

Abstract
We aim to establish a comprehensive COVID-19 autoantigen atlas in order to understand autoimmune diseases caused by SARS-CoV-2 infection. Based on the unique affinity between dermatan sulfate and autoantigens, we identified 348 proteins from human lung A549 cells, of which 198 are known targets of autoantibodies. Comparison with current COVID data identified 291 proteins that are altered at protein or transcript level in SARS-CoV-2 infection, with 191 being known autoantigens. These known and putative autoantigens are significantly associated with viral replication and trafficking processes, including gene expression, ribonucleoprotein biogenesis, mRNA metabolism, translation, vesicle and vesicle-mediated transport, and apoptosis. They are also associated with cytoskeleton, platelet degranulation, IL-12 signaling, and smooth muscle contraction. Host proteins that interact with and that are perturbed by viral proteins are a major source of autoantigens. Orf3 induces the largest number of protein alterations, Orf9 affects the mitochondrial ribosome, and they and E, M, N, and Nsp proteins affect protein localization to membrane, immune responses, and apoptosis. Phosphorylation and ubiquitination alterations by viral infection define major molecular changes in autoantigen origination. This study provides a large list of autoantigens as well as new targets for future investigation, e.g., UBA1, UCHL1, USP7, CDK11A, PRKDC, PLD3, PSAT1, RAB1A, SLC2A1, platelet activating factor acetylhydrolase, and mitochondrial ribosomal proteins. This study illustrates how viral infection can modify host cellular proteins extensively, yield diverse autoantigens, and trigger a myriad of autoimmune sequelae. Our work provides a rich resource for studies into “long COVID” and related autoimmune sequelae.
Keywords: COVID-19, Autoimmunity, Autoantigens, Lung, Atlas, Resource
1. Introduction
To gain better understanding of the transient and chronic autoimmune symptoms caused by SARS-CoV-2 infection, we have embarked on an endeavor to establish a comprehensive autoantigenome for COVID-19. We aim to provide a comprehensive resource and atlas for the investigation of autoimmune sequelae of COVID-19 (“long COVID”). In a previous study, we identified a repertoire of autoantigens (autoAgs) from human fetal lung fibroblast HFL1 cells that are strongly tied to neurological and diverse autoimmune symptoms of COVID-19 [1]. In this study, we aim to identify additional autoAgs from human lung epithelium-like A549 cells, an adenocarcinoma cell line that is frequently used as a model host in SARS-CoV-2 infection studies.
AutoAgs were identified based on the unique affinity between autoAgs and the glycosaminoglycan dermatan sulfate (DS) that we have discovered [2,3]. AutoAgs and DS form affinity complexes that can engage strong dual BCR signaling in autoreactive B1 cells to induce autoantibody production [4]. Hence, any self-molecule capable of forming affinity complexes with DS has a high propensity to become autoantigenic. This unifying mechanism of autoantigenicity explains how seemingly unrelated self-molecules can all induce autoimmune B cell responses via a similar immunological signaling event. Based on DS-autoAg affinity, we have cataloged several hundred autoAgs from various cells and tissues [1,5–7].
COVID-19 is accompanied by a wide range of autoimmune symptoms, including multisystem inflammatory syndrome in children, immune thrombocytopenic purpura, antiphospholipid syndrome, autoimmune cytopenia, immune-mediated neurological syndromes, Guillain-Barré syndrome, connective tissue disease-associated interstitial lung disease, autoimmune hemolytic anemia, autoimmune encephalitis, systemic lupus erythematosus, optic neuritis and myelitis, and acquired hemophilia [8–15]. Numerous autoantibodies have been identified in COVID patients, including the classical ANA (antinuclear antibody) and ENA (extractable nuclear antigen) that are hallmarks of systemic autoimmune diseases, as well as others such as anti-neutrophil cytoplasmic antibody, lupus anticoagulant, antiphospholipid, anti-IFN, anti-myelin oligodendrocyte glycoprotein, and anti-heparin-PF4 complex antibodies [8–15].
SARS-CoV-2, or viruses in general, are opportunistic intracellular pathogens that rely on the host for replication and survival. They hijack the host transcription and translation machinery for their replication, they compromise the host immune defense to evade destruction, and they modulate the host cell cycle and apoptosis for symbiosis. These viral processes are accomplished through extensive modification of host cellular components, which also results in changes in self-molecules and the emergence of autoAgs. In our previous studies, we reported that self-molecules derived from apoptotic cells display strong affinity to DS, becoming a major source of autoAgs [2,3]. In this study, we report several important molecular mechanisms in SARS-CoV-2 infection that change host self-molecules to autoAgs, including direct interaction with viral components, perturbation by viral protein expression, and post-translational protein modification by ubiquitination and phosphorylation from viral infection.
2. Materials and methods
2.1. A549 cell culture
The A549 cell line was obtained from the ATCC (Manassas, VA, USA) and cultured in complete F–12K medium at 37 °C in 75 cm2 flasks to 80% confluency. The growth medium was supplemented with 10% fetal bovine serum and a penicillin-streptomycin-glutamine mixture (Thermo Fisher).
2.2. Protein extraction
About 100 million A549 cells were suspended in 10 ml of 50 mM phosphate buffer (pH 7.4) containing the Roche Complete Mini protease inhibitor cocktail. Cells were homogenized on ice with a microprobe sonicator until the turbid mixture turned nearly clear with no visible cells left. The homogenate was centrifuged at 10,000 g at 4 °C for 20 min, and the total protein extract in the supernatant was collected. Protein concentration was measured by absorbance at 280 nm using a NanoDrop UV–Vis spectrometer (ThermoFisher).
2.3. DS-sepharose resin preparation
The DS-affinity resins were prepared as previously described [3,5]. In brief, 2 ml of EAH Sepharose 4B resins (GE Healthcare Life Sciences) were washed with distilled water three times and mixed with 10 mg of DS (Sigma-Aldrich) in 1 ml of 0.1 M MES buffer, pH 5.0. About 20 mg of N-(3-dimethylaminopropyl)-N′-ethylcarbodiimide hydrochloride (Sigma-Aldrich) powder was added at the beginning of the reaction, and another 20 mg was added after 8 h of reaction. The reaction mixture was mixed by end-over-end rotation at 25 °C for 16 h. The coupled resins were washed with water three times and equilibrated with a low-pH buffer (0.1 M acetate, 0.5 M NaCl, pH 5.0) and a high-pH buffer (0.1 M Tris, 0.5 M NaCl, pH 8.0).
2.4. DS-affinity fractionation
The total proteins extracted from A549 cells were fractionated in a DS-Sepharose column with a BioLogic Duo-Flow system (Bio-Rad). About 40 mg of proteins in 40 ml of 10 mM phosphate buffer (pH 7.4; buffer A) were loaded onto the column at a rate of 1 ml/min. Unbound and weakly proteins were washed off with 60 ml of buffer A and then 40 ml of 0.2 M NaCl in buffer A. The remaining bound proteins were eluted with 40 ml 0.5 M NaCl and then with 40 ml 1.0 M NaCl in buffer A. Fractions were desalted and concentrated to 0.5 ml with 5-kDa cut-off Vivaspin centrifugal filters (Sartorius). Fractionated proteins were separated by 1-D SDS-PAGE in 4–12% Bis-Tris gels, and the gel lanes were divided into two or three sections and subjected to sequencing.
2.5. Mass spectrometry sequencing
Protein sequencing was performed at the Taplin Biological Mass Spectrometry Facility at Harvard Medical School. Proteins in gels were digested with sequencing-grade trypsin (Promega) at 4 °C for 45 min. Tryptic peptides were separated on a nano-scale C18 HPLC capillary column and analyzed in an LTQ linear ion-trap mass spectrometer (Thermo Fisher). Peptide sequences and protein identities were assigned by matching the measured fragmentation pattern with proteins or translated nucleotide databases using Sequest. All data were manually inspected. Only proteins with ≥2 peptide matches were considered positively identified.
2.6. COVID data comparison
DS-affinity proteins were compared with currently available proteomic and transcriptomic data from SARS-CoV-2 infection compiled in the Coronascape database (as of 12/14/2020) [16–36]. These data had been obtained with proteomics, phosphoproteomics, interactome, ubiquitome, and RNA-seq techniques. Up- and down-regulated proteins or genes were identified by comparing COVID-19 patients vs. healthy controls and cells infected vs. uninfected by SARS-CoV-2. Similarity searches were conducted between our data and the Coronascape database to identify DS-affinity proteins (or their corresponding genes) that are up- and/or down-regulated in the viral infection.
2.7. Protein-protein interaction network analysis
Protein-protein interactions were analyzed by STRING [37]. Interactions include both direct physical interaction and indirect functional associations, which are derived from genomic context predictions, high-throughput lab experiments, co-expression, automated text mining, and previous knowledge in databases. Each interaction is annotated with a confidence score from 0 to 1, with 1 being the highest, indicating the likelihood of an interaction to be true. Only interactions with high confidence (a minimum score of 0.7) are shown in the figures.
2.8. Pathway and process enrichment analysis
Pathways and processes enrichment were analyzed with Metascape [16], which utilize various ontology sources such as KEGG Pathway, GO Biological Process, Reactome Gene Sets, Canonical Pathways, CORUM, TRRUST, and DiGenBase. All genes in the genome were used as the enrichment background. Terms with a p-value <0.01, a minimum count of 3, and an enrichment factor (ratio between the observed counts and the counts expected by chance) > 1.5 were collected and grouped into clusters based on their membership similarities. The most statistically significant term within a cluster was chosen to represent the cluster. Hierarchical clustering trees were obtained with ShinyGo [38].
2.9. Autoantigen confirmation literature text mining
Literature searches in Pubmed were performed for every DS-affinity protein identified in this study. Search keywords included the protein name, its gene symbol, alternative names and symbols, and the MeSH keyword “autoantibodies”. Only proteins with their specific autoantibodies reported in PubMed-listed journal articles were considered “confirmed” autoAgs in this study.
3. Results and discussion
3.1. A putative A549 autoantigenome identified by DS-affinity
By DS-affinity fractionation and mass spectrometry sequencing, we identified a global putative autoantigenome of 348 proteins from A549 cellular protein extracts, with 214 protein having strong affinity and 134 having intermediate affinity (Table 1). To find out whether these DS-affinity proteins are known autoAgs, we conducted an extensive literature search and confirmed that 198 (56.0%) proteins are known humoral autoAgs, with their specific autoantibodies reported in a wide spectrum of autoimmune diseases and cancers (see autoAg confirmatory references in Table 1). The remaining 150 proteins may be yet-to-be discovered putative autoAgs and await further investigation. For example, many ribosomal proteins are known autoAgs, but the 24 mitochondrial ribosomal proteins we identified have not yet been reported as autoAgs; given their structural similarity to ribosomal protein autoAgs, it is highly likely that mitochondrial proteins are a group of undiscovered autoAgs.
Table 1.
DS-affinity autoantigenome from human A549 cells.
| # Pep. | Gene | Protein | COVID | A549 infection | Interactome | DS-affinity | AutoAg ref. | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Up | Down | Up | Down | 1.0 M | 0.5 M | |||||
| 2 | A2M | Alpha-2-macroglobulin | D | + | [54] | |||||
| 2 | ACLY | ATP-citrate synthase | U | D | U | D | + | [55] | ||
| 9 | ACTA2 | Actin, aortic smooth muscle | U | D | U | D | + | [56] | ||
| 8 | ACTB | Actin, cytoplasmic 1 | U | D | U | D | + | [57] | ||
| 5 | ACTBL2 | Beta-actin-like protein 2 | U | D | U | D | + | [57] | ||
| 20 | ACTN1 | Alpha-actinin-1 | U | D | U | D | + | [58] | ||
| 20 | ACTN4 | Alpha-actinin-4 | U | D | U | + | [56] | |||
| 2 | ADSS2 | Adenylosuccinate synthetase isozyme 2 | U | U | + | |||||
| 3 | AFP | Alpha-fetoprotein | D | + | [59] | |||||
| 2 | AGRN | Agrin | U | U | + | [60] | ||||
| 6 | AHCY | Adenosylhomocysteinase | D | D | + | |||||
| 5 | AKR1B1 | Aldose reductase (diabetic complication) | U | D | U | D | Orf3 | + | [61] | |
| 6 | ALB | Putative uncharacterized protein ALB | U | D | U | D | + | [62] | ||
| 23 | ALDH1A1 | Retinal dehydrogenase | U | D | U | D | + | [63] | ||
| 5 | ALDH2 | Aldehyde dehydrogenase, mitochondrial | U | D | + | [64] | ||||
| 5 | ALDH3A1 | Aldehyde dehydrogenase, dimeric NADP-preferring, ALDH3 | U | D | U | D | + | |||
| 9 | ALDOA | Fructose-bisphosphate aldolase | U | D | U | D | + | [65] | ||
| 6 | ANP32A | Acidic leucine-rich nuclear phosphoprotein 32 family member A | U | D | D | + | ||||
| 13 | ANP32B | Acidic leucine-rich nuclear phosphoprotein 32 family member B, APRIL | D | D | + | [66] | ||||
| 3 | ANP32C | Acidic leucine-rich nuclear phosphoprotein 32 family member C, PP32R1 | + | |||||||
| 4 | ANP32E | Acidic leucine-rich nuclear phosphoprotein 32 family member E | U | D | U | D | + | |||
| 4 | ANXA2 | Annexin A2 | U | D | U | + | [67] | |||
| 13 | ANXA2P2 | Annexin A2 pseudogene 2 | U | D | U | + | [67] | |||
| 10 | ANXA3 | Annexin A3 | U | D | U | + | [68] | |||
| 5 | ANXA4 | Annexin IV | U | D | U | D | + | [69] | ||
| 15 | ANXA5 | Annexin A5 | U | D | U | D | Orf3 | + | [70] | |
| 8 | ANXA6 | Annexin VI | U | D | + | [71] | ||||
| 8 | AP3B1 | AP-3 complex subunit beta-1 | U | E | + | |||||
| 2 | AP3B2 | AP-3 complex subunit beta-2 | + | [72] | ||||||
| 8 | AP3D1 | AP-3 complex subunit delta-1 | U | D | + | |||||
| 4 | APEX1 | DNA-(apurinic or apyrimidinic site) endonuclease | U | D | U | D | + | [73] | ||
| 4 | ASPH | Aspartyl/asparaginyl beta-hydroxylase | U | D | D | + | ||||
| 2 | ATP2A2 | ATP Sarcoplasmic/ER calcium transporting 2 (cardiac Ca2+ ATPase, heart failure) | U | Nsp4 | + | [74] | ||||
| 13 | ATP5B | ATP synthase subunit beta, mitochondrial, ATP5F1B | U | D | Nsp6 | + | [75] | |||
| 3 | BRIX1 | Ribosome biogenesis protein BRX1 homolog | + | |||||||
| 8 | C1QBP | Complement C1q-binding protein, mitochondrial matrix protein p32 | D | D | + | [76] | ||||
| 10 | CALR | Calreticulin | U | D | U | + | [77] | |||
| 7 | CANX | Calnexin | U | D | U | D | Nsp4 | + | [78] | |
| 2 | CAP1 | Cyclase associated actin cytoskeleton regulatory protein 1 | U | D | Orf3 | + | ||||
| 2 | CAPN1 | Calpain-1 catalytic subunit | + | |||||||
| 2 | CAPZB | F-actin-capping protein subunit beta | D | + | [79] | |||||
| 5 | CCT8 | T-complex protein 1 subunit theta | U | D | U | D | + | [80] | ||
| 3 | CDK11A | Cyclin-dependent kinase 11A, CDC2L2 | U | + | ||||||
| 3 | CEBPZ | CCAAT/enhancer-binding protein zeta | U | + | ||||||
| 4 | CLIC1 | Chloride intracellular channel protein 1 | U | D | + | [81] | ||||
| 4 | CLTC | Clathrin heavy chain 1 | U | D | D | + | [3] | |||
| 2 | CLTCL1 | Clathrin heavy chain 2 | + | |||||||
| 2 | COPA | Coatomer subunit alpha | U | D | U | D | + | [82] | ||
| 2 | CPNE3 | Copine-3 | U | D | + | |||||
| 2 | CTR9 | RNA polymerase-associated protein CTR9 homolog | U | D | D | + | ||||
| 4 | DCAF1 | DDB1- and CUL4-associated factor 1, VPRBP | U | D | U | + | ||||
| 13 | DDB1 | DNA damage-binding protein 1 | U | D | U | D | + | [3] | ||
| 3 | DDX17 | Probable ATP-dependent RNA helicase DDX17 | U | D | U | + | ||||
| 7 | DDX18 | ATP-dependent RNA helicase DDX18 | U | + | ||||||
| 5 | DDX21 | Nucleolar RNA helicase 2 (RH II/Gu) | U | U | N | + | [83] | |||
| 4 | DDX27 | Probable ATP-dependent RNA helicase DDX27 | U | U | + | |||||
| 3 | DDX30 | ATP-dependent RNA helicase DHX30, DDX30 | + | |||||||
| 8 | DDX48 | Eukaryotic initiation factor 4A-III, EIF4A3 | + | |||||||
| 4 | DDX5 | Probable ATP-dependent RNA helicase DDX5 | U | D | + | |||||
| 16 | DDX9 | ATP-dependent RNA helicase A, DHX9 | + | [84] | ||||||
| 12 | DHX15 | Putative pre-mRNA-splicing factor ATP-dependent RNA helicase DHX15 | D | + | ||||||
| 4 | DHX36 | Probable ATP-dependent RNA helicase DHX36 | U | + | ||||||
| 4 | DKC1 | H/ACA ribonucleoprotein complex subunit B | U | D | D | + | ||||
| 3 | DPP3 | Dipeptidyl-peptidase 3 | D | + | ||||||
| 3 | DYNC1I2 | Cytoplasmic dynein 1 intermediate chain 2 | + | |||||||
| 3 | EBP2 | Probable rRNA-processing protein, EBNA1BP2 | + | |||||||
| 4 | ECH1 | Delta(3,5)-Delta(2,4)-dienoyl-CoA isomerase, mitochondrial | U | D | U | + | [85] | |||
| 2 | EEF1A1 | Elongation factor 1-alpha 1 | U | D | U | D | + | [86] | ||
| 2 | EEF1A2 | Elongation factor 1-alpha 2 | U | U | Orf3 | + | [87] | |||
| 2 | EEF1G | Elongation factor 1-gamma | U | D | U | + | ||||
| 3 | EEF2 | Elongation factor 2 | U | D | U | + | [88] | |||
| 16 | EFTUD2 | 116 kDa U5 small nuclear ribonucleoprotein component | D | D | + | [89] | ||||
| 2 | EIF2A | Eukaryotic translation initiation factor 2 subunit 1, EIF2 | + | |||||||
| 6 | EIF3A | Eukaryotic translation initiation factor 3 subunit A | U | D | D | + | [90] | |||
| 4 | EIF3B | Eukaryotic translation initiation factor 3 subunit | U | D | D | + | ||||
| 3 | EIF3CL | Eukaryotic translation initiation factor 3 subunit C-like protein | D | D | + | |||||
| 2 | EIF3E | Eukaryotic translation initiation factor 3 subunit E | U | D | U | + | [91] | |||
| 7 | EIF3L | Eukaryotic translation initiation factor 3, subunit E interacting protein | D | + | ||||||
| 13 | EIF4A1 | Eukaryotic initiation factor 4A-I, DDX2A | U | D | U | D | + | |||
| 2 | EIF5A | Eukaryotic translation initiation factor 5A-1 | U | D | U | D | + | [92] | ||
| 2 | EMG1 | Ribosomal RNA small subunit methyltransferase NEP1 | U | D | + | |||||
| 3 | ENO1 | Isoform alpha-enolase of Alpha-enolase | U | D | U | D | + | [93] | ||
| 6 | EPHX1 | Epoxide hydrolase | D | D | + | [94] | ||||
| 3 | ESYT1 | Extended synaptotagmin-1 | + | [95] | ||||||
| 3 | EZR | Ezrin | U | D | U | D | + | [96] | ||
| 3 | FKBP4 | FK506-binding protein, FKBP52 | + | [97] | ||||||
| 27 | FLNA | Filamin-A | U | D | U | D | + | [98] | ||
| 25 | FLNB | Filamin-B | U | U | + | [3] | ||||
| 9 | FLNC | Filamin-C | U | D | + | [99] | ||||
| 2 | FTH1 | Ferritin heavy chain | U | D | U | D | + | [100] | ||
| 10 | G6PD | Glucose-6-phosphate 1-dehydrogenase (anemia, diabetes) | U | D | U | + | [101] | |||
| 5 | GANAB | Neutral alpha-glucosidase A | D | D | + | [102] | ||||
| 2 | GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | U | D | D | + | [103] | |||
| 2 | GAR1 | H/ACA ribonucleoprotein complex subunit 1 | + | |||||||
| 2 | GBE1 | 1,4-alpha-glucan-branching enzyme | U | + | ||||||
| 4 | GCLC | Glutamate-cysteine ligase catalytic subunit (hemolytic anemia) | Orf3 | + | ||||||
| 6 | GDI1 | Rab GDP dissociation inhibitor alpha | U | D | U | + | [104] | |||
| 7 | GDI2 | cDNA FLJ60299, highly similar to Rab GDP dissociation inhibitor beta | U | D | U | + | [105] | |||
| 2 | GLO1 | Lactoylglutathione lyase (prostasome) | D | Orf3 | + | [106] | ||||
| 2 | GPC1 | Glypican-1 | D | D | + | |||||
| 2 | GPI | Glucose-6-phosphate isomerase | U | D | U | D | Orf3 | + | [107] | |
| 4 | GRWD1 | Glutamate-rich WD repeat-containing protein 1 | + | |||||||
| 6 | GSTP1 | Glutathione S-transferase | U | D | D | + | [108] | |||
| 2 | H2AFR | Histone H2A type 1-A, H2AC1, HIST1H2AA | + | [109] | ||||||
| 3 | H2AFV | Histone H2A.V, H2AZ2 | D | D | + | [109] | ||||
| 11 | H2AFY | H2A histone family, member Y isoform, MacroH2A1 | U | + | [110] | |||||
| 2 | H2AFY2 | Core histone macro-H2A.2, MACROH2A2 | + | [109] | ||||||
| 4 | HADHA | Trifunctional enzyme subunit alpha, mitochondrial | + | |||||||
| 2 | HARS | Histidyl-tRNA synthetase, cytoplasmic, HRS | + | [10] | ||||||
| 4 | HEATR1 | HEAT repeat-containing protein 1 | U | D | U | D | + | |||
| 2 | HIST1H1B | Histone H1.5, H1–5 | U | D | D | + | [111] | |||
| 6 | HIST1H1C | Histone H1.2 | U | D | U | D | + | [111] | ||
| 2 | HIST1H2BL | Histone H2B type 1-L, H2BC13 | U | D | U | D | + | [112] | ||
| 9 | HIST1H4J | Histone H4, H4C1 | U | D | U | D | + | [113] | ||
| 12 | HIST2H2BE | Histone H2B type 2-E, H2BC21 | U | D | D | + | [114] | |||
| 5 | HIST2H3A | HIST2H3C, Histone H3.2, H3C15 | U | D | U | + | [115] | |||
| 3 | HNRNPA2B1 | Heterogenous nuclear ribonucleoproteins A2/B1 | U | D | + | [116] | ||||
| 3 | HNRNPC | Heterogeneous nuclear ribonucleoproteins C1/C2 | U | D | U | D | + | [117] | ||
| 6 | HNRNPCL1 | Heterogeneous nuclear ribonucleoprotein C-like 1 | + | |||||||
| 4 | HNRNPF | Heterogeneous nuclear ribonucleoprotein F | D | D | + | [118] | ||||
| 2 | HNRNPH1 | Heterogeneous nuclear ribonucleoprotein H | U | D | U | + | [118] | |||
| 3 | HNRNPK | Heterogeneous nuclear ribonucleoprotein K | U | + | [119] | |||||
| 3 | HNRNPM | Heterogeneous nuclear ribonucleoprotein M | U | D | D | + | ||||
| 4 | HNRNPR | Heterogeneous nuclear ribonucleoprotein R | U | D | U | + | [120] | |||
| 2 | HNRNPU | Heterogeneous nuclear ribonucleoprotein U | U | D | U | D | + | |||
| 6 | HNRNPUL1 | hnRNP U-like protein 1 | U | D | D | + | ||||
| 4 | HNRNPUL2 | hnRNP U-like protein 2 | U | D | + | |||||
| 4 | HSP70T | Heat shock 70 kDa protein 1-like, HSPA1L | + | |||||||
| 11 | HSP90AA1 | Heat shock 90 kDa protein 1, alpha isoform | U | D | U | + | [121] | |||
| 3 | HSP90AA2 | Heat shock protein HSP 90-alpha A2 | U | D | U | + | [122] | |||
| 5 | HSP90AB1 | Heat shock protein HSP 90-beta | U | D | U | + | [123] | |||
| 23 | HSP90B1 | Endoplasmin | U | D | U | + | [124] | |||
| 7 | HSPA1A | Heat shock 70 kDa protein 1A | U | D | U | D | N, Orf9b | + | [125] | |
| 35 | HSPA5 | HSPA5 protein, BiP, GRP78 | U | D | U | D | Nsp2 Nsp4 |
+ | [126] | |
| 2 | HSP70B | Putative heat shock 70 kDa protein, HSPA7 | U | D | U | + | ||||
| 20 | HSPA8 | Heat shock cognate 71 kDa protein | U | D | U | Nsp2 | + | [127] | ||
| 17 | HSPA9 | Stress-70 protein, mitochondrial | U | D | D | N | + | [127] | ||
| 2 | HSPBP1 | Hsp70-binding protein | U | D | + | [128] | ||||
| 3 | HSPG2 | U | D | U | + | [129] | ||||
| Basement membrane-specific heparan sulfate proteoglycan core | ||||||||||
| protein | ||||||||||
| 4 | HTATSF1 | HIV Tat-specific factor 1 | D | D | + | |||||
| 4 | IDE | Insulin-degrading enzyme (insulin, amyloid) | Nsp4 | + | ||||||
| 2 | IL18 | Interleukin-18 | U | D | U | D | + | [130] | ||
| 5 | ILF2 | Interleukin enhancer-binding factor | U | U | + | [131] | ||||
| 9 | IQGAP1 | Ras GTPase-activating-like protein IQGAP1 | U | + | [132] | |||||
| 4 | ITGB1 | Integrin beta-1 | U | D | D | Nsp4 Orf8 |
+ | [133] | ||
| 2 | IWS1 | Protein IWS1 homolog | U | D | U | D | + | |||
| 3 | KARS | Lysyl-tRNA synthetase | + | [134] | ||||||
| 10 | KPNB1 | Importin subunit beta-1, IPOB, | + | [135] | ||||||
| 2 | KRR1 | KRR1 small subunit processome component homolog, HIV-1 Rev-binding protein | + | [136] | ||||||
| 2 | KYNU | Kynureninase | U | U | Orf3 | + | ||||
| 2 | LAMP2 | Lysosome-associated membrane glycoprotein 2 | U | D | U | D | + | [137] | ||
| 2 | LARS | Leucyl-tRNA synthetase, cytoplasmic | + | [134] | ||||||
| 3 | LCP1 | Plastin-2 | U | D | + | [138] | ||||
| 8 | LDHA | L-lactate dehydrogenase | U | D | U | D | + | [139] | ||
| 9 | LDHB | L-lactate dehydrogenase B chain | U | D | U | D | + | [139] | ||
| 2 | LEO1 | RNA polymerase-associated protein LEO1 | U | U | + | |||||
| 6 | LMNA | Lamin-A/C | U | D | U | D | + | [140] | ||
| 2 | LMNB2 | Lamin-B | U | D | + | [141] | ||||
| 4 | LRPPRC | Leucine-rich PPR motif-containing protein, mitochondrial | D | + | [142] | |||||
| 3 | MAP1B | Microtubule-associated protein 1B | U | D | U | D | + | [143] | ||
| 2 | MARS | Methionyl-tRNA synthetase, cytoplasmic, SLA2 | D | D | + | |||||
| 3 | MDH2 | Malate dehydrogenase, mitochondrial | U | D | + | [144] | ||||
| 4 | MOV10 | Helicase MOV-10, Moloney Leukemia virus 10 protein | U | D | D | N | + | |||
| 5 | MRPL1 | 39S ribosomal protein L1, mitochondrial | + | |||||||
| 3 | MRPL13 | 39S ribosomal protein L13, mitochondrial | D | D | + | |||||
| 2 | MRPL15 | 39S ribosomal protein L15, mitochondrial | U | D | D | + | ||||
| 2 | MRPL17 | 39S ribosomal protein L17, mitochondrial | D | D | + | |||||
| 2 | MRPL18 | 39S ribosomal protein L18, mitochondrial | D | D | + | |||||
| 4 | MRPL19 | 39S ribosomal protein L19, mitochondrial | D | D | + | |||||
| 2 | MRPL2 | 39S ribosomal protein L2, mitochondrial | D | D | + | |||||
| 2 | MRPL23 | 39S ribosomal protein L23, mitochondrial | D | + | ||||||
| 5 | MRPL37 | 39S ribosomal protein L37, mitochondrial | U | D | D | + | ||||
| 5 | MRPL38 | 39S ribosomal protein L38, mitochondrial | D | D | + | |||||
| 2 | MRPL39 | 39S ribosomal protein L39, mitochondrial | D | D | + | |||||
| 3 | MRPL45 | 39S ribosomal protein L45, mitochondrial | D | D | + | |||||
| 2 | MRPL49 | 39S ribosomal protein L49, mitochondrial | D | D | + | |||||
| 4 | MRPS22 | 28S ribosomal protein S22, mitochondrial | + | |||||||
| 4 | MRPS23 | 28S ribosomal protein S23, mitochondrial | + | |||||||
| 6 | MRPS27 | 28S ribosomal protein S27, mitochondrial | Nsp8 | + | ||||||
| 2 | MRPS28 | 28S ribosomal protein S28, mitochondrial, MRPS35 | + | |||||||
| 6 | MRPS29 | 28S ribosomal protein S29, mitochondrial, DAP3 | + | |||||||
| 2 | MRPS30 | 28S ribosomal protein S30, mitochondrial | D | D | + | |||||
| 2 | MRPS34 | 28S ribosomal protein S34, mitochondrial | D | + | ||||||
| 16 | MRPS39 | Pentatricopeptide repeat-containing protein 3, mitochondrial, PTCD3 | + | |||||||
| 3 | MRPS9 | 28S ribosomal protein S9, mitochondrial | + | |||||||
| 4 | MSN | Moesin (membrane organizing extension spike protein) | U | U | Nsp6Orf3 | + | [145] | |||
| 12 | MVP | Major vault protein | U | D | U | + | [146] | |||
| 16 | MYBBP1A | Myb-binding protein 1A | U | D | U | + | ||||
| 2 | MYG1 | UPF0160 protein MYG1, mitochondrial | + | |||||||
| 14 | MYH9 | Myosin-9 | U | D | U | D | + | [147] | ||
| 2 | MYO1E | Unconventional myosin-Ie, MYO1C | U | + | [148] | |||||
| 3 | NAP1L1 | Nucleosome assembly protein 1-like 1 | U | D | D | + | ||||
| 18 | NCL | Nucleolin | U | D | + | [149] | ||||
| 8 | NME1 | Nucleoside diphosphate kinase | U | D | D | + | [150] | |||
| 3 | NME2 | Nucleoside diphosphate kinase 2, NM23 | U | D | + | [151] | ||||
| 2 | NOC2L | Nucleolar complex protein 2 homolog | D | D | + | |||||
| 9 | NOP2 | Probable 28S rRNA (cytosine(4447)-C(5)-methyltransferase | U | U | + | |||||
| 2 | NPEPPS | Puromycin-sensitive aminopeptidase, metalloproteinase MP100 | + | |||||||
| 6 | NPM1 | Nucleophosmin | U | D | U | D | + | [152] | ||
| 2 | NRCAM | Neuronal cell adhesion molecule | U | D | U | D | + | [153] | ||
| 4 | NUDT21 | Cleavage and polyadenylation specificity factor subunit 5 | D | D | + | |||||
| 2 | OLA1 | Obg-like ATPase 1 | U | U | + | |||||
| 11 | P4HB | Protein disulfide-isomerase | U | D | U | D | + | [154] | ||
| 22 | PABPC1 | Polyadenylate-binding protein 1 | D | D | N | + | [155] | |||
| 11 | PABPC4 | Polyadenylate-binding protein 4 | D | D | N | + | [156] | |||
| 4 | PAF1 | RNA polymerase II-associated factor 1 homolog | D | + | ||||||
| 2 | PAFAH1B2 | Platelet-activating factor acetylhydrolase IB subunit beta | U | D | U | D | + | |||
| 3 | PAFAH1B3 | Platelet-activating factor acetylhydrolase IB subunit gamma | U | U | + | |||||
| 6 | PCNA | Proliferating cell nuclear antigen | U | D | U | D | + | [157] | ||
| 12 | PDIA3 | Protein disulfide-isomerase A3 | U | D | U | Orf8 | + | [158] | ||
| 18 | PDIA4 | Protein disulfide-isomerase A4 | U | D | U | + | ||||
| 10 | PDIA6 | Protein disulfide-isomerase A6 | U | D | U | D | + | [159] | ||
| 6 | PELP1 | Proline-, glutamic acid-, leucine-rich protein 1 | D | D | + | |||||
| 2 | PES1 | Pescadillo homolog | D | + | ||||||
| 2 | PFAS | Phosphoribosylformylglycinamidine synthase, GATD8, FGARAT, PURL | + | |||||||
| 2 | PFKP | ATP-dependent 6-phofructokinase, platelet type | U | D | U | Orf7a | + | [160] | ||
| 2 | PGAM2 | Phosphoglycerate mutase, PGAMM | + | |||||||
| 9 | PGD | 6-phosphogluconate dehydrogenase, decarboxylating | U | D | U | + | ||||
| 2 | PLD3 | Phospholipase D3, 5’−3’ exonuclease PLD3 | U | D | D | Nsp2Orf8 Orf7b |
+ | |||
| 39 | PLEC | Plectin-1 | U | D | U | D | + | [161] | ||
| 2 | PLS1 | Plastin-1 | D | + | ||||||
| 5 | PLS3 | Plastin-3 | U | D | + | |||||
| 2 | POP1 | Ribonucleases P/MRP protein subunit POP1 | U | + | [162] | |||||
| 3 | POR | NADPH-cytochrome P450 reductase | U | D | U | Nsp2 | + | |||
| 2 | PPA1 | Inorganic pyrophosphatase (Guillain-Barre syndrome) | U | Orf3 | + | [163] | ||||
| 5 | PPIB | Peptidyl-prolyl cis-trans isomerase | U | D | U | + | [164] | |||
| 6 | PRDX1 | Peroxiredoxin-1 | U | D | U | D | + | [165] | ||
| 4 | PRDX3 | Thioredoxin-dependent peroxide reductase, mitochondrial | U | D | U | + | [166] | |||
| 3 | PRDX4 | Peroxiredoxin-4 | U | D | + | [167] | ||||
| 17 | PRKDC | DNA-dependent protein kinase catalytic subunit | U | D | U | M Nsp4 |
+ | [168] | ||
| 3 | PRMT1 | HMT1, hnRNP methyltransferase-like 2 isoform | D | D | + | |||||
| 19 | PRPF8 | Pre-mRNA-processing-splicing factor 8 | U | D | U | D | + | [3] | ||
| 6 | PSAT1 | Phosphoserine aminotransferase 1 | U | D | U | Orf3 Orf7a |
+ | |||
| 2 | PSMD1 | 26S proteasome non-ATPase regulatory subunit 1 | U | U | + | |||||
| 2 | PSMD6 | 26S proteasome non-ATPase regulatory subunit 6, PFAAP4 | + | |||||||
| 18 | PUM1 | Pumilio homolog 1 | D | D | + | |||||
| 3 | PURA | Transcriptional activator protein Pur-alpha | U | D | U | D | + | |||
| 2 | PURH | Bifunctional purine biosynthesis protein, ATIC | + | [169] | ||||||
| 4 | QARS | Glutaminyl-tRNA synthetase | + | [134] | ||||||
| 3 | RAB1A | Ras-related protein Rab-1A (intracellular membrane trafficking) | D | Nsp7 Orf3 Orf7b |
+ | |||||
| 6 | RALY | RNA-binding protein, autoantigen p542 | U | D | U | D | + | [170] | ||
| 2 | RANGAP1 | Ran GTPase-activating protein 1 | D | + | [171] | |||||
| 3 | RARS | Arginyl-tRNA synthetase, cytoplasmic, RARS1 | U | U | + | |||||
| 5 | RBBP4 | Histone-binding protein RBBP4 | D | + | [172] | |||||
| 3 | RBM8A | RNA-binding protein 8A | U | + | ||||||
| 8 | RDX | Radixin | Nsp13 Orf3 |
+ | [173] | |||||
| 3 | RNPEP | Arginine aminopeptidase, APB | + | |||||||
| 2 | RNPS1 | RNA-binding protein with serine-rich domain 1 | U | D | U | D | + | |||
| 4 | RO60 | 60 kDa SS-A/Ro ribonucleoprotein | U | U | + | [174] | ||||
| 3 | RPF2 | Ribosome production factor 2 homolog, BXDC1 | + | |||||||
| 4 | RPL10A | 60S ribosomal protein L10A, NEDD6 | + | |||||||
| 2 | RPL11 | 60S ribosomal protein L11 | U | U | + | |||||
| 4 | RPL12 | 60S ribosomal protein L12 | U | D | U | + | [175] | |||
| 3 | RPL15 | 60S ribosomal protein L15 | D | D | + | |||||
| 3 | RPL18 | 60S ribosomal protein L18 | D | D | + | |||||
| 2 | RPL26L1 | 60S ribosomal protein L26-like 1, RPL26P1 | + | |||||||
| 2 | RPL35A | 60S ribosomal protein L35a | U | D | U | + | [176] | |||
| 2 | RPL4 | 60S ribosomal protein L4 | U | D | U | D | + | |||
| 17 | RPL5 | 60S ribosomal protein L5 | D | D | + | [177] | ||||
| 11 | RPL6 | 60S ribosomal protein L6 | U | D | U | D | + | [178] | ||
| 9 | RPL7 | 60S ribosomal protein L7, RPL7P32 | U | D | U | D | + | [179] | ||
| 4 | RPL7A | 60S ribosomal protein L7A | U | D | D | + | [176] | |||
| 2 | RPL8 | 60S ribosomal protein L8 | U | D | U | D | + | [171] | ||
| 8 | RPLP0 | 60S acidic ribosomal protein P0 | U | D | U | + | [180] | |||
| 2 | RPLP1 | 60S acidic ribosomal protein P1 | U | D | D | + | [181] | |||
| 3 | RPLP2 | 60S acidic ribosomal protein P2 | U | D | U | D | + | [181] | ||
| 2 | RPS15A | 40S ribosomal protein S15a | U | + | ||||||
| 3 | RPS18 | 40S ribosomal protein S18 | U | D | + | [171] | ||||
| 3 | RPS2 | 40S ribosomal protein S2 | U | D | U | D | + | |||
| 3 | RPS3 | 40S ribosomal protein S3 | U | D | U | + | [182] | |||
| 4 | RPS4X | 40S ribosomal protein S4, X isoform | D | D | + | |||||
| 3 | RPS6 | 40S ribosomal protein S6 | U | D | U | D | + | [176] | ||
| 2 | RPS8 | 40S ribosomal protein S8 | U | D | U | + | ||||
| 8 | RPS9 | 40S ribosomal protein S9 | D | + | [176] | |||||
| 4 | RPSA | 40S ribosomal protein SA, LMAR1 | + | [183] | ||||||
| 2 | RRBP1 | Ribosome-binding protein 1 | U | D | D | + | ||||
| 11 | RRP12 | RRP12-like protein | U | + | ||||||
| 3 | RRP9 | U3 small nucleolar RNA-interacting protein 2 (U3–55K) | U | D | N | + | [184] | |||
| 4 | RRS1 | Ribosome biogenesis regulatory protein homolog | U | U | + | |||||
| 5 | RSL1D1 | Ribosomal L1 domain-containing protein 1 | U | D | U | + | ||||
| 2 | RUVBL1 | RuvB-like 1 | + | [185] | ||||||
| 13 | SAP130 | Histone deacetylase complex subunit SAP130 | U | U | + | |||||
| 5 | SERPINB1 | Leukocyte elastase inhibitor | U | + | ||||||
| 4 | SERPINB6 | Serpin B6, peptidase inhibitor 6 | + | |||||||
| 2 | SERPINC1 | Antithrombin-III | U | + | ||||||
| 6 | SET | Protein SET, phosphatase 2A inhibitor | U | D | D | + | ||||
| 14 | SF3B1 | Splicing factor 3B subunit 1 | U | D | D | + | [186] | |||
| 8 | SFN | 14-3-3 protein sigma | U | D | U | + | [187] | |||
| 2 | SLC1A5 | Neutral amino acid transporter B, Simian type D retrovirus receptor, Baboon M7 virus receptor | U | D | U | + | ||||
| 2 | SLC2A1 | HepG2 glucose transporter, human T-cell leukemia virus receptor, GLUT1 | D | D | Nsp8 | + | [188] | |||
| 17 | SLC3A2 | 4F2 cell-surface antigen heavy chain | U | D | U | + | ||||
| 2 | SND1 | Staphylococcal nuclease domain-containing protein 1 | U | D | D | + | ||||
| 15 | SNRNP200 | U5 small nuclear ribonucleoprotein 200 kDa helicase | D | D | + | [189] | ||||
| 3 | SNRPA | U1 small nuclear ribonucleoprotein A | U | + | [190] | |||||
| 2 | SNRPB | Small nuclear ribonucleoprotein-associated proteins B and B’ | U | D | U | D | + | [191] | ||
| 2 | SNRPD1 | Small nuclear ribonucleoprotein Sm D1 | U | U | + | [192] | ||||
| 4 | SNRPD2 | Small nuclear ribonucleoprotein Sm D2 | D | D | + | [193] | ||||
| 2 | SNRPG | Small nuclear ribonucleoprotein G, PBSCG | + | [194] | ||||||
| 2 | SOD1 | Superoxide dismutase [Cu-Zn] | U | D | + | [195] | ||||
| 46 | SPTAN1 | Spectrin alpha chain, brain | U | D | D | + | [196] | |||
| 29 | SPTBN1 | Spectrin beta chain, brain 1 | U | D | U | D | + | [197] | ||
| 2 | SRP72 | Signal recognition particle 72 kDa protein | D | D | + | [198] | ||||
| 3 | SRSF1 | Serine arginine rich splicing factor 1, ASF, SF2 | U | D | D | + | [199] | |||
| 6 | SSB | Lupus La protein | U | D | U | D | + | [10] | ||
| 9 | SSBP1 | Single-stranded DNA-binding protein, mitochondrial | N | + | ||||||
| 8 | SSRP1 | FACT complex subunit SSRP1 | U | D | U | D | + | [200] | ||
| 2 | ST13 | Hsc70-interacting protein | U | U | + | [201] | ||||
| 9 | SUPT16H | FACT complex subunit SPT16 | D | D | + | |||||
| 2 | SUPT5H | Transcription elongation factor SPT5 | + | |||||||
| 2 | SYNCRIP | Heterogeneous nuclear ribonucleoprotein Q | D | D | + | |||||
| 11 | TALDO1 | Transaldolase | U | D | U | D | + | [202] | ||
| 4 | TEX10 | Testis-expressed protein 10 | + | |||||||
| 3 | TFG | TRK-fused gene protein | + | |||||||
| 4 | TGM2 | Protein-glutamine gamma-glutamyltransferase 2 | U | D | U | D | + | [203] | ||
| 4 | TLN1 | Talin-1 | U | D | U | + | [204] | |||
| 3 | TOP1 | DNA topoisomerase 1 | U | + | [205] | |||||
| 5 | TP53I3 | Quinone oxidoreductase | U | D | U | D | + | |||
| 7 | TPM1 | Tropomyosin 1 alpha chain | U | D | U | D | + | [206] | ||
| 2 | TPM2 | Tropomyosin beta chain | U | D | U | D | + | |||
| 4 | TPM3 | Tropomyosin alpha-3 chain | U | D | U | D | + | [207] | ||
| 8 | TPM4 | Tropomyosin alpha-4 chain | U | D | U | + | [208] | |||
| 3 | TSN | Translin | D | D | + | |||||
| 4 | TUBA1C | Tubulin alpha-1C chain | U | D | U | D | + | [209] | ||
| 5 | TUBA4A | Tubulin alpha-4A chain | U | D | D | + | [210] | |||
| 4 | TUBB4B | Tubulin beta-2C chain, TUBB2C | U | D | U | + | [211] | |||
| 2 | TXNDC5 | Thioredoxin domain-containing protein 5 | U | D | U | D | + | |||
| 15 | TXNRD1 | Thioredoxin reductase 1, cytoplasmic | U | D | U | + | [212] | |||
| 7 | UBA1 | Ubiquitin-like modifier-activating enzyme 1 | U | D | U | D | + | [213] | ||
| 3 | UBC | RPS27A; UBB ubiquitin-40S ribosomal protein S27a precursor | U | D | + | [214] | ||||
| 2 | UBTF | Nucleolar transcription factor 1, autoantigen N0R-90 | D | D | + | [215] | ||||
| 2 | UCHL1 | Ubiquitin carboxyl-terminal hydrolase isozyme L1 | U | D | U | D | Orf3 | + | [216] | |
| 5 | UGDH | UDP-glucose 6-dehydrogenase | U | D | U | + | ||||
| 15 | UPF1 | Regulator of nonsense transcripts 1 | D | N | + | |||||
| 2 | USP7 | Ubiquitin carboxyl-terminal hydrolase (Herpes virus associated) | U | U | + | |||||
| 27 | VCL | Vinculin | U | D | U | + | [217] | |||
| 18 | VCP | Transitional endoplasmic reticulum ATPase | U | D | U | D | + | [218] | ||
| 11 | VIM | Vimentin | U | D | U | D | + | [219] | ||
| 5 | WDR18 | WD repeat-containing protein 18 | D | + | ||||||
| 32 | XRCC5 | ATP-dependent DNA helicase 2 subunit 2 | D | D | + | [220] | ||||
| 30 | XRCC6 | ATP-dependent DNA helicase 2 subunit 1 | U | D | U | D | + | [221] | ||
| 3 | YBX1 | Y-box-binding protein 1 | U | D | U | D | + | [222] | ||
| 6 | YBX3 | Y-box binding protein 3, CSDA | U | D | U | D | + | [223] | ||
| 5 | YWHAB | 14-3-3 protein beta/alpha | U | D | D | + | ||||
| 15 | YWHAE | 14-3-3 protein epsilon | U | D | U | D | + | [187] | ||
| 6 | YWHAG | 14-3-3 protein gamma | U | D | + | [187] | ||||
| 3 | YWHAH | 14-3-3 protein eta | D | + | [224] | |||||
| 7 | YWHAQ | 14-3-3 protein theta | U | D | U | + | [183] | |||
| 7 | YWHAZ | 14-3-3 protein zeta/delta | U | D | D | + | [225] | |||
Notes: # Pep.: number of peptides identified by mass spectrometry; COVID (up/down): protein or gene expression up- and/or down-regulated in SARS-Cov-2 infected cells or patients; A549 infection (up/down): protein or gene expression up- and/or down-regulated in SARS-Cov-2 infected A549 cells; Interactome: host protein interacting with SARS-CoV-2 protein; DS-affinity: concentration of NaCl (1.0 M, strong affinity, or 0.5 M, intermediate affinity) at which a DS-binding protein elutes from DS-affinity resin.
The 348 DS-affinity proteins are highly connected (Fig. 1). They exhibit 6271 interactions, whereas a random set of 348 proteins is expected to have 2536 interactions, as revealed by protein-protein interaction STRING analysis [37]. The tight connections suggest that these known and putative autoAgs are originating from common biological pathways or processes. Our analysis shows that they are indeed pre-dominantly associated with translation, mRNA metabolic process, ribonucleoprotein complex biogenesis, vesicle and vesicle-mediated transport, chromosome, and cytoskeleton (Fig. 1).
Fig. 1.

The autoantigenome from A549 cells identified by DS-affinity. Marked proteins are associated with translation (69 proteins, red), mRNA metabolic processing (69 proteins, pink), ribosome biogenesis (43 proteins, blue), vesicles (87 proteins, green) and vesicle-mediated transport (72 proteins, dark green), chromosomes (40 proteins, aqua), and cytoskeleton (65 proteins, gold).
3.2. COVID-altered proteins among the A549 autoantigenome
To find out whether the known and putative autoAgs identified by DS-affinity may play a role in SARS-CoV-2 infection, we compared our A549 autoantigenome with currently available COVID data compiled in the Coronascape database [16–36]. Of our 348 autoantigenome proteins from A549 cells, 291 (83.6%) have been found to be COVID-altered, i.e., up- and/or down-regulated at protein and/or mRNA level in SARS-CoV-2 infected cells or patient tissues (Table 1). Because the COVID data have been generated from various research labs using different techniques and sources of cells or tissues, 190 proteins are found to be up in some studies but down in others. In total, 231 proteins are found up-regulated, and 252 are found down-regulated in SARS-CoV-2 infection. Based on reported autoantibodies, 191 (65.6%) COVID-altered proteins are confirmed autoAgs (Table 1).
Based on gene ontology (GO) cellular component analysis, proteins of the A549 DS-affinity autoantigenome that are also altered in COVID infection can be located to membrane-bound organelles (247 proteins), nucleus (177 proteins), ribonucleoprotein complex (95 proteins), mitochondrion (46 proteins), endoplasmic reticulum (45 proteins), secretory granules (41 proteins), melanosome (27 proteins), myelin sheath (28 proteins), and axon (16 proteins).
Within the total A549 autoantigenome, the 291 COVID-altered proteins form a tightly interacting network (Fig. 2). At high STRING protein-protein interaction confidence level, these proteins exhibit 2249 interactions, whereas 953 interactions would be expected of a random collection of proteins of the same size. By GO biological process analysis, the COVID-altered proteins are significantly enriched in various biological processes, including translation, peptide biosynthetic process, RNA catabolic process, nucleobase-containing compound catabolic process, SRP-dependent cotranslational protein targeting to membrane, protein localization to organelle, and symbiont process. Among these processes associated with COVID-altered proteins, the hierarchical cluster tree root points to ribonucleoprotein complex biogenesis (Fig. 3).
Fig. 2.
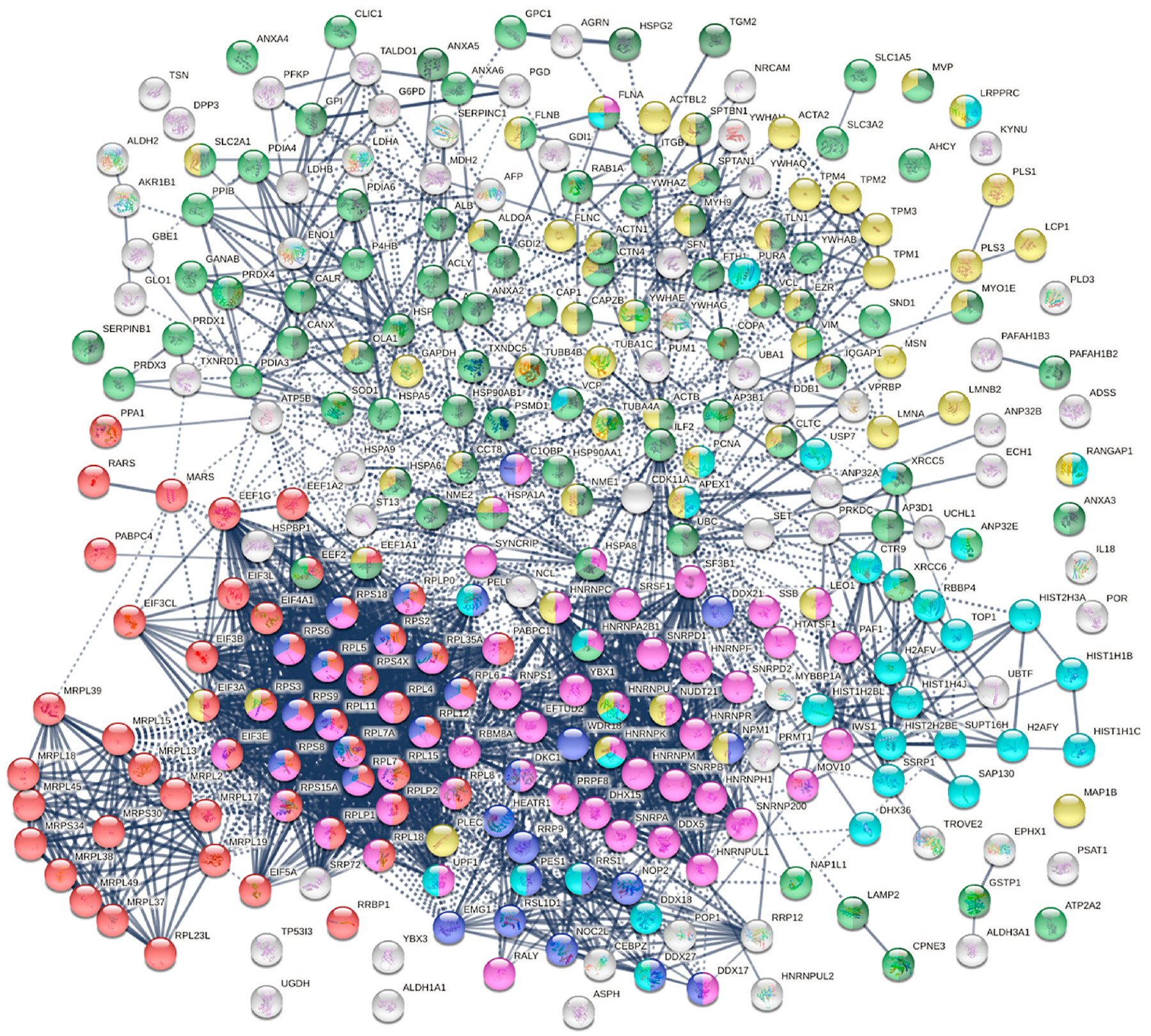
COVID-altered proteins shared with the A549 autoantigenome. Marked proteins are associated with translation (53 proteins, red), mRNA metabolic process (63 proteins, pink), vesicle (79 proteins, green) and vesicle-mediated transport (64 proteins, dark green), cytoskeleton (58 proteins, gold), chromosomes (35 proteins, aqua), and ribosome biogenesis (29 proteins, blue).
Fig. 3.
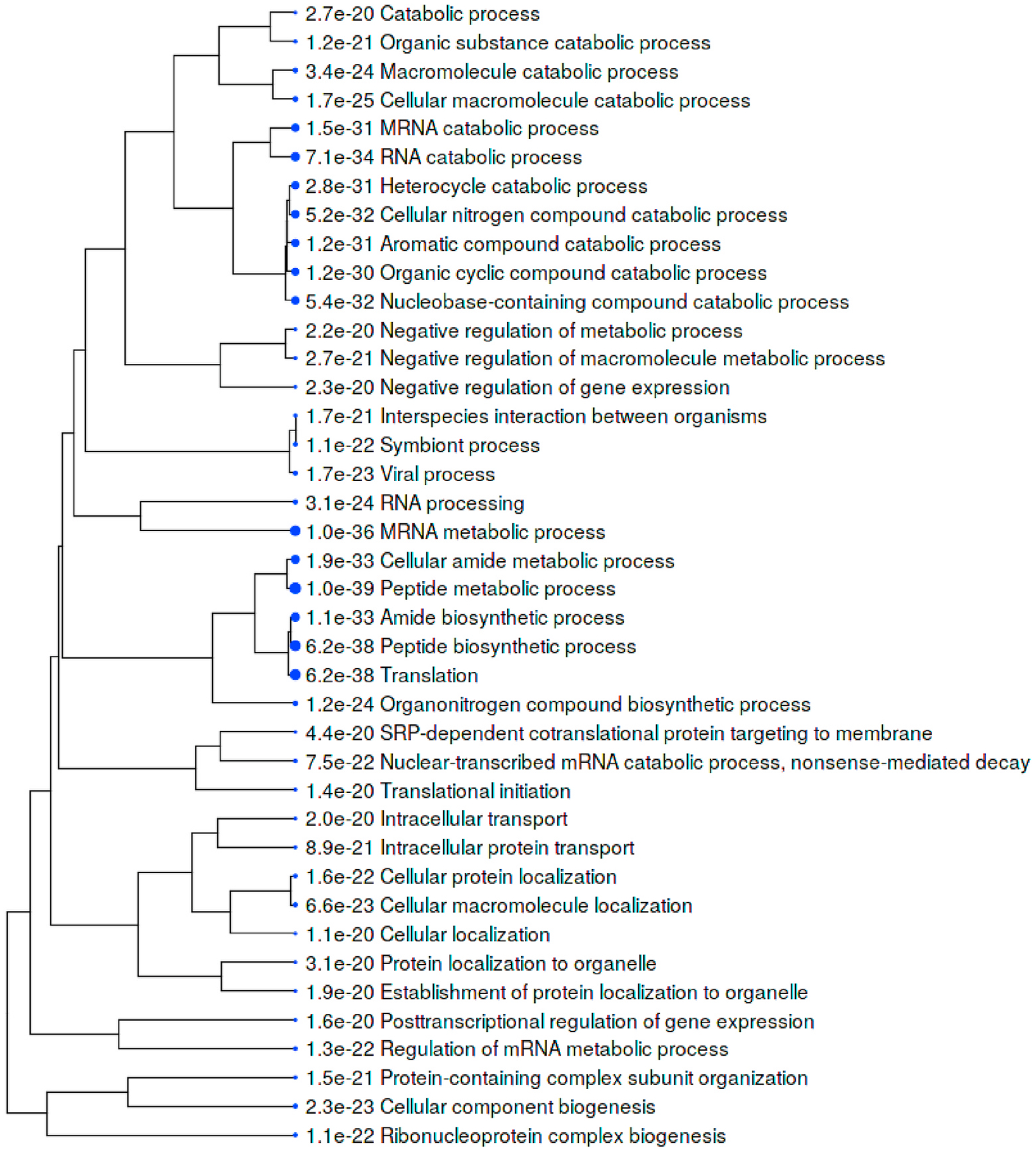
Top 40 enriched GO biological processes among COVID-altered proteins shared with the A549 autoantigenome. Bigger dots indicate more significant p-values.
Combined pathway and process enrichment analyses also show that the COVID-altered DS-affinity proteins are most significantly related to peptide biosynthetic process, metabolism of RNA, and ribonucleoprotein complex biogenesis (Fig. 4). The up-regulated autoAgs are more related to Nop56p-associated pre-rRNA complex, RNA catabolic process, and vesicle-mediated transport, whereas the down-regulated autoAgs are more related to translation and ribosome biogenesis. COVID-altered autoAgs are also significantly associated with regulated exocytosis, platelet degranulation, smooth muscle contraction, and IL-12 signaling.
Fig. 4.
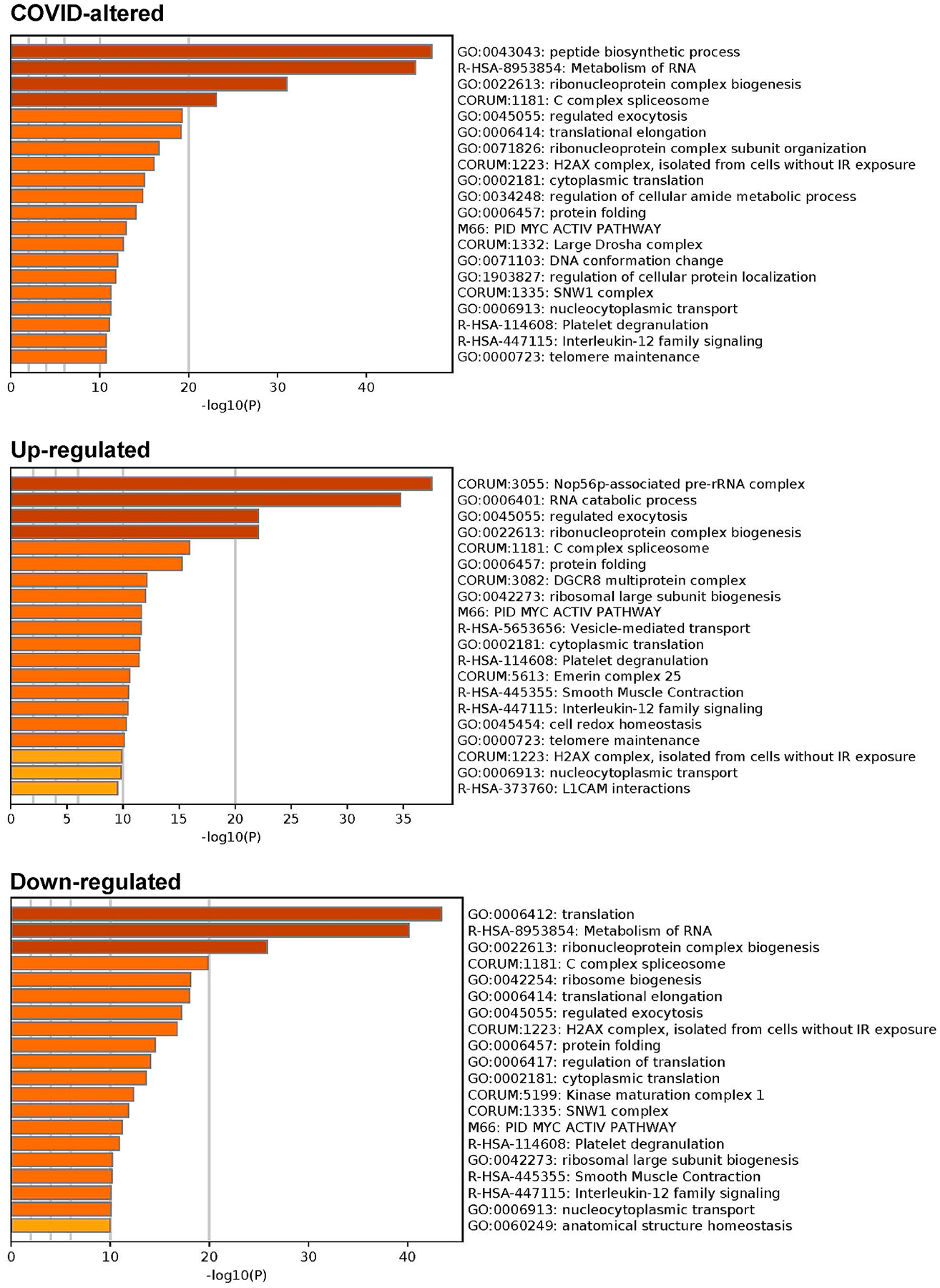
Top 20 enriched pathways and processes among COVID-altered autoAgs. Top: 298 COVID-altered autoAgs. Middle: 231 up-regulated autoAgs in COVID. Bottom: 252 down-regulated autoAgs in COVID.
The molecular functions of the COVID-altered DS-affinity autoAgs include RNA binding (76 proteins), hydrolase activity (58 proteins), purine ribonucleotide triphosphate binding (55 proteins), pyrophosphatase activity (35 proteins), nucleoside-triphosphatase activity (34 proteins), oxidoreductase activity (26 proteins), ATPase activity (25 proteins), ubiquitin protein ligase binding (21 proteins), mRNA 3′-UTR binding (11 proteins) and 5′-UTR binding (3 proteins), and helicase activity (13 proteins).
3.3. AutoAgs that interact with SARS-CoV-2 proteins
To study the origins of COVID-induced autoAg alterations, we examined DS-affinity proteins that are involved in the interactomes of SARS-CoV-2 viral proteins [18,29,33] and found 38 DS-affinity proteins that interact directly with different viral proteins, with 25 of them being known autoAgs, e.g., ATP5B, CANX, DDX21, EEF1A2, PDIA3, and SLC2A1 (Table 1). Orf3 protein is the most striking, as its interactome includes 14 DS-affinity proteins and 9 known autoAgs. N protein interacts with 9 DS-affinity proteins and 6 known autoAgs, including 2 helicases (DDS21 and MOV10), 2 poly(A)-binding proteins (PABPC1 and PABPC4). Nsp4 interact with 6 DS-affinity proteins and 5 known autoAgs. Nsp4 interacts with IDE (insulin degrading enzyme), which is not a known autoAg.
A few host DS-affinity proteins interact with more than one SARS-CoV-2 protein. RAB1A (involved in intracellular membrane trafficking) interacts with 3 viral proteins (Orf3, Orf7b, and Nsp7), and PLD3 (phospholipase) also interacts with 3 viral proteins (Nsp2, Orf7b, and Orf8), but neither RAB1A nor PLD3 has been discovered as autoAgs. HSPA5 (GRP78/BiP) interacts with Nsp2 and Nsp4, HSPA1A interacts with N and Orf9b, and both heat shock proteins are known autoAg. PRKDC, a known autoAg, interacts with M and Nsp. Interestingly, of the ezrin-radixin-moesin protein family that connects the actin cytoskeleton to the plasma membrane, RDX is found to interact with Orf3 and Nsp13, MSN interacts with Orf3 and Nsp6, and EZR is COVID-altered. Furthermore, RDX, MSN, and EZR are all known autoAgs (Table 1).
3.4. AutoAgs from perturbation by viral protein expression
To find out how individual SARS-CoV-2 proteins affect the host, Stukalov et al. conducted extensive proteomic analysis of A549 cells transduced to express individual SARS-CoV-2 proteins [33]. By comparing with their data, we identified 167 DS-affinity proteins that are perturbated by viral protein expression in A549 cells. Among all SARS-CoV-2 proteins, Orf3 expression produced the largest number of potential autoAgs, with 26 up- and 36 down-regulated being DS-affinity proteins (Fig. 5, Supplemental Table 1). Other viral protein expressions affected various numbers of DS-affinity proteins, including E (20 up and 18 down), Orf9b (6 up and 16 down), M (10 up and 11 down), N (10 up and 8 down), Nsp13 (5 up and 11 down), and Nsp12 (3 up and 12 down). Interestingly, S expression yielded only 3 up- and 2 down-regulated DS-affinity proteins, suggesting that it may not have much intracellular activity.
Fig. 5.

DS-affinity proteins that interact with SARS-CoV-2 viral proteins or are perturbed in A549 cells expressing individual viral proteins. Orf3: regulation of gene expression (pink), cytoplasmic vesicle (green), monosaccharide biosynthetic process (blue), response to stress (brown), and hydrolase activity (gold). Orf9b: mitochondrial translation (red), mitochondrion localization (pink), autophagosome maturation (gold). E: establishment of protein localization to membrane (red), translation initiation (pink), mRNA splicing (green), regulation of macroautophagy (brown), post-translational protein modification (gold), RNA polymerase II transcription (blue). M: establishment of protein localization to membrane (red), intracellular protein transport (gold), organelle organization (green), nicotinamide nucleotide metabolic process (aqua), regulation of apoptotic process (blue), and symbiont process (pink). N: maintenance of location in cell (pink), protein localization to ER (red), protein folding (aqua), mitochondrial nucleoid (amber), RNA polymerase II transcription (blue), and RNA-binding (green). Nsp1: protein localization (green), gene expression (red), protein processing in ER (blue), and phagosome (gold). Nsp13: cytoskeleton (gold), regulation of muscle contraction (green), and melanosome (brown).
In total, Orf3 affected 71 DS-affinity proteins identified from A549 cells, which includes those directly interacting with Orf3 and those perturbed by Orf3 protein expression in A549 cells. The large number of Orf3-affected host proteins implicates important roles of Orf3 in SARS-CoV-2 infection. Network analysis reveals these proteins to be mostly associated with gene expression regulation, cytoplasmic vesicles, apoptosis, response to stress, monosaccharide biosynthesis, or hydrolase activity (Fig. 5). Several of these are classical nuclear autoAgs, e.g., PNCA, SSB (Lupus La), XRCC5 (Lupus Ku80, thyroid-lupus autoAg), XRCC6 (Ku70), and SNRPB (SmB/B’). A few are unknown autoAgs but with important relevance to COVID, e.g., PAFAH1B2 and PAFAH1B3 (the alpha catalytic subunits of the cytosolic type I platelet-activating factor (PAF) acetylhydrolase). PAF is produced by a variety of cells involved in host defense, and PAF signaling can trigger inflammatory and thrombotic cascades. The modulation of PAF by SARS-CoV-2 Orf3 may partially explain the frequently occurring thrombotic complications and coagulopathy in COVID-19 patients. PAF also induces apoptosis in a PAF receptor independent pathway that can be inhibited by PAFAH1B2 and PAFAH1B3 [39].
SARS-CoV-2 E protein affects a number of ribonucleoproteins that are related to translation initiation and mRNA splicing, e.g., hnRNP (U and UL1) and ribosomal protein (L7, L8 L11, L12, L35A). E-affected proteins are associated with establishment of protein localization to membrane, regulation of autophagy, and post-translational protein modification (Fig. 5). SARS-CoV-2 M, Nsp1, and N proteins also affect various ribonucleoproteins, whereas Nsp13 appears to affect proteins associated with the cytoskeleton. Overall, the majority of DS-affinity proteins found affected by individual SARS-CoV-2 proteins are known autoAgs (Fig. 5 and Table 1), which indicates that host proteins perturbed by viral components are an important source of autoAgs.
3.5. Mitochondrial perturbation by SARS-CoV-2 Orf9b
By DS-affinity fractionation, we identified 22 mitochondrial ribosomal proteins from A549 cells with strong DS-affinity, including mitochondrial 39S ribosomal proteins (L1, L2, L13, L15, L17, L18, L19, L23, L37, L38, L39, L45, L49) and 28S ribosomal proteins (S9, S22, S23, S27, S28, S29, S30, S34, S39) (Table 1). Remarkably, 14 mitochondrial ribosomal proteins are found down-regulated in SARS-CoV-2 infection, with 2 (L15 and L37) reported both down- and up-regulated. Moreover, MRPS27 is found in the interactome of SAS-Cov-2 Nsp8. Most strikingly, Orf9b-expression in A549 cells caused down-regulation of 16 proteins that we identified by DS-affinity, namely, ALB, ANXA4, LAMP2, MRPL2, MRPL13, MRPL15, MRPL17, MRPL18, MRPL19, MRPS30, MRPL37, MRPL38, MRPL39, MRPL45, MRPL49, and UCHL1 (Fig. 5). Eleven of the Orf9-affected proteins are mitochondrial ribosomal proteins, which may affect the mitochondrial translation machinery. Orf9-affected proteins may also be involved in mitochondrion localization, autophagosome maturation, or other processes. Overall, these findings suggest that SARS-CoV-2 infection may affect mitochondria primarily through Orf9b.
Orf9b of SARS-CoV has been shown to localize to mitochondria, trigger ubiquitination and proteasomal degradation of dynamin-like protein 1, limit host cell interferon signaling by targeting mitochondrial associated adaptor molecule MAVS signalosome, and manipulate the mitochondrial function to help evade host innate immunity [40]. Orf9b of SARS-CoV-2 has been reported to suppress the type I interferon response by targeting TOM70 [41]. In COVID-19 pneumonia patients, monocytes show altered bioenergetics and mitochondrial dysfunction with depolarized and abnormal ultrastructure [42].
Currently, little is known about the involvement of mitochondrial ribosomes or mitochondrial translation in SARS-CoV-2 infection. Expression of mitochondrial ribosomal proteins associated with protein synthesis has been found to be the most striking transcriptional difference among dengue virus-infected children, as revealed by a genome-wide microarray analysis of whole blood RNA from 34 infected children collected on days 3–6 of illness [43]. In human cytomegalovirus infection, proteins involved in biogenesis of the mitochondrial ribosome changed early during the viral replication cycle [44]. Mitochondria are vital to cell survival and apoptosis as they produce the majority of adenosine triphosphate (ATP) that provide chemical energy to cells. Especially for cells such as muscles that require much ATP, mitochondrial dysfunction will certainly lead to problems such as muscle weakness and fatigue. The roles of mitochondrial ribosomal proteins play in COVID and long-term sequelae merit further investigation.
3.6. AutoAgs related to ubiquitination alteration in SARS-CoV-2 infection
Ubiquitination provides a universal signal for protein degradation. By comparing our data with the ubiquitinome of SARS-CoV-2 infected cells, we identified 102 DS-affinity proteins that are altered by ubiquitination during viral infection (Supplemental Table 1). These ubiquitination-altered proteins are significantly associated with gene expression, catabolic process, regulation of apoptotic process, cytoplasmic vesicles, and cytoskeleton (Fig. 6). They include 15 ribosomal proteins, 8 heat shock proteins, 5 hnRNP proteins, 5 histones, 4 translation elongation factors, and 3 translation initiation factors, and a majority of them are known autoAgs (Table 1).
Fig. 6.

Known and putative autoAgs derived from ubiquitination alteration in SARS-CoV-2 infected A549 cells. Marked proteins are associated with catabolic process (37 proteins, blue), ubiquitin protein ligase binding (12 proteins, aqua), gene expression (46 proteins, pink), regulation of apoptotic process (22 proteins, brown), cytoplasmic vesicles (28 proteins, green), and cytoskeleton (26 proteins, gold).
Three ubiquitination/de-ubiquitination enzymes (UBA1, UCHL1, and USP7) are COVID-altered and possess DS-affinity, with UBA1 and UCHL1 being known autoAgs. UBA1 catalyzes the first step in ubiquitin conjugation to mark proteins for degradation through the ubiquitin-proteasome system. USP7 is a hydrolase that deubiquitinates target proteins. UCHL1 is a thiol protease that recognizes and hydrolyzes a peptide bond at the C-terminal glycine of ubiquitin, and is involved in the processing of ubiquitin precursors and of ubiquitinated proteins. UBA1 is found down-regulated by Orf3 expression. UCHL1 is found in the Orf3 interactome, up-regulated by SARS-CoV-2 E protein expression, and down-regulated by Nsp12, Nsp8, Orf8, and Orf9b (Supplemental Table 1).
Ten COVID-altered DS-affinity proteins were identified with ubiquitin protein ligase-binding activity, including HSPA1A, HSPA5, HSPA8, HSP90AA1, HSP90AB1, RPL11, VCP, VCL, YWHAE, and YWHAZ. HSPA1A interacts with SARS-CoV-2 Orf9b and N proteins, HSPA5 (GRP78/BiP) interacts with Nsp2 and Nsp4, and HSPA8 interacts with Nsp2. Except for RPL11, all are known autoAgs (Table 1).
In addition, SARS-CoV-2 and other coronaviruses encode for papain-like proteases (PLP), an important multifunctional enzyme with de-ubiquitination, de-ISGlation, and interferon antagonism activities [45]. PLPs, along with other proteases, are responsible for processing replicase proteins that are required from viral replication. PLP of SARS-CoV-2 is able to reverse host ubiquitination and remove interferon-stimulated gene product 15 (ISG15), and its substrate activity mirrors closely that of PLP of MERS [46].
Ubiquitin modifications can regulate innate immune response and apoptosis, and ISG15 is a ubiquitin-like modifier typically expressed during host cell immune response. Overall, various components of SARS-CoV-2 appear to be able to alter uniquitination of host proteins. The large pool of ubiquitin-altered proteins in SARS-CoV-2 infection indicates that ubiquitin modification, such as differential abundance and dynamic ubiquitination pattern change, may be a major origin of autoAgs.
3.7. AutoAgs related to phosphorylation alteration in SARS-CoV-2 infection
Comparing our data with currently available phosphoproteome data of COVID-19 [25,33], 97 phosphoproteins are found with both DS-affinity and COVID-induced alteration, with a majority (52/97) related to gene expression (Fig. 7). Notably, they include 8 heterogeneous nuclear ribonucleoproteins affected by phosphorylation (HNRNPA2B1, HNRNPC, HNRNPH1, HNRNPK, HNRNPM, HNRNPU, HNRNPUL1, HNRNPUL2), with 4 being known autoAgs (Table 1). HNRNPs are involved in many cellular processes, including gene transcription, cell cycle, DNA damage control, post-transcriptional modification of newly synthesized pre-mRNA, and virus replication. For example, HNRNPA2B1 displays RNA-binding affinity to murine hepatitis coronavirus via binding the viral RNA at the 3’ end and modulate viral RNA synthesis [47].
Fig. 7.
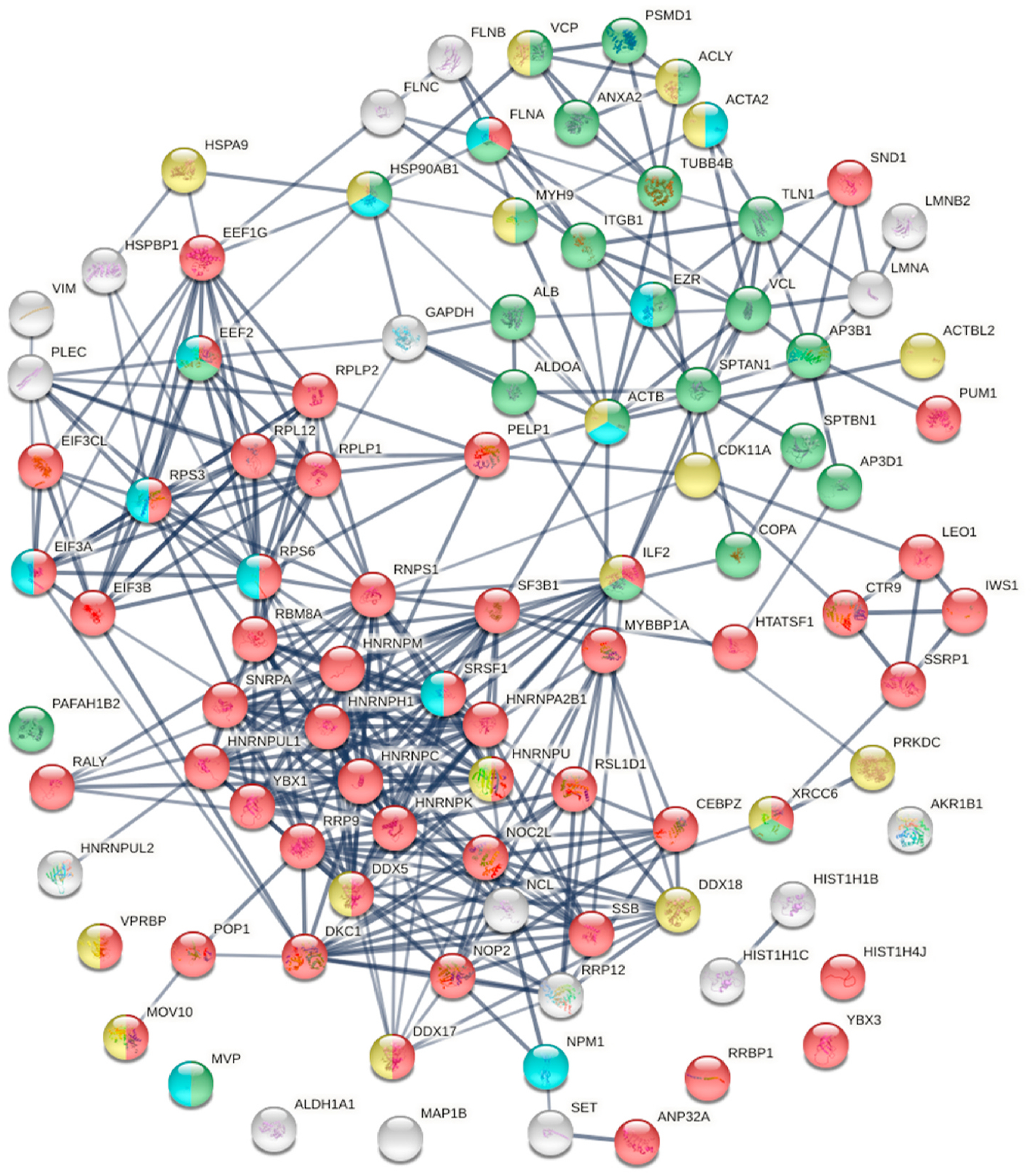
Known and putative autoAgs derived from phosphorylation alteration in SARS-CoV-2 infected cells. Marked proteins are associated with gene expression (52 proteins, red), vesicle-mediated transport (25 proteins, green), ATP binding (18 proteins, gold), and kinase binding (12 proteins, aqua).
There are 25 phosphorylation-altered proteins that are related to vesicle-mediated transport, most of which are known autoAgs, including ACLY, ACTA2, ACTB, ALB, ALDO, ANXA2, FLNA, COPA, SPTAN1, SPTBN1, TLN1, TUBB4, and VCL (Fig. 7 and Table 1). The coatomer, to which COPA (coatomer subunit alpha) belongs, is a cytosolic protein complex that associates with Golgi non-clathrin-coated vesicles and is required for budding from the Golgi membrane. COPA is associated with autoimmune interstitial lung, joint, and kidney disease [48].
There are 18 phosphorylation-altered potential autoAgs with ATP binding activity, and 12 with kinase binding activity (Fig. 7). In particular, PRKDC (DNA-dependent protein kinase catalytic subunit) is identified with strong DS-affinity and is a known autoAg. It is a serine/ threonine-protein kinase that acts as a molecular sensor for DNA damage, with involvement in numerous biological processes such as DNA damage and repair, immunity, innate immunity, ribosome biogenesis, and apoptosis. PRDKC is found in the interactomes of M and Nsp4 proteins of SARS-CoV-2 and up-regulated by expression of Nsp10, Nsp9, Orf7a, or Orf7b protein in A549 cells [18,33]. PRKDC is also found up-regulated at 0 h and 4 h in SARS-CoV-2 infected Vero E6 cells [25] and up-regulated at 24 h in SARS-CoV-2 infected Caco-2 cells [19]. These findings suggest that phosphorylation by PRKDC plays extensive and important roles in COVID.
Proteins phosphorylated during apoptosis are common targets of autoantibodies. For example, the U1–70 snRNP autoAg undergoes specific changes in the phosphorylation/dephosphorylation balance and cellular localization during apoptosis [49], and phosphorylated U1-snRNP complex induced by apoptosis is recognized by autoantibodies in patients with systemic lupus erythematosus [50]. A high degree of phosphorylation of SSB (lupus La autoAg) substantially diminished its poly(U) binding capacity, but its binding to human autoantibodies increased 2-fold with increased phosphorylation [51]. On the other hand, SSB autoAg has also been reported to be dephosphorylated and cleaved during early apoptosis [52]. During apoptosis, ribosomal protein P1 and P2 autoAgs are completely dephosphorylated while P0 autoAg is partially dephosphorylated [53]. Therefore, alterations in phosphorylation, either hyper- or hypo-phosphorylation, may lead to changes in self-molecules and render them autoantigenic.
4. Conclusion
In our quest for a comprehensive autoantigen atlas for COVID-19, we report an autoantigen profile of 191 confirmed autoAgs and 100 putative autoAgs in SARS-CoV-2 infection. These proteins are initially identified from human lung epithelial A549 cells using a unique DS-affinity autoAg enrichment strategy, and then compared with currently available COVID-omics data. Our study reveals that cellular processes and components integral to viral infection are major origins of autoAgs, including gene expression, ribonucleoprotein biogenesis, translation and mitochondrial translation, vesicle and vesicle-mediated transport, and cytoskeleton. Ubiquitination and phosphorylation are particular post-translational modifications that cause changes in self-molecules and render them autoantigenic. Impaired clearance of apoptotic and dead cell material is considered a major pathogenic attribute of autoimmune disease. We have previously shown that DS possesses unique affinity to apoptotic cells and their released autoAgs, and our current study further demonstrates that ubiquitination and phosphorylation associated with apoptosis are possibly major sources of molecular alterations in self-molecule to autoantigen transformation. Overall, our study demonstrates that SARS-CoV-2 causes extensive alterations of host cellular proteins and produces a large number of potential autoAgs, indicating that there may be an intimate relationship between COVID infection and autoimmunity. Our data provide a deep and comprehensive atlas of autoantigens related to COVID and provide a powerful resource for ongoing studies into the pathophysiology and mechanisms of long-term autoimmune sequelae after acute COVID.
Supplementary Material
Acknowledgements
We thank Jung-hyun Rho for technical assistance with experiments. We thank Ross Tomaino and the Taplin Biological Mass Spectrometry facility of Harvard Medical School for expert service with protein sequencing.
Funding statement
This work was partially supported by Curandis, the US NIH, and a Cycle for Survival Innovation Grant (to MHR). MHR acknowledges the NIH/NCI R21 CA251992 and MSKCC Cancer Center Support Grant P30 CA008748. The funding bodies were not involved in the design of the study and the collection, analysis, and interpretation of data.
Declaration of competing interest
JYW is the founder and Chief Scientific Officer of Curandis. WZ was supported by the NIH and declares no competing interests. MWR and VBR are volunteers of Curandis. MHR is a member of the Scientific Advisory Boards of Trans-Hit, Proscia, and Universal DX, but these companies have no relation to the study.
Footnotes
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.jaut.2021.102644.
References
- [1].Wang JY, Zhang W, Roehrl MW, Roehrl VB, Roehrl MH, An Autoantigen Atlas from Human Lung HFL1 Cells Offers Clues to Neurological and Diverse Autoimmune Manifestations of COVID-19, bioRxiv, 2021, 20210124427965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Wang JY, Lee J, Yan M, Rho JH, Roehrl MH, Dermatan sulfate interacts with dead cells and regulates CD5(+) B-cell fate: implications for a key role in autoimmunity, Am. J. Pathol 178 (2011) 2168–2176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Rho JH, Zhang W, Murali M, Roehrl MH, Wang JY, Human proteins with affinity for dermatan sulfate have the propensity to become autoantigens, Am. J. Pathol 178 (2011) 2177–2190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Lee J, Rho J.-h., Roehrl MH, Wang JY, Dermatan Sulfate Is a Potential Master Regulator of IgH via Interactions with Pre-BCR, GTF2I, and BiP ER Complex in Pre-B Lymphoblasts, 2021. bioRxiv 20210118427153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Wang JY, Zhang W, Rho JH, Roehrl MW, Roehrl MH, A proteomic repertoire of autoantigens identified from the classic autoantibody clinical test substrate HEp-2 cells, Clin. Proteonomics 17 (2020) 35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Zhang W, Rho JH, Roehrl MH, Wang JY, A comprehensive autoantigen-ome of autoimmune liver diseases identified from dermatan sulfate affinity enrichment of liver tissue proteins, BMC Immunol 20 (2019) 21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Zhang W, Rho JH, Roehrl MW, Roehrl MH, Wang JY, A repertoire of 124 potential autoantigens for autoimmune kidney diseases identified by dermatan sulfate affinity enrichment of kidney tissue proteins, PloS One 14 (2019), e0219018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Zhou S, Jones-Lopez EC, Soneji DJ, Azevedo CJ, Patel VR, Myelin oligodendrocyte glycoprotein antibody-associated optic neuritis and myelitis in COVID-19, J. Neuro Ophthalmol. : the official journal of the North American Neuro-Ophthalmology Society 40 (2020) 398–402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Bowles L, Platton S, Yartey N, Dave M, Lee K, Hart DP, et al. , Lupus anticoagulant and abnormal coagulation tests in patients with covid-19, N. Engl. J. Med 383 (2020) 288–290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Gruber CN, Patel RS, Trachtman R, Lepow L, Amanat F, Krammer F, et al. , Mapping systemic inflammation and antibody responses in multisystem inflammatory syndrome in children (MIS-C), Cell 183 (2020) 982–995, e14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Gagiannis D, Steinestel J, Hackenbroch C, Schreiner B, Hannemann M, Bloch W, et al. , Clinical, serological, and histopathological similarities between severe COVID-19 and acute exacerbation of connective tissue disease-associated interstitial lung disease (CTD-ILD), Front. Immunol 11 (2020) 587517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Lerma LA, Chaudhary A, Bryan A, Morishima C, Wener MH, Fink SL, Prevalence of autoantibody responses in acute coronavirus disease 2019 (COVID-19), Journal of translational autoimmunity 3 (2020) 100073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Fujii H, Tsuji T, Yuba T, Tanaka S, Suga Y, Matsuyama A, et al. , High levels of anti-SSA/Ro antibodies in COVID-19 patients with severe respiratory failure: a case-based review : high levels of anti-SSA/Ro antibodies in COVID-19, Clin. Rheumatol 39 (2020) 3171–3175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Dragonetti D, Guarini G, Pizzuti M, Detection of anti-heparin-PF4 complex antibodies in COVID-19 patients on heparin therapy, Blood transfusion = Trasfusione del sangue 18 (2020) 328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Bastard P, Rosen LB, Zhang Q, Michailidis E, Hoffmann HH, Zhang Y, et al. , Autoantibodies against Type I IFNs in Patients with Life-Threatening COVID-19, Science, New York, NY), 2020, p. 370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Zhou Y, Zhou B, Pache L, Chang M, Khodabakhshi AH, Tanaseichuk O, et al. , Metascape provides a biologist-oriented resource for the analysis of systems-level datasets, Nat. Commun 10 (2019) 1523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Zhang JY, Wang XM, Xing X, Xu Z, Zhang C, Song JW, et al. , Single-cell landscape of immunological responses in patients with COVID-19, Nat. Immunol 21 (2020) 1107–1118. [DOI] [PubMed] [Google Scholar]
- [18].Davies JP, Almasy KM, McDonald EF, Plate L, Comparative multiplexed interactomics of SARS-CoV-2 and homologous coronavirus nonstructural proteins identifies unique and shared host-cell dependencies, ACS Infect. Dis 6 (2020) 3174–3189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Klann K, Bojkova D, Tascher G, Ciesek S, Münch C, Cinatl J, Growth factor receptor signaling inhibition prevents SARS-CoV-2 replication, Mol. Cell 80 (2020), 164–74.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Sun J, Ye F, Wu A, Yang R, Pan M, Sheng J, et al. , Comparative transcriptome analysis reveals the intensive early stage responses of host cells to SARS-CoV-2 infection, Front. Microbiol 11 (2020) 593857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Bojkova D, Klann K, Koch B, Widera M, Krause D, Ciesek S, et al. , Proteomics of SARS-CoV-2-infected host cells reveals therapy targets, Nature 583 (2020) 469–472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Wilk AJ, Rustagi A, Zhao NQ, Roque J, Martínez-Colón GJ, McKechnie JL, et al. , A single-cell atlas of the peripheral immune response in patients with severe COVID-19, Nat. Med 26 (2020) 1070–1076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Lieberman NAP, Peddu V, Xie H, Shrestha L, Huang ML, Mears MC, et al. , In vivo antiviral host transcriptional response to SARS-CoV-2 by viral load, sex, and age, PLoS Biol 18 (2020), e3000849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Riva L, Yuan S, Yin X, Martin-Sancho L, Matsunaga N, Pache L, et al. , Discovery of SARS-CoV-2 antiviral drugs through large-scale compound repurposing, Nature 586 (2020) 113–119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Bouhaddou M, Memon D, Meyer B, White KM, Rezelj VV, Correa Marrero M, et al. , The global phosphorylation landscape of SARS-CoV-2 infection, Cell 182 (2020) 685–712, e19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Blanco-Melo D, Nilsson-Payant BE, Liu WC, Uhl S, Hoagland D, Møller R, et al. , Imbalanced host response to SARS-CoV-2 drives development of COVID-19, Cell 181 (2020), 1036–45.e9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Shen B, Yi X, Sun Y, Bi X, Du J, Zhang C, et al. , Proteomic and metabolomic characterization of COVID-19 patient sera, Cell 182 (2020) 59–72, e15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Lamers MM, Beumer J, van der Vaart J, Knoops K, Puschhof J, Breugem TI, et al. , SARS-CoV-2 Productively Infects Human Gut Enterocytes, vol. 369, Science, New York, NY: ), 2020, pp. 50–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Gordon DE, Jang GM, Bouhaddou M, Xu J, Obernier K, White KM, et al. , A SARS-CoV-2 protein interaction map reveals targets for drug repurposing, Nature 583 (2020) 459–468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Xiong Y, Liu Y, Cao L, Wang D, Guo M, Jiang A, et al. , Transcriptomic characteristics of bronchoalveolar lavage fluid and peripheral blood mononuclear cells in COVID-19 patients, Emerg. Microb. Infect 9 (2020) 761–770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Vanderheiden A, Ralfs P, Chirkova T, Upadhyay AA, Zimmerman MG, Bedoya S, et al. , Type I and type III interferons restrict SARS-CoV-2 infection of human airway epithelial cultures, J. Virol (2020) 94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Appelberg S, Gupta S, Svensson Akusjärvi S, Ambikan AT, Mikaeloff F, Saccon E, et al. , Dysregulation in Akt/mTOR/HIF-1 signaling identified by proteo-transcriptomics of SARS-CoV-2 infected cells, Emerg. Microb. Infect 9 (2020) 1748–1760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Stukalov A, Girault V, Grass V, Bergant V, Karayel O, Urban C, et al. , Multi-level proteomics reveals host-perturbation strategies of SARS-CoV-2 and SARS-CoV, bioRxiv : the preprint server for biology (2020) 2020, 06.17.156455. [Google Scholar]
- [34].Emanuel W, Kirstin M, Vedran F, Asija D, Theresa GL, Roberto A, et al. , Bulk and single-cell gene expression profiling of SARS-CoV-2 infected human cell lines identifies molecular targets for therapeutic intervention, bioRxiv : the preprint server for biology (2020) 2020, 05.05.079194. [Google Scholar]
- [35].Li Y, Wang Y, Liu H, Sun W, Ding B, Zhao Y, et al. , Urine proteome of COVID-19 patients, medRxiv (2020) 2020, 05.02.20088666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Liao M, Liu Y, Yuan J, Wen Y, Xu G, Zhao J, et al. , Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19, Nat. Med 26 (2020) 842–844. [DOI] [PubMed] [Google Scholar]
- [37].Szklarczyk D, Gable AL, Lyon D, Junge A, Wyder S, Huerta-Cepas J, et al. , STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets, Nucleic Acids Res 47 (2019) D607–D613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Ge SX, Jung D, Yao R, ShinyGO: a graphical gene-set enrichment tool for animals and plants, Bioinformatics 36 (2020) 2628–2629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Bonin F, Ryan SD, Migahed L, Mo F, Lallier J, Franks DJ, et al. , Anti-apoptotic actions of the platelet-activating factor acetylhydrolase I alpha2 catalytic subunit, J. Biol. Chem 279 (2004) 52425–52436. [DOI] [PubMed] [Google Scholar]
- [40].Shi CS, Qi HY, Boularan C, Huang NN, Abu-Asab M, Shelhamer JH, et al. , SARS-coronavirus open reading frame-9b suppresses innate immunity by targeting mitochondria and the MAVS/TRAF3/TRAF6 signalosome, J. Immunol 193 (2014) 3080–3089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Jiang HW, Zhang HN, Meng QF, Xie J, Li Y, Chen H, et al. , SARS-CoV-2 Orf9b suppresses type I interferon responses by targeting TOM70, Cell. Mol. Immunol 17 (2020) 998–1000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Gibellini L, De Biasi S, Paolini A, Borella R, Boraldi F, Mattioli M, et al. , Altered bioenergetics and mitochondrial dysfunction of monocytes in patients with COVID-19 pneumonia, EMBO Mol. Med 12 (2020), e13001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Loke P, Hammond SN, Leung JM, Kim CC, Batra S, Rocha C, et al. , Gene expression patterns of dengue virus-infected children from Nicaragua reveal a distinct signature of increased metabolism, PLoS Neglected Trop. Dis 4 (2010), e710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Karniely S, Weekes MP, Antrobus R, Rorbach J, van Haute L, Umrania Y, et al. , Human cytomegalovirus infection upregulates the mitochondrial transcription and translation machineries, mBio 7 (2016), e00029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Báez-Santos YM, Mielech AM, Deng X, Baker S, Mesecar AD, Catalytic function and substrate specificity of the papain-like protease domain of nsp3 from the Middle East respiratory syndrome coronavirus, J. Virol 88 (2014) 12511–12527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Freitas BT, Durie IA, Murray J, Longo JE, Miller HC, Crich D, et al. , Characterization and noncovalent inhibition of the deubiquitinase and deISGylase activity of SARS-CoV-2 papain-like protease, ACS Infect. Dis 6 (2020) 2099–2109. [DOI] [PubMed] [Google Scholar]
- [47].Shi ST, Yu GY, Lai MM, Multiple type A/B heterogeneous nuclear ribonucleoproteins (hnRNPs) can replace hnRNP A1 in mouse hepatitis virus RNA synthesis, J. Virol 77 (2003) 10584–10593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Watkin LB, Jessen B, Wiszniewski W, Vece TJ, Jan M, Sha Y, et al. , COPA mutations impair ER-Golgi transport and cause hereditary autoimmune-mediated lung disease and arthritis, Nat. Genet 47 (2015) 654–660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Dieker J, Cisterna B, Monneaux F, Decossas M, van der Vlag J, Biggiogera M, et al. , Apoptosis-linked changes in the phosphorylation status and subcellular localization of the spliceosomal autoantigen U1–70K, Cell Death Differ 15 (2008) 793–804. [DOI] [PubMed] [Google Scholar]
- [50].Utz PJ, Hottelet M, van Venrooij WJ, Anderson P, Association of phosphorylated serine/arginine (SR) splicing factors with the U1-small ribonucleoprotein (snRNP) autoantigen complex accompanies apoptotic cell death, J. Exp. Med 187 (1998) 547–560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Pfeifle J, Anderer FA, Franke M, Multiple phosphorylation of human SS-B/LA autoantigen and its effect on poly(U) and autoantibody binding, Biochim. Biophys. Acta 928 (1987) 217–226. [DOI] [PubMed] [Google Scholar]
- [52].Rutjes SA, Utz PJ, van der Heijden A, Broekhuis C, van Venrooij WJ, Pruijn GJ, The La (SS-B) autoantigen, a key protein in RNA biogenesis, is dephosphorylated and cleaved early during apoptosis, Cell Death Differ 6 (1999) 976–986. [DOI] [PubMed] [Google Scholar]
- [53].Zampieri S, Degen W, Ghiradello A, Doria A, van Venrooij WJ, Dephosphorylation of autoantigenic ribosomal P proteins during Fas-L induced apoptosis: a possible trigger for the development of the autoimmune response in patients with systemic lupus erythematosus, Ann. Rheum. Dis 60 (2001) 72–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Saunders RD, Nakajima ST, Rai SN, Pan J, Gercel-Taylor C, Taylor DD, Alterations in antibody subclass immune reactivity to trophoblast-derived fetal fibronectin and α2-macroglobulin in women with recurrent pregnancy loss, Am. J. Reprod. Immunol 68 (2012) 438–449. [DOI] [PubMed] [Google Scholar]
- [55].Petrohai A, Nagy G, Bosze S, Hudecz F, Zsiros E, Paragh G, et al. , Detection of citrate synthase-reacting autoantibodies after heart transplantation: an epitope mapping study, Transpl. Int. : official journal of the European Society for Organ Transplantation 17 (2005) 834–840. [DOI] [PubMed] [Google Scholar]
- [56].Mande PV, Parikh FR, Hinduja I, Zaveri K, Vaidya R, Gajbhiye R, et al. , Identification and validation of candidate biomarkers involved in human ovarian autoimmunity, Reprod. Biomed. Online 23 (2011) 471–483. [DOI] [PubMed] [Google Scholar]
- [57].van Beers JJ, Schwarte CM, Stammen-Vogelzangs J, Oosterink E, Bozic B, Pruijn GJ, The rheumatoid arthritis synovial fluid citrullinome reveals novel citrullinated epitopes in apolipoprotein E, myeloid nuclear differentiation antigen, and beta-actin, Arthritis Rheum 65 (2013) 69–80. [DOI] [PubMed] [Google Scholar]
- [58].Hanrotel-Saliou C, Segalen I, Le Meur Y, Youinou P, Renaudineau Y, Glomerular antibodies in lupus nephritis, Clin. Rev. Allergy Immunol 40 (2011) 151–158. [DOI] [PubMed] [Google Scholar]
- [59].Bei R, Mizejewski GJ, Alpha-fetoprotein is an autoantigen in hepatocellular carcinoma and juvenile Batten disease, Front. Biosci 25 (2020) 912–929. [DOI] [PubMed] [Google Scholar]
- [60].Rivner MH, Quarles BM, Pan JX, Yu Z, Howard JF Jr., Corse A, et al. , Clinical features of LRP4/agrin-antibody-positive myasthenia gravis: a multicenter study, Muscle Nerve 62 (2020) 333–343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Prunotto M, Carnevali ML, Candiano G, Murtas C, Bruschi M, Corradini E, et al. , Autoimmunity in membranous nephropathy targets aldose reductase and SOD2, J. Am. Soc. Nephrol. : JASN (J. Am. Soc. Nephrol.) 21 (2010) 507–519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Nehring J, Schirmbeck LA, Friebus-Kardash J, Dubler D, Huynh-Do U, Chizzolini C, et al. , Autoantibodies against albumin in patients with systemic lupus erythematosus, Front. Immunol 9 (2018) 2090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Edassery SL, Shatavi SV, Kunkel JP, Hauer C, Brucker C, Penumatsa K, et al. , Autoantigens in ovarian autoimmunity associated with unexplained infertility and premature ovarian failure, Fertil. Steril 94 (2010) 2636–2641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Cheng KC, Wu YJ, Cheng KH, Cheng KY, Chen KJ, Wu WC, et al. , Autoantibody against aldehyde dehydrogenase 2 could be a biomarker to monitor progression of Graves’ orbitopathy, Graefe’s archive for clinical and experimental ophthalmology = Albrecht von Graefes Archiv fur klinische und experimentelle Ophthalmologie 256 (2018) 1195–1201. [DOI] [PubMed] [Google Scholar]
- [65].Privitera D, Corti V, Alessio M, Volontè MA, Lampasona V, Comi G, et al. , Proteomic identification of aldolase A as an autoantibody target in patients with atypical movement disorders, Neurol. Sci. : official journal of the Italian Neurological Society and of the Italian Society of Clinical Neurophysiology 34 (2013) 313–320. [DOI] [PubMed] [Google Scholar]
- [66].Pott MC, Frede N, Wanders J, Hammarström L, Glocker EO, Glocker C, et al. , Autoantibodies against BAFF, APRIL or IL21 - an alternative pathogenesis for antibody-deficiencies? BMC Immunol 18 (2017) 34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].Caster DJ, Korte EA, Merchant ML, Klein JB, Wilkey DW, Rovin BH, et al. , Autoantibodies targeting glomerular annexin A2 identify patients with proliferative lupus nephritis, Proteonomics Clin. Appl 9 (2015) 1012–1020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Scholz P, Auler M, Brachvogel B, Benzing T, Mallman P, Streichert T, et al. , Detection of multiple annexin autoantibodies in a patient with recurrent miscarriages, fulminant stroke and seronegative antiphospholipid syndrome, Biochem. Med 26 (2016) 272–278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [69].Satoh A, Suzuki K, Takayama E, Kojima K, Hidaka T, Kawakami M, et al. , Detection of anti-annexin IV and V antibodies in patients with antiphospholipid syndrome and systemic lupus erythematosus, J. Rheumatol 26 (1999) 1715–1720. [PubMed] [Google Scholar]
- [70].Hrycek A, Cieślik P, Annexin A5 and anti-annexin antibodies in patients with systemic lupus erythematosus, Rheumatol. Int 32 (2012) 1335–1342. [DOI] [PubMed] [Google Scholar]
- [71].Seko Y, Matsumoto A, Fukuda T, Imai Y, Fujimura T, Taka H, et al. , A case of neonatal lupus erythematosus presenting delayed dilated cardiomyopathy with circulating autoantibody to annexin A6, Int. Heart J 48 (2007) 407–415. [DOI] [PubMed] [Google Scholar]
- [72].Jarius S, Wildemann B, ‘Medusa head ataxia’: the expanding spectrum of Purkinje cell antibodies in autoimmune cerebellar ataxia. Part 3: anti-Yo/CDR2, anti-Nb/AP3B2, PCA-2, anti-Tr/DNER, other antibodies, diagnostic pitfalls, summary and outlook, J. Neuroinflammation 12 (2015) 168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Katsumata Y, Kawaguchi Y, Baba S, Hattori S, Tahara K, Ito K, et al. , Identification of three new autoantibodies associated with systemic lupus erythematosus using two proteomic approaches, Mol. Cell. Proteomics : MCP 10 (2011). M110.005330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Krishnan B, Massilamany C, Basavalingappa RH, Gangaplara A, Rajasekaran RA, Afzal MZ, et al. , Epitope mapping of SERCA2a identifies an antigenic determinant that induces mainly atrial myocarditis in A/J mice, J. Immunol 200 (2018) 523–537. Baltimore, Md : 1950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Creaney J, Dick IM, Yeoman D, Wong S, Robinson BW, Auto-antibodies to β-F1-ATPase and vimentin in malignant mesothelioma, PloS One 6 (2011), e26515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [76].Beutgen VM, Schmelter C, Pfeiffer N, Grus FH, Autoantigens in the trabecular meshwork and glaucoma-specific alterations in the natural autoantibody repertoire, Clinical & translational immunology 9 (2020), e01101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Kishore U, Sontheimer RD, Sastry KN, Zappi EG, Hughes GR, Khamashta MA, et al. , The systemic lupus erythematosus (SLE) disease autoantigen-calreticulin can inhibit C1q association with immune complexes, Clin. Exp. Immunol 108 (1997) 181–190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [78].Weber CK, Haslbeck M, Englbrecht M, Sehnert B, Mielenz D, Graef D, et al. , Antibodies to the endoplasmic reticulum-resident chaperones calnexin, BiP and Grp94 in patients with rheumatoid arthritis and systemic lupus erythematosus, Rheumatology 49 (2010) 2255–2263. [DOI] [PubMed] [Google Scholar]
- [79].Li WH, Zhao J, Li HY, Liu H, Li AL, Wang HX, et al. , Proteomics-based identification of autoantibodies in the sera of healthy Chinese individuals from Beijing, Proteomics 6 (2006) 4781–4789. [DOI] [PubMed] [Google Scholar]
- [80].Hirai K, Maeda H, Omori K, Yamamoto T, Kokeguchi S, Takashiba S, Serum antibody response to group II chaperonin from Methanobrevibacter oralis and human chaperonin CCT, Pathogens and disease 68 (2013) 12–19. [DOI] [PubMed] [Google Scholar]
- [81].Goto M, Kuribayashi K, Takahashi Y, Kondoh T, Tanaka M, Kobayashi D, et al. , Identification of autoantibodies expressed in acquired aplastic anaemia, Br. J. Haematol 160 (2013) 359–362. [DOI] [PubMed] [Google Scholar]
- [82].Vece TJ, Watkin LB, Nicholas S, Canter D, Braun MC, Guillerman RP, et al. , Copa syndrome: a novel autosomal dominant immune dysregulatory disease, J. Clin. Immunol 36 (2016) 377–387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Garcia MC, Zhou J, Henning D, Arnett FC, Valdez BC, Unique epitopes in RNA helicase II/Gu protein recognized by serum from a watermelon stomach patient, Mol. Immunol 37 (2000) 351–359. [DOI] [PubMed] [Google Scholar]
- [84].Scofield RH, Do we need new autoantibodies in lupus? Arthritis Res. Ther 12 (2010) 120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [85].Dai L, Li J, Tsay JJ, Yie TA, Munger JS, Pass H, et al. , Identification of autoantibodies to ECH1 and HNRNPA2B1 as potential biomarkers in the early detection of lung cancer, OncoImmunology 6 (2017), e1310359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Kim EG, Kwak SH, Hwang D, Yi EC, Park KS, Koo BK, et al. , The level of autoantibodies targeting eukaryote translation elongation factor 1 α1 and ubiquitin-conjugating enzyme 2L3 in nondiabetic young adults, Diabetes Metab. J 40 (2016) 154–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].Mooney CJ, Dunphy EJ, Stone B, McNeel DG, Identification of autoantibodies elicited in a patient with prostate cancer presenting as dermatomyositis, Int. J. Urol 13 (2006) 211–217. [DOI] [PubMed] [Google Scholar]
- [88].Fernández-Madrid F, Tang N, Alansari H, Granda JL, Tait L, Amirikia KC, et al. , Autoantibodies to annexin XI-A and other autoantigens in the diagnosis of breast cancer, Canc. Res 64 (2004) 5089–5096. [DOI] [PubMed] [Google Scholar]
- [89].Bach M, Winkelmann G, Luhrmann R, 20S small nuclear ribonucleoprotein U5 shows a surprisingly complex protein composition, Proc. Natl. Acad. Sci. U. S. A 86 (1989) 6038–6042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Heo CK, Hwang HM, Lee HJ, Kwak SS, Yoo JS, Yu DY, et al. , Serum anti-EIF3A autoantibody as a potential diagnostic marker for hepatocellular carcinoma, Sci. Rep 9 (2019) 11059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Betteridge Z, Chinoy H, Vencovsky J, Winer J, Putchakayala K, Ho P, et al. , Identification of a novel autoantigen eukaryotic initiation factor 3 associated with polymyositis, Rheumatology 59 (2020) 1026–1030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Pagaza-Straffon C, Marchat LA, Herrera L, Díaz-Chávez J, Avante MG, Rodríguez YP, et al. , Evaluation of a panel of tumor-associated antigens in breast cancer, Canc. Biomarkers : section A of Disease markers 27 (2020) 207–211. [DOI] [PubMed] [Google Scholar]
- [93].Moscato S, Pratesi F, Sabbatini A, Chimenti D, Scavuzzo M, Passatino R, et al. , Surface expression of a glycolytic enzyme, alpha-enolase, recognized by autoantibodies in connective tissue disorders, Eur. J. Immunol 30 (2000) 3575–3584. [DOI] [PubMed] [Google Scholar]
- [94].Akatsuka T, Kobayashi N, Ishikawa T, Saito T, Shindo M, Yamauchi M, et al. , Autoantibody response to microsomal epoxide hydrolase in hepatitis C and A, J. Autoimmun 28 (2007) 7–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Leveque C, Hoshino T, David P, Shoji-Kasai Y, Leys K, Omori A, et al. , The synaptic vesicle protein synaptotagmin associates with calcium channels and is a putative Lambert-Eaton myasthenic syndrome antigen, Proc. Natl. Acad. Sci. U. S. A 89 (1992) 3625–3629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [96].Capello M, Cappello P, Linty FC, Chiarle R, Sperduti I, Novarino A, et al. , Autoantibodies to Ezrin are an early sign of pancreatic cancer in humans and in genetically engineered mouse models, J. Hematol. Oncol 6 (2013) 67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [97].Desmetz C, Bascoul-Mollevi C, Rochaix P, Lamy PJ, Kramar A, Rouanet P, et al. , Identification of a new panel of serum autoantibodies associated with the presence of in situ carcinoma of the breast in younger women, Clin. Canc. Res. : an official journal of the American Association for Cancer Research 15 (2009) 4733–4741. [DOI] [PubMed] [Google Scholar]
- [98].Kamhieh-Milz J, Sterzer V, Celik H, Khorramshahi O, Fadl Hassan Moftah R, Salama A, Identification of novel autoantigens via mass spectroscopy-based antibody-mediated identification of autoantigens (MS-AMIDA) using immune thrombocytopenic purpura (ITP) as a model disease, Journal of proteomics 157 (2017) 59–70. [DOI] [PubMed] [Google Scholar]
- [99].Adachi-Hayama M, Adachi A, Shinozaki N, Matsutani T, Hiwasa T, Takiguchi M, et al. , Circulating anti-filamin C autoantibody as a potential serum biomarker for low-grade gliomas, BMC Canc 14 (2014) 452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].Dong X, Yang M, Sun H, Lü J, Zheng Z, Li Z, et al. , Combined measurement of CA 15–3 with novel autoantibodies improves diagnostic accuracy for breast cancer, OncoTargets Ther 6 (2013) 273–279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Terrier B, Tamby MC, Camoin L, Guilpain P, Broussard C, Bussone G, et al. , Identification of target antigens of antifibroblast antibodies in pulmonary arterial hypertension, Am. J. Respir. Crit. Care Med 177 (2008) 1128–1134. [DOI] [PubMed] [Google Scholar]
- [102].Kit Y, Starykovych M, Vajrychova M, Lenco J, Zastavna D, Stoika R, Detection of novel auto-antigens in patients with recurrent miscarriage: description of an approach and preliminary findings, Croat. Med. J 55 (2014) 259–264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [103].Delunardo F, Soldati D, Bellisario V, Berry A, Camerini S, Crescenzi M, et al. , Anti-GAPDH autoantibodies as a pathogenic determinant and potential biomarker of neuropsychiatric diseases, Arthritis & rheumatology (Hoboken, NJ) 68 (2016) 2708–2716. [DOI] [PubMed] [Google Scholar]
- [104].Kiyota A, Iwama S, Sugimura Y, Takeuchi S, Takagi H, Iwata N, et al. , Identification of the novel autoantigen candidate Rab GDP dissociation inhibitor alpha in isolated adrenocorticotropin deficiency, Endocr. J 62 (2015) 153–160. [DOI] [PubMed] [Google Scholar]
- [105].Massa O, Alessio M, Russo L, Nardo G, Bonetto V, Bertuzzi F, et al. , Serological Proteome Analysis (SERPA) as a tool for the identification of new candidate autoantigens in type 1 diabetes, Journal of proteomics 82 (2013) 263–273. [DOI] [PubMed] [Google Scholar]
- [106].Carlsson L, Ronquist G, Nilsson BO, Larsson A, Dominant prostasome immunogens for sperm-agglutinating autoantibodies of infertile men, J. Androl 25 (2004) 699–705. [DOI] [PubMed] [Google Scholar]
- [107].Muraki Y, Matsumoto I, Chino Y, Hayashi T, Suzuki E, Goto D, et al. , Glucose-6-phosphate isomerase variants play a key role in the generation of anti-GPI antibodies: possible mechanism of autoantibody production, Biochem. Biophys. Res. Commun 323 (2004) 518–522. [DOI] [PubMed] [Google Scholar]
- [108].Yang J, Tezel G, Patil RV, Romano C, Wax MB, Serum autoantibody against glutathione S-transferase in patients with glaucoma, Invest. Ophthalmol. Vis. Sci 42 (2001) 1273–1276. [PubMed] [Google Scholar]
- [109].Rubin RL, Bell SA, Burlingame RW, Autoantibodies associated with lupus induced by diverse drugs target a similar epitope in the (H2A-H2B)-DNA complex, J. Clin. Invest 90 (1992) 165–173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [110].Burlingame RW, Boey ML, Starkebaum G, Rubin RL, The central role of chromatin in autoimmune responses to histones and DNA in systemic lupus erythematosus, J. Clin. Invest 94 (1994) 184–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [111].Wesierska-Gadek J, Penner E, Lindner H, Hitchman E, Sauermann G, Autoantibodies against different histone H1 subtypes in systemic lupus erythematosus sera, Arthritis Rheum 33 (1990) 1273–1278. [DOI] [PubMed] [Google Scholar]
- [112].Baranova SV, Dmitrienok PS, Ivanisenko NV, Buneva VN, Nevinsky GA, Antibodies to H2a and H2b histones from the sera of HIV-infected patients catalyze site-specific degradation of these histones, Mol. Biosyst 13 (2017) 1090–1101. [DOI] [PubMed] [Google Scholar]
- [113].Bruschi M, Galetti M, Sinico RA, Moroni G, Bonanni A, Radice A, et al. , Glomerular autoimmune multicomponents of human lupus nephritis in vivo (2): planted antigens, J. Am. Soc. Nephrol. : JASN (J. Am. Soc. Nephrol.) 26 (2015) 1905–1924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [114].van Bavel CC, Dieker J, Muller S, Briand JP, Monestier M, Berden JH, et al. , Apoptosis-associated acetylation on histone H2B is an epitope for lupus autoantibodies, Mol. Immunol 47 (2009) 511–516. [DOI] [PubMed] [Google Scholar]
- [115].Baranova SV, Dmitrenok PS, Zubkova AD, Ivanisenko NV, Odintsova ES, Buneva VN, et al. , Antibodies against H3 and H4 histones from the sera of HIV-infected patients catalyze site-specific degradation of these histones, J. Mol. Recogn. : JMR (J. Mol. Recognit.) 31 (2018), e2703. [DOI] [PubMed] [Google Scholar]
- [116].Konig MF, Giles JT, Nigrovic PA, Andrade F, Antibodies to native and citrullinated RA33 (hnRNP A2/B1) challenge citrullination as the inciting principle underlying loss of tolerance in rheumatoid arthritis, Ann. Rheum. Dis 75 (2016) 2022–2028. [DOI] [PubMed] [Google Scholar]
- [117].Heegaard NH, Larsen MR, Muncrief T, Wiik A, Roepstorff P, Heterogeneous nuclear ribonucleoproteins C1/C2 identified as autoantigens by biochemical and mass spectrometric methods, Arthritis Res 2 (2000) 407–414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [118].Op De Beéck K, Maes L, Van den Bergh K, Derua R, Waelkens E, Van Steen K, et al. , Heterogeneous nuclear RNPs as targets of autoantibodies in systemic rheumatic diseases, Arthritis Rheum 64 (2012) 213–221. [DOI] [PubMed] [Google Scholar]
- [119].Yang L, Fujimoto M, Murota H, Serada S, Fujimoto M, Honda H, et al. , Proteomic identification of heterogeneous nuclear ribonucleoprotein K as a novel cold-associated autoantigen in patients with secondary Raynaud’s phenomenon, Rheumatology 54 (2015) 349–358. [DOI] [PubMed] [Google Scholar]
- [120].Hassfeld W, Chan EK, Mathison DA, Portman D, Dreyfuss G, Steiner G, et al. , Molecular definition of heterogeneous nuclear ribonucleoprotein R (hnRNP R) using autoimmune antibody: immunological relationship with hnRNP P, Nucleic Acids Res 26 (1998) 439–445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [121].Harlow L, Rosas IO, Gochuico BR, Mikuls TR, Dellaripa PF, Oddis CV, et al. , Identification of citrullinated hsp90 isoforms as novel autoantigens in rheumatoid arthritis-associated interstitial lung disease, Arthritis Rheum 65 (2013) 869–879. [DOI] [PubMed] [Google Scholar]
- [122].Qin HY, Mahon JL, Atkinson MA, Chaturvedi P, Lee-Chan E, Singh B, Type 1 diabetes alters anti-hsp90 autoantibody isotype, J. Autoimmun 20 (2003) 237–245. [DOI] [PubMed] [Google Scholar]
- [123].Cid C, Regidor I, Alcazar A, Anti-heat shock protein 90beta antibodies are detected in patients with multiple sclerosis during remission, J. Neuroimmunol 184 (2007) 223–226. [DOI] [PubMed] [Google Scholar]
- [124].Suzuki S, Utsugisawa K, Iwasa K, Satoh T, Nagane Y, Yoshikawa H, et al. , Autoimmunity to endoplasmic reticulum chaperone GRP94 in myasthenia gravis, J. Neuroimmunol 237 (2011) 87–92. [DOI] [PubMed] [Google Scholar]
- [125].Mišunová M, Svitálková T, Pleštilová L, Kryštufková O, Tegzová D, Svobodová R, et al. , Molecular markers of systemic autoimmune disorders: the expression of MHC-located HSP70 genes is significantly associated with autoimmunity development, Clin. Exp. Rheumatol 35 (2017) 33–42. [PubMed] [Google Scholar]
- [126].Shimizu F, Schaller KL, Owens GP, Cotleur AC, Kellner D, Takeshita Y, et al. , Glucose-regulated protein 78 autoantibody associates with blood-brain barrier disruption in neuromyelitis optica, Sci. Transl. Med 9 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [127].Iannaccone A, Giorgianni F, New DD, Hollingsworth TJ, Umfress A, Alhatem AH, et al. , Circulating autoantibodies in age-related macular degeneration recognize human macular tissue antigens implicated in autophagy, immunomodulation, and protection from oxidative stress and apoptosis, PloS One 10 (2015), e0145323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [128].Papp D, Prohászka Z, Kocsis J, Füst G, Bánhegyi D, Raynes DA, et al. , Development of a sensitive assay for the measurement of antibodies against heat shock protein binding protein 1 (HspBP1): increased levels of anti-HspBP1 IgG are prevalent in HIV infected subjects, J. Med. Virol 76 (2005) 464–469. [DOI] [PubMed] [Google Scholar]
- [129].Fillit H, Shibata S, Sasaki T, Spiera H, Kerr LD, Blake M, Autoantibodies to the protein core of vascular basement membrane heparan sulfate proteoglycan in systemic lupus erythematosus, Autoimmunity 14 (1993) 243–249. [DOI] [PubMed] [Google Scholar]
- [130].Tiumentseva M, Morozova V, Zakabunin A, Korobko D, Malkova N, Filipenko M, et al. , Use of the VH6–1 gene segment to code for anti-interleukin-18 autoantibodies in multiple sclerosis, Immunogenetics 68 (2016) 237–246. [DOI] [PubMed] [Google Scholar]
- [131].Bremer HD, Landegren N, Sjoberg R, Hallgren A, Renneker S, Lattwein E, et al. , ILF2 and ILF3 are autoantigens in canine systemic autoimmune disease, Sci. Rep 8 (2018) 4852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [132].Presslauer S, Hinterhuber G, Cauza K, Horvat R, Rappersberger K, Wolff K, et al. , RasGAP-like protein IQGAP1 is expressed by human keratinocytes and recognized by autoantibodies in association with bullous skin disease, J. Invest. Dermatol 120 (2003) 365–371. [DOI] [PubMed] [Google Scholar]
- [133].Idborg H, Zandian A, Sandberg AS, Nilsson B, Elvin K, Truedsson L, et al. , Two subgroups in systemic lupus erythematosus with features of antiphospholipid or Sjögren’s syndrome differ in molecular signatures and treatment perspectives, Arthritis Res. Ther 21 (2019) 62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [134].Targoff IN, Trieu EP, Miller FW, Reaction of anti-OJ autoantibodies with components of the multi-enzyme complex of aminoacyl-tRNA synthetases in addition to isoleucyl-tRNA synthetase, J. Clin. Invest 91 (1993) 2556–2564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [135].Ola TO, Biro PA, Hawa MI, Ludvigsson J, Locatelli M, Puglisi MA, et al. , Importin beta: a novel autoantigen in human autoimmunity identified by screening random peptide libraries on phage, J. Autoimmun 26 (2006) 197–207. [DOI] [PubMed] [Google Scholar]
- [136].Dyachenko L, Havrysh K, Lytovchenko A, Dosenko I, Antoniuk S, Filonenko V, et al. , Autoantibody Response to ZRF1 and KRR1 SEREX Antigens in Patients with Breast Tumors of Different Histological Types and Grades, vol. 2016, Disease markers, 2016, p. 5128720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [137].Kain R, Tadema H, McKinney EF, Benharkou A, Brandes R, Peschel A, et al. , High prevalence of autoantibodies to hLAMP-2 in anti-neutrophil cytoplasmic antibody-associated vasculitis, J. Am. Soc. Nephrol. : JASN (J. Am. Soc. Nephrol.) 23 (2012) 556–566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [138].Ueda K, Nakanishi T, Shimizu A, Takubo T, Matsuura N, Identification of L-plastin autoantibody in plasma of patients with non-Hodgkin’s lymphoma using a proteomics-based analysis, Ann. Clin. Biochem 45 (2008) 65–69. [DOI] [PubMed] [Google Scholar]
- [139].Braunschweig D, Krakowiak P, Duncanson P, Boyce R, Hansen RL, Ashwood P, et al. , Autism-specific maternal autoantibodies recognize critical proteins in developing brain, Transl. Psychiatry 3 (2013) e277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [140].Konstantinov KN, Galcheva-Gargova Z, Hoier-Madsen M, Wiik A, Ullman S, Halberg P, et al. , Autoantibodies to lamins A and C in sera of patients showing peripheral fluorescent antinuclear antibody pattern on HEP-2 cells, J. Invest. Dermatol 95 (1990) 304–308. [DOI] [PubMed] [Google Scholar]
- [141].Brito J, Biamonti G, Caporali R, Montecucco C, Autoantibodies to human nuclear lamin B2 protein. Epitope specificity in different autoimmune diseases, J. Immunol 153 (1994) 2268–2277. Baltimore, Md : 1950. [PubMed] [Google Scholar]
- [142].Tanaka M, Kishimura M, Ozaki S, Osakada F, Hashimoto H, Okubo M, et al. , Cloning of novel soluble gp130 and detection of its neutralizing autoantibodies in rheumatoid arthritis, J. Clin. Invest 106 (2000) 137–144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [143].Gadoth A, Kryzer TJ, Fryer J, McKeon A, Lennon VA, Pittock SJ, Microtubule-associated protein 1B: novel paraneoplastic biomarker, Ann. Neurol 81 (2017) 266–277. [DOI] [PubMed] [Google Scholar]
- [144].Vermeulen N, Vermeire S, Arijs I, Michiels G, Ballet V, Derua R, et al. , Seroreactivity against glycolytic enzymes in inflammatory bowel disease, Inflamm. Bowel Dis 17 (2011) 557–564. [DOI] [PubMed] [Google Scholar]
- [145].Suzuki K, Nagao T, Itabashi M, Hamano Y, Sugamata R, Yamazaki Y, et al. , A novel autoantibody against moesin in the serum of patients with MPO-ANCA-associated vasculitis, Nephrol. Dial. Transplant 29 (2014) 1168–1177. [DOI] [PubMed] [Google Scholar]
- [146].Marinou D, Katsifis G, Barouta G, Liaskos C, Sakkas LI, Tsakris A, et al. , Major vault protein/lung resistance related protein: a novel biomarker for rheumatoid arthritis, Clin. Exp. Rheumatol (2020). https://www.clinexprheumatol.org/abstract.asp?a=15829. [DOI] [PubMed] [Google Scholar]
- [147].von Muhlen CA, Chan EK, Peebles CL, Imai H, Kiyosawa K, Tan EM, Non-muscle myosin as target antigen for human autoantibodies in patients with hepatitis C virus-associated chronic liver diseases, Clin. Exp. Immunol 100 (1995) 67–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [148].Zasońska BA, Hlídková H, Petrovský E, Myronovskij S, Nehrych T, Negrych N, et al. , Monodisperse magnetic poly(glycidyl methacrylate) microspheres for isolation of autoantibodies with affinity for the 46 kDa form of unconventional Myo1C present in autoimmune patients, Mikrochim. Acta 185 (2018) 262. [DOI] [PubMed] [Google Scholar]
- [149].Qin Z, Lavingia B, Zou Y, Stastny P, Antibodies against nucleolin in recipients of organ transplants, Transplantation 92 (2011) 829–835. [DOI] [PubMed] [Google Scholar]
- [150].Le Naour F, Brichory F, Misek DE, Brechot C, Hanash SM, Beretta L, A distinct repertoire of autoantibodies in hepatocellular carcinoma identified by proteomic analysis, Mol. Cell. Proteomics : MCP 1 (2002) 197–203. [DOI] [PubMed] [Google Scholar]
- [151].Liu J, Xing X, Huang H, Jiang Y, He H, Xu X, et al. , Identification of antigenic proteins associated with trichloroethylene-induced autoimmune disease by serological proteome analysis, Toxicol. Appl. Pharmacol 240 (2009) 393–400. [DOI] [PubMed] [Google Scholar]
- [152].Brankin B, Skaar TC, Brotzman M, Trock B, Clarke R, Autoantibodies to the nuclear phosphoprotein nucleophosmin in breast cancer patients, Cancer Epidemiol. Biomark. Prev 7 (1998) 1109–1115. [PubMed] [Google Scholar]
- [153].Devaux JJ, Odaka M, Yuki N, Nodal proteins are target antigens in Guillain-Barré syndrome, J. Peripher. Nerv. Syst. : JPNS 17 (2012) 62–71. [DOI] [PubMed] [Google Scholar]
- [154].Nagayama S, Yokoi T, Tanaka H, Kawaguchi Y, Shirasaka T, Kamataki T, Occurrence of autoantibody to protein disulfide isomerase in patients with hepatic disorder, J. Toxicol. Sci 19 (1994) 163–169. [DOI] [PubMed] [Google Scholar]
- [155].Becker A, Ludwig N, Keller A, Tackenberg B, Eienbroker C, Oertel WH, et al. , Myasthenia gravis: analysis of serum autoantibody reactivities to 1827 potential human autoantigens by protein macroarrays, PloS One 8 (2013), e58095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [156].Houng AK, Maggini L, Clement CY, Reed GL, Identification and structure of activated-platelet protein-1, a protein with RNA-binding domain motifs that is expressed by activated platelets, Eur. J. Biochem 243 (1997) 209–218. [DOI] [PubMed] [Google Scholar]
- [157].Kaneda K, Takasaki Y, Takeuchi K, Yamada H, Nawata M, Matsushita M, et al. , Autoimmune response to proteins of proliferating cell nuclear antigen multiprotein complexes in patients with connective tissue diseases, J. Rheumatol 31 (2004) 2142–2150. [PubMed] [Google Scholar]
- [158].Caorsi C, Niccolai E, Capello M, Vallone R, Chattaragada MS, Alushi B, et al. , Protein disulfide isomerase A3-specific Th1 effector cells infiltrate colon cancer tissue of patients with circulating anti-protein disulfide isomerase A3 autoantibodies, Transl. Res. : J. Lab. Clin. Med 171 (2016) 17–28, e1–2. [DOI] [PubMed] [Google Scholar]
- [159].Zhang X, Shen W, Dong X, Fan J, Liu L, Gao X, et al. , Identification of novel autoantibodies for detection of malignant mesothelioma, PloS One 8 (2013), e72458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [160].Chen C, Liao D, Wang J, Liang Z, Yao Q, Anti-human protein S antibody induces tissue factor expression through a direct interaction with platelet phosphofructokinase, Thromb. Res 133 (2014) 222–228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [161].Wieczorek M, Czernik A, Paraneoplastic pemphigus: a short review, Clin. Cosmet. Invest. Dermatol 9 (2016) 291–295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [162].Mecoli CA, Adler BL, Yang Q, Hummers LK, Rosen A, Casciola-Rosen L, et al. , Cancer in Systemic Sclerosis: Analysis of Antibodies against Components of the Th/To Complex, Arthritis & rheumatology, Hoboken, NJ), 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [163].Loshaj-Shala A, Colzani M, Brezovska K, Poceva Panovska A, Suturkova L, Beretta G, Immunoproteomic identification of antigenic candidate Campylobacter jejuni and human peripheral nerve proteins involved in Guillain-Barré syndrome, J. Neuroimmunol 317 (2018) 77–83. [DOI] [PubMed] [Google Scholar]
- [164].Kratz A, Harding MW, Craft J, Mackworth-Young CG, Handschumacher RE, Autoantibodies against cyclophilin in systemic lupus erythematosus and Lyme disease, Clin. Exp. Immunol 90 (1992) 422–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [165].Karasawa R, Ozaki S, Nishioka K, Kato T, Autoantibodies to peroxiredoxin I and IV in patients with systemic autoimmune diseases, Microbiol. Immunol 49 (2005) 57–65. [DOI] [PubMed] [Google Scholar]
- [166].Lin LH, Xu YW, Huang LS, Hong CQ, Zhai TT, Liao LD, et al. , Serum proteomic-based analysis identifying autoantibodies against PRDX2 and PRDX3 as potential diagnostic biomarkers in nasopharyngeal carcinoma, Clin. Proteonomics 14 (2017) 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [167].Kobayashi S, Hiwasa T, Arasawa T, Kagaya A, Ishii S, Shimada H, et al. , Identification of specific and common diagnostic antibody markers for gastrointestinal cancers by SEREX screening using testis cDNA phage library, Oncotarget 9 (2018) 18559–18569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [168].Schild-Poulter C, Su A, Shih A, Kelly OP, Fritzler MJ, Goldstein R, et al. , Association of autoantibodies with Ku and DNA repair proteins in connective tissue diseases, Rheumatology 47 (2008) 165–171. [DOI] [PubMed] [Google Scholar]
- [169].Heo CK, Hwang HM, Lim WH, Lee HJ, Yoo JS, Lim KJ, et al. , Cyclic peptide mimotopes for the detection of serum anti-ATIC autoantibody biomarker in hepato-cellular carcinoma, Int. J. Mol. Sci (2020) 21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [170].Vaughan JH, Valbracht JR, Nguyen MD, Handley HH, Smith RS, Patrick K, et al. , Epstein-Barr virus-induced autoimmune responses. I. Immunoglobulin M autoantibodies to proteins mimicking and not mimicking Epstein-Barr virus nuclear antigen-1, J. Clin. Invest 95 (1995) 1306–1315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [171].Luo LY, Herrera I, Soosaipillai A, Diamandis EP, Identification of heat shock protein 90 and other proteins as tumour antigens by serological screening of an ovarian carcinoma expression library, Br. J. Canc 87 (2002) 339–343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [172].Doe K, Nozawa K, Hiruma K, Yamada Y, Matsuki Y, Nakano S, et al. , Antibody against chromatin assembly factor-1 is a novel autoantibody specifically recognized in systemic lupus erythematosus, Lupus 23 (2014) 1031–1041. [DOI] [PubMed] [Google Scholar]
- [173].Wagatsuma M, Kimura M, Suzuki R, Takeuchi F, Matsuta K, Watanabe Ezrin H, Radixin and moesin are possible auto-immune antigens in rheumatoid arthritis, Mol. Immunol 33 (1996) 1171–1176. [DOI] [PubMed] [Google Scholar]
- [174].Scofield RH, Harley JB, Autoantigenicity of Ro/SSA antigen is related to a nucleocapsid protein of vesicular stomatitis virus, Proc. Natl. Acad. Sci. U. S. A 88 (1991) 3343–3347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [175].Sato T, Uchiumi T, Kominami R, Arakawa M, Autoantibodies specific for the 20-KDal ribosomal large subunit protein L12, Biochem. Biophys. Res. Commun 172 (1990) 496–502. [DOI] [PubMed] [Google Scholar]
- [176].Absi M, La Vergne JP, Marzouki A, Giraud F, Rigal D, Reboud AM, et al. , Heterogeneity of ribosomal autoantibodies from human, murine and canine connective tissue diseases, Immunol. Lett 23 (1989) 35–41. [DOI] [PubMed] [Google Scholar]
- [177].Guialis A, Patrinou-Georgoula M, Tsifetaki N, Aidinis V, Sekeris CE, Moutsopoulos HM, Anti-5S RNA/protein (RNP) antibody levels correlate with disease activity in a patient with systemic lupus erythematosus (SLE) nephritis, Clin. Exp. Immunol 95 (1994) 385–389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [178].Frampton G, Moriya S, Pearson JD, Isenberg DA, Ward FJ, Smith TA, et al. , Identification of candidate endothelial cell autoantigens in systemic lupus erythematosus using a molecular cloning strategy: a role for ribosomal P protein P0 as an endothelial cell autoantigen, Rheumatology 39 (2000) 1114–1120. [DOI] [PubMed] [Google Scholar]
- [179].Neu E, von Mikecz AH, Hemmerich PH, Peter HH, Fricke M, Deicher H, et al. , Autoantibodies against eukaryotic protein L7 in patients suffering from systemic lupus erythematosus and progressive systemic sclerosis: frequency and correlation with clinical, serological and genetic parameters. The SLE Study Group, Clin. Exp. Immunol 100 (1995) 198–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [180].Elkon K, Bonfa E, Llovet R, Danho W, Weissbach H, Brot N, Properties of the ribosomal P2 protein autoantigen are similar to those of foreign protein antigens, Proc. Natl. Acad. Sci. U. S. A 85 (1988) 5186–5189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [181].Elkon K, Weissbach H, Brot N, Central nervous system function in systemic lupus erythematosus, Neurochem. Res 15 (1990) 401–406. [DOI] [PubMed] [Google Scholar]
- [182].Tycowski KT, Shu MD, Steitz JA, A small nucleolar RNA is processed from an intron of the human gene encoding ribosomal protein S3, Gene Dev 7 (1993) 1176–1190. [DOI] [PubMed] [Google Scholar]
- [183].Qiu J, Choi G, Li L, Wang H, Pitteri SJ, Pereira-Faca SR, et al. , Occurrence of autoantibodies to annexin I, 14-3-3 theta and LAMR1 in prediagnostic lung cancer sera, J. Clin. Oncol. : official journal of the American Society of Clinical Oncology 26 (2008) 5060–5066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [184].Yang JM, Hildebrandt B, Luderschmidt C, Pollard KM, Human scleroderma sera contain autoantibodies to protein components specific to the U3 small nucleolar RNP complex, Arthritis Rheum 48 (2003) 210–217. [DOI] [PubMed] [Google Scholar]
- [185].Kaji K, Fertig N, Medsger TA Jr., Satoh T, Hoshino K, Hamaguchi Y, et al. , Autoantibodies to RuvBL1 and RuvBL2: a novel systemic sclerosis-related antibody associated with diffuse cutaneous and skeletal muscle involvement, Arthritis Care Res 66 (2014) 575–584. [DOI] [PubMed] [Google Scholar]
- [186].Hwang HM, Heo CK, Lee HJ, Kwak SS, Lim WH, Yoo JS, et al. , Identification of anti-SF3B1 autoantibody as a diagnostic marker in patients with hepatocellular carcinoma, J. Transl. Med 16 (2018) 177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [187].Kistner A, Bigler MB, Glatz K, Egli SB, Baldin FS, Marquardsen FA, et al. , Characteristics of autoantibodies targeting 14-3-3 proteins and their association with clinical features in newly diagnosed giant cell arteritis, Rheumatology 56 (2017) 829–834. [DOI] [PubMed] [Google Scholar]
- [188].Chumpitazi BF, Bouillet L, Drouet MT, Kuhn L, Garin J, Zarski JP, et al. , Biological autoimmunity screening in hepatitis C patients by anti-HepG2 lysate and anti-heat shock protein 70.1 autoantibodies, Eur. J. Clin. Microbiol. Infect. Dis 28 (2009) 137–146, official publication of the European Society of Clinical Microbiology. [DOI] [PubMed] [Google Scholar]
- [189].Kubo M, Ihn H, Kuwana M, Asano Y, Tamaki T, Yamane K, et al. , Anti-U5 snRNP antibody as a possible serological marker for scleroderma-polymyositis overlap, Rheumatology 41 (2002) 531–534. [DOI] [PubMed] [Google Scholar]
- [190].Yamamoto AM, Amoura Z, Johannet C, Jeronimo AL, Campos H, Koutouzov S, et al. , Quantitative radioligand assays using de novo-synthesized recombinant autoantigens in connective tissue diseases: new tools to approach the pathogenic significance of anti-RNP antibodies in rheumatic diseases, Arthritis Rheum 43 (2000) 689–698. [DOI] [PubMed] [Google Scholar]
- [191].Huntriss JD, Latchman DS, Williams DG, Lupus autoantibodies discriminate between the highly homologous Sm polypeptides B/B’ and SmN by binding an epitope restricted to B/B, Clin. Exp. Immunol 92 (1993) 263–267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [192].Brahms H, Raymackers J, Union A, de Keyser F, Meheus L, Luhrmann R, The C-terminal RG dipeptide repeats of the spliceosomal Sm proteins D1 and D3 contain symmetrical dimethylarginines, which form a major B-cell epitope for anti-Sm autoantibodies, J. Biol. Chem 275 (2000) 17122–17129. [DOI] [PubMed] [Google Scholar]
- [193].McClain MT, Ramsland PA, Kaufman KM, James JA, Anti-sm autoantibodies in systemic lupus target highly basic surface structures of complexed spliceosomal autoantigens, J. Immunol 168 (2002) 2054–2062. Baltimore, Md : 1950. [DOI] [PubMed] [Google Scholar]
- [194].Satoh M, Chan JY, Ross SJ, Ceribelli A, Cavazzana I, Franceschini F, et al. , Autoantibodies to survival of motor neuron complex in patients with polymyositis: immunoprecipitation of D, E, F, and G proteins without other components of small nuclear ribonucleoproteins, Arthritis Rheum 63 (2011) 1972–1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [195].van Blitterswijk M, Gulati S, Smoot E, Jaffa M, Maher N, Hyman BT, et al. , Anti-superoxide dismutase antibodies are associated with survival in patients with sporadic amyotrophic lateral sclerosis. Amyotrophic lateral sclerosis, in: Official Publication of the World Federation of Neurology Research Group on Motor Neuron Diseases, vol. 12, 2011, pp. 430–438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [196].Garbarz M, Dhermy D, Bournier O, Bezeaud A, Boivin P, Anti-spectrin in sera containing smooth muscle autoantibodies from patients with chronic active hepatitis, Clin. Exp. Immunol 43 (1981) 87–93. [PMC free article] [PubMed] [Google Scholar]
- [197].Zaninoni A, Vercellati C, Imperiali FG, Marcello AP, Fattizzo B, Fermo E, et al. , Detection of red blood cell antibodies in mitogen-stimulated cultures from patients with hereditary spherocytosis, Transfusion 55 (2015) 2930–2938. [DOI] [PubMed] [Google Scholar]
- [198].Utz PJ, Hottelet M, Le TM, Kim SJ, Geiger ME, van Venrooij WJ, et al. , The 72-kDa component of signal recognition particle is cleaved during apoptosis, J. Biol. Chem 273 (1998) 35362–35370. [DOI] [PubMed] [Google Scholar]
- [199].Lipes BD, Keene JD, Autoimmune Epitopes in Messenger RNA, vol. 8, RNA, New York, NY: ), 2002, pp. 762–771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [200].Santoro P, De Andrea M, Migliaretti G, Trapani C, Landolfo S, Gariglio M, High prevalence of autoantibodies against the nuclear high mobility group (HMG) protein SSRP1 in sera from patients with systemic lupus erythematosus, but not other rheumatic diseases, J. Rheumatol 29 (2002) 90–93. [PubMed] [Google Scholar]
- [201].Cortini A, Bembich S, Marson L, Cocco E, Edomi P, Identification of novel non-myelin biomarkers in multiple sclerosis using an improved phage-display approach, PloS One 14 (2019), e0226162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [202].Niland B, Miklossy G, Banki K, Biddison WE, Casciola-Rosen L, Rosen A, et al. , Cleavage of transaldolase by granzyme B causes the loss of enzymatic activity with retention of antigenicity for multiple sclerosis patients, J. Immunol 184 (2010) 4025–4032. Baltimore, Md : 1950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [203].Lee YJ, Ting WH, Yang YW, Lin CJ, Hsieh YT, Huang CY, et al. , HLA-DQ genotype and biochemical characterization of anti-transglutaminase 2 antibodies in patients with type 1 diabetes mellitus in Taiwan, Faseb. J. : official publication of the Federation of American Societies for Experimental Biology 34 (2020) 8459–8474. [DOI] [PubMed] [Google Scholar]
- [204].Muto M, Mori M, Hiwasa T, Takiguchi M, Iwadate Y, Uzawa A, et al. , Novel serum autoantibodies against talin1 in multiple sclerosis: possible pathogenetic roles of the antibodies, J. Neuroimmunol 284 (2015) 30–36. [DOI] [PubMed] [Google Scholar]
- [205].Mariampillai K, Granger B, Amelin D, Guiguet M, Hachulla E, Maurier F, et al. , Development of a new classification system for idiopathic inflammatory myopathies based on clinical manifestations and myositis-specific autoantibodies, JAMA neurology 75 (2018) 1528–1537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [206].Geng X, Biancone L, Dai HH, Lin JJ, Yoshizaki N, Dasgupta A, et al. , Tropomyosin isoforms in intestinal mucosa: production of autoantibodies to tropomyosin isoforms in ulcerative colitis, Gastroenterology 114 (1998) 912–922. [DOI] [PubMed] [Google Scholar]
- [207].Gajbhiye R, Sonawani A, Khan S, Suryawanshi A, Kadam S, Warty N, et al. , Identification and validation of novel serum markers for early diagnosis of endometriosis, Hum. Reprod 27 (2012) 408–417. [DOI] [PubMed] [Google Scholar]
- [208].Kimura A, Sakurai T, Yamada M, Koumura A, Hayashi Y, Tanaka Y, et al. , Anti-endothelial cell antibodies in patients with cerebral small vessel disease, Curr. Neurovascular Res 9 (2012) 296–301. [DOI] [PubMed] [Google Scholar]
- [209].Zhao X, Cheng Y, Gan Y, Jia R, Zhu L, Sun X, Anti-tubulin-alpha-1C autoantibody in systemic lupus erythematosus: a novel indicator of disease activity and vasculitis manifestations, Clin. Rheumatol 37 (2018) 1229–1237. [DOI] [PubMed] [Google Scholar]
- [210].Matthes T, Wolff A, Soubiran P, Gros F, Dighiero G, Antitubulin antibodies. II. Natural autoantibodies and induced antibodies recognize different epitopes on the tubulin molecule, J. Immunol 141 (1988) 3135–3141. Baltimore, Md : 1950. [PubMed] [Google Scholar]
- [211].Prasannan L, Misek DE, Hinderer R, Michon J, Geiger JD, Hanash SM, Identification of beta-tubulin isoforms as tumor antigens in neuroblastoma, Clin. Canc. Res. : an official journal of the American Association for Cancer Research 6 (2000) 3949–3956. [PubMed] [Google Scholar]
- [212].Muro Y, Ogawa Y, Kato Y, Hagiwara M, Autoantibody to thioredoxin reductase in an ovarian cancer patient, Biochem. Biophys. Res. Commun 242 (1998) 267–271. [DOI] [PubMed] [Google Scholar]
- [213].Betteridge ZE, Gunawardena H, Chinoy H, North J, Ollier WE, Cooper RG, et al. , Clinical and human leucocyte antigen class II haplotype associations of autoantibodies to small ubiquitin-like modifier enzyme, a dermatomyositis-specific autoantigen target, in UK Caucasian adult-onset myositis, Ann. Rheum. Dis 68 (2009) 1621–1625. [DOI] [PubMed] [Google Scholar]
- [214].Almamy A, Schwerk C, Schroten H, Ishikawa H, Asif AR, Reuss B, Interactions of antisera to different Chlamydia and Chlamydophila species with the ribosomal protein RPS27a correlate with impaired protein synthesis in a human choroid plexus papilloma cell line, Immunol. Res 65 (2017) 1110–1123. [DOI] [PubMed] [Google Scholar]
- [215].Pluta AF, Earnshaw WC, Specific interaction between human kinetochore protein CENP-C and a nucleolar transcriptional regulator, J. Biol. Chem 271 (1996) 18767–18774. [DOI] [PubMed] [Google Scholar]
- [216].Li X, Sun J, Mu R, Gan Y, Wang G, He J, et al. , The clinical significance of ubiquitin carboxyl hydrolase L1 and its autoantibody in neuropsychiatric systemic lupus erythematosus, Clin. Exp. Rheumatol 37 (2019) 474–480. [PubMed] [Google Scholar]
- [217].Zhou Y, Cui J, Du H, Autoantibody-targeted TAAs in pancreatic cancer: a comprehensive analysis, Pancreatology 19 (2019) 760–768. [DOI] [PubMed] [Google Scholar]
- [218].Miyachi K, Hosaka H, Nakamura N, Miyakawa H, Mimori T, Shibata M, et al. , Anti-p97/VCP antibodies: an autoantibody marker for a subset of primary biliary cirrhosis patients with milder disease? Scand. J. Immunol 63 (2006) 376–382. [DOI] [PubMed] [Google Scholar]
- [219].Li FJ, Surolia R, Li H, Wang Z, Kulkarni T, Liu G, et al. , Autoimmunity to vimentin is associated with outcomes of patients with idiopathic pulmonary fibrosis, J. Immunol 199 (2017) 1596–1605. Baltimore, Md : 1950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [220].Mimori T, Ohosone Y, Hama N, Suwa A, Akizuki M, Homma M, et al. , Isolation and characterization of cDNA encoding the 80-kDa subunit protein of the human autoantigen Ku (p70/p80) recognized by autoantibodies from patients with scleroderma-polymyositis overlap syndrome, Proc. Natl. Acad. Sci. U. S. A 87 (1990) 1777–1781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [221].Hoa S, Hudson M, Troyanov Y, Proudman S, Walker J, Stevens W, et al. , Single-specificity anti-Ku antibodies in an international cohort of 2140 systemic sclerosis subjects: clinical associations, Medicine (Baltim.) 95 (2016), e4713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [222].Morgenroth R, Reichardt C, Steffen J, Busse S, Frank R, Heidecke H, et al. , Autoantibody formation and mapping of immunogenic epitopes against cold-shock-protein YB-1 in cancer patients and healthy controls, Cancers 12 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [223].Braunschweig D, Krakowiak P, Duncanson P, Boyce R, Hansen RL, Ashwood P, et al. , Autism-specific maternal autoantibodies recognize critical proteins in developing brain, Transl. Psychiatry 3 (2013) e277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [224].van Beers-Tas MH, Marotta A, Boers M, Maksymowych WP, van Schaardenburg D, A prospective cohort study of 14-3-3eta in ACPA and/or RF-positive patients with arthralgia, Arthritis Res. Ther 18 (2016) 76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [225].Chakravarti R, Gupta K, Swain M, Willard B, Scholtz J, Svensson LG, et al. , 14-3-3 in Thoracic Aortic Aneurysms: Identification of a Novel Autoantigen in Large Vessel Vasculitis, vol. 67, Arthritis & rheumatology, Hoboken, NJ: ), 2015, pp. 1913–1921. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.