleasman JW and Aldrovandi GM (2017) Association Between Breast Milk Bacterial Communities and Establishment and Development of the Infant Gut Microbiome. JAMA pediatrics 171, 647–654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pusceddu MM, El Aidy S, Crispie F, O’Sullivan O, Cotter P, Stanton C, Kelly P, Cryan JF and Dinan TG (2015) N-3 Polyunsaturated Fatty Acids (PUFAs) Reverse the Impact of Early-Life Stress on the Gut Microbiota. PLOS ONE 10, e0139721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- R_Development_Core_Team (2014) R Foundation for Statistical Computing. R: A language and environment for statistical computing. [Google Scholar]
- Rico E, Fenn B, Abramsky T and Watts C (2011) Associations between maternal experiences of intimate partner violence and child nutrition and mortality: findings from Demographic and Health Surveys in Egypt, Honduras, Kenya, Malawi and Rwanda. Journal of Epidemiology and Community Health 65, 360–367. [DOI] [PubMed] [Google Scholar]
- Rochat TJ, Richter LM, Doll HA, Buthelezi NP, Tomkins A and Stein A (2006) Depression among pregnant rural South African women undergoing HIV testing. JAMA 295, 1376–8. [DOI] [PubMed] [Google Scholar]
- Rumble S, Swartz L, Parry C and Zwarenstein M (1996) Prevalence of psychiatric morbidity in the adult population of a rural South African village. Psychological Medicine 26, 997–1007. [DOI] [PubMed] [Google Scholar]
- Seferovic MD, Belfort B, Major A, Chu D, Racusin D, Castro E, Carroll M, Stewart C, Versalovic J and Aagaard K (2018) 695: Visualization of intact placental microbes in both term and preterm births. American Journal of Obstetrics & Gynecology 218, S417–S418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shamu S, Abrahams N, Temmerman M, Musekiwa A and Zarowsky C (2011) A systematic review of African studies on intimate partner violence against pregnant women: prevalence and risk factors. PLoS One 6, e17591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shannon CE (1948) A Mathematical Theory of Communication. Bell System Technical Journal 27, 379–423. [Google Scholar]
- Sordillo JE, Korrick S, Laranjo N, Carey V, Weinstock GM, Gold DR, O’Connor G, Sandel M, Bacharier LB, Beigelman A, Zeiger R, Litonjua AA and Weiss ST (2019) Association of the Infant Gut Microbiome With Early Childhood Neurodevelopmental Outcomes: An Ancillary Study to the VDAART Randomized Clinical Trial. JAMA Network Open 2, e190905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stein A, Pearson RM, Goodman SH, Rapa E, Rahman A, McCallum M, Howard LM and Pariante CM (2014) Effects of perinatal mental disorders on the fetus and child. The Lancet 384, 1800–1819. [DOI] [PubMed] [Google Scholar]
- Stein DJ, Koen N, Donald KA, Adnams CM, Koopowitz S, Lund C, Marais A, Myers B, Roos A, Sorsdahl K, Stern M, Tomlinson M, van der Westhuizen C, Vythilingum B, Myer L, Barnett W, Brittain K and Zar HJ (2015) Investigating the psychosocial determinants of child health in Africa: The Drakenstein Child Health Study. Journal of Neuroscience Methods 252, 27–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stein DJ, Williams DR and Kessler RC (2009) The South African Stress and Health (SASH) study: a scientific base for mental health policy. South African Medical Journal 99, 337. [PubMed] [Google Scholar]
- Stinson LF, Boyce MC, Payne MS and Keelan JA (2019) The Not-so-Sterile Womb: Evidence That the Human Fetus Is Exposed to Bacteria Prior to Birth. Frontiers in Microbiology 10, 1124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudo N, Chida Y, Aiba Y, Sonoda J, Oyama N, Yu X-N, Kubo C and Koga Y (2004) Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. The Journal of physiology 558, 263–275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson RS, Bonomi AE, Anderson M, Reid RJ, Dimer JA, Carrell D and Rivara FP (2006) Intimate Partner Violence: Prevalence, Types, and Chronicity in Adult Women. American Journal of Preventive Medicine 30, 447–457. [DOI] [PubMed] [Google Scholar]
- Valles-Colomer M, Falony G, Darzi Y, Tigchelaar EF, Wang J, Tito RY, Schiweck C, Kurilshikov A, Joossens M, Wijmenga C, Claes S, Van Oudenhove L, Zhernakova A, Vieira-Silva S and Raes J (2019) The neuroactive potential of the human gut microbiota in quality of life and depression. Nature Microbiology 4, 623–632. [DOI] [PubMed] [Google Scholar]
- van Heyningen T, Honikman S, Myer L, Onah MN, Field S and Tomlinson M (2017) Prevalence and predictors of anxiety disorders amongst low-income pregnant women in urban South Africa: a cross-sectional study. Archives of Women’s Mental Health 20, 765–775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vythilingum B, Roos A, Faure SC, Geerts L and Stein DJ (2012) Risk factors for substance use in pregnant women in South Africa. South African Medical Journal 102, 851–854. [DOI] [PubMed] [Google Scholar]
- Willyard C (2018) Could baby’s first bacteria take root before birth? Nature 553, 264–266. [DOI] [PubMed] [Google Scholar]
- World Heatlh Organization (WHO) (2002) Handbook for Good Clinical Research Practice (GCP) - guidance for implementation [Accessed Dec 2013].
- Zar HJ, Barnett W, Myer L, Stein DJ and Nicol MP (2015) Investigating the early-life determinants of illness in Africa: the Drakenstein Child Health Study. Thorax 70, 592–594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeitlin D, Dhanjal T and Colmsee M (1999) Maternal-foetal bonding: the impact of domestic violence on the bonding process between a mother and child. Archives of Women’s Mental Health 2, 183–189. [Google Scholar]
- Zijlmans MAC, Korpela K, Riksen-Walraven JM, de Vos WM and de Weerth C (2015) Maternal prenatal stress is associated with the infant intestinal microbiota. Psychoneuroendocrinology 53, 233–245. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.