

Abstract
With a constellation of stem cell sources available, researchers hope to utilize their potential for cellular repair as a therapeutic target for disease. However, many lab-to-clinic translational considerations must be given in determining their efficacy, variables such as the host response, effects on native tissue, and potential for generating tumors. This review will discuss the current knowledge of stem cell research in neurological disease, mainly stroke, with a focus on the benefits, limitations, and clinical potential.
Keywords: stem cells, stroke, transplantation, translational biomedical research
1. Translational Gating Items of Stem Cell Therapy
With the increasing diversity of stem cell sources emerging for donor cells in transplantation therapy, many laboratory-to-clinic translational factors must first be considered, dynamics such as the source of the cells, ease of extraction, immunogenicity, capacity for proliferation, and cell yield. These concerns may serve as potential limitations respective to the donor cell origin being considered, proving a particular source to be a more suitable therapy for a specific disease.
Harvesting of stem cells may be divided into two domains, allogenic vs. autologous sources (though xenogeneic cells have been previously tested). Autologous stem cells are acquired from the host in which the cells are intended for use, while allogenic cells are procured from an unrelated donor prior to transplantation. As one may expect, the use of allogenic stem cells may predispose an individual to various immunologic complications upon treatment, giving rise to the significant limitation of graft rejection with this method of treatment. Yet, autologous treatments may be limited by their ability for propagation and cell yield.
The immunological barriers, such as graft vs. host or required immunosuppression of the host, are of constant consideration in the therapeutic benefits and limitations of stem cell transplantation. Prior research of immunocompromised stroke animals demonstrated inhibition neurogenesis in the cortex endogenously via a CD4+ T cell, but not CD25+ T Cell mechanism, supporting the influence of immunodeficiency in reducing stem cell apoptosis [1]. In another study, upon exposure to cyclosporine A, an immunosuppressant, there was enhanced recovery of cortical injury following stroke secondary to endogenous stem cell activity and migration [2]. These papers support the hypothesis that inflammation following a cerebral event may not only damage tissue, but also further disrupt the endogenous neurogenic pathways of repair involving the migration of stem cells from the lateral ventricle regions of the brain, a topic further discussed in the neural stem cell section. This contrasts the proposed mechanism of action for neurotrophic modulators that are believed to furnish a microenvironment conducive to repair, but do not yield new neurons [2].
Because of the potential immunological host response to the transplanted cells, much research has been conducted investigating specific cell line's prospective immunogenicity. Surmounting evidence suggests that the more naive the cell lineage, the less likely the incidence of immunological reaction following transplantation. For instance, umbilical cord blood, due to its immunological immaturity, is less likely to invoke an immunological response and therefore less likely to require immunosuppression. More so, human leukocyte antigen (HLA) matching may be less stringent in umbilical cord blood transplants compared to bone marrow derived transplants and even whole tissue grafts, leading to higher cell viability [3]. Contrasting immunogenicity, some cell lineages may be more immunosuppressive than bone marrow derived stem cells. HLA-G, a contributing factor of immunosuppression [4], is of higher expression in chorionic plate-derived mesenchymal stem cells compared to bone marrow-derived stem cells and adipocyte tissue-derived mesenchymal stem cells [5], suggesting its usefulness as a prognostic indicator of transplant viability in the presence of host immunity [6]. Additionally, placenta-derived mesenchymal stem cells demonstrate less immunomodulation than bone marrow-derived mesenchymal stem cells, suggesting regenerative transplantation potential to be less efficacious [7].
2. Tailoring Stem Cells for Therapeutic Applications in Stroke
Stroke is a major unmet clinical need with only one current Food and Drug Administration (FDA)-approved drug, the tissue plasminogen activator, efficacy limited to 4.5 h after stroke onset. Accordingly, the challenges of proliferation capacity and cell yield are evident with regards to the optimum delivery time of stem cell therapy. With the current clinical trials of cell therapy for acute stroke mostly targeting a window of 48 h, the potential limitation in generating a sufficient number of autologous cells from freshly harvested tissue for therapy in such a short period is apparent [8]. In view of this limitation, allogeneic transplantation is indicated when contemplating with acute stroke therapy. Alternatively, the extended time required for cell amplification with autologous stem cell transplantation renders it more appropriate for chronic stroke therapy. Nonetheless, regardless of autologous or allogeneic stem cell sources, cell harvesting imparts additional technical challenges. For example, acquiring neural stem cells may require invasive procedures that may be disadvantageous despite the therapeutic potential of the cells.
The following sections aim to outline the different tissue sources available for harvesting stem cells, along with their respective benefits, limitations, and prospective use in clinical application as they pertain mainly to neurological diseases, most notably stroke therapy. A review of current stem cells being investigated in neurorestoration has recently been published [9]. We only briefly discuss embryonic and extraembryonic stem cells and focus this paper on our long-standing research interest in adult stem cells.
3. Searching for Safe and Effective Stem Cell Therapy
Embryonic stem cells are pluripotent cells derived from the inner cell mass of the blastocyst that arguably serve as the foundation by which the properties of “stemness” are measured in other cell lines. With the potential to differentiate into all three germ layers, transplantation of embryonic stem cells (ESCs) into animal stroke models has demonstrated repair in both vascular [10] and neuronal damage [11], improved functional recovery of deficits [12–16] and provision of neurotrophic, angiogenic, and anti-apoptotic effects [13–17]. These benefits may extend well into potential translational therapy following a cerebral event, giving the cell line a plentiful array of therapeutic actions in terms of modulating a number of diseases including stroke. The distribution of these cells has been demonstrated with imaging techniques in both the brain and the periphery following transplantation in animal stroke models [18,19]. Although ESCs possess the potential for vast differentiation, there are two predominant concerns limiting their use. The ethics of harvesting embryonic stem cells is widely debated. However, recent advancements, discussed later in this review, may further develop and refine methods for producing these cells through retrograde manipulation of mature cell lines, alleviating some of the tension surrounding their use. Additionally, their naive lineage aligns with the stem cell tenet of the more naive the cell, the greater the potential for tumorigenicity, a topic that will be expanded upon in subsequent sections. Stem cells have the potential to form tumors after transplantation. This tumorigenicity is mostly associated with embryonic stem cells and pluripotent stem (iPS) cells [20]. Shortly after transplantation, a dysregulated differentiation of ESCs was found to cause the formation of teratomas containing all types of somatic tissues of the early embryo [21], due likely to the presence of oncogenes and trisomies which are known to have roles in cancer cell formation [22]. In view of this stem cell tumorigenicity, strategies have been explored including redifferentiation of cells to remove stemness or genetic modification to activate anti-oncogenic genes such as Nurr1 to abrogate the neoplastic state or render the cells post-mitotic [23,24]. While this tumorigenicity has been closely associated with ESCs and iPS cells, recent evidence suggests that safety precautions should also be taken with adult stem cells due to possible ectopic tissue formation seen in grafted mesenchymal stem cells (MSCs) in the Central Nervous System (CNS) [25]. These studies highlight a major hurdle in stem cell therapy, and emphasize the importance of closely monitoring stemness and tumorigenicity as we translate cell therapy to the clinic [22,24,26].
With the concerns surrounding the use of embryonic tissue to harvest stem cells, researchers have looked to sources external to the embryo to harvest stem cells. Wharton's jelly (within the umbilical cord), amnion, placenta, and umbilical cord are all rich stem cell sources [27]. As seen with the neural stem cells and mesenchymal stromal cells, extraembryonic stem cells also relate to different germinal layers. The amniotic epithelium is of ectoderm origin while the amnion-derived mesenchymal stromal cells are derived from the mesoderm [28]. Yet, the amnion-derived mesenchymal stromal cells exhibit less endothelial propensity conferring cell specificity [29]. Transplantation of placenta-derived mesenchymal stromal cells in animal models of stroke are believed to supply a microenvironment favorable for endogenous neural repair and replace damaged tissue [30–32]. Mesenchymal stromal cells derived from umbilical cord lining develop an immunosuppressive effect while demonstrating functional recovery, increased vascular density, increased expression of vascular endothelial growth factor, and basic fibroblast growth factor in rat stroke models [33,34].
Adult stem cells often exist in combination with non-stem cells committed to distinct lineage, creating a heterogeneous environment. Because of this, one challenge in the use of adult-derived stem cells is the purification for the isolation for the particular stem cell of interest. In the following sections we will discuss the variety of adult-derived stem cells (Figure 1), their tissue sources, benefits, limitations, and clinical relevance.
Figure 1.
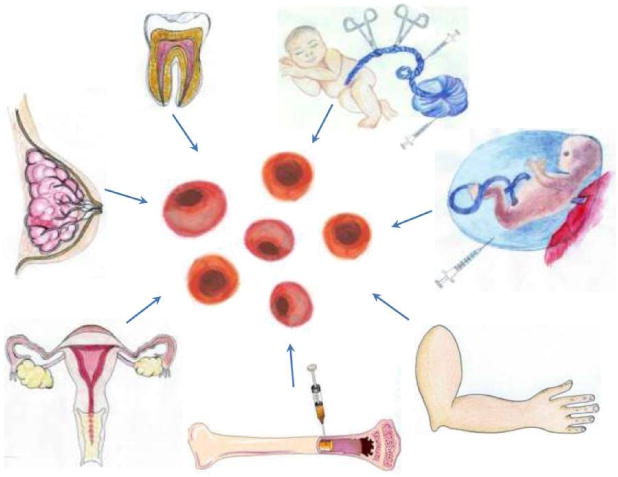
Adult stem sources include umbilical cord blood, placenta, amniotic fluid, bone marrow, menstrual blood, breast milk, dental pup, and skin fibroblasts. Most of these cells have been shown to exert neuroprotective effects in stroke animal models and a few have reached clinical trials in stroke patients.
3.1. A Stem Cell Source with a Long History of Transplant Use: Bone Marrow-Derived Stem Cells
A divergent population stem and blast cells constitute the bone marrow. Thus, these cells may be utilized as an admixture or purified upon harvesting. Emerging research demonstrates the ability of bone marrow-derived stem cells, upon injury, to mobilize from the bone marrow (BM) into the peripheral blood. This feature is very practical for harvesting cells, which is currently employed for many immunologic, hematologic, and oncologic clinical applications. With relation to stroke, once in systemic circulation they may migrate to regions of the central nervous system in response to neuronal injury [35]. Cellular components of bone marrow include: hematopoietic stem cells (HSCs), mesenchymal stem cells (MSCs), endothelial progenitor cells (EPCs), and very small embryonic-like stem cells (VSELs) [36]. Here we will outline the aforementioned cell lines in greater detail.
Hematopoietic stem cells are found primarily in the bone marrow where they give rise to both the myeloid and lymphoid lineages of blood cells. Cytokines produced by the CNS can incite hematopoietic stem cell mobilization into the blood, from the marrow, in response to a cerebrovascular accident (CVA) [37–40]. This mobilization may also be influenced by neurotransmitters, notably catecholamines, either through a paracrine mechanism signaling directly into the bone marrow or through systemic sympathetic release into circulation [41]. This cytokine-mediated recruitment of HSCs is applied clinically through treatment with granulocyte-colony stimulating factor [39,40].
Abundant mobilization of immature hematopoietic CD34+ colony-forming cells and Long-Term Culture-Initiating Cells (LTC-IC) has been observed from clinical data of acute stroke, with the magnitude of this mobilization correlating with recovery of function. Autologous infusions of bone marrow mononuclear cells in human stroke patients during acute, subacute, and chronic phase of stroke have demonstrated no adverse effects of transplantation [42–45]. Transplantation of HSCs into animal models of stroke has greatly elucidated the therapeutic benefits of this type of stem cell in regenerative medicine. HSCs intravenously administered were able to increase the survival rate of stroke mice, accompanied by decreased neuronal cell death, and facilitated recovery from paralysis and forelimb weakness [46]. In an effort to reveal the mechanism of action of HSC therapeutic benefit in stroke, HSCs were intravenously administered at 24 h after ischemic stroke in mice which showed grafted cell migration into the spleen and later into ischemic brain parenchyma, expressing microglial but no neural marker proteins [47]. Moreover, transplanted stroke animals displayed significantly smaller infarct volumes and less apoptotic neuronal cell death in peri-infarct areas accompanied by a reduction of invading T cells and macrophages and a downregulated proinflammatory cytokine and chemokine receptor gene transcription within the spleen [47]. These findings indicate that transplanted HSCs exert therapeutic effects in stroke possibly acting via regulation of both central and peripheral (i.e., spleen) inflammation. Bone marrow (BM) derived HSCs are also being considered as potential treatment for diseases affecting cardiovascular tissue, bone, and cartilage among other tissues.
Mesenchymal stromal cells were first isolated in bone marrow, but have since been found in nearly every tissue of the body. Here we address the therapeutic application of mesenchymal stromal cells, as well as non-bone marrow derived stem cells, for treatment of stroke.
Transplantation of mesenchymal stromal cells into stroke models induces functional recovery of neurological deficits following cerebral ischemia [48–50]. The limited differentiation capacity of mesenchymal stromal cells suggests that observed transplantation benefits may be afforded through activation of endogenous repair pathways by secretion of neurotrophic factors which include brain-derived neurotrophic factor (BDNF) [51], nerve growth factor (NGF) [51], vascular endothelial growth factor (VEGF) [52], basic fibroblast growth factor (bFGF, FGF-2) [52], hepatocyte growth factor (HGF) [48,53], and insulin growth factor-1 (IGF-1) [54]. MSCs may recruit primary stem cells from the subventricular and subgranular zones of the brain to the site of injury, while also dampening apoptosis in the penumbral zone of the lesion [51,52]. A clinical trial of intravenous infusion of autologous BM-derived mesenchymal stromal cells in ischemic stroke patients shows significant functional improvement in infused patients without adverse effects in comparison with non-infused patients [55]. In a long-term 5 year follow up, patients infused with mesenchymal stromal cells demonstrated increased survival rates and greater functional improvement compared to non-infused patients [56]. However, the current transplantation techniques are plagued by very low graft survival rates, and therefore, mode of delivery remains a significant limitation of mesenchymal stromal cell-based therapies for stroke [57].
Mesenchymal stem cells have recently been the focus of many research endeavors, due in large part to their accessibility when compared to other stem cells. Mesoderm-derived mesenchymal stromal cells may be extracted from almost any mesenchymal tissue of the body including: bone marrow, placenta, teeth and adipose tissue. This abundance of harvest sites makes MSCs a preferred line for autologous transplantation. However, evidence indicates that harvest location may impart a specific role to mesenchymal stromal cells as a function of various methods of extraction, isolation, and proliferation [58–62]. To this extent, one site of tissue derived mesenchymal stromal cells may be better qualified for a specific therapy than cells derived from another tissue site.
Despite their limited differentiation capacity and relatively transient life-span after transplantation, evidence shows that mesenchymal stromal cells promote neurogenesis following ischemic injury [52]. As mentioned previously, benefits may stem from secretion of neurotrophic factors such as BDNF and β-NGF, as well as modulation of vasculature from bone marrow, adipose tissue, skeletal muscle, and myocardium [63].
Laboratory findings indicate that neurotrophic factors are involved in the neuroprotective action of stem cells as evidenced by their trophic effects, but additionally their anti-inflammatory and anti-apoptotic effects in animal models of stroke and other neurological disorders [64–66]. That BDNF and NGF have been shown as consistently secreted by transplanted stem cells suggest that targeting these two trophic factors' signaling pathway may further improve the outcome of stem cell therapy. Alternatively, the combination of trophic factor treatment with stem cell transplantation may allow a more robust functional improvement in the clinic. The safety profiles of these trophic factors in the clinic in other disease indications [67,68], and the recognition of their clinical limitations, will guide the clinical trials of this combination therapy for stroke. Of note, BDNF Val(66)Met polymorphism has been implicated in worsened functional outcome in patients with subcortical stroke [69]. Similarly, serum from stroke patients revealed NGF upregulation significantly correlates with clinical and neuroradiological parameters of brain injury [70].
In addition to the many potential benefits of menchymal strem cell-based therapies mentioned above, there are also significant risk factors that must be addressed. As with many types of stem cells, the risk of mesenchymal stem cells developing into tumors must be considered. One study showed that a sarcoma developed in the lungs of mice following transplantation of mesenchymal stem cells [71]. Not only the cells themselves but also their secretions may affect tumors. Interleukin-6 (IL-6) and vascular endothelial growth factor (VEGF) secreted from MSCs increases the migration of breast cancer cell lines [72]. Breast cancer cells stimulate de novo secretion of the chemokine CCL5 from mesenchymal stem cells, which then acts in a paracrine fashion on the cancer cells to enhance their motility, invasion, and metastasis [73]. Accordingly, certain types of mesenchymal stem cells may demonstrate greater tendency toward tumorigenicity and promotion of metastasis.
Endothelial progenitor cells (EPCs) represent a small population of cells present in the blood that give rise to mature endothelium that lines blood vessels. While in circulation, these cells can be recruited to produce new blood vessels, a term called vasculogenesis.
The etiology of stroke is multifaceted. One contributing factor includes the compromise of vascular integrity, leaving a region vulnerable to stroke. With the endothelium regulating the permeability of the blood brain barrier (BBB), the role of endothelial progenitor cells in producing the mature lining of blood vessels is integral in maintaining cerebral homeostasis. Preliminary studies demonstrated that transplanted EPCs were integrated into newly vascularized endothelium of the hind limbs in ischemic animal models [74]. Further research specifies that BM-derived endothelial progenitor cells are likely signaled to sites of new vascularization prior to differentiation [75,76].
A correlational study in human ischemic stroke patients indicates that the level of circulating EPCs relates to improvement on the National Institute of Health Stroke Scale [77]. Animal models of stroke show that intravenous transplantation of EPCs reduces cerebral infarcts in stroke diabetic mice [78]. Moreover, EPCs can incorporate to the BBB microvasculature and delay the stroke onset in an ischemic hemorrhagic stroke model [79]. In addition, intravenous infusion of autologous EPCs after stroke in rabbits produces functional improvement, decreases number of apoptotic cells, increases microvessel density in the ischemic boundary area, and reduces infarct area [80].
The current hypothesis of very small embryonic-like stem cells is that these pluripotent stem cells are deposited early in embryonic development from an epiblast source, where they function as a reserve that can be accessed in response to physiological stress [81,82]. Investigation is underway using VSELs for stroke therapy in the brain, a region rich in VSEL phenotypic cells [83,84]. VSELs are a great candidate in therapy for cerebral vascular incident because of their potential to differentiate into neurons, oligodendrocytes, and microglia to regenerate damaged CNS [35].
However, current restrictions present a challenge in moving forward. Very small embryonic-like stem cells are present in limited quantity, producing a low yield from harvesting. Such an obstacle may be overcome with refining methods of proliferation prior to transplant [35]. An additional challenge is the decreasing population of VSELs present in older age, further contributing to the difficulty of sufficient yield upon harvesting [84].
3.2. Harvesting Neural Stem Cells for Neural Repair in Stroke
With endogenous stem cells being located in the subgranular zone (SGZ) of the dentate gyrus, the subventricular zone (SVZ), and the subependymal zone (SEZ) of the spinal cord, the therapeutic potential of NSCs for cerebrovascular accidents seems obvious. Chemokine signals such as stromal-derived factor-1 (SDF-1), vascular endothelial growth factor (VEGF), and angiopoietin are released from ischemic tissue, influencing the course of the SVZ NSCs toward a path along blood vessels to reach the infarcted area [85–88]. Although endogenous stem cells migrate to the lesion following stroke, there appears to be minimal stem cell survival [89–91]. This supports the hypothesis that endogenous neural stem cells may not exert their effects solely by replacement of neuronal tissue, but rather by secreting growth factors that influence repair. Immunological responses may also influence the differentiation of endogenous stem cells. In ex vivo studies, microglia from ischemic brains prompted the maturation of NSCs into neurons [92].
Although endogenous NSCs are shown to migrate in response to cellular injury, their effects may be augmented by the addition of exogenous neural stem cells. The literature describes transplantation of NSCs inducing further endogenous stem cell production at the site of injury [93–96]. However, another study suggests that intravenous infusion of neural progenitor cells decreased neurogenesis despite increasing dendritic length and the number of branch points [97]. This may further support the hypothesis of neurotrophic factors secreted from stem cells exerting a primary effect.
Neural stem cells are proven in terms of their therapeutic potential; however, they present a few significant limitations. The difficulty of obtaining the cells may be the greatest challenge. Under most circumstances, harvesting neural stem cells would require an invasive procedure for autologous use while allogenic grafts would require a fetal source or manipulation from another cell source. A possibility to circumvent this problem would be harvesting the stem cells for therapy during a surgical procedure already intended for the patient [98], such as during a temporal lobe resection in which subventricular matter, a known source of stem cells, could be harvested. As with many other stem cells, there is constant concern about the potential to illicit aberrant cell growth, producing tumors upon transplantation. Whereas the less differentiated the cell, the less likely it will invoke a host reaction; however, the more naive the stem cell, the greater its propensity for uncontrolled proliferation.
Adult stem cells possess a reduced capacity for proliferation and may be less tumorigenic. However, this presents a problem with producing a sufficient number of cells for transplantation. To traverse these limitations, researchers have developed methods such as: long-term culturing, immortalization, insertion of oncogenes, or even derivation of neural stem cells from other tissues or from pluripotent stem cells.
3.3. Recapitulating Cell Developmental Growth in Other Adult Stem Cells
3.3.1. Mimicking Bone Marrow Therapeutic Transplant Potential: Umbilical Cord Blood (UCB)
With their availability, ease of harvesting, and ability for autologous and allogenic use, the therapeutic potential of umbilical cord blood is expansive. The heterogeneous mixture of cells comprising cord blood includes hematopoietic progenitors, lymphocytes, monocytes, embryonic-like stem cells, and mecenchymal stromal cells. Yet, cord blood is considered immunologically immature and exerts its effects through immune modulation and reducing inflammation [99].
Transplantation of umbilical cord blood-derived stem cells in animal models of stroke has produced encouraging results of functional recovery, reducing infarct size, and higher expression of neuroprotective factors, such as BDNF and VEGF [100–103]. In other studies, human umbilical cord blood has exhibited protective effects in the rat hippocampus in vitro, while promoting dendritic growth. Additional emerging research is investigating the capabilities of human umbilical cord blood hematopoietic stem cells for functional recovery of dopaminergic neuron morphology of the substantia nigra, caudate, and putamen in an 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) Parkinson's disease mouse model. After intracardioventricular injection, there was an increase in size and density of tyrosine hydroxylase staining cells of the substantia nigra [104].
3.3.2. Shedding the Fat for Stem Cells: Adipose Tissue
Adipose tissue derived stem cells have demonstrated the ability to differentiate into neural, glial, and vascular endothelial cells, and also show higher proliferative activity with greater production of VEGF and hepatocyte growth factor in comparison with bone marrow derived stromal cells [98]. In combination with the accessibility, these features make adipocyte-derived stem cells a desirable source for neurovascular therapy. Transplantation of adipose-derived stem cells in ischemic stroke models demonstrates reduction in damage [98]. Additional studies exhibited reduced infarct size, improved neurological function, reduced level of cerebral inflammation, and chronic degeneration in an intracerebral hemorrhage model, substantiating their therapeutic value [105,106].
Yet, stem cells derived from adipose tissue are also subject to limitations. It was considered that spontaneous mutations occur with extensive passaging that foster tumorigenesis, potentially leading to cancer [107,108]. Follow-up studies suggest adipose-derived stem cells promote pre-existing cancerous cells, but do not initiate tumorigenesis. In a human clinical trial of spinal cord injury patients, none of the eight patients experienced any adverse events within the three-month follow-up [109].
3.3.3. Gender-Specific Stem Cells: Menstrual Blood-Derived Stem Cells
Following many of the factors considered in harvesting stem cells, menstrual blood provides a source with many benefits. With the monthly cycling of the endometrium, the ease and availability for harvesting is a large benefit in the research for therapeutic potential. Stem cells collected from menstrual blood demonstrate multipotency and secrete trophic factors such as VEGF, BDNF, and NT-3 in response to oxygen glucose deprivation (OGD) in an in vitro model of stroke. In such studies, the co-culturing of rat primary neurons with menstrual blood, or its conditioned media, improved survival rate [110]. Further, both intracerebral and intravascular transplantation of menstrual blood-derived stem cells in rat stroke models also enhanced survival and behavioral function [110]. Of note, it has been observed that the adherent fraction of menstrual cells do not lose their karyotypic normality or develop tumorigenic potential even after being expanded through 68 doublings [111].
3.3.4. Mother Knows Best: Breast Milk-Derived Stem Cells
Mammary stem cells (MaSCs) present in tissue of the breast, along with differentiated cells, enter the milk through lactating epithelium. Researchers postulate that these cells enter the breast milk through a combination of migration, cell turnover, and mechanical shearing forces [112,113]. The stem cells of breast milk demonstrate pluripotency similar to that of embryonic stem cell morphology and phenotype and thus allow for differentiation into all three germ layers in vitro [113]. Future research may elucidatee therapeutic potentials in line with those of ESCs. Additional benefits results from the noninvasive harvesting of the cells, availability, and potential for autologous transplant. In terms of tumorigenicity of breast milk-derived stem cells, a study has reported that even nine weeks after subcutaneous injection of breast milk-derived stem cells in immunodeficient mice, these cells did not produce tumors [113]. Along this line, subpopulations of pluripotent adult cells and other multilineage stem cells have failed to form teratomas. Further work characterizing breast milk-derived stem cells for any oncogenic activity under different pathological conditions is needed to determine their tumorigenicity.
3.3.5. A Wisdom Tooth: Dental Tissue-Derived Stem Cells
Dental tissue-derived stem cells, such as post-natal dental pulp stem cells (DPSCs) [114], stem cells from exfoliated deciduous teeth (SHED) [115], periodontal ligament stem cells (PDLSCs) [116], stem cells from apical papilla (SCAP) [116,117], and dental follicle precursor cells (DFPCs) [118], which exhibit mesenchymal stromal cell-like capabilities, have been identified (for review, see [119]). Furthermore, dental tissue-derived stem cells have demonstrated differentiation into a variety of cell lines including neural tissue, adipocytes, and odontoblasts [120].
The use of dental tissue-derived stem cells have been utilized in the study of animal model middle cerebral artery occlusion (MCAO), demonstrating improved motor function following transplantation into the dorsolateral striatum [121].
3.3.6. Reverting Differentiated Tissues to Stem Cells: Induced-Pluripotent Stem Cells
Once considered unidirectional, stem cells were thought to progress through a linear maturation process leaving them terminally differentiated. However, current evidence suggests otherwise. Through manipulation, differentiated stem cells may be coerced into a prior state of multipotency. Utilizing the method of transfecting specific transcription factors fibroblasts can be manipulated into their embryonic-like stem cell precursors [122]. This technique has also been applied to umbilical cord blood cells, placental mesenchymal stromal cells, neural stem cells, and adipose-derived precursor cells to increase their potency [123,124]. Further studies in animal models of ischemic stroke demonstrate that some of the benefits in transplanting induced pluripotent stem cells (iPSCs) includes improving sensorimotor functions [125,126], reducing infarct size, reducing pro-inflammatory cytokines, and increasing anti-inflammatory cytokines [125].
As noted above, there is speculation for concern when transplanting less differentiated cells. Of particular apprehension is their potential for tumorigenesis and immunogenicity. The transfection technique used to induce retrograde manipulation utilizes transcription factors of known oncogenicity. The finding that transplantation of iPSCs into ischemic brain tissue produces a higher incidence of tumors than in healthy brain tissue, further supports this notion [127]. Transplantation is also limited by rejection by the host, even when autologous cells are grafted [128].
With the aforementioned concerns in mind, emergent research is demonstrating the therapeutic feasibility of vector-free and transgene-free induced pluripotent cells while reducing their tumor potential. A current study investigates the use of these human iPS cell-derived neural progenitor cells (hiPS-NPCs) in a mouse ischemic stroke model after discovering they differentiated into functional neurons in vitro. There was no evidence of tumor formation for 12 months following in vivo transplantation [129].
4. Stem Cell Therapy is Not a Magic Bullet: Exploring Co-Adjunctive Therapies
Due to distinct therapeutic potential of individual cell lines, the possibility exists to combine their respective benefits in targeting disease. Mounting literature substantiates the potential for synergistic effects on stem cell survival when co-transplanted. One such study established enhanced stem cell survival when delivered with adipose-derived stem cells [130]. Moreover, co-transplantation therapy may also decrease adverse events. The co-transplantation of bone marrow-derived stromal cells with embryonic stem cells reduced the incidence of tumor production and transplanting neural stem cells with epithelial cells enhanced survival while promoting differentiation [131,132].
The ability to enhance therapeutic effects is not limited solely to the use of stem cells. Combination therapy employs the addition of a substrate to enhance the efficacy of the stem cell line being transplanted. Examples include: Combining bone marrow-derived stromal cells with trophic factors to enhance survival and potentiation [133] or providing a scaffold for stem cell adherence [134].
The recognition of immunosuppressant factors secreted by certain cells (such as bone marrow and Sertoli cells) supports the use of co-transplantation with an immune-protective cell to allow better graft survival of the cells [135]. Sertoli cells are the germ cells of the testis and it has been shown that they are able to secrete trophic factors that are highly immunosuppressive, and which serve as neuroprotective factors in different animal models of neurological disorders [135–137]. As discussed previously, with unique sets of growth factors secreted by stem cells, co-transplantation of stem cells should generate a cocktail of growth factors to be secreted and delivered to the injured brain, thereby affording much more improved therapeutic outcomes. Furthermore, following brain injury, different cell types die or succumb to neurodegeneration, thus warranting the need to transplant multiple cell types. In this case co-transplantation of cells that could differentiate into these multiple cell types will be a logical approach towards replacement of the variety of cells damaged after brain injury [138]. A recent study has demonstrated that neural progenitor cell (NPC) survival and therapeutic support can be enhanced when co-grafted with other genetically modified doxycycline NPCs that can provide bFGF when activated [139–141].
As combinations for therapy continue to surface and demonstrate effectiveness, many variables still persist. Factors including: optimal dose, route of administration, and sex of donor/recipient, all of which are likely to be contingent upon the cell type being investigated. To date, we have investigated many of these parameters with umbilical cord blood for conditions such as Alzheimer's disease, Amyotrophic Lateral Sclerosis (ALS), and Sanfilippo syndrome [142], however, there is still much to be ascertained in regards to stroke therapy. To this end, the Stem Cell Therapies as an Emerging Paradigm in Stroke (STEPS) was initiated to resolve these issues and standardize procedures [143–146].
5. Conclusions
Throughout this review we discussed how each cell line under investigation has its own unique benefits and limitations associated with use. Some of those limitations are immunogenicity, tumorigenicity, ease of harvesting, and the ability to proliferate cells. Researchers are currently addressing these issues through many of the techniques reviewed; yet there are still limited clinical trials. More so, current research is expanding beyond a single cell line transplant. It is likely that future clinical therapy may include the use of co-transplantation and combination therapy mentioned. Further studies also aim to explore the molecular mechanism of response by native tissue in the presence of stem cells. This may progress the exploration of unique trophic factors produced by the stem cells and their utilization in these novel therapies. In moving forward, research must still be conducted in assessing factors for optimal transplantation parameters and the efficacy of treatment, however, across the literature, it is evident that stem cells provide a promising niche of therapeutic potential.
Footnotes
Conflicts of Interest: Cesario V. Borlongan holds patents in stem cell technologies for the treatment of neurodegenerative disorders. Cesario V. Borlongan is supported by James and Esther King Foundation for Biomedical Research Program 1KG01-33966 and NIH NINDS RO1 1R01NS071956-01.
Contributor Information
Travis Dailey, Email: tdailey@health.usf.edu.
Christopher Metcalf, Email: cmetcalf@health.usf.edu.
Yusef I. Mosley, Email: ymosley@health.usf.edu.
Robert Sullivan, Email: rsulliv1@health.usf.edu.
Kazutaka Shinozuka, Email: kshinozu@health.usf.edu.
Naoki Tajiri, Email: ntajiri@health.usf.edu.
Mibel Pabon, Email: mpabon@health.usf.edu.
Sandra Acosta, Email: sacosta@health.usf.edu.
Yuji Kaneko, Email: ykaneko@health.usf.edu.
Harry van Loveren, Email: hvanlove@health.usf.edu.
References
- 1.Saino O, Taguchi A, Nakagomi T, Nakano-Doi A, Kashiwamura S, Doe N, Nakagomi N, Soma T, Yoshikawa H, Stern DM, et al. Immunodeficiency reduces neural stem/progenitor cell apoptosis and enhances neurogenesis in the cerebral cortex after stroke. J Neurosci Res. 2010;88:2385–2397. doi: 10.1002/jnr.22410. [DOI] [PubMed] [Google Scholar]
- 2.Erlandsson A, Lin CH, Yu F, Morshead CM. Immunosuppression promotes endogenous neural stem and progenitor cell migration and tissue regeneration after ischemic injury. Exp Neurol. 2011;230:48–57. doi: 10.1016/j.expneurol.2010.05.018. [DOI] [PubMed] [Google Scholar]
- 3.Willing AE, Eve DJ, Sanberg PR. Umbilical cord blood transfusions for prevention of progressive brain injury and induction of neural recovery: An immunological perspective. Regen Med. 2007;2:457–464. doi: 10.2217/17460751.2.4.457. [DOI] [PubMed] [Google Scholar]
- 4.Hunt JS, Petroff MG, McIntire RH, Ober C. HLA-G and immune tolerance in pregnancy. FASEB J. 2005;19:681–693. doi: 10.1096/fj.04-2078rev. [DOI] [PubMed] [Google Scholar]
- 5.Lee JM, Jung J, Lee HJ, Jeong SJ, Cho KJ, Hwang SG, Kim GJ. Comparison of immunomodulatory effects of placenta mesenchymal stem cells with bone marrow and adipose mesenchymal stem cells. Int Immunopharmacol. 2012;13:219–224. doi: 10.1016/j.intimp.2012.03.024. [DOI] [PubMed] [Google Scholar]
- 6.Menier C, Rouas-Freiss N, Favier B, LeMaoult J, Moreau P, Carosella ED. Recent advances on the non-classical major histocompatibility complex class I HLA-G molecule. Tissue Antigens. 2010;75:201–206. doi: 10.1111/j.1399-0039.2009.01438.x. [DOI] [PubMed] [Google Scholar]
- 7.Fazekasova H, Lechler R, Langford K, Lombardi G. Placenta-derived MSCs are partially immunogenic and less immunomodulatory than bone marrow-derived MSCs. J Tissue Eng Regen Med. 2011;5:684–694. doi: 10.1002/term.362. [DOI] [PubMed] [Google Scholar]
- 8.Newcomb JD, Ajmo CT, Jr, Sanberg CD, Sanberg PR, Pennypacker KR, Willing AE. Timing of cord blood treatment after experimental stroke determines therapeutic efficacy. Cell Transpl. 2006;15:213–223. doi: 10.3727/000000006783982043. [DOI] [PubMed] [Google Scholar]
- 9.Huang H, Chen L, Sanberg P. Cell therapy from bench to bedside translation in CNS neurorestoratology era. Cell Med. 2010;1:15–46. doi: 10.3727/215517910X516673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Oyamada N, Itoh H, Sone M, Yamahara K, Miyashita K, Park K, Taura D, Inuzuka M, Sonoyama T, Tsujimoto H, et al. Transplantation of vascular cells derived from human embryonic stem cells contributes to vascular regeneration after stroke in mice. J Transl Med. 2008;6:1–14. doi: 10.1186/1479-5876-6-54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hayashi J, Takagi Y, Fukuda H, Imazato T, Nishimura M, Fujimoto M, Takahashi J, Hashimoto N, Nozaki K. Primate embryonic stem cell-derived neuronal progenitors transplanted into ischemic brain. J Cereb Blood Flow Metab. 2006;26:906–914. doi: 10.1038/sj.jcbfm.9600247. [DOI] [PubMed] [Google Scholar]
- 12.Yanagisawa D, Qi M, Kim DH, Kitamura Y, Inden M, Tsuchiya D, Takata K, Taniguchi T, Yoshimoto K, Shimoama S, et al. Improvement of focal ischemia-induced rat dopaminergic dysfunction by striatal transplantation of mouse embryonic stem cells. Neurosci Lett. 2006;407:74–79. doi: 10.1016/j.neulet.2006.08.007. [DOI] [PubMed] [Google Scholar]
- 13.Wei L, Cui L, Snider BJ, Rivkin M, Yu SS, Lee CS, Adams LD, Gottlieb DI, Johnson EM, Yu SP, et al. Transplantation of embryonic stem cells overexpressing Bcl-2 promotes functional recovery after transient cerebral ischemia. Neurobiol Dis. 2005;19:183–193. doi: 10.1016/j.nbd.2004.12.016. [DOI] [PubMed] [Google Scholar]
- 14.Pignataro G, Studer FE, Wilz A, Simon RP, Boison D. Neuroprotection in ischemic mouse brain induced by stem cell-derived brain implants. J Cereb Blood Flow Metab. 2007;27:919–927. doi: 10.1038/sj.jcbfm.9600422. [DOI] [PubMed] [Google Scholar]
- 15.Theus MH, Wei L, Cui L, Francis K, Hu XY, Keogh C, Yu SP. In vitro hypoxic preconditioning of embryonic stem cells as a strategy of promoting cell survival and functional benefits after transplantation into the ischemic rat brain. Exp Neurol. 2008;210:656–670. doi: 10.1016/j.expneurol.2007.12.020. [DOI] [PubMed] [Google Scholar]
- 16.Yang T, Tsang KS, Poon WS, Ng HK. Neurotrophism of bone marrow stromal cells to embryonic stem cells: Noncontact induction and transplantation to a mouse ischemic stroke model. Cell Transpl. 2009;18:391–404. doi: 10.3727/096368909788809767. [DOI] [PubMed] [Google Scholar]
- 17.Li Z, McKercher SR, Cui J, Nie ZG, Soussou W, Roberts AJ, Sallmen T, Lipton JH, Talantova M, Okamoto SI, et al. Myocyte enhancer factor 2C as a neurogenic and antiapoptotic transcription factor in murine embryonic stem cells. J Neurosci. 2008;28:6557–6568. doi: 10.1523/JNEUROSCI.0134-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hoehn M, Kustermann E, Blunk J, Wiedermann D, Trapp T, Wecker S, Focking M, Arnold H, Hescheler J, Fleischmann BK, et al. Monitoring of implanted stem cell migration in vivo: A highly resolved in vivo magnetic resonance imaging investigation of experimental stroke in rat. Proc Natl Acad Sci USA. 2002;99:16267–16272. doi: 10.1073/pnas.242435499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lappalainen RS, Narkilahti S, Huhtala T, Liimatainen T, Suuronen T, Narvanen A, Suuronen R, Hovatta O, Jolkkonen J. The SPECT imaging shows the accumulation of neural progenitor cells into internal organs after systemic administration in middle cerebral artery occlusion rats. Neurosci Lett. 2008;440:246–250. doi: 10.1016/j.neulet.2008.05.090. [DOI] [PubMed] [Google Scholar]
- 20.Newman MB, Misiuta I, Willing AE, Zigova T, Karl RC, Borlongan CV, Sanberg PR. Tumorigenicity issues of embryonic carcinoma-derived stem cells: Relevance to surgical trials using NT2 and hNT neural cells. Stem Cells Dev. 2005;14:29–43. doi: 10.1089/scd.2005.14.29. [DOI] [PubMed] [Google Scholar]
- 21.Kawai H, Yamashita T, Ohta Y, Deguchi K, Nagotani S, Zhang X, Ikeda Y, Matsuura T, Abe K. Tridermal tumorigenesis of induced pluripotent stem cells transplanted in ischemic brain. J Cereb Blood Flow Metabol. 2010;30:1487–1493. doi: 10.1038/jcbfm.2010.32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Przyborski S. Differentiation of human embryonic stem cells after transplantation in immune-deficient mice. Stem Cells. 2005;23:1242–1250. doi: 10.1634/stemcells.2005-0014. [DOI] [PubMed] [Google Scholar]
- 23.Hovatta O, Jaconi M, Thnen V, Bna F, Gimelli S, Bosman A, Holm F, Wyder S, Zdobnov EM, Irion O, et al. A teratocarcinoma-like human embryonic stem cell (hESC) line and four hESC lines reveal potentially oncogenic genomic changes. PloS One. 2010;5:e10263. doi: 10.1371/journal.pone.0010263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hara K, Yasuhara T, Maki M, Matsukawa N, Masuda T, Yu SJ, Ali M, Yu G, Xu L, Kim SU, et al. Neural progenitor NT2N cell lines from teratocarcinoma for transplantation therapy in stroke. Prog Neurobiol. 2008;3:318–334. doi: 10.1016/j.pneurobio.2008.04.005. [DOI] [PubMed] [Google Scholar]
- 25.Ghosh Z, Huang M, Hu S, Wilson KD, Dey D, Wu JC. Dissecting the oncogenic and tumorigenic potential of differentiated human induced pluripotent stem cells and human embryonic stem cells. Cancer Res. 2011;14:5030–5039. doi: 10.1158/0008-5472.CAN-10-4402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Snyder EY. The risk of putting something where it does not belong: Mesenchymal stem cells produce masses in the brain. Exp Neurol. 2011;1:75–77. doi: 10.1016/j.expneurol.2011.03.012. [DOI] [PubMed] [Google Scholar]
- 27.Marcus AJ, Woodbury D. Fetal stem cells from extra-embryonic tissues: Do not discard. J Cell Mol Med. 2008;12:730–742. doi: 10.1111/j.1582-4934.2008.00221.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yu SJ, Soncini M, Kaneko Y, Hess DC, Parolini O, Borlongan CV. Amnion: A potent graft source for cell therapy in stroke. Cell Transpl. 2009;18:111–118. doi: 10.3727/096368909788341243. [DOI] [PubMed] [Google Scholar]
- 29.Konig J, Huppertz B, Desoye G, Parolini O, Frohlich JD, Weiss G, Dohr G, Sedlmayr P, Lang I. Amnion-derived mesenchymal stromal cells show angiogenic properties but resist differentiation into mature endothelial cells. Stem Cells Dev. 2012;21:1309–1320. doi: 10.1089/scd.2011.0223. [DOI] [PubMed] [Google Scholar]
- 30.Yarygin KN, Kholodenko IV, Konieva AA, Burunova VV, Tairova RT, Gubsky LV, Cheglakov IB, Pirogov YA, Yarygin VN, Skvortsova VI. Mechanisms of positive effects of transplantation of human placental mesenchymal stem cells on recovery of rats after experimental ischemic stroke. Bull Exp Biol Med. 2009;148:862–868. doi: 10.1007/s10517-010-0837-z. [DOI] [PubMed] [Google Scholar]
- 31.Chen J, Shehadah A, Pal A, Zacharek A, Cui X, Cui Y, Roberts C, Lu M, Zeitlin A, Hariri R, et al. Neuroprotective effect of human placenta-derived cell treatment of stroke in rats. Cell Transpl. 2012;22:871–879. doi: 10.3727/096368911X637380. [DOI] [PubMed] [Google Scholar]
- 32.Kranz A, Wagner DC, Kamprad M, Scholz M, Schmidt UR, Nitzsche F, Aberman Z, Emmrich F, Riegelsberger UM, Boltze J. Transplantation of placenta-derived mesenchymal stromal cells upon experimental stroke in rats. Brain Res. 2010;1315:128–136. doi: 10.1016/j.brainres.2009.12.001. [DOI] [PubMed] [Google Scholar]
- 33.Liao WB, Xie J, Zhong J, Liu YJ, Du L, Zhou B, Xu J, Liu PX, Yang SG, Wang JM, et al. Therapeutic effect of human umbilical cord multipotent mesenchymal stromal cells in a rat model of stroke. Transplantation. 2009;87:350–359. doi: 10.1097/TP.0b013e318195742e. [DOI] [PubMed] [Google Scholar]
- 34.Deuse T, Stubbendorff M, Tang-Quan K, Phillips N, Kay MA, Eiermann T, Phan TT, Volk HD, Reichenspurner H, Robbins RC, et al. Immunogenicity and immunomodulatory properties of umbilical cord lining mesenchymal stem cells. Cell Transpl. 2011;20:655–667. doi: 10.3727/096368910X536473. [DOI] [PubMed] [Google Scholar]
- 35.Borlongan CV, Glover LE, Tajiri N, Kaneko Y, Freeman TB. The great migration of bone marrow-derived stem cells toward the ischemic brain: Therapeutic implications for stroke and other neurological disorders. Prog Neurobiol. 2011;95:213–228. doi: 10.1016/j.pneurobio.2011.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Herzog EL, Chai L, Krause DS. Plasticity of marrow-derived stem cells. Blood. 2003;102:3483–3493. doi: 10.1182/blood-2003-05-1664. [DOI] [PubMed] [Google Scholar]
- 37.Lapidot T, Dar A, Kollet O. How do stem cells find their way home? Blood. 2005;106:1901–1910. doi: 10.1182/blood-2005-04-1417. [DOI] [PubMed] [Google Scholar]
- 38.Lapidot T, Kollet O. The brain-bone-blood triad: Traffic lights for stem-cell homing and mobilization. Hematology. 2010;2010:1–6. doi: 10.1182/asheducation-2010.1.1. [DOI] [PubMed] [Google Scholar]
- 39.Nervi B, Link DC, DiPersio JF. Cytokines and hematopoietic stem cell mobilization. J Cell Biochem. 2006;99:690–705. doi: 10.1002/jcb.21043. [DOI] [PubMed] [Google Scholar]
- 40.Papayannopoulou T, Scadden DT. Stem-cell ecology and stem cells in motion. Blood. 2008;111:3923–3930. doi: 10.1182/blood-2007-08-078147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kalinkovich A, Spiegel A, Shivtiel S, Kollet O, Jordaney N, Piacibello W, Lapidot T. Blood-forming stem cells are nervous: Direct and indirect regulation of immature human CD34+ cells by the nervous system. Brain Behav Immun. 2009;23:1059–1065. doi: 10.1016/j.bbi.2009.03.008. [DOI] [PubMed] [Google Scholar]
- 42.Moniche F, Gonzalez A, Gonzalez-Marcos JR, Carmona M, Pinero P, Espigado I, Garcia-Solis D, Cayuela A, Montaner J, Boada C, et al. Intra-arterial bone marrow mononuclear cells in ischemic stroke a pilot clinical trial. Stroke. 2012;43:2242–2244. doi: 10.1161/STROKEAHA.112.659409. [DOI] [PubMed] [Google Scholar]
- 43.Savitz SI, Misra V, Kasam M, Juneja H, Cox CS, Alderman S, Aisiku I, Kar S, Gee A, Grotta JC. Intravenous autologous bone marrow mononuclear cells for ischemic stroke. Ann Neurol. 2011;70:59–69. doi: 10.1002/ana.22458. [DOI] [PubMed] [Google Scholar]
- 44.Battistella V, de Freitas GR, da Fonseca LMB, Mercante D, Gutfilen B, Goldenberg RCS, Dias JV, Kasai-Brunswick TH, Wajnberg E, Rosado-de-Castro PH, et al. Safety of autologous bone marrow mononuclear cell transplantation in patients with nonacute ischemic stroke. Regen Med. 2011;6:45–52. doi: 10.2217/rme.10.97. [DOI] [PubMed] [Google Scholar]
- 45.Friedrich MAG, Martins MP, Araujo MD, Klamt C, Vedolin L, Garicochea B, Raupp EF, El Ammar JS, Machado DC, da Costa JC, et al. Intra-arterial infusion of autologous bone marrow mononuclear cells in patients with moderate to severe middle cerebral artery acute ischemic stroke. Cell Transpl. 2012;21:13–21. doi: 10.3727/096368912x612512. [DOI] [PubMed] [Google Scholar]
- 46.Felfly H, Muotri A, Yao H, Haddad GG. Hematopoietic stem cell transplantation protects mice from lethal stroke. Exp Neurol. 2010;225:284–293. doi: 10.1016/j.expneurol.2010.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Schwarting S, Litwak S, Hao W, Bähr M, Weise J, Neumann H. Hematopoietic stem cells reduce postischemic inflammation and ameliorate ischemic brain injury. Stroke. 2008;39:2867–2875. doi: 10.1161/STROKEAHA.108.513978. [DOI] [PubMed] [Google Scholar]
- 48.Chopp M, Li Y. Treatment of neural injury with marrow stromal cells. Lancet Neurol. 2002;1:92–100. doi: 10.1016/s1474-4422(02)00040-6. [DOI] [PubMed] [Google Scholar]
- 49.Rempe DA, Kent TA. Using bone marrow stromal cells for treatment of stroke. Neurology. 2002;59:486–487. doi: 10.1212/wnl.59.4.486. [DOI] [PubMed] [Google Scholar]
- 50.Song S, Kamath S, Mosquera D, Zigova T, Sanberg P, Vesely DL, Sanchez-Ramos J. Expression of brain natriuretic peptide by human bone marrow stromal cells. Exp Neurol. 2004;185:191–197. doi: 10.1016/j.expneurol.2003.09.003. [DOI] [PubMed] [Google Scholar]
- 51.Li Y, Chen J, Wang L, Lu M, Chopp M. Treatment of stroke in rat with intracarotid administration of marrow stromal cells. Neurology. 2001;56:1666–1672. doi: 10.1212/wnl.56.12.1666. [DOI] [PubMed] [Google Scholar]
- 52.Chen J, Li Y, Katakowski M, Chen X, Wang L, Lu D, Lu M, Gautam SC, Chopp M. Intravenous bone marrow stromal cell therapy reduces apoptosis and promotes endogenous cell proliferation after stroke in female rat. J Neurosci Res. 2003;73:778–786. doi: 10.1002/jnr.10691. [DOI] [PubMed] [Google Scholar]
- 53.Chen J, Li Y, Wang L, Lu M, Chopp M. Caspase inhibition by Z-VAD increases the survival of grafted bone marrow cells and improves functional outcome after MCAo in rats. J Neurol Sci. 2002;199:17–24. doi: 10.1016/s0022-510x(02)00075-8. [DOI] [PubMed] [Google Scholar]
- 54.Zhang J, Li Y, Chen J, Yang M, Katakowski M, Lu M, Chopp M. Expression of insulin-like growth factor 1 and receptor in ischemic rats treated with human marrow stromal cells. Brain Res. 2004;1030:19–27. doi: 10.1016/j.brainres.2004.09.061. [DOI] [PubMed] [Google Scholar]
- 55.Bang OY, Lee JS, Lee PH, Lee G. Autologous mesenchymal stem cell transplantation in stroke patients. Ann Neurol. 2005;57:874–882. doi: 10.1002/ana.20501. [DOI] [PubMed] [Google Scholar]
- 56.Lee JS, Hong JM, Moon GJ, Lee PH, Ahn YH, Bang OY, Collaborators S. A long-term follow-up study of intravenous autologous mesenchymal stem cell transplantation in patients with ischemic stroke. Stem Cells. 2010;28:1099–1106. doi: 10.1002/stem.430. [DOI] [PubMed] [Google Scholar]
- 57.Shen LH, Li Y, Chen J, Zacharek A, Gao Q, Kapke A, Lu M, Raginski K, Vanguri P, Smith A, et al. Therapeutic benefit of bone marrow stromal cells administered 1 month after stroke. J Cereb Blood Flow Metab. 2007;27:6–13. doi: 10.1038/sj.jcbfm.9600311. [DOI] [PubMed] [Google Scholar]
- 58.Barlow S, Brooke G, Chatterjee K, Price G, Pelekanos R, Rossetti T, Doody M, Venter D, Pain S, Gilshenan K, et al. Comparison of human placenta- and bone marrow-derived multipotent mesenchymal stem cells. Stem Cells Dev. 2008;17:1095–1107. doi: 10.1089/scd.2007.0154. [DOI] [PubMed] [Google Scholar]
- 59.Jansen BJ, Gilissen C, Roelofs H, Schaap-Oziemlak A, Veltman JA, Raymakers RA, Jansen JH, Kogler G, Figdor CG, Torensma R, et al. Functional differences between mesenchymal stem cell populations are reflected by their transcriptome. Stem Cells Dev. 2010;19:481–490. doi: 10.1089/scd.2009.0288. [DOI] [PubMed] [Google Scholar]
- 60.Kim SH, Kim YS, Lee SY, Kim KH, Lee YM, Kim WK, Lee YK. Gene expression profile in mesenchymal stem cells derived from dental tissues and bone marrow. J Periodontal Implant Sci. 2011;41:192–200. doi: 10.5051/jpis.2011.41.4.192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Dmitrieva RI, Minullina IR, Bilibina AA, Tarasova OV, Anisimov SV, Zaritskey AY. Bone marrow- and subcutaneous adipose tissue-derived mesenchymal stem cells: Differences and similarities. Cell Cycle. 2012;11:377–383. doi: 10.4161/cc.11.2.18858. [DOI] [PubMed] [Google Scholar]
- 62.Strioga M, Viswanathan S, Darinskas A, Slaby O, Michalek J. Same or not the same? Comparison of adipose tissue-derived versus bone marrow-derived mesenchymal stem and stromal cells. Stem Cells Dev. 2012;21:2724–2752. doi: 10.1089/scd.2011.0722. [DOI] [PubMed] [Google Scholar]
- 63.Lin RZ, Moreno-Luna R, Zhou B, Pu WT, Melero-Martin JM. Equal modulation of endothelial cell function by four distinct tissue-specific mesenchymal stem cells. Angiogenesis. 2012;15:443–455. doi: 10.1007/s10456-012-9272-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Egashira Y, Sugitani S, Suzuki Y, Mishiro K, Tsuruma K, Shimazawa M, Yoshimura S, Iwama T, Hara H. The conditioned medium of murine and human adipose-derived stem cells exerts neuroprotective effects against experimental stroke model. Brain Res. 2012;1461:87–95. doi: 10.1016/j.brainres.2012.04.033. [DOI] [PubMed] [Google Scholar]
- 65.Ribeiro CA, Fraga JS, Graos M, Neves NM, Reis RL, Gimble JM, Sousa N, Salgado AJ. The secretome of stem cells isolated from the adipose tissue and Wharton jelly acts differently on central nervous system derived cell populations. Stem Cell Res Ther. 2012;3:1–7. doi: 10.1186/scrt109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Borlongan C, Hadman M, Sanberg C, Sanberg O. Central nervous system entry of peripherally injected umbilical cord blood cells is not required for neuroprotection in stroke. Stroke. 2004;35:2385–2389. doi: 10.1161/01.STR.0000141680.49960.d7. [DOI] [PubMed] [Google Scholar]
- 67.Aloe L, Rocco ML, Bianchi P, Manni L. Nerve growth factor: From the early discoveries to the potential clinical use. J Transl Med. 2012;10:1–15. doi: 10.1186/1479-5876-10-239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ball S, Marangell LB, Lipsius S, Russell JM. Brain-derived neurotrophic factor in generalized anxiety disorder: Results from a duloxetine clinical trial. Prog Neuropsychopharmacol Biol Psychiatry. 2013;3:217–221. doi: 10.1016/j.pnpbp.2013.01.002. [DOI] [PubMed] [Google Scholar]
- 69.Kim WS, Lim JY, Shin JH, Park HK, Tan SA, Park KU, Paik NJ. Effect of the presence of brain-derived neurotrophic factor val(66)met polymorphism on the recovery in patients with acute subcortical stroke. Ann Rehabil Med. 2013;37:311–319. doi: 10.5535/arm.2013.37.3.311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Stanzani L, Zoia C, Sala G, Appollonio I, Frattola L, de Simoni MG, Ferrarese C. Nerve growth factor and transforming growth factor-beta serum levels in acute stroke patients. Possible involvement of neurotrophins in cerebrovascular disease. Cerebrovasc Dis. 2001;12:240–244. doi: 10.1159/000047710. [DOI] [PubMed] [Google Scholar]
- 71.Tolar J, Nauta AJ, Osborn MJ, Panoskaltsis Mortari A, McElmurry RT, Bell S, Xia L, Zhou N, Riddle M, Schroeder TM, et al. Sarcoma derived from cultured mesenchymal stem cells. Stem Cells. 2007;25:371–379. doi: 10.1634/stemcells.2005-0620. [DOI] [PubMed] [Google Scholar]
- 72.De Luca A, Lamura L, Gallo M, Maffia V, Normanno N. Mesenchymal stem cell-derived interleukin-6 and vascular endothelial growth factor promote breast cancer cell migration. J Cell Biochem. 2012;113:3363–3370. doi: 10.1002/jcb.24212. [DOI] [PubMed] [Google Scholar]
- 73.Karnoub AE, Dash AB, Vo AP, Sullivan A, Brooks MW, Bell GW, Richardson AL, Polyak K, Tubo R, Weinberg RA. Mesenchymal stem cells within tumour stroma promote breast cancer metastasis. Nature. 2007;449:557–563. doi: 10.1038/nature06188. [DOI] [PubMed] [Google Scholar]
- 74.Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, Witzenbichler B, Schatteman G, Isner JM. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275:964–967. doi: 10.1126/science.275.5302.964. [DOI] [PubMed] [Google Scholar]
- 75.Kawamoto A, Losordo DW. Endothelial progenitor cells for cardiovascular regeneration. Trends Cardiovasc Med. 2008;18:33–37. doi: 10.1016/j.tcm.2007.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Masuda H, Asahara T. Post-natal endothelial progenitor cells for neovascularization in tissue regeneration. Cardiovasc Res. 2003;58:390–398. doi: 10.1016/s0008-6363(02)00785-x. [DOI] [PubMed] [Google Scholar]
- 77.Yip HK, Chang LT, Chang WN, Lu CH, Liou CW, Lan MY, Liu JS, Youssef AA, Chang HW. Level and value of circulating endothelial progenitor cells in patients after acute ischemic stroke. Stroke. 2008;39:69–74. doi: 10.1161/STROKEAHA.107.489401. [DOI] [PubMed] [Google Scholar]
- 78.Chen J, Chen SZ, Chen YS, Zhang C, Wang JJ, Zhang WF, Liu G, Zhao B, Chen YF. Circulating endothelial progenitor cells and cellular membrane microparticles in db/db diabetic mouse: Possible implications in cerebral ischemic damage. Am J Physiol Endocrinol Metabol. 2011;301:62–71. doi: 10.1152/ajpendo.00026.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Decano JL, Moran AM, Giordano N, Ruiz-Opazo N, Herrera VL. Analysis of CD45− [CD34+/KDR+] endothelial progenitor cells as juvenile protective factors in a rat model of ischemic-hemorrhagic stroke. PLoS One. 2013;8:e55222. doi: 10.1371/journal.pone.0055222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Chen ZZ, Jiang XD, Zhang LL, Shang JH, Du MX, Xu G, Xu RX. Beneficial effect of autologous transplantation of bone marrow stromal cells and endothelial progenitor cells on cerebral ischemia in rabbits. Neurosci Lett. 2008;445:36–41. doi: 10.1016/j.neulet.2008.08.039. [DOI] [PubMed] [Google Scholar]
- 81.Ratajczak MZ, Machalinski B, Wojakowski W, Ratajczak J, Kucia M. A hypothesis for an embryonic origin of pluripotent Oct-4(+) stem cells in adult bone marrow and other tissues. Leukemia. 2007;21:860–867. doi: 10.1038/sj.leu.2404630. [DOI] [PubMed] [Google Scholar]
- 82.Kucia M, Wysoczynski M, Ratajczak J, Ratajczak MZ. Identification of very small embryonic like (VSEL) stem cells in bone marrow. Cell Tissue Res. 2008;331:125–134. doi: 10.1007/s00441-007-0485-4. [DOI] [PubMed] [Google Scholar]
- 83.Kucia M, Ratajczak J, Ratajczak MZ. Are bone marrow stem cells plastic or heterogenous— That is the question. Exp Hematol. 2005;33:613–623. doi: 10.1016/j.exphem.2005.01.016. [DOI] [PubMed] [Google Scholar]
- 84.Ratajczak J, Shin DM, Wan W, Liu R, Masternak MM, Piotrowska K, Wiszniewska B, Kucia M, Bartke A, Ratajczak MZ. Higher number of stem cells in the bone marrow of circulating low Igf-1 level Laron dwarf mice—Novel view on Igf-1, stem cells and aging. Leukemia. 2011;25:729–733. doi: 10.1038/leu.2010.314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Barkho BZ, Munoz AE, Li X, Li L, Cunningham LA, Zhao X. Endogenous matrix metalloproteinase (MMP)-3 and MMP-9 promote the differentiation and migration of adult neural progenitor cells in response to chemokines. Stem Cells. 2008;26:3139–3149. doi: 10.1634/stemcells.2008-0519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Liu XS, Chopp M, Zhang RL, Hozeska-Solgot A, Gregg SC, Buller B, Lu M, Zhang ZG. Angiopoietin 2 mediates the differentiation and migration of neural progenitor cells in the subventricular zone after stroke. J Biol Chem. 2009;284:22680–22689. doi: 10.1074/jbc.M109.006551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Zhang RL, Chopp M, Gregg SR, Toh Y, Roberts C, Letourneau Y, Buller B, Jia L, Davarani SP, Zhang ZG. Patterns and dynamics of subventricular zone neuroblast migration in the ischemic striatum of the adult mouse. J Cereb Blood Flow Metab. 2009;29:1240–1250. doi: 10.1038/jcbfm.2009.55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Carbajal KS, Schaumburg C, Strieter R, Kane J, Lane TE. Migration of engrafted neural stem cells is mediated by CXCL12 signaling through CXCR4 in a viral model of multiple sclerosis. Proc Natl Acad Sci USA. 2010;107:11068–11073. doi: 10.1073/pnas.1006375107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Arvidsson A, Collin T, Kirik D, Kokaia Z, Lindvall O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat Med. 2002;8:963–970. doi: 10.1038/nm747. [DOI] [PubMed] [Google Scholar]
- 90.Nygren J, Wieloch T, Pesic J, Brundin P, Deierborg T. Enriched environment attenuates cell genesis in subventricular zone after focal ischemia in mice and decreases migration of newborn cells to the striatum. Stroke. 2006;37:2824–2829. doi: 10.1161/01.STR.0000244769.39952.90. [DOI] [PubMed] [Google Scholar]
- 91.Deierborg T, Staflin K, Pesic J, Roybon L, Brundin P, Lundberg C. Absence of striatal newborn neurons with mature phenotype following defined striatal and cortical excitotoxic brain injuries. Exp Neurol. 2009;219:363–367. doi: 10.1016/j.expneurol.2009.05.002. [DOI] [PubMed] [Google Scholar]
- 92.Deierborg T, Roybon L, Inacio AR, Pesic J, Brundin P. Brain injury activates microglia that induce neural stem cell proliferation ex vivo and promote differentiation of neurosphere-derived cells into neurons and oligodendrocytes. Neuroscience. 2010;171:1386–1396. doi: 10.1016/j.neuroscience.2010.09.045. [DOI] [PubMed] [Google Scholar]
- 93.Bachstetter AD, Pabon MM, Cole MJ, Hudson CE, Sanberg PR, Willing AE, Bickford PC, Gemma C. Peripheral injection of human umbilical cord blood stimulates neurogenesis in the aged rat brain. BMC Neurosci. 2008;9 doi: 10.1186/1471-2202-9-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Park DH, Eve DJ, Sanberg PR, Musso J, III, Bachstetter AD, Wolfson A, Schlunk A, Baradez MO, Sinden JD, Gemma C. Increased neuronal proliferation in the dentate gyrus of aged rats following neural stem cell implantation. Stem Cells Dev. 2010;19:175–180. doi: 10.1089/scd.2009.0172. [DOI] [PubMed] [Google Scholar]
- 95.Van Velthoven CT, Kavelaars A, van Bel F, Heijnen CJ. Mesenchymal stem cell treatment after neonatal hypoxic-ischemic brain injury improves behavioral outcome and induces neuronal and oligodendrocyte regeneration. Brain Behav Immun. 2010;24:387–393. doi: 10.1016/j.bbi.2009.10.017. [DOI] [PubMed] [Google Scholar]
- 96.Jin K, Xie L, Mao X, Greenberg MB, Moore A, Peng B, Greenberg RB, Greenberg DA. Effect of human neural precursor cell transplantation on endogenous neurogenesis after focal cerebral ischemia in the rat. Brain Res. 2011;1374:56–62. doi: 10.1016/j.brainres.2010.12.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Chen N, Newcomb J, Garbuzova-Davis S, Davis Sanberg C, Sanberg PR, Willing AE. Human umbilical cord blood cells have trophic effects on young and aging hippocampal neurons in vitro. Aging Dis. 2010;1:173–190. [PMC free article] [PubMed] [Google Scholar]
- 98.Ikegame Y, Yamashita K, Hayashi SI, Mizuno H, Tawada M, You F, Yamada K, Tanaka Y, Egashira Y, Nakashima S, et al. Comparison of mesenchymal stem cells from adipose tissue and bone marrow for ischemic stroke therapy. Cytotherapy. 2011;13:675–685. doi: 10.3109/14653249.2010.549122. [DOI] [PubMed] [Google Scholar]
- 99.Vendrame M, Gemma C, de Mesquita D, Collier L, Bickford PC, Sanberg CD, Sanberg PR, Pennypacker KR, Willing AE. Anti-inflammatory effects of human cord blood cells in a rat model of stroke. Stem Cells Dev. 2005;14:595–604. doi: 10.1089/scd.2005.14.595. [DOI] [PubMed] [Google Scholar]
- 100.Willing AE, Lixian J, Milliken M, Poulos S, Zigova T, Song S, Hart C, Sanchez-Ramos J, Sanberg PR. Intravenous versus intrastriatal cord blood administration in a rodent model of stroke. J Neurosci Res. 2003;73:296–307. doi: 10.1002/jnr.10659. [DOI] [PubMed] [Google Scholar]
- 101.Xiao J, Nan ZH, Motooka Y, Low WC. Transplantation of a novel cell line population of umbilical cord blood stem cells ameliorates neurological deficits associated with ischemic brain injury. Stem Cells Dev. 2005;14:722–733. doi: 10.1089/scd.2005.14.722. [DOI] [PubMed] [Google Scholar]
- 102.Chung DJ, Choi CB, Lee SH, Kang EH, Lee JH, Hwang SH, Han H, Lee JH, Choe BY, Lee SY, et al. Intraarterially delivered human umbilical cord blood-derived mesenchymal stem cells in canine cerebral ischemia. J Neurosci Res. 2009;87:3554–3567. doi: 10.1002/jnr.22162. [DOI] [PubMed] [Google Scholar]
- 103.Vendrame M, Cassady J, Newcomb J, Butler T, Pennypacker KR, Zigova T, Sanberg CD, Sanberg PR, Willing AE. Infusion of human umbilical cord blood cells in a rat model of stroke dose-dependently rescues behavioral deficits and reduces infarct volume. Stroke. 2004;35:2390–2395. doi: 10.1161/01.STR.0000141681.06735.9b. [DOI] [PubMed] [Google Scholar]
- 104.Lu J, Kanji S, Aggarwal R, Das M, Joseph M, Wu LC, Mao HQ, Pompili VJ, Hadjiconstantinou M, Das H. Umbilical cord blood-derived hematopoietic stem cells improve dopaminergic neuron morphology in the MPTP-mice. Front Biosci. 2013;18:970–981. doi: 10.2741/4156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Kim JM, Lee ST, Chu K, Jung KH, Song EC, Kim SJ, Sinn DI, Kim JH, Park DK, Kang KM, et al. Systemic transplantation of human adipose stem cells attenuated cerebral inflammation and degeneration in a hemorrhagic stroke model. Brain Res. 2007;1183:43–50. doi: 10.1016/j.brainres.2007.09.005. [DOI] [PubMed] [Google Scholar]
- 106.Leu S, Lin YC, Yuen CM, Yen CH, Kao YH, Sun CK, Yip HK. Adipose-derived mesenchymal stem cells markedly attenuate brain infarct size and improve neurological function in rats. J Transl Med. 2010;8 doi: 10.1186/1479-5876-8-63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Rubio D, Garcia-Castro J, Martin MC, de la Fuente R, Cigudosa JC, Lloyd AC, Bernad A. Spontaneous human adult stem cell transformation. Cancer Res. 2005;65:3035–3039. doi: 10.1158/0008-5472.CAN-04-4194. [DOI] [PubMed] [Google Scholar]
- 108.De la Fuente R, Bernad A, Garcia-Castro J, Martin MC, Cigudosa JC. Retraction: Spontaneous human adult stem cell transformation. Cancer Res. 2010;70 doi: 10.1158/0008-5472.CAN-10-2451. [DOI] [PubMed] [Google Scholar]
- 109.Ra JC, Shin IS, Kim SH, Kang SK, Kang BC, Lee HY, Kim YJ, Jo JY, Yoon EJ, Choi HJ, et al. Safety of intravenous infusion of human adipose tissue-derived mesenchymal stem cells in animals and humans. Stem Cells Dev. 2011;20:1297–1308. doi: 10.1089/scd.2010.0466. [DOI] [PubMed] [Google Scholar]
- 110.Borlongan CV, Kaneko Y, Maki M, Yu SJ, Ali M, Allickson JG, Sanberg CD, Kuzmin-Nichols N, Sanberg PR. Menstrual blood cells display stem cell-like phenotypic markers and exert neuroprotection following transplantation in experimental stroke. Stem Cells Dev. 2010;19:439–452. doi: 10.1089/scd.2009.0340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Meng X, Ichim T, Zhong J, Rogers A, Yin Z, Jackson J, Wang H, Ge W, Bogin V, Chan K, et al. Endometrial regenerative cells: A novel stem cell population. J Transl Med. 2007;5 doi: 10.1186/1479-5876-5-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Cregan MD, Fan YP, Appelbee A, Brown ML, Klopcic B, Koppen J, Mitoulas LR, Piper KME, Choolani MA, Chong YS, et al. Identification of nestin-positive putative mammary stem cells in human breastmilk. Cell Tissue Res. 2007;329:129–136. doi: 10.1007/s00441-007-0390-x. [DOI] [PubMed] [Google Scholar]
- 113.Hassiotou F, Beltran A, Chetwynd E, Stuebe AM, Twigger AJ, Metzger P, Trengove N, Lai CT, Filgueira L, Blancafort P, et al. Breastmilk is a novel source of stem cells with multilineage differentiation potential. Stem Cells. 2012;30:2164–2174. doi: 10.1002/stem.1188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Gronthos S, Mankani M, Brahim J, Robey PG, Shi S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc Natl Acad Sci USA. 2000;97:13625–13630. doi: 10.1073/pnas.240309797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Seo BM, Miura M, Gronthos S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet. 2004;364:149–155. doi: 10.1016/S0140-6736(04)16627-0. [DOI] [PubMed] [Google Scholar]
- 116.Sonoyama W, Liu Y, Fang DAJ, Yamaza T, Seo BM, Zhang CM, Liu H, Gronthos S, Wang CY, Shi ST, et al. Mesenchymal stem cell-mediated functional tooth regeneration in swine. PloS One. 2006;1:e79. doi: 10.1371/journal.pone.0000079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Sonoyama W, Liu Y, Yamaza T, Tuan RS, Wang S, Shi S, Huang GTJ. Characterization of the apical papilla and its residing stem cells from human immature permanent teeth: A pilot study. J Endod. 2008;34:166–171. doi: 10.1016/j.joen.2007.11.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Morsczeck C, Gotz W, Schierholz J, Zellhofer F, Kuhn U, Mohl C, Sippel C, Hoffmann KH. Isolation of precursor cells (PCs) from human dental follicle of wisdom teeth. Matrix Biol. 2005;24:155–165. doi: 10.1016/j.matbio.2004.12.004. [DOI] [PubMed] [Google Scholar]
- 119.Huang GTJ, Gronthos S, Shi S. Mesenchymal stem cells derived from dental tissues vs. those from other sources: Their biology and role in regenerative medicine. J Dent Res. 2009;88:792–806. doi: 10.1177/0022034509340867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Miura M, Gronthos S, Zhao M, Lu B, Fisher LW, Robey PG, Shi S. SHED: Stem cells from human exfoliated deciduous teeth. Proc Natl Acad Sci USA. 2003;100:5807–5812. doi: 10.1073/pnas.0937635100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Yang KL, Chen MF, Liao CH, Pang CY, Lin PY. A simple and efficient method for generating Nurr1-positive neuronal stem cells from human wisdom teeth (tNSC) and the potential of tNSC for stroke therapy. Cytotherapy. 2009;11:606–617. doi: 10.1080/14653240902806994. [DOI] [PubMed] [Google Scholar]
- 122.Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–676. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 123.Cai J, Li W, Su H, Qin D, Yang J, Zhu F, Xu J, He W, Guo X, Labuda K, et al. Generation of human induced pluripotent stem cells from umbilical cord matrix and amniotic membrane mesenchymal cells. J Biol Chem. 2010;285:11227–11234. doi: 10.1074/jbc.M109.086389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Tat PA, Sumer H, Jones KL, Upton K, Verma PJ. The efficient generation of induced pluripotent stem (iPS) cells from adult mouse adipose tissue-derived and neural stem cells. Cell Transpl. 2010;19:525–536. doi: 10.3727/096368910X491374. [DOI] [PubMed] [Google Scholar]
- 125.Chen SJ, Chang CM, Tsai SK, Chang YL, Chou SJ, Huang SS, Tai LK, Chen YC, Ku HH, Li HY, et al. Functional improvement of focal cerebral ischemia injury by subdural transplantation of induced pluripotent stem cells with fibrin glue. Stem Cells Dev. 2010;19:1757–1767. doi: 10.1089/scd.2009.0452. [DOI] [PubMed] [Google Scholar]
- 126.Jiang M, Lv L, Ji H, Yang X, Zhu W, Cai L, Gu X, Chai C, Huang S, Sun J, et al. Induction of pluripotent stem cells transplantation therapy for ischemic stroke. Mol Cell Biochem. 2011;354:67–75. doi: 10.1007/s11010-011-0806-5. [DOI] [PubMed] [Google Scholar]
- 127.Yamashita T, Kawai H, Tian FF, Ohta Y, Abe K. Tumorigenic development of induced pluripotent stem cells in ischemic mouse brain. Cell Transpl. 2011;20:883–891. doi: 10.3727/096368910X539092. [DOI] [PubMed] [Google Scholar]
- 128.Zhao T, Zhang ZN, Rong Z, Xu Y. Immunogenicity of induced pluripotent stem cells. Nature. 2011;474:212–215. doi: 10.1038/nature10135. [DOI] [PubMed] [Google Scholar]
- 129.Mohamad O, Drury-Stewart D, Song M, Faulkner B, Chen D, Yu SP, Wei L. Vector-free and transgene-free human iPS cells differentiate into functional neurons and enhance functional recovery after ischemic stroke in mice. PLoS One. 2013;8:e64160. doi: 10.1371/journal.pone.0064160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Oh JS, Kim KN, An SS, Pennant WA, Kim HJ, Gwak SJ, do Yoon H, Lim MH, Choi BH, Ha Y. Cotransplantation of mouse neural stem cells (mNSCs) with adipose tissue-derived mesenchymal stem cells improves mNSC survival in a rat spinal cord injury model. Cell Transpl. 2011;20:837–849. doi: 10.3727/096368910X539083. [DOI] [PubMed] [Google Scholar]
- 131.Matsuda R, Yoshikawa M, Kimura H, Ouji Y, Nakase H, Nishimura F, Nonaka J, Toriumi H, Yamada S, Nishiofuku M, et al. Cotransplantation of mouse embryonic stem cells and bone marrow stromal cells following spinal cord injury suppresses tumor development. Cell Transpl. 2009;18:39–54. [PubMed] [Google Scholar]
- 132.Nakagomi N, Nakagomi T, Kubo S, Nakano-Doi A, Saino O, Takata M, Yoshikawa H, Stern DM, Matsuyama T, Taguchi A. Endothelial cells support survival, proliferation, and neuronal differentiation of transplanted adult ischemia-induced neural stem/progenitor cells after cerebral infarction. Stem Cells. 2009;27:2185–2195. doi: 10.1002/stem.161. [DOI] [PubMed] [Google Scholar]
- 133.Zhang W, Yan Q, Zeng YS, Zhang XB, Xiong Y, Wang JM, Chen SJ, Li Y, Bruce IC, Wu W. Implantation of adult bone marrow-derived mesenchymal stem cells transfected with the neurotrophin-3 gene and pretreated with retinoic acid in completely transected spinal cord. Brain Res. 2010;1359:256–271. doi: 10.1016/j.brainres.2010.08.072. [DOI] [PubMed] [Google Scholar]
- 134.Jin K, Mao X, Xie L, Galvan V, Lai B, Wang Y, Gorostiza O, Wang X, Greenberg DA. Transplantation of human neural precursor cells in Matrigel scaffolding improves outcome from focal cerebral ischemia after delayed postischemic treatment in rats. J Cereb Blood Flow Metab. 2010;30:534–544. doi: 10.1038/jcbfm.2009.219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Kaneko Y, Pabon MM, Dailey T, Weinbren NL, Rizzi J, Tamboli C, Vasconcellos J, Kuzmin-Nichols N, Sanberg PR, Eve DJ, et al. The battle of thesexes for stroke therapy: Female- versus male-derived stem cells. CNS Neurol Disord Drug Targets. 2013;12:405–412. doi: 10.2174/1871527311312030013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Sanberg PR, Borlongan CV, Saporta S, Cameron DF. Testis-derived Sertoli cells survive and provide localized immunoprotection for xenografts in rat brain. Nat Biotechnol. 1996;14:1692–1695. doi: 10.1038/nbt1296-1692. [DOI] [PubMed] [Google Scholar]
- 137.Sanberg PR, Othberg AI, Borlongan CV, Saporta S, Anton A, Freeman TB, Cahill DW, Allen RC, Cameron DF. Transplantation of testis-derived Sertoli cells into the mammalian brain. Transpl Proc. 1997;29:1926–1928. doi: 10.1016/s0041-1345(97)00164-4. [DOI] [PubMed] [Google Scholar]
- 138.Saporta S, Cameron DF, Borlongan CV, Sanberg PR. Survival of rat and porcine Sertoli cell transplants in the rat striatum without cyclosporine-A immunosuppression. Exp Neurol. 1997;146:299–304. doi: 10.1006/exnr.1997.6493. [DOI] [PubMed] [Google Scholar]
- 139.Smith GA, Snyder EY. Two cells are better than one: Optimizing stem cell survival by co-grafting “helper” cells that offer regulated trophic support. Exp Neurol. 2013;247:751–754. doi: 10.1016/j.expneurol.2013.07.003. [DOI] [PubMed] [Google Scholar]
- 140.Liang Y, Agren L, Lyczek A, Walczak P, Bulte JW. Neural progenitor cell survival in mouse brain can be improved by co-transplantation of helper cells expressing bFGF under doxycycline control. Exp Neurol. 2013;247:73–79. doi: 10.1016/j.expneurol.2013.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Liang Y, Walczak P, Bulte JW. The survival of engrafted neural stem cells within hyaluronic acid hydrogels. Biomaterials. 2013;34:5521–5529. doi: 10.1016/j.biomaterials.2013.03.095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Sanberg PR, Eve DJ, Cruz LE, Borlongan CV. Neurological disorders and the potential role for stem cells as a therapy. Br Med Bull. 2012;101:163–181. doi: 10.1093/bmb/lds001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Borlongan CV, Chopp M, Steinberg GK, Bliss TM, Li Y, Lu M, Hess DC, Kondziolka D. Potential of stem/progenitor cells in treating stroke: The missing steps in translating cell therapy from laboratory to clinic. Regen Med. 2008;3:249–250. doi: 10.2217/17460751.3.3.249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Borlongan CV. Cell therapy for stroke: Remaining issues to address before embarking on clinical trials. Stroke. 2009;40:146–148. doi: 10.1161/STROKEAHA.108.533091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Chopp M, Steinberg GK, Kondziolka D, Lu M, Bliss TM, Li Y, Hess DC, Borlongan CV. Who's in favor of translational cell therapy for stroke: STEPS forward please? Cell Transpl. 2009;18:691–693. doi: 10.3727/096368909X470883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Borlongan CV, Weiss MD. Baby STEPS: A giant leap for cell therapy in neonatal brain injury. Pediatr Res. 2011;70:3–9. doi: 10.1203/PDR.0b013e31821d0d00. [DOI] [PMC free article] [PubMed] [Google Scholar]