

Abstract
Background & aims
Substance P (SP), a neuropeptide member of the tachykinin family, plays a critical role in colitis. MicroRNAs (miRNAs) are small non-coding RNAs that negatively regulate gene expression. However, whether SP modulates expression of microRNAs in human colonic epithelial cells remains unknown.
Methods
We performed microRNA profiling analysis of SP-stimulated human colonic epithelial NCM460 cells overexpressing neurokinin-1 receptor (NCM460-NK-1R). Targets of SP-regulated microRNAs were validated by real time polymerase chain reaction (RT-PCR). Functions of miRNAs were tested in NCM460-NK-1R cells and the TNBS and DSS models of colitis.
Results
SP stimulated differential expression of 29 microRNAs, including miR-221-5p, the highest up regulated miR (by 12.6-fold) upon SP stimulation. Bioinformatic and luciferase reporter analyses identified interleukin 6 receptor (IL-6R) mRNA as a direct target of miR-221-5p in NCM460 cells. Accordingly, SP exposure of NCM460-NK-1R cells increased IL-6R mRNA expression, while overexpression of miR-221-5p reduced IL-6R expression. NF-κB and JNK inhibition decreased SP-induced miR-221-5p expression. MiR-221-5p expression was increased in both TNBS- and DSS-induced colitis and colonic biopsies from Ulcerative Colitis, but not Crohn’s Disease subjects, compared to controls. In mice, intracolonic administration of a miR-221-5p chemical inhibitor, exacerbated TNBS-and DSS-induced colitis, and increased colonic TNF-α, Cxcl10, and Col2 α 1 mRNA expression. In situ hybridization in TNBS-and DSS-exposed colons revealed increased miR-221-5p expression primarily in colonocytes.
Conclusions
Our results reveal a novel NK-1R-miR-221-5p-IL-6R network that protects from colitis. The use of miR-221-5p mimics may be a promising approach for colitis treatment.
Keywords: SP, microRNA, colitis, inflammation
Introduction
Substance P (SP), an 11 amino acid peptide member of the tachykinin family, expressed in many organs including the intestine1. SP plays a critical role in colitis pathophysiology by interacting with its high-affinity neurokinin-1 receptor (NK-1R)2–4 and activating NF-κB5–7 and JNK8-related signaling pathways in different cell types, including colonocytes5, 8, 9. NK-1R expression is increased in the colon of IBD patients10, 11, further suggesting an important role in IBD pathogenesis.
MicroRNAs (MiRs) represent a class of small non-coding single stranded RNAs that control translation and mRNA degradation by binding to target mRNAs through complementary sequences12. Functional studies demonstrated that miRs play critical roles in many physiological and pathological conditions, including inflammation13 and colitis pathogenesis14–16. There is limited evidence, however, on the interaction between the SP/NK-1R system and miRs. MiR-130a and miR-206 were found to regulate SP synthesis and release in neuronal cells17, while miR-449b and miR-500 modulate NK-1R expression in human astrocytoma cells18. Moreover, a miR-203 mimic blocked SP-mediated increased phospholipase A2 activating protein expression in keratinocytes19. However, whether SP modulates miRs in colonic epithelial cells and if this response is related to the ability of SP to regulate colitis is not known. Here, we performed a microRNA expression analysis to detect the miRNA signature upon SP stimulation of human colonic NCM460 epithelial cells.
Materials and Methods
Cell Studies and reagents
NCM460 human colonic epithelial cells overexpressing NK-1R (NCM460-NK-1R), cultured as we previously described5, were starved in serum free medium overnight, and then stimulated with 0.1 μM SP at the indicated times. CAPE (Caffeic acid phenethyl ester, Cat#C8221), a specific inhibitor of the nuclear factor κB, was from Sigma and the JNK inhibitor SP600125 was from Cell Signaling (Cat#8177). SP was purchased from Sigma (Cat#S6883). Rabbit anti-IL6R (SC-661) was from Santa Cruz Biotechnology. Mouse anti-β actin was from Sigma.
Transfection experiments
Inhibitors of miR-221-5p (Cat#4464084), negative anti-miRNAs controls (Cat#4464076), a miR-221-5p mimic (Cat#4464067), and mimic miRNA controls (Cat#4464058) were purchased from Life Technologies. Mouse anti-miR-221-5p and negative control were purchased from Exiqon (target sequence of anti-miR-221-5p is TGTAACATACGGTCC, target sequence of anti-miR-control is ACGTCTATACGCCCA). Lipid-based siPORTNeoFX Transfection Agent was purchased from Ambion (AM4511), Lipofectamine 2000 was purchased from Life Technologies (Cat#52758). NF-κB p65 siRNA (sc-29140), c-Jun siRNA (sc-29223) and control siRNAs (sc-37007) were purchased from Santa Cruz Biotechnology. NCM460-NK-1R cells were transfected with siRNA using lipofectamine RNAiMAX. For miR-221-5p silencing or overexpression, cells were transfected with antisense-miR-221-5p (as-miR-221-5p) or miR-221-5p mimic, respectively. Cells transfected with siRNA-control, antisense-control miR, or miR-mimic control served as controls.
Microarray Analysis of miRNA expression
The miRNA microarray experiments were performed using TLDA human miRNA v 1.0 system, which contains 365 microRNAs. High-capacity reverse transcription reagent for cDNA was from Applied Biosystems (Cat#4368813). Real time PCR primers were purchased from Life Technologies, except miR-221-5p primers that were from Exiqon (Cat#204302). Total RNA of the NCM460-NK-1R cells were isolated by using Trizol reagents, the RNA concentration were determined by Nanodrop. Data were collected and normalized to nonfunctional small RNA internal controls. Results were validated using quantitative reverse-transcription polymerase chain reaction (qPCR). The miRNA template for real time PCR analysis was prepared using Exiqon reagents. RNU1A1 (Exiqon Cat#203909) expression was used as internal control. The threshold cycle (Ct) value formula was used to calculate the relative expression of selected miRNAs, as we previously reported20.
Human IBD biopsy specimens
Total RNAs from colon tissues from patients with active UC (n=14), active CD (n=15) and normal subjects (n=9–10) were purchased from OriGene (Rockville, Maryland, USA). These biopsies were obtained through strict IRB protocols and with full, documented patient consent, all from accredited US-based medical institutions (www.origene.com). Conversion of cDNA of RNA samples was performed as described above, and levels of NK-1R, IL-6R and miR-221-5p were determined by quantitative RT-PCR analysis.
Luciferase Assays
IL-6R 3′UTR containing the 2 predicted binding sites and mutated sequences were chemically synthesized by GENEWIZ Inc. The wild type and mutants of the IL-6R 3′UTR sequence were then subcloned into the luciferase reporter vector from SwitchGear Genomics (Cat#32011). NCM460-NK-1R cells were transfected using lipofectamine 2000 (Invitrogen, Carlsbad, CA) with luciferase reporter constructs containing the wild type or mutant of 3′UTR of IL-6R and the miR-221-5p mimic. Cell lysates were prepared 24 hours after transfection, and luciferase activity was measured using the LightSwitch Luciferase Assay Kit from SwitchGear Genomics (Cat#LS100) according to manufacturer’s instructions.
Immunoblot analyses
NCM460-NK-1R cells were washed with ice-cold PBS and incubated with radiolabelled immumoprecipitation assay buffer containing the protease inhibitors and sodium orthovanadate (Santa Cruz) for 5 minutes on ice. Equal amount of cell lysates were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to PVDF membrane. Membranes were blocked (phosphate-buffered saline, 5% nonfat dry milk, 0.05% Tween-20) and probed with antibodies followed by corresponding horseradish peroxidase-labeled secondary antiobodies. Blots were developed with enhanced chemiluminescence reagent from Thermo Scientific (Cat#34080).
In situ hybridization
In situ hybridization was performed on mice colon tissue from C57BL/6J mice after 2,4,6-Trinitrobenzenesulfonic acid (TNBS) (7 days) or Dextran Sodium Sulfate(DSS) (6 days) treatment as we previously reported21 and described below. The sequence of the probe specific for mouse miR-221-5p (Exiqon, Cat NO. 99999-15) was as follows, 5′ACAGAAATCTACATTGTATGCCA. The experiments were performed according to the manufacturer’s instructions using miRCURY LNA microRNA ISH optimization Kit (FFPE).
Experimental colitis
TNBS-and DSS-induced colitis were performed by modifications of previously described protocols described by us21. All animal studies were approved by the institutional animal care and use committee. Total RNA from colonic tissues from the TNBS model was purified using the miReasy Mini Kit (Qiagen), while from the DSS model by the Lithium Chloride precipitation method22. Tissue sections were scored for histopathology analyses in a double-blinded manner, as previously reported21.
TNBS-induced colitis
Animals were maintained at UCLA animal research facility and received standard pelleted chow and water ad libitum. to 6–8 week old male C57BL/6J mice (n=5–8 per group) by 50 μl intracolonic injection of 40 mg/kg TNBS (Fluka, Ronkonkoma, NY) in 30% ethanol, using a 1-mL syringe (Becton Dickinson, Laguna Hills, CA, USA) fitted with a polyethylene cannula (Intramedic PE-20 tubing; Becton Dickinson). Control groups were injected with 50 μl of 30% ethanol intracolonically. Mice were held head down for 1 min after the enema administration to ensure the containment of the TNBS solution into the colon. Mice returned to their cages, sacrificed after 7 days by cervical dislocation, and colonic tissues were collected. To assess the effect of miR-221-5p in TNBS induced colitis, mice were intracolonically administrated with 40mg/kg TNBS, and at days 1, 3 and 5, were intracolonically injected with 40 μg of miRCURY ™ LNA Inhibitor probe, in vivo against mmu-miR-221-5p (Exiqon). Briefly, the appropriate amount of oligonucleotides against mmu-miR-221-5p and its respective control were re-suspended in 100 μL Opti-MEM with 2 μL lipofectamine 2000 and administered intracolonicaly. At day 7, mice were sacrificed and the colon tissue was collected for H&E staining and RNA expression analysis.
DSS-induced colitis
To assess the miRNA expression, 1% DSS was dissolved in the drinking water and administrated to C57BL/6J mice for 5 days (n=5–8 mice per group). To test the effect of miR-221-5p in DSS induced colitis, mice were administrated with 1% DSS, then at day 1, 3 and 5, mice were intracolonically injected with 40 μg anti-miR-221-5p or anti-miR-control as above. At day 6, mice were sacrificed and colon tissue was collected for H&E staining and RNA expression analysis.
Statistical analysis
Student t-test for two groups comparisons and ANOVA for multiple groups comparisons were carried out to determine statistically significant differences between the experimental groups. P<0.05 was considered statistically significant. All results were expressed as means ±SD.
Results
SP regulates expression of microRNAs in human colonic epithelial cells
To investigate the SP effects on the miRNAs expression in human colonic epithelial cells, we used 0.1 μM SP to stimulate NCM460-NK-1R cells in serum free medium at 0.5 and 6 hours. This SP concentration was used in previous studies to stimulate NCM460-NK-1R cells9, 23. Total RNA was isolated and miRNA array analysis was carried out as we previously reported24. We found that 29 miRNAs (18 up-regulated and 11 down regulated) had altered expression by SP stimulation (Figure 1A). Hierarchical clustering analysis revealed that 4 different clusters of miRNAs were affected by SP. Specifically, the up-regulated miRNAs were divided into miRNAs that showed an early and stable response (expression was increased at 30 minutes stimulation) and a late response (expression was no different at 30 minutes stimulation, but altered after 6 hours stimulation) (Figure 1B). Similarly, the down regulated miRNAs were divided into early (cluster 3) or late (cluster 4) response clusters (Figure -1C). Among the up-regulated miRNAs, miR-221 and miR-222 showed the highest increase. MiR-221 and miR-222 are encoded in a cluster on the X chromosome. Pre-miR-221 is processed in the cytoplasm and produces two forms of mature miRNA: miR-221-5p (also named as miR-221*) and miR-221-3p (normally referred as miR-221). Real time PCR analysis validated that miR-221-5p, miR-221-3p, miR-222-5p and miR-222-3p were substantially increased by SP at 30 min, with a sustained increase over controls at 6, 12, and 24 hours (Figure 2). Among those 4 four mature miRNAs, miR-221-5p showed a more substantial increase compared to other three mature miRNAs. Thus, we next focused on SP-miR-221-5p interactions.
Figure 1. SP-regulated microRNAs in colon epithelial cells.
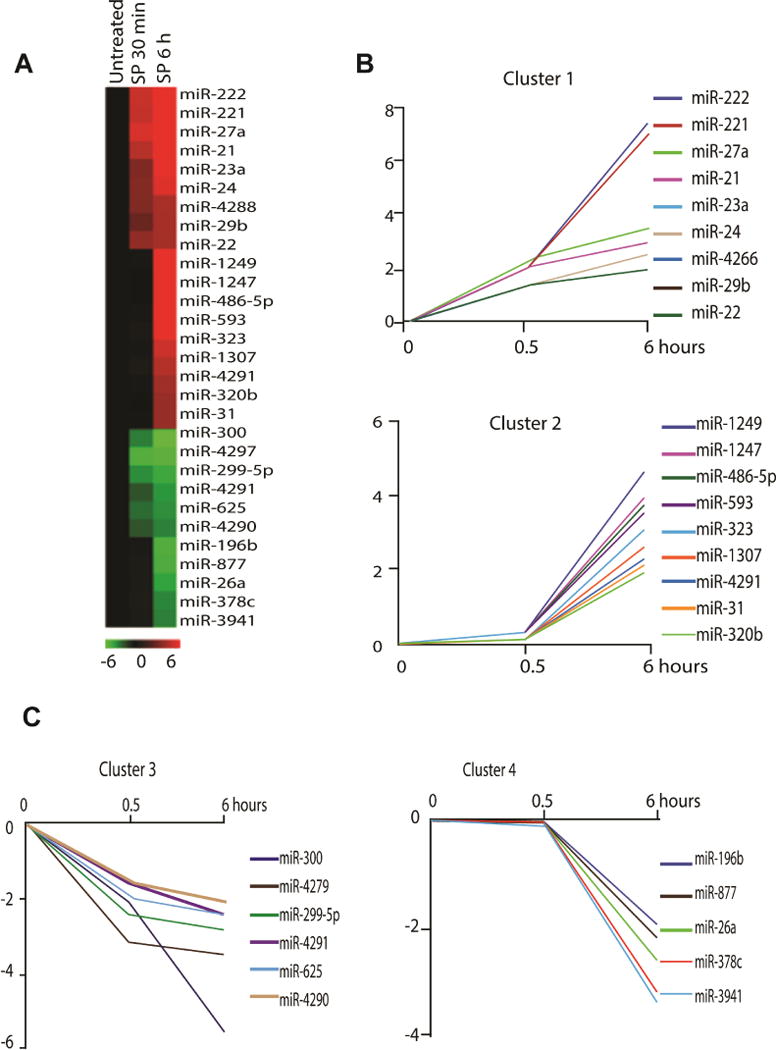
(A). Heat map represents differentially expressed microRNAs after 0.1 μM SP treatment (0.5 and 6 hours) in NCM460-NK-1R cells. Red indicates up-regulated miRNAs, and green indicates down-regulated microRNAs. Cluster of (B) SP-induced and (C) SP-suppressed microRNAs according to their response dynamics.
Figure 2. Quantitative real-time PCR detection of SP induced mature miRNA expression.
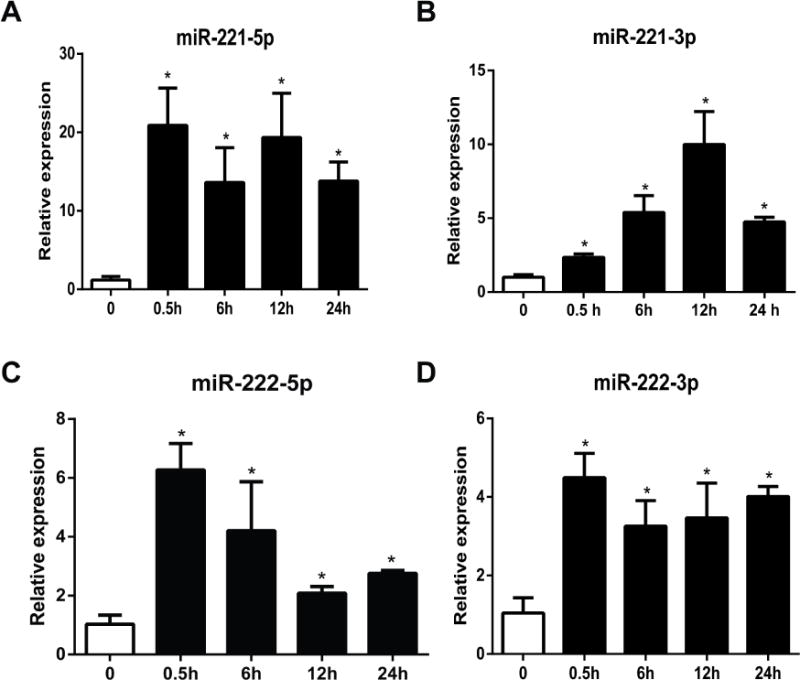
Mature miRNAs expression quantification was performed after 0.1 μM SP treatment (0.5, 6, 12 and 24 hours) of NCM460-NK-1R cells. Expression levels of miR-221-5p, miR-221-3p, miR-222-5p, and miR-222-3p were assessed by RT-PCR analysis. *p<0.05, data show mean ± standard deviation of triplicate samples per experimental condition.
MiR-221-5p and NK-1R are up-regulated in human ulcerative colitis tissues
Our results above indicate that NK-1R signaling is linked to increased miR-221-5p expression in human colonic epithelial cells. Previous results demonstrated increased NK-1R expression in the colon of IBD patients10, 25. Therefore, we next examined whether miR-221-5p expression is also increased in human IBD samples. Our results show increased expression of miR-221-5p in UC biopsies (n= 14), compared to biopsies from control subjects (n= 10, p=0.01, Figure 5A). However, there is no significant differential expression of miR-221-3p, miR-222-5p and miR-221-3p in those samples. However, expression of miR-221-5p was not significantly different in CD tissues (n=15) compared to control biopsies (n=9, p=0.28). Examination of the same UC tissues also revealed increased NK-1R mRNA expression compared to controls (Figure 5A).
Figure 5. Level of miR-221-5p in IBD tissues.
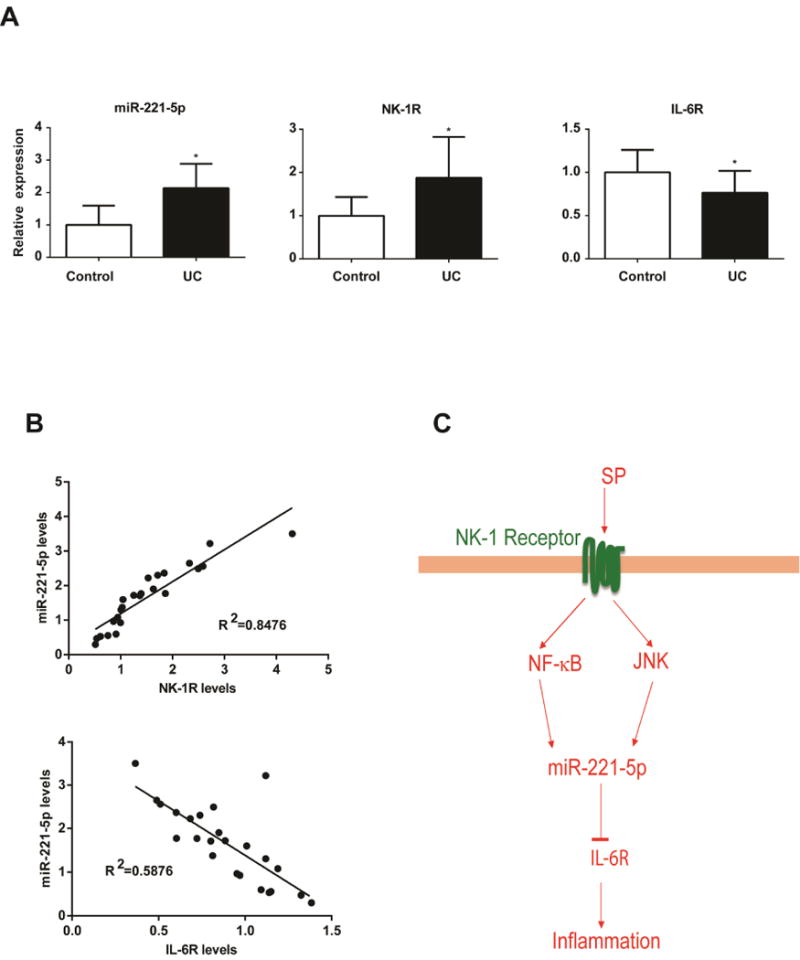
Expression of miR-221-5p, NK-1R, and IL-6R in human UC tissue samples (n=14) compared to control tissue samples (n=10) is shown in A. *p<0.05, data show mean ± standard deviation. B. Correlation of miR-221-5p levels with NK-1R and IL-6R. The correlation coefficient was calculated using Prism6 (GraphPad Software, Inc.). C. Schematic representation of proposed SP-microRNA signaling pathway in human colonic epithelial cells. After binding with NK-1R receptor, SP stimulates miR-221-5p expression via NF-κB and JNK activation in human colonic epithelial cells. MiR-221-5p regulates inflammation through down regulation of IL-6R expression.
SP regulates miR-221-5p expression through NF-κB and JNK pathways
Since NK-1R coupling activates the transcription factor NF-κB and JNK5–8, and these pathways are involved in experimental colitis26, 27, we examined their involvement in SP regulated miR-221-5p expression. Bioinformatics analysis by using online transcription factor search software TFSEARCH indicated the presence of NF-κB binding sites (The sequence is 5′-GGAACGTCCC-3′, from 45750897bp to 45750906 bp of X chromosome, localized in 4740 bp upstream of miR-221, confirming a recent report28). To examine the functional role of NF-κB on miR-221 expression levels, we used the NF-κB inhibitor CAPE that prevents translocation of the p65 subunit of NF-κB to the nucleus29. NCM460-NK-1R cells were treated with 10 μM CAPE for 30 min before SP (0.1 μM) stimulation for 6 hours. Our results showed that CAPE significantly reduced miR-221-5p expression (Figure 3A). Moreover, silencing of the p65 subunit of NF-κB using siRNA substantially decreased SP-induced miR-221-5p expression in NCM460-NK-1R cells, compared to control siRNA (Figure 3B).
Figure 3. SP regulates miR-221-5p expression through NF-κB and JNK pathways.
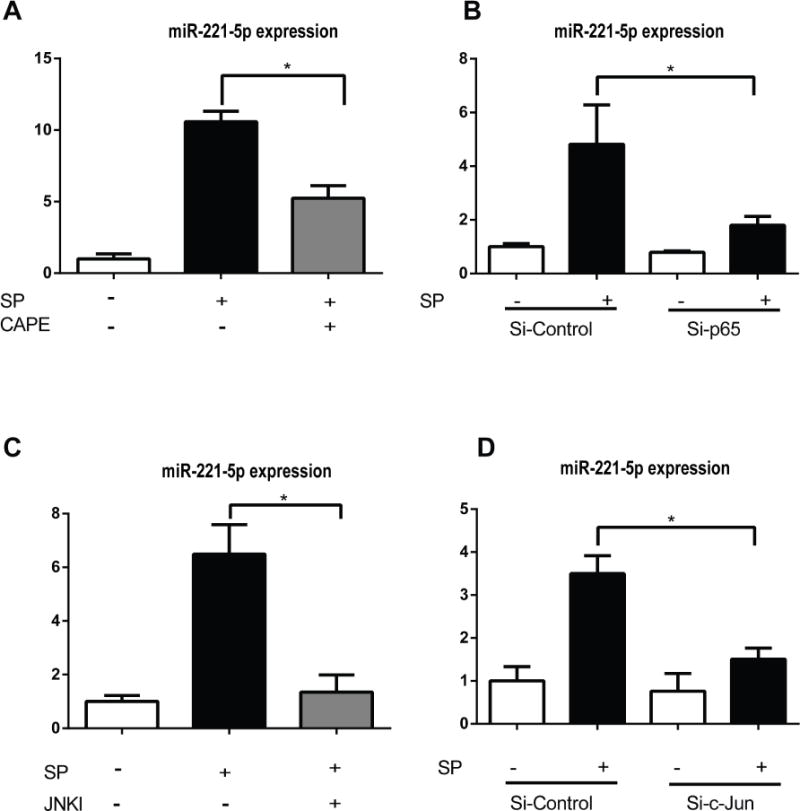
MiR-221-5p expression was assessed by RT-PCR. (A). NCM460-NK-1R cells were pretreated with the NF-κB inhibitor CAPE (10μM) 30 minutes before SP exposure for 6 hours. (B). MiR-221-5p levels in cells transfected with siRNA-p65 for 48 hours before SP exposure of 6 hours. (C). NCM460-NK-1R cells were pretreated with the JNK inhibitor SP600125 30 minutes before SP exposure for 6 hours. (D). MiR-221-5p levels in cells transfected with siRNA-c-Jun for 48 hours before SP exposure for 6 hours. *p<0.05, data show mean ± standard deviation of triplicate.
Previous results indicated that c-Jun regulates miR-221 expression in lung, liver, and prostate cancer cells28, 30. To address the involvement of c-Jun in SP-increased miR-221-5p expression, we pretreated NCM460-NK-1R cells with 50 μM of the C-Jun N-terminal Kinase (JNK) inhibitor SP60012531 and found that SP-stimulated miR-221-5p expression was almost normalized by SP600125 (Figure 3C). To confirm this result we silenced c-Jun using a siRNA approach as described in Methods. As shown in Fig. 3D, c-Jun silencing reduced SP-induced increased miR-221-5p expression. Taken together, these results indicate that SP regulates miR-221-5p expression in human colonic epithelial cells through activation of NF-κB and JNK.
MiR-221-5p regulates directly IL-6R expression in human colonic epithelial cells
MicroRNAs negatively regulate gene expression by binding to complementary sequences in the 3′ untranslated region (3′UTR) of their target genes. Since SP regulates expression of several pro-inflammatory genes, we next determined whether miR-221-5p regulates directly inflammation-related genes using bioinformatics. Our bioinformatics analysis, using microRNA target prediction program from DIANA lab32, showed the presence of 2 binding sites for miR-221-5p in the 3′UTR of human IL-6R mRNA sequence (NM_00565.3) (Figure 4A). A luciferase activity assay using plasmids containing the miR-221-5p binding sites in 3′UTR of IL-6R indicated that cells transfected with a miR-221-5p mimic had lower luciferase activity compared to control-miR-mimic-treated NCM460-NK-1R cells (n=3, p<0.05, Figure 4C), suggesting that IL-6R 3′UTR is sufficient to confer miR-221-5p regulation (Figure 4B,C). Importantly, when both the miR-221-5p seed regions in the IL-6R 3′UTR were mutated, miR-221-5p regulation was abolished (Figure 4B, C).
Figure 4. MiR-221-5p regulates IL-6R expression.
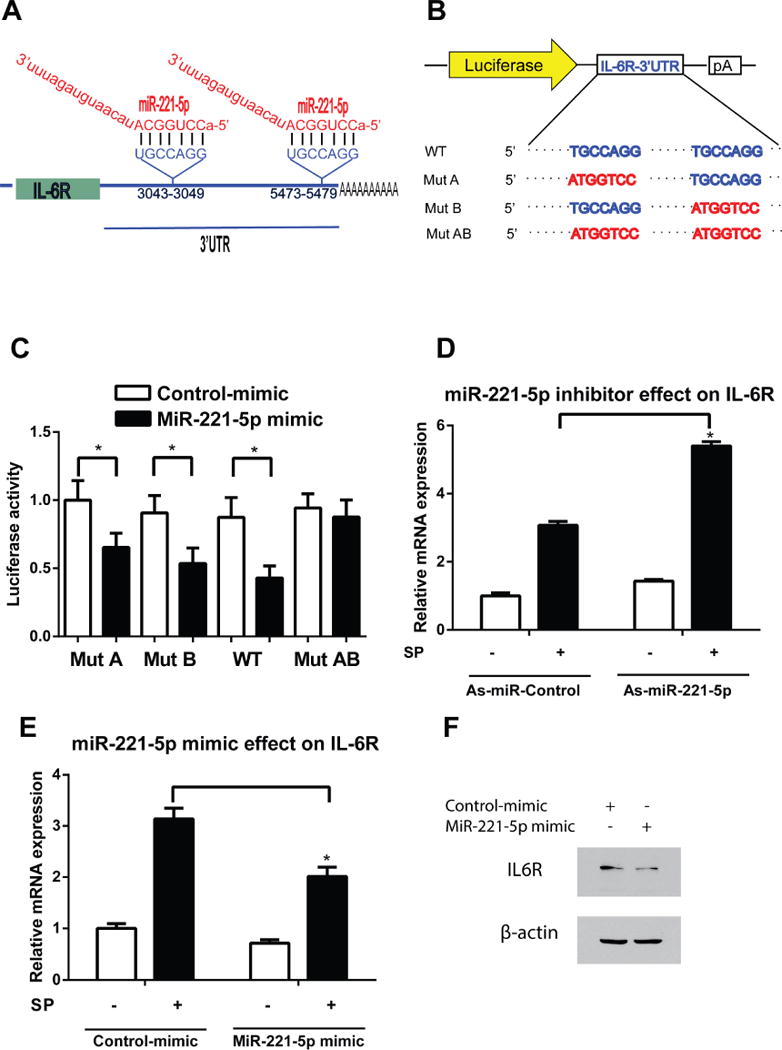
(A) Human IL-6R mRNA contains two predicted miR-221-5p binding sites. (B) A diagram of IL-6R 3′ UTR reporters. (C) WT or mutant IL-6R 3′UTR luciferase constructs were co-transfected with miR-221-5p mimic or control mimic in NCM460-NK-1R cells. Luciferase activity was determined 24 hours after transfection and normalized luciferase activity is shown. *p<0.05, data show mean ± standard deviation of triplicate. (D). MiR-221-5p down regulation increased IL-6R mRNA expression in response to SP stimulation. *p<0.05, data show mean ± standard deviation of triplicate (E). Up-regulation of miR-221-5p decrease IL-6R mRNA expression in response to SP stimulation. *p<0.05, data show mean ± standard deviation of triplicate. (F) IL-6R and β-actin protein levels are assessed by Western blot in miR-221-5p mimic or control-mimic treated NCM460-NK-1R cells.
To elucidate the effects of miR-221-5p on IL-6R mRNA expression levels, NCM460-NK-1R cells were transfected with 0.1 μM of antisense miR-221-5p (as-miR-221-5p), or miR-221-5p mimic, or their controls using siPORTNeoFX. After 48 hours, cells were placed in serum-free medium overnight, and then stimulated with 0.1 μM SP. Total RNA was isolated for RT-PCR analysis. As shown in Figure 4D, miR-221-5p silencing increased IL-6R levels. Conversely, miR-221-5p mimic treatment reduced IL-6R levels compared to the mimic control-treated cells (Figure 4E). Thus miR-221-5p inhibits IL-6R mRNA expression in human colonic epithelial cells. Moreover, treatment of cells with miR-221-5p mimic reduces IL-6R protein level (Figure 4F). Together, these results indicate that in human colonocytes, IL-6R is a downstream target of miR-221-5p.
Based on the negative correlation of miR-221-5p to IL-6R expression shown above, we examined IL-6R mRNA levels in the same colon samples. As shown in Figure 5A, IL-6R expression levels were significantly decreased (by 23%, p<0.05), compared to controls. NK-1R levels were correlated positively with miR-221-5p (R2=0.8476), whereas miR-221-5p levels correlated negatively with IL-6R (R2=0.5876) (Figure 5B). These results indicate that the NK-1R-miR-221-5p signaling is activated and IL-6R expression is decreased in the colon during active UC.
MiR-221-5p is up-regulated in experimental colitis
To start addressing a possible functional role for miR-221-5p in the pathophysiology of colitis we first examined colonic expression of miR-221-5p in two chemical models of experimental colitis. As shown in Figure 6A, colonic expression of miR-221-5p was significantly up-regulated 3 and 7 days after intracolonic administration of TNBS. Moreover, expression of miR-221-5p was also increased by 2-fold 5 days after administration of 1% DSS into the drinking water of C57BL/6J mice (Figure 6C). To examine cellular localization of miR-221-5p we performed in situ hybridization using a miR-221-5p probe in colon tissue sections obtained from control and TNBS- or DSS-treated mice as described in Methods. Our results showed that miR-221-5p expression is increased in the colonic mucosa, primarily in colonic epithelial cells in TNBS- or DSS – exposed mice, compared to controls (Figure 6B and 6D).
Figure 6. MiR-221-5p expression is increased in experimental colitis.
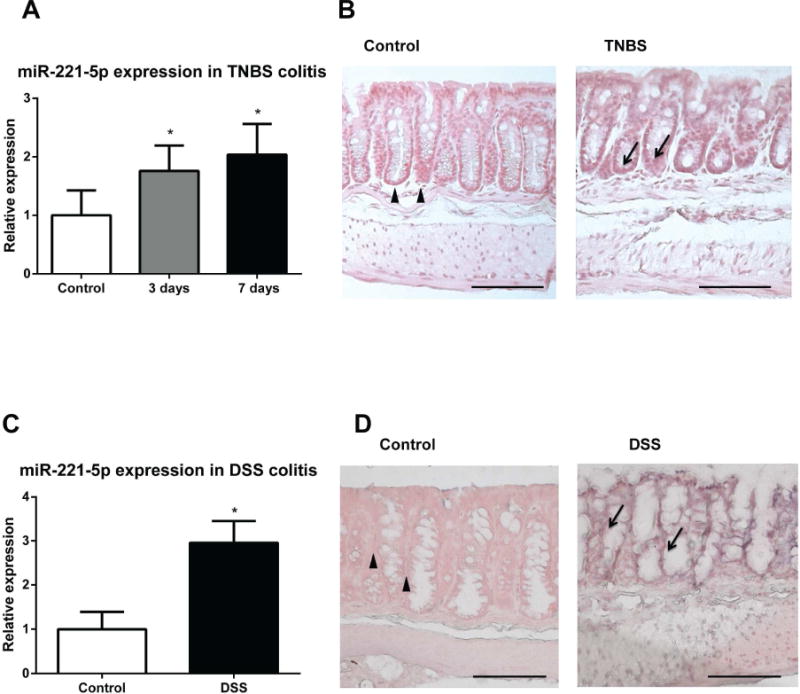
Expression levels of miR-221-5p are assessed by RT-PCR. (A) MiR-221-5p is increased in the colonic mucosa of TNBS-exposed mice. *p<0.05 vs. control (n=8). (B) Representative images of in situ hybridization of miR-221-5p of colon tissues from TNBS-treated C57BL6/J mice and their control counterparts. (C). MiR-221-5p is increased in the colonic mucosa of DSS – administered mice. *p<0.05 vs. control (n=7) and DSS treated mice (n=5). (D). Representative images of in situ hybridization of miR-221-5p of colon tissues from DSS-treated C57BL6/J mice and their control counterparts. Scale bar, 100μm.
Intracolonic silencing of miR-221-5p enhances TNBS-induced colitis
We next tested the importance of miR-221-5p in colitis pathogenesis in the TNBS colitis model. Mice were administered intracolonicaly with 40mg/kg TNBS at day 0 and at day 1, 3 and 5 anti-sense (as) miR-221-5p or as-miRNA-control (40 μg/mouse) were administered intracolonicaly. At day7 colon tissues were collected for further analysis (see experimental design diagram Figure 7A). To verify efficient silencing of endogenous miR-221-5p we measured colonic miR-221-5p expression in the different mouse groups and found reduced endogenous miR-221-5p levels (by 80%) in as-miR-221-5p-exposed mice compared to those exposed to as-control miR (Figure 7D). Mice treated with TNBS showed epithelial destruction and inflammation and increased histopathology score compared to intracolonic administration of vehicle (Figure 7C). We found no difference in histopathology score and levels of miR-221-5p expression in mice treated with either TNBS alone, or TNBS with control as-miR, or TNBS with lipofectamine vehicle (Figure 7-C and D). However, TNBS-exposed mice treated with as-miR-221-5p demonstrated more convoluted crypts and epithelial cell damage than as-miRNA control-treated mice (Figure 7B). These changes are also depicted in increased histopathology score in TNBS-exposed mice following as-miR-221-5p treatment (Figure 7C). We also found a marked increase in the levels of TNF-α expression after silencing of miR-221-5p expression in TNBS-exposed mice, while expression of the neutrophil chemoattractant Cxcl10 was also significantly increased (Figure 7E). Our results also demonstrate increased expression of Col2α1 mRNA (Figure 7E), an important molecule involved in tissue remodeling and fibrosis33, 34, and a trend for increased expression of CXCL1 mRNA (by 2-fold), which was not statistically significant (n=8 per group, p>0.05). We did not find any significant further increase in MPO mRNA expression following si-miR-221-5p treatment (data not shown). Moreover, there was no increase in colonic IL-6R levels in TNBS-exposed as-miR-221-5p-treated mice (data not shown). These results indicated that miR-221-5p regulates experimental colitis by modulating expression of proinflammatory and fibrosis-related genes.
Figure 7. As-miR-221-5p exacerbates TNBS-induced colitis.
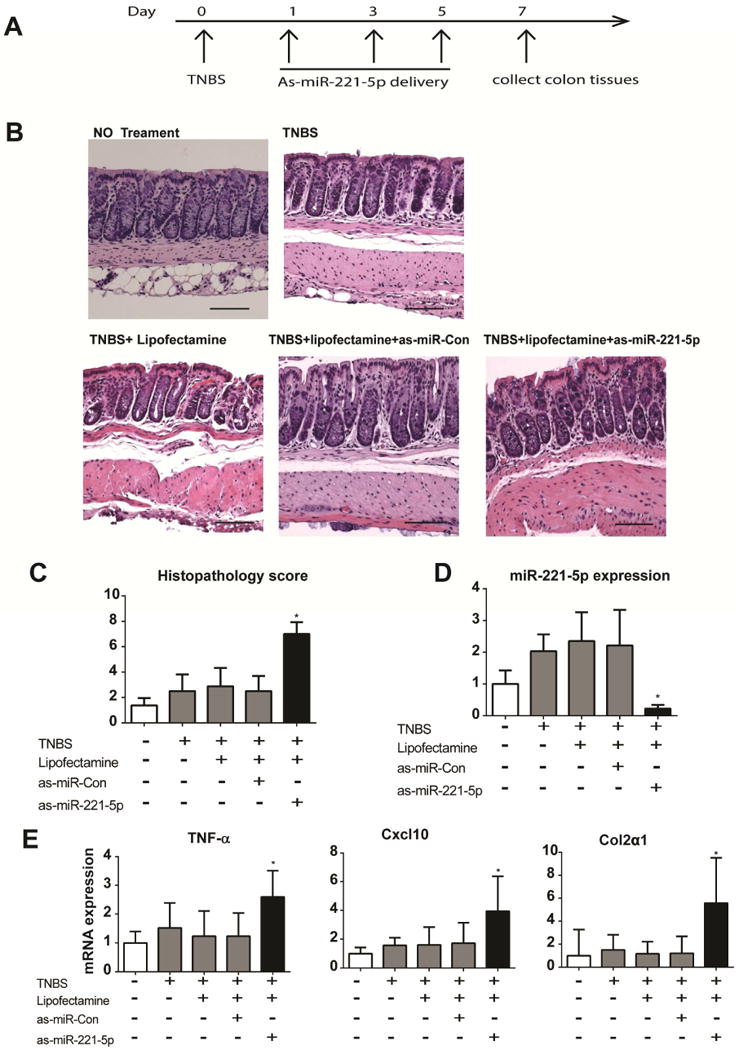
(A) Timeline of as-miR-221-5p treatment in TNBS induced colitis. (B) TNBS-induced histological changes in colons of with controls or as-miR-221-5p treated C57BL6/J mice. Scale bar, 100μm. (C). Histopathology score of as-miR-221-5p treated mice with TNBS-induced colitis. (D). Expression level of miR-221-5p was assessed by real-time PCR after intracolonic administration of as-miR-221-5p. * indicates p<0.05 compared with controls (n=8). (E). Expression of TNF-α, Cxcl10 and Col2α1 in the colon tissue of as-miR-221-5p or as-miR-control treated TNBS mice colitis models. * indicates p<0.05 compared with controls (n=8).
Anti-miR-221-5p exacerbates DSS-induced colitis
Currently, there is no animal colitis model can simulate every aspect of human IBD, and each model has its own limits35. To further confirm the anti-inflammatory effect of anti-miR-221-5p in colitis, we also use the DSS colitis model. Mice were provided with 1% DSS in their drinking water at day 0. As-miR-221-5p or control as-miR was injected intracolonicaly at day 1, 3 and 5; mice were sacrificed at day 6 and their colons were harvested for further analysis. Treatment with anti-miR-221-5p worsened histologic damage and colitis (Figure 8B), histopathology score (Figure 8D), and mucosal PMN infiltrate (Figure 8E), compared to colon of DSS-exposed animals exposed to control as-miR. Consistent with results from TNBS colitis model, anti-miR-221-5p treatment greatly reduced miR-221-5p expression in the colon tissue (Figure 8C), increased the expression of TNF-α, Cxcl10, and Col2 α 1 (Figure 8F), and CXCL1 mRNA (n=8, p>0.05) in the DSS model of colitis with si-miR-221-5p treatment. These data further confirm an anti-inflammatory role for miR-221-5p in vivo.
Figure 8. As-miR-221-5p regulates inflammation in DSS-induced colitis models.
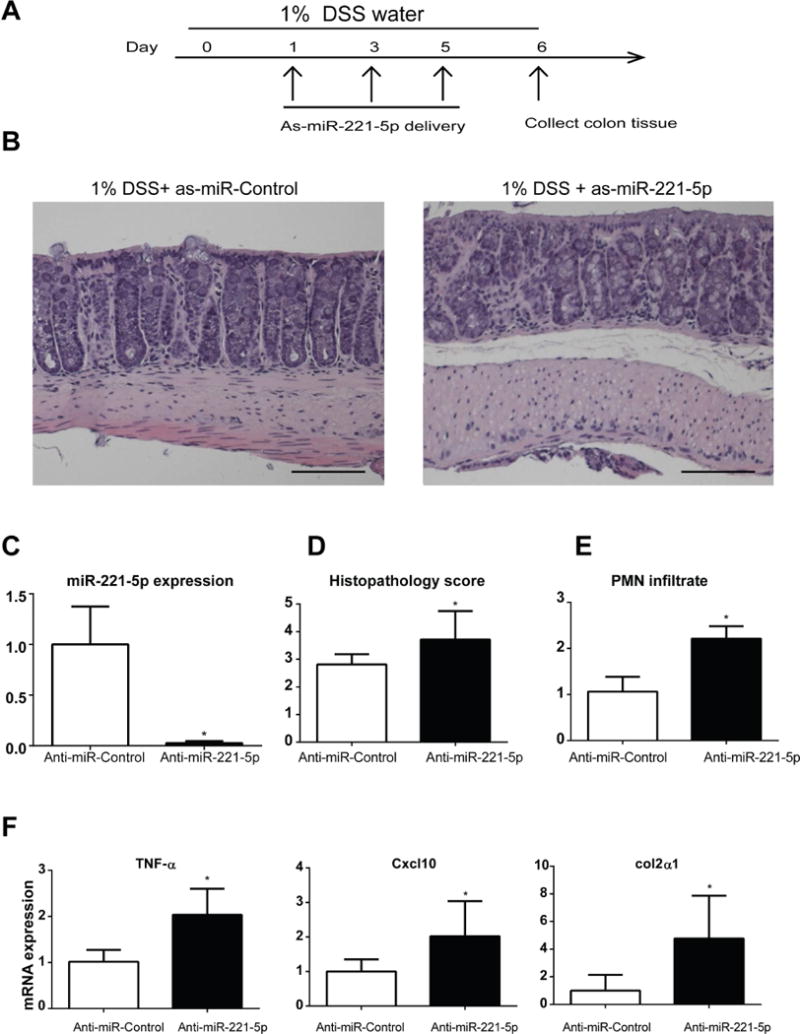
(A) Timeline of as-miR-221-5p treatment in 1% DSS-induced colitis. (B) Histological changes in colons of with controls or as-miR-221-5p treated C57BL6/J mice in DSS colitis model. Scale bar, 100μm. (C). Expression level of miR-221-5p after intracolonic administration of as-MiR-221-5p. * indicates p<0.05 compared with controls (n=6). (D). Histopathology sore of as-miR-221-5p treated mice with DSS-induced colitis. (E). The PMN infiltration sore of as-miR-221-5p treated mice with DSS-induced colitis. (F). Expression of TNF-α, Cxcl10 and Col2α1 in the colon tissue of as-miR-221-5p or anti-miR-control treated DSS-induced colitis models. * indicates p<0.05 compared with controls (n=6).
Discussion
MiRNA expression is deregulated across a broad spectrum of inflammatory disorders13, including IBD15, 16. Levels of SP are elevated in IBD tissues1. SP and its high affinity receptor NK-1R have been also implicated in the pathophysiology of both acute and chronic colitis by regulating several genes involved in the promotion of colitis, as well mucosal healing following colitis1, 36, 37. However, the contribution of miRs and miR-regulated pathways involved in the intestinal inflammatory mechanisms of SP has not been studied. Our results indicate that coupling of SP to NK-1R in human colonic epithelial cells regulates differential expression of 29 miRNAs (Fig. 1), among them miR-21 have been implicated in the pathogenesis of colitis and IBD38–40. We also show that miR-221 and miR-222 represent the highest up-regulated miRs in response to SP (Fig. 1A & B) and miR-221-5p affects the pathophysiology of colitis through stimulation of an anti-inflammatory feedback network (Fig. 5C). Most importantly, our results indicate that this SP-NK-1R-dependent miR-221-5p-IL-6R circuit is activated in human colonic epithelial cells and UC specimens (Fig. 5), suggesting an important role for NK-1R-dependent miRNA regulation in colitis.
We demonstrate that silencing of endogenous colonic miR-221-5p enhances experimental colitis in two different mouse chemical models. Mucosal histologic damage worsened, and colonic mRNA levels of TNF-α, Cxcl10 and Col2α1 were increased following intracolonic silencing of miR-221-5p in both TNBS and DSS-induced colitis (Fig. 7 and 8). Interestingly, TNF-α, CXCL10 and Col2α1 have been associated with the pathophysiology of IBD. Neutralization of TNF-α with monoclonal antibodies represents one of the most promising recent therapies in IBD41. CXCL10 is increased in inflamed colons of IBD patients and stimulates monocyte, natural killer, and T-cell migration42, 43, while Col2α1 is important in tissue remodeling and fibrosis33, 34.
We also present direct molecular and biochemical evidence that IL-6R is a novel downstream target of miR-221-5p that may mediate intestinal anti-inflammatory signaling following SP-miR-221-5p interactions in human colonocytes. IL-6R is implicated in cytokine-cytokine receptor signaling that involves the JAK-STAT signaling pathways, known to be dysregulated in T-cell-mediated, and DSS-, and TNBS-induced colitis20, 44, 45. Additionally, the JAK-STAT pathway is involved in the pathogenesis of UC46, while treatment with antibodies against IL-6R attenuates immune-mediated and chemically induced colitis47.
The ability of SP to activate IL-6R expression and the identification of IL-6R as a downstream target of miR-221-5p in human epithelial cells has not previously recognized. Interestingly, our results indicate that SP induces IL-6R expression, while exposure of NCM460-NK-1R cells to a miR-221-5p mimic inhibits IL-6R expression (Figure 4E and F). This contradictory response is likely due to multiple signaling pathways regulated by SP-NK-1R interactions(reviewed in1). Thus, SP-NK-1R signaling may regulate IL-6R expression not only through miR-221-5p, but also via other transcription factors activated via_NK-1R signaling that can affect IL-6R expression1, while, as shown here, miR-221-5p directly regulates IL-6R expression through binding IL-6R 3′UTR. In addition, our bioinformatics analysis indicates that mouse IL-6R mRNA has no binding sites for miR-221-5p, suggesting that in the mouse other(s) miR-221-5p downstream targets may be involved in the effects of this miR in amelioration of colitis suggested by our in vivo results with miR-221-5p silencing.
A shown in Figure 3, the mechanism by which SP-NK-1R interactions regulate expression of miR-221-5p involves activation of NF-κB and JNK, important signaling pathways known to be regulated by NK-1R activation1. Our results are in line with previous reports demonstrating that NF-κB induces the expression of miR-221 in prostate carcinoma, glioblastoma, and colorectal cancer cells48. Importantly, our finding demonstrates that miR-221-5p act as an anti-inflammatory miRNA, by controlling IL-6R expression in human epithelial cells. IL-6R is implicated in cytokine-cytokine receptor interactions and the JAK-STAT signaling pathways, known to be dysregulated in T-cell-20, DSS-45, and TNBS-induced colitis44.
Compared to controls, IL-6R expression is decreased in inflamed colon biopsies from UC patients, while in the same samples expression of NK-1R and miR-221-5p are increased (Figure 5A and B). These findings combined with our in vitro analysis (Figure 3 and 4) demonstrate a positive correlation between miR-221-5p and NK-1R and an inverse correlation with IL-6R in UC. This NK-1R-miR-221-5p-dependent pathway, its association with the NF-κB and JNK signaling pathways and the IL-6R as a downstream target of this miR are summarized in the diagram under Figure 5C. Previous studies, however, reported increased soluble IL-6R in human IBD serum49, while another study did not find differences in the relative expression of IL-6R in blood T cells and lamina propria T cells between CD, UC and control patients47. These differences in IL-6R expression levels between our study and the studies of Atreya et al47 and Mitsuyama et al49 may be due to different IL-6R measurement methods (ELISA, FACS, vs q-RT-PCR) and/or materials used (serum, and lamina propria T cells, vs mucosal biopsies).
Our results show increased expression of miR-221-5p in colonic biopsies from UC patients (Fig. 5A), a disease state highly associated with colon cancer50, and in the colonic mucosa of mice with experimental colitis (Fig. 6A &6C). MiR-221-5p is also up-regulated in cancer-associated fibroblasts, compared to normal fibroblast cells, in line with a role for miR-221-5p in tumorigenesis51. Yuan et al found that miR-221-5p expression levels correlate negatively with colorectal cancer-associated metastasis by inhibiting MBD2 expression52. Interestingly, Rokavec et al found that IL-6R/STAT3 pathways promote epithelial-to-mesenchymal transition (EMT)-mediated colorectal cancer invasion and metastasis53. These results51–53, together with our findings, suggest that miR-221-5p may regulate colon cancer metastasis through IL-6R/STAT3-related pathways.
In summary, we have identified miR-221-5p as a SP-responsive miRNA that regulates IL-6R mRNA and protein expression in human colonic epithelial cells in vitro and regulates experimental colitis in vivo. Our studies support that the possibility that miR-221-5p may serve as an important anti-“inflamiR” by controlling IL-6R signaling pathways under pathological conditions. Strategies activating miR-221-5p expression may represent a novel therapeutic approach for IBD treatment.
Acknowledgments
Funding: Supported by US Public Health Service grant R01 DK47343, R01 DK60729, Neuroendocrine Assay Core, P50 DK64539 (C.P.), a Crohn’s and Colitis Foundation Fellowship Award (KB), and a CURE:DDRC P30 DK 41301 Pilot and Feasibility study (KF), and Animal Core (CP and KF).
Abbreviations in this paper
- miRNA
microRNA
- mRNA
messenger RNA
- NF-κB
Nuclear factor-κB
- SP
substance P
- NK-1R
neurokinin-1 receptor
- UTR
untranslated region
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Conflicts of interest: The authors disclose no conflicts.
Author contributions: Overall project design and hypotheses were developed by K Fang and C Pothoulakis. K Fang conducted experiments, analyzed the data and wrote the manuscript. K Bakirtzi generated DSS colitis model mouse colon samples. A Sideri, C Polytarchou, and Dimitrios Iliopoulos performed the microRNA array analysis. IKM Law contributed toc experimental design and data analysis. All authors participated in revising the manuscript and agreed to the final version. C Pothoulakis supervised the overall project.
References
- 1.Steinhoff MS, von Mentzer B, Geppetti P, Pothoulakis C, Bunnett NW. Tachykinins and their receptors: contributions to physiological control and the mechanisms of disease. Physiol Rev. 2014;94:265–301. doi: 10.1152/physrev.00031.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Pothoulakis C, Castagliuolo I, LaMont JT, Jaffer A, O’Keane JC, Snider RM, Leeman SE. CP-96,345, a substance P antagonist, inhibits rat intestinal responses to Clostridium difficile toxin A but not cholera toxin. Proc Natl Acad Sci U S A. 1994;91:947–51. doi: 10.1073/pnas.91.3.947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Stucchi AF, Shebani KO, Leeman SE, Wang CC, Reed KL, Fruin AB, Gower AC, McClung JP, Andry CD, O’Brien MJ, Pothoulakis C, Becker JM. A neurokinin 1 receptor antagonist reduces an ongoing ileal pouch inflammation and the response to a subsequent inflammatory stimulus. Am J Physiol Gastrointest Liver Physiol. 2003;285:G1259–67. doi: 10.1152/ajpgi.00063.2003. [DOI] [PubMed] [Google Scholar]
- 4.Castagliuolo I, Riegler M, Pasha A, Nikulasson S, Lu B, Gerard C, Gerard NP, Pothoulakis C. Neurokinin-1 (NK-1) receptor is required in Clostridium difficile-induced enteritis. J Clin Invest. 1998;101:1547–50. doi: 10.1172/JCI2039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zhao D, Kuhnt-Moore S, Zeng H, Pan A, Wu JS, Simeonidis S, Moyer MP, Pothoulakis C. Substance P-stimulated interleukin-8 expression in human colonic epithelial cells involves Rho family small GTPases. Biochem J. 2002;368:665–72. doi: 10.1042/BJ20020950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Karagiannides I, Kokkotou E, Tansky M, Tchkonia T, Giorgadze N, O’Brien M, Leeman SE, Kirkland JL, Pothoulakis C. Induction of colitis causes inflammatory responses in fat depots: evidence for substance P pathways in human mesenteric preadipocytes. Proc Natl Acad Sci U S A. 2006;103:5207–12. doi: 10.1073/pnas.0600821103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lieb K, Fiebich BL, Berger M, Bauer J, Schulze-Osthoff K. The neuropeptide substance P activates transcription factor NF-kappa B and kappa B-dependent gene expression in human astrocytoma cells. J Immunol. 1997;159:4952–8. [PubMed] [Google Scholar]
- 8.Tansky MF, Pothoulakis C, Leeman SE. Functional consequences of alteration of N-linked glycosylation sites on the neurokinin 1 receptor. Proc Natl Acad Sci U S A. 2007;104:10691–6. doi: 10.1073/pnas.0703394104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Koon HW, Zhao D, Zhan Y, Moyer MP, Pothoulakis C. Substance P mediates antiapoptotic responses in human colonocytes by Akt activation. Proc Natl Acad Sci U S A. 2007;104:2013–8. doi: 10.1073/pnas.0610664104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Goode T, O’Connell J, Anton P, Wong H, Reeve J, O’Sullivan GC, Collins JK, Shanahan F. Neurokinin-1 receptor expression in inflammatory bowel disease: molecular quantitation and localisation. Gut. 2000;47:387–96. doi: 10.1136/gut.47.3.387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Koon HW, Zhao D, Xu H, Bowe C, Moss A, Moyer MP, Pothoulakis C. Substance P-mediated expression of the pro-angiogenic factor CCN1 modulates the course of colitis. Am J Pathol. 2008;173:400–10. doi: 10.2353/ajpath.2008.080222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Valencia-Sanchez MA, Liu J, Hannon GJ, Parker R. Control of translation and mRNA degradation by miRNAs and siRNAs. Genes & development. 2006;20:515–24. doi: 10.1101/gad.1399806. [DOI] [PubMed] [Google Scholar]
- 13.Iliopoulos D. MicroRNA circuits regulate the cancer-inflammation link. Sci Signal. 2014;7:pe8. doi: 10.1126/scisignal.2005053. [DOI] [PubMed] [Google Scholar]
- 14.Nguyen HT, Dalmasso G, Muller S, Carriere J, Seibold F, Darfeuille-Michaud A. Crohn’s disease-associated adherent invasive Escherichia coli modulate levels of microRNAs in intestinal epithelial cells to reduce autophagy. Gastroenterology. 2014;146:508–19. doi: 10.1053/j.gastro.2013.10.021. [DOI] [PubMed] [Google Scholar]
- 15.Koukos G, Polytarchou C, Kaplan JL, Morley-Fletcher A, Gras-Miralles B, Kokkotou E, Baril-Dore M, Pothoulakis C, Winter HS, Iliopoulos D. MicroRNA-124 regulates STAT3 expression and is down-regulated in colon tissues of pediatric patients with ulcerative colitis. Gastroenterology. 2013;145:842–52. doi: 10.1053/j.gastro.2013.07.001. e2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wu F, Zikusoka M, Trindade A, Dassopoulos T, Harris ML, Bayless TM, Brant SR, Chakravarti S, Kwon JH. MicroRNAs are differentially expressed in ulcerative colitis and alter expression of macrophage inflammatory peptide-2 alpha. Gastroenterology. 2008;135:1624–1635. doi: 10.1053/j.gastro.2008.07.068. e24. [DOI] [PubMed] [Google Scholar]
- 17.Greco SJ, Rameshwar P. MicroRNAs regulate synthesis of the neurotransmitter substance P in human mesenchymal stem cell-derived neuronal cells. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:15484–9. doi: 10.1073/pnas.0703037104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sanchez Freire V, Burkhard FC, Kessler TM, Kuhn A, Draeger A, Monastyrskaya K. MicroRNAs may mediate the down-regulation of neurokinin-1 receptor in chronic bladder pain syndrome. The American journal of pathology. 2010;176:288–303. doi: 10.2353/ajpath.2010.090552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sun Y, Li XQ, Sahbaie P, Shi XY, Li WW, Liang DY, Clark JD. miR-203 regulates nociceptive sensitization after incision by controlling phospholipase A2 activating protein expression. Anesthesiology. 2012;117:626–38. doi: 10.1097/ALN.0b013e31826571aa. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fang K, Zhang S, Glawe J, Grisham MB, Kevil CG. Temporal genome expression profile analysis during t-cell-mediated colitis: identification of novel targets and pathways. Inflammatory bowel diseases. 2012;18:1411–23. doi: 10.1002/ibd.22842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Law IK, Bakirtzi K, Polytarchou C, Oikonomopoulos A, Hommes D, Iliopoulos D, Pothoulakis C. Neurotensin-regulated miR-133alpha is involved in proinflammatory signalling in human colonic epithelial cells and in experimental colitis. Gut. 2014 doi: 10.1136/gutjnl-2014-307329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Viennois E, Chen F, Laroui H, Baker MT, Merlin D. Dextran sodium sulfate inhibits the activities of both polymerase and reverse transcriptase: lithium chloride purification, a rapid and efficient technique to purify RNA. BMC research notes. 2013;6:360. doi: 10.1186/1756-0500-6-360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Koon HW, Zhao D, Na X, Moyer MP, Pothoulakis C. Metalloproteinases and transforming growth factor-alpha mediate substance P-induced mitogen-activated protein kinase activation and proliferation in human colonocytes. The Journal of biological chemistry. 2004;279:45519–27. doi: 10.1074/jbc.M408523200. [DOI] [PubMed] [Google Scholar]
- 24.Bakirtzi K, Hatziapostolou M, Karagiannides I, Polytarchou C, Jaeger S, Iliopoulos D, Pothoulakis C. Neurotensin signaling activates microRNAs-21 and -155 and Akt, promotes tumor growth in mice, and is increased in human colon tumors. Gastroenterology. 2011;141:1749–61. doi: 10.1053/j.gastro.2011.07.038. e1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Renzi D, Pellegrini B, Tonelli F, Surrenti C, Calabro A. Substance P (neurokinin-1) and neurokinin A (neurokinin-2) receptor gene and protein expression in the healthy and inflamed human intestine. The American journal of pathology. 2000;157:1511–22. doi: 10.1016/S0002-9440(10)64789-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sun P, Zhou K, Wang S, Li P, Chen S, Lin G, Zhao Y, Wang T. Involvement of MAPK/NF-kappaB signaling in the activation of the cholinergic anti-inflammatory pathway in experimental colitis by chronic vagus nerve stimulation. PloS one. 2013;8:e69424. doi: 10.1371/journal.pone.0069424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Assi K, Pillai R, Gomez-Munoz A, Owen D, Salh B. The specific JNK inhibitor SP600125 targets tumour necrosis factor-alpha production and epithelial cell apoptosis in acute murine colitis. Immunology. 2006;118:112–21. doi: 10.1111/j.1365-2567.2006.02349.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Galardi S, Mercatelli N, Farace MG, Ciafre SA. NF-kB and c-Jun induce the expression of the oncogenic miR-221 and miR-222 in prostate carcinoma and glioblastoma cells. Nucleic Acids Res. 2011;39:3892–902. doi: 10.1093/nar/gkr006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Natarajan K, Singh S, Burke TR, Jr, Grunberger D, Aggarwal BB. Caffeic acid phenethyl ester is a potent and specific inhibitor of activation of nuclear transcription factor NF-kappa B. Proceedings of the National Academy of Sciences of the United States of America. 1996;93:9090–5. doi: 10.1073/pnas.93.17.9090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Garofalo M, Di Leva G, Romano G, Nuovo G, Suh SS, Ngankeu A, Taccioli C, Pichiorri F, Alder H, Secchiero P, Gasparini P, Gonelli A, Costinean S, Acunzo M, Condorelli G, Croce CM. miR-221&222 regulate TRAIL resistance and enhance tumorigenicity through PTEN and TIMP3 downregulation. Cancer cell. 2009;16:498–509. doi: 10.1016/j.ccr.2009.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 31.Bennett BL, Sasaki DT, Murray BW, O’Leary EC, Sakata ST, Xu W, Leisten JC, Motiwala A, Pierce S, Satoh Y, Bhagwat SS, Manning AM, Anderson DW. SP600125, an anthrapyrazolone inhibitor of Jun N-terminal kinase. Proceedings of the National Academy of Sciences of the United States of America. 2001;98:13681–6. doi: 10.1073/pnas.251194298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Maragkakis M, Reczko M, Simossis VA, Alexiou P, Papadopoulos GL, Dalamagas T, Giannopoulos G, Goumas G, Koukis E, Kourtis K, Vergoulis T, Koziris N, Sellis T, Tsanakas P, Hatzigeorgiou AG. DIANA-microT web server: elucidating microRNA functions through target prediction. Nucleic acids research. 2009;37:W273–6. doi: 10.1093/nar/gkp292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hanley KP, Oakley F, Sugden S, Wilson DI, Mann DA, Hanley NA. Ectopic SOX9 mediates extracellular matrix deposition characteristic of organ fibrosis. The Journal of biological chemistry. 2008;283:14063–71. doi: 10.1074/jbc.M707390200. [DOI] [PubMed] [Google Scholar]
- 34.Nakken KE, Nygard S, Haaland T, Berge KE, Arnkvaern K, Odegaard A, Labori KJ, Raeder MG. Multiple inflammatory-, tissue remodelling- and fibrosis genes are differentially transcribed in the livers of Abcb4 (−/−) mice harbouring chronic cholangitis. Scandinavian journal of gastroenterology. 2007;42:1245–55. doi: 10.1080/00365520701320521. [DOI] [PubMed] [Google Scholar]
- 35.Hoffmann JC, Pawlowski NN, Kuhl AA, Hohne W, Zeitz M. Animal models of inflammatory bowel disease: an overview. Pathobiology: journal of immunopathology, molecular and cellular biology. 2002;70:121–30. doi: 10.1159/000068143. [DOI] [PubMed] [Google Scholar]
- 36.Koon HW, Pothoulakis C. Immunomodulatory properties of substance P: the gastrointestinal system as a model. Ann N Y Acad Sci. 2006;1088:23–40. doi: 10.1196/annals.1366.024. [DOI] [PubMed] [Google Scholar]
- 37.Gross KJ, Pothoulakis C. Role of neuropeptides in inflammatory bowel disease. Inflamm Bowel Dis. 2007;13:918–32. doi: 10.1002/ibd.20129. [DOI] [PubMed] [Google Scholar]
- 38.Paraskevi A, Theodoropoulos G, Papaconstantinou I, Mantzaris G, Nikiteas N, Gazouli M. Circulating MicroRNA in inflammatory bowel disease. Journal of Crohn’s & colitis. 2012;6:900–4. doi: 10.1016/j.crohns.2012.02.006. [DOI] [PubMed] [Google Scholar]
- 39.Yang Y, Ma Y, Shi C, Chen H, Zhang H, Chen N, Zhang P, Wang F, Yang J, Zhu Q, Liang Y, Wu W, Gao R, Yang Z, Zou Y, Qin H. Overexpression of miR-21 in patients with ulcerative colitis impairs intestinal epithelial barrier function through targeting the Rho GTPase RhoB. Biochemical and biophysical research communications. 2013;434:746–52. doi: 10.1016/j.bbrc.2013.03.122. [DOI] [PubMed] [Google Scholar]
- 40.Shi C, Liang Y, Yang J, Xia Y, Chen H, Han H, Yang Y, Wu W, Gao R, Qin H. MicroRNA-21 knockout improve the survival rate in DSS induced fatal colitis through protecting against inflammation and tissue injury. PloS one. 2013;8:e66814. doi: 10.1371/journal.pone.0066814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nielsen OH, Ainsworth MA. Tumor necrosis factor inhibitors for inflammatory bowel disease. N Engl J Med. 2013;369:754–62. doi: 10.1056/NEJMct1209614. [DOI] [PubMed] [Google Scholar]
- 42.Mendez Samperio P, Trejo A, Miranda E. Role of type I interferon in the bacillus Calmette-Guerin-induced expression of CXCL10 from human monocytes. Mediators of inflammation. 2004;13:343–8. doi: 10.1155/S096293510400050X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Thomas S, Baumgart DC. Targeting leukocyte migration and adhesion in Crohn’s disease and ulcerative colitis. Inflammopharmacology. 2012;20:1–18. doi: 10.1007/s10787-011-0104-6. [DOI] [PubMed] [Google Scholar]
- 44.Kretzmann NA, Fillmann H, Mauriz JL, Marroni CA, Marroni N, Gonzalez-Gallego J, Tunon MJ. Effects of glutamine on proinflammatory gene expression and activation of nuclear factor kappa B and signal transducers and activators of transcription in TNBS-induced colitis. Inflammatory bowel diseases. 2008;14:1504–13. doi: 10.1002/ibd.20543. [DOI] [PubMed] [Google Scholar]
- 45.Fang K, Bruce M, Pattillo CB, Zhang S, Stone R, 2nd, Clifford J, Kevil CG. Temporal genomewide expression profiling of DSS colitis reveals novel inflammatory and angiogenesis genes similar to ulcerative colitis. Physiological genomics. 2011;43:43–56. doi: 10.1152/physiolgenomics.00138.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Sugimoto K. Role of STAT3 in inflammatory bowel disease. World journal of gastroenterology: WJG. 2008;14:5110–4. doi: 10.3748/wjg.14.5110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Atreya R, Mudter J, Finotto S, Mullberg J, Jostock T, Wirtz S, Schutz M, Bartsch B, Holtmann M, Becker C, Strand D, Czaja J, Schlaak JF, Lehr HA, Autschbach F, Schurmann G, Nishimoto N, Yoshizaki K, Ito H, Kishimoto T, Galle PR, Rose-John S, Neurath MF. Blockade of interleukin 6 trans signaling suppresses T-cell resistance against apoptosis in chronic intestinal inflammation: evidence in crohn disease and experimental colitis in vivo. Nature medicine. 2000;6:583–8. doi: 10.1038/75068. [DOI] [PubMed] [Google Scholar]
- 48.Liu S, Sun X, Wang M, Hou Y, Zhan Y, Jiang Y, Liu Z, Cao X, Chen P, Chen X, Tao Y, Xu C, Mao J, Cheng C, Li C, Hu Y, Wang L, Chin YE, Shi Y, Siebenlist U, Zhang X. A microRNA 221- and 222-Mediated Feedback Loop, via PDLIM2, Maintains Constitutive Activation of NFkappaB and STAT3 in Colorectal Cancer Cells. Gastroenterology. 2014 doi: 10.1053/j.gastro.2014.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mitsuyama K, Toyonaga A, Sasaki E, Ishida O, Ikeda H, Tsuruta O, Harada K, Tateishi H, Nishiyama T, Tanikawa K. Soluble interleukin-6 receptors in inflammatory bowel disease: relation to circulating interleukin-6. Gut. 1995;36:45–9. doi: 10.1136/gut.36.1.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Rutter MD, Saunders BP, Wilkinson KH, Rumbles S, Schofield G, Kamm MA, Williams CB, Price AB, Talbot IC, Forbes A. Thirty-year analysis of a colonoscopic surveillance program for neoplasia in ulcerative colitis. Gastroenterology. 2006;130:1030–8. doi: 10.1053/j.gastro.2005.12.035. [DOI] [PubMed] [Google Scholar]
- 51.Zhao L, Sun Y, Hou Y, Peng Q, Wang L, Luo H, Tang X, Zeng Z, Liu M. MiRNA expression analysis of cancer-associated fibroblasts and normal fibroblasts in breast cancer. The international journal of biochemistry & cell biology. 2012;44:2051–9. doi: 10.1016/j.biocel.2012.08.005. [DOI] [PubMed] [Google Scholar]
- 52.Yuan K, Xie K, Fox J, Zeng H, Gao H, Huang C, Wu M. Decreased levels of miR-224 and the passenger strand of miR-221 increase MBD2, suppressing maspin and promoting colorectal tumor growth and metastasis in mice. Gastroenterology. 2013;145:853–64. doi: 10.1053/j.gastro.2013.06.008. e9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Rokavec M, Oner MG, Li H, Jackstadt R, Jiang L, Lodygin D, Kaller M, Horst D, Ziegler PK, Schwitalla S, Slotta-Huspenina J, Bader FG, Greten FR, Hermeking H. IL-6R/STAT3/miR-34a feedback loop promotes EMT-mediated colorectal cancer invasion and metastasis. The Journal of clinical investigation. 2014;124:1853–67. doi: 10.1172/JCI73531. [DOI] [PMC free article] [PubMed] [Google Scholar]