

Abstract
An accurate description of changes in the brain in healthy aging is needed to understand the basis of age-related changes in cognitive function. Cross-sectional magnetic resonance imaging (MRI) studies suggest thinning of the cerebral cortex, volumetric reductions of most subcortical structures and ventricular expansion. However, there is a paucity of detailed longitudinal studies to support the cross-sectional findings. In the present study, 142 healthy elderly participants (60–91 years) were followed with repeated MRI, and were compared to 122 patients with mild to moderate Alzheimer's disease (AD). Volume changes were measured across the entire cortex and in 48 regions of interest (ROIs). Cortical reductions in the healthy elderly were extensive after only one year, especially evident in temporal and prefrontal cortex where annual decline was about 0.5%. All subcortical and ventricular regions except caudate nucleus and the 4th ventricle changed significantly over one year. Some of the atrophy occurred in areas vulnerable to AD, while other changes were observed in areas less characteristic of the disease in early stages. This suggests that the changes are not primarily driven by degenerative processes associated with AD, although it is likely that preclinical changes associated with AD are superposed on changes due to normal aging in some subjects, especially in the temporal lobes. Finally, atrophy was found to accelerate with increasing age, and this was especially prominent in areas vulnerable to AD. Thus, it is possible that the accelerating atrophy with increasing age is due to preclinical AD.
Keywords: MRI, aging, longitudinal, ADNI, cerebral cortex, hippocampus
Introduction
An accurate description of age-related changes in the brain is needed to better understand the neurobiological foundation of cognitive changes in healthy aging. While cross-sectional magnetic resonance imaging (MRI) studies have demonstrated consistent age-effects (Allen et al., 2005; Fjell et al., 2009-a, b; Raz et al., 2004; Raz and Rodrigue, 2006; Salat et al., 2004; Walhovd et al., 2005; Westlye et al., 2009), longitudinal MR studies have often been limited by relatively small numbers of participants and use of either global measures (e.g. whole-brain volume) or only a restricted set of regions of interest (ROIs). Further, while atrophy has been demonstrated in mild cognitive impairment over a year or less (Schuff et al., 2009), it is not known whether brain changes driven solely by normal aging are large enough to be detectable over such short time intervals.
Longitudinal studies have reported annual gross brain volume decreases on the order of 0.2 – 0.5% (Enzinger et al., 2005; Ezekiel et al., 2004; Fotenos et al., 2005; Scahill et al., 2003), while hippocampus, the most studied structure, shows annual atrophy rates from 0.79% to 2.0% (Barnes et al., 2009; Du et al., 2006; Ezekiel et al., 2004; Jack et al., 1998; Raz et al., 2005b; Scahill et al., 2003). This tends to be higher than cross-sectional estimates (Du et al., 2006; Raz et al., 2005a; Scahill et al., 2003). Entorhinal cortex longitudinal decline is about 0.3% to 2.4% (Du et al., 2006; Du et al., 2003; Ezekiel et al., 2004; Raz et al., 2005a). Atrophy rates in other parts of the brain are seldom studied, with a few important exceptions (Driscoll et al., 2009; Pfefferbaum et al., 1998; Raz et al., 2005a; Resnick et al., 2003). These studies describe atrophy in several regions, with prominent decline in prefrontal cortex, as well as decline in caudate nucleus, cerebellum, hippocampus and association cortices (Raz et al., 2005b) and parietal areas (Resnick et al., 2003), in addition to expansion of the ventricles (Pfefferbaum et al., 1998). In a recent study, significant atrophy was found in all 25 regional brain volumes tested, but over time periods extending as long as ten years in some cases (Driscoll et al., 2009). An earlier study found significant ventricular enlargement over one year, but no detectable change in total or regional brain volumes (Resnick et al., 2000).
With newer analyses techniques, we may now possess the tools necessary to detect brain changes over short time intervals in healthy elderly. The aims of the present study were to determine whether brain changes in normal aging can be detected over one (n = 142) or two (n = 89) years, and if so, to determine which areas show the greatest changes, and whether rate of atrophy increases with age. A novel procedure for longitudinal change analysis was used, which enabled measures of change continuously across the cortex, as well as in 48 regions of interest. Atrophy in the healthy elderly was compared to 122 patients with mild to moderate Alzheimer's disease (AD).
Materials and Methods
The raw data used in the preparation of this article were obtained from the ADNI database (www.loni.ucla.edu/ADNI). ADNI was launched in 2003 by the National Institute on Aging (NIA), the National Institute of Biomedical Imaging and Bioengineering (NIBIB), the Food and Drug Administration (FDA), private pharmaceutical companies and non-profit organizations. The primary goal of ADNI has been to test whether serial MRI, PET, other biological markers, and clinical and neuropsychological assessment can be combined to measure the progression of MCI and early AD. The Principal Investigator of this initiative is Michael W. Weiner, VA Medical Center and University of California — San Francisco. There are many co-investigators, and subjects have been recruited from over 50 sites across the U.S. and Canada. The initial goal of ADNI was to recruit 800 adults, including healthy elderly, MCI and AD patients to participate and be followed for 2–3 years. For more information see www.adni-info.org.
Sample
ADNI eligibility criteria are described at http://www.adni-info.org/index.php?option=com_content&task=view&id=9&Itemid=43. Briefly, participants are 55–90 years of age, had an informant able to provide an independent evaluation of functioning, and spoke either English or Spanish. All subjects were willing and able to undergo test procedures including neuroimaging and agreed to longitudinal follow up. Specific psychoactive medications are excluded. General inclusion/exclusion criteria are as follows for normal subjects: Mini-Mental State Examination (MMSE) (Folstein et al., 1975) scores between 24–30 (inclusive), Clinical Dementia Rating (CDR) (Morris, 1993) of 0, non-depressed, non-MCI, and nondemented. In addition, CDR sum of boxes (CDR-sb) was calculated as a measure of clinical functioning (does not need to be zero). To minimize the possibility of including individuals with early, preclinical AD, only participants who had the same or better score on CDR-sb at the time of follow up were included. The subject pool was further restricted to those subjects for whom adequate processed and quality checked MR data were available by February 2009. Sample characteristics are presented in Table 1.
Table 1.
Sample characteristics for the healthy elderly group
| Baseline | 1 year | 2 years | |
|---|---|---|---|
| n | 142 | 142 | 89 |
| Sex (females/males) | 68/ 74 | 68/ 74 | 38/ 51 |
| Age | 75.6 (59.8–90.2) | 76.7 (60.8–91.3) | 78.7 (65.1–92.3) |
| MMSE | 29.2 (25–30) | 29.1 (24–30) | 29.1 (26–30) |
| CDR sum of boxes | 0.03 (0–0.5) | 0 (0–0) | 0 (0–0) |
To be able to compare atrophy in the healthy elderly with AD patients, an AD group was included for selected analyses (n = 122, 57 females/ 65 males, at 1 year follow up: age = 76.5 (56.0–90.0), MMSE = 20.7 (8–29), CDR = 1.0 (0.5–3.0), CDR-sb = 5.88 (1–14)). These patients met the same eligibility criteria as the healthy elderly, had CDR scores of 0.5 or more, and met NINCDS/ ADRDA criteria for probable AD (McKhann et al., 1984). As the mean age (76.5 vs. 76.7 years, respectively) and the female/ male ratio (0.88 vs. 0.92) was almost identical between AD patients and healthy elderly, these variables were not regressed out in the analyses.
MR acquisition and analysis
All scans used for the present paper were from 1.5 T scanners. Data were collected across a variety of scanners with protocols individualized for each scanner, as defined at http://www.loni.ucla.edu/ADNI/Research/Cores/index.shtml. Raw DICOM MRI scans (including two T1-weighted volumes per case) were downloaded from the public ADNI site (http://www.loni.ucla.edu/ADNI/Data/index.shtml) and processed as described elsewhere (Fennema-Notestine et al., 2009). Briefly, these data were reviewed for quality, automatically corrected for spatial distortion due to gradient nonlinearity (Jovicich et al., 2006) and B1 field inhomogeneity (Sled et al., 1998), registered, and averaged to improve signal-to-noise. Scans were segmented as described previously (Fischl et al., 2002, 2004) to yielding data for 11 different volumetric ROIs and four ventricular ROIs (right and left hemisphere regions measured separately), including the hippocampal formation (consisting of the dentate gyrus, CA fields, subiculum/ parasubiculum and the fimbria (Makris et al., 1999)). The cortical surface was reconstructed using a semi-automated approach described elsewhere (Dale et al., 1999; Dale, 1993; Fischl and Dale, 2000; Fischl et al., 1999a; Fischl et al., 1999b; Salat et al., 2004), and parcellated as described in Fischl et al. (Fischl et al., 2004), labelling 33 cortical sulci and gyri (Desikan et al., 2006). The exact anatomical borders for the cortical ROIs can be found in Desikan et al., e.g. for the transverse temporal cortex (the rostral boundary was the rostral extent of the transverse temporal sulcus, the caudal boundary was the caudal portion of the insular cortex, and the lateral fissure and the superior temporal gyrus were utilized as the medial and lateral boundaries, respective) and isthmus of the cingulate cortex (the rostral and caudal boundaries were the posterior division of the cingulate cortex and the parahippocampal gyrus, respectively, and the medial and lateral boundaries were the medial wall (area unknown) and the precuneus, respectively). These surface-based cortical ROIs were thus generated by a different procedure from that used to estimate the other (volumetric) ROIs, which included the hippocampus, the cerebellum GM, nine subcortical ROIs and four ventricular ROIs.
For the analyses of the longitudinal volume changes, each participant's dual 3-D follow-up structural scans were rigid-body aligned, averaged and affine aligned to the baseline scan. Nonlinear registration of the images was then performed, where voxel centers are moved about until a good match between the images is made. Several methods exist for effecting this, including fluid deformation (Christensen et al., 1996; Freeborough and Fox, 1998; Miller et al., 1993) and tensor-based morphometry (TBM) (Ashburner et al., 1999). For the results presented here, however, we applied a method (Holland et al., 2008) based on linear elasticity and closer in spirit to TBM. It proceeds essentially as follows. The images are heavily blurred (smoothed), making them almost identical, and a merit or potential function calculated. This merit function expresses the intensity difference between the images at each voxel, and depends on the displacement field for the voxel centers of the image being transformed; it is also regularized to keep the displacement field spatially smooth. The merit function by design will have a minimum when the displacement field induces a good match between the images. The displacement field in general will turn cubic voxels into displaced irregular hexahedra whose volumes (Grandy, 1997) give the volume change field. The merit function is minimized efficiently using standard numerical methods. Having found a displacement field for the heavily blurred pair of images, the blurring is reduced and the procedure repeated, thus iteratively building up a better displacement field. Two important additions to this are: applying the final displacement field to the image being transformed, then nonlinearly registering the resultant image to the same target, and finally tracing back through the displacement fields thus calculated to find the net displacement field; and restricting to regions of interest and zooming when tissue structures are separated by only a voxel or two. These additional features enable very precise registration involving large or subtle deformations, even at small spatial scales with low boundary contrast. Although large deformations are allowed by multiple nonlinear registration (or relaxation) steps, nonphysical deformations are precluded because at each level of blurring the image undergoing deformation is constrained to conform to the target. Note that calculating the deformation field does not depend on initially segmenting tissue. This deformation field was used to align scans at the sub-voxel level.
The follow-up aligned image underwent skull stripping and volumetric segmentations (subcortical structures, as well as hippocampus and cerebellum GM), with labels applied from the baseline scan. For the cortical reconstructions, surface coordinates for the white matter and pial boundaries were derived from the baseline images and mapped onto the follow-up images using the deformation field. Parcellations from the baseline image were then applied to the follow-up image. This resulted in a one-to-one correspondence between each vertex in the base image and the follow up image. The procedure produces an estimate of the percent cortical volume loss at each vertex and within each ROI. To the extent that regional cortical areas are relatively stable across time points, the volume change is likely driven almost exclusively by changes in thickness. The method has been validated in model studies of complex spherical-shell geometries with low contrast and noise, where a prescribed volume change is numerically estimated to accuracies of within 0.5% (Holland et al., work in progress).
A challenge in cross-sectional MR studies of aging is that the contrast between GM and WM decreases with higher age (Salat et al., 2009). However, the longitudinal registration relies on matching intensities in serial images. A reduction in GM/ WM contrast in elderly relative to younger participants should not impact the quality of the nonlinear registration, as long as the serial images have constant contrast. Change in cortical volume to 1st order arises from change in cortical thickness. To estimate it, the volume change field is sampled midway through the cortical layer. Conceivably, the primary source of error in elderly, relatively to younger participants, would then be in the initial delineation of the cortical surface. However, it has recently been shown that the decrease in GM/ WM contrast can not explain the differences in cortical thickness between older and younger participants (Westlye et al., 2009).
Statistical analyses
Statistical analyses were performed by use of FreeSurfer (http://surfer.nmr.mgh.harvard.edu/) and SPSS 15.0. To reduce the number of comparisons, mean values of the left and right hemisphere were used for the ROI analyses. For the healthy elderly and the AD patients, change in brain morphometry was calculated as percentage change relative to baseline (1 and 2 years for the healthy elderly, 2 years for the AD patients). For the healthy elderly, statistical parametric continuous surface maps were computed by general linear models (GLM) implemented in FreeSurfer and displayed on a semi-inflated template brain. The results were thresholded by a conventional criterion for correction for multiple comparisons (false discovery rate [FDR] < .05). For the healthy elderly and the AD patients, one-sample t-tests were used to test whether longitudinal atrophy rates within each ROI were different from zero. Independent-samples t-tests were used to compare rate of atrophy in healthy controls and AD patients. To visualize the areas that showed more than average and the areas that showed less than average atrophy as surface maps, mean cortical atrophy was calculated for each hemisphere in each group. This value was subtracted from the original surface maps, yielding a new map showing relative atrophy. To test whether the magnitude of atrophy in the temporal lobe vs. the prefrontal cortex differed between the healthy elderly and the AD patients, ANOVA with 2 groups × 2 regions were performed. Temporal lobe atrophy was calculated as mean rate of atrophy in inferior and middle temporal, entorhinal, parahippocampal, temporal pole, and fusiform cortex, as well as hippocampus and amygdala. Prefrontal atrophy was calculated as mean rate of atrophy in caudal and rostral anterior cingulate, caudal and rostral middle frontal, pars opercularis, triangularis and orbitalis (together forming the inferior frontal cortex), lateral, dorsal and medial orbitofrontal, frontal pole and superior frontal cortex (see Figure 4). Finally, Pearson correlations were performed between age and atrophy in each of the 48 ROIs in the healthy elderly.
Figure 4. Frontal vs. temporal effects of healthy aging and Alzheimer's disease.
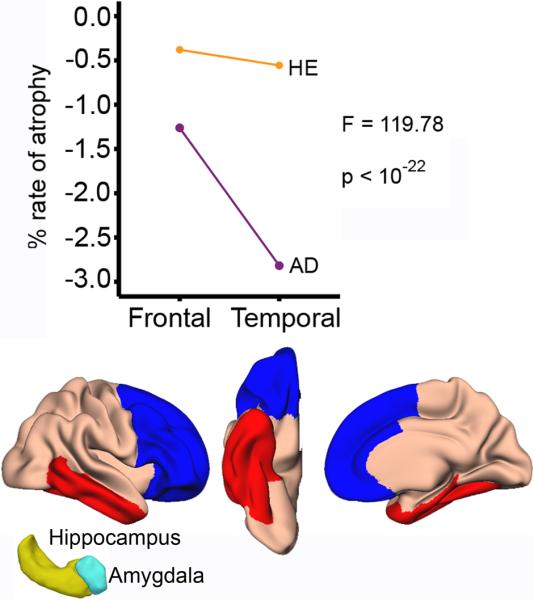
The line graphs show rate of atrophy for the frontal and the temporal regions in healthy elderly and AD patients. The regions are shown below (blue: frontal, red: temporal). In addition to the cortical areas shown, hippocampus and amygdala were included in the temporal region (yellow and cyan 3D figures). ANOVA showed a significant interaction between group and region: While the AD patients showed more atrophy in the temporal (−2.82%, SE = 0.14) than the frontal regions (−1.26%, SE = 0.11), the rate of atrophy between these two regions was comparable in the healthy controls (temporal: −0.56%, SE = 0.07/frontal: −0.38%, SE = 0.07).
Results
Atrophy over 1 and 2 years: Continuous surface analyses
The statistical significance of 1 and 2 year volume changes in cortex across the brain surface is shown in Figure 1. As can be seen, large cortical areas decreased significantly over 1 year. These reductions were most prominent in the temporal lobe, the medial part of the superior frontal gyrus, orbitofrontal, precuneus, supramarginal and inferior parietal, as well as the inferior and part of the middle frontal cortex. Lateral parts of the superior frontal cortex, as well as pre- and post-central gyrus, superior parietal areas, occipital cortex and lingual gyrus were largely spared. Over two years, the effects were stronger and spread to additional areas, involving all cortical areas except lingual cortex and pre- and postcentral gyrus.
Figure 1. Cortical atrophy in healthy elderly.

General linear models were used to test whether rates of atrophy for one and two years were significantly different from zero. The results were thresholded by a conventional criterion for correction for multiple comparisons (false discovery rate < .05) and projected onto a semi-inflated template brain. Blue-cyan areas indicate cortical reduction, while red-yellow areas indicate expansion. As can be seen, significant volume reductions are found across large areas already after one year, and these were especially prominent in temporal and prefrontal areas.
Percentage volume change in cortex over 1 and 2 years is shown in Figure 2. Annual atrophy rates varied across the cortex, but were typically around 0.5 % in affected areas, including most of the lateral and inferior parts of the temporal lobes, supramarginal gyrus, inferior parietal gyrus, precuneus, inferior and middle frontal gyrus as well as the medial parts of the superior frontal gyrus. Atrophy over 2 years exceeded 1.0 % in large areas.
Figure 2. Rate of change in healthy elderly and AD patients.

The left panel shows annual longitudinal atrophy in healthy elderly calculated as percentage volume change relative to baseline, while the middle panel shows longitudinal atrophy over two years. Annual change of about 0.5% can be seen across large sections of the brain surface, e.g. in temporal and prefrontal areas. The right panel shows 1 year atrophy in AD patients. The areas of change in the healthy elderly are overlapping with areas most strongly affected in Alzheimer's disease, e.g. temporal lobe cortex. However, areas that are not especially affected in AD, e.g. prefrontal cortex, also undergo large changes in healthy aging. Please note that to allow optimal visualization of regional variation, the color-scale is different for the HE and the AD groups.
For comparison purposes, rate of atrophy is presented for AD patients in Figure 2. Atrophy in the AD group was much larger than in the healthy elderly, with most areas showing 1% annual change or more. This was especially evident in the lateral temporal cortex and an area around posterior cingulate/ inferior precuneus/ retrosplenial cortex (> 2.5% annual change). Less atrophy was seen around the central sulcus, pericalcarine sulcus and lingual gyrus. To compare the pattern of atrophy between the healthy controls and the AD patients, mean cortical atrophy was calculated for each hemisphere in each group. This value was subtracted from the surface maps, yielding a new map highlighting areas that show greater and lesser rates of atrophy than average for that group. These maps illustrate the parts of the cortex that show relatively higher and relatively lower rates of atrophy within each group (Figure 3). Within the AD group, higher than average atrophy was seen in the inferior and middle temporal gyrus. Within the healthy elderly group, higher than average atrophy was observed in the same areas, but also in the temporal pole, medial orbitofrontal, rostral middle frontal and supramarginal cortex, as well as parts of the medial superior frontal gyrus. Less than average atrophy was seen in cuneus, pericalcarine sulcus, lingual gyrus as well as the central sulcus. Results of formal statistical tests of differences in rates of atrophy between healthy controls and AD patients are described below.
Figure 3. Regional rates of atrophy relative to mean atrophy within group for healthy elderly and AD.

To visualize the areas that showed more than average atrophy and the areas that showed less than average atrophy within each group, mean cortical volume reduction was calculated for each hemisphere in each group. This value was subtracted from the surface maps, yielding a new map highlighting areas that show greater (red) and lesser (blue) rates of atrophy than average for that group. As can be seen, in healthy elderly (left panel) higher than average volume reductions are seen in lateral temporal, inferior parietal, supramarginal and frontal cortex. In contrast, the AD patients show higher than average atrophy mainly in the temporal lobe (middle and inferior temporal gyrus, fusiform gyrus). Please note that to allow optimal visualization of regional variation, the color-scale is different for the HE and the AD groups.
Atrophy over 1 and 2 years: ROI analyses
Percentage annual volume change in each of the 48 ROIs is shown in Table 2 for healthy elderly. Median atrophy for 1 year was −0.38 %, not including the ventricular ROIs. Hippocampus (−0.84 %) and amygdala (−0.81 %) showed the largest reductions, followed by thalamus (−0.69 %). Cerebellum grey matter (−0.35 %), the brain stem (−0.31 %) and caudate nucleus (−0.24 %) showed the least 1 year change of the volumetric ROIs. The inferior lateral (5.47 %), lateral (4.40 %) and the 3rd (3.07 %) ventricles showed the largest changes overall, with no significant change in the 4th (0.71 %) ventricle. Of the cortical ROIs, the frontal pole (−0.59 %), temporal pole (−0.56 %), banks of the superior temporal sulcus (−.55 %), and entorhinal cortex (−0.55 %) changed the most, while post central, pericalcarine, lingual, cuneus and precentral were well preserved (≤ 0.1 %).
Table 2.
Percentage volume change over 1 and 2 years in healthy elderly n.s. indicates p ≥ .01
| % change longitudinal | p < | % change longitudinal | p < | |
|---|---|---|---|---|
| 1 year | 2 years | |||
| Volumetric ROIs | ||||
| Accumbens | −0.57 | 10−8 | −1.40 | 10−19 |
| Amygdala | −0.81 | 10−9 | −2.02 | 10−13 |
| BrainStem | −0.31 | 10−5 | −0.86 | 10−12 |
| Caudate | −0.24 | n.s. | −0.62 | .005 |
| Cerebellum GM | −0.35 | 10−8 | −0.94 | 10−15 |
| Cerebellum WM | −0.57 | 10−14 | −1.22 | 10−20 |
| Cerebral WM | −0.58 | 10−16 | −1.15 | 10−19 |
| Hippocampus | −0.84 | 10−14 | −1.76 | 10−15 |
| Pallidum | −0.40 | 10−11 | −0.75 | 10−13 |
| Putamen | −0.43 | 10−8 | −0.92 | 10−16 |
| Thalamus | −0.69 | 10−16 | −1.42 | 10−21 |
| Ventricular ROIs | ||||
| Lat vent | 4.40 | 10−21 | 9.42 | 10−19 |
| Inf lat vent | 5.47 | 10−15 | 12.11 | 10−18 |
| 3rd ventricle | 3.07 | 10−12 | 6.60 | 10−14 |
| 4th ventricle | 0.71 | n.s. | 2.18 | 10−4 |
| Surface ROIs | ||||
| Cingulate, caudal ant | −0.27 | 0.001 | −0.91 | 10−12 |
| Cingulate, rostral ant | −0.34 | 10−4 | −0.92 | 10−12 |
| Cingulate, posterior | −0.25 | 0.001 | −0.79 | 10−7 |
| Cingulate, isthmus | −0.20 | 0.005 | −0.63 | 10−7 |
| Frontal, superior | −0.31 | 0.005 | −0.93 | 10−11 |
| Frontal, caudal middle | −0.40 | 10−6 | −0.99 | 10−12 |
| Frontal, rostral middle | −0.32 | 10−4 | −0.98 | 10−13 |
| Frontal, pars opercularis | −0.38 | 10−7 | −1.04 | 10−14 |
| Frontal, pars triangularis | −0.37 | 10−6 | −1.08 | 10−15 |
| Frontal, pars orbitalis | −0.38 | 10−4 | −1.11 | 10−13 |
| Frontal, lateral orbital | −0.42 | 10−6 | −1.07 | 10−16 |
| Frontal, medial orbital | −0.39 | 10−5 | −0.98 | 10−14 |
| Frontal, pole | −0.59 | 10−5 | −1.30 | 10−11 |
| Parietal, precentral gyrus | −0.10 | n.s. | −0.48 | 10−4 |
| Parietal, post central gyrus | 0.09 | n.s. | 0.01 | n.s. |
| Parietal, paracentral gyrus | −0.17 | n.s. | −0.52 | .001 |
| Parietal, superior | −0.18 | n.s. | −0.68 | 10−5 |
| Parietal, inferior | −0.40 | 10−7 | −1.03 | 10−13 |
| Parietal, supramarginal | −0.40 | 10−6 | −0.98 | 10−10 |
| Parietal, precuneus | −0.30 | 10−6 | −0.82 | 10−10 |
| Temporal, parahippocampal | −0.36 | 10−6 | −1.08 | 10−18 |
| Temporal, entorhinal | −0.55 | 10−7 | −1.20 | 10−11 |
| Temporal, pole | −0.56 | 10−5 | −1.17 | 10−9 |
| Temporal, superior | −0.43 | 10−7 | −1.00 | 10−13 |
| Temporal, middle | −0.47 | 10−8 | −1.25 | 10−16 |
| Temporal, inferior | −0.47 | 10−9 | −1.18 | 10−17 |
| Temporal, transverse | −0.31 | .005 | −0.79 | 10−8 |
| Temporal, Banks Sup Temp Sulc | −0.55 | 10−11 | −1.32 | 10−15 |
| Temporal, fusiform | −0.38 | 10−9 | −0.98 | 10−18 |
| Occipital, lateral | −0.19 | 0.005 | −0.66 | 10−9 |
| Occipital, pericalcarine | 0.11 | n.s. | −0.18 | n.s. |
| Occipital, lingual | −0.02 | n.s. | −0.35 | 10−4 |
| Occipital, cuneus | −0.04 | n.s. | −0.40 | 10−4 |
For comparison purposes, 1 year volume change in each ROI was calculated for the AD group also (Table 3). All ROIs except the pericalcarine sulcus showed significant change. Median atrophy for 1 year was −1.20 %, not including the ventricular ROIs. Amygdala (−3.79 %) and hippocampus (−3.75 %) largest reductions, followed by inferior and middle temporal gyri (−2.97 and −2.87%, respectively). Also entorhinal and parahippocampal cortex showed well above average atrophy (−2.42 and −2.05%, respectively). Pericalcarine (−0.16%), postcentral (−0.27%) and cuneus (−0.5%) were the three ROIs showing the least atrophy, but of these only pericalcarine did not change at p < .01 (p = .030). The inferior lateral (16.26 %), lateral (10.35 %) and the 3rd (6.82 %) ventricles showed the largest changes overall.
Table 3.
Percentage volume change over 1 year in the AD group n.s. indicates p ≥ .01
| % change longitudinal | p< | |
|---|---|---|
| 1 year | ||
| Volumetric ROIs | ||
| Accumbens | −2.02 | 10−19 |
| Amygdala | −3.79 | 10−19 |
| BrainStem | −0.65 | 10−16 |
| Caudate | −1.49 | 10−13 |
| Cerebellum GM | −0.99 | 10−25 |
| Cerebellum WM | −0.95 | 10−18 |
| Cerebral WM | −1.42 | 10−30 |
| Hippocampus | −3.75 | 10−35 |
| Pallidum | −0.90 | 10−15 |
| Putamen | −1.45 | 10−22 |
| Thalamus | −1.51 | 10−28 |
| Ventricular ROIs | ||
| Lat vent | 10.35 | 10−28 |
| Inf lat vent | 16.26 | 10−33 |
| 3rd ventricle | 6.82 | 10−21 |
| 4th ventricle | 1.61 | .001 |
| Surface ROIs | ||
| Cingulate, caudal ant | −1.14 | 10−17 |
| Cingulate, rostral ant | −1.20 | 10−18 |
| Cingulate, posterior | −1.57 | 10−21 |
| Cingulate, isthmus | −1.54 | 10−23 |
| Frontal, superior | −1.20 | 10−15 |
| Frontal, caudal middle | −1.60 | 10−20 |
| Frontal, rostral middle | −1.23 | 10−15 |
| Frontal, pars opercularis | −1.31 | 10−18 |
| Frontal, pars triangularis | −1.10 | 10−17 |
| Frontal, pars orbitalis | −1.06 | 10−10 |
| Frontal, lateral orbital | −1.47 | 10−18 |
| Frontal, medial orbital | −1.19 | 10−17 |
| Frontal, pole | −1.38 | 10−10 |
| Parietal, precentral gyrus | −0.69 | 10−10 |
| Parietal, post central gyrus | −0.27 | .01 |
| Parietal, paracentral gyrus | −0.76 | 10−11 |
| Parietal, superior | −1.26 | 10−18 |
| Parietal, inferior | −2.10 | 10−27 |
| Parietal, supramarginal | −1.74 | 10−21 |
| Parietal, precuneus | −1.61 | 10−24 |
| Temporal, parahippocampal | −2.05 | 10−34 |
| Temporal, entorhinal | −2.42 | 10−36 |
| Temporal, pole | −2.54 | 10−26 |
| Temporal, superior | −1.63 | 10−23 |
| Temporal, middle | −2.87 | 10−31 |
| Temporal, inferior | −2.97 | 10−32 |
| Temporal, transverse | −0.59 | 10−6 |
| Temporal, Banks Sup Temp Sulc | −2.53 | 10−28 |
| Temporal, fusiform | −2.15 | 10−33 |
| Occipital, lateral | −0.96 | 10−19 |
| Occipital, pericalcarine | −0.16 | n.s. |
| Occipital, lingual | −0.67 | 10−15 |
| Occipital, cuneus | −0.50 | 10−9 |
Differences in rates of atrophy were formally tested between healthy elderly and AD patients. AD patients had significantly (p < .01) higher rates of change than the healthy elderly for all ROIs except 4th ventricle (Supplementary Table 1). In terms of t-scores, inferior temporal (t = 13.25), fusiform (t = 13.15) and middle temporal (t = 12.81), parahippocampal (t = 12.69) and entorhinal (t = 11.93) showed the largest effects of the cortical ROIs, while hippocampus (t = 12.80), the inferior lateral ventricles (t = 9.74) and amygdala (t = 8.32) showed the largest effects for the volumetric ROIs.
Interaction between the groups and temporal vs. frontal atrophy rates were tested by ANOVA with two groups (healthy elderly, AD) × two brain regions (frontal, temporal). A highly significant interaction was found (F [1, 256] = 119.78, p < 10−22), caused by a higher rate of temporal than frontal atrophy in the AD group and a comparable rate of temporal vs. frontal atrophy rate in the healthy elderly (Figure 4).
Effects of age on regional atrophy rates
Correlation analyses between age and 1 year atrophy in each of the ROIs were performed. The results are presented in Table 4 and scatterplots for selected structures in Figure 5. We focus here on the 1 year changes, since the statistical power was lower for 2 years. Significant correlations between age and rates of atrophy were found for 32 ROIs. For the volumetric structures, hippocampus, cerebellum grey and white matter, cerebral white matter, thalamus and pallidum atrophy were related to age, as was expansion of the inferior lateral ventricles. Of the cortical structures, regions demonstrating significant relationships between age and rate of atrophy included entorhinal, transverse temporal, pars opercularis, isthmus cingulate, middle temporal, pars triangularis, banks of the superior temporal sulcus, parahippocampal, superior temporal, posterior cingulate, caudal middle frontal, precentral, fusiform, inferior parietal, temporal pole, precuneus, supramarginal, lateral orbitofrontal, superior parietal, post central, caudal anterior cingulate, medial orbitofrontal, pericalcarine, lateral occipital and inferior temporal cortices.
Table 4.
Pearson correlations between age and 1-year volume change in healthy elderly Negative correlations between age and rate of atrophy, and positive correlations between age and rate of ventricular expansion are generally found, showing that increasing age is associated with more atrophy and increased ventricular size.
| 1 year change | |
|---|---|
| Volumetric ROIs | |
| Accumbens | −0.13 |
| Amygdala | −0.16 |
| BrainStem | −0.15 |
| Caudate | −0.06 |
| Cerebellum GM | −0.20 |
| Cerebellum WM | −0.17 |
| Cerebral WM | −0.17 |
| Hippocampus | −0.33 |
| Pallidum | −0.17 |
| Putamen | −0.14 |
| Thalamus | −0.18 |
| Ventricular ROIs | |
| Lateral ventricles | 0.08 |
| Inf lateral ventricles | 0.21 |
| 3rd ventricle | 0.03 |
| 4th ventricle | 0.08 |
| Surface ROIs | |
| Cingulate, caudal ant | −0.19 |
| Cingulate, rostral ant | −0.11 |
| Cingulate, posterior | −0.22 |
| Cingulate, isthmus | −0.25 |
| Frontal, superior | −0.13 |
| Frontal, caudal middle | −0.21 |
| Frontal, rostral middle | −0.13 |
| Frontal, pars opercularis | −0.26 |
| Frontal, pars triangularis | −0.23 |
| Frontal, pars orbitalis | −0.08 |
| Frontal, lateral orbital | −0.19 |
| Frontal, medial orbital | −0.18 |
| Frontal, pole | −0.11 |
| Parietal, precentral gyrus | −0.21 |
| Parietal, post central gyrus | −0.19 |
| Parietal, paracentral gyrus | −0.13 |
| Parietal, superior | −0.19 |
| Parietal, inferior | −0.20 |
| Parietal, supramarginal | −0.19 |
| Parietal, precuneus | −0.20 |
| Temporal, parahippocampal | −0.23 |
| Temporal, entorhinal | −0.32 |
| Temporal, pole | −0.20 |
| Temporal, superior | −0.22 |
| Temporal, middle | −0.23 |
| Temporal, inferior | −0.17 |
| Temporal, transverse | −0.30 |
| Temporal, Banks Sup Temp Sulc | −0.23 |
| Temporal, fusiform | −0.20 |
| Occipital, lateral | −0.17 |
| Occipital, pericalcarine | −0.18 |
| Occipital, lingual | −0.16 |
| Occipital, cuneus | −0.13 |
Underlined italic indicates p < .05 (r ≥ +/− .17)
Bold indicates p < .01 (r ≥ +/− .21)
Bold underlined italic indicates p < .001 (r ≥ +/− .27)
Figure 5. Relationships between age and rate of atrophy in healthy elderly.

Scatterplots for selected ROIs showing the relationship between age and rates of annual volumetric reductions.
Discussion
Volumetric reductions of the cerebral cortex and subcortical brain structures, as well as expansion of the ventricles, were seen in healthy aging after only one year. While this has been shown previously for MCI and AD patients (McDonald et al., 2009; Schuff et al., 2009), the present report is the first to demonstrate detectable atrophy in a range of brain areas over only one year in healthy participants. Some of the changes seen, e.g. in the temporal lobe, may be due to preclinical AD, while others, e.g. in the prefrontal cortex, likely reflects the non-pathological aging process. This is discussed below.
The present atrophy rates for hippocampus (0.84%) and entorhinal cortex (0.55%) are comparable to previous studies (Barnes et al., 2009; Du et al., 2006; Du et al., 2003; Ezekiel et al., 2004; Jack et al., 1998; Raz et al., 2005a; Raz et al., 2005b; Scahill et al., 2003), supporting the feasibility of pooling data across multiple sites resulting in sensitive and realistic atrophy estimates. Generally, volumetric ROIs showed more decline than surface-based cortical ROIs, with the hippocampus showing the greatest one year reduction. Hippocampal volume has been found to correlate with age in cross-sectional (Good et al., 2001; Raz et al., 2004; Walhovd et al., 2005) and longitudinal studies (Driscoll et al., 2009; Du et al., 2006; Ezekiel et al., 2004; Jack et al., 1998; Raz et al., 2005b; Resnick et al., 2003; Scahill et al., 2003), but usually not more strongly than the other volumetric structures measured (Raz et al., 2005b). However, in a recent cross-sectional multi-sample study, estimated percentage decline per decade in the later half of the life-span was larger for hippocampus than any other volumetric structure (Walhovd et al., in press). In the present study, amygdala, thalamus, putamen and pallidum also showed longitudinal decline. Several cross-sectional studies have shown substantial reductions in the volumes of thalamus, amygdala and putamen (Greenberg et al., 2008; Gunning-Dixon et al., 1998; Raz et al., 2003; Sullivan et al., 2004; Van Der Werf et al., 2001; Walhovd et al., 2005; Walhovd et al., in press), as well as in pallidum after 50 years of age (Walhovd et al., 2005; Walhovd et al., in press), but discrepant findings regarding thalamus and pallidum have been reported (Gunning-Dixon et al., 1998; Jernigan et al., 1991; Jernigan et al., 2001; Luft et al., 1999; Raz et al., 2003).
Hippocampus and amygdala were the two volumetric structures that showed the largest annual decline in the present study, and these are known to show accelerated atrophy early in AD (Barnes et al., 2009; de Leon et al., 1989; Fennema-Notestine et al., 2009; Fischl et al., 2002; Jack et al., 1999; McDonald et al., in press). A recent meta-analysis estimated annual hippocampal reductions to be 4.66% in AD (Barnes et al., 2009). These were also the ROIs showing the largest volume decrease in the AD patients in the present study, with annual rates of decline of 3.75% for hippocampus and 3.79% for amygdala in the AD sample. Of the surface-based cortical ROIs showing substantial decline, entorhinal, parahippocampal and lateral temporal, as well as inferior parietal, supramarginal and fusiform areas, are all found to be vulnerable in early stages of AD (Du et al., 2007; Fennema-Notestine et al., 2009; McDonald et al., in press; McEvoy et al., 2009; Singh et al., 2006). An important question regards the extent to which atrophy in these areas in healthy elderly is an indication of preclinical AD. We cannot rule out with certainty that degenerative processes associated with early AD caused some of the observed atrophy. However, reductions were also seen in areas relatively less affected by early AD. For instance, large effects were found in inferior, middle and superior frontal cortex. Previous studies of the ADNI sample did not find differences in thickness between single domain amnestic MCI and controls in these areas (Fennema-Notestine et al., 2009), nor did these areas show longitudinal increases in atrophy in MCI participants showing mild functional impairment (CDR-SB of 1 or less) (McDonald et al., in press). Still, a recent study found prefrontal cortical volume to predict dementia (Burgmans et al., 2009a), and AD involves frontal atrophy (Dickerson et al., 2008; Du et al., 2007; Fjell et al., 2008; Singh et al., 2006). An early longitudinal study found that while temporal atrophy was seen in presymptomatic and mildly affected patients, frontal involvement occurred later in the disease (Scahill et al., 2002). Another study found demented patients to show annual atrophy rates exceeding 0.4% in large areas of the brain, including the frontal cortex (Buckner et al., 2005). In the present study, more atrophy was observed in AD than in healthy controls in almost all ROIs, including prefrontal cortex. However, temporal lobe atrophy was the most prominent in AD, while healthy aging was characterized by comparable rates of atrophy in the temporal lobe and the frontal cortex. The temporal changes seen in the healthy elderly group may be related to instances of preclinical AD, while the frontal changes are more prominent than what would be expected due to AD. Significant decline was also observed in thalamus, accumbens and cerebral and cerebellar white matter; areas that are not among the most vulnerable in early phases of AD (Fennema-Notestine et al., 2009). In the AD patients included in the present study, cerebellum WM was the 39th most vulnerable region, while thalamus was the 22nd and accumbens the 14th. Thus, it is unlikely that all the brain changes found here in the healthy elderly group can be attributed to preclinical AD. We believe that many of the changes observed in healthy aging are caused by processes unrelated to degenerative disease. Consistent with this, a recent study reported that the transition from healthy elderly to MCI was associated with a unique pattern of longitudinal change, including increased ventricular size, and reductions in temporal grey matter, orbitofrontal areas and temporal association cortices, including the hippocampus (Driscoll et al., 2009). The pattern of atrophy over several years in healthy elderly not developing MCI was widespread but did not resemble the pattern associated with MCI (Driscoll et al., 2009). As further follow-up data become available, the hypothesis that the temporal changes seen in the elderly are related to preclinical AD, while the frontal changes are less associated with incipient disease, can be tested. Since it is possible that structural brain changes due to degenerative disease may be manifested long before cognitive symptoms are detectable, follow-up cognitive and clinical data for several years will be invaluable (Burgmans et al., 2009b).
Accelerated atrophy with higher age
Older age was related to increased atrophy and more ventricular expansion. This is consistent with cross-sectional reports of non-linear volumetric changes (Allen et al., 2005; Fjell et al., In press-b; Jernigan and Gamst, 2005; Walhovd et al., 2005; Walhovd et al., in press). However, cross-sectional studies of cortical thickness have mostly reported linear age-changes over the adult life-span (Fjell et al., in press-a; Salat et al., 2004). It is possible that longitudinal designs are more sensitive to accelerated decline in the oldest old, especially if the effects are of moderate strength and evident only in the latest part of the life-span. The correlations between age and rate of change seldom exceeded +/−.30. Interestingly, the strongest correlations between age and rate of atrophy were found in entorhinal cortex and the hippocampus, and significant correlations were also found in isthmus of the cingulate, middle temporal and parahippocampal gyri, all of which are affected in early stages of AD. Since the occurrence of preclinical AD is much higher among 70- and 80 year olds than among 50 and 60 year olds, it is possible that the pattern of accelerated longitudinal atrophy in aging is related to preclinical AD in a subgroup of the participants, even though cognitive symptoms are not yet manifested. This interpretation is in agreement with another recent study where elderly cognitively normal participants showed a pattern of atrophy more similar to that seen in AD than did younger participants, i.e., more temporal lobe atrophy (Davatzikos et al., 2009).
Conclusion
The present results show that widespread changes in brain structure can be found in healthy elderly over only one year, and that atrophy accelerates with advanced age. Some of the atrophy occurred in areas vulnerable to AD and some in areas less characteristic of the disease in early stages. This suggests that the changes are not primarily driven by degenerative processes associated with AD, although it is likely that preclinical changes associated with AD are superposed on changes due to normal aging in some subjects. Indeed, the pattern of accelerated atrophy with increasing age is probably influenced by occurrence of preclinical AD in a sub-group of the most elderly participants, and this may especially be reflected in the temporal lobe. Nevertheless, this study shows that decreases in brain volume in healthy aging can be observed in almost all brain areas over 1 to 2 years.
Supplementary Material
Acknowledgement
This work was supported by a grant (#U24 RR021382) to the Morphometry Biomedical Informatics Research Network (BIRN, http://www.nbirn.net), that is funded by the National Center for Research Resources at the National Institutes of Health (NIH), U.S.A, and by the National Institute on Aging at the NIH (U01 AG024904; R01 AG031224 and K01AG029218). The work of Walhovd and Fjell was supported by the Norwegian Research Council. Data collection and sharing for this project was funded by the Alzheimer's Disease Neuroimaging Initiative (ADNI; Principal Investigator: Michael Weiner; NIH grant U01 AG024904). ADNI is funded by the National Institute on Aging, the National Institute of Biomedical Imaging and Bioengineering (NIBIB), and through generous contributions from the following: Pfizer Inc., Wyeth Research, Bristol-Myers Squibb, Eli Lilly and Company, GlaxoSmithKline, Merck & Co. Inc., AstraZeneca AB, Novartis Pharmaceuticals Corporation, Alzheimer's Association, Eisai Global Clinical Development, Elan Corporation plc, Forest Laboratories, and the Institute for the Study of Aging, with participation from the U.S. Food and Drug Administration. Industry partnerships are coordinated through the Foundation for the National Institutes of Health. The grantee organization is the Northern California Institute for Research and Education, and the study is coordinated by the Alzheimer's Disease Cooperative Study at the University of California, San Diego. ADNI data are disseminated by the Laboratory of Neuro Imaging at the University of California, Los Angeles. We thank Robin Jennings, Michele Perry, Chris Pung and Elaine Wu for downloading and preprocessing the ADNI MRI data. Data used in the preparation of this article were obtained from the Alzheimer's Disease Neuroimaging Initiative (ADNI) database (www.loni.ucla.edu/ADNI). As such, the investigators within the ADNI contributed to the design and implementation of ADNI and/or provided data but did not participate in analysis or writing of this report. A complete listing of ADNI investigators is available at http://www.loni.ucla.edu/ADNI/Data/ADNI_Authorship_List.pdf
References
- Allen JS, Bruss J, Brown CK, Damasio H. Normal neuroanatomical variation due to age: the major lobes and a parcellation of the temporal region. Neurobiol Aging. 2005;26:1245–1260. doi: 10.1016/j.neurobiolaging.2005.05.023. discussion 1279–1282. [DOI] [PubMed] [Google Scholar]
- Ashburner J, Andersson JL, Friston KJ. High-dimensional image registration using symmetric priors. Neuroimage. 1999;9:619–628. doi: 10.1006/nimg.1999.0437. [DOI] [PubMed] [Google Scholar]
- Barnes J, Bartlett JW, van de Pol LA, Loy CT, Scahill RI, Frost C, Thompson P, Fox NC. A meta-analysis of hippocampal atrophy rates in Alzheimer's disease. Neurobiology of Aging. 2009;30:1711–1723. doi: 10.1016/j.neurobiolaging.2008.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckner RL, Snyder AZ, Shannon BJ, LaRossa G, Sachs R, Fotenos AF, Sheline YI, Klunk WE, Mathis CA, Morris JC, Mintun MA. Molecular, structural, and functional characterization of Alzheimer's disease: evidence for a relationship between default activity, amyloid, and memory. J Neurosci. 2005;25:7709–7717. doi: 10.1523/JNEUROSCI.2177-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgmans S, van Boxtel MP, Smeets F, Vuurman EF, Gronenschild EH, Verhey FR, Uylings HB, Jolles J. Prefrontal cortex atrophy predicts dementia over a six-year period. Neurobiol Aging. 2009a;30:1413–1419. doi: 10.1016/j.neurobiolaging.2007.11.028. [DOI] [PubMed] [Google Scholar]
- Burgmans S, van Boxtel MP, Vuurman EF, Smeets F, Gronenschild EH, Uylings HB, Jolles J. The prevalence of cortical gray matter atrophy may be overestimated in the healthy aging brain. Neuropsychology. 2009b;23:541–550. doi: 10.1037/a0016161. [DOI] [PubMed] [Google Scholar]
- Christensen GE, Rabbitt RD, Miller MI. Deformable templates using large deformation kinematics. IEEE Trans Image Process. 1996;5:1435–1447. doi: 10.1109/83.536892. [DOI] [PubMed] [Google Scholar]
- Dale AM, Fischl B, Sereno MI. Cortical surface-based analysis. I. Segmentation and surface reconstruction. Neuroimage. 1999;9:179–194. doi: 10.1006/nimg.1998.0395. [DOI] [PubMed] [Google Scholar]
- Dale AM, Sereno MI. Improved localization of cortical activity by combining EEG and MEG with MRI cortical surface reconstruction: a linear approach. Journal of Cognitive Neuroscience. 1993;5:162–176. doi: 10.1162/jocn.1993.5.2.162. [DOI] [PubMed] [Google Scholar]
- Davatzikos C, Xu F, An Y, Fan Y, Resnick SM. Longitudinal progression of Alzheimer's-like patterns of atrophy in normal older adults: the SPARE-AD index. Brain. 2009 doi: 10.1093/brain/awp091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Leon MJ, George AE, Stylopoulos LA, Smith G, Miller DC. Early marker for Alzheimer's disease: the atrophic hippocampus. Lancet. 1989;2:672–673. doi: 10.1016/s0140-6736(89)90911-2. [DOI] [PubMed] [Google Scholar]
- Desikan RS, Segonne F, Fischl B, Quinn BT, Dickerson BC, Blacker D, Buckner RL, Dale AM, Maguire RP, Hyman BT, Albert MS, Killiany RJ. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage. 2006;31:968–980. doi: 10.1016/j.neuroimage.2006.01.021. [DOI] [PubMed] [Google Scholar]
- Dickerson BC, Bakkour A, Salat DH, Feczko E, Pacheco J, Greve DN, Grodstein F, Wright CI, Blacker D, Rosas HD, Sperling RA, Atri A, Growdon JH, Hyman BT, Morris JC, Fischl B, Buckner RL. The Cortical Signature of Alzheimer's Disease: Regionally Specific Cortical Thinning Relates to Symptom Severity in Very Mild to Mild AD Dementia and is Detectable in Asymptomatic Amyloid-Positive Individuals. Cereb Cortex. 2008;16:497–510. doi: 10.1093/cercor/bhn113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Driscoll I, Davatzikos C, An Y, Wu X, Shen D, Kraut M, Resnick SM. Longitudinal pattern of regional brain volume change differentiates normal aging from MCI. Neurology. 2009;72:1906–1913. doi: 10.1212/WNL.0b013e3181a82634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du AT, Schuff N, Kramer JH, Rosen HJ, Gorno-Tempini ML, Rankin K, Miller BL, Weiner MW. Different regional patterns of cortical thinning in Alzheimer's disease and frontotemporal dementia. Brain. 2007;130:1159–1166. doi: 10.1093/brain/awm016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du AT, Schuff N, Zhu XP, Jagust WJ, Miller BL, Reed BR, Kramer JH, Mungas D, Yaffe K, Chui HC, Weiner MW. Atrophy rates of entorhinal cortex in AD and normal aging. Neurology. 2003;60:481–486. doi: 10.1212/01.wnl.0000044400.11317.ec. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du AT, Schuff N, Chao LL, Kornak J, Jagust WJ, Kramer JH, Reed BR, Miller BL, Norman D, Chui HC, Weiner MW. Age effects on atrophy rates of entorhinal cortex and hippocampus. Neurobiol Aging. 2006;27:733–740. doi: 10.1016/j.neurobiolaging.2005.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enzinger C, Fazekas F, Matthews PM, Ropele S, Schmidt H, Smith S, Schmidt R. Risk factors for progression of brain atrophy in aging: six-year follow-up of normal subjects. Neurology. 2005;64:1704–1711. doi: 10.1212/01.WNL.0000161871.83614.BB. [DOI] [PubMed] [Google Scholar]
- Ezekiel F, Chao L, Kornak J, Du AT, Cardenas V, Truran D, Jagust W, Chui H, Miller B, Yaffe K, Schuff N, Weiner M. Comparisons between global and focal brain atrophy rates in normal aging and Alzheimer disease: Boundary Shift Integral versus tracing of the entorhinal cortex and hippocampus. Alzheimer Dis Assoc Disord. 2004;18:196–201. [PMC free article] [PubMed] [Google Scholar]
- Fennema-Notestine C, Hagler DJ, Jr., McEvoy LK, Fleisher AS, Wu EH, Karow DS, Dale AM. Structural MRI biomarkers for preclinical and mild Alzheimer's disease. Hum Brain Mapp. 2009 doi: 10.1002/hbm.20744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischl B, Dale AM. Measuring the thickness of the human cerebral cortex from magnetic resonance images. Proc Natl Acad Sci U S A. 2000;97:11050–11055. doi: 10.1073/pnas.200033797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischl B, Sereno MI, Dale AM. Cortical surface-based analysis. II: Inflation, flattening, and a surface-based coordinate system. Neuroimage. 1999a;9:195–207. doi: 10.1006/nimg.1998.0396. [DOI] [PubMed] [Google Scholar]
- Fischl B, Sereno MI, Tootell RB, Dale AM. High-resolution intersubject averaging and a coordinate system for the cortical surface. Hum Brain Mapp. 1999b;8:272–284. doi: 10.1002/(SICI)1097-0193(1999)8:4<272::AID-HBM10>3.0.CO;2-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischl B, Salat DH, Busa E, Albert M, Dieterich M, Haselgrove C, van der Kouwe A, Killiany R, Kennedy D, Klaveness S, Montillo A, Makris N, Rosen B, Dale AM. Whole brain segmentation: automated labeling of neuroanatomical structures in the human brain. Neuron. 2002;33:341–355. doi: 10.1016/s0896-6273(02)00569-x. [DOI] [PubMed] [Google Scholar]
- Fischl B, van der Kouwe A, Destrieux C, Halgren E, Segonne F, Salat DH, Busa E, Seidman LJ, Goldstein J, Kennedy D, Caviness V, Makris N, Rosen B, Dale AM. Automatically parcellating the human cerebral cortex. Cereb Cortex. 2004;14:11–22. doi: 10.1093/cercor/bhg087. [DOI] [PubMed] [Google Scholar]
- Fjell AM, Westlye LT, Amlien I, Espeseth T, Reinvang I, Raz N, Agartz I, Salat D, Greve D, Fischl B, Dale A, Walhovd KB. Minute effects of sex on the aging brain: a multi-sample MRI-study of healthy aging and Alzheimer's disease. Journal of Neuroscience. 2009a;29:8774–8783. doi: 10.1523/JNEUROSCI.0115-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fjell AM, Westlye LT, Amlien I, Espeseth T, Reinvang I, Raz N, Agartz I, Salat D, Greve D, Fischl B, Dale A, Walhovd KB. High consistency of regional cortical thinning in aging across multiple samples. Cerebral Cortex. 2009b;19:2001–2012. doi: 10.1093/cercor/bhn232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fjell AM, Walhovd KB, Amlien I, Bjørnerud A, Reinvang I, Gjerstad L, Cappelen T, Willoch F, Due-Tønnessen P, Grambaite R, Skinningsrud A, Stenset V, Fladby T. Morphometric Changes in the Episodic Memory Network and Tau Pathologic Features Correlate With Memory Performance in Patients With Mild Cognitive Impairment. American Journal of Neuroradiology. 2008;29:1–7. doi: 10.3174/ajnr.A1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Folstein MF, Folstein SE, McHugh PR. “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. J Psychiatr Res. 1975;12:189–198. doi: 10.1016/0022-3956(75)90026-6. [DOI] [PubMed] [Google Scholar]
- Fotenos AF, Snyder AZ, Girton LE, Morris JC, Buckner RL. Normative estimates of cross-sectional and longitudinal brain volume decline in aging and AD. Neurology. 2005;64:1032–1039. doi: 10.1212/01.WNL.0000154530.72969.11. [DOI] [PubMed] [Google Scholar]
- Freeborough PA, Fox NC. Modeling brain deformations in Alzheimer disease by fluid registration of serial 3D MR images. J Comput Assist Tomogr. 1998;22:838–843. doi: 10.1097/00004728-199809000-00031. [DOI] [PubMed] [Google Scholar]
- Good CD, Johnsrude IS, Ashburner J, Henson RN, Friston KJ, Frackowiak RS. A voxel-based morphometric study of ageing in 465 normal adult human brains. Neuroimage. 2001;14:21–36. doi: 10.1006/nimg.2001.0786. [DOI] [PubMed] [Google Scholar]
- Grandy J. Efficient Computation of Volume of Hexahedral Cells. Lawrence Livermore National Laboratory; 1997. [Google Scholar]
- Greenberg DL, Messer DF, Payne ME, Macfall JR, Provenzale JM, Steffens DC, Krishnan RR. Aging, gender, and the elderly adult brain: an examination of analytical strategies. Neurobiol Aging. 2008;29:290–302. doi: 10.1016/j.neurobiolaging.2006.09.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gunning-Dixon FM, Head D, McQuain J, Acker JD, Raz N. Differential aging of the human striatum: a prospective MR imaging study. AJNR Am J Neuroradiol. 1998;19:1501–1507. [PMC free article] [PubMed] [Google Scholar]
- Holland D, Hagler D, Fennema-Notestine C, et al. Longitudinal nonlinear registration to quantitative analysis in whole brain and regions of interest. Alzheimer's & Dementia: The Journal of Alzheimer's Associations. 2008;4 [Google Scholar]
- Jack CR, Jr., Petersen RC, Xu Y, O'Brien PC, Smith GE, Ivnik RJ, Tangalos EG, Kokmen E. Rate of medial temporal lobe atrophy in typical aging and Alzheimer's disease. Neurology. 1998;51:993–999. doi: 10.1212/wnl.51.4.993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jack CR, Jr., Petersen RC, Xu YC, O'Brien PC, Smith GE, Ivnik RJ, Boeve BF, Waring SC, Tangalos EG, Kokmen E. Prediction of AD with MRI-based hippocampal volume in mild cognitive impairment. Neurology. 1999;52:1397–1403. doi: 10.1212/wnl.52.7.1397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jernigan TL, Gamst AC. Changes in volume with age--consistency and interpretation of observed effects. Neurobiol Aging. 2005;26:1271–1274. doi: 10.1016/j.neurobiolaging.2005.05.016. discussion 1275–1278. [DOI] [PubMed] [Google Scholar]
- Jernigan TL, Archibald SL, Berhow MT, Sowell ER, Foster DS, Hesselink JR. Cerebral structure on MRI, Part I: Localization of age-related changes. Biol Psychiatry. 1991;29:55–67. doi: 10.1016/0006-3223(91)90210-d. [DOI] [PubMed] [Google Scholar]
- Jernigan TL, Archibald SL, Fennema-Notestine C, Gamst AC, Stout JC, Bonner J, Hesselink JR. Effects of age on tissues and regions of the cerebrum and cerebellum. Neurobiol Aging. 2001;22:581–594. doi: 10.1016/s0197-4580(01)00217-2. [DOI] [PubMed] [Google Scholar]
- Jovicich J, Czanner S, Greve D, Haley E, van der Kouwe A, Gollub R, Kennedy D, Schmitt F, Brown G, Macfall J, Fischl B, Dale A. Reliability in multi-site structural MRI studies: effects of gradient non-linearity correction on phantom and human data. Neuroimage. 2006;30:436–443. doi: 10.1016/j.neuroimage.2005.09.046. [DOI] [PubMed] [Google Scholar]
- Kuperberg GR, Broome MR, McGuire PK, David AS, Eddy M, Ozawa F, Goff D, West WC, Williams SC, van der Kouwe AJ, Salat DH, Dale AM, Fischl B. Regionally localized thinning of the cerebral cortex in schizophrenia. Arch Gen Psychiatry. 2003;60:878–888. doi: 10.1001/archpsyc.60.9.878. [DOI] [PubMed] [Google Scholar]
- Luft AR, Skalej M, Schulz JB, Welte D, Kolb R, Burk K, Klockgether T, Voight K. Patterns of age-related shrinkage in cerebellum and brainstem observed in vivo using three-dimensional MRI volumetry. Cereb Cortex. 1999;9:712–721. doi: 10.1093/cercor/9.7.712. [DOI] [PubMed] [Google Scholar]
- Makris N, Meyer JW, Bates JF, Yeterian EH, Kennedy DN, Caviness VS. MRI-Based topographic parcellation of human cerebral white matter and nuclei II. Rationale and applications with systematics of cerebral connectivity. Neuroimage. 1999;9:18–45. doi: 10.1006/nimg.1998.0384. [DOI] [PubMed] [Google Scholar]
- McDonald C, McEvoy LK, Gharapetian L, Fennema-Notestine C, Hagler DJ, Jr., Holland D, Koyama A, Dale AM. Regional rates of neocortical atrophy from normal aging to early Alzheimer's Disease. Neurology. 2009;73:457–465. doi: 10.1212/WNL.0b013e3181b16431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McEvoy LK, Fennema-Notestine C, Roddey JC, Hagler DJ, Jr., Holland D, Karow DS, Pung CJ, Brewer JB, Dale AM. Alzheimer disease: quantitative structural neuroimaging for detection and prediction of clinical and structural changes in mild cognitive impairment. Radiology. 2009;251:195–205. doi: 10.1148/radiol.2511080924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKhann G, Drachman D, Folstein M, Katzman R, Price D, Stadlan EM. Clinical diagnosis of Alzheimer's disease: report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer's Disease. Neurology. 1984;34:939–944. doi: 10.1212/wnl.34.7.939. [DOI] [PubMed] [Google Scholar]
- Miller MI, Christensen GE, Amit Y, Grenander U. Mathematical textbook of deformable neuroanatomies. Proc Natl Acad Sci U S A. 1993;90:11944–11948. doi: 10.1073/pnas.90.24.11944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris JC. The Clinical Dementia Rating (CDR): current version and scoring rules. Neurology. 1993;43:2412–2414. doi: 10.1212/wnl.43.11.2412-a. [DOI] [PubMed] [Google Scholar]
- Pfefferbaum A, Sullivan EV, Rosenbloom MJ, Mathalon DH, Lim KO. A controlled study of cortical gray matter and ventricular changes in alcoholic men over a 5-year interval. Arch Gen Psychiatry. 1998;55:905–912. doi: 10.1001/archpsyc.55.10.905. [DOI] [PubMed] [Google Scholar]
- Raz N, Rodrigue KM. Differential aging of the brain: Patterns, cognitive correlates and modifiers. Neuroscience & Biobehavioral Reviews. 2006;30:730. doi: 10.1016/j.neubiorev.2006.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raz N, Rodrigue KM, Kennedy KM, Dahle C, Head D, Acker JD. Differential age-related changes in the regional metencephalic volumes in humans: a 5-year follow-up. Neurosci Lett. 2003;349:163–166. doi: 10.1016/s0304-3940(03)00820-6. [DOI] [PubMed] [Google Scholar]
- Raz N, Gunning-Dixon F, Head D, Rodrigue KM, Williamson A, Acker JD. Aging, sexual dimorphism, and hemispheric asymmetry of the cerebral cortex: replicability of regional differences in volume. Neurobiol Aging. 2004;25:377–396. doi: 10.1016/S0197-4580(03)00118-0. [DOI] [PubMed] [Google Scholar]
- Raz N, Lindenberger U, Rodrigue KM, Kennedy KM, Head D, Williamson A, Dahle C, Gerstorf D, Acker JD. Regional brain changes in aging healthy adults: general trends, individual differences and modifiers. Cereb Cortex. 2005a;15:1676–1689. doi: 10.1093/cercor/bhi044. [DOI] [PubMed] [Google Scholar]
- Raz N, Lindenberger U, Rodrigue KM, Kennedy KM, Head D, Williamson A, Dahle C, Gerstorf D, Acker JD. Regional Brain Changes in Aging Healthy Adults: General Trends, Individual Differences and Modifiers. Cereb Cortex %R 101093/cercor/bhi044. 2005b;15:1676–1689. doi: 10.1093/cercor/bhi044. [DOI] [PubMed] [Google Scholar]
- Resnick SM, Pham DL, Kraut MA, Zonderman AB, Davatzikos C. Longitudinal magnetic resonance imaging studies of older adults: a shrinking brain. J Neurosci. 2003;23:3295–3301. doi: 10.1523/JNEUROSCI.23-08-03295.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Resnick SM, Goldszal AF, Davatzikos C, Golski S, Kraut MA, Metter EJ, Bryan RN, Zonderman AB. One-year age changes in MRI brain volumes in older adults. Cereb Cortex. 2000;10:464–472. doi: 10.1093/cercor/10.5.464. [DOI] [PubMed] [Google Scholar]
- Rosas HD, Liu AK, Hersch S, Glessner M, Ferrante RJ, Salat DH, van der Kouwe A, Jenkins BG, Dale AM, Fischl B. Regional and progressive thinning of the cortical ribbon in Huntington's disease. Neurology. 2002;58:695–701. doi: 10.1212/wnl.58.5.695. [DOI] [PubMed] [Google Scholar]
- Salat DH, Lee SY, van der Kouwe AJ, Greve DN, Fischl B, Rosas HD. Age-associated alterations in cortical gray and white matter signal intensity and gray to white matter contrast. NeuroImage. 48:21–28. doi: 10.1016/j.neuroimage.2009.06.074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salat DH, Buckner RL, Snyder AZ, Greve DN, Desikan RS, Busa E, Morris JC, Dale AM, Fischl B. Thinning of the cerebral cortex in aging. Cereb Cortex. 2004;14:721–730. doi: 10.1093/cercor/bhh032. [DOI] [PubMed] [Google Scholar]
- Scahill RI, Schott JM, Stevens JM, Rossor MN, Fox NC. Mapping the evolution of regional atrophy in Alzheimer's disease: unbiased analysis of fluid-registered serial MRI. Proc Natl Acad Sci U S A. 2002;99:4703–4707. doi: 10.1073/pnas.052587399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scahill RI, Frost C, Jenkins R, Whitwell JL, Rossor MN, Fox NC. A longitudinal study of brain volume changes in normal aging using serial registered magnetic resonance imaging. Arch Neurol. 2003;60:989–994. doi: 10.1001/archneur.60.7.989. [DOI] [PubMed] [Google Scholar]
- Schuff N, Woerner N, Boreta L, Kornfield T, Shaw LM, Trojanowski JQ, Thompson PM, Jack CR, Jr., Weiner MW. MRI of hippocampal volume loss in early Alzheimer's disease in relation to ApoE genotype and biomarkers. Brain. 2009;132:1067–1077. doi: 10.1093/brain/awp007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh V, Chertkow H, Lerch JP, Evans AC, Dorr AE, Kabani NJ. Spatial patterns of cortical thinning in mild cognitive impairment and Alzheimer's disease. Brain. 2006;129:2885–2893. doi: 10.1093/brain/awl256. [DOI] [PubMed] [Google Scholar]
- Sled JG, Zijdenbos AP, Evans AC. A nonparametric method for automatic correction of intensity nonuniformity in MRI data. IEEE Trans Med Imaging. 1998;17:87–97. doi: 10.1109/42.668698. [DOI] [PubMed] [Google Scholar]
- Sullivan EV, Rosenbloom M, Serventi KL, Pfefferbaum A. Effects of age and sex on volumes of the thalamus, pons, and cortex. Neurobiol Aging. 2004;25:185–192. doi: 10.1016/s0197-4580(03)00044-7. [DOI] [PubMed] [Google Scholar]
- Van Der Werf YD, Tisserand DJ, Visser PJ, Hofman PA, Vuurman E, Uylings HB, Jolles J. Thalamic volume predicts performance on tests of cognitive speed and decreases in healthy aging. A magnetic resonance imaging-based volumetric analysis. Brain Res Cogn Brain Res. 2001;11:377–385. doi: 10.1016/s0926-6410(01)00010-6. [DOI] [PubMed] [Google Scholar]
- Walhovd KB, Fjell AM, Reinvang I, Lundervold A, Dale AM, Eilertsen DE, Quinn BT, Salat D, Makris N, Fischl B. Effects of age on volumes of cortex, white matter and subcortical structures. Neurobiol Aging. 2005;26:1261–1270. doi: 10.1016/j.neurobiolaging.2005.05.020. discussion 1275–1268. [DOI] [PubMed] [Google Scholar]
- Walhovd KB, Westlye LT, Amlien I, Espeseth T, Reinvang I, Raz N, Agartz I, Salat D, Greve D, Fischl B, Dale A. Consistent neuroanatomical age-related volume differences across multiple samples. Neurobiology of Aging. doi: 10.1016/j.neurobiolaging.2009.05.013. (in press) doi:10.1016/j.neurobiolaging.2009.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westlye LT, Walhovd KB, Dale A, Espeseth T, Reinvang I, Raz N, Agartz I, Greve D, Fischl B, Fjell AM. Increased sensitivity to effects of normal aging and Alzheimer's disease on cortical thickness by adjustment for local variability in gray/ white contrast: A multi-sample MRI study. NeuroImage. 2009;47:1545–1557. doi: 10.1016/j.neuroimage.2009.05.084. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.