

Abstract
Cone photoreceptors are the primary initiator of visual transduction in the human retina. Dysfunction or death of rod photoreceptors precedes cone loss in many retinal and macular degenerative diseases, suggesting a rod-dependent trophic support for cone survival. Rod differentiation and homeostasis are dependent on the basic motif leucine zipper transcription factor NRL. The loss of Nrl (Nrl−/−) in mice results in a retina with predominantly S-opsin containing cones that exhibit molecular and functional characteristics of WT cones. Here we report that Nrl−/− retina undergoes a rapid but transient period of degeneration in early adulthood, with cone apoptosis, retinal detachment, alterations in retinal vessel structure, and activation and translocation of retinal microglia. However, cone degeneration stabilizes by four months of age, resulting in a thinner but intact outer nuclear layer with residual cones expressing S- and M-opsins and a preserved photopic ERG. At this stage, microglia translocate back to the inner retina and reacquire a quiescent morphology. Gene profiling analysis during the period of transient degeneration reveals misregulation of genes related to stress response and inflammation, implying their involvement in cone death. The Nrl−/− mouse illustrates the long-term viability of cones in the absence of rods and RPE defects in a rodless retina. We propose that Nrl−/− retina may serve as a model for elucidating mechanisms of cone homeostasis and degeneration that would be relevant to understanding diseases of the cone-dominant human macula.
Introduction
Retinal neurodegeneration is a common feature of many blinding diseases in the developed world. Among inherited retinal diseases (http://www.sph.uth.tmc.edu/retnet/), a majority is associated with the dysfunction or death of photoreceptors (Jackson et al., 2002; Bramall et al., 2010; Swaroop et al., 2010; Wright et al., 2010). The rod photoreceptors allow vision under dim light and are capable of catching a single photon, while cone photoreceptors mediate day light vision, color perception and visual acuity (Luo et al., 2008; Mustafi et al., 2009). Any impediment in the photoreceptor metabolism or function, caused by genetic defects or microenvironment, can lead to cell death.
While rod photoreceptors die first in retinitis pigmentosa (RP) and age-related macular degeneration (AMD), the death of cones follows (Jackson et al., 2002; Punzo et al., 2009; Wright et al., 2010) suggesting a non-cell-autonomous mechanism of cone survival and/or death. The secondary cone death could result from the absence of a neurotrophic factor or a rod-derived survival factor (Faktorovich et al., 1990; Leveillard et al., 2004) or nutritional imbalance including compromised glucose uptake (Punzo et al., 2009). The study of cone photoreceptors has been difficult because of their relatively low proportion in the retina in mice and humans (Curcio et al., 1990; Mustafi et al., 2009) and/or other significant limitations (including the breeding of study models, antibody availability, and genetics) (Kryger et al., 1998; Hendrickson and Hicks, 2002; Bobu et al., 2006).
The Nrl gene encodes a basic motif leucine zipper transcription factor that is necessary for rod cell fate determination during retinal development (Swaroop et al., 2010). The loss of Nrl (Nrl−/−) in mice leads to a cone-only retina with the complete absence of rods and photoreceptors with S-cone like characteristics but shorter outer segments (OS) (Mears et al., 2001). The photoreceptors in Nrl−/− mice express cone-specific genes (Yoshida et al., 2004; Akimoto et al., 2006), have morphological and physiological features of cones (Daniele et al., 2005; Nikonov et al., 2005), yet seem to establish synaptic connections with rod bipolar cells (Strettoi et al., 2004). The analyses of Rpe65−/− and Grk1−/− mice on Nrl−/− background have revealed novel insights into cone visual cycle (Zhu et al., 2003; Wenzel et al., 2007; Feathers et al., 2008; Kunchithapautham et al., 2009).
Given the relative importance of cones in human vision and the utility of Nrl−/− mice for delineating cone biology, we undertook a comprehensive evaluation of cone-only Nrl−/− retina. Here we report our unusual findings of rapid yet transient cone cell death between one and four months of age, with a subsequent stabilization of the remaining cone number and function. This observation is different from progressive photoreceptor degeneration in rodent models of RP, yet highly significant since macular cones are preserved for longer periods in many RP patients. Our studies also show that cones can survive and function for extended periods even in the absence of rod photoreceptors.
Materials and Methods
Animal and tissue collection
Nrl−/− mice (Mears et al., 2001) were established on C57Bl/6J background. Mice of either sex were used in the study and euthanized by CO2 inhalation. Mouse eyeballs were fixed in 4% paraformaldehyde for 1 hr on ice, incubated in an increasing concentration of PBS/sucrose (10%, 20%, 30%), embedded in Tissue-Tek CRYO-OCT Compound (Fisher Scientific, Pittsburgh, PA) for cryosection. For plastic sections, eyeballs were first fixed in 4% gluteraldehyde for 30 min before overnight fixation in 4% paraformaldehyde. After embedding in methacrylate, five micron-thick sections were cut and stained with hematoxylin and eosin. For RNA extraction, the retinas were excised rapidly, frozen on dry ice and stored at −80°C. Experimental procedures involving animals were performed according to animal protocols approved by the Animal Care and Use Committee at the National Eye Institute. Mice of either sex were used for experimental procedures.
Antibodies and dyes
The following primary antibodies were used: anti-cone arrestin, anti-Sox9 and anti-GFAP polyclonal antibodies (Chemicon, Billerica, MA); anti-Brn3a monoclonal antibody (Santa-Cruz Biotechnology, Santa Cruz, CA); Acrolein polyclonal antibody (Abcam, Cambridge, MA); Peanut agglutinin conjugated to Alexa Fluor® 568 (Invitrogen, Carlsbad, CA); anti-Neurofilament-L polyclonal antibody (Millipore, Billerica, MA); AK-Fluor - fluorescein (Akorn Inc. Decatur, IL), Iba-1 (1:800, Wako Chemicals, Richmond, VA), AlexaFluor-633-conjugated phalloidin (1:100; Invitrogen, Carlsbad, CA), RPE65 polyclonal antibody (TM Redmond, National Eye Institute, Bethesda, MD); anti-S-opsin and anti-M-opsin polyclonal antibodies (CM Craft, University of Pennsylvania, Philadelphia, PA). Secondary antibodies included goat-anti-rabbit and anti-mouse antibodies conjugated with Alexa fluor 488, 568 (Molecular Probes, Invitrogen, Carlsbad, CA).
Immunohistochemistry and TUNEL assay
Cryosections were probed with specific antibodies, as previously described (Roger et al., 2006) and visualized using an Olympus FluoView FV1000 confocal laser scanning unit and Olympus BX61WI upright microscope (Olympus America Inc., Center Valley, PA). Apoptosis was detected by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assays using in situ cell death detection kit per manufacturer instructions (Roche, Indianapolis, IN).
Fundus examination, angiography, and optical coherence tomography (OCT) imaging
Experimental mice were anaesthetized, subjected to pupillary dilation with topical 0.5% tropicamide and 1% cyclopentolate hydrochloride, and photographed with a customized fundus imaging system (Paques et al., 2007). Briefly, we connected a 5-cm-long tele-otoscope (1218AA; Karl Storz, Tuttlingen, Germany) and digital camera (D80; Nikkon; Tokyo, Japan) with an additional +5.00 magnifying lens. A Xenon lamp (201315-20; Karl Storz) connected through an optic fiber to the tele-otoscope was used as the light source. Fundus angiography was performed following intraperitoneal injection of fluorescein and imaged with Micron II Rodent Fundus Imaging System (Phoenix Research Labs, Pleasanton, CA), equipped with a 390–490 nm excitation filter and a 500 nm long-pass emission filter. Still-frame images and video-rate sequences were acquired with Streampix III image acquisition software (NorPix, Montreal, Canada). Ultrahigh-Resolution Spectral-Domain Optical Coherence Tomography Imaging (SD-OCT) (Bioptigen, Durham, NC) was used to obtain cross-sectional views of the retina in vivo (right eye). Four individual B-scan images were averaged to generate a final image for analysis.
Flat mount analyses of microglia and retinal pigment epithelium (RPE) cells
Nrl−/− mice (from two wk to 11 mon) were euthanized by CO2 asphyxiation and enucleated. The globes were fixed in 4% paraformaldehyde overnight, rinsed in PBS, and then incubated in a buffer containing 1X PBS, 0.5% BSA, 0.2% Tween 20, 0.05% sodium azide, at pH 7.3. Retinal and sclerochoroidal tissues were dissected intact from the globe, flat-mounted, and processed for immunohistochemistry. Sclerochoroidal flat-mounts (with the RPE layer uppermost) were used for analysis of microglia and RPE cell morphology and distribution. Retinal flat-mounts were used to analyze the organization of photoreceptors and ganglion cells. Separately fixed eyecups from Nrl−/− mice were embedded in 7% agarose and 100 μm-thick retinal sections prepared using a vibratome (Leica VT1000S, Bannockburn, IL).
Electroretinography (ERG)
ERG was performed as previously described (Pang et al., 2005). Only photopic ERGs were recorded since Nrl−/− mice have a cone-only retina (Mears et al., 2001).
MDA assay
ThioBarbituric Acid Reactive Substances (TBAR) assay kit (Cayman Chemical Company, Ann Arbor, MI) was used to measure the concentration of malondialdehyde (MDA), a product of lipid peroxidation. For each condition, five retinas were used in two independent experiments for TBAR assays and processed according to the manufacturer’s instructions.
RNA Isolation
For microarray analysis, frozen Nrl−/− mouse retinas from four biological replicates at 1, 2, 4, 6, and 10-mon of age were lysed with a mortar and pestle in RLT buffer containing DTT (Qiagen, Valencia CA), followed by a QiaShredder column (Qiagen, Valencia, CA). RNA was isolated using the RNeasy Mini Kit employing an on-column DNase procedure with Rnase-Free DNase (Qiagen, Valencia, CA). For whole transcriptome sequencing, frozen Nrl−/− mouse retinas from 1, 2, and 4 months of age were lysed with a mortar and pestle in Trizol Reagent (Invitrogen, Carlsbad, CA). RNA quality and quantity were assessed using RNA 6000 Nano Kit (Agilent, Santa Clara, CA).
Microarray hybridization
Ribosomal reduction reactions were performed on total RNA (1 μg) from each sample using the RiboMinus Kit (Invitrogen, Carlsbad, CA). rRNA reduced RNA was used for sense target construction using the GeneChip Whole Transcript Sense Target Labeling Assay Kit (Affymetrix, Santa Clara, CA) per the manufacture’s protocol. Sense targets were hybridized to GeneChip Mouse Exon 1.0 ST arrays (Affymtrix, Santa Clara, CA), processed per manufacturer’s instructions, and scanned on a GC Scanner 3000 7G running AGCC (Affymetrix Santa Clara, CA) for .CEL file generation.
Microarray data analysis
The values of individual probes belonging to one probe set (four probes/set in 90% of the cases) were averaged and normalized using Partek Genomics Suite 6.5 (Partek Inc., St. Louis, MO). The average fluorescence intensity was calculated for annotated genes using Robust Multichip Average (RMA) algorithm. The updated mapping files were downloaded from Affymetrix’s web site and core meta-probe set list having strong annotation support (e.g., from RefSeq and other GenBank alignments of ‘complete CDS’ transcripts) was used for further analysis. Oneway Analysis of Variance (ANOVA) was performed to identify differentially expressed genes. To correct for false positives, we used the Benjamini-Hochberg false discovery rate procedure (FDR) with a minimum p-val ≤ 0.05. Only genes showing a greater than two fold change with respect to 1-mon expression data were considered for further analysis.
Whole transcriptome sequencing (RNAseq)
Transcriptome libraries were constructed using 1 μg of total RNA with the mRNA-seq Sample Preparation Kit (Illumina, San Diego, CA), according to manufacture’s protocols with only minor modifications. Briefly, ligated library DNA were electrophoresed on a 2% agarose gel and stained with SYBR Gold. The 200 bp band was isolated using a 1 mm GeneCatcher (Gel Company, San Francisco, CA), and DNA recovered using QIAquick Gel Extraction Kit (Qiagen, Valecia, CA). The library DNA was quantitated using DNA-1000 Kit (Agilent, Santa Clara, CA). Single-read cluster generation of 10 pM libraries was performed using the Single-Read Cluster Generation Kit v4 (Illumina, San Diego, CA). Sequence-by-synthesis of 76 base length was carried out on a Genetic Analyzer IIx running SCS2.6 (Illumina, San Diego, CA).
RNA-Seq data analysis
Raw sequence reads were mapped onto the University of California, Santa Cruz mm9 mouse genome using the public domain Efficient Large Scale alignment of Nucleotide Database (ELAND). This data was analyzed with Partek software (Partek Inc., St. Louis, MO) and normalized using the method that calculated reads per 1 kb of exon model per million mapped reads (RPKM) (Marioni et al., 2008). RefSeq Transcripts database (http://www.ncbi.nlm.nih.gov/RefSeq/) was used to map the sequence reads to the mouse genome. One-way ANOVA analysis was performed to identify differentially expressed genes.
Results
Transient cone photoreceptor cell death in Nrl−/− mouse retina
Fundus examination of the Nrl−/− mice revealed abnormalities with age (Figure 1A). White spots were observed in 1-mon old Nrl−/− but not in age-matched wild type (WT) C57Bl6 mouse retina; these white lesions correspond in size and location to whorls and rosettes reported in the outer nuclear layer (ONL) of Nrl−/− retina (Mears et al., 2001; Daniele et al., 2005; Oh et al., 2007). In addition, an accumulation of pigmentary abnormalities and increased attenuation of the retinal vessels were observed in 2-mon old Nrl−/− mice. At six months, papillary atrophy was evident, suggesting optic nerve or retinal ganglion cell loss (Figure 1A).
Figure 1. Cone degeneration in Nrl−/− mice.

(A) Fundus image of 1-mon old WT and 1, 2, 6, 10-mon old Nrl−/− mice. White dots corresponding to pseudorosettes were observed in Nrl−/− mice at 1 and 2-mon of age (white arrowheads). Attenuated blood vessels were observed in 10-mon old Nrl−/− mice and optic atrophy increased at 6-mon (white arrows). (B) Light adapted Nrl−/− mice ERG recording showed a decrease between 1 and 4-mon of age of a-and b-waves. (C) Average of the maximum amplitude for a- and b-wave in Nrl−/− mice at 1, 2, 4, 6 and 10-mon of age. The maximum decrease is observed between 2- and 4-mon. Error bars show ±SEM from four independent mice (D) Methacrylate sections followed by H&E staining on 1, 2, 4 and 10-mon old Nrl−/− mouse retina showed a loss of nuclei in the outer nuclear layer between 1 and 4 months. Rpe, Retinal pigment epithelium; onl, outer nuclear layer; inl, inner nuclear layer; gcl, ganglion cell layer. Scale bar = 20μm. (E) Transmission EM images of Nrl−/− mouse retina in the dorsal-ventral midline plane, taken through the central retina, at 1, 4, and 10-mon of age. Rudimentary OS are evident at 1-mon but not at 4 and 10 months. Abnormal accumulation of IS material is detected in the ONL at 4 and 10 months; the ONL in wild-type mice contains primarily nuclei (data not shown). The stacked membranous structures at the RPE-subretinal interface correspond to apical microvilli of the RPE, identified by their position, spacing, and length. Scale bar = 2 μm. Outer segments (OS) in 1-mon old retina are shown at higher magnification. Note that while regular stacks of discs begin to form, they do not elongate (as in WT mice). Some areas of abnormal membrane vesicle formation are present (asterisks). Scale bar = 500 nm. Abbreviations: RPE, retinal pigment epithelium; apMV, apical microvilli of the RPE; OS, outer segment; IS, inner segments; OLM, outer limiting membrane; ONL, outer nuclear layer. (F) OCT images of 2-mon old WT retina and 2-and 10-mon old Nrl−/− retina. Center and periphery areas of the retina were imaged. In old animals, ONL is thinner, and large arteries were observed by OCT Doppler (red and blue color indicated by white arrows). Areas of retinal detachment (asterisk) and pseudorosettes (arrowheads) were observed only in the central part and periphery of the 2-month old Nrl−/− retina.
As previously described (Mears et al., 2001; Daniele et al., 2005), photopic ERG recordings (corresponding to cone function) in Nrl−/− mice at 1- and 2-mon of age showed supranormal amplitudes for both a- and b-wave (Figure 1B–C). However, repeated recordings at different ages between 1 and 4 months demonstrated a drastic reduction in the mean a-wave amplitude, from 50.6±1.1μV at 1-mon to 16±2.4μV, at 4-mon age (Figure 1C). The b-wave amplitudes at maximum light stimulation were relatively stable between 1- and 2-mon (189±11μV and 179±19μV, respectively) (Figure 1B, C), but decreased significantly (to 109±9μV) by 4-mon. Histological analysis in methacrylate sections of the Nrl−/− retina demonstrated massive photoreceptor cell loss retina between 2- and 4-mon of age (Figure 1D). During this period, the ONL decreased in thickness from 8–10 rows of nuclei to about 4 rows. Interestingly, ONL thickness remained relatively stable (2–4 rows) at 6 mon and later. However, at 10-mon, the ganglion cell layer showed progressive thinning compared to younger Nrl−/− mice (Figure 1D), suggesting a cause for optic atrophy observed by fundoscopy at this age (Figure 1A). Even in 18-mon old mice, the thickness of the ONL was unchanged (2–4 rows of nuclei; data not shown), indicating that the complete absence of rod photoreceptors in Nrl−/− mouse retina did not lead to further loss of cones.
Transmission electron microscopy (TEM) images of Nrl−/− mouse retina in the dorsal-ventral midline plane and in the dorso-central retina demonstrated the presence of short OS at 1-mon of age (Figure 1E). While regular stacks of OS discs were present, they did not elongate as in WT mice, and some areas of abnormal membrane vesicle formation were observed (asterisks, Figure 1E). By 4-mon, OS have progressive shortened such that the distinction between OS and inner segment (IS) could no longer be discerned. In addition, abnormal accumulation of IS material appeared in the ONL at 10-mon. Membrane stacks corresponding to RPE apical microvilli (as identified by their position, spacing and length) were also observed (Figure 1E).
OCT imaging in 1-mon old live animals showed broad areas of shallow retinal detachment from the RPE, distinct from the pseudorosettes located in the ONL (Figure 1F). These detachments were detected both in the center and the periphery of the retina without a preferential locus of occurrence. However, all areas of retinal detachment had resolved at 4-mon of age. By OCT Doppler imaging, large retinal blood vessels of 2- and 10-mon old Nrl−/− mice had increased cross-sectional areas (Figure 1F) compared to WT mice.
Preservation of S-cones and M-cones following the period of retinal cell death in the Nrl−/− retina
Immunolabeling with peanut agglutinin (PNA), a marker of cone extracellular matrix, in 1-mon old Nrl−/− mice showed prominent co-localization with whorls and rosettes that were previously noted in Nrl−/− and rd7 mice (Figure 2A) (Akhmedov et al., 2000; Mears et al., 2001). The PNA-stained rosette structures in the retina decreased drastically in number from 968±41 per retina at 1-mon to 141±16 per retina at 4-mon, and were completely absent at 10-mon. However, PNA-positive staining continued to be present in the distal cone segments at all ages, indicating a persistence of the cone extracellular matrix in that location (Figure 2A,B). S-opsin, M-opsin and cone arrestin were also uniformly expressed at all ages (Figure 2C). Interestingly, M-opsin was relocalized in the IS in old mice in contrast with its presence in the ONL observed in younger animals. The counting of TUNEL-positive cells confirmed that most cones underwent apoptosis between 2- and 4-mon (Figure 2D). We did not observe significant differences in the repartition of the TUNEL-positive cells between areas of retinal detachments and rosettes. However, the cell death in the rosettes could be due to the lack of contact of cone outer segments with RPE, as suggested in a previous study (Daniele et al., 2005). Surprisingly, a significant number of apoptotic cells were also detected in the inner nuclear layer (INL) and ganglion cell layer (GCL). Thus, a majority of cell death in Nrl−/− retina is transitory during the first 4-mon of life and occurs in all retinal layers.
Figure 2. Transitory cell death between 1 and 4-mon in Nrl−/− retina.

(A) Identification of pseudorosettes with PNA on flat-mount retina and cryosections on 1 and 10 months old Nrl−/− mice. (B) Average of the number of pseudorosettes in Nrl−/− mice counted after labeling with PNA on flat mount retina at 1, 2, 4, 6 and 10-mon of ages. The maximum decrease is observed between 1 and 4 months. Error bars show ±SEM from four independent mice. (C) S-opsin, M-opsin and cone arrestin immunostaining (green) in 1-and 10-mon old Nrl−/− retina show maintenance of expression in old mice. (D) By TUNEL assay, quantification of apoptotic cells per section in the outer nuclear layer (dashed line), inner nuclear layer (solid line) and ganglion cell layers (dotted line), separately in 1, 2, 4, 6 and 10-mon old Nrl−/− mice. Error bars show ± SEM from four independent mice. The data shows cell death in all retinal layers between 2 and 4 months. The cell death was more pronounced in the inner nuclear layer compared to outer nuclear layer. (A, C) ONL, outer nuclear layer; INL, inner nuclear layer; GCL, ganglion cell layer. Nuclei were visualized by DAPI staining. Scale bar = 20μm.
Alterations in ganglion and Muller cells in Nrl−/− retina
As cell death was detected in all neuronal layers, we examined the markers for different retinal cell types in Nrl−/− mice. We observed the expression of Brn3a and NF-L, two ganglion cell markers, in the INL of 1- and 2-mon old Nrl−/− retina (Figure 3A), suggesting the abnormal location of displaced retinal ganglion cells. Interestingly, a fraction of the Sox9-positive Muller cell nuclei were also displaced to the ONL at 4-mon (Figure 3B). Notably, Muller cells showed an evidence of gliosis (as revealed by GFAP immunostaining) as early as 1-mon (Figure 3C) when cell death is initiated.
Figure 3. Altered cell types in 1, 2, 4 and 10-mon Nrl−/− retina.

(A) Ganglion cells were immunolabeled with anti-Brn3a (red) and anti NF-L (green) and showed presence of displaced ganglion cells in the INL of young animal (1 and 2-mon old). These were not present in 4 and 10-mon old animals. (B) Muller cells, immunolabeled with anti-Sox9 antibody (green), migrated first around the remaining pseudorosettes (at 4-mon) then in the entire outer nuclear layer (10-mon). (C) GFAP immunolabeling (red) showed strong activation of Muller cells at 2, 4 and 10-mon of age.
Since microglial cell activation is reported in many neurodegenerative diseases and models of retinal degeneration (Hanisch and Kettenmann, 2007; Langmann, 2007), we assessed changes in microglial morphology and distribution in Nrl−/− mice during the period of transitory cell death and with age (Figure 4). At 2-wk of age, retinal microglia (as indicated by labeling with Iba1 immunostaining) demonstrated ramified morphologies and a typical distribution (i.e., in the GCL, inner plexiform layer (IPL), and outer plexiform layer (OPL), and were excluded from the outer retina and subretinal space) in Nrl−/− retina, similar to those in WT mice. By 3-wk, a few microglial cells in the ventral retina could be seen migrating into the subretinal space coming into contact with RPE, and assuming more rounded and less ramified morphologies, consistent with an activated status (Figure 4C). The influx of microglia into the outer retina and subretinal space increased with age, peaking at 2-mon (Figure 4K, M), exhibiting close association with cone photoreceptor OS including those located in pseudorosettes (Figure 4M). Beginning at 4-mon of age, a progressive decrease was detected in the number of microglia in the subretinal space (Figure 4K), with a resumption of microglia distribution to OPL, IPL, and GCL (Figure 4N). Microglia appeared to be recruited to the areas of cone degeneration early in the process, prior to extensive cone loss, and returned to a typical inner retinal distribution after the transitory phase of retinal cell death.
Figure 4. Migration of retinal microglia into the outer retina during the period of photoreceptor degeneration.

Microglia migration into the subretinal space onto the RPE monolayer were imaged in sclerochoroidal flat-mounts in the ventral and dorsal quadrants respectively at different ages: 2 weeks (A, B); 3 weeks (C, D); 2 months (E, F); 4 months (G, H); 11 months (I, J). (K) Quantification of microglia in the subretinal space demonstrates that microglia accumulation begins at 3 weeks and peaks at 2 months, with larger numbers accumulating in the ventral quadrant compared to the dorsal quadrant. RPE cell density and morphology are not significantly changed during this period. (L, M, N) Cross-sectional views of microglia distribution in vibratome sections of the retina at different ages. At 2 months (M), microglia are closely associated with photoreceptor rosette structures (arrow) and also in the subretinal space (arrowheads). At 4 months (N), microglial density in the outer retina decreases as the photoreceptor pseudorosettes resolve and disappear. Scale bars = 50 μm
Absence of rod photoreceptor affects RPE integrity, retinal vasculature and ganglion cell survival
As interaction of the RPE with photoreceptors is essential for visual function, we investigated the contact of these two layers on 100 μm thick vibratome sections (Figure 5A) and RPE integrity in sclerochoroidal flat mounts (Figure 5B). In the RPE layer of WT mice, RPE65, a key enzyme of the visual cycle, could be immunolocalized to the cytoplasm of all RPE cells, while ezrin was detected in the apical RPE cell surface. In Nrl−/− retina, despite continuous PNA localization in the cones, expression of RPE65 and ezrin was absent in some regions of the RPE monolayer, suggesting a patchy loss of RPE cell function. As early as 1-mon of age, we could identify abnormalities in the RPE mosaic (revealed by phalloidin immunolabelling) of Nrl−/− retina. Some cells did not express RPE65, and others showed internal staining with phalloidin in the focal plane, which likely corresponded to the microvilli. By 10-mon age, the RPE integrity appeared to be disrupted, as shown by the presence of some pyknotic nuclei and loss of junctions between RPE cells (Figure 5B). Our data is consistent with a recent report demonstrating defective phagocytosis in RPE from Nrl−/− retina (Mustafi et al., 2011).
Figure 5. Changes in RPE in Nrl−/− retina.

(A) Immunolabeling on vibratome sections of RPE with anti-RPE65 (green), cones with PNA (red) and RPE apical side with anti-Ezrin (grey) on 4-month old WT and Nrl−/− mouse retina showed weaker RPE65 expression and absence of ezrin in front of cone photoreceptors in Nrl−/− mice compared to WT mice. Scale bar: 40μm (B) Top Panel: RPE-sclerochoroidal whole mount of 1 month WT mice and 1- and 10-mon old Nrl−/− mice after immunolabeling of RPE cellular outlines stained with Phalloidin (red) and anti-RPE65 (green). Compared to RPE from WT mice, RPE from Nrl−/− mice showed abnormalities in their junctions with presence of large patches positive for phalloidin. Nuclei are visualized by DAPI staining. Arrowhead indicates apoptotic bodies. If in WT mice, RPE65 showed homogenous expression all RPE cells, in contrast RPE65 expressions were absent in a large number of RPE cells and even among the cells with phalloidin staining in their cell body. Bottom Panel: High magnification of methacrylate sections followed by H&E staining showed a loss of RPE cells in aged (10-mon) Nrl−/− retina. Scale bar = 20μm.
We also evaluated the structure and permeability of the retinal vasculature using fluorescein angiography. Two-mon old Nrl−/− mice demonstrated dilated retinal vessels and also a denser network of retinal capillaries (Fig 6B), supporting the observations made by OCT imaging (Figure 1F). Some abnormal retinal capillary permeability was also found, as evidenced by mild leakage of fluorescein in the retinal periphery (Figure 6C). A 3D reconstruction from flat mount retina of 2-mon old Nrl−/− mice after dextran-fluorescein injection demonstrated that the leakage observed by angiography came in part from blood vessels present on the internal retinal ganglion cell layer side (Figure 6D). This observation was confirmed in 5-mon old Nrl−/− retina (data not shown). With increasing age, the retinal vessels showed a converse luminal narrowing and an increasing extent of fluorescein leakage (Figure 6E, F).
Figure 6. Age-associated changes in vascular structure and permeability in Nrl−/− retina.
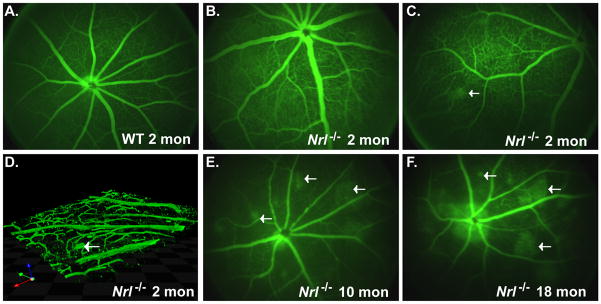
(A–C, E–F) Angiography after intraperitoneal injection of fluorescein in 2-mon old WT mice (A) and in Nrl−/− mice at 2-mon (B–C), 10-mon (E) and 18-mon (F). Leakage of fluorescein was observed first at 2-mon with increase of the area of leakage during aging. (D) 3-D reconstruction form 0.2 μm z-section taken from flat mount retina of 2-mon old Nrl−/− mice after injection of fluorescein showed the presence of abnormal aneurysmal dilatations budding from the blood vessels in the ganglion cell layer.
As GFAP-expressing astrocytes in the GCL have a close relationship with the retinal vasculature (Gariano et al., 1996; West et al., 2005), we evaluated the effects of retinal degeneration on vasculature integrity using flat mount retina (Figure 7). We observed a profound change in both large arteries and capillaries in 18-mon old Nrl−/− retina that in part explained the massive blood leakage detected by angiography (Figure 7).
Figure 7. Age-associated alterations of the superficial retinal vasculature and ganglion cell death in Nrl−/− retina.

Immunohistochemistry on whole-flat mount retina with anti-GFAP (green) and anti-Brn3a (red) antibodies labeling astrocytes, activated Muller cells and ganglion cells, respectively. GFAP-positive cells aligning blood vessels on the ganglion cell layer side showed profound disorganization of the retinal vasculature, with large areas with no staining in 18-mon old Nrl−/− mice compared to WT control. Presence of ganglion cells was assessed with anti-Brn3a immunolabeling. Nrl−/− retina did not show high level of Brn3a expression at all time points. Loss of ganglion cells occurred in 18-mon old Nrl−/− mice but was more pronounced at the periphery compared to central retina.
In parallel, we immunolabeled ganglion cells on flat mount retina with anti-Brn3a, a transcription factor specifically expressed in ganglion cells. Consistently, the expression level of Brn3a was lower in Nrl−/− retina compared to WT (Figure 7). Thus, if ganglion cells were still present in the central retina of 18-mon old Nrl−/− mice, they would have low Brn3a expression levels. In contrast, only few ganglion cells were detected at the periphery of the retina, confirming the death of ganglion cells as indicated (Figure 1A, D; Figure 3A). Interestingly, we also observed an increase in small capillaries, as revealed by the secondary anti-mouse IgG that bind to the endogenous IgG in the blood contained in mouse retinal capillaries.
Our findings reveal that transitory cone cell death in Nrl−/− mice appears to cause profound changes in the retina, including RPE cell death, changes in the retinal vasculature, ganglion cell death, and fluid leakage from the superficial blood vessels on the inner retinal ganglion cell side. However, loss of RPE might also disrupt the retinal blood barrier, resulting in fluid leakage from the choroid side.
Oxidative stress associated with retinal cell degeneration in Nrl−/− mice
The oxygen stress in Nrl−/− mouse retina is predicted to be higher than in WT as illustrated by the fact that the overall photoreceptor circulating current in the Nrl−/− cones at 1-mon (when cones are still healthy) is roughly one-third that of WT (as reflected by the saturated a-wave amplitude), and therefore the outer retina of the Nrl−/− mice will require only about one-third as much oxygen supplied by the choroid, leading to elevated oxygen tension (Daniele et al., 2005). We therefore investigated the oxidative stress in Nrl−/− retina by assessing the presence of acrolein, a toxic product formed by lipid peroxidation (Figure 8A). While staining for acrolein was absent in 1-mon old retina, prominent staining was observed at 2-mon, particularly in the ganglion cell layer and around the whorls and rosettes of the ONL, before decreasing in overall intensity at 10-mon. We also measured the concentration of MDA, a marker of oxidative stress, in Nrl−/− and WT retinas at different ages. In Nrl−/− retinas, levels of MDA were low and similar to those found in WT retinas at 1-mon, but increased to a peak at 4-mon (2.5-fold increase) before decreasing at 10-mon (Figure 8B). However, we noticed that the peak of MDA is delayed with significant cone cell loss occurring between 2- and 4-mon of age. The concordance between increased acrolein staining and MDA levels at this age implicates a role of oxidative stress in the cone cell death.
Figure 8. Lipid peroxidation in Nrl−/− retina.
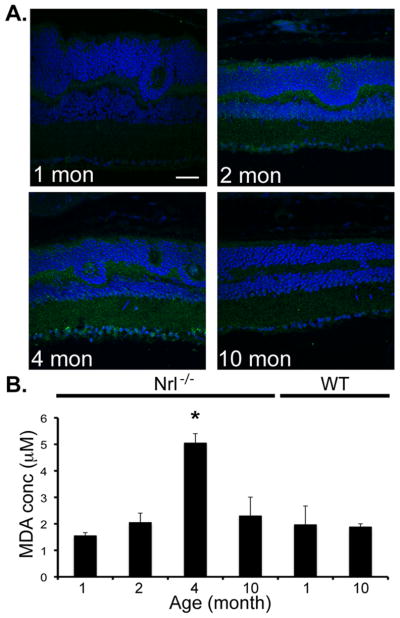
(A) Immunolabeling of acrolein (geen), a toxic product formed from lipid peroxidation and used as a marker of oxidative stress, was increased at 2- and 4-mon in Nrl−/− mice, relative to 1- and 10-mon. Scale bar = 20 μm (B) Thiobarbituric acid reactive substances (TBARS) assay was used to assessed lipid peroxidation in retinal lysates of 1, 2, 4, 10 months-old Nrl−/− mice and of 1- and 10-mon old WT mice. The assay shows a significant increase of malondialdehyde (MDA) in 4-mon old Nrl−/− mouse retina. Error bars indicate ±SD. Standard t-test was used to determine significance. * indicates p value <0.05.
Changes in gene expression profile during cone cell death in Nrl−/− retina
To gain insights into cellular pathways underlying transitory cone death in Nrl−/− retina, we performed microarray analysis of four biological replicates at five different time points (1, 2, 4, 6 and 10-mon). We compared 2, 4, 6- and 10-mon microarray data to 1-mon, when all cones are still present, and selected a minimum 2-fold change (FC) cut-off for further analysis. The data revealed statistically significant differences (P ≤ 0.05) for 350 mapped genes in at least one of the four comparisons (data not shown). As most cone death occurs between 1- and 4-mon of age in Nrl−/− retina, we grouped together genes with FC ≥2 at 2- or 4-mon and those differentially expressed at 6- or 10-mon with respect to 1-mon. At 6- or 10-mon, 337 genes were significantly differentially expressed compared to 1-mon and 88 at 2- or 4-mon compared to 1-mon. The fact that the vast majority of genes are differentially expressed at 6- or 10-mon might reflect the remodeling of the retina after a period of transitory cone cell death. We then focused on 88 genes that are potentially involved in cone cell death and exhibit FC >2. Statistically significant down-regulation was detected for 53 genes with an average fold change (AFC) of −2.4 and −2.6 at 2- and 4-mon, respectively (Figure 9A). Thirty-five genes with statistically significant upregulation demonstrated AFC of 2.2 and 2.5 at 2- and 4-mon, respectively (Figure 9B).
Figure 9. Gene expression analysis of Nrl−/− retina.

(A–B) Hierarchical clustering dendrogram of 350 genes having a False Discovery Rate (FDR) p≤0.05 and a minimum fold change 2 between 2- or 4-mon compared to 1-mon. Bright blue indicates lowest signal with increasing expression by yellow to bright red. (C) Chart pie representation of the main biological function representing 88 genes differentially expressed. (D) qRT-PCR validation of Cryαa, Cryβa1, Cryβb1, Cryγc and Cryγd using 4 individual biological replicates at 1, 2, 4, 6 and 10-mon. Bars indicate ±SEM. (E) Network reconstruction using Ingenuity Pathway Analysis (IPA) with 13 genes having a False Discovery Rate (FDR) p≤0.05 and a minimum fold change 2 only between 2 or 4-mon compared to 1-mon. (F) Network reconstruction using IPA with 39 genes identified as differentially expressed between 2 or 4-mon compared to 1 month after comparison with microarray and RNAseq analysis. Red indicates upregulation of the gene of interest at 2-mon compared to 1-mon; green indicates downregulation of the gene of interest at 2-mon compared to 1-mon.
Meta analysis of biological functions of 88 differentially expressed genes at 2-and 4-mon compared to 1-mon identified three major categories – immune response, signal transduction and stress response (Figure 9C). Among these, Crystallins correspond to a large family of chaperones involved in response to stress and inflammation, and their expression is increased in several retinal diseases (Fort and Lampi, 2011) including AMD (Umeda et al., 2005), uveitis (Rao et al., 2008) and diabetic retinopathy (Kumar et al., 2005; Fort et al., 2009). Interestingly, Cryαa, Cryβa1, Cryβb1, Cryγc and Cryγd were highly expressed at 1, 2- and 4-mon of age in Nrl−/− retina (Table 1). We confirmed their down-regulation with age by qPCR (Figure 9D), corresponding to the transitory cell death. However, Crystallin expression is almost undetectable thereafter (6- and 10-mon). Cryαa, Cryβa1 and Cryγd are among the most highly expressed crystallins between 1- and 4-mon, indicating their potential role in cone homeostasis and response to stress.
Table 1. Differentially expressed genes at two and four months compared to one month in common between microarray and RNAseq analysis.
Microarray analysis identified 88 genes with FC2 differentially expressed between two and four months compared to one month. Among them, 39 were present in RNAseq data analysis. These genes are potentially involved in the transient cell death occurring in Nrl−/− retina between one and four months of age.
| Microarray | RNAseq | |||||
|---|---|---|---|---|---|---|
|
| ||||||
| Gene Symbol | RefSeq | Entrez Gene Name | FC 2 vs 1 | FC 4 vs 1 | FC 2 vs 1 | FC 4 vs 1 |
| Immune response | ||||||
| Cxcl13 | NM_018866 | chemokine (C-X-C motif) ligand 13 | 3.01 | 1.82 | 8.32 | 4.32 |
| Lcn2 | NM_008491 | lipocalin 2 | 2.24 | 1.73 | 6.90 | 5.93 |
| Ctss | NM_021281 | cathepsin S | 2.12 | 1.60 | 2.30 | 1.81 |
| Ifitm3 | NM_025378 | interferon induced transmembrane protein 3 | 2.07 | 1.43 | 2.60 | 1.39 |
| Serping1 | NM_009776 | serpin peptidase inhibitor, clade G, member 1 | 2.04 | 2.11 | 3.27 | 2.48 |
| A2m | NM_175628 | alpha-2-macroglobulin | 1.90 | 2.51 | 2.86 | 2.62 |
| Anxa1 | NM_010730 | annexin A1 | 1.83 | 2.26 | 1.71 | 2.14 |
| Cfl | NM_007686 | complement factor I | 1.74 | 2.01 | 3.22 | 2.45 |
| Cd93 | NM_010740 | CD93 molecule | −1.52 | −2.01 | −1.61 | −3.10 |
| Tnfsf13 | NM_023517 | tumor necrosis factor superfamily, member 13 | −1.59 | −2.03 | −2.25 | −2.65 |
| Glmn | NM_133248 | glomulin, FKBP associated protein | −2.10 | −2.20 | −1.83 | −2.01 |
| Signal transduction | ||||||
| Drd4 | NM_007878 | dopamine receptor D4 | 2.71 | 2.31 | 4.05 | 3.54 |
| Ednrb | NM_007904 | endothelin receptor type B | 2.36 | 2.75 | 1.85 | 2.79 |
| Egfr | NM_207655 | epidermal growth factor receptor | 1.93 | 2.15 | 1.84 | 2.47 |
| Gna14 | NM_008137 | guanine nucleotide binding protein, alpha 14 | 1.83 | 2.40 | 2.56 | 3.08 |
| Arrdc2 | NM_027560 | arrestin domain containing 2 | 1.39 | 2.10 | 2.51 | 6.47 |
| Pde6b | NM_008806 | phosphodiesterase 6B, cGMP-specific, rod, beta | −1.11 | −2.08 | −1.12 | −2.26 |
| Mtnr1a | NM_008639 | melatonin receptor 1A | −1.53 | −2.08 | −1.23 | −5.23 |
| Tnfrsf18 | NM_009400 | tumor necrosis factor receptor superfamily, member 18 | −2.00 | −2.05 | −2.07 | −1.59 |
| Response to stress anti-apoptosis visual perception | ||||||
| Crygs | NM_009967 | crystallin, gamma S | −2.29 | −2.30 | −3.5E+16 | −4.4E+15 |
| Crygd | NM_007776 | crystallin, gamma D | −2.64 | −2.33 | −7.4E+16 | −5.7E+15 |
| Crygc | NM_001082573 | crystallin, gamma C | −2.70 | −2.78 | −7.2E+15 | −6.2E+15 |
| Cryba1 | NM_009965 | crystallin, beta A1 | −4.46 | −4.71 | −7.5E+15 | −4.4E+15 |
| Crybb1 | NM_023695 | crystallin, beta B1 | −4.95 | −4.18 | −11.34 | −15.34 |
| Cryaa | NM_013501 | crystallin, alpha A | −14.55 | −14.54 | −4.4E+15 | −4.4E+15 |
| Regulation of transcription | ||||||
| Esrrb | NM_001159500 | estrogen-related receptor beta | 2.85 | 3.44 | 2.30 | 2.82 |
| En2 | NM_010134 | engrailed homeobox 2 | 1.85 | 2.11 | 1.82 | 2.89 |
| Klf15 | NM_023184 | Kruppel-like factor 15 | 1.51 | 2.01 | 1.90 | 2.03 |
| Cell adhesion | ||||||
| Col6a1 | NM_009933 | collagen, type VI, alpha 1 | −1.88 | −2.06 | −2.79 | −2.73 |
| Cldn5 | NM_013805 | claudin 5 | −1.90 | −2.38 | −1.55 | −2.64 |
| Metabolic process | ||||||
| Hs6st2 | NM_001077202 | heparan sulfate 6-O-sulfotransferase 2 | 2.87 | 4.95 | 2.40 | 3.44 |
| Pnpla3 | NM_054088 | patatin-like phospholipase domain containing 3 | −2.05 | −2.31 | −2.51 | −2.67 |
| Regulation of cell growth | ||||||
| Fam107a | NM_183187 | family with sequence similarity 107, member A | 2.36 | 2.64 | 3.77 | 3.79 |
| Transport | ||||||
| Tc2n | NM_001082976 | tandem C2 domains, nuclear | 2.74 | 2.65 | 8.35 | 8.38 |
| Scn7a | NM_009135 | sodium channel, voltage-gated, type VII, alpha | 2.12 | 2.83 | 1.85 | 2.30 |
| Unknown | ||||||
| Fam159b | NM_029984 | family with sequence similarity 159, member B | 2.10 | 3.10 | 2.43 | 5.30 |
| Samd7 | NM_029489 | sterile alpha motif domain containing 7 | 2.09 | 1.69 | 2.02 | 1.81 |
| Agxt2l1 | NM_027907 | alanine-glyoxylate aminotransferase 2-like 1 | 2.06 | 2.73 | 1.61 | 2.44 |
| Acrbp | NM_016845 | acrosin binding protein | −2.35 | −2.28 | −2.11 | −1.97 |
Only thirteen genes were differentially expressed between 1- and 4-mon, with no statistically significant expression change between 6- and 10-mon compared to 1-mon:Cxcl13, Ifit1, Serpina3n, Asf1b, Ctss, Samd7, Ifitm3, Serping1, Cfi, Hnrpdl, Col6a1, Abhdl4a, Pcbp4. Ingenuity Pathways Analysis (IPA) revealed that 12 out of 13 genes act as hubs with a majority associated with immune response: Cfi, Ctss, Cxcl13, Ifit1, Iftm3, Serpina3n and Serping1 (Figure 9E).
We then performed whole transcriptome analysis by NextGen Sequencing (RNA-Seq) of 1, 2- and 4-mon Nrl−/− retina and identified 39 genes (Table 1) that were also present in 88 differentially expressed genes revealed by microarray analysis. Network reconstruction with these 39 genes (Figure 9F) identified Tnf (Tumor Necrosis Factor) as a central hub for 12 differentially expressed genes, confirming the importance of immune response in cone death observed in Nrl−/− retina. The presence of SerpinG1 up-regulation is notable as its expression is associated with AMD (Mullins et al., 2009; Lu et al., 2010).
Discussion
The studies of cone function are critical for elucidating normal visual process and defining pathogenic mechanisms of retinal diseases. Photoreceptors in Nrl−/− retina share many features with “normal” cones (Daniele et al., 2005), including chromatin clumping, cone matrix sheath, gene expression, mitochondrial length, open discs, and ERG pattern. Here we report novel aspects of cone biology by comprehensive examination of Nrl−/− retina, which at 1 to 4 month age shares fundamental features (e.g., photoreceptor degeneration, RPE changes, Muller cell hypertrophy) with rodent models of retinal degeneration and human RP (Milam et al., 1998). The rapid cone cell death in Nrl−/− retina is similar to what is observed in Pde6b−/− and Pde6g−/− mouse retina between one and four months of age (Punzo et al., 2009). However, unlike rodent and human RP, cone photoreceptor death stabilizes in the Nrl−/− retina, with a population of cones for the remainder of life. Notably, macular cones, though not functional, are preserved in many RP and LCA patients (John et al., 2000; Cideciyan et al., 2007; Jacobson et al., 2010).
An important question is why the cones in the Nrl−/− retina do not undergo a constant rate of degeneration like their counterparts in the macula of an RP patient? Large areas of retinal detachment, loss of ganglion cells and altered vasculature in Nrl−/− retina at a young age indicate neuronal degeneration and remodeling (Milam et al., 1998; Marc et al., 2003). Unlike Nrl−/− retina, inner retinal neurons are more preserved and send processes into aberrant regions in human RP retina (Stone et al., 1992; Santos et al., 1997; Humayun et al., 1999). The remodeling of synaptic connections between photoreceptors and bipolar cells is also observed in animals carrying rhodopsin or Pdeb mutation (Peng et al., 2000) or after retinal detachment (Lewis et al., 1998). In Nrl−/− retina, the rod bipolar cells connect to cone terminals in the absence of rods (Strettoi et al., 2004), whereas cone bipolars are connected to rods in the absence of cones in Crxp-Nrl mouse (Oh et al., 2007). The axon and dendritic terminals of horizontal cells are also affected in absence of rod or cone photoreceptors (Raven et al., 2007).
Ablation of cones by cone-DTA transgene in rd7/rd7 mouse retina, caused by the loss of Nr2e3 (a transcriptional target of NRL (Oh et al., 2008)) function, reduces Muller cell activation, indicating a contribution of excess cones to pseudorosette formation (Chen and Nathans, 2007). The cone death between 1- and 4-mon in Nrl−/− retina results in complete loss of pseudorosettes without preventing Muller cell activation. We suggest that loss of rosettes in older Nrl−/− retina allows appropriate positioning of surviving cones with RPE, which can explain the arrest of cone cell death. Notably, prompt surgical reattachment after retinal detachment in humans prevents visual acuity loss and allows vision restoration (Fisher and Lewis, 2003). In monkeys that had detachment/reattachment surgery, the cones degenerate while detached but their OS length partially recovers when reattached (Guerin et al., 1989). Retinal detachment in Nrl−/− retina does not significantly alter S- or L/M-opsin expression, as in a cat model of retinal detachment (Rex et al., 2002). Additionally, formation of glial scar and retinal remodeling (Jones and Marc, 2005; Lewis et al., 2010) are undetectable in Nrl−/− retina. Experimental retinal detachment (Lewis et al., 2010) leads to proliferation of Muller cells and migration of their cell bodies to ONL. Thus, retinal detachment early in Nrl−/− retina can partly explain the presence of Muller cell bodies in the ONL and may reflect a consequence of dedifferentiation (Fischer and Reh, 2003; Bernardos et al., 2007; Karl et al., 2008). A few BrdU-positive cells were visible in the ONL of 12-mon but not 2-mon old Nrl−/− retina (data not shown). Further investigations are required to delineate the identity of these cells.
RPE plays a critical role in the maintenance of photoreceptor integrity (Strauss, 2005; Sparrow et al., 2010). The use of Rpe65−/−;Nrl−/− mice has demonstrated the importance of RPE65 in both rods and cones (Wenzel et al., 2007; Feathers et al., 2008; Kunchithapautham et al., 2009). Surprisingly, we observed early RPE defects, including the loss of RPE65 expression in patches, RPE atrophy and loss of tight junctions between one and four months in Nrl−/− retina. We did not observe RPE65 in the whirls and rosettes as reported in one (Feathers et al., 2008) but not the other study (Wenzel et al., 2007). Thus, an absence of rod photoreceptors or secretion of a toxic product by excess cones appears to compromise the integrity of RPE. However, the remaining RPE65-positive RPE cells can renew the pool of 11-cis retinal and maintain some light sensitivity at high intensity stimulation. These observations can also be explained by the existence of a distinct visual pigment recycling pathway revealed in cone-rich retina of chicken and ground squirrels (Mata et al., 2002; Trevino et al., 2005). Indeed, Muller cells may contribute to maintenance of cone function in Nrl−/− retina and compensate for RPE defect. We also noticed that the absence of RPE did not lead to lack of PNA staining, suggesting that cones secrete their own extracellular matrix (Mieziewska et al., 1993). The early RPE defect in Nrl−/− retina might also indicate the existence of a rod-derived RPE survival factor.
In mouse models with no or short rod OS, such as Crx−/− (Furukawa et al., 1999) or Rho−/− mice (Humphries et al., 1997), rod photoreceptors die within weeks. In fact, all rods are lost in most mouse retinal degeneration mutants, independent of cell death kinetics (Punzo et al., 2009; Bramall et al., 2010). In Nrl−/− mice, however, though distinct cone OS are not detected by 4-mon, many cones survive for an extended period of time. We suggest that cone death in Nrl−/− mice is not intrinsic to photoreceptors but reflects an adaptive homeostatic mechanism. Rapid cell death allows the cone-only Nrl−/− retina to accommodate the steep requirement of oxygen (Perkins et al., 2003; Daniele et al., 2005; Shen et al., 2005) by surviving cones.
Prolonged survival of many cones (2–4 layers of nuclei even at 18-mon) in Nrl−/− retina, in the absence of rods since birth, demonstrates that rod photoreceptors are not essential for cone survival and may not be the sole source of trophic factors. For instance, Rdcvf, first identified as a rod-derived cone viability factor able to preserve cone function (Leveillard et al., 2004; Yang et al., 2009), is also expressed in the INL (Lambard et al.; Reichman et al.). In Nrl−/− retina, Rdcvf1 expression does not change with age though Rdcvf2 shows decreased expression between one and ten months (data not shown). Even if the numbers of nuclei are preserved at older ages, we observed a slight second decline after 8-mon in the ERG b-wave that may be attributed to cone aging, consistent with a small age-associated decline in rod and cone function in 12-mon old C57Bl/6 mouse retina (Gresh et al., 2003; Parapuram et al., 2010).
The higher concentration of acrolein and MDA in 4-mon old Nrl−/− retina corroborates the hypothesis that the average circulating current and oxygen demand in Nrl−/− retina should be less compared to WT due to the difference in a-wave amplitude generated by rod and cone photoreceptors (Daniele et al., 2005). Thus, oxygen stress in Nrl−/− mice should be higher, and hyperoxia may be toxic for photoreceptors. Interestingly, we observed attenuated retinal vasculature in older Nrl−/− retina, leading to a decrease in oxygen supply that could be beneficial for cones. This provides further support for the hypothesis that Nrl−/− retina are able to adapt and maintain a certain number of cones for prolonged period. The use of antioxidants in Nrl−/− mice might also reduce photoreceptor cell death, as described in several models of retinitis pigmentosa (Cao and Phillis, 1995; Kowluru and Odenbach, 2004; Komeima et al., 2006).
Retinal detachment, oxidative stress due to retinal hyperoxia (Stone et al., 1999), and activation of microglia cells (Gupta et al., 2003; Glybina et al., 2009) have been associated with RP and are observed in Nrl−/− retina. We suggest that these changes indicate a central role for rods in retinal homeostasis and not a direct impact on cone survival. As rods constitute almost 70% of cells in mouse and human retina, the loss of rods is expected to have a major influence on retinal architecture. Loss of rods compromises the fragile homeostasis in the retina of Nrl−/− mice, leading to RPE atrophy, Muller cell activation, altered retinal vasculature, and loss or dysfunction of other neuronal cell types. While additional investigations are necessary to delineate the underlying molecular pathways of cone survival, we suggest that functionally relevant contacts with RPE and trophic factors from Muller glia and microglia cells contribute in prolonged survival of cone photoreceptors even in the absence of rods. As Nrl−/− mouse exhibits many features of retinal remodeling in other retinal degenerations, our studies may provide new insights into pathogenic mechanisms of retinal neurodegenerative diseases and allow evaluation of alternative therapies.
Acknowledgments
These studies are supported by intramural program of the National Eye Institute. We thank Harsha Rajasimha for help with RNAseq data and Chun Y. Gao for technical assistance.
References
- Akhmedov NB, Piriev NI, Chang B, Rapoport AL, Hawes NL, Nishina PM, Nusinowitz S, Heckenlively JR, Roderick TH, Kozak CA, Danciger M, Davisson MT, Farber DB. A deletion in a photoreceptor-specific nuclear receptor mRNA causes retinal degeneration in the rd7 mouse. Proc Natl Acad Sci U S A. 2000;97:5551–5556. doi: 10.1073/pnas.97.10.5551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akimoto M, Cheng H, Zhu D, Brzezinski JA, Khanna R, Filippova E, Oh EC, Jing Y, Linares JL, Brooks M, Zareparsi S, Mears AJ, Hero A, Glaser T, Swaroop A. Targeting of GFP to newborn rods by Nrl promoter and temporal expression profiling of flow-sorted photoreceptors. Proc Natl Acad Sci U S A. 2006;103:3890–3895. doi: 10.1073/pnas.0508214103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernardos RL, Barthel LK, Meyers JR, Raymond PA. Late-stage neuronal progenitors in the retina are radial Muller glia that function as retinal stem cells. J Neurosci. 2007;27:7028–7040. doi: 10.1523/JNEUROSCI.1624-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bobu C, Craft CM, Masson-Pevet M, Hicks D. Photoreceptor organization and rhythmic phagocytosis in the nile rat Arvicanthis ansorgei: a novel diurnal rodent model for the study of cone pathophysiology. Invest Ophthalmol Vis Sci. 2006;47:3109–3118. doi: 10.1167/iovs.05-1397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bramall AN, Wright AF, Jacobson SG, McInnes RR. The genomic, biochemical, and cellular responses of the retina in inherited photoreceptor degenerations and prospects for the treatment of these disorders. Annu Rev Neurosci. 2010;33:441–472. doi: 10.1146/annurev-neuro-060909-153227. [DOI] [PubMed] [Google Scholar]
- Cao X, Phillis JW. The free radical scavenger, alpha-lipoic acid, protects against cerebral ischemia-reperfusion injury in gerbils. Free Radic Res. 1995;23:365–370. doi: 10.3109/10715769509065257. [DOI] [PubMed] [Google Scholar]
- Chen J, Nathans J. Genetic ablation of cone photoreceptors eliminates retinal folds in the retinal degeneration 7 (rd7) mouse. Invest Ophthalmol Vis Sci. 2007;48:2799–2805. doi: 10.1167/iovs.06-0922. [DOI] [PubMed] [Google Scholar]
- Cideciyan AV, Aleman TS, Jacobson SG, Khanna H, Sumaroka A, Aguirre GK, Schwartz SB, Windsor EA, He S, Chang B, Stone EM, Swaroop A. Centrosomal-ciliary gene CEP290/NPHP6 mutations result in blindness with unexpected sparing of photoreceptors and visual brain: implications for therapy of Leber congenital amaurosis. Hum Mutat. 2007;28:1074–1083. doi: 10.1002/humu.20565. [DOI] [PubMed] [Google Scholar]
- Curcio CA, Sloan KR, Kalina RE, Hendrickson AE. Human photoreceptor topography. J Comp Neurol. 1990;292:497–523. doi: 10.1002/cne.902920402. [DOI] [PubMed] [Google Scholar]
- Daniele LL, Lillo C, Lyubarsky AL, Nikonov SS, Philp N, Mears AJ, Swaroop A, Williams DS, Pugh EN., Jr Cone-like morphological, molecular, and electrophysiological features of the photoreceptors of the Nrl knockout mouse. Invest Ophthalmol Vis Sci. 2005;46:2156–2167. doi: 10.1167/iovs.04-1427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faktorovich EG, Steinberg RH, Yasumura D, Matthes MT, LaVail MM. Photoreceptor degeneration in inherited retinal dystrophy delayed by basic fibroblast growth factor. Nature. 1990;347:83–86. doi: 10.1038/347083a0. [DOI] [PubMed] [Google Scholar]
- Feathers KL, Lyubarsky AL, Khan NW, Teofilo K, Swaroop A, Williams DS, Pugh EN, Jr, Thompson DA. Nrl-knockout mice deficient in Rpe65 fail to synthesize 11-cis retinal and cone outer segments. Invest Ophthalmol Vis Sci. 2008;49:1126–1135. doi: 10.1167/iovs.07-1234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer AJ, Reh TA. Potential of Muller glia to become neurogenic retinal progenitor cells. Glia. 2003;43:70–76. doi: 10.1002/glia.10218. [DOI] [PubMed] [Google Scholar]
- Fisher SK, Lewis GP. Muller cell and neuronal remodeling in retinal detachment and reattachment and their potential consequences for visual recovery: a review and reconsideration of recent data. Vision Res. 2003;43:887–897. doi: 10.1016/s0042-6989(02)00680-6. [DOI] [PubMed] [Google Scholar]
- Fort PE, Lampi KJ. New focus on alpha-crystallins in retinal neurodegenerative diseases. Exp Eye Res. 2011;92:98–103. doi: 10.1016/j.exer.2010.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fort PE, Freeman WM, Losiewicz MK, Singh RS, Gardner TW. The retinal proteome in experimental diabetic retinopathy: up-regulation of crystallins and reversal by systemic and periocular insulin. Mol Cell Proteomics. 2009;8:767–779. doi: 10.1074/mcp.M800326-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furukawa T, Morrow EM, Li T, Davis FC, Cepko CL. Retinopathy and attenuated circadian entrainment in Crx-deficient mice. Nat Genet. 1999;23:466–470. doi: 10.1038/70591. [DOI] [PubMed] [Google Scholar]
- Gariano RF, Sage EH, Kaplan HJ, Hendrickson AE. Development of astrocytes and their relation to blood vessels in fetal monkey retina. Invest Ophthalmol Vis Sci. 1996;37:2367–2375. [PubMed] [Google Scholar]
- Glybina IV, Kennedy A, Ashton P, Abrams GW, Iezzi R. Photoreceptor neuroprotection in RCS rats via low-dose intravitreal sustained-delivery of fluocinolone acetonide. Invest Ophthalmol Vis Sci. 2009;50:4847–4857. doi: 10.1167/iovs.08-2831. [DOI] [PubMed] [Google Scholar]
- Gresh J, Goletz PW, Crouch RK, Rohrer B. Structure-function analysis of rods and cones in juvenile, adult, and aged C57bl/6 and Balb/c mice. Vis Neurosci. 2003;20:211–220. doi: 10.1017/s0952523803202108. [DOI] [PubMed] [Google Scholar]
- Guerin CJ, Anderson DH, Fariss RN, Fisher SK. Retinal reattachment of the primate macula. Photoreceptor recovery after short-term detachment. Invest Ophthalmol Vis Sci. 1989;30:1708–1725. [PubMed] [Google Scholar]
- Gupta N, Brown KE, Milam AH. Activated microglia in human retinitis pigmentosa, late-onset retinal degeneration, and age-related macular degeneration. Exp Eye Res. 2003;76:463–471. doi: 10.1016/s0014-4835(02)00332-9. [DOI] [PubMed] [Google Scholar]
- Hanisch UK, Kettenmann H. Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nat Neurosci. 2007;10:1387–1394. doi: 10.1038/nn1997. [DOI] [PubMed] [Google Scholar]
- Hendrickson A, Hicks D. Distribution and density of medium- and short-wavelength selective cones in the domestic pig retina. Exp Eye Res. 2002;74:435–444. doi: 10.1006/exer.2002.1181. [DOI] [PubMed] [Google Scholar]
- Humayun MS, Prince M, de Juan E, Jr, Barron Y, Moskowitz M, Klock IB, Milam AH. Morphometric analysis of the extramacular retina from postmortem eyes with retinitis pigmentosa. Invest Ophthalmol Vis Sci. 1999;40:143–148. [PubMed] [Google Scholar]
- Humphries MM, Rancourt D, Farrar GJ, Kenna P, Hazel M, Bush RA, Sieving PA, Sheils DM, McNally N, Creighton P, Erven A, Boros A, Gulya K, Capecchi MR, Humphries P. Retinopathy induced in mice by targeted disruption of the rhodopsin gene. Nat Genet. 1997;15:216–219. doi: 10.1038/ng0297-216. [DOI] [PubMed] [Google Scholar]
- Jackson GR, Owsley C, Curcio CA. Photoreceptor degeneration and dysfunction in aging and age-related maculopathy. Ageing Res Rev. 2002;1:381–396. doi: 10.1016/s1568-1637(02)00007-7. [DOI] [PubMed] [Google Scholar]
- Jacobson SG, Roman AJ, Aleman TS, Sumaroka A, Herrera W, Windsor EA, Atkinson LA, Schwartz SB, Steinberg JD, Cideciyan AV. Normal central retinal function and structure preserved in retinitis pigmentosa. Invest Ophthalmol Vis Sci. 2010;51:1079–1085. doi: 10.1167/iovs.09-4372. [DOI] [PubMed] [Google Scholar]
- John SK, Smith JE, Aguirre GD, Milam AH. Loss of cone molecular markers in rhodopsin-mutant human retinas with retinitis pigmentosa. Mol Vis. 2000;6:204–215. [PubMed] [Google Scholar]
- Jones BW, Marc RE. Retinal remodeling during retinal degeneration. Exp Eye Res. 2005;81:123–137. doi: 10.1016/j.exer.2005.03.006. [DOI] [PubMed] [Google Scholar]
- Karl MO, Hayes S, Nelson BR, Tan K, Buckingham B, Reh TA. Stimulation of neural regeneration in the mouse retina. Proc Natl Acad Sci U S A. 2008;105:19508–19513. doi: 10.1073/pnas.0807453105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Komeima K, Rogers BS, Lu L, Campochiaro PA. Antioxidants reduce cone cell death in a model of retinitis pigmentosa. Proc Natl Acad Sci U S A. 2006;103:11300–11305. doi: 10.1073/pnas.0604056103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kowluru RA, Odenbach S. Effect of long-term administration of alpha-lipoic acid on retinal capillary cell death and the development of retinopathy in diabetic rats. Diabetes. 2004;53:3233–3238. doi: 10.2337/diabetes.53.12.3233. [DOI] [PubMed] [Google Scholar]
- Kryger Z, Galli-Resta L, Jacobs GH, Reese BE. The topography of rod and cone photoreceptors in the retina of the ground squirrel. Vis Neurosci. 1998;15:685–691. doi: 10.1017/s0952523898154081. [DOI] [PubMed] [Google Scholar]
- Kumar PA, Haseeb A, Suryanarayana P, Ehtesham NZ, Reddy GB. Elevated expression of alphaA- and alphaB-crystallins in streptozotocin-induced diabetic rat. Arch Biochem Biophys. 2005;444:77–83. doi: 10.1016/j.abb.2005.09.021. [DOI] [PubMed] [Google Scholar]
- Kunchithapautham K, Coughlin B, Crouch RK, Rohrer B. Cone outer segment morphology and cone function in the Rpe65−/− Nrl−/− mouse retina are amenable to retinoid replacement. Invest Ophthalmol Vis Sci. 2009;50:4858–4864. doi: 10.1167/iovs.08-3008. [DOI] [PubMed] [Google Scholar]
- Lambard S, Reichman S, Berlinicke C, Niepon ML, Goureau O, Sahel JA, Leveillard T, Zack DJ. Expression of rod-derived cone viability factor: dual role of CRX in regulating promoter activity and cell-type specificity. PLoS One. 2010;5:e13075. doi: 10.1371/journal.pone.0013075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langmann T. Microglia activation in retinal degeneration. J Leukoc Biol. 2007;81:1345–1351. doi: 10.1189/jlb.0207114. [DOI] [PubMed] [Google Scholar]
- Leveillard T, Mohand-Said S, Lorentz O, Hicks D, Fintz AC, Clerin E, Simonutti M, Forster V, Cavusoglu N, Chalmel F, Dolle P, Poch O, Lambrou G, Sahel JA. Identification and characterization of rod-derived cone viability factor. Nat Genet. 2004;36:755–759. doi: 10.1038/ng1386. [DOI] [PubMed] [Google Scholar]
- Lewis GP, Linberg KA, Fisher SK. Neurite outgrowth from bipolar and horizontal cells after experimental retinal detachment. Invest Ophthalmol Vis Sci. 1998;39:424–434. [PubMed] [Google Scholar]
- Lewis GP, Chapin EA, Luna G, Linberg KA, Fisher SK. The fate of Muller’s glia following experimental retinal detachment: nuclear migration, cell division, and subretinal glial scar formation. Mol Vis. 2010;16:1361–1372. [PMC free article] [PubMed] [Google Scholar]
- Lu F, Zhao P, Fan Y, Tang S, Hu J, Liu X, Yang X, Chen Y, Li T, Lei C, Yang J, Lin Y, Ma S, Li C, Shi Y, Yang Z. An association study of SERPING1 gene and age-related macular degeneration in a Han Chinese population. Mol Vis. 2010;16:1–6. [PMC free article] [PubMed] [Google Scholar]
- Luo DG, Xue T, Yau KW. How vision begins: an odyssey. Proc Natl Acad Sci U S A. 2008;105:9855–9862. doi: 10.1073/pnas.0708405105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marc RE, Jones BW, Watt CB, Strettoi E. Neural remodeling in retinal degeneration. Prog Retin Eye Res. 2003;22:607–655. doi: 10.1016/s1350-9462(03)00039-9. [DOI] [PubMed] [Google Scholar]
- Marioni JC, Mason CE, Mane SM, Stephens M, Gilad Y. RNA-seq: an assessment of technical reproducibility and comparison with gene expression arrays. Genome Res. 2008;18:1509–1517. doi: 10.1101/gr.079558.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mata NL, Radu RA, Clemmons RC, Travis GH. Isomerization and oxidation of vitamin a in cone-dominant retinas: a novel pathway for visual-pigment regeneration in daylight. Neuron. 2002;36:69–80. doi: 10.1016/s0896-6273(02)00912-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mears AJ, Kondo M, Swain PK, Takada Y, Bush RA, Saunders TL, Sieving PA, Swaroop A. Nrl is required for rod photoreceptor development. Nat Genet. 2001;29:447–452. doi: 10.1038/ng774. [DOI] [PubMed] [Google Scholar]
- Mieziewska K, Van Veen T, Aguirre GD. Development and fate of interphotoreceptor matrix components during dysplastic photoreceptor differentiation: a lectin cytochemical study of rod-cone dysplasia 1. Exp Eye Res. 1993;56:429–441. doi: 10.1006/exer.1993.1056. [DOI] [PubMed] [Google Scholar]
- Milam AH, Li ZY, Fariss RN. Histopathology of the human retina in retinitis pigmentosa. Prog Retin Eye Res. 1998;17:175–205. doi: 10.1016/s1350-9462(97)00012-8. [DOI] [PubMed] [Google Scholar]
- Mullins RF, Faidley EA, Daggett HT, Jomary C, Lotery AJ, Stone EM. Localization of complement 1 inhibitor (C1INH/SERPING1) in human eyes with age-related macular degeneration. Experimental eye research. 2009;89:767–773. doi: 10.1016/j.exer.2009.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mustafi D, Engel AH, Palczewski K. Structure of cone photoreceptors. Prog Retin Eye Res. 2009;28:289–302. doi: 10.1016/j.preteyeres.2009.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mustafi D, Kevany BM, Genoud C, Okano K, Cideciyan AV, Sumaroka A, Roman AJ, Jacobson SG, Engel A, Adams MD, Palczewski K. Defective photoreceptor phagocytosis in a mouse model of enhanced S-cone syndrome causes progressive retinal degeneration. Faseb J. 2011;25:3157–3176. doi: 10.1096/fj.11-186767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nikonov SS, Daniele LL, Zhu X, Craft CM, Swaroop A, Pugh EN., Jr Photoreceptors of Nrl−/− mice coexpress functional S- and M-cone opsins having distinct inactivation mechanisms. J Gen Physiol. 2005;125:287–304. doi: 10.1085/jgp.200409208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh EC, Khan N, Novelli E, Khanna H, Strettoi E, Swaroop A. Transformation of cone precursors to functional rod photoreceptors by bZIP transcription factor NRL. Proc Natl Acad Sci U S A. 2007;104:1679–1684. doi: 10.1073/pnas.0605934104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh EC, Cheng H, Hao H, Jia L, Khan NW, Swaroop A. Rod differentiation factor NRL activates the expression of nuclear receptor NR2E3 to suppress the development of cone photoreceptors. Brain Res. 2008;1236:16–29. doi: 10.1016/j.brainres.2008.01.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pang JJ, Chang B, Hawes NL, Hurd RE, Davisson MT, Li J, Noorwez SM, Malhotra R, McDowell JH, Kaushal S, Hauswirth WW, Nusinowitz S, Thompson DA, Heckenlively JR. Retinal degeneration 12 (rd12): a new, spontaneously arising mouse model for human Leber congenital amaurosis (LCA) Mol Vis. 2005;11:152–162. [PubMed] [Google Scholar]
- Paques M, Guyomard JL, Simonutti M, Roux MJ, Picaud S, Legargasson JF, Sahel JA. Panretinal, high-resolution color photography of the mouse fundus. Invest Ophthalmol Vis Sci. 2007;48:2769–2774. doi: 10.1167/iovs.06-1099. [DOI] [PubMed] [Google Scholar]
- Parapuram SK, Cojocaru RI, Chang JR, Khanna R, Brooks M, Othman M, Zareparsi S, Khan NW, Gotoh N, Cogliati T, Swaroop A. Distinct signature of altered homeostasis in aging rod photoreceptors: implications for retinal diseases. PLoS One. 2010;5:e13885. doi: 10.1371/journal.pone.0013885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng YW, Hao Y, Petters RM, Wong F. Ectopic synaptogenesis in the mammalian retina caused by rod photoreceptor-specific mutations. Nat Neurosci. 2000;3:1121–1127. doi: 10.1038/80639. [DOI] [PubMed] [Google Scholar]
- Perkins GA, Ellisman MH, Fox DA. Three-dimensional analysis of mouse rod and cone mitochondrial cristae architecture: bioenergetic and functional implications. Mol Vis. 2003;9:60–73. [PubMed] [Google Scholar]
- Punzo C, Kornacker K, Cepko CL. Stimulation of the insulin/mTOR pathway delays cone death in a mouse model of retinitis pigmentosa. Nat Neurosci. 2009;12:44–52. doi: 10.1038/nn.2234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao NA, Saraswathy S, Wu GS, Katselis GS, Wawrousek EF, Bhat S. Elevated retina-specific expression of the small heat shock protein, alphaA-crystallin, is associated with photoreceptor protection in experimental uveitis. Invest Ophthalmol Vis Sci. 2008;49:1161–1171. doi: 10.1167/iovs.07-1259. [DOI] [PubMed] [Google Scholar]
- Raven MA, Oh EC, Swaroop A, Reese BE. Afferent control of horizontal cell morphology revealed by genetic respecification of rods and cones. J Neurosci. 2007;27:3540–3547. doi: 10.1523/JNEUROSCI.0372-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reichman S, Kalathur RK, Lambard S, Ait-Ali N, Yang Y, Lardenois A, Ripp R, Poch O, Zack DJ, Sahel JA, Leveillard T. The homeobox gene CHX10/VSX2 regulates RdCVF promoter activity in the inner retina. Hum Mol Genet. 2010;19:250–261. doi: 10.1093/hmg/ddp484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rex TS, Lewis GP, Geller SF, Fisher SK. Differential expression of cone opsin mRNA levels following experimental retinal detachment and reattachment. Mol Vis. 2002;8:114–118. [PubMed] [Google Scholar]
- Roger J, Brajeul V, Thomasseau S, Hienola A, Sahel JA, Guillonneau X, Goureau O. Involvement of Pleiotrophin in CNTF-mediated differentiation of the late retinal progenitor cells. Dev Biol. 2006;298:527–539. doi: 10.1016/j.ydbio.2006.07.003. [DOI] [PubMed] [Google Scholar]
- Santos A, Humayun MS, de Juan E, Jr, Greenburg RJ, Marsh MJ, Klock IB, Milam AH. Preservation of the inner retina in retinitis pigmentosa. A morphometric analysis. Arch Ophthalmol. 1997;115:511–515. doi: 10.1001/archopht.1997.01100150513011. [DOI] [PubMed] [Google Scholar]
- Shen J, Yang X, Dong A, Petters RM, Peng YW, Wong F, Campochiaro PA. Oxidative damage is a potential cause of cone cell death in retinitis pigmentosa. J Cell Physiol. 2005;203:457–464. doi: 10.1002/jcp.20346. [DOI] [PubMed] [Google Scholar]
- Sparrow JR, Hicks D, Hamel CP. The retinal pigment epithelium in health and disease. Curr Mol Med. 2010;10:802–823. doi: 10.2174/156652410793937813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stone J, Maslim J, Valter-Kocsi K, Mervin K, Bowers F, Chu Y, Barnett N, Provis J, Lewis G, Fisher SK, Bisti S, Gargini C, Cervetto L, Merin S, Peer J. Mechanisms of photoreceptor death and survival in mammalian retina. Prog Retin Eye Res. 1999;18:689–735. doi: 10.1016/s1350-9462(98)00032-9. [DOI] [PubMed] [Google Scholar]
- Stone JL, Barlow WE, Humayun MS, de Juan E, Jr, Milam AH. Morphometric analysis of macular photoreceptors and ganglion cells in retinas with retinitis pigmentosa. Arch Ophthalmol. 1992;110:1634–1639. doi: 10.1001/archopht.1992.01080230134038. [DOI] [PubMed] [Google Scholar]
- Strauss O. The retinal pigment epithelium in visual function. Physiol Rev. 2005;85:845–881. doi: 10.1152/physrev.00021.2004. [DOI] [PubMed] [Google Scholar]
- Strettoi E, Mears AJ, Swaroop A. Recruitment of the rod pathway by cones in the absence of rods. J Neurosci. 2004;24:7576–7582. doi: 10.1523/JNEUROSCI.2245-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swaroop A, Kim D, Forrest D. Transcriptional regulation of photoreceptor development and homeostasis in the mammalian retina. Nat Rev Neurosci. 2010;11:563–576. doi: 10.1038/nrn2880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trevino SG, Villazana-Espinoza ET, Muniz A, Tsin AT. Retinoid cycles in the cone-dominated chicken retina. J Exp Biol. 2005;208:4151–4157. doi: 10.1242/jeb.01881. [DOI] [PubMed] [Google Scholar]
- Umeda S, Suzuki MT, Okamoto H, Ono F, Mizota A, Terao K, Yoshikawa Y, Tanaka Y, Iwata T. Molecular composition of drusen and possible involvement of anti-retinal autoimmunity in two different forms of macular degeneration in cynomolgus monkey (Macaca fascicularis) Faseb J. 2005;19:1683–1685. doi: 10.1096/fj.04-3525fje. [DOI] [PubMed] [Google Scholar]
- Wenzel A, von Lintig J, Oberhauser V, Tanimoto N, Grimm C, Seeliger MW. RPE65 is essential for the function of cone photoreceptors in NRL-deficient mice. Invest Ophthalmol Vis Sci. 2007;48:534–542. doi: 10.1167/iovs.06-0652. [DOI] [PubMed] [Google Scholar]
- West H, Richardson WD, Fruttiger M. Stabilization of the retinal vascular network by reciprocal feedback between blood vessels and astrocytes. Development. 2005;132:1855–1862. doi: 10.1242/dev.01732. [DOI] [PubMed] [Google Scholar]
- Wright AF, Chakarova CF, Abd El-Aziz MM, Bhattacharya SS. Photoreceptor degeneration: genetic and mechanistic dissection of a complex trait. Nat Rev Genet. 2010;11:273–284. doi: 10.1038/nrg2717. [DOI] [PubMed] [Google Scholar]
- Yang Y, Mohand-Said S, Danan A, Simonutti M, Fontaine V, Clerin E, Picaud S, Leveillard T, Sahel JA. Functional cone rescue by RdCVF protein in a dominant model of retinitis pigmentosa. Mol Ther. 2009;17:787–795. doi: 10.1038/mt.2009.28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida S, Mears AJ, Friedman JS, Carter T, He S, Oh E, Jing Y, Farjo R, Fleury G, Barlow C, Hero AO, Swaroop A. Expression profiling of the developing and mature Nrl−/− mouse retina: identification of retinal disease candidates and transcriptional regulatory targets of Nrl. Hum Mol Genet. 2004;13:1487–1503. doi: 10.1093/hmg/ddh160. [DOI] [PubMed] [Google Scholar]
- Zhu X, Brown B, Li A, Mears AJ, Swaroop A, Craft CM. GRK1-dependent phosphorylation of S and M opsins and their binding to cone arrestin during cone phototransduction in the mouse retina. J Neurosci. 2003;23:6152–6160. doi: 10.1523/JNEUROSCI.23-14-06152.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]