

Abstract
Glucocorticoids have major effects on adipose tissue metabolism. To study tissue mRNA expression changes induced by chronic elevated endogenous glucocorticoids, we performed RNA sequencing on subcutaneous adipose tissue from patients with Cushing’s disease (n=5) compared to patients with non-functioning pituitary adenomas (n=11). We found higher expression of transcripts involved in several metabolic pathways, including lipogenesis, proteolysis and glucose oxidation as well as decreased expression of transcripts involved in inflammation and protein synthesis. To further study this in a model system, we subjected mice to dexamethasone treatment for 12 weeks and analyzed their inguinal (subcutaneous) fat pads, which led to similar findings. Additionally, mice treated with dexamethasone showed drastic decreases in lean body mass as well as increased fat mass, further supporting the human transcriptomic data. These data provide insight to transcriptional changes that may be responsible for the co-morbidities associated with chronic elevations of glucocorticoids.
Keywords: Cushing’s Syndrome, lipolysis, insulin resistance, glucocorticoid, lipogenesis, RNA sequencing, transcriptome
Introduction
Cushing’s disease, or persistently high circulating levels of cortisol secondary to a pituitary adenoma, leads to significant truncal obesity and diabetes (Cushing 1932). Obesity and diabetes are major factors in morbidity and mortality in Cushing’s disease (Ntali et al. 2015). Cushing's disease is very rare, with an incidence of 1.2–2.4 per million (Lindholm et al. 2001), but iatrogenic Cushing's syndrome, caused by chronic glucocorticoid treatment, is very common and leads to similar clinical manifestations.
Numerous studies have shown that glucocorticoids have profound effects on adipose tissue metabolism, including promotion of adipocyte differentiation (Hauner et al. 1987) and induction of lipolysis and lipogenesis (Divertie et al. 1991; Samra et al. 1998; Kršek et al. 2006; Campbell et al. 2011). Glucocorticoids, through binding to the glucocorticoid receptor, exert transcriptional induction and repression of numerous genes (Reddy et al. 2009; Surjit et al. 2011). Despite the widespread chronic glucocorticoid exposure, there have been no human in vivo studies on global gene expression changes in adipose tissue in response to long-term exposure to glucocorticoids.
To study the effect of excess endogenous glucocorticoids on adipose tissue, we used RNA sequencing of adipose tissue biopsies from Cushing's disease patients and controls with non-secreting adenomas. We found a distinctive pattern of changes in many transcripts that are highly associated with Cushing's disease. Many of these genes explain previously observed metabolic effects of excess glucocorticoids described in vitro, in both animal models and in humans. These include enhanced fatty acid and triglyceride biosynthesis, protein degradation, activation of glycolysis and reductions in immune responses.
Materials and Methods
Patient Recruitment
The study was approved by the institutional review board of the University of Michigan Medical System. Written informed consent was obtained from all patients. Patients were recruited consecutively from those undergoing transsphenoidal adenomectomy at the University of Michigan for Cushing's disease or non-functioning pituitary adenoma over a 12 month period. Exclusion criteria were age <18, current hormone treatment including glucocorticoids, malignancy, inflammatory disease, diabetes type 1 and established pituitary hormone deficiencies. For each patient, a data sheet was completed including, age, sex, anthropometric measurements, diagnosis of hypertension, diabetes, results of blood tests and medications. Fasting blood samples were assayed for glucose (Siemens Advia 1800) and insulin (Life Technologies) as instructed by the manufacturers.
Subcutaneous Fat Biopsy
During the course of pituitary surgery, a routine subcutaneous fat graft for sealing the surgical field is taken immediately after anesthesia, but before glucocorticoid treatment. Approximately 500 mg of this fat graft was used in this study. ~100 mg fresh adipose tissue was utilized for ex vivo lipolysis assay, ~200 mg was snap frozen in liquid nitrogen and stored at −80°C for RNA preparation and ceramide analysis.
Lipolysis Assay
25 mg pieces of adipose tissue were pre-incubated for 15 minutes in KRBH buffer (sigma) at 37°C and then incubated for 1 hour at 37°C in 300 ml KRBH in duplicate. Glycerol was assayed in supernatants using a glycerol assay kit (Sigma) as instructed by manufacturer.
Treatment of Animals with Dexamethasone
Twenty-four C57BL/6 adult male mice were purchased from The Jackson Laboratory at nine weeks of age. Following a one-week acclimation period, mice were either treated with 1 mg/kg/day of dexamethasone (Sigma-Aldrich) in their drinking water (N=12) or used as controls (N=12). All animal procedures were approved by the University of Tennessee Health Science Center Institutional Animal Care and Use Committee. Animal body weight and body composition was determined weekly using an echoMRI 2100. Food was weighed weekly, with food intake determined as the decrease in food weight per mouse per week per cage. All mice were provided with ad libitum access to water and a standard rodent diet throughout the study. After 12 weeks of treatment, mice were fasted for 16h and were sacrificed by cervical dislocation at ZT3 after isoflurane anaesthesia. Following cervical dislocation, a sagittal incision was made along the medioventral surface of each mouse and the skin carefully pulled back to expose the subcutaneous fat depots. The incision was extended along the anterior surface of each hindlimb to allow careful dissection of the inguinal fat pads. A small incision was then made into the rectus abdominus muscle to expose the abdominal cavity. The epididymal fats pads were identified and carefully dissected out. The right fat pads from each mouse were weighed and snap frozen in liquid nitrogen for later analysis.
Insulin Tolerance Test
Insulin tolerance was assessed after 12 weeks of dexamethasone treatment (21 weeks of age). Following a six-hour fast, mice were given intraperitoneal injections of insulin (Humulin R, Lily) at a concentration of 1 mU/g. Blood glucose was determined at 15 minute intervals post-injection using a One Touch Ultra Glucometer (Lifescan).
Grip Test
Grip strength was measured at baseline, 4, 8 and 12 weeks following treatment using a Chatillon digital force gauge (AMETEK). Mice were placed on the grid having all four paws in contact with the apparatus and slowly pulled backwards by the tail. Mice were given five trials with about 10 seconds rest in between trials. Grip strength was measured by the average peak torque (N) over the five trials.
Quantitative Real-Time PCR
RNA was extracted with the PureLink RNA mini kit (Life Technologies). Synthesis of cDNA from 1 ug of RNA was performed using the High Capacity Reverse Transcription Kit (Life Technologies). cDNA and primers were added to Power SYBR Green PCR Master Mix (Life Technologies) in accordance with the manufacturer’s guidelines and subjected to quantitative real-time PCR as previously described (Lu et al., 2014). The primer sequences used are listed in Table 1. mRNA expression levels of all genes were normalized to Actb for adipose tissue and Gapdh for muscle tissue after confirming that these mRNAs are unaffected by dexamethasone treatment. Statistical tests were performed as described below based on tests of normality and homoscedasticity, then p-values were adjusted for multiple comparisons based on the number of genes tested for each tissue across this manuscript.
Table 1.
Primer sequences used for qPCR analyses.
| Gene | Forward Sequence | Reverse Sequence |
|---|---|---|
| Acaca | GCTAAACCAGCACTCCCGAT | GTATCTGAGCTGACGGAGGC |
| Aco1 | AACACCAGCAATCCATCCGT | GGTGACCACTCCACTTCCAG |
| Acsl1 | GCCTCACTGCCCTTTTCTGA | GCAGAATTCATCTGTGCCATCC |
| Acss2 | CGTTCTGTGGAGGAGCCAC | GGCATGCGGTTTTCCAGTAA |
| Actb | ATGTGGATCAGCAAGCAGGA | AAGGGTGTAAAACGCAGCTCA |
| Agpat2 | CGTGTATGGCCTTCGCTTTG | TCCATGAGACCCATCATGTCC |
| Dgat2 | AACACGCCCAAGAAAGGTGG | GTAGTCTCGGAAGTAGCGCC |
| Dhcr7 | ATGGCTTCGAAATCCCAGCA | GAACCAGTCCACTTCCCAGG |
| Dhcr24 | AGCTCCAGGACATCATCCCT | TACAGCTTGCGTAGCGTCTC |
| Fasn | GGAGGTGGTGATAGCCGGTAT | TGGGTAATCCATAGAGCCCAG |
| Fbxo32 | CTTCTCGACTGCCATCCTGG | GTTCTTTTGGGCGATGCCAC |
| Gapdh | CACTTGAAGGGTGGAGCCAA | ACCCATCACAAACATGGGGG |
| Gpam | AGCAAGTCCTGCGCTATCAT | CTCGTGTGGGTGATTGTGAC |
| Gpd1 | GTGAGACGACCATCGGCTG | TTGGGTGTCTGCATCAGGT |
| Idh1 | CTCAGAGCTCTCTTGGACCGA | CATCTCCTTGCATCTCCACCA |
| Ldhb | AAAGGCTACACCAACTGGGC | GCCGTACATTCCCTTCACCA |
| Mdh1 | GGAACCCCAGAGGGAGAGTT | TGGGGAGGCCTTCAACAAAC |
| Me1 | GGACCCGCATCTCAACAAG | TCGAAGTCAGAGTTCAGTCGTT |
| Psmd1 | TGCCAATCATGGTGGTGACA | ACACATCCTGACGTGCAGTT |
| Psmd8 | ACGAGTGGAACCGGAAGAAC | CCGTGGTTGGCAGGAAATTG |
| Rplp0 | GAAACTGCTGCCTCACATCCG | GCTGGCACAGTGACCTCACACG |
| Rplp13a | GCGGATGAATACCAACCCCT | CCTGGCCTCTCTTGGTCTTG |
| Scd1 | CACTCGCCTACACCAACGG | GAACTGGAGATCTCTTGGAGCA |
| Trim63 | GAGGGCCATTGACTTTGGGA | TTTACCCTCTGTGGTCACGC |
Ceramide Determination
Ceramide analysis of tissue samples was performed by liquid chromatography-triple quadrupole mass spectrometry according to a modified version of the protocol reported by Kasumov et al. (2010). Briefly, frozen tissue samples were pulverized under liquid nitrogen, then 20 mg portions were extracted using 1.6 mL of a 2:1:0.8 mixture of chloroform:methanol:water containing internal standards (50 ng each of C17 and C25 ceramide and C12 glucosylceramide per sample). The organic layer of the extract was dried under nitrogen gas and reconstituted in 100 uL of 60:40 acetonitrile: isopropanol (Bligh & Dyer 1959). The re-constituted extract was analyzed by electrospray ionization LC-MS/MS on an Agilent (Santa Clara, CA) 6410 triple quadrupole instrument operating in positive ion multiple reaction monitoring mode. The LC column used was a Waters (Milford, MA) Xbridge C18 2.5 µ, 50 mm × 2.1 mm i.d. Mobile phase A was 5mM ammonium acetate, adjusted to pH 9.9 with ammonium hydroxide; mobile phase B was 60:40 acetonitrile:isopropanol. The gradient consisted of a linear ramp from 50 to 100%B over 5 minutes, a 20 minute hold at 100%B, and re-equilibration at 50%B for 10 minutes. Injection volume was 25 µL. Ceramides and glucosylceramides were identified by retention time and by MS/MS fragmentation parameters, and were quantitated by peak area relative to the closest-matching internal standard using Agilent MassHunter Quantitative Analysis software.
Transcriptomic Analysis
Total RNA was extracted from adipose tissue using the RNEasy kit (Qiagen) and its quality was verified using the Agilent 2100 Bioanalyzer (Agilent Technologies). At the University of Michigan DNA Sequencing Core, cDNA libraries from polyA mRNA were prepared using TruSeq cDNA synthesis kit and sequenced using a HiSeq 2000 (Illumina). Samples were run on 2 lanes of a HiSeq 2000 (Illumina) generating 8,612,682 to 16,469,501 single-ended 50 bp reads per sample. These were aligned to the human genome (Enembl GRCh37.74, Genbank Assembly ID GCA_000001405.14) using TopHat version 2.0.10 (Kim et al. 2013), Bowtie 2 version 2.1.0 (Langmead & Salzberg 2012) and Samtools version 0.1.18. Reads were mapped to known genes using HTseq (Anders et al. 2014). Gene expression was analyzed using DESeq2 version 1.2.10 (Love et al. 2014). These subjects corresponded to the patients described in Table 2, with the exception of subjects 29 and 31 (both Cushing's disease patients), which had clinical data but no RNAseq data.
Table 2. Clinical characteristics of Cushing’s disease and control patients.
Data represents mean +/− standard error.
| Cushing's disease (n=5) |
Controls (n=11) |
p-value | |
|---|---|---|---|
| Height (cm) | 166 ± 4.3 | 169 ± 2.4 | 0.47 |
| Weight (kg) | 91 ± 9.1 | 89 ± 6.7 | 0.89 |
| BMI | 33 ± 3.8 | 30 ± 1.8 | 0.52 |
| Abdominal circumference (cm) | 112.4 ± 6.4 | 100.65 ± 4.4 | 0.16 |
| Tumor size (cm) | 0.95 ± 0.3 | 1.96 ± 0.14 | 0.01 |
| Age (years) | 39.8 ± 4.5 | 63.4 ± 2.7 | 0.0003 |
Statistics
Descriptive statistics including means and standard errors were determined for clinical measurements. All statistical tests were performed using the R package (version 3.0.2,(R Core Team 2013)). Normality assumption was checked via Shapiro-Wilk test. Wilcoxon rank sum tests were used when data were not normally distributed. Welch’s t-test was performed if the equal variance assumption was rejected by Levene's test (car package version 2.0–19), otherwise a Student’s t-test was used. Longitudinal measurements such as body weight, food intake, body composition and insulin tolerance tests were analyzed via mixed linear models and a χ2 test between models with and without dexamethasone treatment as a covariate. This used the lme4 package, version 1.1–7 (Bates et al. 2014). Statistical significance in this study was defined as a p/q-value of less than 0.05. To correct for multiple hypotheses, p-values were adjusted by the method of Benjamini and Hochberg (Benjamini & Hochberg 1995). The DESeq2 algorithm excludes genes with very high variance to improve statistical power (Love et al. 2014). The analysis we focused on in this manuscript was without adjustment for BMI or age and is presented in Supplementary Table 1, with GSEA analyses in Supplementary Tables 2–3. A model controlled for BMI as a linear covariate or stratified into obese or non-obese subjects is presented in Supplementary Tables 4 and 5. A model controlled for both BMI and age was also constructed and is presented in Supplementary Table 6. To ensure that we did not miss any genes that had a high fold change, but that DESeq2 did not perform statistical tests for, we manually inspected genes that had a expression at >50 reads, fold change >2.5 but no p-value calculated. These genes included FADS1, FADS2, ELOVL6, SPP1, BMP3 and AACS (see Supplementary Table 1). All data are presented as mean +/− standard error of the mean.
We used Gene Set Enrichment Analysis (GSEA v2.0.13 (Subramanian et al. 2005; Clark & Ma’ayan 2011)) to determine whether our rank-ordered gene list for the comparison of Cushing's disease versus control patients is enriched in genes from gene ontology, KEGG, transcription factor or microRNA target gene sets (MSigDB version 4.0). The gene list was ranked based on t-statistics and the statistical significance of the enrichment score was determined by performing 1000 phenotype permutation. Other settings for GSEA were left to the software defaults. All GSEA results are in Supplementary Tables 2–3 and summarized in Table 3. All code and raw data from this study are available through the Gene Expression Omnibus (GSE66446) and at http://bridgeslab.github.io/CushingAcromegalyStudy (Hochberg et al. 2015)
Table 3. Summarized gene set enrichment analysis of pathways.
Selected pathway enriched in subcutaneous adipose tissue from Cushing’s disease patients via GSEA analysis. NES is the net enrichment score, asterisk indicates q<0.25. For a complete list see Supplementary Tables 2–3.
| Pathway | Dataset | NES |
|---|---|---|
| M_PHASE_OF_MITOTIC_CELL_CYCLE | Gene Ontology | 2.60* |
| KEGG_CITRATE_CYCLE_TCA_CYCLE | KEGG | 2.41* |
| KEGG_BIOSYNTHESIS_OF_UNSATURATED_FATTY_ACIDS | KEGG | 2.41* |
| REACTOME_TRIGLYCERIDE_BIOSYNTHESIS | Reactome | 2.24* |
| PYRUVATE_METABOLISM | Gene Ontology | 2.24* |
| KEGG_VALINE_LEUCONE_AND_ISOLEUCINE_DEGRADATION | KEGG | 2.16* |
| STEROID_BIOSYNTHETIC_PROCESS | Gene Ontology | 2.11* |
| KEGG_STARCH_AND_SUCROSE_METABOLISM | KEGG | 2.08* |
| PROTEASOME_COMPLEX | Gene Ontology | 1.78* |
| KEGG_ALLOGRAFT_REJECTION | KEGG | −1.87* |
| KEGG_BASAL_CELL_CARCINOMA | KEGG | −1.86* |
| KEGG_RIBOSOME | KEGG | −2.33* |
Results
Patient Characteristics
Clinical and metabolic measurements were obtained for five Cushing's disease patients and 11 control subjects, who were admitted with non-secreting adenomas. Patient characteristics are shown in Table 2. Our Cushing’s disease patients were in general younger and had smaller tumors than the patients with non-secreting adenomas. In the Cushing’s disease cohort there was a non-significant elevation in body weight (p=0.47), body mass index (BMI) (p=0.27) and abdominal circumference (p=0.07, Figure 1A), consistent with Cushing’s disease patients having elevated fat mass and truncal obesity (Lamberts & Birkenhäger 1976).
Figure 1. Metabolic characteristics of Cushing’s disease patients in our study.
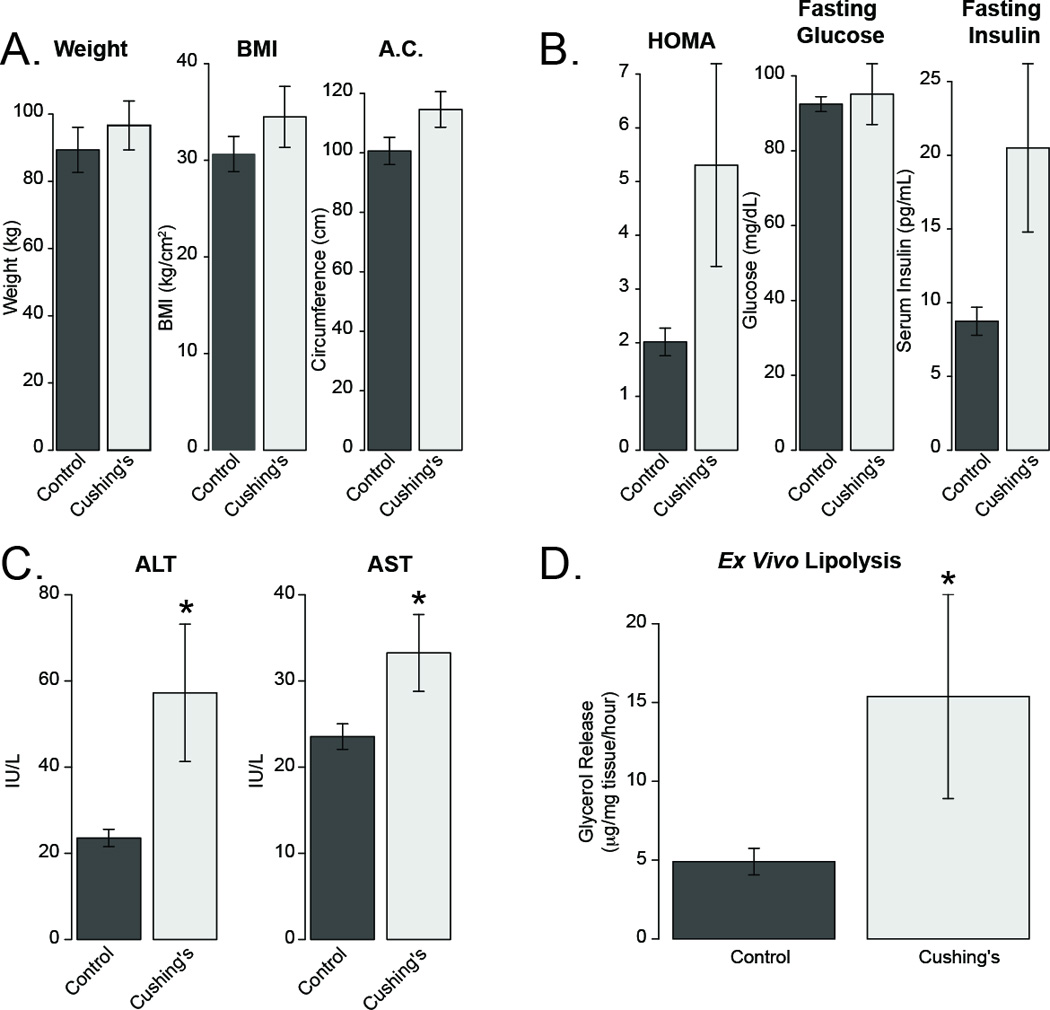
A) Morphometric data from control (non-secreting adeoma) and Cushing’s disease subjects. A.C indicates abdominal circumference. B) HOMA-IR score, fasting insulin and fasting blood glucose from subjects. C) Liver enzymes from subjects D) Glycerol release from isolated subcutaneous adipose tissue. Asterisks indicates p<0.05.
We detected a non-significant elevation in HOMA-IR score (2.6 Fold, p=0.67 by Wilcoxon test, Figure 1B), driven largely by increases in fasting insulin levels (p=0.30). Three out of the five Cushing's disease patients had diabetes, while only 1 of the 11 controls had diabetes (p=0.03 via χ2 test). These data are consistent with elevated glucose intolerance in patients with Cushing’s syndrome. We observed significant elevations in both ALT and AST in serum from Cushing’s disease patients. To evaluate lipolysis in explants from these patients we measured glycerol release from isolated subcutaneous adipose tissue and found a 3.1 fold elevation in glycerol release from these tissues (p=0.049 via Student’s t-test). These data support previous studies which implicate elevated lipolysis (Kršek et al. 2006) and higher rates of non-alcoholic fatty liver disease in Cushing’s disease patients (Rockall et al. 2003).
Dexamethasone Treatment of Mice as a Model of Cushing’s Syndrome
To validate the gene expression changes observed in human subjects, we treated C67BL/6J mice with dexamethasone, a synthetic glucocorticoid in their drinking water to mimic the systemic effects of cortisol overproduction. These mice had an initial catabolic phase in which their body weight was rapidly reduced (Figure 2A), an effect that was primarily in due to a reduction in lean body mass (Figure 2B). This is consistent with previously reported effects of glucocorticoids on muscle atrophy (Pleasure et al. 1970). After approximately 5 weeks, we observed an elevation in both total fat mass, and percent adiposity in the dexamethasone treated mice (Figure 2C–D). We did not detect any differences in food intake between the groups throughout the study (Figure 2E). To evaluate insulin sensitivity, we performed insulin tolerance tests on these mice after 12 weeks of dexamethasone treatment, and found that while they had reduced fasting glucose at this stage, they were resistant to insulin-induced reductions in blood glucose (Figure 2E). Upon sacrifice after 12 weeks of dexamethasone treatment, adipose tissue was dissected and weighed. As shown in Figure 2F, we observed elevated subcutaneous fat mass in dexamethasone treated animals.
Figure 2. Dexamethasone treatment results in decreased lean mass and increased fat mass in mice.
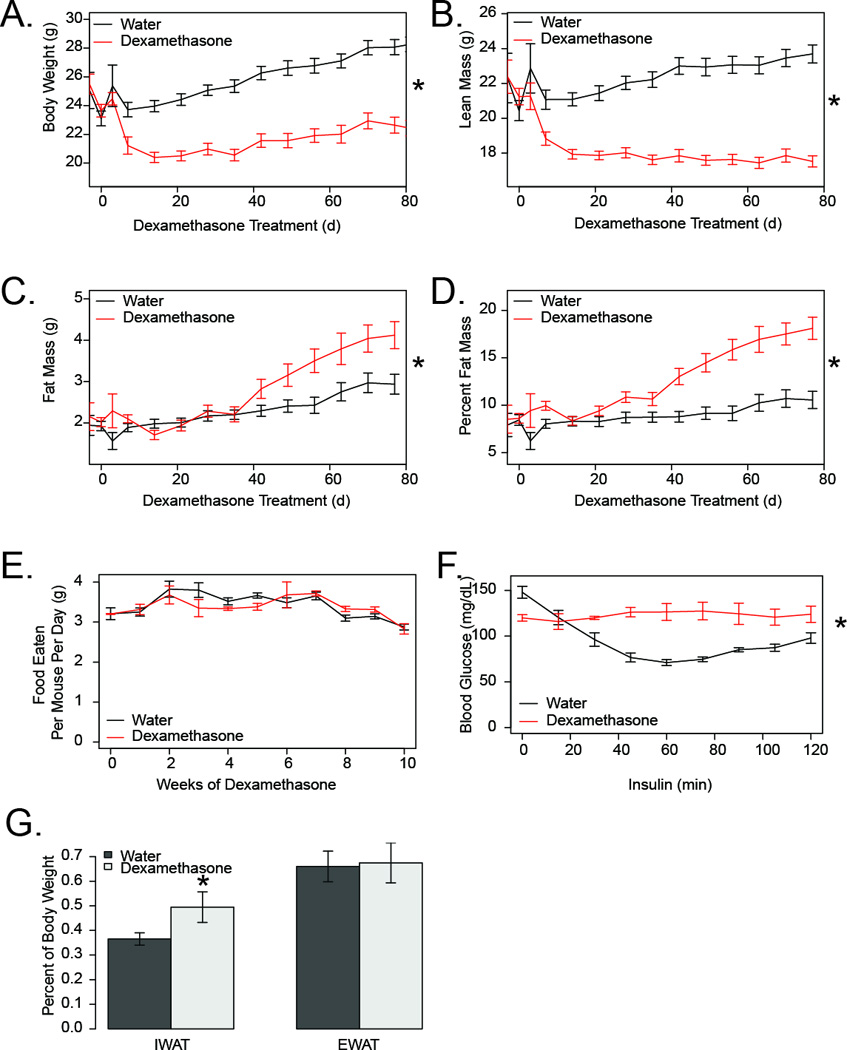
Weekly body weight (A), lean mass (B), fat mass (C) and percent fat (D) from control (black) and dexamethasone (red) treated mice. E) Average food consumption per mouse per day. F) Insulin tolerance test. Following a 6 hour fast, insulin (1 mU/g) was administered via IP injection and blood glucose was measured at baseline and the indicated time post injection. G) Inguinal (IWAT) and epididymal (EWAT) fat pad weights, for right fat pads only. Asterisks indicate p<0.05.
Transcriptomic Analysis of Human Adipose Tissue From Cushing’s patients
To determine which genes and pathways are altered in adipose tissue in the human Cushing's disease subjects, we analyzed the transcriptome from subcutaneous adipose tissue mRNA from the 5 Cushing's disease patients and 11 controls. We identified 473 genes that had significantly different expression in Cushing's disease patients, of these 192 genes were expressed at a lower level and 281 at a higher level in the adipose tissue from the disease patients. These transcripts form a signature identifying transcriptional differences in adipose tissue in response to long-term exposure to glucocorticoids (Figure 3A).
Figure 3. Differentially expressed transcripts in subcutaneous adipose tissue from Cushing’s disease subjects.
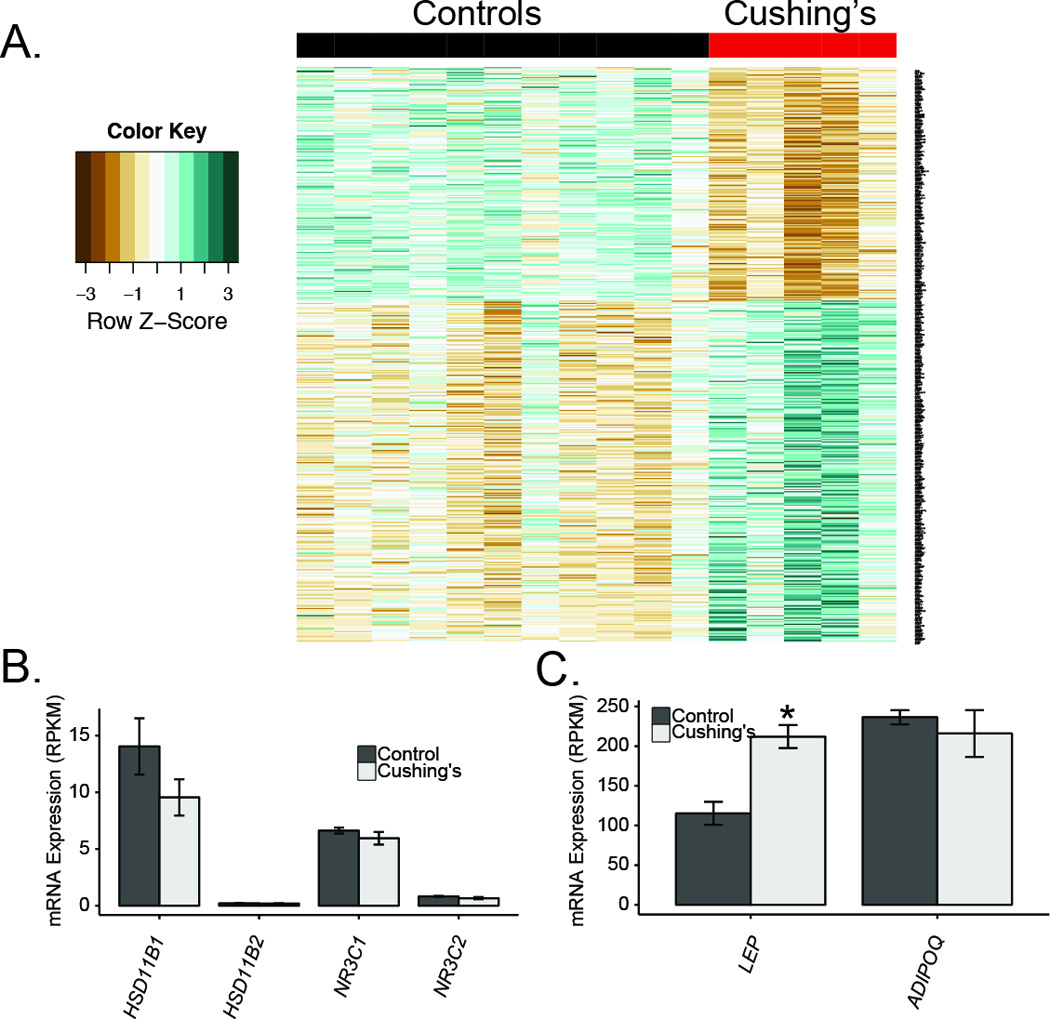
A) Heatmap of genes with significant differential expression. The bar on the top indicates control subjects (non-secreting adenoma; black) and Cushing’s subjects (red). B) Genes involved in cortisol signaling. C) Leptin and Adiponectin mRNA levels. Asterisks indicate q<0.05.
To identify conserved pathways underlying these changes, gene set enrichment analysis was performed on these data. As summarized in Table 3, we detected enrichment of genes in several categories involved in metabolism, including higher expression of gene sets involved in lipid biosynthesis, glucose metabolism, activation of amino acid degradation, protein degradation, and reductions in protein synthesis. We also observed reduced expression of transcripts involved in immune function. These will be discussed in subsequent sections.
We next evaluated the levels of the glucocorticoid receptor (NR3C1) and the mineralocorticoid receptor (NR3C2) and observed no significant downregulation of these receptors at the mRNA level in Cushing’s patients (Figure 3B). Another potential mechanism for negative feedback of glucocorticoid signaling is through the enzymatic activities of 11β-HSD1/2 which control the local concentrations of cortisol in adipose tissues. We observed a non-significant reduction in HSD11B1 mRNA levels (24% reduced, padj=0.49), potentially desensitizing adipose tissue to cortisol by reducing the conversion of cortisone to cortisol. Induction of leptin by glucocorticoids has been previously reported in human adipocytes (Halleux et al. 1998) and in human adipose tissue in vivo (Papaspyrou-Rao et al. 1997). We observed 1.8 fold higher level of Leptin (LEP) expression, and non-significantly higher resistin (RETN) expression but no significant changes in adiponectin mRNA levels (ADIPOQ, padj=0.94; Figure 3C).
Lipogenesis Genes are Upregulated in Response to Elevated Glucocorticoids
Increased subcutaneous fat mass is a hallmark of Cushing’s syndrome, and could potentially be mediated through activation of adipogenesis or lipogenesis. Our transcriptomic data support the hypothesis that lipogenesis is activated in these tissues via transcriptional activation of fatty acid synthesis and triglyceride synthesis. All the major genes involved in the synthesis of fatty acids were expressed at higher levels including ACACA, FASN, ACSL1/3/4, and ELOVL1/5/6 (Figure 4A). Desaturation of fatty acids is an essential aspect of de novo fatty acid synthesis, and we also observed elevations in all fatty acid desaturases SCD, FADS1, FADS2 and HSD17B12 (Figure 4B). The triglyceride synthesis genes GPAM, DGAT1, DGAT2, AGPAT2/3, and GPD1 were also all upregulated in subcutaneous adipose tissue from Cushing’s disease patients (Figure 4C).
Figure 4. Elevated glucocorticoids result in elevated fatty acid and triglyceride synthesis genes.

A) Fatty acid synthesis genes in Cushing’s disease and control patients. B) Fatty acid desaturases in Cushing’s disease patients. C) Triglyceride synthesis genes. D) Lipolysis genes. E) Steroid biogenesis genes. F) Evaluation of lipogenic genes in mouse subcutaneous adipose tissue. Asterisks indicate q<0.05.
In spite of increased lipid deposition and elevations of lipogenesis genes in Cushing’s disease patients' adipose tissue, there have been several studies linking elevated glucocorticoids to increased lipolysis. In our patients, this was observed in ex vivo explants of subcutaneous adipose tissue (Figure 1D). Among genes that may liberate fatty acids from triglycerides, Lipoprotein Lipase (LPL) was induced 1.45 fold (padj=0.055) in the Cushing’s disease subjects, but neither Hormone Sensitive Lipase (LIPE) nor Adipose Triglyceride Lipase (PNPLA2) were significantly changed at the transcriptional level (Figure 4D). It is possible that insulin resistance due to glucocorticoids caused decreased repression of lipolysis leading to its upregulation. However, our data supports an insulin-independent activation as well, since in our explants insulin was not present during the lipolysis assay. We detected an elevation of Perilipin 4 (PLIN4) which is one of the proteins that coat intracellular lipid storage droplets (induced 1.45 fold, padj=0.056, Supplementary Table 1).
Several genes that regulate steroid biogenesis were elevated in adipose tissue from Cushing’s disease patients as described in Figure 4E. These include several cytochrome P450 family members, steroid reductases (SRD5A1, SRD5A3), Aldo-keto reductase family 1 member C1 (AKR1C1), 7-dehydrocholesterol reductase (DHCR7) and HMG-CoA synthase (HMGCS1).
To examine whether lipogenesis genes are activated in the dexamethasone treated mice, we tested several of these genes in subcutaneous adipose tissue from dexamethasone treated mice, and observed elevations in Fasn, Gpam, Gpd1, Acss2, Acs1, Dgat, Agpat2, Dhcr7/24 and Acaca1. (Figure 4F). In contrast to the human samples, did not observe an elevation in the mouse isoform of SCD, but saw instead a reduction in Scd1 mRNA.
Genes Controlling Glucose Oxidation Are Elevated in Cushing's Disease Patients
Several glucose metabolism genes, and specifically glycolysis and TCA cycle genes were expressed at higher levels in Cushing's disease patients (Figure 5). Strongly induced genes included HK3, FBP1, ALDOC, ENO1, IDH1, ME1 and DLAT. Upregulations in Idh1 and Me1 were also noted in mouse adipose tissue, along with other transcripts involved in glucose oxidation such as Aco1, Ldhb and Mdh1 (Figure 5C).
Figure 5. Glycolysis and glucose oxidation genes are upregulated with elevated glucocorticoids.
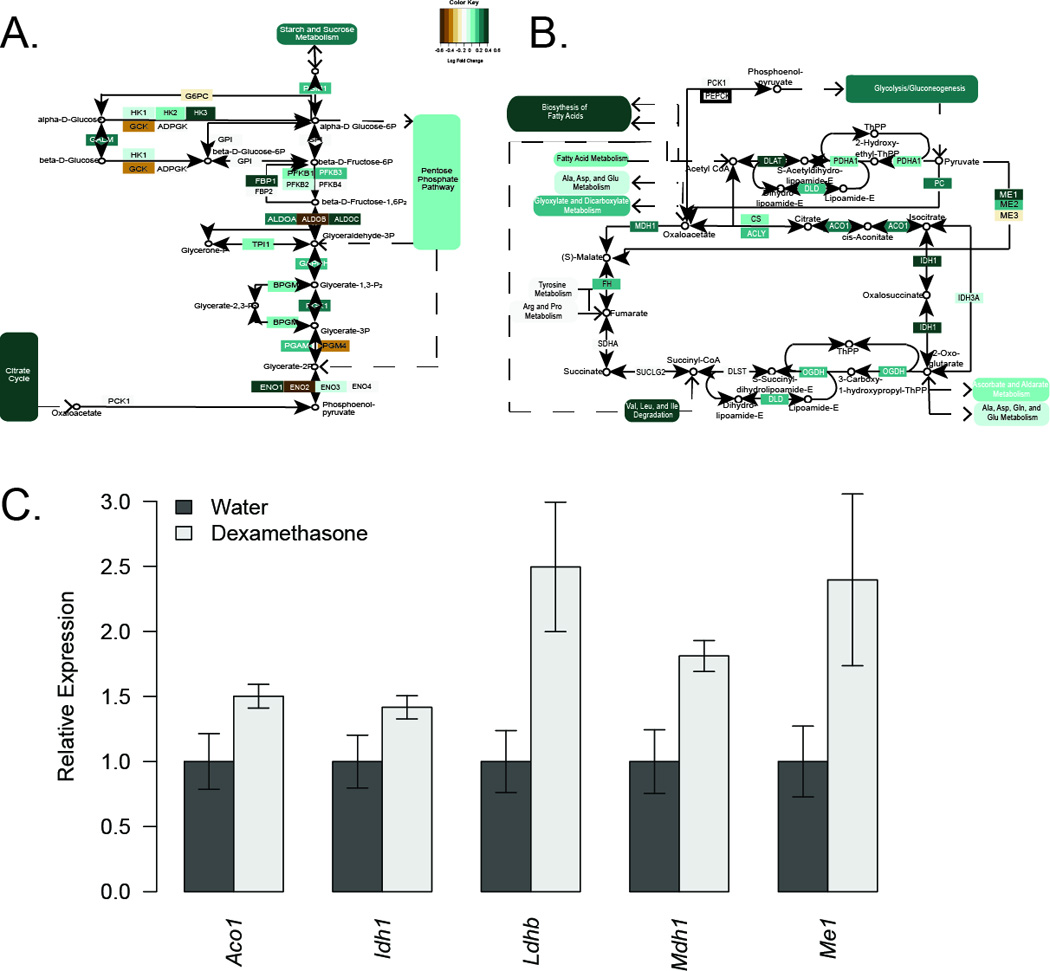
Schematic of A) glycolysis and B) the TCA cycle, colored by gene expression changes in subcutaneous adipose tissue from Cushing’s disease subjects. C) qPCR analysis of selected glucose oxidation genes from mouse subcutaneous adipose tissue after 12 weeks of dexamethasone treatment. Asterisks indicate q<0.05.
The major glycogen synthesis transcripts were also induced, including GYS2, UGP2 and GBE1. This agrees with biochemical studies which implicate glucocorticoid treatment in elevated hepatic and adipose tissue glycogenesis (Engel & Scott 1951; Segal & Gonzalez Lopez 1963; Baqué et al. 1996). The relevance of this effect in adipose tissue has not yet been explored.
Genes That Regulate Protein Catabolism are Upregulated in Adipose Tissue from Glucocorticoid Exposed Subjects
We found two major pathways of protein homeostasis are altered in response to glucocorticoids. In concert with reductions in lean body (including muscle) mass (Figure 2B), we observed substantial muscle weakness in mice treated with dexamethasone (Figure 6A). In a separate cohort of mice, after one week of dexamethasone treatment skeletal muscle, mRNA levels of the E3 ligases (Atrogin-1 and MuRF1) were induced as was the proteasomal gene Psmd8 (Figure 6B). These effects did not reach statistical significance due to variability in dexamethasone responsiveness. Similar inductions of the proteasomal genes were observed in subcutaneous adipose tissue from the cohort of mice treated with dexamethasone for 12 weeks (Figure 6C).
Figure 6. Increased glucocorticoids are associated with increased protein degradation and decreased strength.

A) Mouse grip strength (N) assessed at baseline, 4, 8 and 12 weeks of dexamethasone treatment. Muscle atrogene (B) and proteasomal transcript expression changes in gastrocnemius muscles from mice following 1 week of dexamethasone treatment. C) Proteosomal mRNA levels from subcutaneous adipose tissue of mice treated with dexamethasone for 12 weeks. Proteasomal (D) and protein catabolism (E) transcript expression changes in subcutaneous adipose tissue from Cushing’s disease and control subjects. F) Heatmap of differentially expressed ribosomal transcripts in Cushing’s disease and control subjects.
In adipose tissue from Cushing’s disease patients, we observed inductions of both the proteasomal pathways (via KEGG, net enrichment score 1.76, padj=0.01; Figure 6D), and genes involved in amino acid catabolism (Figure 6E) and a general downregulation of ribosomal genes (Figure 6F). Among the amino acid catabolism genes, AOX1 (96% increase, padj=0.03), OXCT1 (40% increase, padj=0.04) and BCAT1 (80% increase, p=0.048) were all significantly upregulated. Together these data support the hypothesis that protein catabolism and a reduction in protein synthesis also occur in adipose tissue in response to glucocorticoid exposure.
Genes Involved in Proximal Insulin Signaling are Unchanged in Adipose Tissue from Cushing’s Disease Patients
As described in Figures 1B and 2F, we observed insulin resistance in concert with elevated glucocorticoid levels in both mice and humans. Several mechanisms have been proposed linking glucocorticoids to insulin sensitivity including elevated lipolysis. As shown in Figure 7A, There was a slightly higher expression of insulin pathway transcripts including FOXO1, insulin receptor (INSR), the insulin receptor substrates IRS1 or IRS2 and p85 regulatory subunit of phosphoinositide-3-kinase (PIK3R1), consistent with previous studies (Gathercole et al. 2007; Tomlinson et al. 2010; Hazlehurst et al. 2013), though in our hands none of these genes reached statistical significance. The insulin pathway was generally expressed at significantly higher levels in the Cushing's disease patients compared to controls (KEGG pathway, net enrichment score 1.84, padj=0.006). These data do not support transcriptional downregulation of proximal insulin signaling genes as mediating insulin resistance in subcutaneous adipose tissue.
Figure 7. Expression of insulin signaling transcripts, ceramides and inflammatory transcripts in control vs. Cushing’s disease subjects.
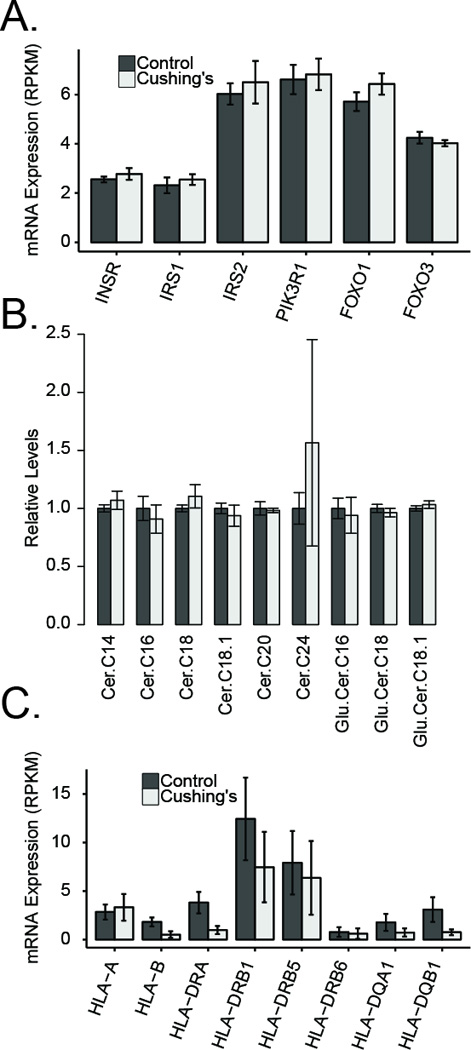
A) Insulin signaling transcript expression levels. B) Ceramide levels. C) MHC complex transcript expression levels.
Changes in cell ceramide and glucosylceramide have been suggested to be important in vitro and in obesity- and glucocorticoid-induced insulin resistance in skeletal muscle (Adams et al. 2004; Aerts et al. 2007; Holland et al. 2007). To test biochemically whether ceramides may play a role in the Cushing's disease associated insulin resistance in adipose tissue, we took a lipidomics approach to analyze ceramide species from the adipose tissue explants of the same patients. We observed no statistically significant changes in any ceramide species (Figure 7B, q>0.25).
Inflammation
Several pathways involved in immune function were downregulated in adipose tissue from Cushing’s disease patients. This is consistent with the effects of cortisol in suppressing immune function generally. Adipose tissue leukocyte infiltration both relies on an intact immune system and also responds to changes in adiposity (Lumeng & Saltiel 2011). Among immune genes, we detected reductions in several genes that form the class II major histocompatibility complex, notably HLA-DPB2, HLA-DRA, HLA-DRB9 and HLADQA1 (Figure 7C). These genes normally present antigens for T-cell recruitment. Consistent with this, we observed reductions in the mRNA of IL32, a hormone secreted by Natural Killer and T lymphocytes (Dinarello & Kim 2006). We also observed a downregulation in transcripts that are interferon gamma dependent. Together, these data support the hypothesis that the decreased T-cell activation observed with cortisol signaling also impacts adipose tissue.
Modifying Effect of Obesity on Glucocorticoid Responsiveness
In our small cohort of Cushing’s disease subjects, we examined whether some of the dramatic transcriptional changes were modified by the obesity status of the patients (based on a BMI cutoff of 30; Supplementary Tables 4 and 5). We were surprised to note that many genes that had strongly elevated transcripts in non-obese Cushing’s disease patients had blunted effects in obese Cushing’s disease patients. Some examples of this include FASN, PSMD8 and IDH8 (Figure 8A–C). Among genes that were more strongly induced in obese patients, most of these are involved in lysosomal function, including the cathepsins (CTSB, CTSD, CTSZ), LAPTM5 and LIPA (Figure 8D). Although the small number of obese and non-obese Cushing’s patients in our study makes these provocative observations, preliminary, it is suggestive of both a general reduction of glucocorticoid sensitivity in obese subjects but also potentially an underappreciated role of lysosomes in obese patients with elevated cortisol levels.
Figure 8. Transcript expression changes in Cushing’s disease are less robust after adjusting for obesity.
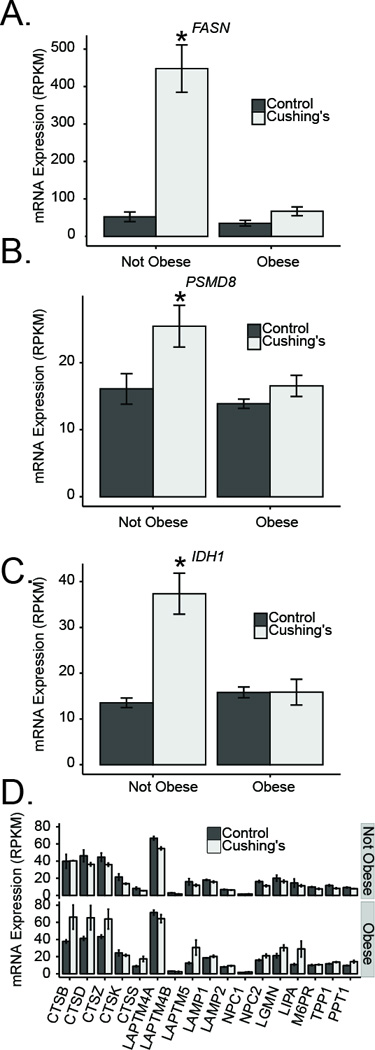
FASN (A), PSMD8 (B), IDH1 (C), and lysosomal (D) transcripts in non-obese and obese Cushing’s subjects.
Discussion
In this study we have described a transcriptional signature in adipose tissue from subjects with Cushing's disease and verified several of these changes using a mouse model of glucocorticoid treatment. We have identified several pathways that are significantly changed in response to chronic glucocorticoid exposure. Broadly, these changes reflect a shift towards more rapid metabolism of glucose through glycolysis and the TCA cycle, and shifting of glucose and protein metabolites towards lipogenic pathways in adipose tissue. This is indicated by significant increases in glycolytic (ALDOC, ENO1, IDH1, ME1, GALM and GAPDH), proteolytic (PSMD1/14) and lipogenic (ACACA, FASN, ACSL1, ELOVL5, GPAM and DGAT2) transcripts in human adipose tissue, with similar transcript expression changes seen in mouse adipose and muscle tissue when treated with dexamethasone. A limitation of our human data is the difference in age between non-secreting adenoma and Cushing’s disease subjects. Cushing’s disease is diagnosed and treated much more rapidly, which leads to these differences. We therefore confirmed many of our human findings in a mouse model of excessive glucocorticoid treatment, wherein the mice were treated under more controlled conditions. Studies using a Hsd11b1 knockout mouse showed similar findings to our data including increased fat mass and decreased lean mass and strength, along with reduced insulin sensitivity (Morgan et al. 2014). Transcriptionally both of our studies report increases in Dgat mRNA, though we observed no effects of Cushing’s disease on lipolytic genes (Figure 4D) as that study did. In our study we did observe induction of fatty acid synthesis genes in both humans and mice (Figure 4A/F) which was not observed in the Morgan et al. study. Three differences could potentially explain these discrepancies. One is that in our case, dexamethasone is already active and cannot be further activated by 11-HSD1, whereas in their study corticosterone can be both inactivated by 11-HSD2 and reactivated by 11-HSD1. Another key difference is the duration of treatment, which for our study was three months and for the Morgan et al. study was just over one month. Finally they determined mRNA levels from gonadal adipose tissue, not subcutaneous adipose tissue, as we did in our work.
Cushing's disease patients have a significant change in fat distribution (Mayo-Smith et al. 1989), and higher lipogenesis, as measured by conversion of glucose to neutral lipid ex vivo in subcutaneous adipose tissue from Cushing's disease patients compared to obese controls (Galton & Wilson 1972). Higher triglyceride synthesis has also been found in animal models of Cushing's disease, including CRH overproducing mice, which also have elevated glucocorticoid levels (Harris et al. 2013) and in dexamethasone treated mice (Roohk et al. 2013). These findings are consistent with our observed elevations of lipogenesic mRNA transcripts in human and mouse subcutaneous adipose tissue. Key transcripts in this category found to be significantly upregulated include Acetyl-CoA carboxylase alpha (ACACA), responsible for the first step of lipogenesis (the irreversible conversion of acetyl-CoA to malonyl-CoA) and Glycerol-3-phospahte acyltransferase (GPAM) is responsible for the first step in the synthesis of glycerolipids. In addition to a shift towards lipid storage, we also observed elevated expression of glycogen synthesis mRNA transcripts in the Cushing's disease patients. Most notably of these are significantly elevated mRNA transcripts Glycogen synthase 2 (GYS2) and UDP-glucose pyrophosphorylase 2 (UGP2), both of which are required for glycogen synthesis.
Muscle wasting is a well recognized adverse event of excess glucocorticoids caused by both increased muscle proteolysis and decreased protein synthesis (Deng et al. 2004; Menconi et al. 2007). Exposure of rats to glucocorticoids activates the muscle ubiquitin-proteasome system (Wing & Goldberg 1993; Price et al. 1994), increasing muscle expression of proteases (cathepsins B and D, calpain) and components of the ubiquitin-proteasome pathway (Dardevet et al. 1995) along with inhibition of muscle protein synthesis (Long et al. 2001). A study in healthy humans also found that prednisone (a synthetic corticosteroid) increases leucine oxidation supporting our observation of elevated amino acid catabolic genes (Beaufrere et al. 1989). We found higher expression of both proteasomal and the amino acid degradation pathways in adipose tissue, suggesting that a similar induction occurs in adipose tissue from our Cushing's disease subjects. We also observe elevations in lysosomal genes, though these changes appear to be restricted to obese Cushing’s disease patients. The metabolic relevance of activated proteolysis in adipose tissue has not been widely explored and warrants further study.
There are several limitations to our evaluation of insulin sensitivity in this study. One aspect is that two of the three patients with Cushing’s syndrome and diabetes were treated with antidiabetic medications. Secondly, it is possible that insulin resistance in these subjects/mice is mainly due to muscle or liver insulin resistance and that adipose tissue may respond to insulin in a relatively normal fashion. Glucocorticoid-induced insulin resistance is thought to be mostly secondary to the increase in free fatty acids caused by the induction of lipolysis (Geer et al. 2014). Results from a recent study imply that glucocorticoids do not induce insulin resistance in subcutaneous adipose tissue in vivo in healthy subjects (Hazlehurst et al. 2013), suggesting that peripheral insulin resistance may not occur in adipocytes and that whole-body insulin resistance may primarily occur in muscle and liver tissues. This is consistent with our observations of a lack of changes in proximal insulin signaling transcripts (Figure 7A) or ceramides in our subcutaneous adipose tissue lysates (Figure 7B).
Another limitation in our study is the small sample size, especially the number of biological replicates in Cushing’s group (n=5). Adding a covariate such as BMI or age in the model further reduces the sample size to 2 or 3 replicates. Although this sample size is small, it is reasonable for a rare disease such as Cushing’s. Realizing our limitation, we chose DESeq2 as the statistical method for our RNAseq data. DESeq2 overcomes the small sample size problem by pooling information across genes. Maximum likelihood estimation is applied to estimate the dispersion or variance of a gene across all replicates in a group. An empirical Bayes approach is then used to get maximum a posterior as the final dispersion estimate. This method utilizes the available data to the maximum extent; therefore, helps avoiding potential false positives (Love et al. 2014).
These data provide a variety of novel transcriptional changes that may be causative of the co-morbidities associated with Cushing's disease. Further studies in animals and cells using knockout or overexpression of specific transcripts may verify which of the changes is crucial in the metabolic effects of glucocorticoid effects in adipose tissue.
Supplementary Material
{kind=link}
Acknowledgements
We thank Charlotte Gunden, Elizabeth Walkowiak and Eric Vasbinder for their valuable help in the study. The authors would like to thank Ian Brooks and the UTHSC-ORNL Center for Biomedical Informatics for provisioning the RStudio server used in this analysis. We would also like to thank the University of Tennessee Health Science Center Neuroscience Institute for use of the grip strength monitor. We would also like to thank the Molecular Resource Center at the University of Tennessee Health Science Center for qPCR facilities.
Funding
This work was supported by Motor City Golf Classic (MCGC) Grant # G010640 and Le Bonheur Grant #650700. This work utilized Metabolomics Core Services supported by grant U24 DK097153 of the NIH Common Fund to the University of Michigan.
Footnotes
Declaration of Interest
The authors have no conflict of interest.
Author Contributions
IHo conceived of the study, and DB, ARS and IHo provided funding. WFC and ALB recruited the patients and obtained clinical data. WFC supplied the biopsies and serum samples. IHo assayed the tissues for lipolysis and performed the serum measurements. QT, DB, IHa and IHo analyzed the RNAseq data. IHa generated the mouse data with assistance from EJS. This was analyzed by IHa, DB and QT. IHo and DB wrote the manuscript
References
- Adams JM, Pratipanawatr T, Berria R, Wang E, DeFronzo RA, Sullards MC, Mandarino LJ. Ceramide content is increased in skeletal muscle from obese insulin-resistant humans. Diabetes. 2004;53:25–31. doi: 10.2337/diabetes.53.1.25. [DOI] [PubMed] [Google Scholar]
- Aerts JM, Ottenhoff R, Powlson AS, Grefhorst A, van Eijk M, Dubbelhuis PF, Aten J, Kuipers F, Serlie MJ, Wennekes T, et al. Pharmacological inhibition of glucosylceramide synthase enhances insulin sensitivity. Diabetes. 2007;56:1341–1349. doi: 10.2337/db06-1619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anders S, Pyl PT, Huber W. HTSeq - A Python framework to work with high-throughput sequencing data. Bioinformatics. 2014:1–4. doi: 10.1093/bioinformatics/btu638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baqué S, Roca a, Guinovart JJ, Gómez-Foix aM. Direct activating effects of dexamethasone on glycogen metabolizing enzymes in primary cultured rat hepatocytes. European Journal of Biochemistry. 1996;236:772–777. doi: 10.1111/j.1432-1033.1996.t01-1-00772.x. [DOI] [PubMed] [Google Scholar]
- Bates D, Mächler M, Bolker B, Walker S. Fitting Linear Mixed-Effects Models using lme4. ArXiv. 2014;1406.5823:1–51. [Google Scholar]
- Beaufrere B, Horber FF, Schwenk WF, Marsh HM, Matthews D, Gerich JE, Haymond MW. Glucocorticosteroids Increase Leucine Oxidation and Impair Leucine Balance in Humans. The American Journal of Physiology. 1989 doi: 10.1152/ajpendo.1989.257.5.E712. [DOI] [PubMed] [Google Scholar]
- Benjamini Y, Hochberg Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. Journal of the Royal Statistical Society. Series B. 1995;57:289–300. [Google Scholar]
- Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology. 1959;37:911–917. doi: 10.1139/o59-099. [DOI] [PubMed] [Google Scholar]
- Campbell JE, Peckett AJ, D’souza AM, Hawke TJ, Riddell MC. Adipogenic and lipolytic effects of chronic glucocorticoid exposure. American Journal of Physiology. Cell Physiology. 2011;300:C198–C209. doi: 10.1152/ajpcell.00045.2010. [DOI] [PubMed] [Google Scholar]
- Clark NR, Ma’ayan A. Introduction to statistical methods for analyzing large data sets: gene-set enrichment analysis. Science Signaling. 2011;4 doi: 10.1126/scisignal.2001966. tr4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cushing H. The basophil adenomas of the pituitary body and their clinical manifestations. Bulletin of the Johns Hopkins Hospital. 1932;50:157–158. [Google Scholar]
- Dardevet D, Sornet C, Taillandier D, Savary I, Attaix D, Grizard J. Sensitivity and protein turnover response to glucocorticoids are different in skeletal muscle from adult and old rats. Lack of regulation of the ubiquitin-proteasome proteolytic pathway in aging. Journal of Clinical Investigation. 1995;96:2113–2119. doi: 10.1172/JCI118264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng X, Elam MB, Wilcox HG, Cagen LM, Park Ea, Raghow R, Patel D, Kumar P, Sheybani A, Russell JC. Dietary olive oil and menhaden oil mitigate induction of lipogenesis in hyperinsulinemic corpulent JCR:LA-cp rats: microarray analysis of lipid-related gene expression. Endocrinology. 2004;145:5847–5861. doi: 10.1210/en.2004-0371. [DOI] [PubMed] [Google Scholar]
- Dinarello Ca, Kim S-H. IL-32, a novel cytokine with a possible role in disease. Annals of the Rheumatic Diseases. 2006;65(Suppl 3):iii61–iii64. doi: 10.1136/ard.2006.058511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Divertie GD, Jensen MD, Miles JM. Stimulation of lipolysis in humans by physiological hypercortisolemia. Diabetes. 1991;40:1228–1232. doi: 10.2337/diab.40.10.1228. [DOI] [PubMed] [Google Scholar]
- Engel FL, Scott JL. The role of hormones in adipose tissue glycogen synthesis in the rat; the adrenal cortex. Endocrinology. 1951;48:56–69. doi: 10.1210/endo-48-1-56. [DOI] [PubMed] [Google Scholar]
- Galton DJ, Wilson JP. Lipogenesis in adipose tissue of patients with obesity and Cushing’s disease. Clinical Science. 1972;43:17P. doi: 10.1042/cs043017pa. [DOI] [PubMed] [Google Scholar]
- Gathercole LL, Bujalska IJ, Stewart PM, Tomlinson JW. Glucocorticoid modulation of insulin signaling in human subcutaneous adipose tissue. The Journal of Clinical Endocrinology and Metabolism. 2007;92:4332–4339. doi: 10.1210/jc.2007-1399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geer EB, Islam J, Buettner C. Mechanisms of glucocorticoid-induced insulin resistance: Focus on adipose tissue function and lipid metabolism. Endocrinology and Metabolism Clinics of North America. 2014;43:75–102. doi: 10.1016/j.ecl.2013.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halleux CM, Servais I, Reul BA, Detry R, Brichard SM. Multihormonal control of ob gene expression and leptin secretion from cultured human visceral adipose tissue: Increased responsiveness to glucocorticoids in obesity. Journal of Clinical Endocrinology and Metabolism. 1998;83:902–910. doi: 10.1210/jcem.83.3.4644. [DOI] [PubMed] [Google Scholar]
- Harris C, Roohk DJ, Fitch M, Boudignon BM, Halloran BP, Hellerstein MK. Large increases in adipose triacylglycerol flux in Cushingoid CRH-Tg mice are explained by futile cycling. American Journal of Physiology. Endocrinology and Metabolism. 2013;304:E282–E293. doi: 10.1152/ajpendo.00154.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauner H, Schmid P, Pfeiffer EF. Glucocorticoids and insulin promote the differentiation of human adipocyte precursor cells into fat cells. Journal of Clinical Endocrinology and Metabolism. 1987;64:832–835. doi: 10.1210/jcem-64-4-832. [DOI] [PubMed] [Google Scholar]
- Hazlehurst JM, Gathercole LL, Nasiri M, Armstrong MJ, Borrows S, Yu J, Wagenmakers AJM, Stewart PM, Tomlinson JW. Glucocorticoids fail to cause insulin resistance in human subcutaneous adipose tissue in vivo. The Journal of Clinical Endocrinology and Metabolism. 2013;98:1631–1640. doi: 10.1210/jc.2012-3523. [DOI] [PubMed] [Google Scholar]
- Hochberg I, Tran Q, Harvey I, Stephenson EJ, Barkan AR, Chander WF, Saltiel AR, Bridges D. Dataset for Cushing’s and Acromegaly Studies. 2015 [Google Scholar]
- Holland WL, Brozinick JT, Wang L-PP, Hawkins ED, Sargent KM, Liu Y, Narra K, Hoehn KL, Knotts Ta, Siesky A, et al. Inhibition of ceramide synthesis ameliorates glucocorticoid-, saturated-fat-, and obesity-induced insulin resistance. Cell Metabolism. 2007;5:167–179. doi: 10.1016/j.cmet.2007.01.002. [DOI] [PubMed] [Google Scholar]
- Kasumov T, Huang H, Chung Y-M, Zhang R, McCullough AJ, Kirwan JP. Quantification of ceramide species in biological samples by liquid chromatography electrospray ionization tandem mass spectrometry. Analytical Biochemistry. 2010;401:154–161. doi: 10.1016/j.ab.2010.02.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim D, Pertea G, Trapnell C, Pimentel H, Kelley R, Salzberg SL. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biology. 2013;14:R36. doi: 10.1186/gb-2013-14-4-r36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kršek M, Rosická M, Nedvídková J, Kvasničková H, Hána V, Marek J, Haluzík M, Lai EW, Pacák K. Increased lipolysis of subcutaneous abdominal adipose tissue and altered noradrenergic activity in patients with Cushing’s syndrome: An in-vivo microdialysis study. Physiological Research. 2006;55:421–428. doi: 10.33549/physiolres.930832. [DOI] [PubMed] [Google Scholar]
- Lamberts SW, Birkenhäger JC. Body composition in Cushing’s disease. The Journal of Clinical Endocrinology and Metabolism. 1976;42:864–868. doi: 10.1210/jcem-42-5-864. [DOI] [PubMed] [Google Scholar]
- Langmead B, Salzberg S. Fast gapped-read alignment with Bowtie 2. Nature Methods. 2012;9:357–360. doi: 10.1038/nmeth.1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindholm J, Juul S, Jørgensen JOL, Astrup J, Bjerre P, Feldt-Rasmussen U, Hagen C, Jørgensen J, Kosteljanetz M, Kristensen L, et al. Incidence and late prognosis of Cushing’s syndrome: A population-based study. Journal of Clinical Endocrinology and Metabolism. 2001;86:117–123. doi: 10.1210/jcem.86.1.7093. [DOI] [PubMed] [Google Scholar]
- Long W, Wei L, Barrett EJ. Dexamethasone inhibits the stimulation of muscle protein synthesis and PHAS-I and p70 S6-kinase phosphorylation. American Journal of Physiology. Endocrinology and Metabolism. 2001;280:E570–E575. doi: 10.1152/ajpendo.2001.280.4.E570. [DOI] [PubMed] [Google Scholar]
- Love MI, Huber W, Anders S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq 2. 2014 doi: 10.1186/s13059-014-0550-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lumeng CN, Saltiel AR. Inflammatory links between obesity and metabolic disease. The Journal of Clinical Investigation. 2011;121:2111–2117. doi: 10.1172/JCI57132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayo-Smith W, Hayes CW, Biller BM, Klibanski A, Rosenthal H, Rosenthal DI. Body fat distribution measured with CT: correlations in healthy subjects, patients with anorexia nervosa, and patients with Cushing syndrome. Radiology. 1989;170:515–518. doi: 10.1148/radiology.170.2.2911678. [DOI] [PubMed] [Google Scholar]
- Menconi M, Fareed M, O’Neal P, Poylin V, Wei W, Hasselgren P-O. Role of glucocorticoids in the molecular regulation of muscle wasting. Critical Care Medicine. 2007;35:S602–S608. doi: 10.1097/01.CCM.0000279194.11328.77. [DOI] [PubMed] [Google Scholar]
- Morgan SA, McCabe EL, Gathercole LL, Hassan-Smith ZK, Larner DP, Bujalska IJ, Stewart PM, Tomlinson JW, Lavery GG. 11β-HSD1 is the major regulator of the tissue-specific effects of circulating glucocorticoid excess. Proceedings of the National Academy of Sciences of the United States of America. 2014;111:E2482–E2491. doi: 10.1073/pnas.1323681111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ntali G, Grossman A, Karavitaki N. Clinical and biochemical manifestations of Cushing’s. Pituitary. 2015 doi: 10.1007/s11102-014-0631-4. [DOI] [PubMed] [Google Scholar]
- Papaspyrou-Rao S, Schneider SH, Petersen RN, Fried SK. Dexamethasone increases leptin expression in humans in vivo. Journal of Clinical Endocrinology and Metabolism. 1997;82:1635–1637. doi: 10.1210/jcem.82.5.3928. [DOI] [PubMed] [Google Scholar]
- Pleasure DE, Walsh GO, Engel WK. Atrophy of skeletal muscle in patients with Cushing’s syndrome. Archives of Neurology. 1970;22:118–125. doi: 10.1001/archneur.1970.00480200024002. [DOI] [PubMed] [Google Scholar]
- Price SR, England BK, Bailey JL, Van Vreede K, Mitch WE. Acidosis and glucocorticoids concomitantly increase ubiquitin and proteasome subunit mRNAs in rat muscle. The American Journal of Physiology. 1994;267:C955–C960. doi: 10.1152/ajpcell.1994.267.4.C955. [DOI] [PubMed] [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. 2013 [Google Scholar]
- Reddy TE, Pauli F, Sprouse RO, Neff NF, Newberry KM, Garabedian MJ, Myers RM. Genomic determination of the glucocorticoid response reveals unexpected mechanisms of gene regulation. Genome Research. 2009;19:2163–2171. doi: 10.1101/gr.097022.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rockall a, Sohaib S, Evans D, Kaltsas G, Isidori a, Monson J, Besser G, Grossman a, Reznek R. Hepatic steatosis in Cushing’s syndrome: a radiological assessment using computed tomography. European Journal of Endocrinology. 2003;149:543–548. doi: 10.1530/eje.0.1490543. [DOI] [PubMed] [Google Scholar]
- Roohk DJ, Mascharak S, Khambatta C, Leung H, Hellerstein M, Harris C. Dexamethasone-mediated changes in adipose triacylglycerol metabolism are exaggerated, not diminished, in the absence of a functional GR dimerization domain. Endocrinology. 2013;154:1528–1539. doi: 10.1210/en.2011-1047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samra JS, Clark ML, Humphreys SM, Macdonald IA, Bannister PA, Frayn KN. Effects of physiological hypercortisolemia on the regulation of lipolysis in subcutaneous adipose tissue. Journal of Clinical Endocrinology and Metabolism. 1998;83:626–631. doi: 10.1210/jcem.83.2.4547. [DOI] [PubMed] [Google Scholar]
- Segal HL, Gonzalez Lopez C. Early Effects of Glucocorticoids on Precursor Incorporation into Glycogen. Nature. 1963;200:143–144. doi: 10.1038/200143a0. [DOI] [PubMed] [Google Scholar]
- Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub TR, Lander ES, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proceedings of the National Academy of Sciences of the United States of America. 2005;102:15545–15550. doi: 10.1073/pnas.0506580102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Surjit M, Ganti KP, Mukherji A, Ye T, Hua G, Metzger D, Li M, Chambon P. Widespread negative response elements mediate direct repression by agonist-liganded glucocorticoid receptor. Cell. 2011;145:224–241. doi: 10.1016/j.cell.2011.03.027. [DOI] [PubMed] [Google Scholar]
- Tomlinson JJ, Boudreau A, Wu D, Abdou Salem H, Carrigan A, Gagnon A, Mears AJ, Sorisky A, Atlas E, Haché RJG. Insulin sensitization of human preadipocytes through glucocorticoid hormone induction of forkhead transcription factors. Molecular Endocrinology (Baltimore, Md.) 2010;24:104–113. doi: 10.1210/me.2009-0091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wing SS, Goldberg AL. Glucocorticoids activate the ATP-ubiquitin-dependent proteolytic system in skeletal muscle during fasting. The American Journal of Physiology. 1993;264:E668–E676. doi: 10.1152/ajpendo.1993.264.4.E668. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.