

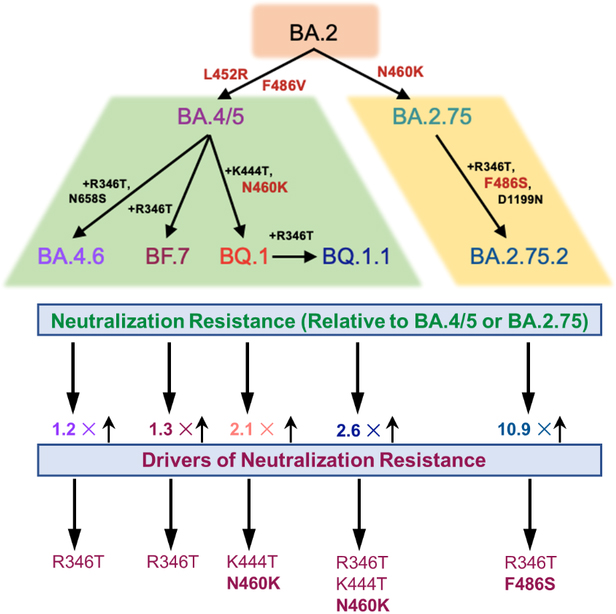
SUMMARY
The continued evolution of SARS-CoV-2 has led to the emergence of several new Omicron subvariants, including BQ.1, BQ.1.1, BA.4.6, BF.7 and BA.2.75.2. Here we examine the neutralization resistance of these subvariants against sera from 3-dose vaccinated health care workers, hospitalized BA.1-wave patients, and BA.4/5-wave patients. We found enhanced neutralization resistance in all new subvariants, especially the BQ.1 and BQ.1.1 subvariants driven by the N460K and K444T mutations, as well as the BA.2.75.2 subvariant driven largely by its F486S mutation. All Omicron subvariants maintained their weakened infectivity in Calu-3 cells, with the F486S mutation driving further diminished titer for the BA.2.75.2 subvariant. Molecular modelling revealed the mechanisms of antibody-mediated immune evasion by the R346T, K444T, F486S, and D1199N mutations. Altogether, these findings shed light on the evolution of newly emerging SARS-CoV-2 Omicron subvariants.
Blurbs
Numerous Omicron subvariants have emerged following the BA.4/5 and BA.2.75 subvariants. Qu and colleagues investigate the neutralizing antibody resistance of these subvariants and their ancestral variants. BQ.1, BQ.1.1, BA.4.6, BF.7 and BA.2.75.2 exhibit enhanced neutralizing antibody escape, with BQ.1/BQ.1.1 and BA.2.75.2 driven by N460K/K444T and F486S mutations, respectively.
Graphical Abstract
INTRODUCTION
Since its emergence in late 2021, the Omicron variant of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has led to the emergence of numerous subvariants that continue to evade vaccine- and infection-induced immunity and alter the virus biology1–7. The initial BA.1 Omicron subvariant drove a large wave of coronavirus disease 2019 (COVID-19) cases and exhibited strong immune escape from 2-mRNA vaccine dose-induced immunity that was recovered by booster mRNA vaccine administration1,8–14. In addition, the BA.1 subvariant exhibited reduced cell-cell fusogenicity, impaired replication in lower airway epithelial cells and lung derived CaLu-3 cells, as well as reduced plasma membrane entry through transmembrane protease serine 2 (TMPRSS2)15–20. These features correlated with reduced replication capacity of Omicron in lung tissues, enhanced nasopharyngeal tropism, and decreased pathogenicity in vivo15,18,20–22. Importantly, these characteristics were largely maintained by subsequent Omicron subvariants.
The BA.2 subvariant overtook BA.1 due to its slightly enhanced transmissibility and immune evasion, with an ability to reinfect individuals who were previously infected with BA.123–25. From BA.2, several subvariants emerged in quick succession, often with concurrent circulations; these included the BA.4 and BA.5 subvariants (bearing identical S proteins, referred to as BA.4/5 hereafter) that next rose to dominance, and exhibited further immune escape4,26–31. In addition, BA.2 gave rise to the BA.2.75 subvariant, which is currently increasing in proportion of COVID-19 cases32, but does not exhibit as substantial immune escape compared with BA.4/53,33–35. Notably, the BA.4/5 and BA.2.75 subvariants have driven further diversification of the circulating SARS-CoV-2, with the emergence of several additional subvariants including the BA.4.6, BF.7, BQ.1, and BQ.1.1 (derived from BA.4/5), as well as BA.2.75.2 (derived from BA.2.75). Additionally, these new subvariants are currently increasing in frequency32,36 and may be the next major dominant Omicron subvariant.
The extent of immune evasion and functional alterations to the spike protein (S) in these emerging Omicron subvariants currently remains unclear. To address this, we examine the sensitivity of the BA.4.6, BF.7, BQ.1, BQ.1.1, and BA.2.75.2 subvariants to neutralization by serum from recipients of 3 mRNA vaccine doses, as well as COVID-19 patients infected with the BA.1 or BA.4/5 variants. Additionally, we examined the contribution of subvariant lineage defining mutations to neutralization escape. We observe strong neutralization resistance in the BQ.1 and BQ.1.1 subvariants driven largely by their N460K mutation, as well as in the BA.2.75.2 subvariant driven largely by its F486S mutation. Notably, the F486S diminishes BA.2.75.2 infectivity in Calu-3 cells. Finally, structural modeling showed that the F486S mutation reduces binding affinity for both the angiotensin-converting enzyme 2 (ACE2) receptor and class I and II antibodies, while the R346T and K444T mutation are likely responsible for evasion of class III antibody recognition.
RESULTS
New Omicron subvariants exhibit similarly increased infectivity in HEK293T-ACE2 cells but comparably decreased infectivity in Calu-3 cells
Derived from the BA.4/5 subvariant, BF.7 acquires an additional R346T mutation whereas BA.4.6 and BQ.1 contain R346T/N658S mutations and K444T/N460K mutations, respectively; BQ.1.1 also bears an additional R346T mutation compared with BQ.1 (Fig. 1A). The BA.2.75.2 subvariant acquires R346T, F486S and D1199N mutations in contrast to BA.2.75 (Fig. 1A). These mutations, particularly those on the receptor-binding domain (RBD), have caused increasing concern over further immune escape37.
Figure 1: Omicron subvariant-defining mutations and their impacts on pseudotyped viral infectivity in HEK293T-ACE2 and Calu-3 cells.
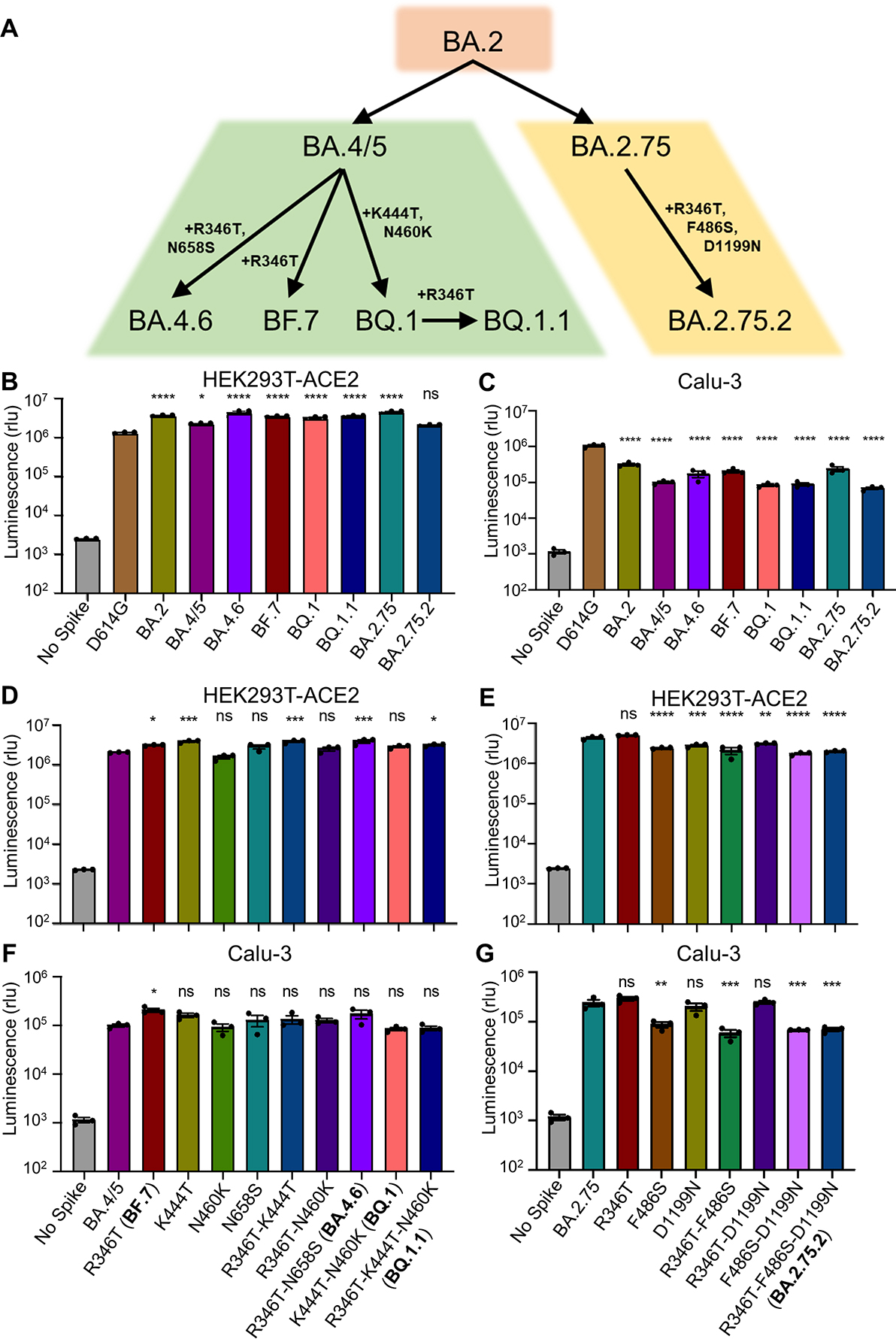
(A) Displayed is a schematic of SARS-CoV-2 Omicron subvariant evolution indicating the mutations acquired by the BA.4.6, BF.7, BQ.1, BQ.1.1, and BA.2.75.2 subvariants. (B-C) Infectivity of lentivirus pseudotyped with the indicated Omicron subvariant S constructs in HEK293T-ACE2 cells (B) (n = 3) or in Calu-3 cells (C) (n = 3). Bars represent means +/− standard error. Significance relative to D614G was determined by one-way ANOVA with Bonferroni’s multiple testing correction. P values are represented as ns for p ≥ 0.05, *p < 0.05, and ****p < 0.0001. (D-G) Infectivity of lentivirus pseudotyped with the indicated BA.4/5-derived mutant S constructs (D, F) (n = 3) or BA.2.75-derived mutant S constructs (E, G) (n = 3) in HEK293T-ACE2 or Calu-3 cells. Bars represent means +/− standard error. Significance relative to BA.4/5 or BA.2.75 was determined by one-way ANOVA with Bonferroni’s multiple testing correction. P values are represented as ns for p ≥ 0.05, *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001.
We first examined the infectivity of these subvariants, along with the individual mutants and D614G by producing lentivirus pseudotyped with S from each of the critical S constructs (Fig. 1B–G). All Omicron subvariant-pseudotyped viruses exhibited modestly enhanced infectivity in HEK293T-ACE2 cells over the D614G variant with the exception of BA.2.75.2 (Fig. 1B). Additionally, all Omicron subvariants exhibited comparably poor infectivity in lung-derived Calu-3 cells (Fig. 1C) compared to D614G, consistent with prior Omicron subvariants and the weak lung tropism observed for Omicron15–18. We found that these panels of mutants also exhibited similar infectivity compared to their parental BA.4/5 or BA.2.75 variants in HEK293T-ACE2 cells (Fig. 1D–E) and Calu-3 cells (Fig. 1F–G), although F486S-containing/BA.2.75-derived mutants, including the triple mutant R346T-F486S-D1199N (i.e., BA.2.75.2), had reduced titers in Calu-3 cells relative to the parental BA.2.75 (Fig. 1G).
BQ.1, BQ.1.1, and BA.2.75.2 exhibit potent neutralization resistance to vaccine- and infection-induced sera.
Using our previously reported pseudotyped lentivirus neutralization assay38, we first examined the resistance of these emerging Omicron subvariants to sera from health care workers (HCWs) collected 2–13 weeks after vaccination with a homologous booster dose of monovalent Moderna mRNA-1273 (n = 3) or Pfizer/BioNTech BNT162b2 vaccine (n = 12). Compared to D614G, the BA.4.6, BF.7, BQ.1, and BQ.1.1 subvariants exhibited a 10.6-fold (p < 0.0001), 11.0-fold (p < 0.0001), 18.7-fold (p < 0.0001), and 22.9-fold (p < 0.0001) higher neutralization resistance, respectively, while BA.4/5 exhibited an 8.7-fold (p < 0.0001) higher neutralization resistance (Fig. 2A and Fig. S1A). Similarly, compared to D614G, the BA.2.75.2 subvariant exhibited a 48.4-fold (p < 0.0001) higher neutralization resistance while BA.2.75 exhibited a 4.4-fold (p < 0.0001) higher neutralization resistance (Fig. 2A). These data indicate further neutralization escape in emerging Omicron subvariants, with BA.Q.1, BA.Q.1.1, and BA.2.75.2, especially the latter, showing the most substantial neutralization resistance.
Figure 2: Omicron subvariants, especially BQ.1, BQ.1.1, and BA.2.75.2, exhibit strong neutralization resistance.
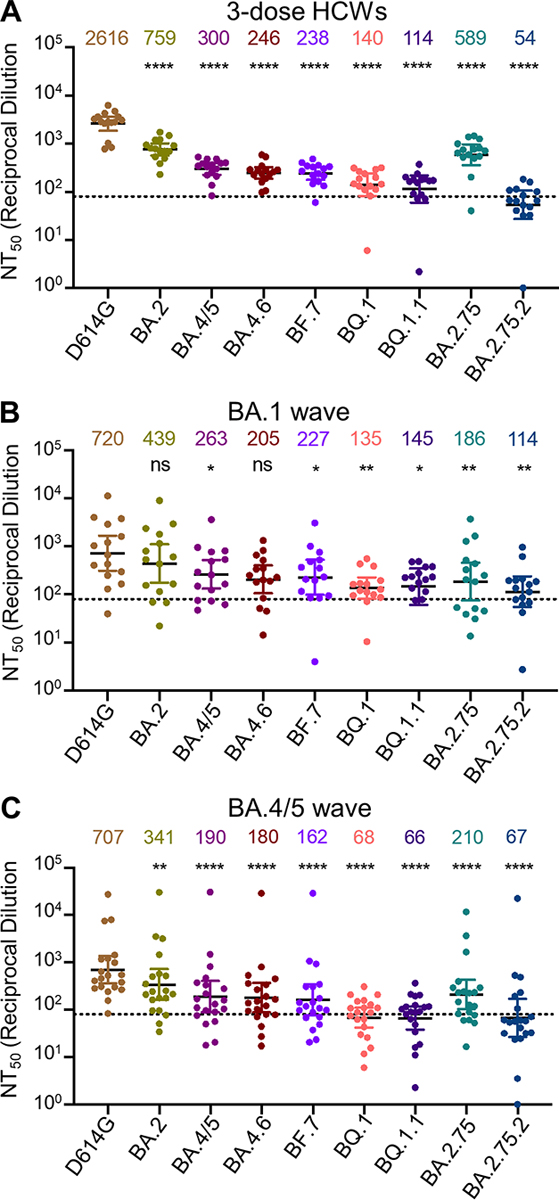
Neutralizing antibody titers were determined against lentivirus pseudotyped with S from the Omicron subvariants and ancestral D614G variant for sera from health care workers (HCWs) (n = 15) who received a single homologous monovalent Moderna mRNA-1273 (n = 3) or Pfizer/BioNTech BNT162b2 (n = 12) mRNA booster vaccination (A); for sera from BA.1-wave hospitalized COVID-19 patients (n = 15) (B); or for sera from BA.4/5-wave SARS-CoV-2 infected Columbus, Ohio first responders and household contacts (n = 20) (C). Bars represent geometric means with 95% confidence intervals. Geometric means are displayed at the top of the plots. Significance relative to D614G was determined by one-way repeated measures ANOVA with Bonferroni’s multiple testing correction. P values are displayed as ns for p ≥ 0.05, *p < 0.05, **p < 0.01, and ****p < 0.0001.
We also examined the resistance of these Omicron subvariants to neutralization by sera from hospitalized COVID-19 patients (n = 15) infected during the BA.1 wave of the pandemic. These include 6 unvaccinated patients, 5 patients vaccinated with 2 doses of Pfizer/BioNTech BNT162b2 or Moderna mRNA-1273, and 4 patients with 3 doses of Pfizer/BioNTech BNT162b2. BA.1-wave patient sera exhibited a similar pattern with the BA.4.6, BF.7, BQ.1, BQ.1.1, and BA.2.75.2 subvariants exhibiting the strongest immune evasion, with neutralization resistance 3.5-fold (p > 0.05), 3.2-fold (p < 0.05), 5.3-fold (p < 0.01), 5.0-fold (p < 0.05), and 6.3-fold (p < 0.01) higher than D614G, respectively (Fig. 2B and Fig. S1B). As would be expected, BA.1 patient sera neutralized BA.2 with a higher efficiency compared to these BA.2-derived subvariants (Fig. 2B and Fig. S1B).
To determine the breadth of immunity from individuals infected with more recent Omicron subvariants, we next examined sera from Columbus, Ohio first responders and household contacts testing positive for COVID-19 during the BA.4/5 wave of the pandemic, with 11/20 subjects having the infecting variant confirmed by sequencing. Compared to D614G, the BA.4.6, BF.7, BQ.1, and BQ.1.1 subvariants exhibited a 3.9-fold (p < 0.0001), 4.4-fold (p < 0.0001), 10.4-fold (p < 0.0001), and 10.7-fold (p < 0.0001) higher neutralization resistance, respectively, while BA.4/5 exhibited a 3.7-fold (p < 0.0001) higher neutralization resistance than D614G (Fig. 2C and Fig. S1C). Additionally, compared to D614G, the BA.2.75.2 subvariant exhibited a 10.6-fold (p < 0.0001) higher neutralization resistance whereas BA.2.75 showed a 3.4-fold (p < 0.0001) higher neutralization resistance (Fig. 2C). Interestingly, BA.2 exhibited less resistance to sera from BA.4/5-wave infected patients than BA.4/5, with 2.1-fold reduced neutralization sensitivity compared to D614G (p < 0.01) (Fig. 2C and Fig. S1C). Similar trends were observed for both BA.1-wave and BA.4/5-wave patients regardless of vaccination status (Fig. S2). Together, these results showed that BQ.1, BQ.1.1, and BA.2.75.2 are strongly resistant to neutralization by sera from subjects infected with the recently dominant BA.4/5 variant.
The N460K mutation is the most critical for the enhanced neutralization resistance of BA.4.6, BF.7, BQ.1 and BQ.1.1
We further examined the amino acid residues critical for the neutralization resistance to sera from health care workers (HCWs), BA.1 infection, as well as and BA.4/5-infected individuals. Compared to the parental BA.4/5 subvariant, introduction of the N460K mutation reduced neutralization sensitivity by an additional 2.6-fold (p < 0.0001), with a similar 2.8-fold (p < 0.0001) reduction for the R346T/N460K double mutant (Fig. 3A). Consistent with this finding, the N460K-bearing subvariants BQ.1 and BQ.1.1 showed the strongest neutralization resistance with a 2.1-fold (p < 0.01) and 2.6-fold (p < 0.01) reduction in neutralization sensitivity compared to BA.4/5 (Fig. 3A and Fig. S3A). Though not significant, two other individual mutations, R346T and K444T, were associated with a milder 1.3-fold (p = 0.0782) and 1.4-fold (p = 0.1886) reduction in neutralization sensitivity, respectively (Fig. 3A). Of note, the N658S mutation did not appear to be strongly associated with neutralization resistance (Fig. S3A), with a 1.1-fold (p > 0.05) reduction compared to the parental BA.4/5 (Fig. 3A).
Figure 3: Mutations N460K and F486S, and to a lesser extent, R346T and K444T, drive Omicron subvariant neutralization resistance.
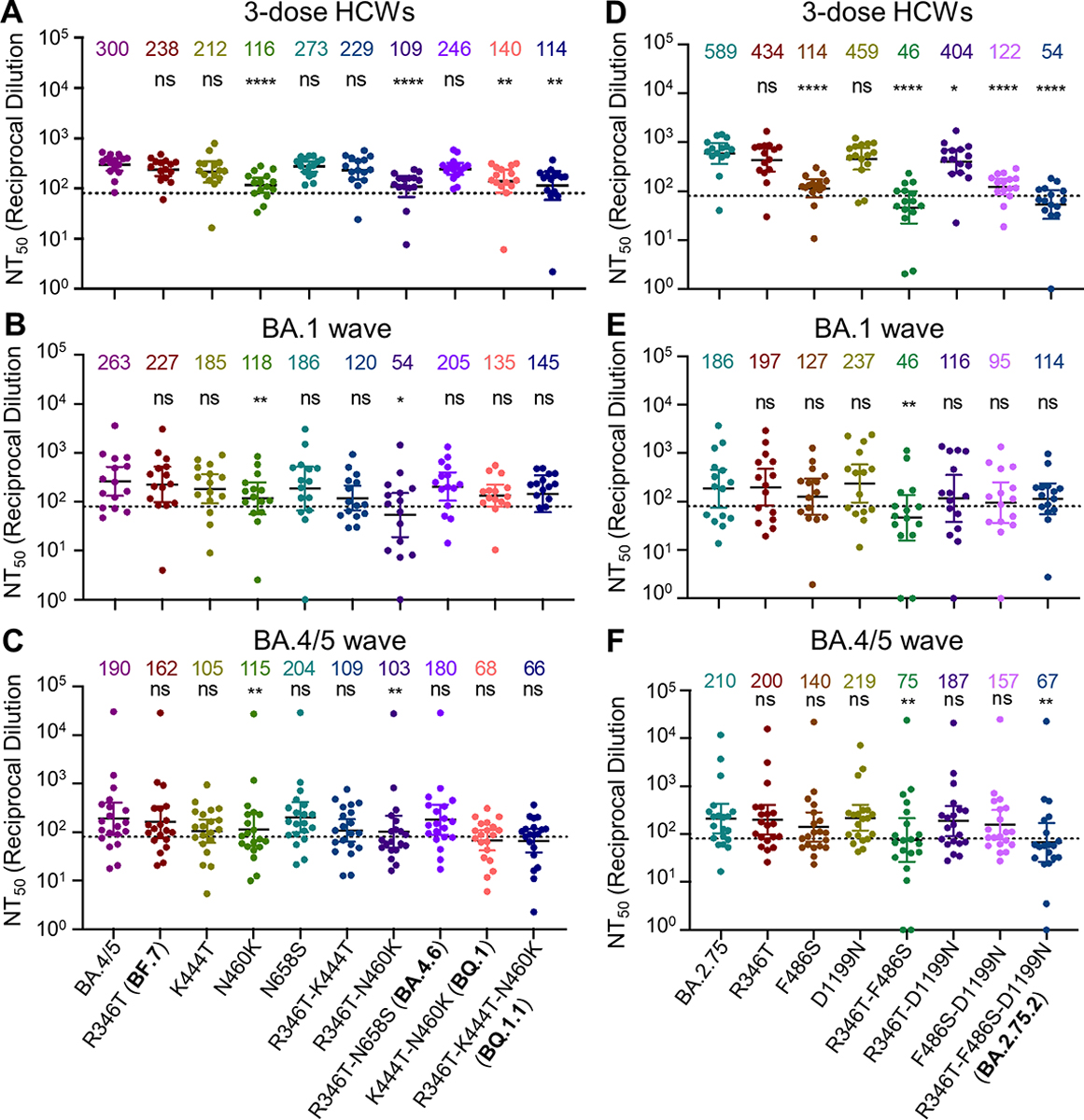
Neutralizing antibody titers were determined against lentivirus pseudotyped with S from the BA.4/5-derived (A, B and C) or BA.2.75 (D, E, and F) mutants for sera from health care workers (HCWs) (A and D) (n = 15) who received a single homologous monovalent Moderna mRNA-1273 (n = 3) or Pfizer/BioNTech BNT162b2 (n = 12) mRNA booster vaccination; for sera from BA.1-wave hospitalized COVID-19 patients (B and E) (n = 15); or for sera from BA.4/5-wave SARS-CoV-2 infected Columbus, Ohio first responders and household contacts (C and F) (n = 20). Bars represent geometric means with 95% confidence intervals. Geometric means are displayed at the top of the plots. Significance relative to parental variants was determined by one-way repeated measures ANOVA with Bonferroni’s multiple testing correction. P values are displayed as ns for p ≥ 0.05, *p < 0.05, **p < 0.01, and ****p < 0.0001.
We next examined the determinants of resistance of these Omicron subvariant mutants to neutralization by sera from hospitalized COVID-19 patients infected during the BA.1 wave of the pandemic. In addition, we tested sera from Columbus, Ohio first responders and household contacts testing positive for COVID-19 during the BA.4/5 wave of the pandemic. We found that N460K and R346T/N460K mutations in the context of BA.4/5 reduced neutralization sensitivity to BA.1 sera 2.2-fold (p < 0.01) and 4.9-fold (p < 0.05), respectively (Fig. 3B and Fig. S3B), and to BA.4/5 sera 1.7-fold (p < 0.01) and 1.8-fold (p < 0.01), respectively (Fig. 3C and Fig. S3C). Notably, two BA.4/5-wave patients (BA.4/5_P5 and BA.4/5_P17) and one BA.1-wave patient (BA.1_P11) exhibited starkly reduced neutralization of K444T containing mutants, indicating the K444T mutation may be critical for evading the neutralizing antibody response for a subset of Omicron infected patients (Fig. S3B–C). Together with the results from HCWs, we concluded that the N460K mutation, and to a lesser extent R346T, K444T, and N658S, are critical for the enhanced resistance of the BQ.1 and BQ.1.1 subvariants.
The F486S mutation drives the enhanced neutralization resistance of BA.2.75.2
We also interrogated the critical amino acid residues responsible for the substantially enhanced nAb resistance of the BA.2.75.2 subvariant to sera from the same cohorts of HCWs, BA.1-wave patients, as well as BA.5-infected individuals mentioned above. We found that introduction of the F486S, R346T/F486S, or F486S/D1199N mutations to the parental BA.2.75 reduced neutralization sensitivity to HCW sera 5.2-fold (p < 0.0001), 12.8-fold (p < 0.0001), 4.8-fold (p < 0.0001), respectively; these were in comparison with the 10.9-fold (p < 0.0001) reduction seen for the BA.2.75.2 subvariant which contains all three mutations, i.e., R346T, F486S and D1199N (Fig. 3D). Notably, introduction of the R346T, D1199N, or R346T/D1199N resulted in only a 1.4-fold (p = 0.1074), 1.3-fold (p > 0.05), and 1.5-fold (p < 0.05) reduction in neutralization sensitivity, respectively (Fig. 3D and Fig. S3D), indicating that R346T and D1199N mutations play less important roles compared to F486S in conferring the neutralization resistance. We noted that R346T/F486S double mutations reduced neutralization sensitivity to BA.1 wave patient sera 4.0-fold (p < 0.01) (Fig. 3E and Fig.S3E), although the F486S mutation alone only conferred a 1.5-fold (p = 0.3698) reduction in neutralization sensitivity for this cohort (Fig. 3E). Importantly, introduction of the R346T/F486S mutations into BA.2.75 reduced neutralization sensitivity to BA.4/5-infected patient sera 2.8-fold (p < 0.01) (Fig. 3F and Fig. S3F). Overall, these results indicated that the F486S mutation in BA.2.75.2 is the major driver of enhanced nAb resistance, for both sera from HCWs and COVID-positive patients.
Structural modeling reveals mechanism of mutation-mediated antibody evasion
To further understand the functional impact of the mutations in these Omicron subvariants, we performed homology modeling-based structural analyses. Neutralizing antibodies targeting the SARS-CoV-2 S protein can largely be divided into four classes based on the region of the receptor binding domain (RBD) targeted by the antibodies39,40. Specifically, class I antibodies target RBDs in the “up” conformation, class II and class III antibodies target RBDs in either their “up” or “down” conformation, and class IV antibodies generally fail to block interactions with ACE2 40. Notably, R346 and K444 are located outside of SARS-CoV-2 receptor binding motif (RBM) and are within the epitope of class III neutralizing antibodies. Structural analysis indicated an interference of antibody recognition introduced by R346S (Fig. 4A) and K444T (Fig. 4B), where hydrogen bonds and salt-bridge can be abolished. In contrast, F486 is located within the RBM and is a key residue for binding to both the ACE2 receptor and to neutralizing antibodies; F486 interacts hydrophobically with M82 and Y83 on ACE2 and residues on antibody complementarity-determining regions region, whereas the F486S mutation negatively impacts the interaction with ACE2 (Fig. 4C), as well as the recognition by some monoclonal antibodies in the class I and II categories (Fig. 4D). Further structural analysis showed that residue D1199 is located in the heptad repeat 2 (HR2) region on a solvent-accessible surface close to the transmembrane domain or membrane (Fig. 4E). Electrostatic surface potential (Fig. 4E inset) of this region reveals a strong negative surface charge, which repulses the negatively charged membrane and could help keep the spike in an up-right position. However, the D1199N mutation in the S2 subunit would reduce the electrostatic repulsion, resulting in a more tilted spike and may impact receptor utilization. These analyses provided molecular mechanisms of mutation-mediated antibody evasion and alteration in receptor utilization by the BQ.1, BQ.1.1, BA.4.6, BF.7, and BA.2.75.2 subvariants.
Figure 4: Homology modeling-based structural analyses of mutations essential for nAb resistance and fusogenicity.
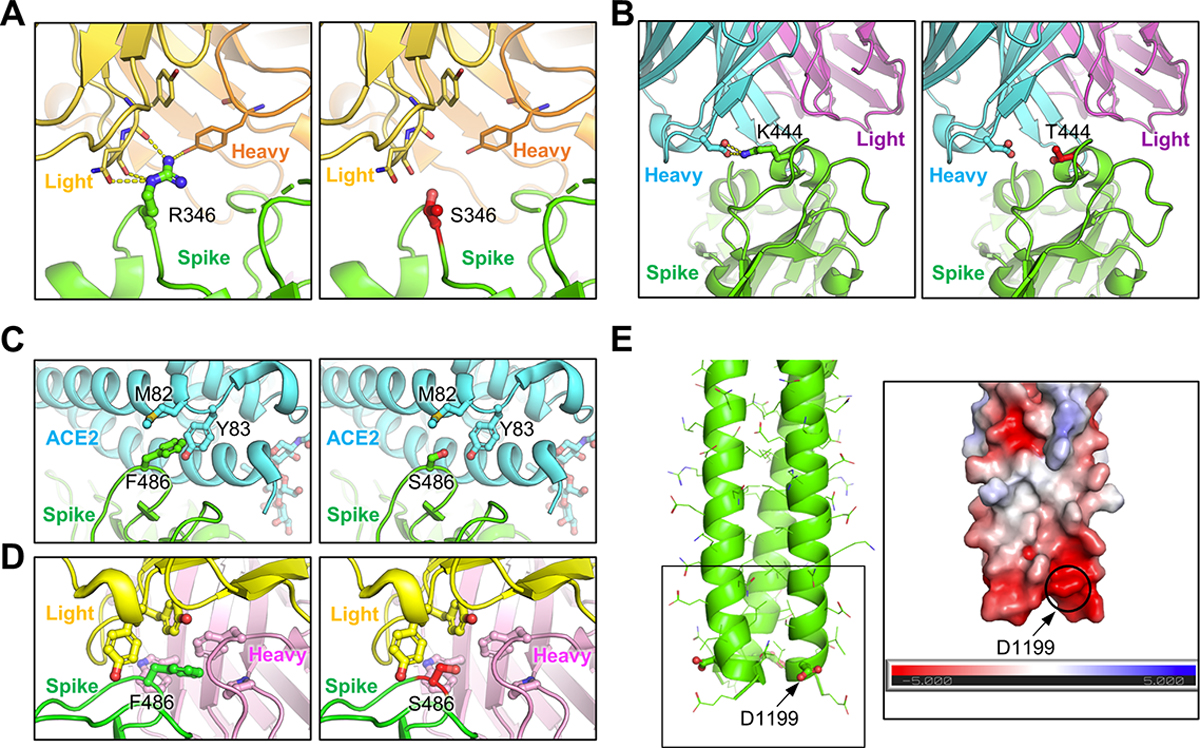
(A) and (B) Structures of spike-antibody binding interface shown as ribbons. Spike recognition of class III neutralizing antibodies C1365 (A) and SW186 (B) are interfered by R346S and K444T mutations, where multiple hydrogen bonds and salt-bridge (shown as yellow dot lines) are abolished. (C) Structure of spike-ACE2 binding interface shown as ribbon. F486 interacts hydrophobically with M82 and Y83 on ACE2, whereas F486S impedes this interaction. (D) Structure of class I antibody (AZD8895) recognition focusing on residue F486, with multiple antibody residues forming a surrounding hydrophobic cage. The F486S mutation abolishes this interaction. (E) Structural model of HR2 region of SARS-CoV2 S. Inset: Electrostatic surface potential of HR2 membrane proximal region. D1199 contributes to the overall negative charge of this region.
DISCUSSION
We examined the neutralization resistance of the emerging BA.4.6, BF.7, BQ.1, BQ.1.1, and BA.2.75.2 Omicron subvariants, as well as the impact on neutralization resistance of their lineage-defining mutations. All of these subvariants exhibit some degree of enhanced neutralization resistance over their parental BA.4/5 or BA.2.75.2 subvariants, with BQ.1, BQ.1.1, and BA.2.75.2 exhibiting the strongest resistance. Notably, this pattern is consistent for sera collected from HCWs following a homologous mRNA booster vaccination, from BA.1-wave hospitalized COVID-19 patients, as well as from BA.4/5-wave SARS-CoV-2 COVID-19 positive first responders and household contacts. Critically, we find that the neutralization resistance of BQ.1 and BQ.1.1 is driven largely by their N460K mutation, while the neutralization resistance of the BA.2.75.2 subvariant is determined by the F486S mutation. Our previous structural modeling analyses have shown that the N460K mutation present in BA.2.75 may potentiate ACE2 interactions3, thus introduction of the N460K mutation into the BA.4/5-derived subvariants may contribute to enhanced cell-cell fusion and neutralization resistance. Our new structural modeling and additional analyses in this work revealed crucial roles of residues R346, K444 and F486 in antibody recognition, as well as the potential mechanisms of immune evasion through R346T, K444T and F486S mutations present in Omicron subvariants. Notably, these mutations mediate resistance not only to 3-dose mRNA vaccine induced sera, but also BA.1- and BA.4/5-infection-induced sera. We also found that BA.4/5-wave patient sera exhibited weaker neutralization of BA.4/5 than of BA.2, which could be related to prior exposure to SARS-CoV-2 variant antigen biasing patient neutralizing antibody response to BA.4/5 infection. In addition, conformational alterations to the S protein have been demonstrated to alter neutralization sensitivity41–43.
It is well established that the Omicron subvariants exhibit diminished TMPRSS2-mediated plasma membrane entry resulting in reduced infectivity in Calu-3 cells, while retaining cathepsin-mediated endosomal entry capacity allowing robust entry in HEK293T-ACE2 cells15,17,20. This altered preference of entry route has been shown to be associated with the reduced lung tropism and reduced pathogenicity of the Omicron subvariants15,17,18,20. Here we show that the emerging Omicron subvariants BA.4.6, BF.7, BQ.1, BQ.1.1, and BA.2.75.2 retain this diminished infectivity in Calu-3 cells, but noticeably, the F486S mutation resulted in further diminished infectivity in Calu-3 cells. If this reduced infectivity in Calu-3 cell remains correlated with reduced lung tropism, the F486S mutation would induce further reduced pathogenicity of SARS-CoV-2. However, we have previously found increased syncytia formation capacity in several Omicron subvariants including BA.4/5, BA.2.75, BQ.1, BQ.1.1, and BA.2.75.2 compared to their ancestral Omicron strains3,44. This may indicate a continuing shift towards more efficient TMPRSS2 utilization to allow for plasma membrane-mediated fusion. A thorough investigation of the entry route preferences and in vivo pathogenicity of emerging Omicron subvariants is warranted.
The perpetual emergence of SARS-CoV-2 variants with enhanced immune escape continues to threaten public health. Monitoring the immune escape of emerging variants will be critical to improving mRNA vaccine reformulation, assessing new broader coronavirus vaccine candidates, as well as directing ongoing public health measures. Further, emerging variants, especially those containing the N460K, F486S, R346T, and K444T mutations, must be monitored closely for enhanced neutralization resistance.
Limitations of this work include the lack of experiments using authentic viruses for the neutralization assay; however, prior reports from ours and other groups have confirmed that results of pseudotyped viruses can faithfully reflect the neutralization of infectious SARS-CoV-2 and variants38,45. Additionally, whether or not these subvariants, as well as associated mutants, would use entry pathways distinct from the prototype Omicron and their parental lineages37, is unknown, which will be determined in future studies. Lastly, the impact of Omicron-defining mutations on ACE2 binding, as well as spike conformational changes warrants further biochemical and structural characterization.
STAR METHODS
RESOURCE AVAILABILITY
Lead Contact
Further information and requests for resources and reagents should be directed to the lead contact, Dr. Shan-Lu Liu (liu.6244@osu.edu).
Materials Availability
Plasmids generated in this study are available upon request made to the lead contact.
Data and Code Availability
NT50 values and de-identified patient information will be deposited to the National Cancer Institute SeroNet Coordinating Center. Additionally, NT50 values and de-identified patient information reported in this paper will be shared by the lead contact upon request.
This paper does not report original code.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
EXPERIMENTAL MODEL AND SUBJECT DETAILS
Samples and Patient Information
Sera samples were collected from HCWs at the Ohio State University Wexner Medical Center in Columbus, Ohio with approval from an institutional review board46 (IRB 2020H0228, IRB 2020H0527, and IRB 2017H0292). These HCWs samples were collected 2–13 weeks after vaccination with a third homologous dose of the monovalent Moderna mRNA-1273 (n = 3) or Pfizer BioNTech BNT162b2 (n = 12) vaccines. HCWs included 10 male and 5 female subjects with ages ranging from 26 to 61 (median 33).
Sera from BA.1-wave COVID-19 patients hospitalized in Columbus, Ohio were collected with approval from an IRB (IRB 2020H0527). The patient samples were collected 1–7 days after hospitalization with COVID-19. Hospitalizations occurred between the end of January and the end of February of 2022, a BA.1 dominant period in Columbus, Ohio. Patients included 12 male and 3 female patients with ages ranging from 29 to 78 (median 57). Patients included 6 unvaccinated patients, 5 patients vaccinated with 2 doses of Pfizer/BioNTech BNT162b2 (n = 2) or Moderna mRNA-1273 (n = 3), and 4 patients vaccinated and boosted with Pfizer/BioNTech BNT162b2.
Sera from BA.4/5-wave Columbus, Ohio first responders and household contacts who tested positive for SARS-CoV-2 infection were collected with IRB approval (IRB 2020H0527, 2020H0531, and 2020H0240). 11 patient nasal swab samples were sequenced to confirm infection with BA.4, BA.5, or derivative variants, with 4 patients infected with BA.4, 7 with BA.5, and 9 patients could not have their variant determined. Of those who could not have the specific variant identified, their samples were collected between late July and late September of 2022, a BA.4/5 dominant period based on CDC Ohio database. Except for one patient whose gender and age are unknown, patients included 4 male and 15 female with ages ranging from 27 to 58 (median 44). Patients included 17 unvaccinated, and 3 vaccinated and boosted with Pfizer/BioNTech BNT162b2 (n = 1) or Moderna mRNA-1273 (n = 2).
Cell Lines and Maintenance
Human female embryonic kidney cell lines HEK293T (ATCC CRL-11268, RRID: CVCL_1926) and HEK293T cells stably expressing human ACE2 (BEI NR-52511, RRID: CVCL_A7UK) were maintained in DMEM (Gibco, 11965–092) with 10% FBS (Sigma, F1051) and 1% penicillin-streptomycin (HyClone, SV30010) added. Human male adenocarcinoma lung epithelial cell line Calu-3 (RRID:CVCL_0609) were maintained in EMEM (ATCC, 30–2003) with 10% FBS and 1% penicillin-streptomycin added. All cells were passaged first by washing with Dulbecco’s phosphate buffered saline (Sigma, D5652–10X1L) then incubating in 0.05% Trypsin + 0.53 mM EDTA (Corning, 25-052-CI) until cells were completely detached. Cells were maintained at 37°C and 5.0% CO2 in 10 cm cell culture dishes (Greiner Bio-one, 664160).
METHOD DETAILS
Plasmids
The pNL4-3 inGluc lentiviral vector has been reported on in our previous publications38. Briefly, the vector is in the HIV-1 pNL4-3 backbone with a deletion of Env and an addition of a Gaussia luciferase reported gene that is expressed in target cells without premature expression in producer cells. The S variant constructs were cloned into the pcDNA3.1 vector by GenScript Biotech (Piscataway, NJ) using restriction enzyme cloning by Kpn I and BamH I; alternatively, they were produced by PCR mutagenesis. The constructs bear FLAG tags on the N- and C-terminal ends. All constructs were confirmed by sequencing.
Pseudotyped lentivirus production and infectivity
Pseudotyped lentiviral vectors were produced as previously described38. HEK293T cells were transfected with the pNL4-3-inGluc and S constructs in a 2:1 ratio using polyethyleneimine transfection (Transporter 5 Transfection Reagent, Polysciences) in order to generate viral particles. Virus was harvested 24, 48, and 72 hours post-transfection. Relative infectivity was determined by infection of HEK293T-ACE2 or Calu-3 cells and measurement of Gaussia luciferase activity 48 and 72 hours post-infection. Gaussia luciferase activity was measured by combining equal volumes of cell culture media and Gaussia luciferase substrate (0.1 M Tris pH 7.4, 0.3 M sodium ascorbate, 10 μM coelenterazine) with luminescence measured immediately by a BioTek Cytation5 plate reader.
Lentivirus neutralization assay
Neutralization assays with pseudotyped lentiviral vectors were performed as described previously38. HCW and COVID-19 patient samples were 4-fold serially diluted and equal amounts of SARS-CoV-2 pseudotyped virus were added to the diluted sera. Final dilutions were 1:80, 1:320, 1:1280, 1:5120, 1:20480, and no serum control. The virus and sera mixture was incubated for 1 hour at 37°C then added to HEK293T-ACE2 cells to allow for infection. Gaussia luciferase activity was measured at 48 and 72 hours post-infection by combining equal volumes of cell culture media and Gaussia luciferase substrate with luminescence measured immediately by a BioTek Cytation5 plate reader. The 50% neutralization titers (NT50) were determined by least-squares-fit, non-linear regression in GraphPad Prism 9 (San Diego, CA).
Structural modeling and analysis
Homology modeling of Omicron spike protein complexes with either ACE2 receptor or neutralizing antibodies was performed on SWISS-MODEL server with published X-ray crystallography and cryo-EM structures as templates (PDB IDs: 7K8Z, 8DT3, 7XB0, 7L7D, 2FXP). Molecular contacts of Omicron mutants were examined and illustrated with the programs PyMOL.
Quantification and statistical analysis
NT50 values were determined by least-squares-fit, non-linear-regression in GraphPad Prism 9 (San Diego, CA). NT50 values were log10 transformed for hypothesis testing to better approximate normality, and multiplicity was addressed by the use of Bonferroni corrections. The statistical analysis was performed using GraphPad Prism 9 and are referenced in the figure legends, including one-way ANOVA (Figs. 1B–G), one-way repeated measures ANOVA (Figs. 2A–C, Fig. 3A–F, and Fig. S2A–B), and two-way repeated measures ANOVA (Fig. S2A–B) for significance analysis between vaccination status.
Supplementary Material
Figure S1: Omicron subvariants display enhanced immune escape, related to Figure 2. (A-C) Neutralizing antibody tiers against Omicron subvariants and ancestral D614G are displayed as heat maps for sera from health care workers (HCWs, “H#”) (n = 15) who received a single homologous monovalent Moderna mRNA-1273 (n = 3) or Pfizer/BioNTech BNT162b2 (n = 12) mRNA booster vaccination (A); for sera from BA.1-wave hospitalized COVID-19 patients (“BA.1_P#”) (n = 15) (B), and for sera from BA.4/5-wave SARS-CoV-2 infected Columbus, Ohio first responders and household contacts (“BA.4/5_P#”) (n = 20) (C).
Figure S2: Omicron subvariants escape sera from vaccinated and unvaccinated COVID-19 patients, related to Figure 2.
(A) NT50 data from Fig. 2B is plotted by vaccination status with unvaccinated (n = 6), 2-dose (n = 5), and 3-dose (n = 4) individuals. (B) NT50 data from Fig. 1C is plotted by vaccination status with unvaccinated (n = 17) and 3-dose (n = 3) individuals. Geometric mean NT50 values are displayed at the top of plots; bars represent geometric mean ± 95% confidence interval and significance was determined by one-way repeated measures ANOVA or two-way repeated measures ANOVA with Bonferroni’s multiple testing correction. p-values are represented as *p<0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns, not significant.
Figure S3: BA.4/5- and BA.2.75-derived mutants display distinct immune escapes, related to Figure 3.
(A-F) Neutralizing antibody tiers against BA.4/5-derived (A, B and C) or BA.2.75-derived (D, E and F) mutants are displayed as heat maps for sera from health care workers (HCWs, “H#”) (n = 15) who received a single homologous monovalent Moderna mRNA-1273 (n = 3) or Pfizer/BioNTech BNT162b2 (n = 12) mRNA booster vaccination (A, D); for sera from BA.1-wave hospitalized COVID-19 patients (“BA.1_P#”) (n = 15) (B, E), and for sera from BA.4/5-wave SARS-CoV-2 infected Columbus, Ohio first responders and household contacts (“BA.4/5_P#”) (n = 20) (C and F).
KEY RESOURCES TABLE.
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
|
| ||
| Biological samples | ||
|
| ||
| 3-dose HCWs Sera | Evans et al.1, Qu et al. 4 | N/A |
| Omicron BA.1-wave Infected Patient Sera | Evans et al.1, Qu et al. 4 | N/A |
| Omicron BA.4/5-wave Infected First Responders and Household Contacts Sera | This paper | N/A |
|
| ||
| Chemicals, peptides, and recombinant proteins | ||
|
| ||
| Polyethylenimine (PEI) | Thermo Fisher Scientific | Cat# BMS1003 |
| Dulbecco’s Modified Eagles Medium (DMEM) | Sigma-Aldrich | Cat#: 11965-092 |
| Fetal Bovine Serum (FBS) | Thermo Fisher Scientific | Cat#: F1051 |
| 0.05% Trypsin + 0.53 mM EDTA | Corning | Cat# 25-052-CI |
| Penicillin-Streptomycin | HyClone | Cat#: SV30010 |
| Immobilon Crescendo Western HRP substrate | Millipore | Cat# WBLUR0500 |
| QIAprep Spin Miniprep Kit | QIAGEN | Cat# 27106 |
| Coelenterazine | GoldBio | Cat#: CZ2.5, CAS: 55779-48-1 |
|
| ||
| Deposited data | ||
|
| ||
| NT50 Values and De-identified patient data | SeroNet Coordinating Center, NCI, NIH | N/A |
|
| ||
| Experimental models: Cell lines | ||
|
| ||
| HEK293T | ATCC | Cat#: CRL-11268, RRID: CVCL_1926 |
| HEK293T-ACE2 | BEI Resources | Cat#: NR-52511, RRID: CVCL_A7UK |
|
| ||
| Calu-3 | ATCC | RRID: CVCL_0609 |
|
| ||
| Recombinant DNA | ||
|
| ||
| pNL4-3-inGluc | David Derse, NIH Maruzov et al. 47 |
N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_D614G | GenScript Biotech Zeng et al. 48 |
N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.2 | GenScript Biotech Evans et al.1 |
N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.4/5 | GenScript Biotech Qu et al. 4 |
N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.4.6 | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BF.7 | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BQ.1 | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BQ.1.1 | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.2.75 | GenScript Biotech Qu et al. 3 |
N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.2.75.2 | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.4/5_K444T | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.4/5_N460K | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.4/5_N658S | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.4/5_R346T_K444T | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.4/5_R346T_N460K | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.2.75_R346T | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.2.75_F486S | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.2.75_D1199N | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.2.75_R346T_F486S | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.2.75_R346T_D1199N | This paper | N/A |
| pcDNA3.1-SARS-CoV-2-Flag-S-Flag_BA.2.75_F486S_D1199N | This paper | N/A |
|
| ||
| Software and algorithms | ||
|
| ||
| GraphPad Prism | GraphPad Version 9.0.0 | www.graphpad.com |
| SWISS-MODEL | Guex et al. 49 | https://swissmodel.expasy.org/ |
| PyMOL | Warren DeLano and Sarin Bromberg | https://pymol.org/ |
|
| ||
| Other | ||
|
| ||
| Cytation 5 Imaging Reader | BioTek | N/A |
Highlights.
Enhanced neutralization resistance of BQ.1 and BQ.1.1 is driven by N460K and K444T
Enhanced neutralization resistance of BA.2.75.2 is driven by F486S
R346T and K444T contribute to evasion of class III antibody recognition
Modeling reveals F486S reduces binding for both ACE2 and class I and II antibodies
ACKNOWLEDGEMENTS
We thank the NIH AIDS Reagent Program and BEI Resources for providing important reagents for this work. We also thank the Clinical Research Center/Center for Clinical Research Management of The Ohio State University Wexner Medical Center and The Ohio State University College of Medicine in Columbus, Ohio, specifically Francesca Madiai, Dina McGowan, Breona Edwards, Evan Long, and Trina Wemlinger, for logistics, collection and processing of samples. In addition, we thank Sarah Karow, Madison So, Preston So, Daniela Farkas, and Finny Johns in the clinical trials team of The Ohio State University for sample collection and other supports. This work was supported by a fund provided by an anonymous private donor to OSU. S.-L.L., F.S., D. J., G.L., A.P., R.J.G., L.J.S. and E.M.O. were supported by the National Cancer Institute of the NIH under award no. U54CA260582. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. J.P.E. was supported by Glenn Barber Fellowship from the Ohio State University College of Veterinary Medicine. R.J.G. was additionally supported by the Robert J. Anthony Fund for Cardiovascular Research and the JB Cardiovascular Research Fund, and L.J.S. was partially supported by NIH R01 HD095881. K.X. was supported by the Ohio State University James Cancer Center and a Path to K award from the Ohio State University Office of Health Sciences and the Center for Clinical & Translational Science. The content is solely the responsibility of the authors and does not necessarily represent the official views of the university, or the Center for Clinical & Translational Science.
Footnotes
DECLARATION OF INTERESTS
The authors declare no competing interests.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Evans JP, Zeng C, Qu P, Faraone J, Zheng YM, Carlin C, Bednash JS, Zhou T, Lozanski G, Mallampalli R, et al. (2022). Neutralization of SARS-CoV-2 Omicron sub-lineages BA.1, BA.1.1, and BA.2. Cell Host Microbe 30, 1093–1102.e1093. 10.1016/j.chom.2022.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kurhade C, Zou J, Xia H, Cai H, Yang Q, Cutler M, Cooper D, Muik A, Jansen KU, Xie X, et al. (2022). Neutralization of Omicron BA.1, BA.2, and BA.3 SARS-CoV-2 by 3 doses of BNT162b2 vaccine. Nat Commun 13, 3602. 10.1038/s41467-022-30681-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Qu P, Evans JP, Zheng YM, Carlin C, Saif LJ, Oltz EM, Xu K, Gumina RJ, and Liu SL (2022). Evasion of neutralizing antibody responses by the SARS-CoV-2 BA.2.75 variant. Cell Host Microbe. 10.1016/j.chom.2022.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Qu P, Faraone J, Evans JP, Zou X, Zheng YM, Carlin C, Bednash JS, Lozanski G, Mallampalli RK, Saif LJ, et al. (2022). Neutralization of the SARS-CoV-2 Omicron BA.4/5 and BA.2.12.1 Subvariants. The New England Journal of Medicine 386, 2526–2528. 10.1056/NEJMc2206725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Xia S, Wang L, Zhu Y, Lu L, and Jiang S (2022). Origin, virological features, immune evasion and intervention of SARS-CoV-2 Omicron sublineages. Signal Transduct Target Ther 7, 241. 10.1038/s41392-022-01105-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Yamasoba D, Kimura I, Nasser H, Morioka Y, Nao N, Ito J, Uriu K, Tsuda M, Zahradnik J, Shirakawa K, et al. (2022). Virological characteristics of the SARS-CoV-2 Omicron BA.2 spike. Cell 185, 2103–2115.e2119. 10.1016/j.cell.2022.04.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yu J, Collier AY, Rowe M, Mardas F, Ventura JD, Wan H, Miller J, Powers O, Chung B, Siamatu M, et al. (2022). Neutralization of the SARS-CoV-2 Omicron BA.1 and BA.2 Variants. The New England Journal of Medicine 386, 1579–1580. 10.1056/NEJMc2201849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Abu-Raddad LJ, Chemaitelly H, Ayoub HH, AlMukdad S, Yassine HM, Al-Khatib HA, Smatti MK, Tang P, Hasan MR, Coyle P, et al. (2022). Effect of mRNA Vaccine Boosters against SARS-CoV-2 Omicron Infection in Qatar. The New England Journal of Medicine 386, 1804–1816. 10.1056/NEJMoa2200797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gruell H, Vanshylla K, Tober-Lau P, Hillus D, Schommers P, Lehmann C, Kurth F, Sander LE, and Klein F (2022). mRNA booster immunization elicits potent neutralizing serum activity against the SARS-CoV-2 Omicron variant. Nat Med 28, 477–480. 10.1038/s41591-021-01676-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cao Y, Wang J, Jian F, Xiao T, Song W, Yisimayi A, Huang W, Li Q, Wang P, An R, et al. (2022). Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature 602, 657–663. 10.1038/s41586-021-04385-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Xia H, Zou J, Kurhade C, Cai H, Yang Q, Cutler M, Cooper D, Muik A, Jansen KU, Xie X, et al. (2022). Neutralization and durability of 2 or 3 doses of the BNT162b2 vaccine against Omicron SARS-CoV-2. Cell Host & Microbe 30, 485–488.e483. 10.1016/j.chom.2022.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zou J, Xia H, Xie X, Kurhade C, Machado RRG, Weaver SC, Ren P, and Shi PY (2022). Neutralization against Omicron SARS-CoV-2 from previous non-Omicron infection. Nat Commun 13, 852. 10.1038/s41467-022-28544-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Liu L, Iketani S, Guo Y, Chan JF, Wang M, Liu L, Luo Y, Chu H, Huang Y, Nair MS, et al. (2022). Striking antibody evasion manifested by the Omicron variant of SARS-CoV-2. Nature 602, 676–681. 10.1038/s41586-021-04388-0. [DOI] [PubMed] [Google Scholar]
- 14.Cerutti G, Guo Y, Liu L, Liu L, Zhang Z, Luo Y, Huang Y, Wang HH, Ho DD, Sheng Z, and Shapiro L (2022). Cryo-EM structure of the SARS-CoV-2 Omicron spike. Cell Rep 38, 110428. 10.1016/j.celrep.2022.110428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shuai H, Chan JF, Hu B, Chai Y, Yuen TT, Yin F, Huang X, Yoon C, Hu JC, Liu H, et al. (2022). Attenuated replication and pathogenicity of SARS-CoV-2 B.1.1.529 Omicron. Nature 603, 693–699. 10.1038/s41586-022-04442-5. [DOI] [PubMed] [Google Scholar]
- 16.Wang Q, Anang S, Iketani S, Guo Y, Liu L, Katsamba PS, Shapiro L, Ho DD, and Sodroski JG (2022). Functional properties of the spike glycoprotein of the emerging SARS-CoV-2 variant B.1.1.529. Cell Rep 39, 110924. 10.1016/j.celrep.2022.110924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Meng B, Abdullahi A, Ferreira I, Goonawardane N, Saito A, Kimura I, Yamasoba D, Gerber PP, Fatihi S, Rathore S, et al. (2022). Altered TMPRSS2 usage by SARS-CoV-2 Omicron impacts infectivity and fusogenicity. Nature 603, 706–714. 10.1038/s41586-022-04474-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Barut GT, Halwe NJ, Taddeo A, Kelly JN, Schön J, Ebert N, Ulrich L, Devisme C, Steiner S, Trüeb BS, et al. (2022). The spike gene is a major determinant for the SARS-CoV-2 Omicron-BA.1 phenotype. Nat Commun 13, 5929. 10.1038/s41467-022-33632-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cui Z, Liu P, Wang N, Wang L, Fan K, Zhu Q, Wang K, Chen R, Feng R, Jia Z, et al. (2022). Structural and functional characterizations of infectivity and immune evasion of SARS-CoV-2 Omicron. Cell 185, 860–871.e813. 10.1016/j.cell.2022.01.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Suzuki R, Yamasoba D, Kimura I, Wang L, Kishimoto M, Ito J, Morioka Y, Nao N, Nasser H, Uriu K, et al. (2022). Attenuated fusogenicity and pathogenicity of SARS-CoV-2 Omicron variant. Nature 603, 700–705. 10.1038/s41586-022-04462-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Su W, Choy KT, Gu H, Sia SF, Cheng KM, Nizami SIN, Krishnan P, Ng YM, Chang LDJ, Liu Y, et al. (2022). Omicron BA.1 and BA.2 sub-lineages show reduced pathogenicity and transmission potential than the early SARS-CoV-2 D614G variant in Syrian hamsters. J Infect Dis. 10.1093/infdis/jiac276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.McMahan K, Giffin V, Tostanoski LH, Chung B, Siamatu M, Suthar MS, Halfmann P, Kawaoka Y, Piedra-Mora C, Jain N, et al. (2022). Reduced pathogenicity of the SARS-CoV-2 omicron variant in hamsters. Med (New York, N.Y.) 3, 262–268.e264. 10.1016/j.medj.2022.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lyngse FP, Kirkeby CT, Denwood M, Christiansen LE, Mølbak K, Møller CH, Skov RL, Krause TG, Rasmussen M, Sieber RN, et al. (2022). Transmission of sars-cov-2 omicron voc subvariants BA.1 and BA.2: Evidence from danish households. MedRxiv. [Google Scholar]
- 24.Stegger M, Edslev SM, Sieber RN, Ingham AC, Ng KL, Tang M-HE, Alexandersen S, Fonager J, Legarth R, Utko M, et al. (2022). Occurrence and significance of Omicron BA. 1 infection followed by BA. 2 reinfection. medRxiv. [Google Scholar]
- 25.Iketani S, Liu L, Guo Y, Liu L, Chan JF, Huang Y, Wang M, Luo Y, Yu J, Chu H, et al. (2022). Antibody evasion properties of SARS-CoV-2 Omicron sublineages. Nature 604, 553–556. 10.1038/s41586-022-04594-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Khan K, Karim F, Ganga Y, Bernstein M, Jule Z, Reedoy K, Cele S, Lustig G, Amoako D, Wolter N, et al. (2022). Omicron BA.4/BA.5 escape neutralizing immunity elicited by BA.1 infection. Nat Commun 13, 4686. 10.1038/s41467-022-32396-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tuekprakhon A, Nutalai R, Dijokaite-Guraliuc A, Zhou D, Ginn HM, Selvaraj M, Liu C, Mentzer AJ, Supasa P, Duyvesteyn HME, et al. (2022). Antibody escape of SARS-CoV-2 Omicron BA.4 and BA.5 from vaccine and BA.1 serum. Cell 185, 2422–2433.e2413. 10.1016/j.cell.2022.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hachmann NP, Miller J, Collier AY, Ventura JD, Yu J, Rowe M, Bondzie EA, Powers O, Surve N, Hall K, and Barouch DH (2022). Neutralization Escape by SARS-CoV-2 Omicron Subvariants BA.2.12.1, BA.4, and BA.5. The New England journal of medicine 387, 86–88. 10.1056/NEJMc2206576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Cao Y, Yisimayi A, Jian F, Song W, Xiao T, Wang L, Du S, Wang J, Li Q, Chen X, et al. (2022). BA.2.12.1, BA.4 and BA.5 escape antibodies elicited by Omicron infection. Nature 608, 593–602. 10.1038/s41586-022-04980-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kimura I, Yamasoba D, Tamura T, Nao N, Suzuki T, Oda Y, Mitoma S, Ito J, Nasser H, Zahradnik J, et al. (2022). Virological characteristics of the SARS-CoV-2 Omicron BA.2 subvariants, including BA.4 and BA.5. Cell 185, 3992–4007.e3916. 10.1016/j.cell.2022.09.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wang Q, Guo Y, Iketani S, Nair MS, Li Z, Mohri H, Wang M, Yu J, Bowen AD, Chang JY, et al. (2022). Antibody evasion by SARS-CoV-2 Omicron subvariants BA.2.12.1, BA.4 and BA.5. Nature 608, 603–608. 10.1038/s41586-022-05053-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Centers for Disease Control and Prevention (2022). COVID Data Tracker. Atlanta, GA: US Department of Health and Human Services, CDC; 2022, October 16. https://covid.cdc.gov/covid-data-tracker.
- 33.Cao Y, Song W, Wang L, Liu P, Yue C, Jian F, Yu Y, Yisimayi A, Wang P, Wang Y, et al. (2022). Characterization of the enhanced infectivity and antibody evasion of Omicron BA. 2.75. Cell Host Microbe. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wang Q, Iketani S, Li Z, Guo Y, Yeh AY, Liu M, Yu J, Sheng Z, Huang Y, Liu L, and Ho DD (2022). Antigenic characterization of the SARS-CoV-2 Omicron subvariant BA.2.75. Cell Host Microbe. 10.1016/j.chom.2022.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Saito A, Tamura T, Zahradnik J, Deguchi S, Tabata K, Anraku Y, Kimura I, Ito J, Yamasoba D, Nasser H, et al. Virological characteristics of the SARS-CoV-2 Omicron BA. 2.75 variant. Cell Host Microbe. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Iacobucci G (2022). Covid-19: Hospital admissions rise in England as some trusts reinstate mask requirements. Bmj 379, o2440. 10.1136/bmj.o2440. [DOI] [PubMed] [Google Scholar]
- 37.Cao Y, Jian F, Wang J, Yu Y, Song W, Yisimayi A, An R, Zhang N, Wang Y, Wang P, et al. (2022). Imprinted SARS-CoV-2 humoral immunity induces converging Omicron RBD evolution. bioRxiv. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zeng C, Evans JP, Pearson R, Qu P, Zheng YM, Robinson RT, Hall-Stoodley L, Yount J, Pannu S, Mallampalli RK, et al. (2020). Neutralizing antibody against SARS-CoV-2 spike in COVID-19 patients, health care workers, and convalescent plasma donors. JCI Insight 5. 10.1172/jci.insight.143213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Greaney AJ, Starr TN, Barnes CO, Weisblum Y, Schmidt F, Caskey M, Gaebler C, Cho A, Agudelo M, Finkin S, et al. (2021). Mapping mutations to the SARS-CoV-2 RBD that escape binding by different classes of antibodies. Nat Commun 12, 4196. 10.1038/s41467-021-24435-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Barnes CO, Jette CA, Abernathy ME, Dam KA, Esswein SR, Gristick HB, Malyutin AG, Sharaf NG, Huey-Tubman KE, Lee YE, et al. (2020). SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies. Nature 588, 682–687. 10.1038/s41586-020-2852-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Weissman D, Alameh MG, de Silva T, Collini P, Hornsby H, Brown R, LaBranche CC, Edwards RJ, Sutherland L, Santra S, et al. (2021). D614G Spike Mutation Increases SARS CoV-2 Susceptibility to Neutralization. Cell Host Microbe 29, 23–31.e24. 10.1016/j.chom.2020.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Plante JA, Liu Y, Liu J, Xia H, Johnson BA, Lokugamage KG, Zhang X, Muruato AE, Zou J, Fontes-Garfias CR, et al. (2021). Spike mutation D614G alters SARS-CoV-2 fitness. Nature 592, 116–121. 10.1038/s41586-020-2895-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Yurkovetskiy L, Wang X, Pascal KE, Tomkins-Tinch C, Nyalile TP, Wang Y, Baum A, Diehl WE, Dauphin A, Carbone C, et al. (2020). Structural and Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant. Cell 183, 739–751.e738. 10.1016/j.cell.2020.09.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Qu P, Evans JP, Faraone J, Zheng Y-M, Carlin C, Anghelina M, Stevens P, Fernandez S, Jones D, Lozanski G, et al. (2022). Distinct Neutralizing Antibody Escape of SARS-CoV-2 Omicron Subvariants BQ.1, BQ.1.1, BA.4.6, BF.7 and BA.2.75.2. bioRxiv, 2022.2010.2019.512891. 10.1101/2022.10.19.512891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Xiong HL, Wu YT, Cao JL, Yang R, Liu YX, Ma J, Qiao XY, Yao XY, Zhang BH, Zhang YL, et al. (2020). Robust neutralization assay based on SARS-CoV-2 S-protein-bearing vesicular stomatitis virus (VSV) pseudovirus and ACE2-overexpressing BHK21 cells. Emerg Microbes Infect 9, 2105–2113. 10.1080/22221751.2020.1815589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gordon DE, Jang GM, Bouhaddou M, Xu J, Obernier K, White KM, O’Meara MJ, Rezelj VV, Guo JZ, Swaney DL, et al. (2020). A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 583, 459–468. 10.1038/s41586-020-2286-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Mazurov D, Ilinskaya A, Heidecker G, Lloyd P, and Derse D (2010). Quantitative comparison of HTLV-1 and HIV-1 cell-to-cell infection with new replication dependent vectors. PLoS Pathog 6, e1000788. 10.1371/journal.ppat.1000788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zeng C, Evans JP, Faraone JN, Qu P, Zheng YM, Saif L, Oltz EM, Lozanski G, Gumina RJ, and Liu SL (2021). Neutralization of SARS-CoV-2 Variants of Concern Harboring Q677H. mBio 12, e0251021. 10.1128/mBio.02510-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Guex N, Peitsch MC, and Schwede T (2009). Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: a historical perspective. Electrophoresis 30 Suppl 1, S162–173. 10.1002/elps.200900140. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1: Omicron subvariants display enhanced immune escape, related to Figure 2. (A-C) Neutralizing antibody tiers against Omicron subvariants and ancestral D614G are displayed as heat maps for sera from health care workers (HCWs, “H#”) (n = 15) who received a single homologous monovalent Moderna mRNA-1273 (n = 3) or Pfizer/BioNTech BNT162b2 (n = 12) mRNA booster vaccination (A); for sera from BA.1-wave hospitalized COVID-19 patients (“BA.1_P#”) (n = 15) (B), and for sera from BA.4/5-wave SARS-CoV-2 infected Columbus, Ohio first responders and household contacts (“BA.4/5_P#”) (n = 20) (C).
Figure S2: Omicron subvariants escape sera from vaccinated and unvaccinated COVID-19 patients, related to Figure 2.
(A) NT50 data from Fig. 2B is plotted by vaccination status with unvaccinated (n = 6), 2-dose (n = 5), and 3-dose (n = 4) individuals. (B) NT50 data from Fig. 1C is plotted by vaccination status with unvaccinated (n = 17) and 3-dose (n = 3) individuals. Geometric mean NT50 values are displayed at the top of plots; bars represent geometric mean ± 95% confidence interval and significance was determined by one-way repeated measures ANOVA or two-way repeated measures ANOVA with Bonferroni’s multiple testing correction. p-values are represented as *p<0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns, not significant.
Figure S3: BA.4/5- and BA.2.75-derived mutants display distinct immune escapes, related to Figure 3.
(A-F) Neutralizing antibody tiers against BA.4/5-derived (A, B and C) or BA.2.75-derived (D, E and F) mutants are displayed as heat maps for sera from health care workers (HCWs, “H#”) (n = 15) who received a single homologous monovalent Moderna mRNA-1273 (n = 3) or Pfizer/BioNTech BNT162b2 (n = 12) mRNA booster vaccination (A, D); for sera from BA.1-wave hospitalized COVID-19 patients (“BA.1_P#”) (n = 15) (B, E), and for sera from BA.4/5-wave SARS-CoV-2 infected Columbus, Ohio first responders and household contacts (“BA.4/5_P#”) (n = 20) (C and F).
Data Availability Statement
NT50 values and de-identified patient information will be deposited to the National Cancer Institute SeroNet Coordinating Center. Additionally, NT50 values and de-identified patient information reported in this paper will be shared by the lead contact upon request.
This paper does not report original code.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.