

Abstract
Background
Alzheimer neuropathology (AD) is found in almost half of patients with non-semantic primary progressive aphasia (PPA). This study examined hippocampal abnormalities in PPA to determine similarities to those described in amnestic AD.
Methods
In 37 PPA patients and 32 healthy controls, we generated hippocampal subfield surface maps from structural MRIs and administered a face memory test. We analyzed group and hemisphere differences for surface shape measures and their relationship with test scores and ApoE genotype.
Results
The hippocampus in PPA showed inward deformity (CA1 and subiculum subfields) and outward deformity (CA2-4+DG subfield) and smaller left than right volumes. Memory performance was related to hippocampal shape abnormalities in PPA patients, but not controls, even in the absence of memory impairments.
Conclusions
Hippocampal deformity in PPA is related to memory test scores. This may reflect a combination of intrinsic degenerative phenomena with transsynaptic or Wallerian effects of neocortical neuronal loss.
Keywords: frontotemporal dementia, lobar degeneration, multi-atlas mapping, structural magnetic resonance imaging (MRI), memory, neuroanatomy, primary progressive aphasia (PPA), Alzheimer’s disease (AD)
1. Introduction
The clinical course of primary progressive aphasia (PPA) is characterized by initial progressive loss of language abilities and relative preservation of other cognitive functions, such as episodic memory.[1] There are three subtypes of PPA, each based on the most prominent type of language deficit in the clinical profile,[2] but the preservation of memory is most easily demonstrated in non-semantic PPA subtypes, namely, the agrammatic and logopenic variants, since they have relatively preserved language comprehension. Neuropathologically, PPA is heterogeneous: while the amyloid plaques and neurofibrillary tangles of Alzheimer’s disease (AD) are predominant in the brains of some PPA patients, frontotemporal lobar degeneration (FTLD) pathology is predominant in others.[3–5] Post-mortem studies have revealed that the prevalence of AD pathology in non-semantic PPA subtypes ranges from 30–60%,[4–7] and is highest in the logopenic subtype.[4, 6, 8] Although significant accumulation of neurofibrillary AD pathology can be seen in the hippocampus/entorhinal cortex of PPA patients, its entorhinal-to-neocortical ratio is lower in PPA when compared to this ratio in post mortem analysis of individuals with the more typical amnestic dementia of AD.[3]
Medial temporal neurofibrillary pathology is the most characteristic feature of AD pathophysiology.[9, 10] In AD associated with the amnestic dementia of the AD type (DAT), the neurofibrillary tangles accumulate in the entorhino-hippocampal complex at the earliest stages of the disease, even prior to symptom onset,[11–13] and lead to regional atrophy of hippocampal subfields[14] and entorhinal[15] cortices as the disease progresses. In DAT, deficits in visual and verbal memory are known to correlate with both post-mortem neuropathological measures[16] and in vivo atrophy of the entorhinal cortex and hippocampus.[11]
While neuroimaging has revealed atrophy of the left hemisphere language network in PPA,[17] less consistency is found in the hippocampus. For example, van de Pol et al.[18] found no significant hippocampal atrophy in patients with progressive non-fluent aphasia, but Gorno-Tempini et al.[19] found atrophy of the anterior left hippocampus in a non-semantic logopenic subtype. A study of 9 patients with non-semantic progressive non-fluent aphasia and 21 controls found no overall total hippocampal volume difference between groups, although the left was smaller than the right in the patient group.[20] Using spherical harmonics, the same study found inward deformity in the patients’ left hippocampus relative to the controls. These studies tended to have small sample sizes and all but one study examined only volumes but not shape, factors that may have contributed to inconsistent results.
The goal of the present study was to compare the shape of the hippocampal surface including its subfields between PPA subjects and matched controls. We predicted a spectrum of hippocampal deformity in PPA based on the fact that some might have AD neuropathology whereas others might not. We also predicted a greater degree of deformity in the left hippocampus, reflective of the left hemispheric focus of PPA. Study methodology involved a new multi-atlas FreeSurfer-initiated Large-Deformation Diffeomorphic Metric Mapping (ma-FSLDDMM) procedure for mapping of the hippocampal structure, and the assessment of nonverbal memory to correlate with hippocampal morphology. ma-FSLDDMM was based on our previously published single-atlas technique (sa-FSLDDMM),[21, 22] for automated brain segmentation in high resolution structural scans which combined initial FreeSurfer segmentation of gray and white matter structures (http://surfer.nmr.mgh.harvard.edu) with a smoothed approximation via Large-Deformation Diffeomorphic Metric Mapping (LDDMM).[23] Compared with single-atlas methods, approaches that combine maps from multiple atlases that best match any individual subject’s scan features have been shown to improve segmentation accuracy and reduce biases.[24] We chose the nonverbal Faces subtest in the Wechsler Memory Scale III (WMS-III)[ 25] to assess episodic memory. Although even an apparently nonverbal memory test can elicit internal verbalization [26], this choice avoided the pitfalls of using story or word-list recall to test episodic memory in patients with PPA. [19, 27] Finally, the ε4 allele of the apoliprotein ε (ApoE) gene has been revealed as a risk factor for amnestic but not aphasic dementias [28]. Because genotype does not predict AD pathology in PPA, we hypothesized that the presence of the ApoE ε4 allele would not influence a patient’s hippocampal shape or memory performance.
2. Methods
2.1 Participants and assessments
The present study consisted of analysis of data derived from a larger PPA Research Program at Northwestern University Feinberg School of Medicine, and included 37 PPA and 32 control participants. The protocol for recruitment, comprehensive assessment of language and non-language cognitive functions, MRI scanning, and ApoE genotyping was approved by the Institutional Review Board of Northwestern University; informed consent was obtained prior to evaluation.
All participants were right-handed. Diagnoses were established by consensus from experienced clinicians (MM, SW) according to previously published criteria[2] based on clinical interview, cognitive testing with the Uniform Data Set of the NIA Alzheimer Disease Centers program,[29] and review of prior diagnostic tests such as MRI and PET scans. ApoE genotyping was completed at Northwestern University using previously described methods [28]. PPA participants had obtained diagnoses of PPA-G (nonfluent, agrammatic) and PPA-L (logopenic) variants, referred to in this paper as non-semantic variants. Aphasia severity was assessed using the Aphasia Quotient score from the Western Aphasia Battery (WAB)[30] containing subtests of auditory comprehension, naming, repetition, and spontaneous speech.
Episodic memory was assessed using the Wechsler Memory Scale-III Faces subtest of visual-nonverbal recognition memory.[25] A similar test, the Warrington Recognition Memory Test (RMT), which assesses immediate recognition of both words and faces, has been shown to be sensitive to hippocampal damage.[31] The WMS-III Faces was chosen due to the addition of a delayed recognition condition while the RMT tests only immediate recognition. On tests of visual memory, particularly memory for faces, participants benefit from mental verbalization of stimuli [26]. However, because WMS-III Faces is primarily visually-mediated, this test may to a degree circumvent verbal deficits of PPA. Because face encoding involves primarily the right medial temporal lobe and right dorsal frontal cortex [32], PPA patients’ greater left-sided cortical atrophy [33] may disrupt the face encoding system to a lesser extent than other modalities. Participants first studied 24 faces, presented serially for 2 seconds each. Immediately afterwards, these faces were presented serially amongst 24 novel faces, and the participant selected the previously presented faces. Then, following a 25–35 minute delay, participants were given a similar test of the 24 original faces, to be discriminated from yet a third set of novel faces. Accuracy scores (hits) from these immediate and delayed recognition tests were recorded.
2.2 MR scanning and ma-FSLDDMM
MPRAGE sequences (TR=2.3sec, TE=2.86msec, flip angle=9°, FOV=256mm, 160 slices, resolution=1×1×1mm3) were obtained on a Siemens TIM TRIO 3-Tesla system using a 12-channel head coil.
Hippocampal segmentations were generated using ma-FSLDDMM, which consisted of a 38-scan multi-atlas library and their expert manual segmentations. The manual segmentations followed the delineation protocol described in Haller et al.[34] See Table 1 for descriptions of the multi-atlas library and Figure 1 for descriptions of the ma-FSLDDMM procedure.
Table 1. Multi-atlas library for ma-FSLDDMM.
This library is composed of 38 scans and their expert manual segmentations. Manual segmentations followed the delineation protocol described in Haller et al.[34]
| Study Source/Reference | Brief Description, Scan Type | N (group) | Demographics Mean (SD) Age, M/F |
|---|---|---|---|
| Current study | 3T scans manually segmented by one of the authors (AC/JR) | 14 (Control) | Age=61.9 (7.2), M/F=7/7 |
| Alzheimer’s Disease Neuroimaging Initiative[54] | 1.5T scans manually segmented by one of the authors (AC) | 10 (3 Control/3 MCI/4 AD) | Control Age=75.3 (2.1), M/F=0/3 MCI Age=67.7 (10.2), M/F=1/2 AD, Age=76.8 (5.9), M/F=1/3 |
| Unpublished Data | 3T scans with manual segmentation (LW) | 8 (Control) | Age= 75.4 (8.3), M/F=6/2 |
| University of New South Wales Memory and Ageing Study[55] | 3T scans with manual segmentation (Sachdev et al) | 6 (Control) | Age= 77.1 (4.5), M/F=4/2 |
Abbreviations: ma, multi-atlas; FSLDDMM, FreeSurfer-initiated Large-Deformation Diffeomorphic Metric Mapping; MCI, mild cognitive impairment patients; AD, Alzheimer’s disease patients
Figure 1. Atlas-Based Segmentation ma-FSLDDMM with Selection, Illustrated for a Single Subject.

A, 38 individual sa-FSLDDMM hippocampal segmentations and a voxel-wise average were first generated for the subject. B, surfaces with corresponding vertices were created for each of the 38 segmentations and the average, using a previously developed template surface injection procedure.[35, 36] C, sum-of-squared errors between the average surface and each sa-FSLDDMM surface were calculated. D, Across all subjects, 14 sa-FSLDDMM maps consistently showed substantially larger error than others and were subsequently removed from further analysis. E, new average segmentation.
Abbreviations: ma, multi-atlas; sa, single-atlas; FSLDDMM, FreeSurfer-initiated Large-Deformation Diffeomorphic Metric Mapping
In the ma-FSLDDMM procedure, 38 individual sa-FSLDDMM hippocampal segmentations and a voxel-wise average were first generated for each subject (Figure 1-A, B). Next, surfaces with corresponding vertices were created for each of the 38 segmentations and the average, using a previously developed template surface injection procedure (Figure 1-B).[35, 36] Then, sum-of-squared errors between the average surface and each sa-FSLDDMM surface were calculated (Figure 1-C). Across all subjects, 14 sa-FSLDDMM maps consistently showed substantially larger error than others and were subsequently removed from further analysis (Figure 1-D), giving rise to a new average segmentation (Figure 1-E). Finally, an expert rater (AC) inspected the new average segmentation embedded in the MR scan, and made minor manual edits whenever necessary. We separately demonstrated high reliability for manual editing in 10 randomly selected scans (5 patients and 5 controls) – volume intraclass correlation coefficient was 0.945 (AC/JR). A final hippocampal surface was generated by injection.[35, 36]
The final surface was adjusted for individual head size by multiplying vertex coordinates by a scaling factor, defined as the cubic root of (population average intracranial volume/individual intracranial volume),[37] where intracranial volumes were provided by FreeSurfer. Adjusted hippocampal volume was calculated as volume enclosed within the hippocampal surface. Adjusted hippocampal surfaces were used in subsequent shape analysis.
2.3 Hippocampal shape and volume comparison
To visualize between-group shape differences, whole hippocampal surfaces were first rigidly aligned into a previously established template space,[36] and a population average was generated. Perpendicular displacements were computed between corresponding vertices from this average to each subject. T-scores between the patient and control displacements were calculated and visualized. The CA1, subiculum, and combined CA2, 3, 4 and dentate gyrus (CA2-4+DG) hippocampal subfields were delineated on the surface along previously defined and validated borders during the template injection procedure.[38] Previously, a team of neuroanatomy experts had manually segmented the CA1, CA2, CA3, CA4, DG, and subiculum subfields in coronal sections of the template MR scan[38] using reference sections based on the Duvernoy neuroanatomical atlas.[39] These subfield segmentations were projected onto the template surface. CA2, CA3, CA4, and DG were combined into a single CA2-4+DG as in prior studies because of their limited size and hence limited accuracy and reliability of defining each individually.[12, 40] However, this combination restricts the ability to make specific inferences for these individual subfields. Shape measures were calculated for the whole hippocampus, and individually for each of the three subfields via principal component analysis (PCA) of the displacements. Ten principal components (PCs) accounting for at least 90% of the variance in whole or subfield displacements were used in statistical analyses.
The data for the present study consisted of volume, shape PC scores for the whole hippocampus, shape PC scores for each subfield, memory test scores, and demographic variables. Age and education were used as covariates because normative scores on cognitive tests decline with age, and are frequently higher for those with advanced education.[25] Statistical analyses were performed using SPSS.[41]
Main effects of group (between-subjects factor), hemisphere (within-subjects repeated factor), and group-by-hemisphere interactions on hippocampal volume and on whole hippocampus and subfield shape PC scores were tested using repeated-measures analysis of variance (RM-ANOVA)(within subject repeated PC factor was not tested.). Significance was corrected for multiple comparisons of 3 subfields by setting the significance level at p=0.017 (i.e., 0.05/3). WMS-III Faces scores were analyzed with analysis of covariance (ANCOVA) and independent sample t-tests (ApoE present/absent).
Correlations between measures of shape and cognitive performance were explored within the PPA group. To do this, shape summary scores were generated for each subject, separately for the whole hippocampus and each subfield in each hemisphere, by applying stepwise logistic regression procedures to the respective shape PC scores. These scalar summary scores represented a continuum of overall shape variation (i.e., as opposed to individual PCs which represented dimensions of shape variation) such that a more positive shape summary score would indicate a more abnormal shape. Pearson correlation coefficients between the summary scores and WMS-III Faces Immediate and Delay scores were computed within the PPA group and within the control group separately. Significance was not corrected for number of subfields and type of WMS-III Faces scores (immediate or delayed) due to the exploratory nature of the analysis.[42]
3. Results
3.1 Participants and assessments
PPA and controls did not differ in age, gender, and education, nor in WMS-III Faces raw scores and scaled scores (the latter based on comparison to published norms)[25] (Table 2). Across groups, the raw scores on the WMS-III Faces tests were positively correlated with education as expected (Immediate: Pearson’s r=.35, p=.005; Delay: r=.40, p=.001), and negatively correlated with age only for delayed memory (Immediate: r=−.16, p=.20; Delay: r=−.25, p=.049). Normatively, on the WMS-III Faces test, three PPA patients performed in the mildly impaired range (i.e., equivalent to the 9th percentile) for Immediate Recognition, and one of those three also for Delayed Recognition. Two control subjects also performed in the mildly impaired range for Immediate Recognition, and one for Delayed Recognition.
Table 2. Subject characteristics.
Demographic information of the study sample and WMS-III Faces raw and scaled scores for Immediate and Delayed recognition. On the WMS-III Faces test, three PPA patients performed in the mildly impaired range for Immediate Recognition, with one of those three also for Delayed Recognition. Two control subjects also performed in the mildly impaired range for Immediate Recognition, and one for Delayed Recognition (normal: scaled score greater than 6, mild impairment: 5–6, moderate: 3–4, severe: 1–2). The WAB was administered only to PPA patients, because healthy adults are expected to obtain a perfect score of 100. Lower scores signify increasing language deficits. [56]
| PPA n = 37 | Control n = 32 | |
|---|---|---|
| M/F (% Male) | 16/21 (43.2%) | 16/16 (50%) |
| Mean (SD) Age in years | 64.9 (7.2) | 62.5 (7.0) |
| Mean (SD) Education in years | 16.1 (2.1) | 15.8 (2.5) |
| Race (Caucasian/Asian/African American) | 37/0/0 | 26/1/5* |
| Mean (SD) [Range] WAB Aphasia Quotient |
86.9 (7.5) [73.9–97.2] | -- |
| Mean (SD) [Range] | n = 34 | n = 28 |
|---|---|---|
| Immediate Raw Score | 35.9 (5.0) [26–44] | 37.4 (3.8) [27–43] |
| Delay Raw Score | 37.3 (4.3) [28–45] | 37.6 (3.5) [29–45] |
| Immediate Scaled Score | 11.1 (3.3) [6–17] | 11.7 (2.7) [6–17] |
| Delay Scaled Score | 12.5 (3.3) [6–18] | 12.3 (2.6) [6–18] |
p < .01
Abbreviations: PPA, primary progressive aphasia; WMS-III, Wechsler Memory Scale, Third Edition; SD, standard deviation, WAB, Western Aphasia Battery
3.2 Volume and shape comparison
There was no group difference for whole hippocampal volume [mean(SD) = PPA: left=1912(240) mm3, right=1992(238) mm3; Control: left=1993(204) mm3, right=1977(188) mm3; F(1,65)=2.4, p=.13]. Whole hippocampal shape comparison using PC scores showed significant group difference [F(9,57)=6.9, p<.001], with both inward (indicated by cooler shades) and outward deformities (indicated by warmer shades) observed in PPA (Figure 2). CA1 and subiculum subfields showed mostly inward deformity [F(9,57)=3.2, p<.01; F(9,57)=2.8, p<.01, respectively], and CA2-4+DG showed mostly outward deformity [F(9,57)=4.3, p<.001].
Figure 2. Shape comparison (T-score) between PPA vs. control subjects.
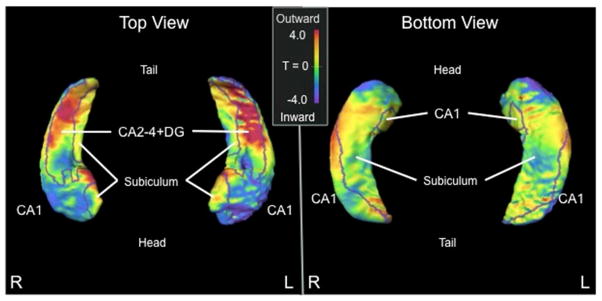
Cooler shades represent greater inward deformity of the PPA group relative to controls, while warmer shades represent greater outward deformity of the PPA group relative to controls. Borders on the surfaces indicate the CA1, subiculum, and CA2-4+DG subfield divisions.
Abbreviations: PPA, primary progressive aphasia; CA2-4+DG, CA2, 3, 4 and dentate gyrus
With regard to left-right asymmetry, a significant group-by-hemisphere interaction was found in volume [F(1,65)=9.7, p=.003], primarily driven by a leftward asymmetry of volume loss within the PPA group. Also, significant group-by-hemisphere interactions were found in the shape of the whole hippocampus [F(9,57)=4.5, p<.001], and in CA1 [F(9,57)=3.0, p<.01] and CA2-4+DG [F(9,57)=4.0, p<.001] subfields, driven by a greater effect of left hippocampal deformity (e.g., whole hippocampus mean PC effect size: left ηp2 = .06, right ηp2 = .03).
3.3 Correlation of hippocampal shape and volume with visual memory scores
Hippocampal shape summary scores were derived from discriminant function analyses (i.e., stepwise logistic regression) on the PC scores, and were positive for the PPA (i.e., diseased) and negative for the controls across all PCs (Table 3). Because these summary scores represent a continuum of overall shape variation, they were used to examine the relationship between shape and cognition.
Table 3. Mean hippocampal shape summary scores in PPA and control subjects.
Note that the mean scores for the PPA subjects are positive and for controls are negative. This signifies that scores that are more positive represented hippocampal shape that deviated farther from the control group and are indicative of PPA characteristics, while negative scores are indicative of control characteristics.
| Mean (SD) | PPA | Control |
|---|---|---|
| Whole Hippocampus, Left | 0.97 (1.26) | −0.67 (1.24) |
| Whole Hippocampus, Right | 0.49 (0.76) | −0.27 (1.00) |
| CA1, Left | 1.09 (1.56) | −0.68 (1.24) |
| CA1, Right | 0.97 (1.25) | −0.72 (1.43) |
| Subiculum, Left | 0.68 (1.18) | −0.41 (0.92) |
| Subiculum, Right | 0.43 (0.75) | −0.15 (0.79) |
| CA2-4+DG, Left | 1.44 (1.75) | −1.06 (1.50) |
| CA2-4+DG, Right | 0.64 (0.85) | −0.47 (1.26) |
Abbreviations: PPA, primary progressive aphasia; CA2-4+DG, CA2, 3, 4 and dentate gyrus
Within the PPA group, lower immediate visual memory scores were correlated with increased shape deformity in the right whole hippocampus (r=−.50, p<.01), CA1 (r=−.48, p=.01), and subiculum (r=−.34, p=.05) (Figure 3A–C). There were trends toward significance in correlation of the shape abnormalities of the left CA1 and CA2-4+DG with immediate visual memory performance (r=−.31, p=.08; r=−.33, p=.06; respectively) (Figure 3F,H). There was no correlation between hippocampal volume and immediate visual memory scores. No significant correlations were observed within the control group.
Figure 3. Correlation between hippocampal shape scores and visual memory (WMS-III Faces) performance within the PPA group.

Shape scores that are more positive represent a hippocampal shape that deviates farther from the average control. A–D. Correlation between hippocampal shape scores in the whole right hippocampus and its subfields and immediate visual memory (WMS-III Faces: Immediate scores) within the PPA group. A: Whole hippocampal shape score vs. WMS-III Faces Immediate score (p<.01). B: CA1 subfield shape score vs. WMS-III Faces Immediate score (p=.01). C: Subiculum subfield shape score vs. WMS-III Faces Immediate score (p=.05). D: CA2-4+DG subfield shape score vs. WMS-III Faces Immediate score (p=.20). Declines in immediate visual memory performance were correlated with increased shape deformity in the right whole hippocampus, CA1, and subiculum. E–H. Correlation between hippocampal shape scores in the whole left hippocampus and its subfields and immediate visual memory (WMS-III Faces: Immediate scores). E: Whole hippocampal shape score vs. WMS-III Faces Immediate score (p=.99). F: CA1 subfield shape score vs. WMS-III Faces Immediate score (p=.08). G: Subiculum subfield shape score vs. WMS-III Faces Immediate score (p=.11). H: CA2-4+DG subfield shape score vs. WMS-III Faces Immediate score (p=.06). There were trends toward significance in the left CA1 and CA2-4+DG. I–L. Correlation between hippocampal shape scores in the whole right hippocampus and its subfields and delayed visual memory (WMS-III Faces: Delay scores). I: Whole hippocampal shape score vs. WMS-III Faces Delay score (p=.10). J: CA1 subfield shape score vs. WMS-III Faces Delay score (p=.25). K: Subiculum subfield shape score vs. WMS-III Faces Delay score (p=.12). L: CA2-4+DG subfield shape score vs. WMS-III Faces Delay score (p=.09). There were trends toward significance in the right whole hippocampus and CA2-4+DG. M–P. Correlation between hippocampal shape scores in the whole left hippocampus and its subfields and delayed visual memory (WMS-III Faces: Delay scores). M: Whole hippocampal shape score vs. WMS-III Faces Delay score (p=.77). N: CA1 subfield shape score vs. WMS-III Faces Delay score (p=.03). O: Subiculum subfield shape score vs. WMS-III Faces Delay score (p=.28). P: CA2-4+DG subfield shape score vs. WMS-III Faces Delay score (p=.15). Declines in delayed visual memory performance were correlated with increased shape deformity in the left CA1.
Abbreviations: PPA, primary progressive aphasia; WMS-III, Wechsler Memory Scale, Third Edition; CA2-4+DG, CA2, 3, 4 and dentate gyrus
Lower delayed visual memory scores were correlated with the measure of increased shape deformity in the left CA1 (r=−.38, p =.03) (Figure 3N). There were trends toward significance in the correlation of shape of the right whole hippocampus and CA2-4+DG with reduced delayed visual memory (r=−.30 p=.10; r=−.31, p=.09; respectively) (Figure 3I,L). There was no correlation between hippocampal volume and delayed visual memory scores. No significant correlations were observed within the control group.
3.4 ApoE genotyping
Of the 33 PPA patients in the present study with both ApoE genotyping and memory data, 21% (n=7) had the AD risk factor allele, ε4. T-tests yielded no significant difference in memory scores between those with the allele present and absent [Raw Scores: Immediate, t(31)=1.2, p=.24; Delay, t(31)=0.7, p=.49; Scaled Scores: Immediate, t(31)=1.5, p=.15; Delay, t(31)=0.8, p=.40) (Table 4). In fact, compared to normative data (WMS scaled scores), the three PPA patients in the study with mild impairment in memory all did not possess an ε4 allele, and those with an ε4 allele had average or better memory. T-tests on shape summary scores for the whole hippocampus and each subfield in both hemispheres yielded no significant differences between ApoE present/absent groups (all p>.25).
Table 4.
ApoE Genotyping in PPA.
| ε4 Allele Absent n = 26 (79%) |
ε4 Allele Present n = 7 (21%) |
|
|---|---|---|
| ApoE Allele | ε2/ε2 (1) ε3/ε3 (25) | ε3/ε4 (6) ε4/ε4 (1) |
|
| ||
|
Mean (SD) [Range]
| ||
| Immediate Raw Score | 35.5 (5.1) [26–44] | 38.0 (4.5) [30–43] |
| Delay Raw Score | 37.0 (4.7) [28–45] | 38.3 (2.2) [35–41] |
| Immediate Scaled Score | 10.7 (3.1) [6–17] | 12.7 (3.1) [8–17] |
| Delay Scaled Score | 12.2 (3.5) [6–18] | 13.4 (2.2) [11–17] |
Abbreviations: apoliprotein ε, ApoE; PPA, primary progressive aphasia; WMS-III, Wechsler Memory Scale, Third Edition; SD, standard deviation
4. Discussion
Post-mortem studies suggest significant heterogeneity in the neuropathologic diagnosis associated with non-semantic PPA, with 30–60% having atypically distributed AD pathology.[4–7] For this study, we hypothesized that because AD pathology would be present in a portion of the patients studied, hippocampal changes would be variable and some would share common features with those reported in typical amnestic forms of AD.[12, 43] In fact, compared to typical hippocampal patterns reported in studies of DAT,[14] we found similarities, in the inward deformity of CA1 and subiculum, but also differences, in the outward deformity of CA2-4+DG.
The PPA patients showed asymmetry in a pattern of smaller left than right hippocampi that was not observed in the controls, and bilateral shape deformities that were more pronounced in the left hippocampus than the right compared to the controls. Moreover, the volume and shape asymmetry indicates that shape deformity in the left hippocampus is accompanied by volume loss, and abnormality of the left hippocampus is a consistent finding in previous PPA studies.[18, 20, 44, 45] Further, although there was no absolute episodic memory impairment, inter-individual variations in delayed face recognition memory performance correlated with left CA1 shape measures in PPA whereas variations in the immediate memory performance correlated with right whole hippocampal shape deformity. In both instances, increased shape deformity was correlated with worse episodic memory performance, and no such correlations were found in controls. These observed patterns in delayed memory are consistent with the known dominant role of the left hippocampus in all types of episodic memory, and the patterns for immediate memory are consistent with the role of the right temporal lobe in the working memory functions underlying face processing [32, 46]. Specifically, positron emission tomography of healthy subjects has revealed that while the right hippocampus is essential for face processing, the left is recruited during episodic memory judgments of face stimuli[46]. Furthermore, imaging of healthy subjects making judgments on face stimuli revealed early right hippocampal involvement and gradually increasing left hippocampal involvement only with greater memory load or time interval. [47, 48]
We also found that PPA patients with the ApoE ε4 allele did not show impaired memory or different hippocampal shape compared to PPA patients with other ApoE genotypes. A pattern of memory impairment and hippocampal atrophy consistent with amnestic dementia was not expected in the PPA patients with an ε4 allele, because the ε4 allele in this diagnostic group does not increase the accuracy of predicting underlying AD neuropathology. [28] Although postmortem data were not available in this study, we would expect that the neuropathologic source of localized hippocampal changes in PPA may vary. In some PPA patients with AD pathology, the hippocampus may contain neurofibrillary tangles and amyloid plaques that cause neuronal death and morphological distortion within the hippocampus.[3] In AD cases as well as in those with FTLD pathology, peak neurodegeneration sites in the PPA patients may encompass neocortical regions that project to the hippocampus. Prefrontal and temporal cortices in PPA patients have shown atrophy as the disease progresses.[17] Tracing studies of hippocampal connectivity in the rhesus monkey have revealed corresponding afferent and efferent cortical projections in the hippocampal formation, particularly througout the CA1 and subicular subfields.[49, 50] Diffusion tensor imaging studies in PPA have also suggested damage to white matter tracts that project to atrophic cortical areas.[51, 52] The particular mechanism whereby cortical atrophy, white matter damage, and hippocampal CA1/subiculum deformity correspond has yet to be determined. Alterations of shape through Wallerian or transsynaptic mechanisms are potential explanations. Furthermore, while some studies have examined overall cortical and subcortical densities in aphasic and amnestic AD,[3, 53] differences in the focal distribution of pathology amongst hippocampal subfields has yet to be determined. Therefore, future work in these areas will be integral to the understanding of hippocampal shape change in PPA.
The greater abnormality of the left hippocampus may reflect the asymmetrically greater degeneration of the language-dominant (usually left) hemisphere in PPA. Regardless of the underlying mechanisms, the resultant hippocampal abnormalities seem to have been functionally compensated since the patients on average did not evidence recognition memory impairments compared to healthy individuals. However, the morphological abnormalities were not inconsequential since they were related to interindividual variations in performance on memory tasks. In the future such in vivo investigations can be combined with post-mortem or biomarker data concerning the underlying pathology, such as tau and amyloid PET imaging, in order to explore the cellular basis of hippocampal deformities and their functional impact.
Finally, our analysis indicated that while overall hippocampal volume could not distinguish PPA from controls, hippocampal shape measures revealed strong group differences in different subfields. This indicates that hippocampal subfield shape can be used as an in vivo marker of structural changes that are not reflected in volumetric measures in patients with PPA. Many of the individuals in this study have agreed to brain donation at the time of death which will allow for further validation of these findings.
Research in Context.
Systematic review
We reviewed the literature on in vivo neuroimaging findings on the hippocampus in non-semantic primary progressive aphasia. While the hippocampus has been well-studied in typical Alzheimer’s disease, detailed examinations have not been previously conducted for non-semantic PPA.
Interpretation
Hippocampal shape abnormalities in PPA showed both differences and similarities to the typical AD pattern. Furthermore, damage to hippocampal subregions that are related to memory impairment in typical AD are also related to level of memory ability in PPA, despite PPA patient’s preserved functional memory.
Future directions
Combining our computational anatomy methods with cellular measures from post-mortem or biomarker data will advance our understanding of specific effects of the varied pathology in this disease. Use of statistical learning algorithms on imaging data will also allow for examination of potential pathological subsets in PPA.
Acknowledgments
This study was supported in part by a grant titled Language in PPA (DC008552) from the National Institute of Deafness and Other Communication Disorders, the National Institute of Neurological Disorders and Stroke (NINDS; R01 NS075075) and by the Alzheimer Disease Core Center grant from the National Institute of Aging (AG13854) both at the Northwestern University Cognitive Neurology and Alzheimer’s Disease Center, and NIH grants R01 NR014182, P50 AG05681, P01 AG03991, P01 AG026276. The Northwestern PPA Research Program and Cognitive Neurology and Alzheimer’s Disease Center at Northwestern University’s Feinberg School of Medicine conducted clinical and cognitive examinations and collected neuroimaging data for all PPA and control subjects. Some atlas images were provided from work that was in part funded by the Alzheimer’s Association: “A Harmonized Protocol for Hippocampal Volumetry: an EADC-ADNI Effort”, grant n. IIRG -10-174022; project PI Giovanni B Frisoni, IRCCS Fatebenefratelli, Brescia, Italy; co-PI Clifford R. Jack, Mayo Clinic, Rochester, MN. We would like to thank Ashley Chong and Hande Ozergin for discussions on manual editing procedures.
Footnotes
Disclosures: Mr. Christensen: None, Ms. Alpert: None, Dr. Rogalski: NIH grant during the course of the study, Dr. Cobia: None, Dr. Rao: None, Dr. Beg: None, Dr. Weintraub: NIH grant during the course of the study, Dr. Mesulam: None, Dr. Wang: None
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Mesulam MM. Primary progressive aphasia--a language-based dementia. The New England journal of medicine. 2003;349:1535–42. doi: 10.1056/NEJMra022435. [DOI] [PubMed] [Google Scholar]
- 2.Gorno-Tempini ML, Hillis AE, Weintraub S, Kertesz A, Mendez M, Cappa SF, et al. Classification of primary progressive aphasia and its variants. Neurology. 2011;76:1006–14. doi: 10.1212/WNL.0b013e31821103e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gefen T, Gasho K, Rademaker A, Lalehzari M, Weintraub S, Rogalski E, et al. Clinically concordant variations of Alzheimer pathology in aphasic versus amnestic dementia. Brain : a journal of neurology. 2012;135:1554–65. doi: 10.1093/brain/aws076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mesulam M, Wicklund A, Johnson N, Rogalski E, Leger GC, Rademaker A, et al. Alzheimer and frontotemporal pathology in subsets of primary progressive aphasia. Annals of neurology. 2008;63:709–19. doi: 10.1002/ana.21388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Alladi S, Xuereb J, Bak T, Nestor P, Knibb J, Patterson K, et al. Focal cortical presentations of Alzheimer’s disease. Brain : a journal of neurology. 2007;130:2636–45. doi: 10.1093/brain/awm213. [DOI] [PubMed] [Google Scholar]
- 6.Hu WT, McMillan C, Libon D, Leight S, Forman M, Lee VM, et al. Multimodal predictors for Alzheimer disease in nonfluent primary progressive aphasia. Neurology. 2010;75:595–602. doi: 10.1212/WNL.0b013e3181ed9c52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Knibb JA, Xuereb JH, Patterson K, Hodges JR. Clinical and pathological characterization of progressive aphasia. Annals of neurology. 2006;59:156–65. doi: 10.1002/ana.20700. [DOI] [PubMed] [Google Scholar]
- 8.Gorno-Tempini ML, Brambati SM, Ginex V, Ogar J, Dronkers NF, Marcone A, et al. The logopenic/phonological variant of primary progressive aphasia. Neurology. 2008;71:1227–34. doi: 10.1212/01.wnl.0000320506.79811.da. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hyman BT, Vanhoesen GW, Damasio AR, Barnes CL. Alzheimers-Disease - Cell-Specific Pathology Isolates the Hippocampal-Formation. Science. 1984;225:1168–70. doi: 10.1126/science.6474172. [DOI] [PubMed] [Google Scholar]
- 10.Braak H, Braak E. Evolution of the neuropathology of Alzheimer’s disease. Acta neurologica Scandinavica Supplementum. 1996;165:3–12. doi: 10.1111/j.1600-0404.1996.tb05866.x. [DOI] [PubMed] [Google Scholar]
- 11.Apostolova LG, Morra JH, Green AE, Hwang KS, Avedissian C, Woo E, et al. Automated 3D mapping of baseline and 12-month associations between three verbal memory measures and hippocampal atrophy in 490 ADNI subjects. NeuroImage. 2010;51:488–99. doi: 10.1016/j.neuroimage.2009.12.125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wang L, Miller JP, Gado MH, McKeel DW, Rothermich M, Miller MI, et al. Abnormalities of hippocampal surface structure in very mild dementia of the Alzheimer type. NeuroImage. 2006;30:52–60. doi: 10.1016/j.neuroimage.2005.09.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Price JL, Morris JC. Tangles and plaques in nondemented aging and “preclinical” Alzheimer’s disease. Annals of neurology. 1999;45:358–68. doi: 10.1002/1531-8249(199903)45:3<358::aid-ana12>3.0.co;2-x. [DOI] [PubMed] [Google Scholar]
- 14.Csernansky JG, Hamstra J, Wang L, McKeel D, Price JL, Gado M, et al. Correlations between antemortem hippocampal volume and postmortem neuropathology in AD subjects. Alzheimer disease and associated disorders. 2004;18:190–5. [PubMed] [Google Scholar]
- 15.Price JL, Ko AI, Wade MJ, Tsou SK, McKeel DW, Morris JC. Neuron number in the entorhinal cortex and CA1 in preclinical Alzheimer disease. Archives of neurology. 2001;58:1395–402. doi: 10.1001/archneur.58.9.1395. [DOI] [PubMed] [Google Scholar]
- 16.Guillozet AL, Weintraub S, Mash DC, Mesulam MM. Neurofibrillary tangles, amyloid, and memory in aging and mild cognitive impairment. Archives of neurology. 2003;60:729–36. doi: 10.1001/archneur.60.5.729. [DOI] [PubMed] [Google Scholar]
- 17.Rogalski E, Cobia D, Harrison TM, Wieneke C, Weintraub S, Mesulam MM. Progression of language decline and cortical atrophy in subtypes of primary progressive aphasia. Neurology. 2011;76:1804–10. doi: 10.1212/WNL.0b013e31821ccd3c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.van de Pol LA, Hensel A, van der Flier WM, Visser PJ, Pijnenburg YA, Barkhof F, et al. Hippocampal atrophy on MRI in frontotemporal lobar degeneration and Alzheimer’s disease. Journal of neurology, neurosurgery, and psychiatry. 2006;77:439–42. doi: 10.1136/jnnp.2005.075341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gorno-Tempini ML, Dronkers NF, Rankin KP, Ogar JM, Phengrasamy L, Rosen HJ, et al. Cognition and anatomy in three variants of primary progressive aphasia. Annals of neurology. 2004;55:335–46. doi: 10.1002/ana.10825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lindberg O, Walterfang M, Looi JC, Malykhin N, Ostberg P, Zandbelt B, et al. Hippocampal shape analysis in Alzheimer’s disease and frontotemporal lobar degeneration subtypes. Journal of Alzheimer’s disease : JAD. 2012;30:355–65. doi: 10.3233/JAD-2012-112210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Khan AR, Wang L, Beg MF. Multistructure large deformation diffeomorphic brain registration. IEEE transactions on bio-medical engineering. 2013;60:544–53. doi: 10.1109/TBME.2012.2230262. [DOI] [PubMed] [Google Scholar]
- 22.Wang L, Khan A, Csernansky JG, Fischl B, Miller MI, Morris JC, et al. Fully-Automated, Multi-Stage Hippocampus Mapping in Very Mild Alzheimer Disease. Hippocampus. 2009;19:541–8. doi: 10.1002/hipo.20616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Beg MF, Miller MI, Trouve A, Younes L. Computing Large Deformation Metric Mappings via Geodesic Flows of Diffeomorphisms. International Journal of Computer Vision. 2005;61:139. [Google Scholar]
- 24.Aljabar P, Heckemann RA, Hammers A, Hajnal JV, Rueckert D. Multi-atlas based segmentation of brain images: atlas selection and its effect on accuracy. NeuroImage. 2009;46:726–38. doi: 10.1016/j.neuroimage.2009.02.018. [DOI] [PubMed] [Google Scholar]
- 25.Wechsler D. Wechsler Memory Scale-third edition. San Antonio, TX: The Psychological Corporation; 1997. [Google Scholar]
- 26.Brown C, Lloyd-Jones TJ. Beneficial effects of verbalization and visual distinctiveness on remembering and knowing faces. Memory & cognition. 2006;34:277–86. doi: 10.3758/bf03193406. [DOI] [PubMed] [Google Scholar]
- 27.Weintraub S, Rogalski E, Shaw E, Sawlani S, Rademaker A, Wieneke C, et al. Verbal and nonverbal memory in primary progressive aphasia: the Three Words-Three Shapes Test. Behavioural neurology. 2013;26:67–76. doi: 10.3233/BEN-2012-110239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Rogalski EJ, Rademaker A, Harrison TM, Helenowski I, Johnson N, Bigio E, et al. ApoE E4 is a susceptibility factor in amnestic but not aphasic dementias. Alzheimer disease and associated disorders. 2011;25:159–63. doi: 10.1097/WAD.0b013e318201f249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Weintraub S, Salmon D, Mercaldo N, Ferris S, Graff-Radford NR, Chui H, et al. The Alzheimer’s Disease Centers’ Uniform Data Set (UDS): the neuropsychologic test battery. Alzheimer Dis Assoc Disord. 2009;23:91–101. doi: 10.1097/WAD.0b013e318191c7dd. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kertesz A. Western aphasia battery. San Antonio: The Psychological Corporation; 1982. [Google Scholar]
- 31.Miller LA, Lai R, Munoz DG. Contributions of the entorhinal cortex, amygdala and hippocampus to human memory. Neuropsychologia. 1998;36:1247–56. doi: 10.1016/s0028-3932(97)00139-5. [DOI] [PubMed] [Google Scholar]
- 32.Kelley WM, Miezin FM, McDermott KB, Buckner RL, Raichle ME, Cohen NJ, et al. Hemispheric specialization in human dorsal frontal cortex and medial temporal lobe for verbal and nonverbal memory encoding. Neuron. 1998;20:927–36. doi: 10.1016/s0896-6273(00)80474-2. [DOI] [PubMed] [Google Scholar]
- 33.Rogalski E, Cobia D, Harrison TM, Wieneke C, Thompson CK, Weintraub S, et al. Anatomy of language impairments in primary progressive aphasia. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2011;31:3344–50. doi: 10.1523/JNEUROSCI.5544-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Haller JW, Banerjee A, Christensen GE, Gado M, Joshi S, Miller MI, et al. Three-dimensional hippocampal MR morphometry with high-dimensional transformation of a neuroanatomic atlas. Radiology. 1997;202:504–10. doi: 10.1148/radiology.202.2.9015081. [DOI] [PubMed] [Google Scholar]
- 35.Qiu A, Miller MI. Multi-structure network shape analysis via normal surface momentum maps. Neuroimage. 2008;42:1430–8. doi: 10.1016/j.neuroimage.2008.04.257. [DOI] [PubMed] [Google Scholar]
- 36.Csernansky JG, Wang L, Joshi S, Miller JP, Gado M, Kido D, et al. Early DAT is distinguished from aging by high-dimensional mapping of the hippocampus. Dementia of the Alzheimer type. Neurology. 2000;55:1636–43. doi: 10.1212/wnl.55.11.1636. [DOI] [PubMed] [Google Scholar]
- 37.Styner M, Oguz I, Xu S, Brechbuhler C, Pantazis D, Levitt JJ, et al. Framework for the Statistical Shape Analysis of Brain Structures using SPHARM-PDM. The insight journal. 2006:242–50. [PMC free article] [PubMed] [Google Scholar]
- 38.Wang L, Swank JS, Glick IE, Gado MH, Miller MI, Morris JC, et al. Changes in hippocampal volume and shape across time distinguish dementia of the Alzheimer type from healthy aging. NeuroImage. 2003;20:667–82. doi: 10.1016/S1053-8119(03)00361-6. [DOI] [PubMed] [Google Scholar]
- 39.Duvernoy HM. The human hippocampus : an atlas of applied anatomy. München: J.F. Bergmann; 1988. [Google Scholar]
- 40.Csernansky JG, Wang L, Swank J, Miller JP, Gado M, McKeel D, et al. Preclinical detection of Alzheimer’s disease: hippocampal shape and volume predict dementia onset in the elderly. NeuroImage. 2005;25:783–92. doi: 10.1016/j.neuroimage.2004.12.036. [DOI] [PubMed] [Google Scholar]
- 41.IBM. IBM SPSS Statistics for Windows, Version 21.0. Armonk, NY: IBM Corp; 2012. [Google Scholar]
- 42.Bender R, Lange S. Adjusting for multiple testing--when and how? Journal of clinical epidemiology. 2001;54:343–9. doi: 10.1016/s0895-4356(00)00314-0. [DOI] [PubMed] [Google Scholar]
- 43.Apostolova LG, Dinov ID, Dutton RA, Hayashi KM, Toga AW, Cummings JL, et al. 3D comparison of hippocampal atrophy in amnestic mild cognitive impairment and Alzheimer’s disease. Brain. 2006;129:2867–73. doi: 10.1093/brain/awl274. [DOI] [PubMed] [Google Scholar]
- 44.Rohrer JD, Ridgway GR, Crutch SJ, Hailstone J, Goll JC, Clarkson MJ, et al. Progressive logopenic/phonological aphasia: erosion of the language network. NeuroImage. 2010;49:984–93. doi: 10.1016/j.neuroimage.2009.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Seeley WW, Matthews BR, Crawford RK, Gorno-Tempini ML, Foti D, Mackenzie IR, et al. Unravelling Bolero: progressive aphasia, transmodal creativity and the right posterior neocortex. Brain : a journal of neurology. 2008;131:39–49. doi: 10.1093/brain/awm270. [DOI] [PubMed] [Google Scholar]
- 46.Kapur N, Friston KJ, Young a, Frith CD, Frackowiak RSJ. Activation of Human Hippocampal Formation During Memory for Faces: A Pet Study. Cortex; a journal devoted to the study of the nervous system and behavior. 1995;31:99–108. doi: 10.1016/s0010-9452(13)80108-6. [DOI] [PubMed] [Google Scholar]
- 47.Paller KA, Ranganath C, Gonsalves B, LaBar KS, Parrish TB, Gitelman DR, et al. Neural correlates of person recognition. Learn Mem. 2003;10:253–60. doi: 10.1101/lm.57403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.McIntosh AR, Grady CL, Haxby JV, Ungerleider LG, Horwitz B. Changes in limbic and prefrontal functional interactions in a working memory task for faces. Cerebral cortex. 1996;6:571–84. doi: 10.1093/cercor/6.4.571. [DOI] [PubMed] [Google Scholar]
- 49.Blatt GJ, Rosene DL. Organization of direct hippocampal efferent projections to the cerebral cortex of the rhesus monkey: projections from CA1, prosubiculum, and subiculum to the temporal lobe. The Journal of comparative neurology. 1998;392:92–114. doi: 10.1002/(sici)1096-9861(19980302)392:1<92::aid-cne7>3.0.co;2-k. [DOI] [PubMed] [Google Scholar]
- 50.Barbas H, Blatt GJ. Topographically specific hippocampal projections target functionally distinct prefrontal areas in the rhesus monkey. Hippocampus. 1995;5:511–33. doi: 10.1002/hipo.450050604. [DOI] [PubMed] [Google Scholar]
- 51.Zhang Y, Tartaglia MC, Schuff N, Chiang GC, Ching C, Rosen HJ, et al. MRI signatures of brain macrostructural atrophy and microstructural degradation in frontotemporal lobar degeneration subtypes. Journal of Alzheimer’s disease : JAD. 2013;33:431–44. doi: 10.3233/JAD-2012-121156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Catani M, Mesulam MM, Jakobsen E, Malik F, Martersteck A, Wieneke C, et al. A novel frontal pathway underlies verbal fluency in primary progressive aphasia. Brain : a journal of neurology. 2013;136:2619–28. doi: 10.1093/brain/awt163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Josephs KA, Dickson DW, Murray ME, Senjem ML, Parisi JE, Petersen RC, et al. Quantitative neurofibrillary tangle density and brain volumetric MRI analyses in Alzheimer’s disease presenting as logopenic progressive aphasia. Brain and language. 2013;127:127–34. doi: 10.1016/j.bandl.2013.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Boccardi M, Bocchetta M, Ganzola R, Robitaille N, Redolfi A, Duchesne S, et al. Operationalizing protocol differences for EADC-ADNI manual hippocampal segmentation. Alzheimer’s & dementia : the journal of the Alzheimer’s Association. 2013 doi: 10.1016/j.jalz.2013.03.001. [DOI] [PubMed] [Google Scholar]
- 55.Sachdev PS, Brodaty H, Reppermund S, Kochan NA, Trollor JN, Draper B, et al. The Sydney Memory and Ageing Study (MAS): methodology and baseline medical and neuropsychiatric characteristics of an elderly epidemiological nondemented cohort of Australians aged 70–90 years. Int Psychogeriatr. 2010;22:1248–64. doi: 10.1017/S1041610210001067. [DOI] [PubMed] [Google Scholar]
- 56.Shewan CM, Kertesz A. Reliability and validity characteristics of the Western Aphasia Battery (WAB) The Journal of speech and hearing disorders. 1980;45:308–24. doi: 10.1044/jshd.4503.308. [DOI] [PubMed] [Google Scholar]