

Abstract
Money is a secondary reinforcer that acquires its value through social communication and interaction. In everyday human behavior and laboratory studies, money has been shown to influence appetitive or reward learning. It is unclear, however, if money has a similar impact on aversive learning. The goal of this study was to investigate the efficacy of money in aversive learning, comparing it with primary reinforcers that are traditionally used in fear conditioning paradigms. A series of experiments were conducted in which participants initially played a gambling game that led to a monetary gain. They were then presented with an aversive conditioning paradigm, with either shock (primary reinforcer) or loss of money (secondary reinforcer) as the unconditioned stimulus. Skin conductance responses and subjective ratings indicated that potential monetary loss modulated the conditioned response. Depending on the presentation context, the secondary reinforcer was as effective as the primary reinforcer during aversive conditioning. These results suggest that stimuli that acquire reinforcing properties through social communication and interaction, such as money, can effectively influence aversive learning.
Keywords: reward, punishment, striatum, amygdala, incentive, valence, primary reinforcer, fear conditioning, Pavlovian conditioning, neuroeconomics
INTRODUCTION
Research focused on understanding the mechanisms of affective learning have led to the development of elegant animal models that can be translated to humans. Yet, such models are limited in reflecting everyday human function in a social and cultural context. For instance, animal studies have traditionally utilized inherently appetitive or aversive stimuli to modulate learning. In human culture, however, the more common reinforcers that influence behavior are socially defined. One example is money, a reinforcer that acquires value through social communication and interaction. Although some research exists to show the modulatory influence of money as a reward on the behavioral and neural correlates of appetitive learning, less is known about the effectiveness of money during aversive conditioning, where primary reinforcers such as shock are traditionally used. The goal of this study is to first investigate the efficacy of money, a secondary reinforcer, in aversive conditioning; and second, to compare the influence of primary and secondary reinforcers on aversive conditioning.
Early theories of appetitive conditioning identified two different categories of reinforcers, primary and secondary reinforcers, that are capable of influencing motivational behavior (Skinner, 1938; Hull, 1943). For instance, hunger was defined as a primary drive, an innate or biologically preprogrammed state which elicits food seeking behavior and food consumption, in consequence, labeling food as a primary reinforcer. Studies of appetitive conditioning in animals have demonstrated the effect of primary reinforcers such as food, liquid or odors on behavior (Pavlov and Anrep, 1927; Skinner, 1953; Rescorla and Wagner, 1972) and its associated neural mechanisms (Parkinson et al., 2000; Everitt et al. 2003; Cardinal and Everitt, 2004). Similarly in humans, primary reinforcers can influence behavior and act through common neural structures to induce appetitive conditioning (Breiter et al., 1997; Gottfried et al., 2002; O’Doherty et al., 2002; McClure et al., 2003).
In contrast, a secondary drive was defined as a learned or acquired state. Money, as previously mentioned, is an example of a secondary reinforcer, which acquires its reinforcing properties through its association with primary reinforcers (i.e. money can be used to acquire food). Due to societal and cultural factors, money has evolved to become a powerful incentive in driving human behavior, perhaps equally as important as primary reinforcers. In the laboratory, this has been primarily observed during appetitive learning, where secondary reinforcers such as money lead to successful behavioral learning of contingencies, further eliciting activation in similar neural structures as primary reinforcers (Breiter and Rosen, 1999; Delgado et al., 2000; Elliott et al., 2000; Knutson et al., 2001a, b; Delgado et al., 2003, 2005).
Less is known about the role of money as a secondary reinforcer in aversive classical conditioning. Typically in a fear conditioning session, for example, an aversive primary reinforcer, such as mild shock or puff of air in the eye is used as the unconditioned stimulus (US). Studies using mild shocks as reinforcers have found robust conditioned responses expressed through measures such as freezing, for instance, in rats (Davis, 1992; LeDoux, 2000; Pare, et al., 2004) and skin conductance responses in humans (Adolphs et al., 1995; LaBar et al., 1995; LaBar et al., 1998; Phelps et al., 2004). Nevertheless, it is unclear how effective secondary reinforcers such as money can be during aversive classical conditioning, and how such reinforcers compare with aversive primary reinforcers.
The current study aims to investigate the efficacy of money as an unconditioned stimulus in an aversive conditioning paradigm, comparing its efficacy with a primary reinforcer. Specifically, two questions are asked: (1) can loss of money serve as an aversive unconditioned stimulus during a classical conditioning paradigm; and (2) how does it compare to a primary reinforcer, namely shock, at inducing conditioned fear? Three experiments were designed to investigate these issues (Figure 1). The first experiment probes the effectiveness of loss of money as an unconditioned stimulus in a simple aversive or fear conditioning paradigm. In the second experiment, primary and secondary reinforcers within the same conditioning session are contrasted. Finally, the third experiment compares the efficacy of loss of money and shock as unconditioned stimuli when the presentation context is manipulated and separate fear conditioning sessions are used.
Fig. 1.
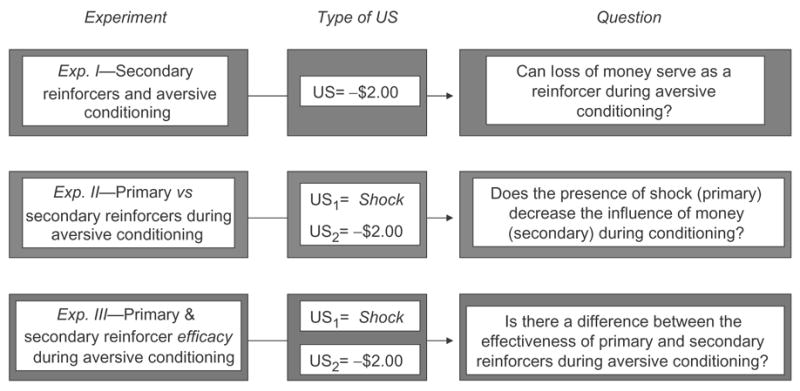
Experimental breakdown—question and goals of each of the experiments, along with type of unconditioned stimulus (US) used in aversive conditioning sessions. Experiments I and II involved one conditioning session each, while Experiment III contained two separate aversive conditioning sessions.
EXPERIMENT I
Methods
Participants
Twenty-two volunteers participated in this study (7 males, and 15 females). Two participants were removed due to technical issues with the physiological recording equipment and failure to comply with instructions. Final analysis was therefore conducted on 20 participants (6 males, and 14 females). Participants responded to posted advertisement (average age: M = 19.85, s.d. = 1.79), and all participants gave informed consent.
Procedure
Participants were instructed that the experiment consisted of two parts (Figure 2). The first part was a gambling session (adapted from Delgado et al., 2004). In this session, participants were told they were playing a computerized ‘card-guessing’ game, where the objective was to determine if the value of a given card was higher or lower than the number 5 (Figure 2A). During each trial, a question mark was presented in the center of the ‘card’ (a white rectangle projected in a black screen), indicating that participants had 3.5 s to make a response. Participants could chose either higher (possible outcomes: 6, 7, 8 and 9) or lower (possible outcomes: 1, 2, 3 and 4) than 5 by using their right hand’s index and middle finger, respectively, and pressing the corresponding button in the keyboard. The outcome was then displayed for 500 ms, followed by a feedback arrow (which indicated positive or negative feedback) for another 500 ms and an inter-trial interval of 4.5 s before the onset of the next trial.
Fig. 2.
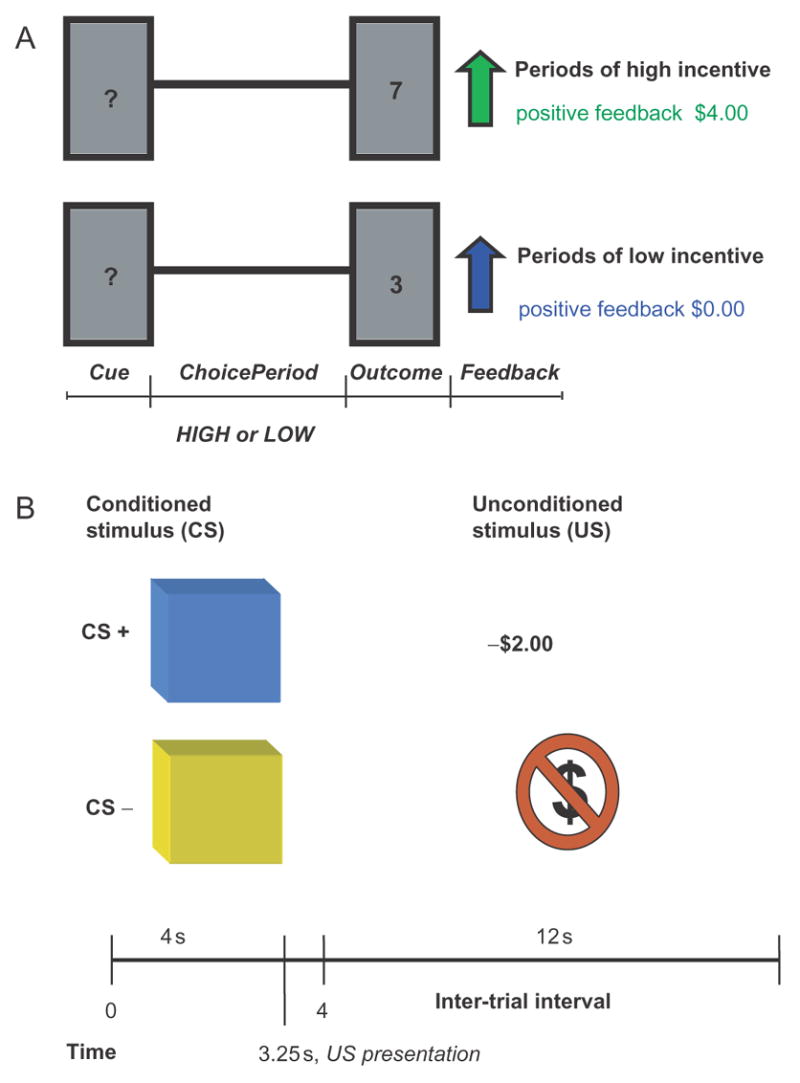
(A) The gambling task—adapted from Delgado et al. (2004). Participants were presented with a card and were asked if its value was higher or lower than the number 5 and were given feedback on their choice. The question mark was the cue for participants to make their choice (either high or low). During a subsequent outcome phase, participants were presented with the actual value of the card and either positive or negative feedback depending on their choice. (B) Aversive conditioning paradigm with money as an unconditioned stimulus—adapted from Phelps et al. (2004). Participants viewed two squares of different colors (blue, yellow) which served as the conditioned stimuli (CS). One of the squares (CS+) was paired with occasional presentations of a visual display indicating a monetary loss of $2.00, the unconditioned stimulus.
Participants were told they would be playing trials of the game during alternating blocks called ‘money’ and ‘no money’ blocks. During ‘money’ blocks, or high incentive trials, a green feedback arrow pointing up indicated a correct response and a monetary reward of $4.00, while a red feedback arrow pointing down indicated an incorrect response and a monetary loss of −$2.00. In contrast, during ‘no money’ blocks or low incentive trials, no monetary incentive was available, only feedback as a blue feedback arrow pointing up indicated a correct response while a blue feedback arrow pointing down indicated an incorrect response. Participants played three blocks of high incentive trials (12 per block) intermixed with three blocks of low incentive trials (12 per block) for a total of six counterbalanced blocks of trials. Unbeknownst to participants, the outcomes were predetermined ensuring a 50% reinforcement rate and a total monetary gain of $36.00 at the end of the first part. This profit was an endowment which could be drawn upon in the fear conditioning session.
Each participant was informed of their total upon completion of the gambling session. The second part of the experiment consisted of a fear conditioning session (adapted from Phelps et al., 2004). During this session, participants viewed repeated presentations of blue and yellow squares projected on a white screen (Figure 2B). The squares served as conditioned stimuli (CS) and were presented for 4 s, followed by a 12 s inter-trial interval. The unconditioned stimulus (US) was loss of money, depicted by the symbol −$2.00 written in red font and projected inside the square for 500 ms co-terminating with the presentation of the CS. For each fear conditioning session, one colored square was paired with the monetary loss (CS+), while another colored square was never paired with the US (CS−) in a partial reinforcement design. There were 24 presentations of the CS− and 32 presentations of the CS+, 8 of which were paired with the US. Participants were instructed to watch the screen and the presentation of different colored squares. They were also told that there was a possibility they would see a −$2.00 sign, and that if it appeared, that sum would be extracted from their $36.00 total. At the end of the experiment, participants were debriefed and paid $20.00 in compensation.
Physiological set-up, assessment & behavioral analysis
Skin conductance responses (SCR) were acquired from the participant’s middle phalanges of the second and third fingers in the left hand using BIOPAC systems skin conductance module. Shielded Ag–AgCl electrodes were grounded through an RF filter panel and served to acquire data. AcqKnowledge software was used to analyze SCR waveforms. The level of SCR response was assessed as the base to peak difference for an increase in the 0.5–4.5 s window following the onset of a CS, the blue or yellow square (LaBar et al., 1995). A minimum response criterion of 0.02 μS was used with all other responses scored as 0. Responses were square-root transformed prior to statistical analysis to reduce skewness (LaBar et al. 1998). Acquired SCRs for each participant were then averaged per participant, per type of trial.
During the first part of the experiment, the gambling session, physiological responses were acquired throughout the task. Based on analysis and results from Delgado et al. (2004), where behavioral and imaging data suggested that participants were more engaged in the task during periods of high incentive than periods of low incentive, SCRs acquired at the onset of the trial (initial question mark) were analyzed. Two-tailed paired t-tests were used to compare activity at the onset of high incentive vs low incentive trials to determine the levels of arousal elicited by the gambling session (when an opportunity to earn money was presented). During the second part of the experiment, the fear conditioning session, physiological responses were also acquired throughout the task. Based on analysis and results from Phelps et al. (2004), SCRs acquired at the onset of CS presentations were analyzed. Two-tailed paired t-tests were used to compare activity of CS+ vs CS- trials. Finally, to further assess the efficacy of money during conditioning, a two-tailed correlation between the levels of arousal during gambling (the difference between high and low incentive trials) and the levels of conditioning (the difference between CS+ and CS− trials) was conducted.
Results and Discussion
During the gambling session, participants showed higher SCRs during high incentive trials when compared to low incentive trials [t(19) = 3.4, P < 0.003] suggesting they were more engaged during task performance when the incentive was monetary (Figure 3A). This result is consistent with and further supports a previous neuroimaging study (Delgado et al., 2004) that found behavioral and neural differences while participants played a gambling game that varied in the motivational context (i.e. high −low incentive trials).
Fig. 3.
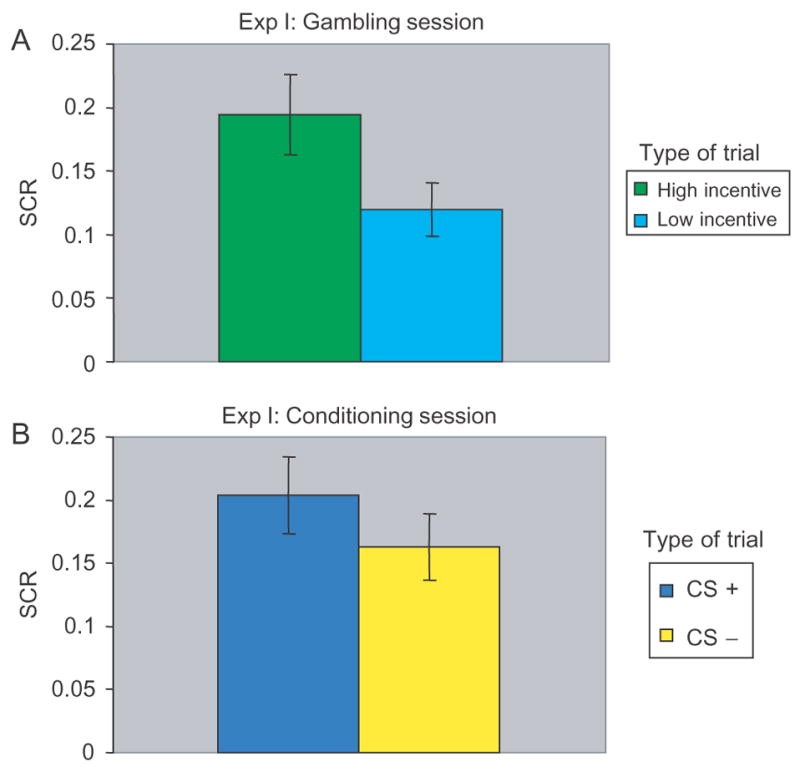
Experiment I results. (A) Skin conductance responses (SCRs) obtained during the gambling session. Participants showed higher SCRs during periods of high incentive as compared to periods of low incentive. (B) SCRs obtained during the aversive or fear conditioning session. Error bars represent standard error.
During the fear conditioning session, loss of money (−$2.00) was successful in inducing conditioning, as SCRs were higher for CS+ compared to CS− trials [t(19) = 3.09, P < 0.006; Figure 3B]. Interestingly, a correlation between the levels of conditioning (CS+ - CS− trials) and levels of arousal during gambling (high – low incentive trials) yielded a trend [r(18) = 0.43, P < 0.06], suggesting that the more aroused participants were to winning money during the gambling session, the more they ‘feared’ losing the money during the aversive conditioning session.
These results mirror previous studies of acquisition of conditioned fear in humans where the association of a neutral stimulus with an aversive event (i.e. a mild shock) leads to increased SCRs to the presentation of the previously neutral stimulus (Adolphs et al., 1995; LaBar et al., 1995, 1998; Phelps et al., 2004). More interestingly, this experiment suggests that money, a secondary reinforcer, can be used during both appetitive and aversive conditioning. It is possible, however, that a secondary reinforcer is only effective in the absence of a potentially more salient primary reinforcer. A second experiment investigated the efficacy of a secondary reinforcer, such as money, as an unconditioned stimulus during aversive conditioning when compared with a primary reinforcer, such as shock.
EXPERIMENT II
Methods
Participants
Thirty-three volunteers participated in this study (16 males, and 17 females). Two participants were removed due to technical issues with the SCR equipment and failure to comply with instructions. Final analysis was conducted on 31 participants (15 males, and 16 females). Three of these participants did not fully complete their post-questionnaires, thus self-ratings data for the remaining 28 participants are included in analysis. Participants responded to posted advertisement (average age: M = 21.26, s.d. = 3.54), and all participants gave informed consent.
Procedure
Participants were instructed that the experiment consisted of two parts. The first part consisted of a gambling session and was identical to that described the first part of Experiment I (Figure 2A). Upon completion of the gambling session, each participant was informed of their total ($36.00) and that they were about to start the second part of the experiment, which consisted of a fear conditioning session. During the session, participants viewed repeated presentations of blue, yellow and red squares projected on a white screen (Figure 4). The squares served as CS and were presented for 6 s, followed by a 10 s inter-trial interval. There were two types of US: (1) loss of money (−$2.00), lasting 750 ms and co-terminating with the CS; and (2) mild shock to the wrist, lasting 250 ms and co-terminating with the CS. For each fear conditioning session, one colored square was paired with the monetary loss (CS+), while another colored square was paired with the aversive shock (CS++) and a remaining square was never paired with any US (CS−) in a partial reinforcement setting.
Fig. 4.
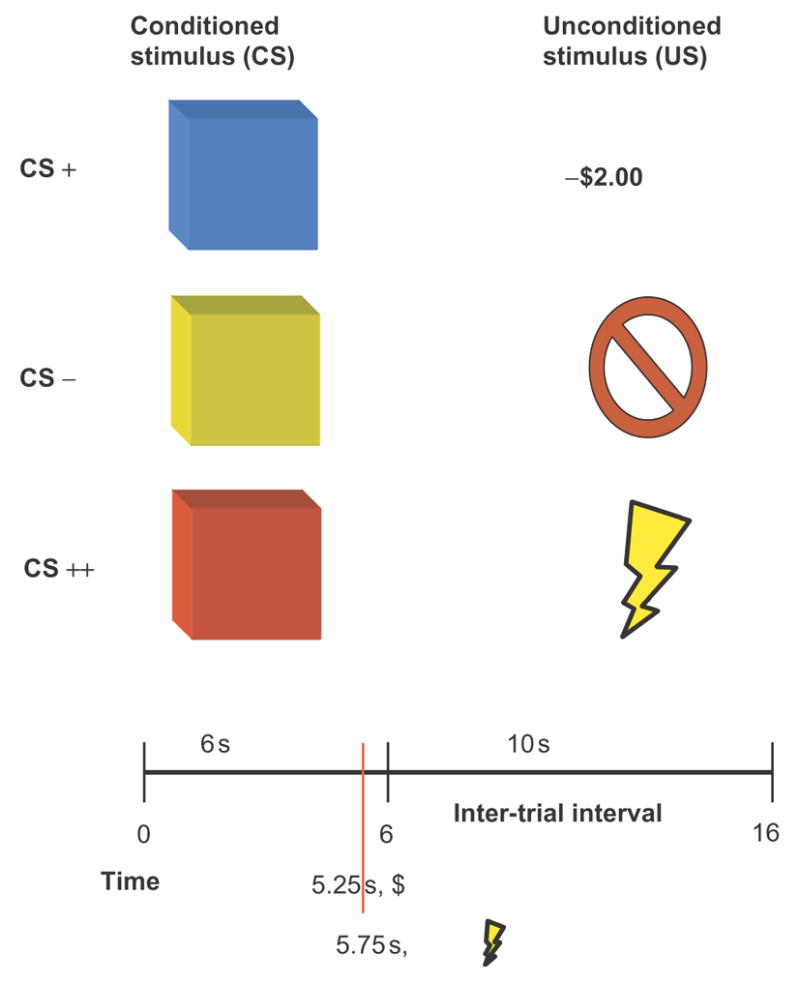
Aversive conditioning procedure for Experiment II. CS+ trials predicted potential monetary loss, CS++ trials predicted a potential shock, and CS- trials were the baseline.
There were 26 presentations of the CS− and 35 presentations of the CS+ and CS++ each, with eight of these trials being paired with their respective US. Participants were instructed to watch the screen and the presentation of different colored squares. They were also told that there was a possibility they would see a −$2.00 sign and that if it appeared, that sum would be extracted from their $36.00 total, and that in some instances they might receive a shock. At the end of the experiment, participants filled out a post-experiment questionnaire. Participants were asked to rate each square on a scale from 1 to 7 in terms of intensity and valence. They were then debriefed and paid $20.00 in compensation.
Physiological set-up, assessment and behavioral analysis
Skin conductance response (SCR) was acquired and analyzed as described in Experiment I. In addition, a Grass Instruments stimulator was used to administer mild shocks to participants during the second part. The stimulator was shielded for magnetic interference and grounded through an RF filter. A bar electrode attached to the right wrist delivered the shocks. Prior to experimental session, participants received a mild shock (200 ms duration, 50 pulses/s) which was gradually increased according to the participant’s self account. They were instructed to set their own level where the shock would feel ‘uncomfortable, but not painful’ (maximum = 50 volts).
Analysis for the gambling session was identical as Experiment I, where two-tailed paired t-tests were used to compare activity at the onset of high incentive vs low incentive trials. For the fear conditioning session, physiological responses acquired at the onset of CS presentations were analyzed. Three two-tailed paired t-tests were used to compare activity of CS++ vs CS−, CS+ vs CS− and CS++ vs CS+ trials. Similar analyses were conducted using the self-report data (both for intensity and valence) acquired through the post-experiment questionnaire. Finally, correlations between the levels of conditioning (CS+ vs CS- trials) acquired during the fear conditioning session and levels of arousal during gambling (high vs low incentive trials) were investigated, and also correlated with the subjective ratings data to further probe the efficacy of money, a secondary reinforcer, in driving aversive conditioning.
Results and Discussion
As expected, a replication of results was observed during the gambling session. Similar to Experiment I, participants showed higher SCRs during high incentive trials when compared to low incentive trials [t(30) = 5.36, P < 0.0001] suggesting they were more engaged during task performance when the incentive was monetary. After attaining $36.00 from the gambling session, participants were subjected to the fear conditioning session. A main effect of type of CS (CS+ vs CS−) was supported by a one-way ANOVA [F(2, 91) = 9.59, P < 0.003]. Additionally, post hoc t-tests suggested that both primary (shock) and secondary (money) reinforcers were successful in inducing conditioning (Figure 5A), as SCRs were higher for both CS++ [t(30) = 6.25, P < 0.0001] and CS+ [t(30) = 3.78, P < 0.007] when compared with CS− trials, respectively. Despite both reinforcers leading to conditioning, there was a significant difference between shock and money in the role of an effective US, as CS++ trials led to significantly higher SCRs than CS+ trials [t(30) = 5.85, P < 0.0001].
Fig. 5.
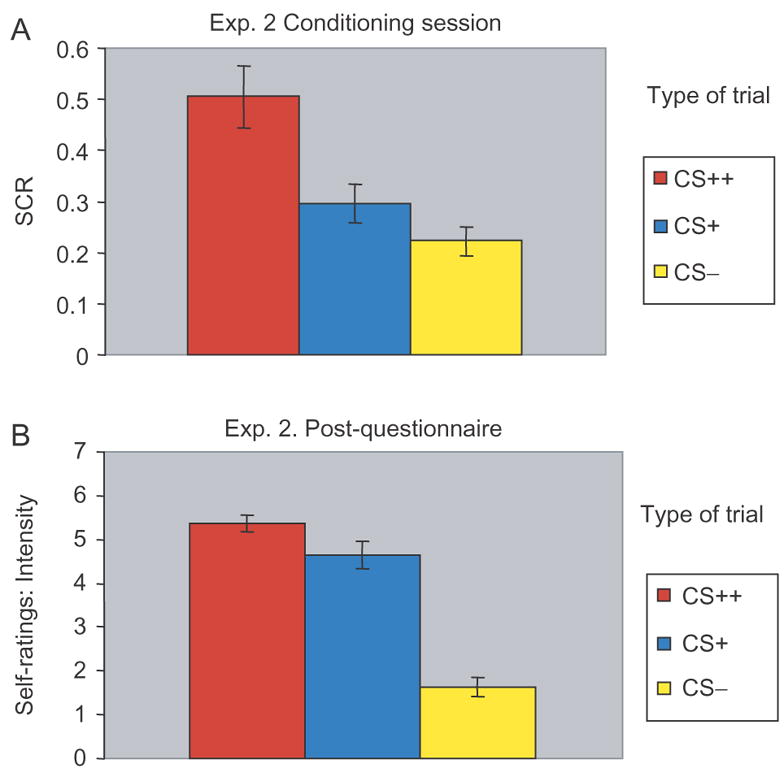
Experiment II results. (A) SCRs obtained during aversive conditioning session. (B) Self-rating of intensity. Error bars represent standard error.
At the end of the experiment participants also rated each type of CS based on intensity and valence (Figure 5B). Participants rated CS++, or shock trials, (intensity: M = 5.36, s.d. = 1.03; valence: M = 4.96, s.d. = 1.75) higher than CS- trials (intensity: M = 1.64, s.d. = 1.16; valence: M = 2.54, s.d. = 2.05) for both intensity [t(27) = 11.30, P < 0.0001] and valence [t(27) = 4.23, P < 0.0002]. Similarly, they rated CS+, or money trials, (intensity: M = 4.64, s.d. = 1.66; valence: M = 5.04, s.d. = 2.27) higher than CS− trials for both intensity [t(27) = 7.24, P < 0.0001] and valence [t(27) = 3.62, P < 0.001]. Interestingly, participants did not rate CS++ and CS+ trials differently from each other for both intensity [t(27) = 1.63, P = 0.12] and valence [t(27) = −0.13, P = 0.9].
A correlation between the levels of conditioning (CS+ or CS++ trials − CS− trials) and levels of arousal during gambling (high − low incentive trials) was observed for both money [r(28) = 0.42, P < 0.05] and shock trials [r(28) = 0.36, P < 0.05]. Interestingly, participants who showed the greatest index of arousal during gambling (as measured by differential SCR) also rated the CS+ or money trials as more intense [r(28) = 0.39, P < 0.05], but not the shock or CS++ trials [r(28) = −0.14, P = 0.49] or the CS− trials [r(28) = −0.11, P = 0.59]. Furthermore, participants who showed the greatest levels of conditioning with a secondary reinforcer also rated money trials as more intense [r(28) = 0.44, P < 0.05] and of higher valence [r(28) = 0.46, P < 0.05].
The results of Experiment II support the suggestion that a secondary reinforcer such as money can be an effective unconditioned stimulus during an aversive conditioning paradigm, although perhaps not as effective as a primary reinforcer such as shock, which elicited a higher conditioned response (as measured by SCR). Interestingly, participant’s subjective ratings of intensity and valence of both types of CSs presented during the fear conditioning session did not differ. This is peculiar because of the obvious disagreement between participant’s physiological and explicit expressions of feelings regarding the two CSs. It is possible that because both money and shock were presented during the same session, presentation context modulated the strength of the physiological conditioning (CS–US contingency). To examine this possibility, Experiment III was designed to test the same participants in two separate sessions of aversive conditioning, one with primary US and another with secondary US, each with its own neutral baseline (CS-) to facilitate comparison across the two conditions.
EXPERIMENT III
Methods
Participants
Twenty-six volunteers participated in this study (13 males and 13 females). Two participants were removed due to technical issues with the SCR equipment. Final analysis was conducted on 24 participants (12 males, 12 females). Participants responded to posted advertisement (average age: M = 21.46, s.d. = 3.40), and all participants gave informed consent.
Procedure
Participants were instructed that the experiment consisted of three parts. The first part consisted of a gambling session and was identical to that described in Experiment I (Figure 2a). Upon completion of the gambling session, each participant was informed of their total ($36.00) and that they were about to start the next two parts of the experiment (Figure 6). Both the second and third part of the experiment were separate fear conditioning sessions. During both sessions, participants viewed repeated presentations of two colored squares (e.g. blue and yellow for one session, purple and grey for the other session) projected on a white screen. The squares served as CSs and one of the squares was paired with a US (CS+), while another square was not (CS−). The main difference between the two fear conditioning sessions was that the type of US was different across sessions. For one of the sessions, a primary reinforcer such as an aversive shock, served as the US, while in the other session, a secondary reinforcer, such as loss of money (−$2.00), served as the US in a partial reinforcement design.
Fig. 6.
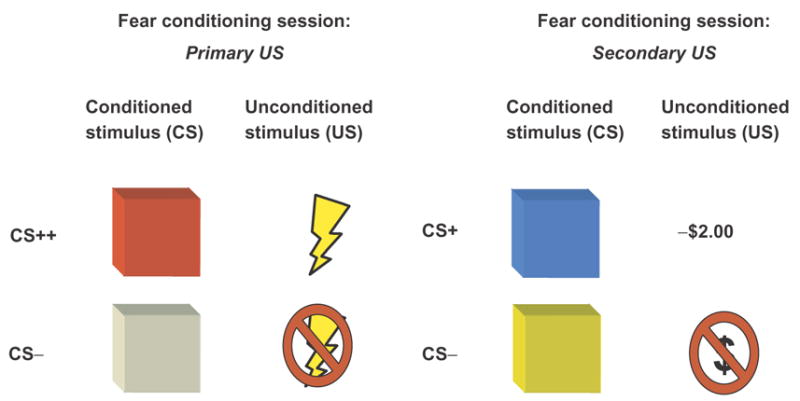
Aversive conditioning procedure Experiment III. Two separate conditioning sessions followed the gambling task. In the first session, a primary reinforcer (i.e. shock) was used as a US. In the second session, a secondary reinforcer (i.e. loss of money) was used as a US. Both sessions contained a CS- baseline.
For each conditioning session, there were 12 presentations of the CS− and 20 presentations of the CS+ each, with eight of these trials being paired with either a primary or secondary US (dependent upon session). As in Experiment II, the CS presentation was 6 s, followed by a 10 s inter-trial interval. The monetary US was 750 ms and the shock US lasted 250 ms, both co-terminating with the CS. The color of the squares and the order in which the two conditioning sessions were administered were counterbalanced across subjects. Participants were instructed to watch the screen and the presentation of different colored squares. For one fear conditioning session (primary reinforcer), they were told that they may or may not receive a shock during the session, but that they would not lose money. For the other fear conditioning session (secondary reinforcer), participants were told that there was a possibility they would see a −$2.00 sign and that if so, that sum would be extracted from their $36.00 total, but that they would not receive a shock during that session. At the end of the experiment, participants filled out a post-experiment questionnaire. Participants were asked to rate each square on a scale from 1 to 7 in terms of intensity and valence. They were then debriefed and paid $20.00 in compensation.
Physiological set-up, assessment and behavioral analysis
Skin conductance response (SCR) was acquired during all experimental parts (gambling and both fear conditioning sessions). Mild shocks were administered during one of the fear conditioning sessions as described in Experiment II. Assessment of SCRs was conducted as previously described.
Analysis for the gambling session, was identical as Experiment I, where two-tailed paired t-tests were used to compare activity at the onset of high incentive vs low incentive trials. For the fear conditioning sessions, physiological responses acquired at the onset of CS presentations were analyzed. A repeated measures ANOVA was conducted with type of CS (CS+ and CS−) and type of reinforcer (primary and secondary) as within-subjects factor to investigate the effects of primary and secondary reinforcers on conditioned fear. Similar analyses were conducted using the self-report data (both for intensity and valence) acquired through the post-experiment questionnaire.
Results and Discussion
A replication of results was once again observed during the first part of the experiment, the gambling session. Similar to the previous two experiments, participants showed higher SCRs during high incentive trials when compared to low incentive trials [t(23) = 4.22, P < 0.0003] suggesting they were more engaged during task performance when the incentive was monetary. After attaining $36.00 from the gambling session, participants were subjected to the second part (i.e. a fear conditioning session where the US was shock), followed by the third part (i.e. a fear conditioning session where the US was loss of money), with counterbalanced orders. A repeated measures ANOVA revealed a main effect of type of CS [CS+ and CS−: F(1, 23) = 19.54, P < 0.0002], suggesting that conditioning was successful irrespective of session, and a main effect of type of reinforcer (primary and secondary: F(1, 23) = 6.41, P < 0.02), suggesting that overall SCRs were higher during the fear conditioning session with a primary US (Figure 7A). However, no interaction between type of CS and reinforcer was observed [F(1, 23) = 1.58, P = 0.22], suggesting that a secondary reinforcer (i.e. money) can be as effective as a primary reinforcer (i.e. shock) in driving aversive conditioning when presented in the appropriate context.
Fig. 7.
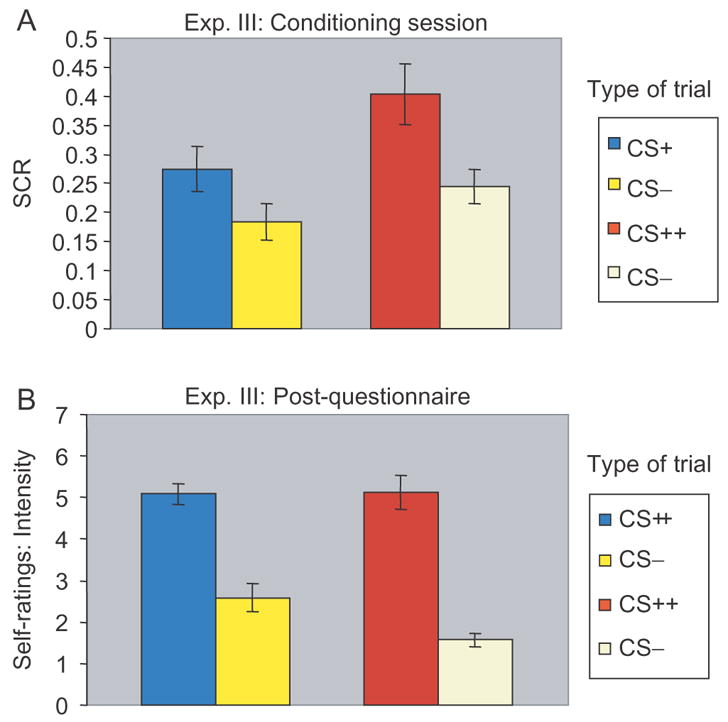
Experiment III Results. (A) SCRs obtained during both aversive conditioning sessions. (B) Self-rating of intensity. Error bars represent standard error.
As in Experiment II, participants also rated each type of CS based on intensity and valence at the end (Figure 7B). For example, during the fear conditioning session with shock as a US, participants rated both CS+, (intensity: M = 5.13, s.d. = 1.96; valence: M = 5.08, s.d. = 1.64) and CS− trials (intensity: M = 3.29, s.d. = 2.14; valence: M = 2.5, s.d. = 1.89). Similarly, during the fear conditioning session with money as an US, they rated both CS+ (intensity: M = 5.08, s.d. = 1.18; valence: M = 4.83, s.d. = 2.16) and CS− trials (intensity: M = 2.58, s.d. = 1.67; valence: M = 2.92, s.d. = 2.04). To investigate differences between subjective ratings for the separate conditioning sessions, a repeated measures ANOVA was conducted. In accordance with the SCR measures, a main effect of type of CS for both intensity [F(1, 23) = 23.81, P < 0.0001] and valence [F(1, 23) = 16.20, P < 0.001] was observed. Unlike the SCR data, however, no main effect of reinforcer was present for both intensity [F(1, 23) = 1.41, P = 0.25] and valence [F(1, 23) = 0.05, P = 0.82]. Consistent with the physiological findings, an interaction was not observed for either intensity [F(1, 23) = 1.06, P = 0.31] or valence [F(1, 23) = 0.91, P = 0.35].
As in the previous experiments, the correlation between levels of arousal during gambling (high – low incentive trials) and levels of conditioning approached significance for the money conditioning session [r(22) = 0.34, P = 0.10], although it was not observed during the shock conditioning session [r(22) = −0.24, P = 0.26]. Participants with higher levels of arousal during gambling (as measured by SCR) also rated money CS+ trials higher in terms of valence [r(22) = 0.48, P < 0.05].
Interestingly, the magnitude of physiological responses during the conditioning session with a particular reinforcer (money or shock) was correlated with the subjective ratings of that reinforcer’s valence, and negatively correlated with the intensity experienced by the opposite reinforcer. That is, participants who showed a greater conditioned response with money as a US also felt worse about trials that predicted monetary loss [money CS+ trials: valence correlation, r(22) = 0.48, P < 0.09], and felt that shock CS+ trials were of low intensity [r(22) = −0.44, P < 0.05]. In contrast, the magnitude of physiological responses during the conditioning session with shock as a reinforcer was correlated with the subjective ratings of shock valence, and negatively correlated with money intensity. That is, participants who showed a greater conditioned response with money as a US felt worse about trials that predicted a potential shock [shock CS+ trials: valence correlation, r(22) = 0.41, P < 0.05] and felt that money CS+ trials were of low intensity [r(22) = −0.41, P < 0.05].
The results of Experiment III support the suggestion that a secondary reinforcer such as money can be an effective unconditioned stimulus during an aversive conditioning paradigm. Furthermore, both physiological and subjective ratings suggest that money can be as effective as a primary reinforcer such as shock at driving conditioning when presented in the appropriate context.
GENERAL DISCUSSION
The goal of this study was to determine if money, a secondary reinforcer that acquires its reinforcing properties through social communication and interaction, could influence aversive conditioning, and to compare its efficacy with a primary reinforcer, namely shock. In three separate experiments, participants’ physiological and subjective responses to a stimulus that predicted a monetary loss were higher than responses to a non-predictive conditioned stimulus. Furthermore, loss of money was as effective in driving aversive conditioning as shock, when the reinforcers were presented in separate aversive conditioning sessions. Together, these experiments suggest that money is a powerful incentive that can have similar effects on aversive conditioning as a primary reinforcer when presented in the appropriate context, potentially highlighting the strength of reinforcers learned through association (e.g. money, social rewards) in day to day behavior.
The role of secondary reinforcers, such as money, in influencing behavior has been well characterized in society. This can be illustrated by the simple example of going to work, where the incentive of a paycheck or interaction with co-workers is evident. In the laboratory, the influence of money on behavioral, physiological and neural correlates of conditioning has also been observed for appetitive conditioning tasks (Bechara et al., 1996; Delgado et al., 2004; Zink et al., 2004). In society, however, aversive secondary reinforcers such as losing money can also impact behavior, as exemplified by risk aversion during simple decision making (Kahneman and Tversky, 2000). The current results extend our understanding of money as a reinforcer for human behavior by demonstrating that loss of money can be used as an unconditioned stimulus during aversive conditioning paradigms, being comparable to shock depending on the presentation context. These findings suggest that money is a possible common metric that can be used to compare appetitive and aversive affective learning.
Although the current results demonstrate that loss of money is as effective as shock as an unconditioned stimulus in certain circumstances, there was a discrepancy between the subjective and physiological measures of conditioning when comparing the primary and secondary reinforcers. In Experiment II, in which shock and loss of money conditioning trails were intermixed, the conditioned response to the CS+ indicating potential shock was higher than for the CS+ indicating potential loss of money. This difference was not apparent in the subjective ratings of intensity and valence for the two CSs. This suggests a potential differentiation between the implicit conditioned response to a potential negative outcome and the explicit experience attached to it, suggesting the relative intensity of conditioned response acquired with a primary or secondary reinforcer may vary when presented close in time, but only when assessed implicitly. This discrepancy, however, may not have been observed if subjective ratings were collected at the single trial level like SCR, rather than at the end of the experiment. In addition, this difference may have occurred because of unbalanced elevations in general arousal levels. When shock and money unconditioned stimuli were presented in separate conditioning sessions in Experiment III, both subjective and physiological measures of the conditioned response were equivalent. However, the general arousal levels, as measured by SCR, were higher during the shock conditioning session for both the CS+ and the CS−. Since the conditioned response was assessed as the differential SCR to the CS+ and CS−, having a common CS− baseline against which to compare the shock and loss of money CS+ responses in Experiment II may have played a role in the difference observed in the physiological assessment of the conditioned response. These results suggest that the presentation context, the baseline for comparison, and the means of assessment may be important factors in understanding the effectiveness of a secondary reinforcer in aversive conditioning. In the appropriate context, loss of money can influence aversive affective learning in a similar manner as a primary reinforcer.
As a group, the participants demonstrated robust aversive conditioning when loss of money was the unconditioned stimulus. However, there was individual variability in the strength of the conditioned response. In all three experiments, participants who were more aroused when gambling for an opportunity to earn money also showed greater SCRs when presented with a CS that represented a potential monetary loss. The same participants also felt (as assessed through subjective ratings) that the money trials were either more intense (Experiment II) or loss of money was a worse outcome (Experiment III). This suggests a potential link between ‘earning’ or working for money and being sensitive to signs that mean potential money loss. This data also illustrates that the subjective value of money varies and this variability may be related to its effectiveness as a conditioned reinforcer, as expressed both subjectively and physiologically. It would be interesting to examine if this same correlation would be apparent with a gambling population that may be insensitive to losses attained during gambling due to discounting (Rachlin, 1990).
In the current study, we specifically examined if the presence of a primary reinforcer altered the effectiveness of the secondary reinforcer in aversive conditioning. Another factor that may have played a role in the magnitude of this effect that was not examined was the instrumental action, or the act of earning the money, that preceded the aversive conditioning procedure. Before each conditioning session, participants earned money through a gambling task to create an endowment for future losses. It is possible this resulted in an endowment effect and heightened loss aversion (Kahneman and Tversky, 2002). It is unclear if participants were merely handed the money at the onset of the study whether the conditioning paradigm would have been as effective. In fact, some neuroimaging studies suggest that the perception of control regarding the outcome of an action, or ‘earning’ money through behavior, recruits the neural circuitry of reward-related processes more robustly than situations where money is just received (Tricomi et al., 2004; Zink et al., 2004). Such discrepancy might exemplify a potential difference between primary and secondary reinforcers. Along these lines, it might be interesting to compare primary and secondary reinforcers in an instrumental aversive conditioning paradigm.
Another issue raised by this current study is how to properly titrate different reinforcers when comparing primary and secondary reinforcers in aversive conditioning. While the $2.00 money reinforcer was fixed throughout participants in our study, the shock level was set by each participant based on their subjective measure of tolerance. As observed in Experiment II, the general arousal levels were higher during the prediction of a potential shock. This may not have been apparent if the potential monetary loss was greater, or the study was conducted in a population that was more sensitive to small losses in money. Although we were able to find evidence for aversive conditioning with loss of money as an unconditioned stimulus, a proper comparison between the relative effectiveness of primary and socially defined, secondary reinforcers may be difficult because of the appropriate titration of perceived negative impact and individual differences in the sensitivity to social and cultural reinforcers.
These experiments and other studies that use money as a reinforcer also raise the question of whether money, due to its societal importance, may not be a typical example of a secondary reinforcer, and may act more like a primary reinforcer. Perhaps money, because it is an overlearned reinforcer in society, is in a class by itself. It is unclear, for example, how learning through the potential loss of money is related to other types of secondary reinforcers. How might money compare with different types of socially defined secondary reinforcers (e.g. pat on the back, compliments) and how such reinforcers interact with motivation levels (e.g. intrinsic −extrinsic)?
In order to understand potential overlaps or differences between primary and socially defined, secondary reinforcers in aversive conditioning, a comparison of their underlying neural systems would be helpful. If both shock and money are successful in modulating aversive conditioning then they should both rely in similar neural mechanisms, most notably the amygdala. However, studies have shown that the amygdala is involved in processing information about stimulus intensity (Anderson et al., 2003; Small et al., 2003), suggesting perhaps a possible dissociation between primary and secondary reinforcers that is difficult to remedy. In contrast, other brain regions such as the human striatum, involved in updating representations of values during appetitive and aversive learning (Gottfried et al., 2002; Jensen et al., 2003; McClure et al., 2003; O’Doherty et al., 2003; O’Doherty 2004; Seymour et al., 2004; Delgado et al., 2005; Knutson and Cooper, 2005), may not differentiate between primary and secondary reinforcers if their effect on behavior is similar.
In spite of all the remaining questions for future investigations, the current results clearly demonstrate that money, a secondary reinforcer and a stimulus that acquires its reinforcing properties through social and cultural means, can influence aversive conditioning. Depending on the context of presentation, money can be as effective as a primary reinforcer in driving conditioning, suggesting a socially defined reinforcer’s properties may be malleable depending on the situation.
Footnotes
Conflict of Interest
None declared.
The authors would like to acknowledge Joe LeDoux for useful discussion and the support of the NIMH (MH62104) and the James S. McDonnell Foundation to EAP.
References
- Adolphs R, Tranel D, Damasio H, Damasio AR. Fear and the human amygdala. Journal of Neuroscience. 1995;15:5879–91. doi: 10.1523/JNEUROSCI.15-09-05879.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson AK, Christoff K, Stappen I, et al. Dissociated neural representations of intensity and valence in human olfaction. Nature Neuroscience. 2003;6:196–202. doi: 10.1038/nn1001. [DOI] [PubMed] [Google Scholar]
- Bechara A, Tranel D, Damasio H, Damasio AR. Failure to respond autonomically to anticipated future outcomes following damage to prefrontal cortex. Cerebral Cortex. 1996;6:215–25. doi: 10.1093/cercor/6.2.215. [DOI] [PubMed] [Google Scholar]
- Breiter HC, Gollub RL, Weisskoff RM, et al. Acute effects of cocaine on human brain activity and emotion. Neuron. 1997;19:591–611. doi: 10.1016/s0896-6273(00)80374-8. [DOI] [PubMed] [Google Scholar]
- Breiter HC, Rosen BR. Functional magnetic resonance imaging of brain reward circuitry in the human. Annals of the New York Academy of Sciences. 1999;877:523–47. doi: 10.1111/j.1749-6632.1999.tb09287.x. [DOI] [PubMed] [Google Scholar]
- Cardinal RN, Everitt BJ. Neural and psychological mechanisms underlying appetitive learning: links to drug addiction. Current Opinion in Neurobiology. 2004;14:156–62. doi: 10.1016/j.conb.2004.03.004. [DOI] [PubMed] [Google Scholar]
- Davis M. The role of the amygdala in fear and anxiety. Annual Review of Neuroscience. 1992;15:353–75. doi: 10.1146/annurev.ne.15.030192.002033. [DOI] [PubMed] [Google Scholar]
- Delgado MR, Locke HM, Stenger VA, Fiez JA. Dorsal striatum responses to reward and punishment: effects of valence and magnitude manipulations. Cognitive, Affective & Behavioural Neuroscience. 2003;3:27–38. doi: 10.3758/cabn.3.1.27. [DOI] [PubMed] [Google Scholar]
- Delgado MR, Miller MM, Inati S, Phelps EA. An fMRI study of reward-related probability learning. Neuroimage. 2005;24:862–73. doi: 10.1016/j.neuroimage.2004.10.002. [DOI] [PubMed] [Google Scholar]
- Delgado MR, Nystrom LE, Fissell C, Noll DC, Fiez JA. Tracking the hemodynamic responses to reward and punishment in the striatum. Journal of Neurophysiology. 2000;84:3072–7. doi: 10.1152/jn.2000.84.6.3072. [DOI] [PubMed] [Google Scholar]
- Delgado MR, Stenger VA, Fiez JA. Motivation-dependent Responses in the Human Caudate Nucleus. Cerebral Cortex. 2004;14:1022–30. doi: 10.1093/cercor/bhh062. [DOI] [PubMed] [Google Scholar]
- Elliott R, Friston KJ, Dolan RJ. Dissociable neural responses in human reward systems. Journal of Neuroscience. 2000;20:6159–65. doi: 10.1523/JNEUROSCI.20-16-06159.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Everitt BJ, Cardinal RN, Parkinson JA, Robbins TW. Appetitive behavior: impact of amygdala-dependent mechanisms of emotional learning. Annals of the New York Academy of Sciences. 2003;985:233–50. [PubMed] [Google Scholar]
- Gottfried JA, O’Doherty J, Dolan RJ. Appetitive and aversive olfactory learning in humans studied using event-related functional magnetic resonance imaging. Journal of Neuroscience. 2002;22:10829–37. doi: 10.1523/JNEUROSCI.22-24-10829.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hull CL. Principles of Behavior, an Introduction to Behavior Theory. New York: D. Appleton-Century Company; 1943. [Google Scholar]
- Jensen J, McIntosh AR, Crawley AP, Mikulis DJ, Remington G, Kapur S. Direct activation of the ventral striatum in anticipation of aversive stimuli. Neuron. 2003;40:1251–7. doi: 10.1016/s0896-6273(03)00724-4. [DOI] [PubMed] [Google Scholar]
- Kahneman D, Tversky A. Choices, Values, and Frames. New York, and Cambridge, UK: Russell Sage Foundation and Cambridge University Press; 2000. [Google Scholar]
- Knutson B, Adams CM, Fong GW, Hommer D. Anticipation of increasing monetary reward selectively recruits nucleus accumbens. Journal of Neuroscience. 2001a;21:RC159. doi: 10.1523/JNEUROSCI.21-16-j0002.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knutson B, Cooper JC. Functional magnetic resonance imaging of reward prediction. Current Opinion in Neurology. 2005;18:411–7. doi: 10.1097/01.wco.0000173463.24758.f6. [DOI] [PubMed] [Google Scholar]
- Knutson B, Fong GW, Adams CM, Varner JL, Hommer D. Dissociation of reward anticipation and outcome with event-related fMRI. Neuroreport. 2001b;12:3683–7. doi: 10.1097/00001756-200112040-00016. [DOI] [PubMed] [Google Scholar]
- LaBar KS, Gatenby JC, Gore JC, LeDoux JE, Phelps EA. Human amygdala activation during conditioned fear acquisition and extinction: a mixed-trial fMRI study. Neuron. 1998;20:937–45. doi: 10.1016/s0896-6273(00)80475-4. [DOI] [PubMed] [Google Scholar]
- LaBar KS, LeDoux JE, Spencer DD, Phelps EA. Impaired fear conditioning following unilateral temporal lobectomy in humans. Journal of Neuroscience. 1995;15:6846–55. doi: 10.1523/JNEUROSCI.15-10-06846.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LeDoux JE. Emotion circuits in the brain. Annual Review of Neuroscience. 2000;23:155–84. doi: 10.1146/annurev.neuro.23.1.155. [DOI] [PubMed] [Google Scholar]
- McClure SM, Berns GS, Montague PR. Temporal prediction errors in a passive learning task activate human striatum. Neuron. 2003;38:339–46. doi: 10.1016/s0896-6273(03)00154-5. [DOI] [PubMed] [Google Scholar]
- O’Doherty JP. Reward representations and reward-related learning in the human brain: insights from neuroimaging. Current Opinion in Neurobiology. 2004;14:769–76. doi: 10.1016/j.conb.2004.10.016. [DOI] [PubMed] [Google Scholar]
- O’Doherty JP, Dayan P, Friston K, Critchley H, Dolan RJ. Temporal difference models and reward-related learning in the human brain. Neuron. 2003;38:329–37. doi: 10.1016/s0896-6273(03)00169-7. [DOI] [PubMed] [Google Scholar]
- O’Doherty JP, Deichmann R, Critchley HD, Dolan RJ. Neural responses during anticipation of a primary taste reward. Neuron. 2002;33:815–26. doi: 10.1016/s0896-6273(02)00603-7. [DOI] [PubMed] [Google Scholar]
- Pare D, Quirk GJ, Ledoux JE. New vistas on amygdala networks in conditioned fear. Journal of Neurophysiology. 2004;92:1–9. doi: 10.1152/jn.00153.2004. [DOI] [PubMed] [Google Scholar]
- Parkinson JA, Cardinal RN, Everitt BJ. Limbic cortical-ventral striatal systems underlying appetitive conditioning. Progress in Brain Research. 2000;126:263–85. doi: 10.1016/S0079-6123(00)26019-6. [DOI] [PubMed] [Google Scholar]
- Pavlov IP, Anrep GV. Conditioned Reflexes. An Investigation of the Physiological Activity of the Cerebral Cortex. London: Oxford University Press; Humphrey Milford; 1927. [Google Scholar]
- Phelps EA, Delgado MR, Nearing KI, LeDoux JE. Extinction learning in humans: role of the amygdala and vmPFC. Neuron. 2004;43:897–905. doi: 10.1016/j.neuron.2004.08.042. [DOI] [PubMed] [Google Scholar]
- Rachlin H. Why do people gamble and keep gambling despite heavy losses? Psychological Science. 1990;1:294–7. [Google Scholar]
- Rescorla RA, Wagner AR. A theory of Pavlovian conditioning: variations in the effectiveness of reinforcement and nonreinforcement. In: Black A, Prokasy WF, editors. Classical Conditioning II: Current Research and Theory. New York: Appleton-Century-Crofts; 1972. pp. 64–99. [Google Scholar]
- Seymour B, O’Doherty JP, Dayan P, et al. Temporal difference models describe higher-order learning in humans. Nature. 2004;429:664–7. doi: 10.1038/nature02581. [DOI] [PubMed] [Google Scholar]
- Skinner BF. The Behavior of Organisms: An Experimental Analysis. New York, London: D. Appleton-Century Company; 1938. [Google Scholar]
- Skinner BF. Science and Human Behavior. New York: Macmillan; 1953. [Google Scholar]
- Small DM, Gregory MD, Mak YE, Gitelman D, Mesulam MM, Parrish T. Dissociation of neural representation of intensity and affective valuation in human gustation. Neuron. 2003;39:701–11. doi: 10.1016/s0896-6273(03)00467-7. [DOI] [PubMed] [Google Scholar]
- Tricomi EM, Delgado MR, Fiez JA. Modulation of caudate activity by action contingency. Neuron. 2004;41:281–92. doi: 10.1016/s0896-6273(03)00848-1. [DOI] [PubMed] [Google Scholar]
- Zink CF, Pagnoni G, Martin-Skurski ME, Chappelow JC, Berns GS. Human striatal responses to monetary reward depend on saliency. Neuron. 2004;42:509–17. doi: 10.1016/s0896-6273(04)00183-7. [DOI] [PubMed] [Google Scholar]