

Abstract
Background:
Lipoprotein-related traits have been consistently identified as risk factors for atherosclerotic cardiovascular disease, largely on the basis of studies of coronary artery disease (CAD). The relative contributions of specific lipoproteins to risk of peripheral artery disease (PAD) have not been well-defined. We leveraged large-scale genetic association data to investigate effects of circulating lipoprotein-related traits on PAD risk.
Methods:
Genome-wide association study summary statistics for circulating lipoprotein-related traits were used in the MR Bayesian model averaging framework to prioritize the most likely causal major lipoprotein and subfraction risk factors for PAD and CAD. MR was used to estimate the effect of ApoB-lowering on PAD risk using gene regions proxying lipid-lowering drug targets. Genes relevant to prioritized lipoprotein subfractions were identified using transcriptome-wide association studies.
Results:
ApoB was identified as the most likely causal lipoprotein-related risk factor for both PAD (marginal inclusion probability [MIP] 0.86, p = 0.003) and CAD (MIP 0.92, p = 0.005). Genetic proxies for ApoB (apolipoprotein B)-lowering medications were associated with reduced risk of both PAD (OR 0.87 per 1 standard deviation decrease in ApoB, 95% CI 0.84 to 0.91, p = 9 × 10−10) and CAD (OR 0.66, 95% CI 0.63 to 0.69, p = 4 × 10−73), with a stronger predicted effect of ApoB-lowering on CAD (ratio of effects 3.09, 95% CI 2.29 to 4.60, p < 1 × 10−6). Extra-small-VLDL particle concentration (XS.VLDL.P) was identified as the most likely subfraction associated with PAD risk (MIP 0.91, p = 2.3 × 10−4), while large-LDL particle concentration (L.LDL.P) was the most likely subfraction associated with CAD risk (MIP 0.95, p = 0.011). Genes associated with XS.VLDL.P and L.LDL.P included canonical ApoB-pathway components, although gene-specific effects were variable. Lipoprotein(a) was associated with increased risk of PAD, independent of ApoB (OR 1.04, 95% CI 1.03 to 1.04, 95% CI 1.0 × 10−33).
Conclusion:
ApoB was prioritized as the major lipoprotein fraction causally responsible for both PAD and CAD risk. However, ApoB-lowering drug targets and ApoB-containing lipoprotein subfractions had diverse associations with ASCVD, and distinct subfraction-associated genes suggest possible differences in the role of lipoproteins in the pathogenesis of PAD and CAD.
Keywords: Lipoproteins, Atherosclerosis, Peripheral Artery Disease, Coronary Artery Disease, Genomics
INTRODUCTION
Atherosclerotic cardiovascular disease (ASCVD) is the most common cause of morbidity and mortality worldwide.1 Most research has focused on ASCVD in the coronary arteries (coronary artery disease, CAD). However, peripheral artery disease (PAD) represents another common and often underrecognized manifestation of ASCVD that is also associated with significant morbidity (eg. pain, tissue-loss, amputation) and mortality, affecting more than 5% of the global adult population.2,3 Dyslipidemia has been a long-established risk factor for ASCVD, with the strongest evidence derived from large studies primarily focused on CAD endpoints. Although low-density lipoprotein cholesterol (LDL-C)-reducing medications like statins are commonly employed in the prevention and treatment of PAD, evidence on the relationship between LDL-C and PAD risk has been inconsistent.4 Observational studies with modest event rates have suggested that components of the atherogenic dyslipidemia profile (elevated levels of triglyceride-rich lipoproteins, small LDL-C, and the ratio of total cholesterol to HDL-cholesterol [HDL-C] along with low concentrations of HDL-C) may be more strongly associated with PAD than CAD, although the relative contribution of the major circulating lipoproteins (LDL-C, HDL-C, triglycerides, apolipoprotein B [ApoB], and apolipoprotein A1 [ApoA1]) and associated lipoprotein subfractions to PAD specifically has remained poorly defined.4–7
Over the past 15 years, genome-wide association studies have identified hundreds of genetic loci associated with ASCVD traits, major lipoproteins, and related subfractions.8–12 The large datasets arising from these studies include genetic associations estimated in hundreds of thousands of participants. An array of analytic methods enables the analysis of these genetic datasets to provide insights into the underlying biology of diseases. Mendelian randomization (MR) uses genetic variants as instrumental variables to infer the effect of an exposure on an outcome, under the assumption that genetic associations with the outcome are mediated via the exposure for selected variants.13 MR has been used to implicate ApoB as an important risk factor for CAD, and to validate the effects of drug targets on disease outcomes.14–19 Integrating genetic data with gene transcription datasets in transcriptome-wide association studies (TWAS) has been used to identify tissue-level gene expression associations with disease (such as between hepatic expression of SORT1 and risk of CAD).20
We aimed to integrate large-scale genetic datasets to 1) prioritize the role of circulating lipoproteins and subfractions on PAD risk; 2) identify genes that may represent novel lipoprotein-pathway targets in the prevention and treatment of PAD; and 3) estimate the effects of current/potential lipid-lowering medications on PAD risk.
METHODS
Data Availability
The data that support the findings of this study are available from the corresponding author upon reasonable request. GWAS summary statistics for PAD are available by application in dbGaP (phs001672). GWAS summary statistics for GLGC lipids, UKB lipids, NMR lipids, and CARDIoGRAMplusC4D CAD are available for download from the IEU Open GWAS Project (https://gwas.mrcieu.ac.uk/).
Ethical Approval
The VA Central Institutional Review Board approved the MVP study protocol.
Study Population and Outcomes
Our primary outcome was PAD. Genetic associations with PAD were derived from a 2019 genome-wide association study by Klarin et al.10 Full summary data are available by application to dbGaP (phs001672). This study included 31,307 PAD cases (24,009 European-ancestry, 5,373 African-ancestry, 1,925 Hispanic-ancestry) and 211,753 controls among participants of the Veterans Affairs Million Veteran Program, which recruited individuals aged 19 from Veterans Affairs Medical Centers across the United States.21 PAD diagnoses were ascertained from electronic health records using International Classification of Diseases (ICD)-9/10 and Current Procedural Terminology (CPT) codes. Genetic associations were performed separately by ancestry groups using logistic regression adjusted for age, sex, and five ancestry-specific genetic principal components, and then combined using an inverse-variance weighted fixed-effects method.
CAD was included as an outcome in our analysis to help contextualize the PAD results, as most of the observational, Mendelian randomization, and randomized control trial data relating to ASCVD have focused on CAD outcomes. Genetic associations with CAD were derived from the CARDIoGRAMplusC4D 1000 Genomes GWAS.11 This is a meta-analysis of 48 studies, including 60,801 CAD cases and 123,504 controls primarily of European ancestries (77%), including a combination incident and prevalent CAD among the cases. CAD case/control status was determined at the individual study level, with CAD cases included based on the presence of myocardial infarction, acute coronary syndrome, chronic stable angina, or coronary stenosis of >50%.
Prioritizing the Role of Major Lipoprotein-related Traits and Lipoprotein Subfractions on PAD
We performed two Mendelian randomization Bayesian model averaging (MR-BMA)14 analyses as illustrated in Figure 1A. MR-BMA is an extension of multivariable MR that applies a Bayesian variable selection method and aims to identify true causal risk factors (rather than the magnitude of effect) by jointly considering correlated exposures (in this case lipoprotein-related traits).14,22,23 Details of the MR-BMA method are available in the Supplemental Methods. First, to identify causal relationships between major lipoprotein-related traits and PAD, our exposures of interest were circulating lipoproteins: low-density lipoprotein cholesterol (LDL-C), high-density lipoprotein cholesterol (HDL-C); their primary constituent apolipoproteins: ApoA1 and ApoB; and triglycerides (TG) (Figure 1A). Second, in order to investigate whether the predicted effect of ApoB-lowering on PAD may be influenced by specific lipoprotein subfractions, we performed a further MR-BMA analysis focusing on ten ApoB-containing lipoprotein subfractions (Figure 1A). For comparison, we also performed these analyses for CAD.
Figure 1: Overview of Risk Factor Prioritization, Drug-Target, and TWAS Analyses.
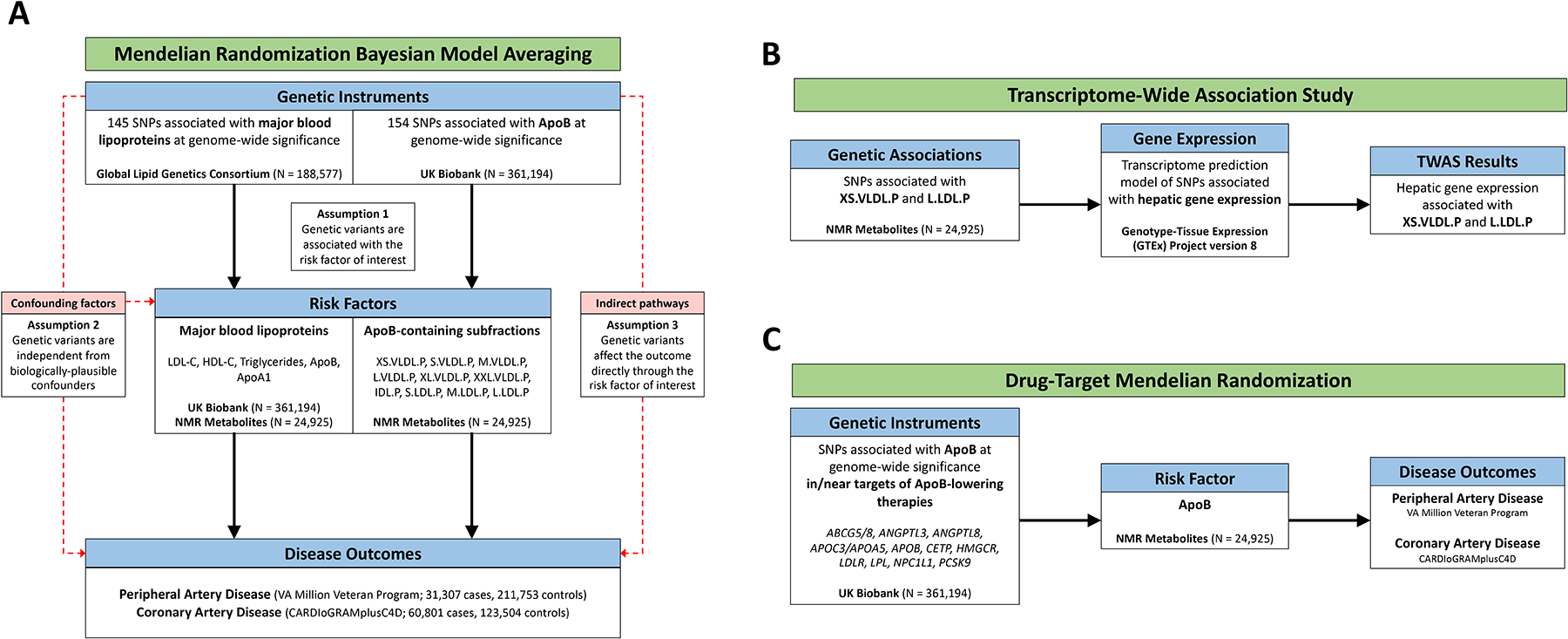
Overview of main analyses. A) Risk factor prioritization was performed using MR-BMA to prioritize the contribution of 1) major lipoproteins, and 2) ApoB-containing subfractions to PAD and CAD risk. The primary MR assumptions are denoted, with red dashed lines representing violations of the MR assumptions. B) A Transcriptome-Wide Association Study integrating gene expression and genetic association data was performed to identify putative genes involved in regulation of the prioritized ApoB-containing subfractions. C) Drug-target MR was performed to identify the effect of genes encoding targets of ApoB-lowering medications on PAD and CAD outcomes.
The instrumental variables consisted of 145 independent (r2 < 0.001 in the 1000 Genomes European-ancestry Reference Panel) genetic variants associated with any major lipoprotein-related trait (total cholesterol, LDL-cholesterol, HDL-cholesterol, or triglycerides) at a genome-wide significance level (p < 5 × 10−8) in the Global Lipids Genetics Consortium, a GWAS comprising 60 individual studies of primarily European-ancestry participants.8 Genetic associations with circulating levels of major lipoprotein-related traits in blood were estimated in the UK Biobank study based on 361,194 European-ancestry participants (http://www.nealelab.is/uk-biobank/). Genetic associations were adjusted for age, sex, and twenty principal components. Genetic associations with lipoprotein subfractions were estimated from a GWAS of circulating lipoproteins and subfractions in 24,925 European-descent participants.9 In this dataset, lipoproteins and subfractions were measured using nuclear magnetic resonance (NMR) spectroscopy. Genetic associations were adjusted for age, sex, and the first 10 genetic principal components. As a replication analysis, we repeated the analysis for major lipoprotein-related traits using genetic associations from the NMR dataset.
Transcriptome-Wide Association Study
After prioritizing the role of lipoprotein subfractions in PAD, we next sought to identify genes associated with those subfractions, which may ultimately serve as therapeutic targets (Figure 1B). To identify genes associated with circulating levels of lipoprotein subfractions, transcriptome-wide association studies (TWAS) were performed using S-PrediXcan.20 This tool enables the integration of tissue-level expression quantitative trait loci (eQTL) datasets with GWAS summary statistics to prioritize genes associated with traits of interest. As the liver plays a critical role in lipoprotein metabolism, we obtained a pre-trained transcriptome prediction model for liver gene expression derived from the Genotype-Tissue Expression (GTEx) Project version 8 (http://predictdb.org). Predicted liver gene expression and GWAS summary statistics for lipoprotein subfractions were then correlated using S-PrediXcan to identify genes significantly associated with circulating levels of lipoprotein subfractions. The significance of differences in the effect of each gene on each outcome were determined by: with p-values derived from the normal distribution. We performed gene ontology enrichment analysis using ShinyGO to identify Gene Ontology Biological Processes significantly associated with the genes prioritized by S-PrediXcan.24 Finally, we performed a combined multivariate and collapsing test to investigate the effect of rare damaging mutations in genes prioritized by the TWAS on risk of PAD among participants of the UK Biobank who underwent whole exome sequencing (Supplemental Methods).25
Genetically-proxied ApoB-lowering and PAD Risk
To predict the impact of ApoB-lowering on PAD and CAD risk, we performed further MR analyses (Figure 1C). We performed gene-based analyses using variants associated with ApoB in gene regions that proxy specific lipid-lowering drugs (licensed or proposed), and polygenic analyses using all such variants. Genetic variants associated with ApoB levels at genome-wide significance (p < 5 × 10−8) were identified from the UK Biobank (http://www.nealelab.is/uk-biobank/) and pruned at r2<0.1 to exclude highly correlated variants. This set of variants was further narrowed into two biologically-informed sets. First, we examined variants located in or near (+/− 200kb) genes encoding previously-identified regulators of ApoB metabolism (ABCG5/8, ANGPTL3, ANGPTL4, ANGPTL8, APOC3/APOA5, APOB, CETP, DGAT, HMGCR, LDLR, LPL, MTTP, NPC1L1, PCSK9, and PPARA), representing the targets of current or proposed therapeutics.16 Next, we examined variants in or near (+/− 200kb) genes associated with extra-small VLDL particle concentration (XS.VLDL.P) in the TWAS analysis (FDR q < 0.05).26 Genetic associations with ApoB were taken from the NMR dataset to avoid winner’s curse and sample overlap.9 MR estimates were obtained from the random effects inverse-variance weighted method performed using the MendelianRandomization package in R, accounting for linkage disequilibrium correlation among variants using the 1000 Genomes Phase 3 European reference panel. The MR-Egger method, which makes different assumptions about the presence of pleiotropy at the cost of decreased statistical power, was performed as a sensitivity analysis when >2 SNPs were present in the genetic instrument.27
Lipoprotein(a) and PAD Risk
To examine the relationship between Lp(a) and PAD, we performed two-sample MR using summary statistics. As genetic instruments for Lp(a), we utilized fifteen common genetic variants which are conditionally independent predictors of Lp(a), had been previously shown to explain >40% of the variance in circulating Lp(a) levels, and were present in GWAS summary statistics for both PAD and CAD. We obtained genetic association effect estimates from a prior analysis.28 We performed inverse-variance weighted MR, considering PAD as the primary outcome, with CAD presented for comparison. We also performed multivariable MR, accounting for the associations between these genetic variants and ApoB in the UK Biobank biomarker GWAS (http://www.nealelab.is/uk-biobank/).
Statistical Analysis
For the main MR-BMA analyses of major lipoprotein-related traits and PAD, FDR correction was performed to account for multiple testing, with FDR-corrected q < 0.05 set as the predetermined significance threshold.15 For the MR-BMA analysis of lipoprotein subfractions, the Nyholt procedure of effective tests was used to account for the strong correlation among the subfractions, with a multiple testing adjusted p-value of 0.05 set as the significance threshold.29 For the drug target MR analysis, p < 0.05 was the predetermined significance threshold. For the TWAS and gene ontology enrichment analyses, the false-discovery rate was used to account for multiple testing, with FDR-corrected p < 0.05 set as the predetermined significance threshold. To compare the influence of lipoprotein risk factors on PAD vs. CAD we calculated the ratio of effects (log-odds), with 95% confidence intervals obtained by bootstrap resampling. All statistical analyses were performed using R version 4.0.3 (R Foundation for Statistical Computing (https://www.R-project.org/). This study is reported in accordance with the STROBE guidelines for reporting observational studies.30
RESULTS
Prioritizing the Role of Major Lipoprotein-related Traits and Lipoprotein Subfractions on PAD
In the MR-BMA analysis for major lipoprotein-related traits, ApoB was the top-ranked risk factor for PAD (marginal inclusion probability 0.86, p = 0.003). (Table 1; Tables I–III in the Supplement). In the replication analysis, where genetic association estimates for the five major lipoprotein-related traits were derived from the NMR metabolite GWAS, ApoB was again the top-ranked risk factor for PAD with a marginal inclusion probability of 0.68 (p = 0.001) (Tables IV–VI in the Supplement). Similarly, ApoB was identified as the prioritized risk factor for CAD in the primary (marginal inclusion probability 0.92, p = 0.005) (Table 1) and replication (marginal inclusion probability 0.80, p = 0.004) analyses, in keeping with the previously established role of ApoB in CAD (Tables VII–VIII in the Supplement)15,16. These results provide strong, consistent support for the role of ApoB as the primary lipoprotein risk factor for PAD and CAD.
Table 1:
Prioritization of Causal Risk Factors Amongst Major Lipoproteins
| Peripheral Artery Disease | Marginal Inclusion Probability | Uncorrected p-value |
FDR-corrected p-value |
|---|---|---|---|
| ApoB | 0.856 | 0.003 | 0.015 |
| ApoA1 | 0.529 | 0.025 | 0.052 |
| HDL-C | 0.467 | 0.031 | 0.052 |
| LDL-C | 0.461 | 0.168 | 0.21 |
| Triglycerides | 0.238 | 0.386 | 0.386 |
| Coronary Artery Disease | |||
| ApoB | 0.922 | 0.005 | 0.027 |
| HDL-C | 0.457 | 0.022 | 0.054 |
| LDL-C | 0.33 | 0.746 | 0.746 |
| ApoA1 | 0.284 | 0.123 | 0.206 |
| Triglycerides | 0.152 | 0.58 | 0.726 |
Ranking of most likely causal exposures amongst major lipoproteins for peripheral artery disease and coronary artery disease prioritized using multivariable Mendelian randomization in the Mendelian randomization Bayesian Model Averaging (MR-BMA) framework. False discovery rate (FDR) corrected p-values are derived in an empirical permutation procedure. LDL-C: low-density lipoprotein cholesterol, HDL-C: high-density lipoprotein cholesterol, ApoA1: apolipoprotein A1, ApoB: apolipoprotein B.
In the MR-BMA analysis for lipoprotein subfractions, extra-small VLDL particle concentration (XS.VLDL.P) was prioritized as the primary ApoB-containing risk factor for PAD (marginal inclusion probability 0.91, p = 2.3 × 10−4) (Table 2; Table IX in the Supplement). In contrast, large LDL particle concentration (L.LDL.P) was prioritized as the primary ApoB-containing risk factor for CAD (marginal inclusion probability 0.95, p = 0.011) (Table 2; Table X in the Supplement).
Table 2:
Prioritization of Causal Risk Factors Amongst Apolipoprotein B-containing Lipid Subfractions
| Peripheral Artery Disease | Marginal Inclusion Probability | Uncorrected p-value |
FDR-corrected p-value |
|---|---|---|---|
| XS.VLDL.P | 0.912 | 0.0023 | 0.0012 |
| IDL.P | 0.078 | 0.963 | 1 |
| L.LDL.P | 0.023 | 0.999 | 1 |
| S.VLDL.P | 0.022 | 0.901 | 1 |
| M.LDL.P | 0.021 | 0.999 | 1 |
| XL.VLDL.P | 0.021 | 0.996 | 1 |
| S.LDL.P | 0.02 | 1 | 1 |
| XXL.VLDL.P | 0.017 | 1 | 1 |
| L.VLDL.P | 0.014 | 0.999 | 1 |
| M.VLDL.P | 0.014 | 0.997 | 1 |
| Coronary Artery Disease | |||
| L.LDL.P | 0.613 | 0.008 | 0.04 |
| M.LDL.P | 0.382 | 0.554 | 1 |
| IDL.P | 0.071 | 0.614 | 1 |
| S.LDL.P | 0.045 | 0.981 | 1 |
| XS.VLDL.P | 0.036 | 0.294 | 1 |
| S.VLDL.P | 0.027 | 0.765 | 1 |
| M.VLDL.P | 0.023 | 0.9 | 1 |
| L.VLDL.P | 0.016 | 0.948 | 1 |
| XL.VLDL.P | 0.015 | 0.986 | 1 |
| XXL.VLDL.P | 0.014 | 0.95 | 1 |
Ranking of most likely causal exposures amongst apolipoprotein B-containing lipid subfractions for peripheral artery disease and coronary artery disease prioritized using multivariable Mendelian randomization in the Mendelian randomization Bayesian Model Averaging (MR-BMA) framework. False discovery rate (FDR) corrected p-values are derived using the Nyholt procedure of effective tests to account for the strong correlation among the subfractions. XXL.VLDL.P: Extra-extra large very-large density lipoprotein particles, XL.VLDL.P: Extra large very-large density lipoprotein particles, L.VLDL.P: Large very-large density lipoprotein particles, M.VLDL.P: Medium very-large density lipoprotein particles, S.VLDL.P: Small very-large density lipoprotein particles, XS.VLDL.P: Extra small very-large density lipoprotein particles, L.LDL.P: Large large-density lipoprotein particles, M.LDL.P: Medium large-density lipoprotein particles, S.LDL.P: Small large-density lipoprotein particles, IDL.P: Intermediate-density lipoprotein particles.
Identification of Genes Associated with ApoB-containing Lipoprotein Subfractions
Having prioritized XS.VLDL.P and L.LDL.P as important ApoB-containing lipoprotein subfractions for PAD and CAD respectively, we explored whether specific genes may influence the circulating levels of these subfractions. Given the key role of the liver in lipoprotein metabolism, we integrated hepatic gene expression data from the Genotype-Tissue Expression (GTEx) Project with the XS.VLDL.P and L.LDL.P GWAS summary statistics to identify genes associated with circulating levels of each ApoB-containing lipoprotein subfraction.
Using TWAS, we identified 31 genes associated with XS.VLDL.P and 23 genes associated with L.LDL.P, for a total of 40 unique genes (FDR <0.05 for either subfraction) (Tables XI–XII in the Supplement). Of these 40 genes, 17 were uniquely associated with XS.VLDL.P levels, 9 were uniquely associated with L.LDL.P levels, and 14 were associated with circulating levels of both subfractions (Figure 2A–B). As expected, genes associated with these lipoprotein subfractions were significantly enriched for membership in cholesterol metabolism and related pathways (Table XIII in the Supplement). Among the genes associated with both subfractions were several canonical genes involved in lipoprotein metabolism, including PCSK9, ABCG8, LIPC, and APOA5. The 17 genes uniquely associated with XS.LDL.P levels were enriched for clusters of biological processes involving triglyceride-rich lipoprotein metabolism (Table XIV in the Supplement), while the 9 genes uniquely associated with L.LDL.P levels were not significantly enriched for specific biological processes.
Figure 2: Genes Associated with Circulating Levels of XS.VLDL.P and L.LDL.P Lipoprotein Subfractions.
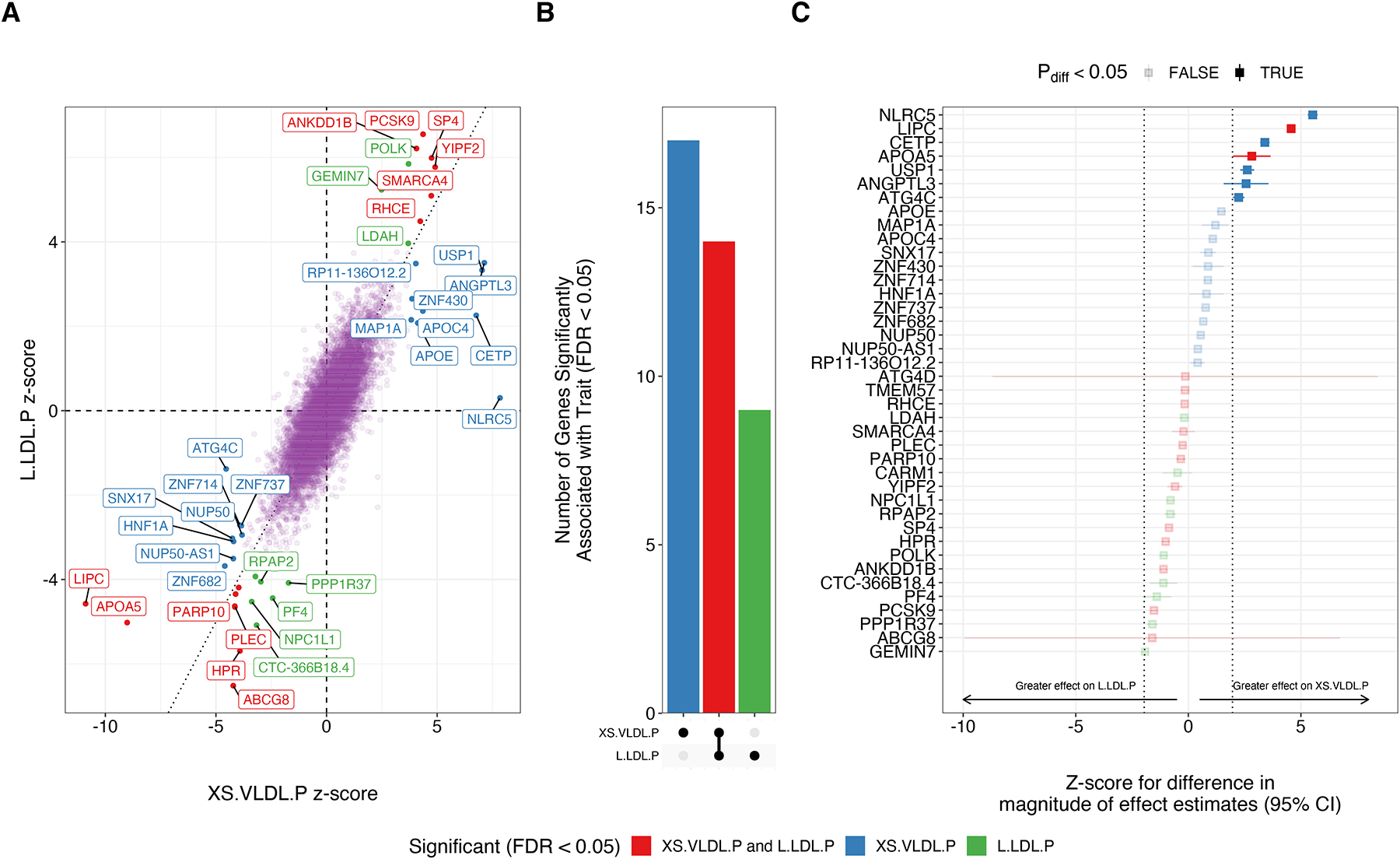
Transcriptome-wide association studies were performed integrating liver gene-expression data from GTEX v8 with GWAS summary statistics for XS.VLDL.P and L.LDL.P to identify genes associated with circulating levels of each lipoprotein subfraction. A) Genes significantly (FDR < 0.05) associated with either subfraction are labeled, and colors represent the subfraction associations. B) The bar plot depicts the number of unique and shared genes between the two subfractions. C) The forest plot depicts the z-score for the difference in the effect magnitude for each gene on each subfraction (). Dotted lines represent z-scores of +/− 1.96, with point estimates outside this range representing significant (pdiff < 0.05) differential effects. Errors represent 95% confidence intervals for the z-score.
Across the genome, there was overall strong correlation between the estimated effects of gene expression on circulating XS.VLDL.P and L.LDL.P levels (Pearson correlation = 0.80, p < 2.2 × 10−16). However, among the 40 genes significantly associated with either lipoprotein subfraction, we identified potentially heterogenous effects. For example, NLRC5, LIPC, CETP, APOA5, USP1, ANGPTL3, and ATGC4 expression was predicted to more strongly impact circulating XS.VLDL.P levels in comparison to L.LDL.P (Figure 2C).
To investigate the impact of these genes on PAD risk, we examined whether damaging mutations in XS.VLDL.P-associated genes might influence PAD risk among UK Biobank participants. We identified rare loss-of-function variants in 29 of 31 XS.VLDL.P-associated genes among 154,584 UK Biobank participants (1,668 PAD cases and 152,916 controls). After accounting for multiple testing, only damaging variants in SP4 were associated with prevalent PAD (FDR q = 0.049) (Table XV in the Supplement).
Genetically-predicted ApoB-lowering and PAD Risk
Because Mendelian randomization has previously been utilized to predict the impact of current and proposed ApoB-lowering therapies on CAD risk, we sought to explore the effect of these treatments on PAD risk.16 We first performed polygenic and gene-based MR analyses to determine whether ApoB-associated genetic variants located within/near genes encoding these therapeutic targets were associated with risk of PAD. In polygenic analyses, genetically-proxied ApoB-lowering was associated with reduced risk of PAD (OR 0.87 per 1 standard deviation reduction in ApoB, 95% CI 0.84 to 0.91, p = 9 × 10−9) (Figure 3A). As a comparison, the association of genetically-proxied ApoB-lowering with CAD risk using the same genetic variants was greater (OR 0.66 per 1 standard deviation decrease in circulating ApoB, 95% CI 0.63 to 0.69, p = 4 × 10−73; ratio of effects 3.09, 95% CI 2.29 to 4.60, p < 1 × 10−6). Associations were also significant using the MR-Egger method (PAD OR 0.91, 95% CI 0.86 to 0.98, p = 0.009; CAD OR 0.70, 95% CI 0.65 to 0.76, p = 8 × 10−20).
Figure 3: Associations Between Genetically-Predicted ApoB levels and PAD.

Estimates represent associations between genetically-proxied ApoB and PAD or CAD risk, scaled to the change in disease risk per 1 standard deviation (1-SD) decrease in ApoB. Genetic variants used to proxy reductions in ApoB only included those located near/within genes important in ApoB metabolism. A) Polygenic analysis including all such variants. OR = odds ratio, CI = confidence interval. B) Gene-based analyses considering variants for each gene region. The dotted line with slope 1 represents the scenario where the association of genetically-proxied ApoB with disease risk is equal for both CAD and PAD. C) Relative effects of each gene region on CAD vs. PAD as determined by the ratio of effects. Error bars represent 95% confidence intervals.
Next, we compared the associations between genetically-proxied ApoB-lowering and ASCVD outcomes in gene-based analyses, identifying potential heterogenous effects on PAD and CAD risk. We identified protective effects on CAD and/or PAD for 11 of 12 ApoB target genes (Figure 3B; Table XVI in the Supplement). While several associations did not achieve statistical significance for PAD specifically, associations were generally in the risk decreasing direction, with the exception of HMGCR locus which trended toward increased PAD risk (although 95% confidence intervals did not exclude a small protective effect on PAD). Consistent with the overall polygenic analysis, genetically-proxied ApoB-lowering at the ABCG5, PCSK9, APOB, LDLR, and ANGPTL8 loci had significantly greater protective effects on CAD in comparison to PAD (ratio of effect estimates > 1, FDR < 0.05) (Figure 3C). Results were similar, though with wider confidence intervals, when considering the MR-Egger method (Table XVI in the Supplement).
We also performed polygenic and gene-specific MR analyses to explore whether XS.VLDL.P-associated genes identified in the TWAS analysis were associated with PAD risk. In the polygenic analysis, ApoB-lowering proxied by genetic variants located within/near XS.VLDL.P-associated genes was associated with reduced risk of PAD (OR 0.89 per 1 SD reduction in ApoB, 95% CI 0.86 to 0.92, p = 3 × 10−11) (Figure 4; Table XVII in the Supplement). In gene-specific analyses, ApoB-lowering at the CETP, NLRC5, and YIPF2 loci were significantly associated with decreased PAD risk, while ApoB-lowering at the ANKDD1B locus was associated with increased PAD risk (Figure 4). Confidence intervals were wider with the MR-Egger method (Table XVII in the Supplement), although the association remained significant at the polygenic level.
Figure 4: Associations between XS.VLDL.P-associated Genes and PAD.

Polygenic and gene-specific MR were performed to estimate the association between the ApoB-lowering effect of XS.VLDL.P-associated genes and PAD. Gene-specific MR examining the impact of ApoB-associated variants in/near (+/− 200kb) each XS.VLDL.P-associated gene, with Polygenic Targets denoting the aggregate impact of variants in/near these genes. ORs represent the change in disease risk per 1 standard deviation (1-SD) decrease in ApoB.
Circulating Lipoprotein(a) and PAD Risk
Finally, as prior studies have identified an association between genetically-predicted Lp(a) and CAD,28,31 we investigated the relationship between circulating Lp(a) and PAD risk. Unlike the other major lipoprotein-related traits which are genetically-correlated and share a complex polygenic architecture, circulating Lp(a) levels are primarily regulated by genetic variants acting specifically at the LPA locus on chromosome 6.32 Consistent with this architecture, we did not identify significant genetic correlation between Lp(a) and other lipoprotein-related traits (Table XVIII in the Supplement). In MR analyses, increases in genetically-predicted Lp(a) was associated with increased risk of PAD (OR 1.04 per 10 mg/dL increase in Lp(a), 95% CI 1.03 to 1.04, p = 3 × 10−32), and the effect appeared linear across the range of Lp(a) (Table XIX in the Supplement; Figure I in the Supplement). This association was not attenuated in multivariable MR accounting for the associations of these genetic variants with ApoB (OR 1.04, 95% CI 1.03 to 1.04, 95% CI 1.0 × 10−33). For comparison, genetically-predicted Lp(a) was also associated with increased risk of CAD (OR 1.06, 95% CI 1.05 to 1.06, p = 1 × 10−94). Genetically-predicted Lp(a) was more strongly associated with CAD in comparison to PAD (ratio of effects 1.62, 95% CI 1.35–1.98, p < 1 × 10−6).
DISCUSSION
We integrated several large genetic datasets and an array of statistical genetics, molecular epidemiology, and bioinformatic techniques to uncover novel causal relationships between circulating lipoprotein-related traits and PAD, and compared these findings with CAD. First, we identified ApoB as the primary major circulating lipoprotein-related trait responsible for risk of PAD, similar to CAD. Next, we prioritized XS.VLDL.P as the ApoB-associated subfraction most strongly associated with PAD risk, in contrast to CAD where L.LDL.P was the most strongly associated lipoprotein subfraction. We identified genes involved in the regulation of important ApoB-containing lipoprotein subfractions, which may represent directed targets for novel PAD prevention and treatment strategies. We explored the impact of ApoB-lowering on PAD and uncovered the potential for the XS.VLDL.P pathway to be targeted to reduce PAD risk. Finally, we identified an association between Lp(a) and PAD, independent of ApoB.
These results highlight similarities and differences in the roles of circulating lipoproteins for PAD and CAD. Our primary analysis identified ApoB as the major lipoprotein-related trait responsible for both PAD and CAD risk. This finding is consistent with a recent meta-analysis of 22 studies (including 1,892 PAD cases and 30,937 controls) which found significantly higher ApoB levels among PAD cases compared to controls.33 A nested case-control study within the Physicians’ Health Study (PHS) similarly identified baseline ApoB levels (in addition to several other lipid fractions) as a significant predictor of incident PAD.34 In contrast, a large observational study including 31,657 participants of 5 prospective Finnish cohorts did not detect an association between ApoB levels and incident PAD, but may have been limited by a low incidence of PAD (498 cases) and by defining PAD based on hospitalization codes.35 Similarly, while the Women’s Health Study (WHS) did not identify an association between baseline ApoB and incident PAD,5 differences in PAD case definitions, ascertainment, and demographics (incident PAD in WHS vs. prevalent in MVP; women in WHS vs. predominately men in MVP and PHS), may account for these differences. Our prioritization of ApoB as the most important lipoprotein-related risk factor for CAD is consistent with recent MR studies establishing ApoB as the primary risk factor for CAD.15,16 We also identified a modest but significant association between Lp(a) and PAD that was independent of ApoB, consistent with prior MR studies linking Lp(a) to CAD.28,36 In the setting of strong epidemiologic and genetic correlation between CAD and PAD, it is not surprising that these two manifestations of ASCVD share ApoB and Lp(a) as common risk factors. However, we identified a stronger effect of ApoB on CAD than PAD, which has implications for risk stratification and treatment.
ApoB-containing particles exist on a spectrum of varying sizes, densities, and particle compositions, and identification of specific subfractions that contribute to different forms of atherosclerotic cardiovascular disease may have implications for pathophysiology, molecular mechanisms, risk stratification, and treatment.37 Variability in the distribution of ApoB within lipoprotein subfractions may contribute to differential risk of PAD compared to CAD. Indeed, clinical observations have suggested that Type III hyperlipoproteinemia (familial dysbetalipoproteinemia), a disorder specifically associated with VLDL remnant particles, may be a greater risk factor for PAD than for CAD.37 In our genetic analyses of ApoB-containing lipoprotein subfractions using MR-BMA, extra-small VLDL particle concentration (XS.VLDL.P) was the primary ApoB-containing subfraction contributing to PAD risk. In contrast, large LDL particle concentration (L.LDL.P) was the primary ApoB-containing subfraction contributing to CAD risk. These results are consistent with a recent observational study exploring the effect of circulating lipoproteins and metabolites on incident PAD and CAD among 31,657 participants of five prospective Finnish cohorts, which found a strong association between XS.VLDL.P and incident PAD, with no significant association between L.LDLP and incident PAD.35 The primary effect of XS.VLDL.P on PAD implicates an important role for triglyceride-rich lipoproteins and remnant particles in the pathogenesis of PAD. This stands in contrast to the effect of L.LDL.P on CAD, which suggests that LDL-associated lipoprotein fractions may play a more important role in the pathogenesis of coronary artery disease.
Epidemiologically, small (rather than large) LDL particles have been traditionally associated with CAD risk, serving as a marker of atherogenic dyslipidemia, particularly in the context of metabolic syndrome.38 Various mechanisms have been proposed to explain this relationship, including increased ability of small LDL particles to penetrate the arterial wall and increased susceptibility to oxidation. While a recent systematic review and meta-analysis found both small dense LDL particles and concentration were associated with risk of CAD across 21 studies representing more than 30,000 subjects, there was substantial heterogeneity across studies, and observational findings more broadly may be limited by residual environmental confounding.39 In contrast, two recent observational studies have suggested that remnant cholesterol is more strongly predictive of ASCVD events than other lipoprotein-related traits.40,41 The prioritization of L.LDL.P does not preclude the possibility that small LDL or remnant particles also cause ASCVD. Whether small LDL and/or triglyceride-rich lipoproteins represent causal risk factors rather than a consequence of other metabolic derangements, and the specific mechanisms by which different ApoB-containing subfractions contribute to ASCVD risk requires further study. Intriguingly, a prior MR analysis proposed a mechanism by which remnant particles causally increase inflammation (as measured by C-reactive protein level), whereas no inflammatory effect was detected in the setting of genetically-proxied elevations in LDL cholesterol.42
While both PAD and CAD are manifestations of ASCVD, there are pathophysiologic differences between the two diseases that may provide a basis for targeted treatment strategies.43 Observational and genetic studies have suggested that the influence common cardiovascular risk factors may vary across ASCVD outcomes, for example, with smoking more strongly associated with PAD, and blood pressure more strongly associate with CAD, among others.44–47 Similarly, although ApoB represents a common lipoprotein risk factor for PAD and CAD, we identified differences in predicted response to ApoB-lowering treatment, underscoring potential differences in the role of circulating lipoproteins on these ASCVD outcomes. We demonstrate that while ApoB-lowering is expected to have favorable effects on both PAD and CAD risk, the relative benefit is expected to be significantly greater for CAD risk-reduction in comparison to PAD risk-reduction. Although large randomized controlled trials, genetic studies, and observational evidence have highlighted the importance of LDL-cholesterol and ApoB-lowering in reducing ASCVD outcomes overall, these studies have primarily focused on major adverse cardiovascular events and CAD outcomes.7,48,49 Our results reveal that the ApoB-lowering effect of several clinically approved and clinical trial-stage drug targets is predicted to differ between PAD and CAD, a finding which may have implications for both drug discovery and treatment paradigms. Although we identified varied effects across ApoB-lowering targets, these results should not at this point be used to guide treatment decisions, and MR and clinical trial estimates of treatment effects may vary.50 While overall these results support guideline recommendations for the use of ApoB-lowering medications to reduce PAD risk, our results also argue for PAD-specific outcomes to be measured in cardiovascular outcomes trials, as the absence of a treatment effect for CAD (or a combined endpoint) may not exclude PAD-specific effects.1,49 For example, the CETP-inhibitor torcetrapib was associated with a substantial decrease in the incidence of PAD in a large, placebo-controlled Phase III clinical trial, although was overall associated with increased risk of cardiovascular morbidity and mortality.51 The ongoing PROMINENT (Pemafibrate to Reduce Cardiovascular Outcomes by Reducing Triglycerides in Patients with Diabetes) Study will evaluate the impact of triglyceride-rich lipoprotein-lowering on major cardiovascular events with adjudicated PAD events as a secondary endpoint of the trial.52
Finally, our polygenic and gene-based drug target analyses highlighted potential targets for directed treatment strategies. As lipid-lowering medications induce specific changes in the circulating lipoprotein profile, our results suggest that specific drug-target identification may play an important role in identifying PAD-focused treatments.53 For example, genes associated with circulating XS.VLDL.P were enriched in pathways related to triglyceride-rich lipoprotein metabolism. Although our polygenic and gene-specific MR analyses suggested that both currently available and proposed ApoB-lowering therapies would be expected to reduce PAD risk, we used MR to further highlight the XS.VLDL.P pathway as a potentially novel therapeutic target. Whether these genes and pathways represent pharmacologic targets that ultimately impact PAD outcomes warrants further study.
Limitations
This study should be interpreted within the context of its limitations. First, this study focused on prevalent PAD outcomes ascertained from electronic health records. The effect of lipoprotein-related traits may vary across specific incident PAD outcomes including intermittent claudication, rest pain, tissue-loss, and amputation. Second, PAD outcomes were studied among primarily male participants of the Veterans Affairs Million Veteran Program, and although participants were of diverse ancestries, further studies among other populations are warranted to improve generalizability of these findings. Third, MR effect estimates reflect lifelong genetic exposures, and may not accurately reflect the magnitude of benefit of shorter-term pharmacologic interventions.54 Similarly, although we did not detect pleiotropic effects in MR-Egger analyses, we cannot exclude the possibility that genetic variants located within targets of lipid-lowering therapies may influence other cardiometabolic traits. Thus, our drug-target MR findings should not be used to guide clinical decisions regarding lipid-lowering therapies at this stage. Fourth, when correlated exposures exist within a common pathway, the MR-BMA method identifies the most proximate risk factor to the outcome. Circulating levels of each lipid fraction are composed of several subcomponents (for example, ApoB, triglycerides, and cholesterol are all found in VLDL, IDL, and LDL compartments and subfractions). Prioritization of ApoB over LDL-C for PAD and CAD risk does not nullify LDL-C as a causal risk factor, but indicates that effects of lipid-lowering therapies are likely to be proportional to the change in ApoB rather than LDL-C. Detailed MR analysis of other lipid subfractions may prioritize additional PAD risk factors and therapeutic targets. Finally, larger population-scale GWAS of circulating lipoprotein-related traits and subfractions may enable the development of more robust genetic instruments, which may provide additional insights regarding the relationships between circulating lipoproteins-related traits and ASCVD.
Overall, this analysis of large genetic datasets identified ApoB as the primary causal lipoprotein-related risk factor for PAD. Diverse effects of ApoB-lowering drug targets and ApoB-containing lipoprotein subfractions on PAD in comparison to CAD suggests possible biologic differences in the pathogenesis of these diseases, with gene expression analyses revealing potential targets for novel PAD therapies.
Supplementary Material
CLINICAL PERSPECTIVE.
What is new?
Apolipoprotein B was identified as the most likely casual lipoprotein-related risk factor for PAD, and ApoB-lowering medications were predicted to reduce risk of PAD.
The effect of ApoB (and ApoB-lowering medications) was predicted to be approximately 3 times greater for CAD than PAD.
XS.VLDL.P was prioritized as the most likely ApoB-containing subfraction associated with PAD risk, while L.LDL.P was prioritized as the most likely subfraction associated with CAD.
What are the clinical implications?
Interventions targeting ApoB are likely to reduce risk of PAD.
Future studies may be warranted to test whether interventions targeting specific lipoprotein-related subfractions may reduce risk of PAD and CAD.
ACKNOWLEDGEMENTS
The authors thank the participants of the VA Million Veterans Program, UK Biobank, CARDIoGRAMplusC4D, and NMR metabolomics studies.
SOURCES OF FUNDING
This research is based on data from the MVP, Office of Research and Development, Veterans Health Administration and was supported by award no. MVP003/MVP028 (I01-BX003362). A list of MVP collaborators is included in the supplement. This work was supported by US Department of Veterans Affairs grants IK2-CX001780 (SMD). This publication does not represent the views of the Department of Veterans Affairs or the United States government. VZ is supported by UK Dementia Research Institute at Imperial College London, which is funded by the Medical Research Council, Alzheimer’s Society and Alzheimer’s Research UK (MC_PC_17114). AWA is supported by NIH K23 HL151871. DG is supported by the British Heart Foundation Centre of Research Excellence (RE/18/4/34215) at Imperial College London and a National Institute for Health Research Clinical Lectureship at St. George’s, University of London (CL-2020-16-001). SB is supported by a Sir Henry Dale Fellowship jointly funded by the Wellcome Trust and the Royal Society (Grant Number 204623/Z/16/Z).
DISCLOSURES
SMD receives research support to his institution from RenalytixAI and reports consulting fees from Calico Labs, all outside the current work. DG is employed part-time by Novo Nordisk. DJR serves on scientific advisory boards for Alnylam, Novartis, Pfizer and Verve and is a co-founder of Staten Biotechnology. AWA reports consulting fees from OptumCare outside the current work.
Non-Standard Abbreviations and Acronyms
- ApoA1
apolipoprotein A1
- ApoB
apolipoprotein B
- ASCVD
atherosclerotic cardiovascular disease
- CAD
coronary artery disease
- eQTL
expression quantitative trait loci
- GTEx
Genotype-Tissue Expression Project
- GWAS
genome wide association study
- HDL
high density lipoprotein
- IDL
intermediate density lipoprotein
- L.LDL.P
large low density lipoprotein particle concentration
- LDL
low density lipoprotein
- Lp(a)
lipoprotein(a)
- MIP
marginal inclusion probability
- MR
Mendelian randomization
- MR-BMA
Mendelian randomization Bayesian Model Averaging
- MVP
Veterans Affairs Million Veteran Program
- NMR
nuclear magnetic resonance
- PAD
peripheral artery disease
- PHS
Physicians’ Health Study
- TG
triglycerides
- TWAS
transcriptome-wide association study
- VLDL
very low density lipoprotein
- WHS
Women’s Health Study
- XS.VLDL.P
extra-small very low density lipoprotein particle concentration
Footnotes
SUPPLEMENTAL MATERIAL
Supplemental Methods
Supplemental Figure I
Supplemental Excel File I (Supplemental Tables 1–19)
References 55–60
REFERENCES
- 1.Gerhard-Herman MD, Gornik HL, Barrett C, Barshes NR, Corriere MA, Drachman DE, Fleisher LA, Fowkes FGR, Hamburg NM, Kinlay S, et al. 2016 Lower Extremity Peripheral Arterial Disease Guidelines. Journal of the American College of Cardiology. 2017;69:e71–e126. [DOI] [PubMed] [Google Scholar]
- 2.Criqui MH, Aboyans V. Epidemiology of Peripheral Artery Disease. Circulation Research. 2015;116:1509–1526. [DOI] [PubMed] [Google Scholar]
- 3.Song P, Rudan D, Zhu Y, Fowkes FJI, Rahimi K, Fowkes FGR, Rudan I. Global, regional, and national prevalence and risk factors for peripheral artery disease in 2015: an updated systematic review and analysis. The Lancet Global Health. 2019;7:e1020–e1030. [DOI] [PubMed] [Google Scholar]
- 4.Aday AW, Everett BM. Dyslipidemia Profiles in Patients with Peripheral Artery Disease. Current Cardiology Reports. 2019;21:42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Aday AW, Lawler PR, Cook NR, Ridker PM, Mora S, Pradhan AD. Lipoprotein Particle Profiles, Standard Lipids, and Peripheral Artery Disease Incidence: Prospective Data From the Women’s Health Study. Circulation. 2018;138:2330–2341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Duran EK, Aday AW, Cook NR, Buring JE, Ridker PM, Pradhan AD. Triglyceride-Rich Lipoprotein Cholesterol, Small Dense LDL Cholesterol, and Incident Cardiovascular Disease. Journal of the American College of Cardiology. 2020;75:2122–2135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Aung PP, Maxwell HG, Jepson RG, Price JF, Leng GC. Lipid-lowering for peripheral arterial disease of the lower limb. Cochrane Database of Systematic Reviews. 2007;2007:CD000123. doi: 10.1002/14651858.CD000123.pub2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Willer CJ, Schmidt EM, Sengupta S, Peloso GM, Gustafsson S, Kanoni S, Ganna A, Chen J, Buchkovich ML, Mora S, et al. Discovery and refinement of loci associated with lipid levels. Nature Genetics. 2013;45:1274–1285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kettunen J, Demirkan A, Würtz P, Draisma HHM, Haller T, Rawal R, Vaarhorst A, Kangas AJ, Lyytikäinen L-P, Pirinen M, et al. Genome-wide study for circulating metabolites identifies 62 loci and reveals novel systemic effects of LPA. Nat Commun. 2016;7:11122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Klarin D, Lynch J, Aragam K, Chaffin M, Assimes TL, Huang J, Lee KM, Shao Q, Huffman JE, Natarajan P, et al. Genome-wide association study of peripheral artery disease in the Million Veteran Program. Nature Medicine. 2019;25:1274–1279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Nikpay M, Goel A, Won HH, Hall LM, Willenborg C, Kanoni S, Saleheen D, Kyriakou T, Nelson CP, CHopewell J, et al. A comprehensive 1000 Genomes-based genome-wide association meta-analysis of coronary artery disease. Nature Genetics. 2015;47:1121–1130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Malik R, Chauhan G, Traylor M, Sargurupremraj M, Okada Y, Mishra A, Rutten-Jacobs L, Giese A-K, van der Laan SW, Gretarsdottir S, et al. Multiancestry genome-wide association study of 520,000 subjects identifies 32 loci associated with stroke and stroke subtypes. Nature Genetics. 2018;50:524–537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Burgess S, Foley CN, Zuber V. Inferring Causal Relationships Between Risk Factors and Outcomes from Genome-Wide Association Study Data. Annual Review of Genomics and Human Genetics. 2018;19:303–327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zuber V, Colijn JM, Klaver C, Burgess S. Selecting likely causal risk factors from high-throughput experiments using multivariable Mendelian randomization. Nature Communications. 2020;11:1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Zuber V, Gill D, Ala-Korpela M, Langenberg C, Butterworth A, Bottolo L, Burgess S. High-throughput multivariable Mendelian randomization analysis prioritizes apolipoprotein B as key lipid risk factor for coronary artery disease. Int J Epidemiol. 2020; November 1;dyaa216. doi: 10.1093/ije/dyaa216. Online ahead of print. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ference BA, Kastelein JJP, Ray KK, Ginsberg HN, Chapman MJ, Packard CJ, Laufs U, Oliver-Williams C, Wood AM, Butterworth AS, et al. Association of Triglyceride-Lowering LPL Variants and LDL-C-Lowering LDLR Variants with Risk of Coronary Heart Disease. JAMA. 2019;321:364–373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gill D, Georgakis MK, Koskeridis F, Jiang L, Feng Q, Wei W-Q, Theodoratou E, Elliott P, Denny JC, Malik R, et al. Use of Genetic Variants Related to Antihypertensive Drugs to Inform on Efficacy and Side Effects. Circulation. 2019;140:270–279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Walker VM, Kehoe PG, Martin RM, Davies NM. Repurposing antihypertensive drugs for the prevention of Alzheimer’s disease: a Mendelian randomization study. International Journal of Epidemiology. 2020;49:1132–1140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Schmidt AF, Finan C, Gordillo-Marañón M, Asselbergs FW, Freitag DF, Patel RS, Tyl B, Chopade S, Faraway R, Zwierzyna M, et al. Genetic drug target validation using Mendelian randomisation. Nature Communications. 2020;11:3255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Barbeira AN, Dickinson SP, Bonazzola R, Zheng J, Wheeler HE, Torres JM, Torstenson ES, Shah KP, Garcia T, Edwards TL, et al. Exploring the phenotypic consequences of tissue specific gene expression variation inferred from GWAS summary statistics. Nature Communications. 2018;9:1825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gaziano JM, Concato J, Brophy M, Fiore L, Pyarajan S, Breeling J, Whitbourne S, Deen J, Shannon C, Humphries D, et al. Million Veteran Program: A mega-biobank to study genetic influences on health and disease. Journal of Clinical Epidemiology. 2016;70:214–223. [DOI] [PubMed] [Google Scholar]
- 22.Sanderson E, Davey Smith G, Windmeijer F, Bowden J. An examination of multivariable Mendelian randomization in the single-sample and two-sample summary data settings. International Journal of Epidemiology. 2019;48:713–727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Burgess S, Thompson SG. Multivariable Mendelian Randomization: The Use of Pleiotropic Genetic Variants to Estimate Causal Effects. American Journal of Epidemiology. 2015;181:251–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ge SX, Jung D, Jung D, Yao R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics. 2020;36:2628–2629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Li B, Leal SM. Methods for Detecting Associations with Rare Variants for Common Diseases: Application to Analysis of Sequence Data. The American Journal of Human Genetics. 2008;83:311–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gill D, Georgakis MK, Walker VM, Schmidt AF, Gkatzionis A, Freitag DF, Finan C, Hingorani AD, Howson JMM, Burgess S, et al. Mendelian randomization for studying the effects of perturbing drug targets. Wellcome Open Research. 2021;6:16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Slob EAW, Burgess S. A comparison of robust Mendelian randomization methods using summary data. Genetic Epidemiology. 2020;44:313–329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Burgess S, Ference BA, Staley JR, Freitag DF, Mason AM, Nielsen SF, Willeit P, Young R, Surendran P, Karthikeyan S, et al. Association of LPA variants with risk of coronary disease and the implications for lipoprotein(a)-lowering therapies: A mendelian randomization analysis. JAMA Cardiology. 2018;3:619–627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Nyholt DR. A simple correction for multiple testing for single-nucleotide polymorphisms in linkage disequilibrium with each other. American journal of human genetics. 2004;74:765–769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Von Elm E, Altman DG, Egger M, Pocock SJ, Gøtzsche PC, Vandenbroucke JP. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: Guidelines for reporting observational studies. PLoS Medicine. 2007;4:1623–1627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Clarke R, Peden JF, Hopewell JC, Kyriakou T, Goel A, Heath SC, Parish S, Barlera S, Franzosi MG, Rust S, et al. Genetic Variants Associated with Lp(a) Lipoprotein Level and Coronary Disease. New England Journal of Medicine. 2009;361:2518–2528. [DOI] [PubMed] [Google Scholar]
- 32.Barlera S, Specchia C, Farrall M, Chiodini BD, Franzosi MG, Rust S, Green F, Nicolis EB, Peden J, Assmann G, et al. Multiple QTL influence the serum Lp(a) concentration: A genome-wide linkage screen in the PROCARDIS study. European Journal of Human Genetics. 2007;15:221–227. [DOI] [PubMed] [Google Scholar]
- 33.Forte F, Calcaterra I, Lupoli R, Orsini RC, Chiurazzi M, Tripaldella M, Iannuzzo G, Di Minno MND. Association of apolipoprotein levels with peripheral arterial disease: a meta-analysis of literature studies. European Journal of Preventive Cardiology. 2020. September 18;zwaa029. doi: 10.1093/eurjpc/zwaa029. Online ahead of print. [DOI] [PubMed] [Google Scholar]
- 34.Ridker PM, Stampfer MJ, Rifai N. Novel risk factors for systemic atherosclerosis: A comparison of C-reactive protein, fibrinogen, homocysteine, lipoprotein(a), and standard cholesterol screening as predictors of peripheral arterial disease. Journal of the American Medical Association. 2001;285:2481–2485. [DOI] [PubMed] [Google Scholar]
- 35.Tikkanen E, Jägerroos V, Rodosthenous R, Holmes MV, Sattar N, Ala-Korpela M, Jousilahti P, Lundqvist A, Perola M, Salomaa V, et al. Metabolic biomarkers for peripheral artery disease compared with coronary artery disease: Lipoprotein and metabolite profiling of 31,657 individuals from five prospective cohorts. medRxiv. 2020;2020.07.24.20158675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Trinder M, Zekavat SM, Uddin MM, Pampana A, Natarajan P. Apolipoprotein B is an insufficient explanation for the risk of coronary disease associated with lipoprotein(a). Cardiovascular Research. 2021;117:1245–1247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Rader DJ, Kathiresan S. Disorders of Lipoprotein Metabolism. In: Jameson JL, Fauci AS, Kasper DL, Hauser SL, Longo DL, Loscalzo J, editors. Harrison’s Principles of Internal Medicine, 20e. New York, NY: McGraw-Hill Education; 2018. [Google Scholar]
- 38.Xiao C, Dash S, Morgantini C, Hegele RA, Lewis GF. Pharmacological Targeting of the Atherogenic Dyslipidemia Complex: The Next Frontier in CVD Prevention Beyond Lowering LDL Cholesterol. Diabetes. 2016;65:1767–1778. [DOI] [PubMed] [Google Scholar]
- 39.Liou L, Kaptoge S. Association of small, dense LDL-cholesterol concentration and lipoprotein particle characteristics with coronary heart disease: A systematic review and meta-analysis. PLOS ONE. 2020;15:e0241993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Balling M, Afzal S, Varbo A, Langsted A, Davey Smith G, Nordestgaard BG. VLDL Cholesterol Accounts for One-Half of the Risk of Myocardial Infarction Associated With apoB-Containing Lipoproteins. Journal of the American College of Cardiology. 2020;76:2725–2735. [DOI] [PubMed] [Google Scholar]
- 41.Castañer O, Pintó X, Subirana I, Amor AJ, Ros E, Hernáez Á, Martínez-González MÁ, Corella D, Salas-Salvadó J, Estruch R, et al. Remnant Cholesterol, Not LDL Cholesterol, Is Associated With Incident Cardiovascular Disease. Journal of the American College of Cardiology. 2020;76:2712–2724. [DOI] [PubMed] [Google Scholar]
- 42.Varbo A, Benn M, Tybjærg-Hansen A, Nordestgaard BG. Elevated remnant cholesterol causes both low-grade inflammation and ischemic heart disease, whereas elevated low-density lipoprotein cholesterol causes ischemic heart disease without inflammation. Circulation. 2013;128:1298–309. [DOI] [PubMed] [Google Scholar]
- 43.Narula N, Olin JW, Narula N. Pathologic Disparities Between Peripheral Artery Disease and Coronary Artery Disease. Arteriosclerosis, thrombosis, and vascular biology. 2020;40:1982–1989. [DOI] [PubMed] [Google Scholar]
- 44.Ding N, Sang Y, Chen J, Ballew SH, Kalbaugh CA, Salameh MJ, Blaha MJ, Allison M, Heiss G, Selvin E, et al. Cigarette Smoking, Smoking Cessation, and Long-Term Risk of 3 Major Atherosclerotic Diseases. Journal of the American College of Cardiology. 2019;74:498–507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Levin MG, Klarin D, Assimes TL, Freiberg MS, Ingelsson E, Lynch J, Natarajan P, O’Donnell C, Rader DJ, Tsao PS, et al. Genetics of Smoking and Risk of Atherosclerotic Cardiovascular Diseases: A Mendelian Randomization Study. JAMA network open. 2021;4:e2034461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Levin MG, Klarin D, Walker VM, Gill D, Lynch J, Hellwege JN, Keaton JM, Lee KM, Assimes TL, Natarajan P, et al. Association Between Genetic Variation in Blood Pressure and Increased Lifetime Risk of Peripheral Artery Disease. ATVB. 2021;41:2027–2034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kannel WB. Risk factors for atherosclerotic cardiovascular outcomes in different arterial territories. European Journal of Cardiovascular Prevention & Rehabilitation. 1994;1:333–339. [PubMed] [Google Scholar]
- 48.Stone NJ, Robinson JG, Lichtenstein AH, Bairey Merz CN, Blum CB, Eckel RH, Goldberg AC, Gordon D, Levy D, Lloyd-Jones DM, et al. 2013 ACC/AHA guideline on the treatment of blood cholesterol to reduce atherosclerotic cardiovascular risk in adults: A report of the american college of cardiology/american heart association task force on practice guidelines. Circulation. 2014;129:S1–S45. [DOI] [PubMed] [Google Scholar]
- 49.Arnett DK, Blumenthal RS, Albert MA, Buroker AB, Goldberger ZD, Hahn EJ, Himmelfarb CD, Khera A, Lloyd-Jones D, McEvoy JW, et al. 2019 ACC/AHA Guideline on the Primary Prevention of Cardiovascular Disease: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation. 2019;140:e596–e646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Burgess S, Butterworth A, Malarstig A, Thompson SG. Use of Mendelian randomisation to assess potential benefit of clinical intervention. BMJ. 2012;345:e7325–e7325. [DOI] [PubMed] [Google Scholar]
- 51.Barter PJ, Caulfield M, Eriksson M, Grundy SM, Kastelein JJP, Komajda M, Lopez-Sendon J, Mosca L, Tardif J-C, Waters DD, et al. Effects of Torcetrapib in Patients at High Risk for Coronary Events. New England Journal of Medicine. 2007;357:2109–2122. [DOI] [PubMed] [Google Scholar]
- 52.Pradhan AD, Paynter NP, Everett BM, Glynn RJ, Amarenco P, Elam M, Ginsberg H, Hiatt WR, Ishibashi S, Koenig W, et al. Rationale and design of the Pemafibrate to Reduce Cardiovascular Outcomes by Reducing Triglycerides in Patients with Diabetes (PROMINENT) study. American Heart Journal. 2018;206:80–93. [DOI] [PubMed] [Google Scholar]
- 53.Kofink D, Eppinga RN, van Gilst WH, Bakker SJL, Dullaart RPF, van der Harst P, Asselbergs FW. Statin Effects on Metabolic Profiles. Circulation: Cardiovascular Genetics. 2017;10:e001759. [DOI] [PubMed] [Google Scholar]
- 54.Davies NM, Holmes MV., Davey Smith G. Reading Mendelian randomisation studies: A guide, glossary, and checklist for clinicians. BMJ (Online). 2018;362:k601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Zuber V, Gill D, Ala-Korpela M, Langenberg C, Butterworth A, Bottolo L, Burgess S. High-throughput multivariable Mendelian randomization analysis prioritizes apolipoprotein B as key lipid risk factor for coronary artery disease. International Journal of Epidemiology. 2020. doi: 2020.02.10.20021691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Staples J, Qiao D, Cho MH, Silverman EK, University of Washington Center for Mendelian Genomics, Nickerson DA, Below JE. PRIMUS: rapid reconstruction of pedigrees from genome-wide estimates of identity by descent. Am J Hum Genet. 2014;95:553–564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ioannidis NM, Rothstein JH, Pejaver V, Middha S, McDonnell SK, Baheti S, Musolf A, Li Q, Holzinger E, Karyadi D, et al. REVEL: An Ensemble Method for Predicting the Pathogenicity of Rare Missense Variants. Am J Hum Genet. 2016;99:877–885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Karczewski KJ, Francioli LC, Tiao G, Cummings BB, Alföldi J, Wang Q, Collins RL, Laricchia KM, Ganna A, Birnbaum DP, et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature. 2020;581:434–443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.McLaren W, Gil L, Hunt SE, Riat HS, Ritchie GRS, Thormann A, Flicek P, Cunningham F. The Ensembl Variant Effect Predictor. Genome Biol. 2016;17:122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhan X, Hu Y, Li B, Abecasis GR, Liu DJ. RVTESTS: an efficient and comprehensive tool for rare variant association analysis using sequence data. Bioinformatics. 2016;32:1423–1426. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request. GWAS summary statistics for PAD are available by application in dbGaP (phs001672). GWAS summary statistics for GLGC lipids, UKB lipids, NMR lipids, and CARDIoGRAMplusC4D CAD are available for download from the IEU Open GWAS Project (https://gwas.mrcieu.ac.uk/).