

Abstract
Dietary saturated fats have recently been appreciated for their ability to modify innate immune cell function, including monocytes, macrophages, and neutrophils. Many dietary saturated fatty acids (SFAs) embark on a unique pathway through the lymphatics following digestion, and this makes them intriguing candidates for inflammatory regulation during homeostasis and disease. Specifically, palmitic acid (PA), and diets enriched in PA, have recently been implicated in driving innate immune memory in mice. PA has been shown to induce long-lasting hyperinflammatory capacity against secondary microbial stimuli in vitro and in vivo, and PA-enriched diets alter the developmental trajectory of stem cell progenitors in the bone marrow. Perhaps the most relevant finding is the ability of exogenous PA to enhance clearance of fungal and bacterial burdens in mice, yet the same PA treatment enhances endotoxemia severity and mortality. Westernized countries are becoming increasingly dependent on SFA-enriched diets, and a deeper understanding of SFA regulation of innate immune memory is imperative in this pandemic era.
Keywords: Saturated fatty acid, monocytes, macrophages, palmitic acid, innate immune memory, Western Diet, Ketogenic Diet, trained immunity, priming, metabolism, epigenetics, inflammation, chylomicron, ceramide, toll-like receptor, CD36, hematopoietic stem cell, oleic acid
1. Introduction
The innate immune system acts as an efficient non-species-specific shield against infection that thwarts a diverse array of pathogens, and has recently been shown to be highly influenced by the nutritional milieu of the host. Specifically, in humans, the innate immune response has been adapting to constantly changing dietary patterns and, more recently, a variety of diets with enriched saturated fatty acids (SFAs). Decades of research have been dedicated to understanding how SFAs modulate innate immune cell function and inflammatory capacity (1, 2); however, it has only recently been described that enriched dietary SFAs can induce innate immune memory.
Innate immune memory is characterized by epigenetic and metabolic changes within macrophages and monocytes induced by a primary inflammatory stimulus that leads to an enhanced (trained immunity or priming) or decreased (tolerance) response to a secondary inflammatory stimulus (3). Trained immunity and priming are primitive adaptations of innate host defense that results from exposure to a primary inflammatory stimulus, and leads to a faster and greater inflammatory response to a secondary homologous or heterologous challenge (4). Hallmark features of trained immunity specifically include metabolic alterations that induce long-lasting epigenetic changes within innate immune cells, remodeling of the hematopoietic stem cell (HSC) compartment that allows for a sustained augmented response from developing myeloid cells, an amplified inflammatory response to pathogenic infection, and transmission of epigenetic memory across generations (5–9). In contrast, the effects of priming do not involve long-term epigenetic modifications or HSC remodeling; however, priming does allow a cell to respond more strongly to secondary inflammatory stimuli, but only if the secondary stimulus takes place before the cell returns to basal inflammation and homeostasis (3). The most studied inducers of trained immunity are the Bacillus Calmette-Guerin (BCG) vaccine, and the fungal antigen, β-glucan. BCG and β-glucan induce cross-protection against heterologous pathogens in humans and in mice respectively, including cross-protection against Sars-CoV-2 (4, 10). Trained immunity is also known for its dual nature, and can be beneficial or detrimental to the host depending on the disease context and inflammatory status.
There is little known about the impact of dietary SFAs on innate immune memory, however, SFA-enriched diets have been shown to induce long-lasting impacts on innate immune inflammation and microbial infection in mice, via remodeling of the HSC compartment in the bone marrow (11, 12). Specifically, enriched dietary palmitic acid (PA; C16:0), the most prevalent SFA found in the diet and circulating in human blood, has been shown to induce trained immunity within myeloid cells, and plays an important role in both homeostasis and infection in vivo (1, 12, 16). PA is known to enhance toll-like receptor (TLR)-dependent inflammation by inducing ceramide metabolism, and sensitizes innate immune cells to subsequent TLR stimulation (12–15). During endotoxemia [lipopolysaccharide (LPS)-induced acute systemic shock], mice pre-exposed to PA exhibit enhanced circulating inflammation, disease severity, and mortality compared to vehicle (Veh)-treated mice and showed significantly greater clearance of fungal and bacterial infections compared to Veh-treated mice (12, 16). The molecular details of SFA- and/or PA-induced trained immunity in vitro and in vivo have not been fully defined; however, in light of these exciting results, we will review here the specific relationship between PA and innate immune memory, and discuss the implications of our known data and the need for future investigations that will elucidate the intricate dynamics of this relationship.
2. Palmitic Acid mediates macrophage metabolism and function
Trained immunity in macrophages is accompanied by direct crosstalk between metabolites from the Krebs cycle, and histone modifying enzymes necessary for enhanced transcription of pro-inflammatory cytokines via epigenetic mechanisms (17). It is well known that specific metabolic pathways mediate the transcription and release of pro- and anti-inflammatory cytokines by macrophages (18, 19). Glycolytic metabolism in the cytosol is upregulated in pro-inflammatory macrophage responses, and is accompanied by breaks in the Krebs Cycle that allow for accumulation of metabolites that also support the Warburg effect (i.e. aerobic glycolysis) (20–22). In contrast, oxidative phosphorylation in the mitochondria is upregulated, and the Krebs cycle remains intact, in order to support anti-inflammatory macrophage responses (18). The plasticity of macrophage inflammatory polarization is governed by tightly regulated metabolic pathways that can be disrupted by excessive exposure to dietary SFAs, including PA (12, 15, 23).
PA is the most common SFA found in the human body (20-30% total FAs) and is enriched in meat, dairy products (50-60% of total fats) and is nearly 30% of total fats in breast milk (2, 24). The Western Diet (WD) contains high levels of FAs, specifically PA, and sucrose; the Ketogenic Diet (KD) is exclusively enriched in FAs, and depending on the dietary structure, it may contain excess PA content. In addition to dietary sources, PA can be synthesized endogenously throughout the mammalian body from other FAs, carbohydrates, and amino acids (2). PA is known as an immunomodulatory molecule, and it has the capacity to regulate inflammatory processes of innate immune cells, including monocytes, macrophages, and neutrophils (12, 15, 25).
Historically, it was believed that PA was a ligand for TLR4 and induced TLR4-dependent inflammatory cytokine production (26). However, an elegant study has recently shown that PA is not a ligand for TLR4, but enhances activation of multiple TLR signaling pathways and subsequent NF-κB-dependent transcription of inflammatory cytokines through c-Jun N-terminal kinase (JNK) activation that is dependent on mitochondrial metabolic regulation via mammalian target of rapamycin (mTOR) (23). These studies were conclusive in determining PA is not a TLR4 ligand, but much still remains to understand how PA is metabolically enhancing TLR-dependent inflammation in macrophages.
Importantly, when there is an excess of PA in the diet, it is reflected in an increase of free PA systemically (1, 27). When macrophages and monocytes are exposed to excess free PA, it is taken up through the membrane scavenger receptor, CD36. CD36-dependent intake of PA can lead to lipid accumulation and modulation of signaling through metabolic dysfunction, including suppressing AMPK activation (28, 29). It has been found that genetic loss of Cd36 renders murine macrophages insensitive to some TLR2 ligands and induces hyper-susceptibility of mice to Staphylococcus aureus infection (30). Considering this, inhibition of free PA uptake by genetic deletion of Cd36 leading to a dampened inflammatory response may contribute to enhanced susceptibility to infection and decreased TLR2-mediated inflammation. Currently, it is unknown if CD36 plays a role in PA-induced inflammation, but it is clearly required for PA-dependent inflammatory phenotypes.
After CD36-dependent uptake of PA by the macrophage, PA is converted into phospholipids, diacylglycerol (DAG), and ceramides reviewed here: (1); however, in the presence of excess PA, TAG synthesis is stalled at the DAG stage causing accumulation in the cell (1). Both DAG and ceramides have been shown to enhance TLR-mediated signaling cascades in macrophages and subsequent activation of NF-κB (1). Specifically, in the presence of excess PA, macrophages enhance expression of adipose fatty acid binding protein (A-FABP), which acts as a PA chaperone during uptake to enhance ceramide synthesis (31).
Ceramide is a bioactive sphingolipid with cell signaling capabilities, and there are three metabolic pathways that can lead to intracellular ceramide synthesis: the de novo pathway, sphingomyelin (SM) hydrolysis, and the endosomal salvage pathway (32). Stimulating macrophages with LPS induces ceramide synthesis via SM hydrolysis (33). In contrast, de novo ceramide synthesis in the presence of excess PA is important for increasing TLR4 activation and cytokine production in primary peritoneal macrophages, however, ceramide produced via SM hydrolysis was shown to regulate LPS-induced IL-6 secretion in PA-treated RAW macrophages. (13, 14). In addition, our lab recently showed, using primary bone marrow-derived macrophages (BMDMs), that de novo ceramide synthesis is required for PA-induced hyperinflammation in response to LPS; moreover, inhibiting de novo ceramide synthesis completely abolished the hyperinflammatory impact with respect to TNF, however only partially inhibited IL-6-and IL-1β-mediated hyperinflammation (12). Future studies will benefit from determining precisely which pathway(s) of ceramide synthesis may be responsible for driving innate immune cell reprogramming and which proteins and signaling pathways are targeted by ceramide, as any of these have the potential to be therapeutic targets for inflammatory diseases driven by ceramide.
Together, these data conclude PA is not a TLR ligand, but enters macrophages through CD36, and subsequently mediates metabolism to enhance TLR-dependent cytokine production, which may play a critical role in regulating host response to infection and inflammatory diseases. Importantly, the term “excess” dietary PA is subjective, and likely depends on disease context, and the ability of the host to regulate homeostasis between endogenous and exogenous PA levels; however, physiologically relevant serum PA concentrations have been mimicked in vitro and in vivo to show a significant impact on metabolic pathways that alter macrophage inflammation (12–14, 23, 34). In order to target PA metabolism of macrophages therapeutically, future studies should determine the specific threshold of intracellular PA levels that, when exceeded, can lead to metabolic and inflammatory dysregulation.
3. Exogenous palmitic acid and trained immunity
It has recently been proposed that exogenous PA is capable of altering the metabolic and epigenetic landscape of monocytes and macrophages in order to initiate a non-specific, hyperinflammatory memory response to secondary stimulation with a microbial ligand (12). Although the precise impact of PA-mediated metabolism on the monocyte/macrophage epigenome has yet to be described, numerous features of innate immune memory, specifically trained immunity, have been revealed by studies involving exogenous PA treatments in vitro and in vivo.
Many studies show the inflammatory impact of combined PA and TLR ligand treatment on monocytes and macrophages, however, few studies show the effect of pre-treating cells with PA followed by TLR agonist or microbial challenge (Table 1). Pre-treatment experiments allow to understand if the PA-dependent metabolic and epigenetic changes can alter short- or long-term inflammatory response to secondary stimulation with a microbial challenge (trained immunity). Pre-treatment can be a model for a host that is exposed to excess dietary PA and then challenged by an infection, and these studies can inform how PA may be involved in induction of innate immune memory.
Table 1.
The impact of PA on innate immune inflammatory responses and memory.
| Conditions | Responses | Biological effects | Models used | References |
|---|---|---|---|---|
| Simultaneous PA + LPS (in vitro) | Priming | Enhanced ceramide; enhanced TNF/IL-6 secretion; JNK dependent | Primary & immortalized mouse macrophages | Schilling et al., 2013; Jin et al., 2013; Nishiyama et al., 2018 |
| PA pre-treatment + LPS (in vitro) | Trained immunity | Enhanced TNF/IL-6/IL-1β secretion, all ceramide dependent; role of mitochondria and MAPK signaling | Primary mouse macrophages; immortalized human & mouse macrophages | Seufert et al., 2022; Fang et al., 2022; Caslin et al., 2023; Schwartz et al., 2010 |
| PA pre-treatment + LPS (in vivo) | Trained immunity | Enhanced circulating inflammation; decreased survival | Wild type female BALB/c mice | Seufert et al., 2022 |
| PA pre-treatment + infection (in vivo) | Trained immunity | Enhanced microbial clearance | Rag−/− mice + Candida albicans infection; wild type female BALB/c mice + Brucella abortus infection | Seufert et al., 2022; Reyes et al., 2021 |
It was recently shown that immortalized human and mouse macrophage cell lines that were pre-treated with exogenous PA exhibited enhanced LPS-induced IL-6 and TNF secretion, respectively (15). Schwartz et. al. showed in THP-1 monocytes, that the hyperinflammatory response to LPS following PA-pretreatment was dependent on ceramide-mediated activation of protein kinase C (PKC) and mitogen activated protein kinase (MAPK) signaling pathways (34). While the mechanism for augmented TNF secretion was not defined by Zhang et al., the hyperinflammatory effect of PA-pretreatment was associated with enhanced phosphorylation of p38 and JNK, and enhanced expression of carnitine palmitoyltransferase1A (CPT1A), an enzymatic shuttle on the outer membrane of mitochondria that facilitates uptake of activated fatty acids into the mitochondrial matrix for β-oxidation (35). These data suggest that mitochondrial metabolism plays an important role in the hyperinflammatory impact of PA-pretreatment and subsequent challenge with LPS in macrophages (15, 34). These studies, however, did not distinguish if PA pre-treatment was inducing priming or trained immunity, but they did suggest that PA can alter macrophages to respond more acutely to secondary LPS challenge.
Our team recently showed that PA-pretreatment of BMDMs subsequently challenged with LPS enhanced TNF, IL-6, and IL-1β release; this was dependent on ceramide synthesis, and reversible when BMDMs were pre-treated simultaneously with both PA and the monounsaturated FA that diverts ceramide synthesis in the presence of PA, Oleic Acid (OA) (12). Additionally, the synergistic effect of PA and LPS was shown in primary mouse peritoneal macrophages, however, only simultaneous treatment was used to show significantly enhanced de novo ceramide synthesis, and significantly enhanced TNF and IL-6 secretion (13). Thus, they could not conclude if this was trained immunity. This effect of combined PA and LPS treatment was recapitulated in RAW murine macrophages to show that ceramide mediates LPS-induced IL-6 secretion via JNK phosphorylation; interestingly, this process was regulated by fatty acid transporter 1 (FATP1), and the requirement of CD36 was not shown (36).
Though these studies have been immensely important in describing the effect of PA on inflammation, it is still unclear if PA is inducing priming or trained immunity induced in macrophages. Thus, future studies regarding the role of PA in innate immune memory should determine the following: (1) The time point of initial inflammatory release in primary stimulation of macrophages with PA, (2) a return to basal inflammation, and (3) hyperinflammation upon secondary stimulation. Together, these outcomes would bolster the hypothesis that PA induces macrophage trained immunity in vitro.
In vivo, exogenous free PA has been shown to play an important role during infection. Specifically, in Rag1−/− mice that lack adaptive immunity, an intraperitoneal (i.p.) injection of a PA solution 12 h prior to intravenous (i.v.) infection with Candida albicans lead to a significant decrease in kidney fungal burden, compared to infected mice only pretreated with a vehicle solution (12). This suggests that PA-induced memory mediates microbial clearance, and this is dependent on innate immune cells. Further, mice injected i.p. with a PA solution 12 h prior to LPS-induced endotoxemia show significantly enhanced Tnf and Il-6 expression in the blood within 5 h post-LPS (12). Importantly, blood draws were taken immediately prior to LPS injections to show that baseline inflammation was not upregulated in PA-treated mice, suggesting that LPS-induced cytokine expression in the blood was not a priming effect induced by PA, but rather a trained immunity phenomenon (12).
Further, in a study by Reyes et. al., defining the role of exogenous PA in Brucella abortus infection in mice, oral gavage of PA was shown to significantly reduce splenic bacterial burden when administered for 3 days prior to, and during 14 days of B. abortus infection following an i.p. challenge (16). This was accompanied by a suppression of serum IL-10, however the mechanism underlying enhanced bacterial clearance induced by PA remains unknown. It is not entirely clear whether these data represent PA-induced priming or trained immunity, because there was no resting period after 3 day PA exposure, or determination of basal inflammatory status prior to infection with B. abortus. The results are still compelling, and to our knowledge they are the first to show the impact of dietary PA on the clearance of pathogenic bacteria in vivo.
Lastly, as mentioned previously, canonical trained immunity induced by β-glucans or BCG vaccine leads to long-term metabolic and functional reprogramming of myeloid cells, and relies on epigenetic alterations for sustained inflammatory capacity. Remarkably, PA-induced memory was shown to elicit long-lasting immune reprogramming in vivo using an LPS-induced endotoxemia mouse model (12). Specifically, 9 daily i.p. injections of PA followed by a 7-day resting period and subsequent LPS challenge led to enhanced endotoxemia severity and mortality compared to mice injected with a vehicle (Veh) solution (12). PA-injected mice showed significantly increased hypothermia compared to Veh-injected mice, and while mortality was enhanced in PA-injected mice, this survival defect was not significant compared to mice injected with a Veh. Thus, PA exacerbates endotoxemia severity, but this is not sufficient to significantly decrease survival in an endotoxemia mouse model (12). These data suggest that PA regulates epigenetic modifications that persist beyond the time points of PA exposure, and these alterations adversely impact the ability of the host to regulate body temperature in response to LPS challenge. Thus, PA induces trained immunity in this context, and not priming. Additional experiments will be required to determine if PA depends solely on innate immune memory mechanisms to exert the long-term adverse effects described here.
Together, these studies build a compelling case that exogenous PA modulates microbial-induced inflammation and clearance in vitro and in vivo. Importantly, the effects of PA exposure in vivo are long-lasting, and PA-exposed mice do not exhibit heightened circulating inflammation prior to infection, indicating that PA is inducing trained immunity, and not priming. The intracellular mechanisms of PA-induced memory, and the subsequent augmenting effects on microbial ligand stimulation are not fully characterized. While intracellular ceramide has been shown to mediate PA-induced memory in macrophages, it is still unclear how the metabolism of PA may lead to epigenetic alterations that ultimately modify the expression of inflammatory response genes.
4. Enriched dietary palmitic acid and trained immunity
Thus far we have described experimental data involving exogenous PA and its effects on monocyte and macrophage metabolism, inflammation, and memory, in addition to in vivo models of PA injections (i.p.) and oral gavage. Next, we outline the impacts of enriched dietary PA consumption on the development of innate immune cells in the bone marrow, and how this may contribute to long-term innate immune memory, specifically trained immunity.
Recent studies from our group and others have begun to understand the effect of excess chronic dietary PA on systemic response to microbial challenge and macrophage function ex vivo. Christ et. al., found that atherosclerotic mice (Ldlr−/− C57BL/6) fed WD for 4 weeks resulted in a hyper-inflammatory response when challenged with LPS ex vivo (11), suggesting exposure to excess dietary PA may be a factor in influencing the inflammatory capacity of myeloid cell populations in vivo. More recently, we have reported WT mice fed WD and KD, both enriched in PA, exhibit increased systemic inflammation in response to endotoxemia, a single i.p. injection of LPS (12, 37). Importantly, we show that this enhanced systemic inflammation in response to LPS is independent of glycolytic shock and the diet-induced microbiome, further suggesting that enriched dietary PA is leading to an enhanced response to TLR4 agonist LPS (12). Together, these studies suggest a direct link between enriched dietary PA and trained immunity in vivo.
A hallmark of canonical trained immunity in vivo is the skewing of the HSC compartment towards increased myeloid cell production and enhanced inflammatory capacity (8, 11). It has been shown, 4 weeks of WD administration was sufficient to alter HSC populations within the bone marrow of atherosclerotic mice; and after reverting back to a standard chow (SC) for an additional 4 weeks, stem cell progenitors remained skewed toward developing monocytes with hyper-inflammatory potential (11). More recently, we published that WT Balb/C mice on WD for 2 weeks do not exhibit altered HSC populations (12). Importantly, these studies may disagree due to the use of different mouse models (Ldlr−/− C57BL/6 versus wildtype BALB/c) and the length of diet administration; more follow-up studies are required to understand this bifurcation. However, we have additionally shown that exposure to a PA-enriched KD skews the HSC compartment to develop significantly enhanced populations of long- and short-term HSCs, and multipotent progenitors (12). Importantly, no one has shown whether PA is responsible for the HSC skewing in either of these models.
As mentioned previously, trained immunity can last days to years after initial induction with a primary inflammatory stimulus. While the endurance of dietary PA-induced memory remains unknown, the impacts exhibit hallmark features of trained immunity, including (1) the induction of long-term hyperinflammatory capacity toward microbial stimuli, (2) the low basal inflammatory status shown in vitro and in vivo following PA treatments prior to secondary stimulation, (3) the enhanced clearance of microbial infection in vivo, and (4) the induction of HSC remodeling that mimics HSC remodeling in BCG-induced trained immunity (8).
5. PA from the mouth to the bone marrow
In order to determine the holistic impact of dietary PA on innate immune memory, we must also consider the pathway that PA follows after digestion, and the byproducts of PA metabolism that may impact innate immune cells enroute to various tissues for energy or storage purposes. PA and its metabolic byproducts encounter innate immune cells prior to being distributed to tissues, because many fats, depending on their size and diffusion capability, circulate in lipoprotein structures throughout the lymphatics even before entering the bloodstream (38). Dietary PA digestion, absorption, and mobilization to tissues is complex compared to that of carbohydrates and proteins, which are transported directly to the liver via the superior mesenteric and hepatic portal veins immediately following digestion, and absorption from the small intestine into the blood circulation (Figure 1) (39, 40).
Figure 1. Pathway of dietary fatty acids following digestion.
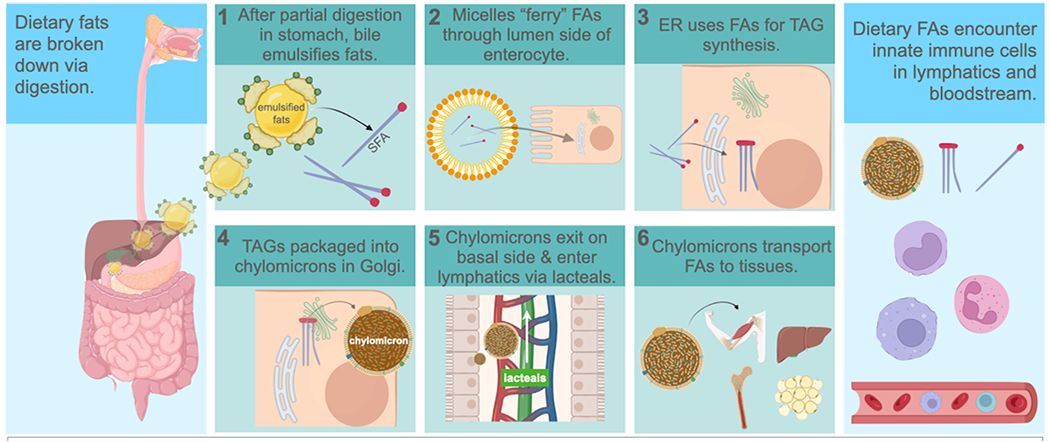
Following enzymatic digestion in the stomach, (1) bile salts from the gall bladder emulsify FAs and MAGs during entry into the duodenum. (2) FAs greater than 14-carbons in length are packaged into micelles that “ferry” them into enterocytes from the lumen. (3) FAs are resynthesizes into TAGs in the ER, following by (4) repackaging into chylomicrons within the Golgi. (5) Chylomicrons are carried in vesicles through the basal end of the enterocyte, entering lymphatic lacteals within the villi of the small intestine. (6) FAs are transported through the lymphatics within chylomicrons, and may encounter monocytes macrophages, and neutrophils. FA=fatty acid; MAG=monoacylglyceride; TAG=triacylglyceride; ER=endoplasmic reticulum. Created with BioRender.com
The basic structure of PA is a 16-carbon chain saturated with hydrogen, and thus too large to diffuse from the small intestine directly into capillaries leading to the blood (41). PA requires the emulsification action of bile, and “ferrying” within bile micelles towards the luminal epithelium of the small intestine (42) (Figure 1). Diffusion can then occur from the micelles that act as PA carriers, into the enterocytes of the small intestine, where they are broken down, reassembled into triglycerides (TAGs), and packaged into chylomicrons before being taken up by lymphatic lacteals (43). Chylomicrons are structures with a lipid membrane that contain hydrophobic lipids and proteins internally, and they enter into the lymphatics via lacteals, small ducts at the ends of lymphatic vessels within microvilli of the small intestine. After circulating through the lymph, PA-carrying chylomicrons enter the venous circulation via the thoracic duct, and lymphatic ducts near the subclavian veins. Then, the chylomicrons travel through the heart and are distributed to the remaining peripheral tissues including adipose tissue, skeletal muscle, bone marrow, and finally the liver, where they can be used for energy for various metabolic processes, incorporated into lipid membranes for cells and organelles, or stored as TAGs for later use (43). The CO2 waste from PA digestion is excreted via the respiratory system, and the term chylomicron “remnants” is often used to describe the final remaining TAGs that are taken to the liver after distribution throughout the rest of the body.
There are many opportunities for innate immune cells to encounter PA, which can be in the form of free fatty acids (FAs), TAGs, phosphatidylcholines (PCs), and sphingolipids (SGLs) while circulating in the lymphatics and blood to be distributed to skeletal muscle, adipose, bone marrow, liver, and spleen tissues for energy use or storage (44). Of particular importance is in the bone marrow, where HSCs may be reprogrammed by dietary PA, and then later differentiate into more specific progenitor cells with enhanced inflammatory capacity (11, 12, 45). It is important to consider the likelihood that reprogrammed HSCs may then seed tissues during stress or a localized infection, and exacerbate inflammation potentially leading to disruption of inflammatory homeostasis within a tissue, or whole organ.
This unique pathway that PA embarks on following digestion, and its ability to be transported within chylomicrons to the bone marrow, is especially important for the long-term nature of innate immune memory. Due to the short lifespan of circulating monocytes (1-3 days) prior to their differentiation into tissue-specific macrophages, it seems unlikely that these cell types would retain long-term, non-specific memory; however, PA-induced HSC reprogramming may have the potential to harbor epigenetically modified progenitor cells within the bone marrow that can be subsequently recruited to infected tissues, and respond with hyperinflammatory output to quickly clear infection. Moreover, in the context of dysregulated inflammation, such as in a septic response, trained HSCs would likely provide too much inflammation, and exacerbate disease via tissue damage. Considering that dietary SFAs can be transported to the bone marrow and alter HSC populations, the role of dietary PA in bone marrow remodeling is crucial to consider when determining therapeutic intervention strategies for infection and inflammatory diseases among populations that consume PA-enriched diets.
6. Plasticity of PA-dependent innate immune memory
There is evidence that PA-dependent innate immune memory induces metabolic and inflammatory changes that are reversible, and in contrast to PA exerting pro-inflammatory effects, certain unsaturated FAs (UFAs) are known to promote an anti-inflammatory outcome. It is now appreciated that PA accumulation can lead to pathophysiological changes and inflammation when there is an imbalance of dietary PA/Monounsaturated FAs (MUFAs). Oleic acid (OA) is the second most prevalent FA in the blood next to PA, and it has been shown to induce low levels of inflammation in macrophages in vitro at certain concentrations (46). However, OA is considered to be an anti-inflammatory MUFA, due to its ability to counteract or reverse the inflammatory impacts of SFAs in vitro (46). For example, in mouse peritoneal macrophages, OA reverses PA- and stearic acid-induced IL-1β secretion (47). In vivo, OA injections have been shown to mitigate the endotoxemia severity and mortality that is exacerbated in KD-fed mice (12). The polyunsaturated FA (PUFA), docosahexaenoic acid (DHA), has also been shown to counteract the inflammatory effects of PA and LA in RAW macrophages and BMDMs, displaying a specific target against de novo synthesis of ceramide (48–50). Thus, specific ratios of SFAs to MUFAs, or SFAs to PUFAs in the blood, bone marrow, and peripheral tissues such as liver, spleen, adipose, and skeletal muscle, will be important to consider when studying the regulatory impact of FAs on innate immune inflammation in vivo.
It is tempting to consider the possibility that a metabolic rheostat exists between trained immunity and inflammatory homeostasis, that can be controlled and balanced with specific ratios of dietary SFAs to UFAs; if inflammation is required to clear microbial burden, then SFAs may work to upregulate it, and if tissue homeostasis requires restoration after a hyperinflammatory event, then UFAs may serve to downregulate inflammation before it becomes damaging. If innate immune cells must adapt to a changing dietary environment, they may be required to increase or decrease their uptake of SFAs and UFAs, and perhaps this function is perturbed during excessive and prolonged exposure to SFAs.
7. Conclusion: Where to next?
The studies we present here provide compelling evidence that PA is an immunomodulating SFA that reprograms the innate immune cell response to secondary inflammatory stimuli in vitro and in vivo, i.e., PA induces innate immune memory. Although some outcomes suggest that PA induces priming, more recent investigations show the long-term impact of PA on innate immune inflammatory regulation. This long-term nature of PA reprogramming, in addition to the ability of PA to alter secondary inflammatory outcomes even after basal inflammation has been reached, strongly supports the hypothesis that PA induces trained immunity.
For future in vivo experiments, more long-term studies need to be done to collectively elucidate the following: (1) The physiologically relevant PA concentrations that induce the initial inflammatory response—are they relevant to consumers of SFA-enriched diets? (2) The initial inflammatory response to dietary PA—is it systemic or localized to specific tissues? (3) The minimum and maximum rest periods from PA exposure required to return to inflammatory homeostasis and still maintain innate immune memory reprogramming—how long does the memory last? (4) The specific metabolites, and histone modifications that are required for epigenetic reprogramming of innate immune cells—what is the mechanism behind the augmented inflammatory response to secondary stimulation? Addressing these questions will advance our knowledge of PA-mediated trained immunity and how it regulates innate immune homeostasis, inflammation, and the response to infection.
PA constitutes 20-25% of human breast milk, and if PA induces trained immunity, it is interesting to postulate that enhanced maternal dietary intake of PA may induce trained immunity in nursing neonates (51). Enhanced PA within maternal milk may influence the capacity of a neonate to induce inflammation and or enhance protection against microbial challenge. Interestingly, a study from Du et. al. shows that maternal WD consumption in mice causes the production of milk that contains excessive PA, ceramide accumulation, and inflammation in nursing neonates (52). Additionally, this ceramide accumulation and inflammation in nursing neonates was TLR4/2 dependent; however, they did not look into how enhanced PA milk effected neonatal 1) protection against microbes, 2) neonate HSC composition, or 3) neonatal macrophage response to LPS challenge.
While most immunological studies highlight the impact of PA on monocytes and macrophages, neutrophils may also play a role in SFA-induced inflammation and innate immune memory. For example, neutrophil recruitment and inflammation are enhanced by PA-treated bone marrow-derived macrophages (BMDMs) through a chemotactic response mechanism. Specifically, PA induces pannexin channel formation and nucleotide release, which then attracts neutrophils to the high-SFA region that macrophages were exposed to (53). Other dietary SFAs are also known to stimulate the release of neutrophil precursors from the bone marrow of mice into the bloodstream (54). Importantly, the direct impact that PA has on neutrophil metabolism and subsequent inflammation has not been fully studied. Future studies should focus on illustrating the impact that PA-enhanced neutrophil recruitment may have on inflammatory homeostasis, and the host response to microbial triggers.
The discovery that PA induces innate immune memory, and modulates inflammatory and infection outcomes in mice, is just the beginning of a much greater appreciation for the nuances of dietary-regulated immune function. A deeper understanding of the crosstalk between dietary SFAs and innate immune inflammation is imperative for understanding the long-term impact of diets enriched in SFAs on the host response to infection and inflammatory disease. This will contribute to effective prevention and treatment of diseases exacerbated by excessive SFA intake, advanced personalized nutrition, and a greater sense of agency over our immune health while living in societies heavily dependent on dietary SFAs.
Funding:
This study was supported by National Institute of General Medical Sciences (NIGMS) grant 5R35GM133804-02 to B.A.N.
Abbreviations:
- SFA
saturated fatty acid
- PA
palmitic acid
- WD
Western Diet
- KD
Ketogenic Diet
- TLR
toll-like receptor
- HSC
hematopoietic stem cell
- OA
oleic acid
References
- 1.Korbecki J, Bajdak-Rusinek K, The effect of palmitic acid on inflammatory response in macrophages: an overview of molecular mechanisms. Inflamm Res 68, 915–932 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Carta G, Murru E, Banni S, Manca C, Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Front Physiol 8, 902 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Divangahi M. et al. Trained immunity, tolerance, priming and differentiation: distinct immunological processes. Nat Immunol 22, 2–6 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Netea MG et al. Defining trained immunity and its role in health and disease. Nat Rev Immunol 20, 375–388 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Arts RJ et al. Glutaminolysis and Fumarate Accumulation Integrate Immunometabolic and Epigenetic Programs in Trained Immunity. Cell Metab 24, 807–819 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Saeed S. et al. Epigenetic programming of monocyte-to-macrophage differentiation and trained innate immunity. Science 345, 1251086 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mitroulis I, Kalafati L, Hajishengallis G, Chavakis T, Myelopoiesis in the Context of Innate Immunity. J Innate Immun 10, 365–372 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kaufmann E. et al. BCG Educates Hematopoietic Stem Cells to Generate Protective Innate Immunity against Tuberculosis. Cell 172, 176–190 e119 (2018). [DOI] [PubMed] [Google Scholar]
- 9.Katzmarski N. et al. Transmission of trained immunity and heterologous resistance to infections across generations. Nat Immunol 22, 1382–1390 (2021). [DOI] [PubMed] [Google Scholar]
- 10.Covian C, Retamal-Diaz A, Bueno SM, Kalergis AM, Could BCG Vaccination Induce Protective Trained Immunity for SARS-CoV-2? Front Immunol 11, 970 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Christ A. et al. Western Diet Triggers NLRP3-Dependent Innate Immune Reprogramming. Cell 172, 162–175.e114 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Seufert AL et al. Enriched dietary saturated fatty acids induce trained immunity via ceramide production that enhances severity of endotoxemia and clearance of infection. Elife 11, (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Schilling JD et al. Palmitate and Lipopolysaccharide Trigger Synergistic Ceramide Production in Primary Macrophages. Journal of Biological Chemistry 288, 2923–2932 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jin J. et al. Acid sphingomyelinase plays a key role in palmitic acid-amplified inflammatory signaling triggered by lipopolysaccharide at low concentrations in macrophages. Am J Physiol Endocrinol Metab 305, E853–867 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Fang C et al. Differential regulation of lipopolysaccharide-induced IL-1β and TNF-α production in macrophages by palmitate via modulating TLR4 downstream signaling. Int Immunopharmacol, (2022). [DOI] [PubMed] [Google Scholar]
- 16.Reyes AWB et al. Protection of palmitic acid treatment in RAW264.7 cells and BALB/c mice during Brucella abortus 544 infection. Journal of Veterinary Science 22, (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ryan DG, O’Neill LAJ, Krebs Cycle Reborn in Macrophage Immunometabolism. Annu Rev Immunol 38, 289–313 (2020). [DOI] [PubMed] [Google Scholar]
- 18.Viola A, Munari F, Sanchez-Rodriguez R, Scolaro T, Castegna A, The Metabolic Signature of Macrophage Responses. Front Immunol 10, 1462 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Curi R. e. a., A past and present overview of macrophage metabolism and functional outcomes. Clinical Science, (2017). [DOI] [PubMed] [Google Scholar]
- 20.Jha A. K. e. a., Network integration of parallel metabolic and transcriptional data reveals metabolic modules that regulate macrophage polarization. Immunity, (2015). [DOI] [PubMed] [Google Scholar]
- 21.O’Neill L, A broken krebs cycle in macrophages. Immunity, (2015). [DOI] [PubMed] [Google Scholar]
- 22.Warburg O, The metabolism of tumors in the body. Journal of General Physiology, (1927). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lancaster GI et al. Evidence that TLR4 Is Not a Receptor for Saturated Fatty Acids but Mediates Lipid-Induced Inflammation by Reprogramming Macrophage Metabolism. Cell Metab 27, 1096–1110.e1095 (2018). [DOI] [PubMed] [Google Scholar]
- 24.Innis SM, Palmitic Acid in Early Human Development. Critical Reviews in Food Science and Nutrition, (2016). [DOI] [PubMed] [Google Scholar]
- 25.Hidalgo MA, Caretta MD, Burgos RA, Long chain fatty acids as modulators of immune cells function: Contribution of FFA1 and FFA4 Receptors. Frontiers in Physiology, (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Morris A, Palmitic acid is not a TLR4 agonist. Nature Reviews Endocrinology, (2018). [DOI] [PubMed] [Google Scholar]
- 27.Abdelmagid SA et al. Comprehensive Profiling of Plasma Fatty Acid Concentrations in Young Healthy Canadian Adults. PLoS One, (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Pepino MY, Kuda O, Samovski D, Abumrad NA, Structure-function of CD36 and importance of fatty acid signal transduction in fat metabolism. Annual Review of Nutrition, (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Samovski D. et al. Regulation of AMPK activation by CD36 links fatty acid uptake to β-oxidation. Diabetes, (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hoebe K. et al. CD36 is a sensor of diacylglycerides. Nature, (2005). [DOI] [PubMed] [Google Scholar]
- 31.Zhang Y. et al. Adipose Fatty Acid Binding Protein Promotes Saturated Fatty Acid-Induced Macrophage Cell Death through Enhancing Ceramide Production. J Immunol 198, 798–807 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Spiegel S, Milstien S, Sphingosine-1-phosphate: An enigmatic signaling lipid. Nature Reviews Molecular Cell Biology, (2003). [DOI] [PubMed] [Google Scholar]
- 33.Ruysschaert J-M, Lonez C, Role of lipid microdomains in TLR-mediated signaling. Biochimica et Biophysica Acta (BBA) - Biomembranes, (2015). [DOI] [PubMed] [Google Scholar]
- 34.Schwartz EA et al. Nutrient modification of the innate immune response: a novel mechanism by which saturated fatty acids greatly amplify monocyte inflammation. Arterioscler Thromb Vasc Biol 30, 802–808 (2010). [DOI] [PubMed] [Google Scholar]
- 35.Divakaruni AS et al. Etomoxir inhibits macrophage polarization by disrupting CoA homeostasis. Cell Metabolism, (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Nishiyama K. et al. Fatty acid transport protein 1 enhances the macrophage inflammatory response by coupling with ceramide and c-Jun N-terminal kinase signaling. Int Immunopharmacol 55, 205–215 (2018). [DOI] [PubMed] [Google Scholar]
- 37.Napier BA et al. Western diet regulates immune status and the response to LPS-driven sepsis independent of diet-associated microbiome. Proc Natl Acad Sci U S A 116, 3688–3694 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Black DD, Development and Physiological Regulation of Intestinal Lipid Absorption. I. Development of intestinal lipid absorption: cellular events in chylomicron assembly and secretion. Am J Physiol Gastrointest Liver Physiol 293, (2007). [DOI] [PubMed] [Google Scholar]
- 39.Mansbach CM II, Gorelick F, Development and physiological regulation of intestinal lipid absorption. II. Dietary lipid absorption, complex lipid synthesis, and the intracellular packaging and secretion of chylomicrons. Am J Physiol Gastrointest Liver Physiol 293, (2007). [DOI] [PubMed] [Google Scholar]
- 40.Dixon BJ, Mechanisms of chylomicron uptake into lacteals. Annals of the New York Academy of Sciences, (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tso P, Nauli A, Lo C-M, Enterocyte fatty acid uptake and intestinal fatty acid-binding protein. Biochemical Society Transactions, (2004). [DOI] [PubMed] [Google Scholar]
- 42.Hall JE, Textbook of Medical Physiology 12th edition. (Saunders, 2010). [Google Scholar]
- 43.Hussain MM et al. Chylomicron assembly and catabolism: role of apolipoproteins and receptors. Biochimica et Biophysica Acta, (1996). [DOI] [PubMed] [Google Scholar]
- 44.Hubler MJ, Kennedy AJ, Role of lipids in the metabolism and activation of immune cells. J Nutr Biochem, (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Mitroulis I. et al. Modulation of Myelopoiesis Progenitors Is an Integral Component of Trained Immunity. Cell 172, 147–161 e112 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Palomer X, Pizarro-Delgado J, Barroso E, Vázquez-Carrera M, Palmitic and Oleic Acid: The Yin and Yang of Fatty Acids in Type 2 Diabetes Mellitus. Trends in Endocrinology & Metabolism 29, 178–190 (2018). [DOI] [PubMed] [Google Scholar]
- 47.Karasawa T. e. a., Saturated fatty acids undergo intracellular crystallization and activate the NLRP3 inflammasome in macrophages. Arterioscler Thromb Vasc Biol, (2018). [DOI] [PubMed] [Google Scholar]
- 48.Lee JY, Sohn KH, Rhee SH, Hwang D, Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through toll-like receptor 4. Journal of Biological Chemistry, (2001). [DOI] [PubMed] [Google Scholar]
- 49.Lee J. Y. e. a., Saturated fatty acid activates but polyunsaturated fatty acid inhibits toll-like receptor 2 dimerized with toll-like receptor 6 or 1. The Journal of Biological Chemistry, (2004). [DOI] [PubMed] [Google Scholar]
- 50.Jin J. et al. Docosahexaenoic acid antagonizes the boosting effect of palmitic acid on LPS inflammatory signaling by inhibiting gene transcription and ceramide synthesis. PLoS One 13, e0193343 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Havlicekova Z, Jesenak M, Banovcin P, Kuchta M, Beta-palmitate - a natural component of human milk in supplemental milk formulas. Nutrition Journal, (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Du Y. e. a., Maternal western diet causes inflammatory milk and TLR2/4-dependent neonatal toxicity. Genes Dev, (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Tam TH, Nucleotides released from palmitate-activated murine macrophages attract neutrophils. The Journal of Biological Chemistry, (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ortega-Gomez A. e. a., New evidence for dietary fatty acids in the neutrophil traffic between the bone marrow and the peripheral blood. Food Chemistry, (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]