

Abstract
Objective:
Use a highly potent and selective small molecule inhibitor of interleukin-1 associated kinase (IRAK) 4, PF-06650833, to demonstrate its role in autoimmune pathophysiology in vitro, in vivo and in the clinic.
Methods:
Rheumatoid arthritis (RA) inflammatory pathophysiology was modeled in vitro through stimulation of primary human macrophages (MΦ) with anti-citrullinated protein antibody (ACPA) immune complexes (IC), RA fibroblast-like synoviocyte (-FLS) cultures stimulated with toll-like receptor (TLR) ligands, as well as additional human primary cell co-cultures. Systemic lupus erythematosus (SLE) pathophysiology was simulated in human neutrophils, dendritic cells (DC), B cells and PBMC stimulated with TLR ligands and SLE patient IC. PF-06650833 was evaluated in vivo in the rat collagen-induced arthritis (CIA) model and the mouse pristane-induced and MRL/lpr models of lupus. Finally, RNASeq data generated with whole blood samples from a Phase 1 multiple ascending dose clinical trial of PF-06650833 were used to test in vivo human pharmacology.
Results:
In vitro, PF-06650833 inhibited human primary cell inflammatory responses to physiologically relevant stimuli generated with RA and SLE patient plasma. In vivo, PF-06650833 reduced circulating autoantibody levels in the pristane-induced and MRL/lpr murine models of lupus and protected rats from CIA. In a phase 1 clinical trial (NCT02485769), PF-06650833 demonstrated in vivo pharmacology pertinent to SLE by reducing whole blood interferon (IFN) gene signature expression in healthy volunteers.
Conclusion:
These data demonstrate that inhibition of IRAK4 kinase activity can reduce markers of inflammation in humans and provide confidence in the rationale for clinical development of IRAK4 inhibitors for rheumatologic indications.
Introduction
Rheumatoid arthritis affects approximately 1% of the population, manifesting in joint pain and tissue destruction characterized in seropositive cases by antibodies against post-translationally modified proteins, as well as IgM (rheumatoid factor, RF)(1, 2). Cellular players in RA inflammation include not only lymphocytes but also neutrophils, macrophages, osteoclasts, and synovial fibroblasts. Treatments for RA include both small molecule disease-modifying anti-rheumatic drugs (DMARDs) as well as biologics(3). The ability to sustain remission rates over time in RA remains disappointingly low, highlighting the need for additional therapeutic mechanisms(4).
Systemic lupus erythematosus is also a systemic disease mediated by autoantibodies, and is characterized by tissue inflammation and damage to multiple organ systems including joints, skin and kidney(5). Defects in the clearance of apoptotic and necrotic cells have been demonstrated, allowing access to nuclear antigens by autoantibodies(6). Resulting immune complexes activate numerous immune cell types, including DC and B lymphocytes. Treatments for SLE include corticosteroids and anti-malarials, but efficacy is limited and long-term use is associated with toxicity. The BAFF neutralizing antibody belimumab was approved in 2015, and while it provides benefit to some SLE patients, there is clear medical need remaining(5).
Interleukin-1 receptor activated kinase 4 is a central regulator of the innate immune response. IRAK4 transmits signals from TLRs and IL1R by binding the adaptor protein MyD88 and inducing signals through IRAK1 and IRAK2(7, 8). The downstream result of myddosome assembly is activation of nuclear factor kappa B (NFΚB), interferon regulatory factor 5 (IRF5) and mitogen activated protein kinase (MAPK) (9). Deletion of IRAK4 or inactivation of IRAK4 kinase activity in mice prevents the development of inflammation in multiple models of inflammatory disease(10–14). Cells from IRAK4-deficient humans also show no response to TLR or IL1R family ligands that signal through MyD88(15).
Several studies have recently highlighted the efficacy of IRAK4 inhibitors in human cells and pre-clinical models of inflammation(16–20). However, clinical development has been hampered by debate over the role of IRAK4 kinase activity in disease, as there is a significant role for kinase-independent signaling by IRAK4(21, 22). We have shown that inhibition of the kinase activity of IRAK4 does not significantly affect IL1- or TLR-induced NFkB or MAP-kinase activation and show minimal suppression of IL1 induced cytokines, whereas IRAK4 null cells are devoid of IL1 or TLR activation of NFkB, MAP-kinase activation and production of cytokines(7, 23). Additionally, several in vitro studies have suggested that IRAK4 inhibitors, which are efficacious in rodent cells, are not efficacious in human cells(22, 24, 25). It has also been speculated that inhibition of both IRAK4 and IRAK1 kinase activities may be required for efficacy in human cells(21, 22). A new report using an IRAK4 inhibitor has confirmed the requirement for IRAK4 kinase activity in mouse and human dendritic cells and B cell activation as well as in several in vivo models of SLE(20). However, translation of in vitro efficacy of IRAK4 inhibitors to reduction of inflammation in humans is lacking.
We have recently developed potent and selective inhibitors of IRAK4 with little activity against IRAK1(18). Here, we demonstrate that PF-06650833 reduces responses to disease-relevant stimuli in human cells and in animal models of RA and SLE. Importantly, we show that administration of PF-06650833 to humans resulted in the suppression of an interferon gene signature in a Phase 1 multiple ascending dose (MAD) clinical trial(26). Thus, selective IRAK4 inhibitors reduce signals of inflammation in humans and are potential therapeutics for autoimmune disease.
Materials and Methods
All details of materials and methods are supplied in Supplementary Materials. The identification of PF-06650833 and methods used to define its pharmacology were previously described(18).
Human and rodent cytokine release assays.
Human, mouse and rat whole blood collected under IRB- and IACUC-approved protocols and anti-coagulated with heparin, or PBMCs derived therefrom, were incubated with PF-06650833, exposed to TLR ligands, and cytokines assayed by Mesoscale Discovery assay kits (MSD).
IRAK4 Occupancy.
Occupancy of the IRAK4 ATP binding site was quantified via blockade of covalent labeling with a biotin-labeled ATP probe followed by immunoprecipitation and western blotting for IRAK4.
Macrophage ACPA-IC induced TNFα.
Human MΦ were exposed to inhibitors, then incubated with ACPA-IC and supernatant cytokine measured by MSD.
Biomap Diversity PLUS Panel.
The BioMAP Diversity PLUS Panel protocols have previously been published (27, 28) and were performed at Eurofins.
Human RA-FLS.
Human FLS were incubated with inhibitors, and the noted stimulatory ligands added. Supernatants were analyzed by MSD.
The rat collagen-induced arthritis model.
Female Lewis rats were immunized with collagen and then dosed orally with PF-06650833 or Tofacitnib for 7 days. Paw volume and body weight were measured daily. All procedures were reviewed and approved by the Pfizer IACUC.
Netosis assay.
Neutrophils were isolated from human volunteers and SLE patients under IRB-approved protocols. Healthy volunteer neutrophils were primed with IFNα2β and stimulated with R837 alone or in combination with PF-06650833. Neutrophils derived from SLE patient blood samples were cultured in media 8 to 12 hrs. dsDNA was quantified using the Quant-iT™ PicoGreen™ Kit.
Monocyte IRF5 assay.
PBMC were isolated from healthy human volunteers, incubated with PF-06650833, and stimulated with R848 or SLE patient sera. Nuclear localization of IRF5 was measured using an Amnis Imaging Cytometer(7).
B cell maturation assay.
Human B cells were isolated from leukopacks, primed with IFNα, incubated with PF-06650833, and stimulated with R848. Supernatants were analyzed for cytokines and the cells for plasmablast differentiation.
pDC activation assay.
The total DC fraction was obtained from healthy volunteer buffy coats. PDCs were then isolated by FACS and cultured with 40% SLE neutrophil supernatants with or without PF-06650833. IFN-α levels were measured by ELISA.
SLE-IC assay.
SLE-IC were generated from patient plasma and used to stimulate human PBMC with or without PF-06650833. IFNα were measured by ELISA. IFN-responsive gene expression was analyzed by quantitative RT-PCR (qRT-PCR).
Pristane and MRL-lpr in vivo models of SLE.
Procedures were conducted according to the appropriate IACUC regulations. Following pristane administration to Balb/c mice, treatment with PF-06650833 was dosed in chow weeks 8–20, while prednisone was orally administered once daily weeks 1–20. Serum was collected at weeks, 4, 8, 12, and 20. Anti-ds DNA, anti-SSA and anti-RNP were quantified by ELISA. Kidney specimens were prepared as described (29) and evaluated by a pathologist blinded to the treatment group.
Female MRL/lpr were dosed with standard chow or PF-06650833 for 12–13 weeks. Body weights were taken once a week, proteinuria assessments made every other week, and blood collection was performed every 4 weeks.
Interferon signature modulation in HV.
The Phase 1 studies of PF-06650833 have been previously described(26). RNA was extracted from whole blood and subjected to RNASeq. An equally weighted 21 gene signature(30) was analyzed for percent change at day 14 vs day 0.
Results
Pharmacological properties of PF-06650833
PF-06650833 was identified as described previously (Fig 1A)(18). The compound is selective as measured by its ability to compete with a covalent analog of ATP in monocyte cell lysates by ActivX (La Jolla, CA) (31). Twelve kinases, other than IRAK4, have IC50 values less than 1 uM (Fig 1B and Table S1), and PF-06650833 is nearly 7000 times more selective for IRAK4 than for IRAK1 (Fig. 1B). In PBMCs stimulated with the TLR 7/8 ligand R848, PF-06650833 had an IC50 of 2.4 nM for the inhibition of TNFα release and is potent in human whole blood, giving an IC50 of 8.8 nM (Fig 1C). The free IC50 values for PF-06650833 in rodent whole blood were determined to be 5–10 fold less potent than in human whole blood, in contrast to previous reports indicating lack of IRAK4 inhibitor activity in human cells (24, 25).
Fig. 1.
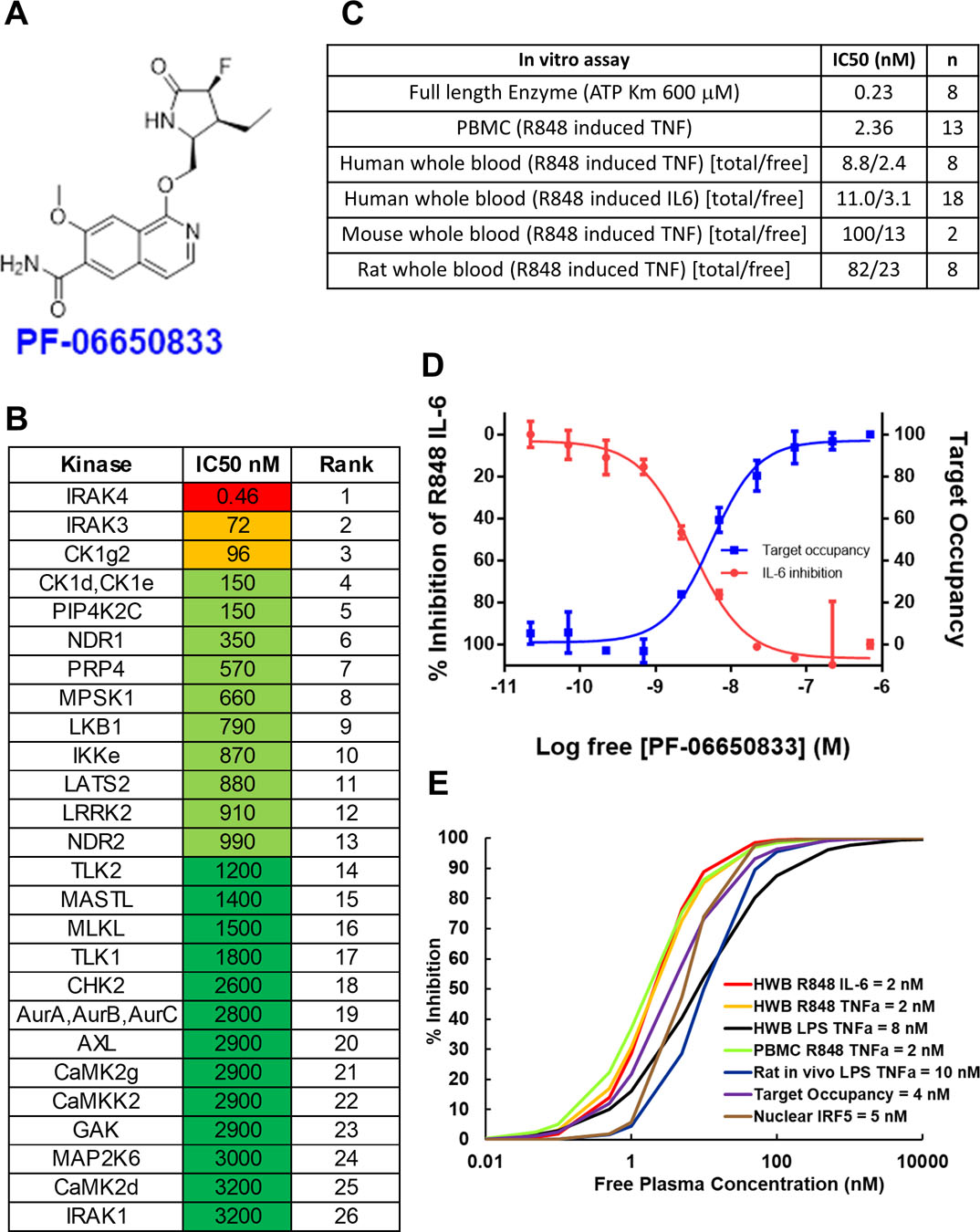
Pharmacological properties of PF-06650833. (A) Structure of PF-06650833. (B) Selectivity of PF-06650833 in ActivX ATP occupancy assay in THP1 lysates. The top 26 out of 200+ kinases are shown. IC50s were determined by a 5-point dose response curve (full data set in table S1) (C) Potency of PF-06650833 in enzyme and cell-based assays. IC50 values for enzyme and PBMC assays are a single value, whereas whole blood (wb) assays for human, rat and mouse are denoted as total concentration of compound over the non-protein bound (free=f) concentration of compound. (wb IC50*[fu/(B/P)] = IC50 free, where fu = 0.22, B/P = Blood/plasma ratio = 0.91). (D) Demonstration of the relationship between free drug concentration, IRAK4 ATP binding site occupancy, and inhibition of downstream pharmacology by PF-06650833 in a sample of human SLE whole blood. (E) EC50 data for each pre-clinical assay were used to determine that 100 nM was the target free compound concentration was required to maintain greater than 90% IRAK4 inhibition across species and assays.
Modeling of PF-06650833 IRAK4 pharmacology to select efficacious dose
Pfizer has established the “Three Pillars of Survival” (32) concept to predict clinical success based on the ability to measure candidate pharmacology. For PF-06650833, we established a system that captured all 3 pillars- free drug concentration (Pillar I), target occupancy (Pillar II), and down-stream biological effect (Pillar III)- in an in vitro assay performed in SLE patient whole blood (Fig 1D). Increasing concentrations of PF-06650833 (Pillar I, X axis) resulted in increased target occupancy (Pillar II, right Y axis), as measured by blockade of the IRAK4 binding site from covalent modification by a probe. With a very similar EC50 value to target occupancy, R848-induced IL-6 release (Pillar III, left Y axis) was inhibited by PF-06650833. Similar results were obtained using blood provided by a healthy volunteer (Fig S1). Thus, target occupancy and cytokine inhibition were proportional and predictable from compound concentration, allowing us to infer Pillar II, target occupancy, from Pillar III assay results. Using experimentally generated EC50, Hill slope, and protein binding values, we then examined the relationship between free drug concentration and target inhibition for the pre-clinical assays (Figure 1E). The EC50 values for PF-06650833 varied by only 5-fold in these assays- from 2 to 10 nM- such that achieving greater than 100 nM free drug concentration would be expected to inhibit more than 90% of any down-stream biology dependent on IRAK4 kinase activity. Thus, our target efficacious concentration (Ceff) for pre-clinical experiments was set at 100 nM.
PF-06650833 inhibits pathophysiological processes central to RA.
In the case of seropositive RA, formation of anti-citrullinated protein antibodies (ACPA) precedes the clinical diagnosis of RA, are a specific marker of autoimmunity in RA, and indicate more rapid progression of disease and poorer treatment response (33, 34). ACPA immune complexes (IC) activate MΦ to release TNFα(35). Thus, we tested the ability of PF-06650833 to block ACPA-IC induced TNFα. As shown in Figure 2A, 100 nM PF-06650833 inhibited TNFα release to 57.5 +/− 17.9 %, p = 0.003 for raw values, paired T test, n = 11 donors. As a positive control, we included a bruton’s tyrosine kinase (BTK) inhibitor, as this kinase has been previously implicated in MΦ TNF responses to both TLR and FcγR ligands(36).
Fig. 2.
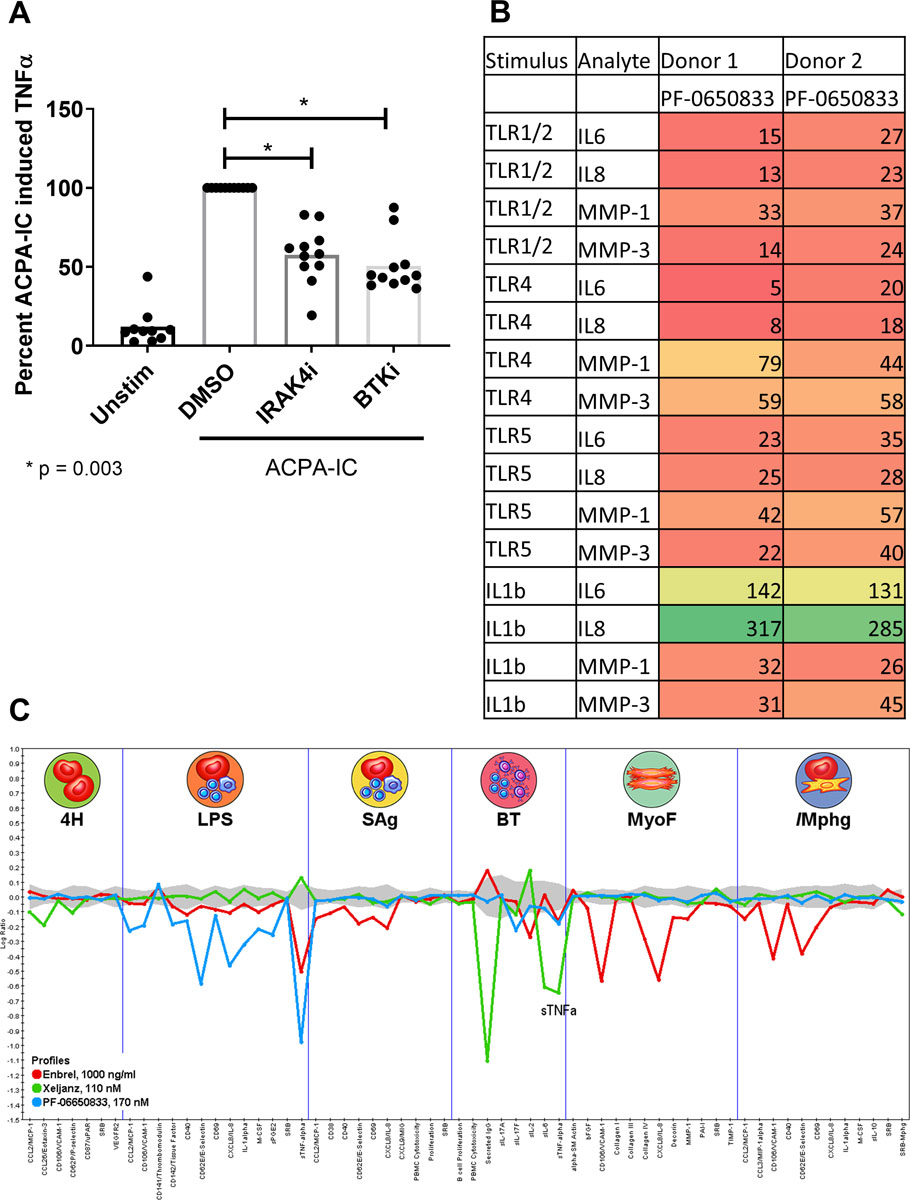
PF-06650833 inhibits RA pathophysiology.(A) Human MΦ were exposed to ACPA-IC formed with CCP+ RA sera as described in Methods in the presence or absence of 100 nM IRAK4i (PF-06650833) or 100 nM BTKi (PF-303). Supernatants were analyzed for TNFα by ELISA. (B) Human RA-FLS from were stimulated with 10 ug/ml PAM3Cys (TLR1/2), 10 ng/ml LPS (TLR4), 100 ng/ml flagellin (TLR5), or 0.1 ng/ml IL1b in the presence or absence of 100 nM PF-06650833. Supernatants were assayed for cytokine and MMP content by MSD. Values represent percentage of vehicle control. (C) Compounds at the noted concentrations were profiled by DiscoverRx on the BioMAP® platform. Data represent the log ratio of compound treated samples to controls, with negative values indicating inhibition, and points outside of the gray shading demonstrating a statistically significant difference based on assay variability. Each point on the X axis represents the result of a different assay endpoint in each assay system: venular endothelial cells (4H), peripheral blood mononuclear cells plus venular endothelial cells stimulated with LPS (LPS), peripheral blood mononuclear cells plus venular endothelial cells stimulated with super antigen (Sag), B cells plus peripheral blood mononuclear cells (BT), lung fibroblasts (MyoF), and macrophages plus venular endothelial cells (/Mphg).
Synovial fibroblasts from RA patients display increased inflammatory responses to TLR ligands and IL1β (37, 38). We exposed RA-FLS to inflammatory stimuli in the presence or absence of 100 nM PF-06650833 and quantified their inflammatory response. As depicted in the heatmap (Fig 2B), PF-06650833 substantially reduced cytokine and matrix metalloprotease (MMP) release in response to all ligands profiled. However, PF-06650833 did not inhibit IL1β-induced cytokines in RA-FLS. This is similar to our previous findings with dermal fibroblasts stimulated with IL1β, where cytokine release was only weakly affected by IRAK4 inhibition(8, 23). Similar results were also generated using FLS from non-RA donors (n = 2), although the magnitude of inflammatory mediators released was less (not shown). Intriguingly, PF-06650833 was able to block MMP induction by IL1β, the first response to IL1β in non-hematopoietic cells we have found to be IRAK4 kinase-dependent.
We next tested the ability of PF-06650833 to inhibit inflammatory processes in more physiologic tissue culture models involving multiple human primary cells using the DiscoveRx BioMap® platform and compared the effect to 2 approved RA therapies, the TNFα inhibitor Enbrel and the JAK inhibitor Xeljanz. As depicted in Figure 2C, PF-06650833 showed the greatest inhibition of inflammatory read-outs in assays of innate immunity, notably the LPS-stimulated PBMC plus endothelium assay, with modest activity in a small number of endpoints in the B-T cell assay. By contrast and as expected, the JAK inhibitor had no impact on assays of innate immunity (LPS, MyoF, /MΦ), blocking only responses in the B-T cell assay. Enbrel had yet a third pattern, with predominant activity in the myofibroblast and macrophage containing assays, and a modest effect on some readouts in the LPS and bacterial superantigen-stimulated PBMC assays. These results indicate that an IRAK4 inhibitor would block a distinct pattern of multicellular inflammatory processes in RA to the approved RA treatments.
PF-06650833 reduces inflammation in the rat CIA model.
To model RA in vivo, we profiled PF-06650833 in the rat CIA model. Vehicle, 10 mg/kg tofacitinib QD or 3 mg/kg PF-06650833 BID were administered therapeutically for the last 7 days of the study. The kinetic data from one representative study of 3 is presented in Figure 3, demonstrating significant inhibition versus vehicle treated animals (0.786 +/− 0.043 mL, MN +/− SEM) of paw swelling on Day 7 for both PF-06650833 (0.555 +/− 0.034 mL, MN +/− SEM, n = 10 animals per group, p = 0.0005, t-test) and tofacitinib (0.371 +/− 0.044 mL, p < 0.0001). Thus PF-06650833 is capable of inhibiting inflammation in vivo, even when administered weeks after the initial inflammatory insult.
Fig. 3.
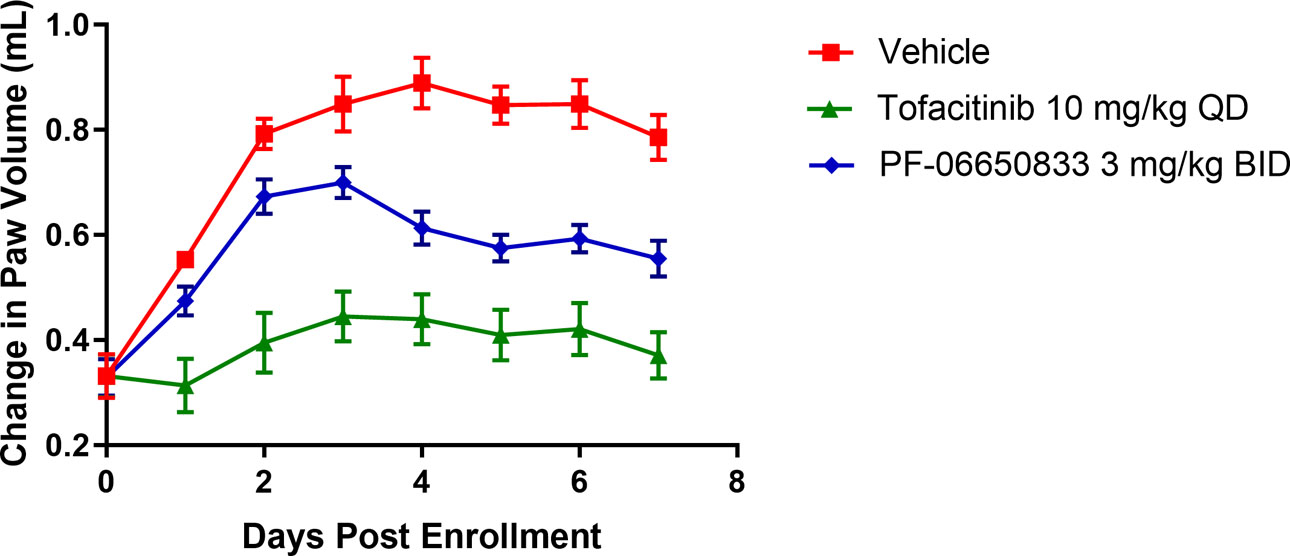
PF-06650833 is efficacious in rat CIA. PF-06650833 was administered 3mg/kg BID or Tofacitinib was administered 10 mg/kg PO QD in the rat CIA model for 7 days following enrollment. Change in paw volume on each day are presented (Representative data from one of 3 experiments are shown).
PF-06650833 inhibits pathophysiological processes central to SLE.
High titers of anti-nuclear antibodies (ANA) are diagnostic for SLE and formation of IC contributes to the multi-organ inflammation characteristic of SLE(39). Exposure of the immune system to nuclear antigens results from impaired clearance of apoptotic cells and/or enhanced release from neutrophils via NETosis(40–42). Lupus immune complexes can then induce cytokine release from monocytes, T cell independent B cell maturation, as well as activate plasmacytoid DC (pDC) to release type 1 interferons, resulting in the characteristic interferon gene signature of SLE(43, 44). We stimulated DNA release from human neutrophils by application of the TLR7 agonist R837 for 15h. DNA content in the supernatants was significantly higher than from neutrophils incubated in medium alone (43.3 +/− 9.6 versus 113.5 +/− 10.7 ng/mL, p = 0.0001 paired T test, n = 3), and was completely reversed by pre-incubation with PF-06650833 (45.83 +/− 9.7 ng/mL, p = 0.0017) (Figure 4A). Monocytes from SLE patients show higher IRF5 protein expression and nuclear localization(45). After confirming the ability of PF-06650833 to inhibit nuclear localization of IRF5 stimulated by R848 (23.0 +/− 13.0% vs. 6.8 +/− 4.4%, p = 0.012, 2-way ANOVA, n = 3), we demonstrated its capacity to inhibit IRF5 nuclear localization in response to SLE patient sera (38.6 +/− 18.4% vs 24.2 +/− 13.0%, p < 0.0001, 2-way ANOVA, n = 12) (Figure 4B).
Fig. 4.
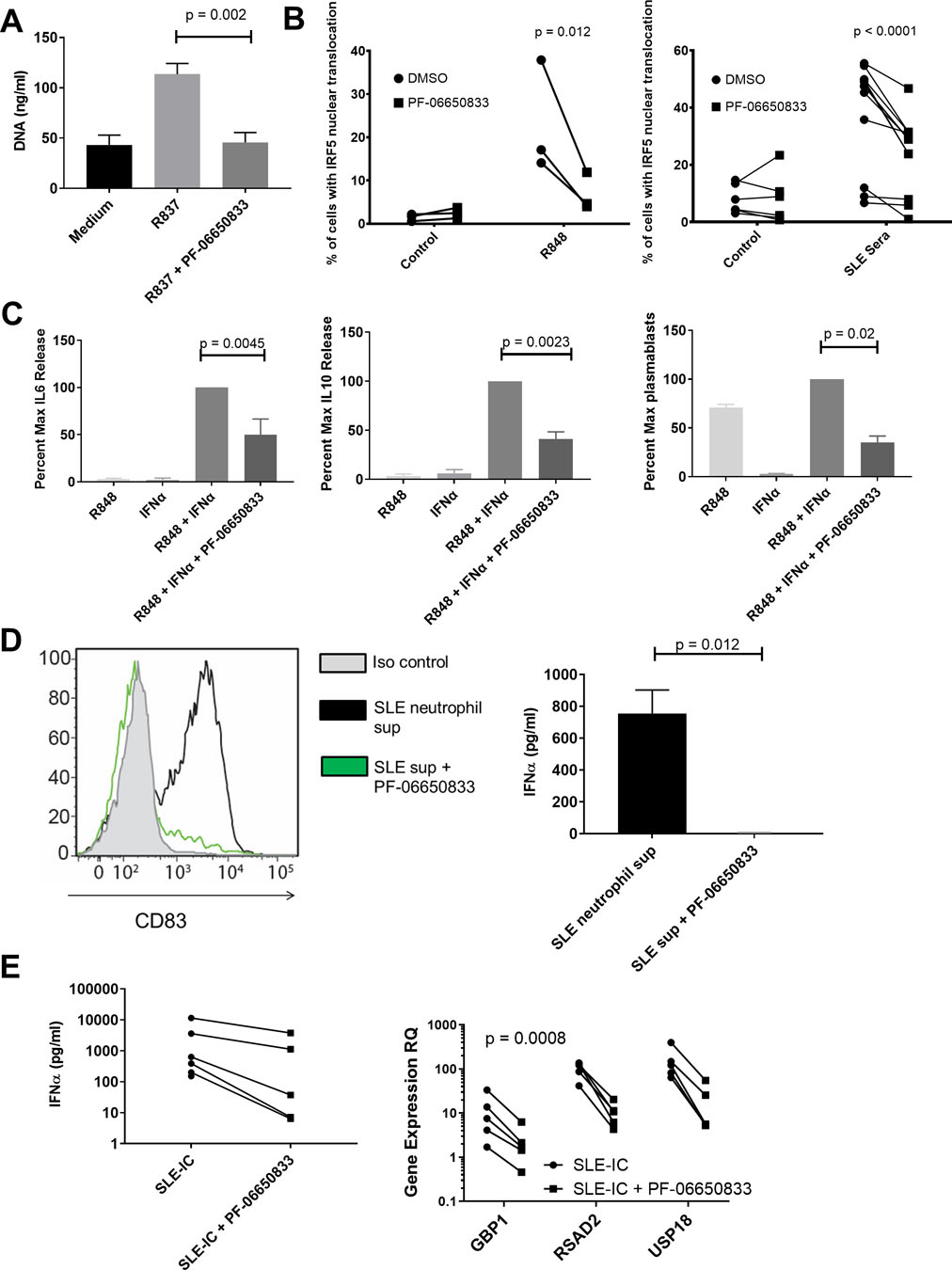
PF-06650833 inhibits SLE human pathophysiology. (A) DNA release was measured in neutrophil supernatant following exposure to R837 in the presence or absence of 100 nM PF-06650833 (MN +/− SD, n = 3 donors) (B) PBMC were exposed to either R848 or SLE sera in the presence or absence of 100 nM PF-06650833, incubated for 2 hours at 37oC, then analyzed for IRF5 nuclear translocation by Amnis imaging cytometry as described in Methods. (C) B cells were exposed to IFNα + R848 in the presence or absence of 100 nM PF-06650833. Supernatants were harvested at 24 hours for IL6 release and 72h for IL10 release. Plasmablasts were quantified by flow cytometry after 6 days. (D) Neutrophils were isolated from SLE patients and netosis induced with R837. Supernatants were used to stimulate pDC for 24 hours in the presence or absence of 200 nM PF-06650833 prior to quantification of CD83 induction by flow cytometry (n=5) or IFNα release by ELISA (n=3). (E) PBMC were exposed to SLE-IC in the presence or absence of 100 nM PF-06650833. Supernatant was analyzed after 24 hours for IFNα by ELISA and IFN-induced gene expression in cells using qRT-PCR.
Exposure to IFN and TLR7 ligands results in B cell cytokine release and T cell independent B cell maturation into plasmablasts(46, 47). Incubation of human B cells with 100 nM PF-06650833 prior to IFNα and R848 inhibited B cell IL6 production at 24 h to 50.0 +/− 16.7 % of control (p = 0.0045, paired T test of raw values, n = 6 donors); IL10 production at 72 h to 41.3 +/− 7.1% of control (p = 0.0023, paired T test of raw values, n = 6 donors); and CD27+/CD38+ plasmablast differentiation at 7 d to 35.1 +/− 6.6 % of control (p = 0.02 paired T test of raw values, n = 5 donors) (Figure 4C). Although PF-06650833’s potency (IC50) for inhibiting these activities in primary human B cells (not shown) is comparable to that for inflammatory cytokine release by other cell types, inhibition is less complete than for inflammatory cytokine production by monocytes (48), suggesting IRAK4 kinase-independent signaling.
Plasmacytoid DC are a rare population in peripheral blood, but are responsible for the majority of IFNα release(44, 49). Neutrophil DAMPs induced pDC to upregulate CD83, indicating maturation, and this response was significantly inhibited by pre-incubation with 200 nM PF-06650833, and IFNα release from pDC was significantly inhibited by compound (755 +/− 147 vs 5.7 +/− 2.5 pg/mL, p = 0.012, paired T test, n = 3) (Figure 4D). SLE-IC formed by purifying IgG from plasma of an anti-RNP/anti-dsDNA positive SLE patient and mixing with apoptotic cells were used to stimulate human PBMC. PF-06650833 exposure inhibited IFNα release in all donors, as well as the induction of 3 interferon responsive genes- RSAD2, USP18 and GBP1 (p = 0.0008 for treatment, 2-way ANOVA, n = 5) (Figure 4E). In summary, numerous SLE pathophysiological processes, including neutrophil DAMP release, monocyte inflammatory cytokine response, B cell cytokine release and plasmablast differentiation, pDC maturation, and IFNα release as well as the resulting interferon gene signature are dependent on IRAK4 kinase activity.
PF-06650833 is therapeutic in mouse models of SLE.
The pristane-induced model of SLE is dependent on TLR activation and results in an anti-nuclear autoantibody repertoire similar to that for human SLE(50). Balb/c mice were dosed intraperitoneally (IP) with pristane to induce disease, and standard chow was replaced 8 weeks later with compound-containing chow for the remaining 12 weeks of the study. The results from 1 of 2 representative studies are shown in Figure 5. Longitudinal quantitation of anti-dsDNA titers revealed a significant increase in anti-dsDNA for pristane-treated animals versus control beginning at 12 weeks (10, 977 +/− 1,503 U/mL vs 5294 +/− 678, MN +/− SEM, n = 10 animals per group, p = 0.0023) that continued at week 20 (17,645 +/− 3527 vs 5103 +/− 816 U/mL, MN +/− SEM, p < 0.0001 for comparison by 2-way ANOVA). At week 20, therapeutic dosing with PF-06650833 had significantly reduced anti-dsDNA titers compared to control (8159 +/− 1282 vs 17,645 +/− 3527, p <0.0001), as had prednisone treatment (8608 +/− 1637, p <0.0001). At week 20 there also were significant reductions in anti-SSA IgG (2726 +/− 494 vs 7693 +/− 1241 U/mL, MN +/− SEM, p < 0.004, Mann-Whitney test) and anti-RNP IgG (3730 +/− 2861 vs 18,127 +/− 7902, MN +/− SEM, p = 0.015, Mann-Whitney test) with PF-06650833 treatment.
Fig. 5.
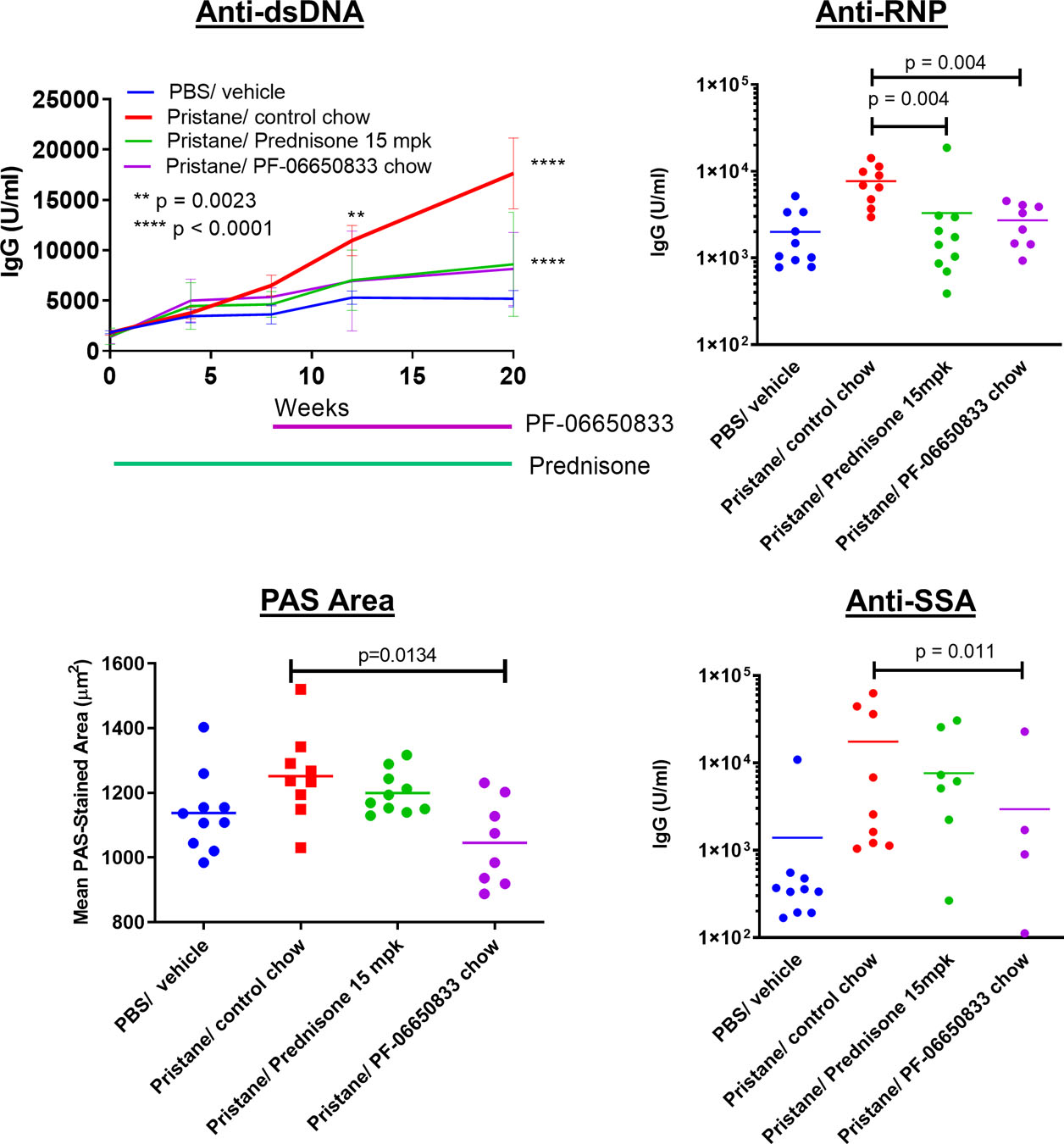
Effect of PF-06650833 (weeks 8–20) on pristane-induced SLE: PF-06650833 was administered to Balb/c mice by chow dosing during weeks 8–20 following pristane administration. Anti-dsDNA was quantified by ELISA on weeks 4, 8, 12, and 20. Additional auto-antibody titers as well as immunohistochemical determinations were made at termination at week 20. (Representative data shown for 1 of 2 experiments)
Kidney inflammation in the pristane model was mild and did not result in proteinuria. As there was no increase in renal inflammation, tubular injury nor proteinuria/cast scores in response to pristane exposure, the effects of the IRAK4 inhibitor were not evaluated on these endpoints. Kidneys in all groups were also evaluated by quantitative image analysis (QIA), as described previously(51), for glomerular tuft area, periodic acid Schiff (PAS) staining area per tuft, glomerular nuclear area, IgG immunohistochemistry (IHC) and C3 IHC scores. Of these, only mean glomerular tuft area was significantly increased by pristane, and only mean PAS area per tuft was significantly reduced by PF-06650833(1045 +/− 47 μm2 vs 1252 +/− 45 μm2, MN +/− SEM, n = 8 animals per group, p = 0.013 for comparison Kruskal-Wallis test) (Figure 5).
Chow dosing of PF-06650833 was also explored in the MRL/lpr model of SLE. In two studies, PF-06650833 significantly reduced lymphadenopathy, anti-histone IgG, histological kidney inflammation, glomerular nephropathy, C3 and IgG deposition by IHC, and glomerular tuft area and glomerular nuclear area by image analysis (Fig. S2, S3), while showing trends toward reductions in splenomegaly, proteinuria, anti-dsDNA, anti-SSA, and tubular injury. As BTK inhibitors have previously shown efficacy in this model (52–54), and we previously showed in vivo efficacy of the BTK inhibitor PF-06250112 in the NZB/NZW murine model of lupus(29), it was included as a positive control. PF-06250112 significantly reduced proteinuria, autoantibody titers, and all renal histological pathology endpoints other than PAS area per tuft (Fig. S2, S3). It may well be that in the pristane-induced SLE model whereby autoantibodies are induced via TLR activation, IRAK4 kinase activity is essential, while in the MRL/lpr model, where autoantibody production is initiated by faulty B cell apoptosis, IRAK4 kinase activity is more important for the resulting inflammation than the production of autoantibodies themselves.
Selective IRAK4 inhibition reduces interferon signature genes in healthy volunteers.
We have completed two randomized, double-blind, sponsor-open Phase 1 studies of the safety, pharmacokinetics, and pharmacodynamics of single- and multiple-ascending doses (MAD) of PF-06650833(26). Since the 300 mg dose of a modified release (MR) formulation showed maximum pharmacology in reducing CRP at Day 14 of dosing, we evaluated the effect of PF-06650833 on an IFN signature (comprised of the normalized expression of 21 genes, (30)) as a biomarker of systemic inflammation relevant to SLE pathogenesis at this timepoint, in comparison to the respective gene signature for each volunteer on Day 0 prior to dosing. As demonstrated in Figure 6, the IFN gene signature in the placebo group had changed positively a median of 9.1%, while that of the PF-06650833-treated trial participants was reduced by a median of 28.8% (delta = −37.9%, p=0.015 Wilcoxon test). The paired gene score values for each volunteer at Day 0 and Day 14 are also plotted in Figure S4, showing minimal regulation in gene scores for placebo treated individuals- 6 of 11 positive and 5 of 11 negative- whereas the magnitude of change between Day 0 and Day 14 is consistently higher and 6 of 7 are negative for the PF-06650833 treated individuals. Thus, we have demonstrated that in humans, an IRAK4 kinase inhibitor reduces biomarkers of inflammation relevant to RA and SLE pathophysiology.
Fig. 6.
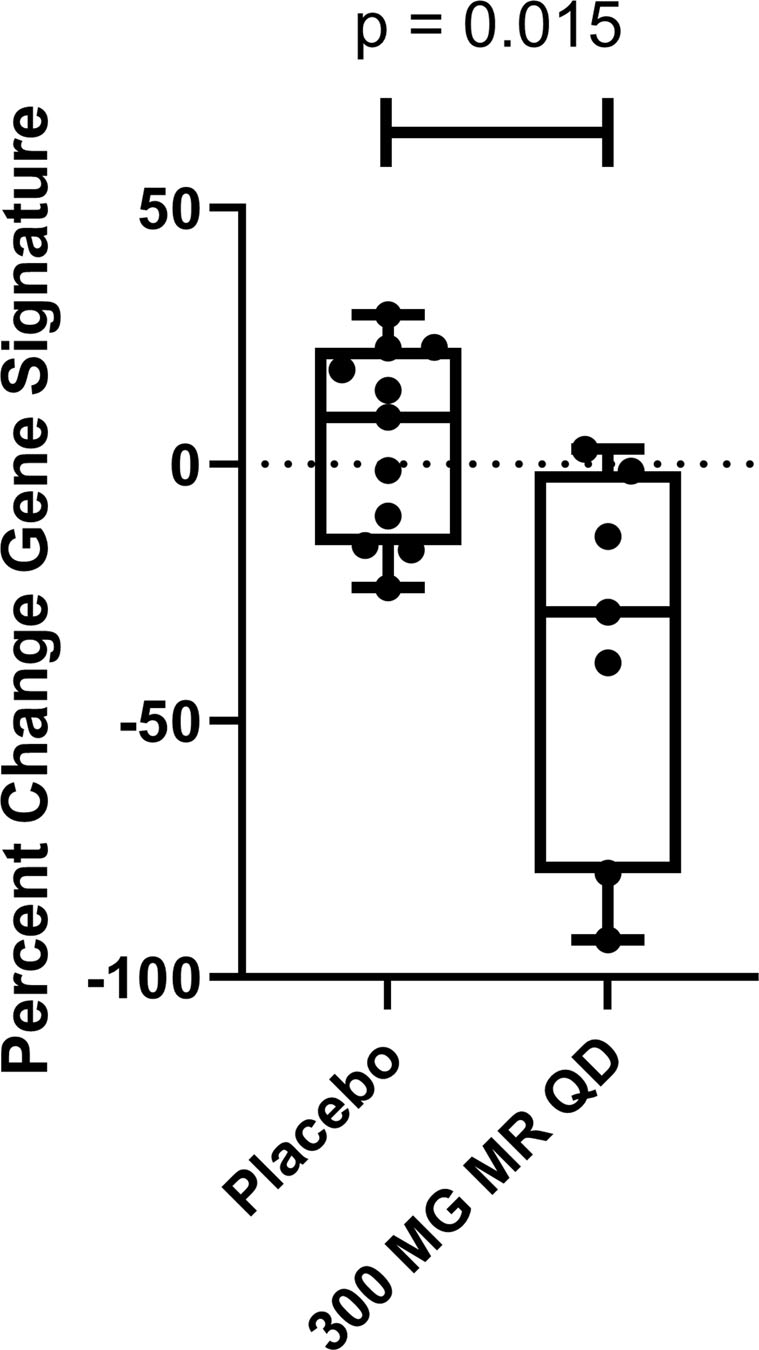
PF-06650833 inhibits type I interferon signature in vivo in humans. 300 mg of MR PF-06650833 was administered QD for 14 days in a Phase 1 MAD trial in healthy human volunteer subjects. A PAXgene whole blood tube was collected on day 0 prior to the first dose and on day 14 prior to administration of the last dose, RNA extracted, and a composite gene signature calculated as described in Methods. Plotted is the percentage change in the composite gene signature for each individual participant between the two timepoints. (Black line indicates median, box indicates 2 center quartiles, n = 11 placebo, n = 7 300 MG MR QD)
Discussion
Inhibitors of innate immune signaling pathways are potential therapeutic targets for treatment of autoimmune diseases(55, 56). In this report, we demonstrate that PF-06650833 effectively inhibits cytokines produced by MΦ activated by ACPA-IC from RA patients as well as pDCs activated by SLE-IC. We also show that PF-06650833 effectively inhibits cytokines induced by TLR ligands on RA-FLS but is not effective against IL1β-induced cytokines. However, PF-06650833 is effective at blocking MMPs induced by IL1β on these cells. The reason for this difference is not known but may result from the differential effects of IRAK4 kinase inhibition on early versus late myddosome formation and/or differential effects on transcription factor activation. Previously, we have shown that inhibition of IRAK4 kinase activity affects IL1β signaling by stabilizing the early form of the myddosome that signals primarily through IRAK1 and NFΚB (8, 57). It is possible that IL6/8 secretion is induced primarily by signaling via the early myddosome, whereas activation of MMPs might be mediated by the late myddosome, which signals through IRAK2(58). Regarding transcription factors, IRAK4 kinase activity is known to block IRF5, but not NFΚB or MAPK activation(7) and that these effects are cell type specific(38). It is possible that in RA-FLS, chromatin remodeling makes the MMP promoters more dependent on IRF5 or other IRAK4 kinase dependent transcription factors.
We also demonstrate that PF-06650833 is efficacious in a rat model of arthritis that did not use complete Freund’s adjuvant to ensure it was not driven by TLR. While it is encouraging that PF-06650833 significantly reduces severity in the rat CIA model, it is notable that the degree of inhibition was not as great as that observed for tofacitinib. It is possible that the rapid clearance of PF-06650833 from rodent species (rat t1/2= 0.6 hours(18)), resulted in transient loss of IRAK4 target inhibition in rat CIA. We have shown previously that efficacy for Tofacitinib is driven by average target occupancy (Cave)(56), while the evidence thus far suggests that minimum target occupancy (Cmin) is more relevant for IRAK4 mediated efficacy. Thus, the respective efficacy for PF-06650833 and Tofacitinib in rat CIA may have to do with target occupancy, mechanism of inhibition, or the respective contribution of each kinase to disease pathophysiology. Of greater relevance to the treatment of human RA patients, we recently disclosed significant effects on disease activity from a Phase 2 study of the efficacy and safety of PF-06650833 in patients with active rheumatoid arthritis and inadequate response to methotrexate(57). The clinical response rate at 12 weeks in that study were consistent with those reported for Tofacitinib, and the study itself included a cohort treated with Tofacitinib. Thus, the respective efficacy of the two molecules will be revealed by the publication of data from this clinical trial.
We show here that PF-06650833 can block type-I IFN induced by SLE serum and by neutrophil DAMPs in primary human pDCs. We also show that it can block plasmablast differentiation and B cell activation induced by TLR ligands and activation of the transcription factor IRF5 by SLE serum in monocytes(59, 60). Therapeutic dosing of PF-06650833 was efficacious in reducing the induction of all ANA tested in the pristane model of SLE, while only significantly reducing one of 3 reactivities assayed in the MRL/lpr model. Interestingly, PF-06650833 did show robust inhibition of kidney inflammatory histology in the MRL/lpr model. It is unknown whether this difference has to do with the different mechanisms of disease initiation in the 2 models or differences in our ability to cover the target, as PF-06650833 is rapidly cleared from mouse circulation.
Despite lower potency in rodent cells and rapid clearance from rodent circulation, the in vitro and in vivo results generated with PF-06650833 presented here go a long way towards settling some of the controversies regarding the relative importance of IRAK1 and IRAK4 kinase activity in various cell types in mouse and human (16, 19, 21, 22, 24, 25). IRAK4 kinase activity is necessary for DAMP induced inflammatory signaling in rodent and human leukocytes, and IRAK1 kinase activity does not provide a sufficient substitute. A recent publication is consistent with our data providing confirmation of the importance of IRAK4 kinase activity to lupus pathophysiological signaling in dendritic cells, B cells, and in vivo models using an unrelated small molecule inhibitor of IRAK4(20). There is further work to be done in understanding the role of IRAK4 kinase activity in stromal cells, as well as in IL1β-induced inflammation. We have shown that IL1β-induced cytokines from human fibroblasts treated with an IRAK4 inhibitor were only weakly inhibited even though IRAK4 autophosphorylation was completely inhibited (8, 61), while inhibition of TLR7/8 (R848)-induced cytokine production by IRAK4 inhibition coincided with the inhibition of IRAK4 autophosphorylation in primary human monocytes(7, 61) Likewise, in the results presented here, IL1β-induced cytokines in RA-FLSs were not inhibited by PF-06650833, in contrast to MMPs, which were inhibited.
To demonstrate proof of pharmacology in humans, we assessed markers of inflammation in a phase 1 clinical trial in healthy volunteers on an exploratory basis. These markers were IFN signature, increased in SLE(62), and CRP levels, increased in RA(63, 64). Remarkably, basal levels of these markers were present in healthy volunteers and, at a daily dose of 300 mg MR PF-06650833, we observed statistically significant reductions in both endpoints, demonstrating that an IRAK4 inhibitor reduces markers of inflammation in humans. To our knowledge, this is the first demonstration of modulating IFN regulated genes in healthy volunteers. Together with the CRP data(26), data reported herein represent the first proof of pharmacology of a selective IRAK4 inhibitor on inflammatory signaling pathways in humans. Altogether, these data strongly support the clinical utility of IRAK4 inhibitors for the treatment of multiple human inflammatory autoimmune diseases.
Supplementary Material
Acknowledgments:
The authors wish to thank Tatyana Souza and Varenka Rodriguez for optimizing the plasmablast differentiation assay; Jill Wright and Yanyu Zhang for human IgG purification from plasma; Alison O’Mahoney of Eurofins for providing the DiscoverRx figure; the scientists at ActivX for performing kinase selectivity assays and at Washington Biotechnology Inc for performing the pristane SLE model; our clinical collaborators for the Phase 1 MAD study and especially all the human volunteers who contributed to every phase of this work.
Conflict of interest statement: This research performed at all institutions was fully funded by Pfizer, Inc.; BJB is also supported by NIAMS, NIH 1R21AR065959-01. All authors except Saurav De, SA, VP and BJB were employees of Pfizer, Inc., which has an interest in the clinical development of PF-06650833, when these studies were performed. Saurav De is now an employee of Pfizer, Inc. SA, VP, and BJB declare no conflicts of interest.
Footnotes
ClinicalTrials.gov identifier: NCT02485769
Data-Sharing Statement: Upon request, and subject to review, Pfizer will provide the data that support the findings of this study. Subject to certain criteria, conditions, and exceptions, Pfizer may also provide access to the related individual anonymized participant data. See https://www.pfizer.com/science/clinical-trials/trial-data-and-results for more information.
References:
- 1.Malmström V, Catrina AI, and Klareskog L. The immunopathogenesis of seropositive rheumatoid arthritis: from triggering to targeting. Nature Reviews Immunology. 2016;17:60. [DOI] [PubMed] [Google Scholar]
- 2.Smolen JS, Aletaha D, Barton A, Burmester GR, Emery P, Firestein GS, et al. Rheumatoid arthritis. Nature Reviews Disease Primers. 2018;4:18001. [DOI] [PubMed] [Google Scholar]
- 3.Aletaha D, and Smolen JS. Diagnosis and Management of Rheumatoid Arthritis: A ReviewDiagnosis and Management of Rheumatoid ArthritisDiagnosis and Management of Rheumatoid Arthritis. JAMA. 2018;320(13):1360–72. [DOI] [PubMed] [Google Scholar]
- 4.Smolen JS, and Aletaha D. Rheumatoid arthritis therapy reappraisal: strategies, opportunities and challenges. Nature Reviews Rheumatology. 2015;11:276. [DOI] [PubMed] [Google Scholar]
- 5.Dörner T, and Furie R. Novel paradigms in systemic lupus erythematosus. The Lancet. 2019;393(10188):2344–58. [DOI] [PubMed] [Google Scholar]
- 6.Tsokos GC, Lo MS, Reis PC, and Sullivan KE. New insights into the immunopathogenesis of systemic lupus erythematosus. Nature Reviews Rheumatology. 2016;12:716. [DOI] [PubMed] [Google Scholar]
- 7.Cushing L, Winkler A, Jelinsky SA, Lee K, Korver W, Hawtin R, et al. IRAK4 kinase activity controls Toll-like receptor–induced inflammation through the transcription factor IRF5 in primary human monocytes. Journal of Biological Chemistry. 2017;292(45):18689–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.De S, Karim F, Kiessu E, Cushing L, Lin LL, Ghandil P, et al. Mechanism of dysfunction of human variants of the IRAK4 kinase and a role for its kinase activity in interleukin-1 receptor signaling. The Journal of biological chemistry. 2018;293(39):15208–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Balka KR, and De Nardo D. Understanding early TLR signaling through the Myddosome. Journal of leukocyte biology. 2019;105(2):339–51. [DOI] [PubMed] [Google Scholar]
- 10.Suzuki N, Suzuki S, Duncan GS, Millar DG, Wada T, Mirtsos C, et al. Severe impairment of interleukin-1 and Toll-like receptor signalling in mice lacking IRAK-4. Nature. 2002;416(6882):750–6. [DOI] [PubMed] [Google Scholar]
- 11.Kim TW, Staschke K, Bulek K, Yao J, Peters K, Oh KH, et al. A critical role for IRAK4 kinase activity in Toll-like receptor-mediated innate immunity. The Journal of experimental medicine. 2007;204(5):1025–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Koziczak-Holbro M, Littlewood-Evans A, Pöllinger B, Kovarik J, Dawson J, Zenke G, et al. The critical role of kinase activity of interleukin-1 receptor-associated kinase 4 in animal models of joint inflammation. Arthritis and rheumatism. 2009;60(6):1661–71. [DOI] [PubMed] [Google Scholar]
- 13.Nanda SK, Lopez-Pelaez M, Arthur JS, Marchesi F, and Cohen P. Suppression of IRAK1 or IRAK4 Catalytic Activity, but Not Type 1 IFN Signaling, Prevents Lupus Nephritis in Mice Expressing a Ubiquitin Binding-Defective Mutant of ABIN1. Journal of immunology (Baltimore, Md : 1950). 2016;197(11):4266–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kawagoe T, Sato S, Jung A, Yamamoto M, Matsui K, Kato H, et al. Essential role of IRAK-4 protein and its kinase activity in Toll-like receptor-mediated immune responses but not in TCR signaling. The Journal of experimental medicine. 2007;204(5):1013–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Picard C, Puel A, Bonnet M, Ku CL, Bustamante J, Yang K, et al. Pyogenic bacterial infections in humans with IRAK-4 deficiency. Science (New York, NY). 2003;299(5615):2076–9. [DOI] [PubMed] [Google Scholar]
- 16.Kelly PN, Romero DL, Yang Y, Shaffer AL 3rd, Chaudhary D, Robinson S, et al. Selective interleukin-1 receptor-associated kinase 4 inhibitors for the treatment of autoimmune disorders and lymphoid malignancy. The Journal of experimental medicine. 2015;212(13):2189–201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chaudhary D, Robinson S, and Romero DL. Recent advances in the discovery of small molecule inhibitors of interleukin-1 receptor-associated kinase 4 (IRAK4) as a therapeutic target for inflammation and oncology disorders. J Med Chem. 2015;58(1):96–110. [DOI] [PubMed] [Google Scholar]
- 18.Lee KL, Ambler CM, Anderson DR, Boscoe BP, Bree AG, Brodfuehrer JI, et al. Discovery of Clinical Candidate 1-{[(2S,3S,4S)-3-Ethyl-4-fluoro-5-oxopyrrolidin-2-yl]methoxy}−7-methoxyisoquinoline-6-carboxamide (PF-06650833), a Potent, Selective Inhibitor of Interleukin-1 Receptor Associated Kinase 4 (IRAK4), by Fragment-Based Drug Design. Journal of Medicinal Chemistry. 2017;60(13):5521–42. [DOI] [PubMed] [Google Scholar]
- 19.Dudhgaonkar S, Ranade S, Nagar J, Subramani S, Prasad DS, Karunanithi P, et al. Selective IRAK4 Inhibition Attenuates Disease in Murine Lupus Models and Demonstrates Steroid Sparing Activity. Journal of immunology (Baltimore, Md : 1950). 2017;198(3):1308–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Corzo CA, Varfolomeev E, Setiadi AF, Francis R, Klabunde S, Senger K, et al. The kinase IRAK4 promotes endosomal TLR and immune complex signaling in B cells and plasmacytoid dendritic cells. Science signaling. 2020;13(634). [DOI] [PubMed] [Google Scholar]
- 21.Qin J, Jiang Z, Qian Y, Casanova JL, and Li X. IRAK4 kinase activity is redundant for interleukin-1 (IL-1) receptor-associated kinase phosphorylation and IL-1 responsiveness. The Journal of biological chemistry. 2004;279(25):26748–53. [DOI] [PubMed] [Google Scholar]
- 22.Song KW, Talamas FX, Suttmann RT, Olson PS, Barnett JW, Lee SW, et al. The kinase activities of interleukin-1 receptor associated kinase (IRAK)-1 and 4 are redundant in the control of inflammatory cytokine expression in human cells. Molecular immunology. 2009;46(7):1458–66. [DOI] [PubMed] [Google Scholar]
- 23.Cushing L, Stochaj W, Siegel M, Czerwinski R, Dower K, Wright Q, et al. Interleukin 1/Toll-like Receptor-induced Autophosphorylation Activates Interleukin 1 Receptor-associated Kinase 4 and Controls Cytokine Induction in a Cell Type-specific Manner. Journal of Biological Chemistry. 2014;289(15):10865–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chiang EY, Yu X, and Grogan JL. Immune complex-mediated cell activation from systemic lupus erythematosus and rheumatoid arthritis patients elaborate different requirements for IRAK1/4 kinase activity across human cell types. Journal of immunology (Baltimore, Md : 1950). 2011;186(2):1279–88. [DOI] [PubMed] [Google Scholar]
- 25.Sun J, Li N, Oh KS, Dutta B, Vayttaden SJ, Lin B, et al. Comprehensive RNAi-based screening of human and mouse TLR pathways identifies species-specific preferences in signaling protein use. Science signaling. 2016;9(409):ra3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Danto SI, Shojaee N, Singh RSP, Li C, Gilbert SA, Manukyan Z, et al. Safety, tolerability, pharmacokinetics, and pharmacodynamics of PF-06650833, a selective interleukin-1 receptor-associated kinase 4 (IRAK4) inhibitor, in single and multiple ascending dose randomized phase 1 studies in healthy subjects. Arthritis research & therapy. 2019;21(1):269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.O’Mahony A, John MR, Cho H, Hashizume M, and Choy EH. Discriminating phenotypic signatures identified for tocilizumab, adalimumab, and tofacitinib monotherapy and their combinations with methotrexate. Journal of translational medicine. 2018;16(1):156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Shah F, Stepan AF, O’Mahony A, Velichko S, Folias AE, Houle C, et al. Mechanisms of Skin Toxicity Associated with Metabotropic Glutamate Receptor 5 Negative Allosteric Modulators. Cell chemical biology. 2017;24(7):858–69.e5. [DOI] [PubMed] [Google Scholar]
- 29.Rankin AL, Seth N, Keegan S, Andreyeva T, Cook TA, Edmonds J, et al. Selective inhibition of BTK prevents murine lupus and antibody-mediated glomerulonephritis. Journal of immunology (Baltimore, Md : 1950). 2013;191(9):4540–50. [DOI] [PubMed] [Google Scholar]
- 30.Yao Y, Higgs BW, Richman L, White B, and Jallal B. Use of type I interferon-inducible mRNAs as pharmacodynamic markers and potential diagnostic markers in trials with sifalimumab, an anti-IFNα antibody, in systemic lupus erythematosus. Arthritis research & therapy. 2010;12 Suppl 1(Suppl 1):S6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Patricelli MP, Nomanbhoy TK, Wu J, Brown H, Zhou D, Zhang J, et al. In situ kinase profiling reveals functionally relevant properties of native kinases. Chemistry & biology. 2011;18(6):699–710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Morgan P, Van Der Graaf PH, Arrowsmith J, Feltner DE, Drummond KS, Wegner CD, et al. Can the flow of medicines be improved? Fundamental pharmacokinetic and pharmacological principles toward improving Phase II survival. Drug Discovery Today. 2012;17(9):419–24. [DOI] [PubMed] [Google Scholar]
- 33.Castelar-Pinheiro GdR, and Xavier RM. The Spectrum and Clinical Significance of Autoantibodies in Rheumatoid Arthritis. Frontiers in Immunology. 2015;6(320). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Catrina AI, Svensson CI, Malmström V, Schett G, and Klareskog L. Mechanisms leading from systemic autoimmunity to joint-specific disease in rheumatoid arthritis. Nature Reviews Rheumatology. 2016;13:79. [DOI] [PubMed] [Google Scholar]
- 35.Sokolove J, Zhao X, Chandra PE, and Robinson WH . Immune complexes containing citrullinated fibrinogen costimulate macrophages via Toll-like receptor 4 and Fcγ receptor. Arthritis & Rheumatism. 2011;63(1):53–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Horwood NJ, Page TH, McDaid JP, Palmer CD, Campbell J, Mahon T, et al. Bruton’s tyrosine kinase is required for TLR2 and TLR4-induced TNF, but not IL-6, production. Journal of immunology (Baltimore, Md : 1950). 2006;176(6):3635–41. [DOI] [PubMed] [Google Scholar]
- 37.Falconer J, Murphy AN, Young SP, Clark AR, Tiziani S, Guma M, et al. Review: Synovial Cell Metabolism and Chronic Inflammation in Rheumatoid Arthritis. Arthritis & Rheumatology. 2018;70(7):984–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Doody KM, Bottini N, and Firestein GS. Epigenetic alterations in rheumatoid arthritis fibroblast-like synoviocytes. Epigenomics. 2017;9(4):479–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pisetsky DS. Anti-DNA antibodies — quintessential biomarkers of SLE. Nature Reviews Rheumatology. 2015;12:102. [DOI] [PubMed] [Google Scholar]
- 40.Lee KH, Kronbichler A, Park DD-Y, Park Y, Moon H, Kim H, et al. Neutrophil extracellular traps (NETs) in autoimmune diseases: A comprehensive review. Autoimmunity Reviews. 2017;16(11):1160–73. [DOI] [PubMed] [Google Scholar]
- 41.Bouts YM, Wolthuis DFGJ, Dirkx MFM, Pieterse E, Simons EMF, Van Boekel AM, et al. Apoptosis and NET formation in the pathogenesis of SLE. Autoimmunity. 2012;45(8):597–601. [DOI] [PubMed] [Google Scholar]
- 42.Mahajan A, Herrmann M, and Muñoz LE. Clearance Deficiency and Cell Death Pathways: A Model for the Pathogenesis of SLE. Frontiers in Immunology. 2016;7(35). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Muñoz LE, Janko C, Schulze C, Schorn C, Sarter K, Schett G, et al. Autoimmunity and chronic inflammation — Two clearance-related steps in the etiopathogenesis of SLE. Autoimmunity Reviews. 2010;10(1):38–42. [DOI] [PubMed] [Google Scholar]
- 44.Crow MK. Type I Interferon in the Pathogenesis of Lupus. The Journal of Immunology. 2014;192(12):5459–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Stone RC, Feng D, Deng J, Singh S, Yang L, Fitzgerald-Bocarsly P, et al. Interferon regulatory factor 5 activation in monocytes of systemic lupus erythematosus patients is triggered by circulating autoantigens independent of type I interferons. Arthritis & Rheumatism. 2012;64(3):788–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kiefer K, Oropallo MA, Cancro MP, and Marshak-Rothstein A. Role of type I interferons in the activation of autoreactive B cells. Immunology & Cell Biology. 2012;90(5):498–504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Douagi I, Gujer C, Sundling C, Adams WC, Smed-Sörensen A, Seder RA, et al. Human B Cell Responses to TLR Ligands Are Differentially Modulated by Myeloid and Plasmacytoid Dendritic Cells. The Journal of Immunology. 2009;182(4):1991–2001. [DOI] [PubMed] [Google Scholar]
- 48.Cushing L, Winkler A, Jelinsky SA, Lee K, Korver W, Hawtin R, et al. IRAK4 kinase activity controls Toll-like receptor-induced inflammation through the transcription factor IRF5 in primary human monocytes. The Journal of biological chemistry. 2017;292(45):18689–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Colonna M, Trinchieri G, and Liu Y-J. Plasmacytoid dendritic cells in immunity. Nature Immunology. 2004;5:1219. [DOI] [PubMed] [Google Scholar]
- 50.Zhuang H, Szeto C, Han S, Yang L, and Reeves WH. Animal Models of Interferon Signature Positive Lupus. Frontiers in Immunology. 2015;6(291). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Homer BL, and Dower K. 41-Week Study of Progressive Diabetic Nephropathy in the ZSF1 fa/fa(CP) Rat Model. Toxicologic pathology. 2018;46(8):976–7. [DOI] [PubMed] [Google Scholar]
- 52.Honigberg LA, Smith AM, Sirisawad M, Verner E, Loury D, Chang B, et al. The Bruton tyrosine kinase inhibitor PCI-32765 blocks B-cell activation and is efficacious in models of autoimmune disease and B-cell malignancy. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(29):13075–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kim YY, Park KT, Jang SY, Lee KH, Byun JY, Suh KH, et al. HM71224, a selective Bruton’s tyrosine kinase inhibitor, attenuates the development of murine lupus. Arthritis research & therapy. 2017;19(1):211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Chalmers SA, Wen J, Doerner J, Stock A, Cuda CM, Makinde HM, et al. Highly selective inhibition of Bruton’s tyrosine kinase attenuates skin and brain disease in murine lupus. Arthritis research & therapy. 2018;20(1):10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Shi FD, Ljunggren HG, and Sarvetnick N. Innate immunity and autoimmunity: from self-protection to self-destruction. Trends in immunology. 2001;22(2):97–101. [DOI] [PubMed] [Google Scholar]
- 56.Waldner H The role of innate immune responses in autoimmune disease development. Autoimmun Rev. 2009;8(5):400–4. [DOI] [PubMed] [Google Scholar]
- 57.De Nardo D, Balka KR, Cardona Gloria Y, Rao VR, Latz E, and Masters SL. Interleukin-1 receptor–associated kinase 4 (IRAK4) plays a dual role in myddosome formation and Toll-like receptor signaling. Journal of Biological Chemistry. 2018;293(39):15195–207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Pauls E, Nanda SK, Smith H, Toth R, Arthur JSC, and Cohen P. Two Phases of Inflammatory Mediator Production Defined by the Study of IRAK2 and IRAK1 Knock-in Mice. The Journal of Immunology. 2013;191(5):2717–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Lazzari E, and Jefferies CA. IRF5-mediated signaling and implications for SLE. Clin Immunol. 2014;153(2):343–52. [DOI] [PubMed] [Google Scholar]
- 60.Ban T, Sato GR, and Tamura T. Regulation and role of the transcription factor IRF5 in innate immune responses and systemic lupus erythematosus. Int Immunol. 2018;30(11):529–36. [DOI] [PubMed] [Google Scholar]
- 61.Cushing L, Stochaj W, Siegel M, Czerwinski R, Dower K, Wright Q, et al. Interleukin 1/Toll-like receptor-induced autophosphorylation activates interleukin 1 receptor-associated kinase 4 and controls cytokine induction in a cell type-specific manner. The Journal of biological chemistry. 2014;289(15):10865–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Crow MK. Type I interferon in the pathogenesis of lupus. Journal of immunology. 2014;192(12):5459–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Otterness IG. The value of C-reactive protein measurement in rheumatoid arthritis. Semin Arthritis Rheum. 1994;24(2):91–104. [DOI] [PubMed] [Google Scholar]
- 64.Tishler M, Caspi D, and Yaron M. C-reactive protein levels in patients with rheumatoid arthritis: the impact of therapy. Clin Rheumatol. 1985;4(3):321–4. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.