

Abstract
The pharyngeal mesoderm of developing embryos contributes to broad regions of head and heart musculature. We have developed a novel method to study head and heart progenitor cell development with pharyngeal arches (also known as branchial arches) ex vivo. Using this method, we have recently described that the second pharyngeal arch contains self-renewing heart progenitors and serves as a microenvironment for expansion of the progenitors during mouse heart development. The progenitor cells remain undifferentiated and expansive inside the arch, but quickly become functional cardiomyocytes as they migrate out of the arch. We also reported that first pharyngeal arch contains muscle progenitors giving rise to myotubes after leaving the arch. Here, we demonstrate the procedure for the dissection and ex-vivo culture of first and second pharyngeal arches from developing mouse embryos. The method enables one to study head and heart progenitor/muscle development, including cardiomyocyte and myotube formation in detail ex vivo.
Keywords: Cardiac Progenitors, Microenvironment, Pharyngeal arch, Cardiogenesis, Head muscle progenitors, Stem cells
Introduction
Pharyngeal mesoderm cells give rise to parts of the heart and the pharyngeal muscles. During embryonic development, multipotent cardiac progenitor cells from the second heart field migrate from the pharyngeal mesoderm and populate the cardiac outflow tract and right ventricle, and their abnormal development is closely associated with congenital heart disease—the leading cause of birth defects and birth defect-related deaths in humans1-3. Recent studies have demonstrated that pharyngeal mesoderm contributes to head muscles, in addition to the heart, making the mesoderm a critical part of the cardio-craniofacial development4. Thus, the developmental processes including induction, proliferation, and differentiation of head and heart 5-7 progenitors in the pharyngeal mesoderm are under active investigation. Until recently, it remained unknown whether cardiac progenitor cells undergo expansion without differentiation, partly due to the lack of information on their cellular environment. Our recent study suggests that pharyngeal arches serve as a microenvironment for the renewal of cardiac and muscle progenitors and can be cultured ex-vivo over several weeks8. This explant method offers a novel and unique opportunity to study the development of cardio-craniofacial progenitors ex vivo.
Protocol
All mice were maintained at an American Association for the Accreditation of Laboratory Animal Care (AAALAC)-accredited animal facility at the Johns Hopkins University and housed in accordance with the procedures outlined in the Guide for the Care and Use of Laboratory Animals. The Institutional Animal Care and Use Committee (IACUC) approved all experimental protocols.
1. Experimental Preparation
1.1. Coat 12-well plate with 10% Fetal Bovine Serum (FBS) in Phosphate Buffered Solution (PBS) for 60 min.
1.22. Remove coating solution and add 200 μL of Serum-Free Media (SFM). Twirl plate to make sure that a film of media covers the entire surface area
2. Surgical Procedures
2.1. Euthanize pregnant mice using the institution's animal care committee-approved protocol followed by cervical dislocation to ensure complete euthanasia.
2.2. Spray and clean the belly region of the mouse with 70% ethanol. Maintain strict sterile techniques to avoid contamination.
2.3. Use forceps and scissors to make a 0.5 cm2 incision at the navel region.
2.4. Use fingers to pinch/grab the skin above and below the incision and gently pull the skin in opposite directions (head and tail).
2.5. Use forceps and scissors to make an initial incision in the membrane at the navel region. Gently cut the membrane in V-shape from the navel to the ovaries on each side (2 × 1.5 cm2), thereby revealing the uterus.
2.6. Use forceps to pinch the oviduct connecting the uterus to the ovary and scissors to pinch the oviduct on the ovary side, thus freeing the uterus from the ovary.
2.7. Gently pull up the uterus by the oviduct with the forceps and cut the uterus free from the bladder region with scissors.
2.8. Pull the uterus further up by the oviduct with the forceps, and cut the oviduct with scissors on the other side, thereby releasing the uterus from the mouse.
2.9. Transfer the uterus to a 50 mL tube and add 20 mL of cold sterile PBS with Ca2+ and Mg2+. PBS must contain Ca2+ and Mg2+ during embryo dissection.
2.10. Gently shake the tube for 10 seconds to wash the uterus for blood.
2.11. Transfer the uterus from the tube with forceps and place it in a 10 mL dish and add 5 mL of cold PBS on top of the uterus.
2.12. Place the dish with the uterus under a stereomicroscope.
Note: Steps 2.13-2.19 requires a stereomicroscope.
2.13. Use two pairs of forceps to gently open the uterus and dissect out amniotic sacs with embryos from the uterus one by one.
2.13.1. Use two pairs of forceps to carefully open the amniotic sac that surrounds the embryo, pinch the sac with one forceps and gently remove it from the embryo, use the other forceps to cut and release the amniotic sac from the embryo. If a specific genotype needed, keep amniotic sac for genotyping.
2.14. Place the embryo on the side and use forceps to cut the 1st and 2nd arch posteriorly to the heart between the arch and the pharyngeal pouch (Figure 1A″).
Figure 1. Mouse embryo (Mesp1cre, Ai9) 9.5 days post fertilization (E.9.5).
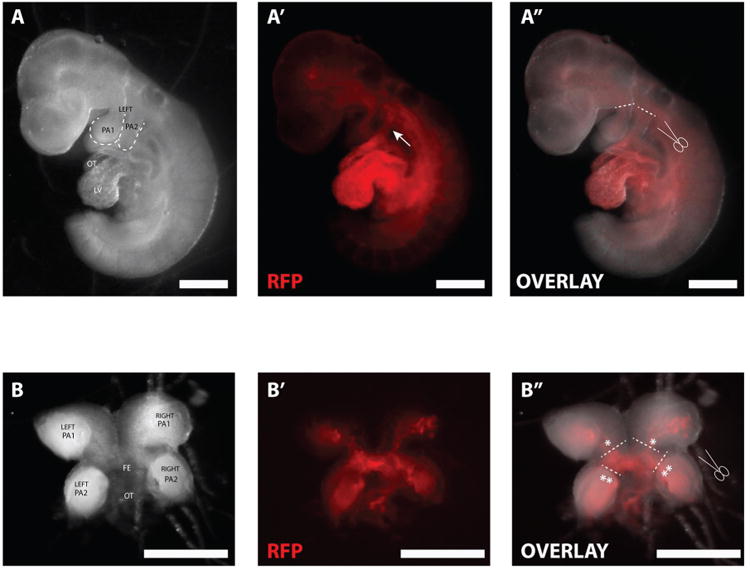
A) Left side of embryo. (PA: Pharyngeal Arch, OT: Outflow tract, LV: Left ventricle. A′) Mesp1 linage-trace; RFP marks Mesp1+ cells and their progeny. Arrow indicates RFP+ Mesp1-progeny in PA2 that is continuous with the OT. A″) Dashed line indicate where to cut PA1 and PA2 posterior to the heart. B.) Butterfly-like right and left PA1 and PA2 after dissection from the embryo. FE: Foregut endoderm. B′: RFP marks Mesp1 progeny in PA1 and PA2. B″) * and ** along with the dashed lines indicates where to cut and release PA1 and PA2 from the OT and FE. Scale bar: 500 μm.
2.15. Gently flip the embryo to the other side and use forceps to cut the 1st and 2nd arch posteriorly to the heart between the arch and the pharyngeal pouch.
2.16. Use forceps to cut the cardiac outflow tract that is still connecting the 1st and 2nd pharyngeal arches to the embryo. Remove the arches, which are still attached together in the shape of a butterfly (Figure 1B), from the embryo.
2.17. Use forceps to pinch and release the 1st pharyngeal arches from the remaining cardiac outflow tract and foregut endoderm (Figure 1B″ indicated with dashed line and *).
2.18. Use forceps to pinch and release the 2nd pharyngeal arches from the remaining cardiac outflow tract and foregut endoderm (Figure 1B″ indicated with dashed line and **).
2.19. Transfer each pair of arches separately using forceps and place both arches in the middle of a designated well. Plate 1st and 2nd pharyngeal arches in separate wells. Ensure that the arches are in contact with the film of media added in step 1.2. It is crucial that arches are NOT covered by medium, as they need to make contact with the surface of the well for attachment during this period.
3. Incubation and Imaging
3.1 Incubate arches plated in the 12-well plate in 37 °C/5% CO2 for 2 h for arches to attach to the well surface area.
3.2. Gently add 200 μL of 37 °C SFM down the side of the well. Make sure that arches remain attached and incubate overnight.
3.3. Next day, visually check that pharyngeal arches are attached and gently add 200 μL of 37 °C SFM down the side of the well. If arches remain unattached and floating, gently remove media with a pipette without removing arches until only a film of media is left. Use pipette tip to place arches in the middle of the well and repeat step 3.1.
3.4. Monitor daily; add ∼100-200 μL of media every 2nd day to replace any evaporated media.
Note: 24-48 h after migrating cells appear around the arch and will proliferate and migrate from arch over the next 5-8 days. After 36-72 h formation of beating cardiomyocytes can be observed from the 2nd pharyngeal arch (Figure 2B). Myotube formation can be observed from the 1st pharyngeal arch 3-7 days after attachment (Figure 2A).
Figure 2. Cultured pharyngeal arches.
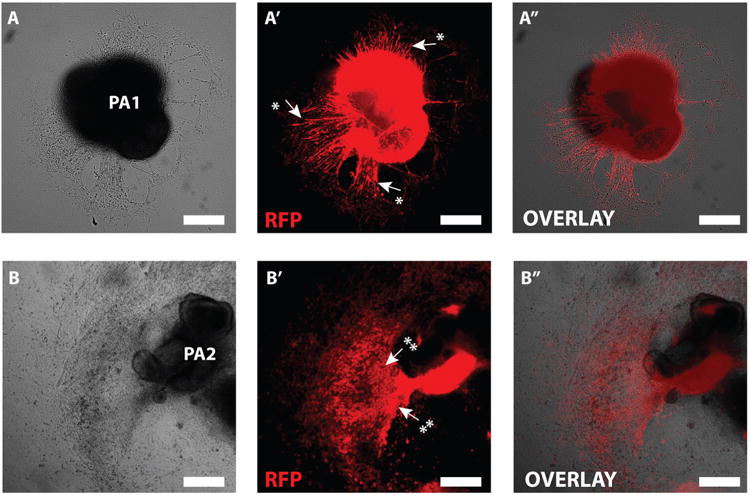
A) Cultured PA1 (3 days of culture). A′) RFP marks Mesp1 progeny in PA1. Arrows and * indicate early myotube formation. A″) Overlay of RFP expression in PA1. B) Cultured PA2 (8 days of culture). B′) RFP marks Mesp1 progeny in PA2. Arrows and ** indicate beating area of cardiomyocytes (modified from Shenje et al. 8). B″ Overlay of RFP expression in PA2. Scale bar: 250 μm.
Representive Results
During development, facial muscle and heart progenitors can be traced as they proliferate and migrate from the 1st and 2nd pharyngeal arch, to become head and heart musculature, respectively (figure 1A, A′ and A″). Culturing pharyngeal arches offers a unique way to study heart and muscle development in detail ex vivo. After dissection and attachment of pharyngeal arches, migrating cells from attached arches can be observed within 24-48 of culture. Within 3 days of culture myotube formation can be observed from 1st pharyngeal arches and cardiomyocyte formation from the 2nd pharyngeal arches (Figure 2A and 2B). Cardiomyocyte formation can be visually confirmed by spontaneously contracting clusters of migrating cells and/or analysis of specific cardiomyocyte markers (e.g. Cardiac Troponin T). Myotube/facial muscle formation can be visually observed by long elongated (up to 500 μm in length) spontaneously twitching cells and/or analysis of specific muscle markers (e.g. Myogenin). By using the cre-lox system in mice, mesoderm progeny can be traced by fluorescent reporters upon cre-expression during ex vivo culture (Figure 2A′and 2B′). Likewise, with this method it is possible to study the fate of progeny wherein a specific gene of interest is conditionally knocked out or overexpressed, which otherwise would result in early embryonic lethality as we previously described8.
Discussion
In this video, we demonstrate how to isolate and culture first and second pharyngeal arches of 9.5 days old mouse embryos. Pharyngeal arches are transient, segmented bulges that appear on the craniolateral side of developing embryos 9, which contain multi-potent cardiac progenitor cells—building blocks to make the heart during embryogenesis 10,11—in second arches and head muscle progenitors in first arches 8.
Dissection of pharyngeal arches requires advanced mouse embryo dissection skills. Once arches have been successfully dissected using this stepwise protocol and transferred to pre-coated wells, it is critical that arches attach to the bottom area. In this protocol, we coat wells with 10% FBS followed by culture in SFM resulting in attachment and growth of >75% of arches, however other coating materials can be used (matrigel, gelatin, fibronectin etc.) ´as well as serum containing media.
The pharyngeal arches serves as primordia for a multitude of structures during development. After initial formation of the pharyngeal arches (8 days post fertilization in mice), CPCs expand in the PA2 and migrate into the cardiac outflow tract where they differentiate into cardiac cells, thereby serving as a renewable source of CPCs that supplies cells needed to sustain heart growth. As the pharyngeal arches are temporal developmental structures (E8-11.5) and migration of CPCs into the outflow tract occurs from approximately E8-E10) this brings certain limitations to the technique. For CPC studies, we recommend using pharyngeal arches dissected between E9-10, at this stage each PA2 contain approximately 5-800 RFP+ cells (Mesp1 linage trace).
With fluorescent labeling, this ex-vivo culture allows us to monitor developing progenitors and their differentiation into beating cardiomyocytes or myotubes in real-time. In addition, this ex-vivo culture system can be used to study roles of cell-autonomous and microenvironmental factors affecting cardiac/muscle progenitor cell proliferation, migration, and differentiation using existing Cre/loxP lines. As such, this system is expected to facilitate cardiac/muscle progenitor– niche studies, which may lead to a better understanding of heart/muscle formation and disease.
Acknowledgments
We thank Kwon laboratory members for helpful discussions. This work was supported by grants from NHLBI/NIH (R01HL111198) and Maryland Stem Cell Research Fund (MSCRF).
Footnotes
A complete version of this article that includes the video component is available at http://dx.doi.org/10.3791/52876.
Disclosures: The authors have nothing to disclose.
Contributor Information
Peter Andersen, Email: pander31@jhmi.edu, Division of Cardiology, Institute for Cell Engineering, Johns Hopkins University School of Medicine, Baltimore, MD.
Chulan Kwon, Division of Cardiology, Institute for Cell Engineering, Johns Hopkins University School of Medicine, Baltimore, MD.
References
- 1.Kelly RG. The second heart field. Curr Top Dev Biol. 2012;100:33–65. doi: 10.1016/B978-0-12-387786-4.00002-6. [DOI] [PubMed] [Google Scholar]
- 2.CDC. Congenital Heart Defects. 2013 http://www.cdc.gov/ncbddd/heartdefects/index.html.
- 3.Bruneau BG. The developmental genetics of congenital heart disease. Nature. 2008;451:943–948. doi: 10.1038/nature06801. [DOI] [PubMed] [Google Scholar]
- 4.Tzahor E, Evans SM. Pharyngeal mesoderm development during embryogenesis: implications for both heart and head myogenesis. Cardiovasc Res. 2011;91:196–202. doi: 10.1093/cvr/cvr116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Uosaki H, et al. Direct Contact with Endoderm-like Cells Efficiently Induces Cardiac Progenitors from Mouse and Human Pluripotent Stem Cells. PLoS One. 2012;7(10):e46413. doi: 10.1371/journal.pone.0046413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cheng P, et al. Fibronectin mediates mesendodermal cell fate decisions. Development. 2013;140:2587–2596. doi: 10.1242/dev.089052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kwon C, et al. A regulatory pathway involving Notch1/beta-catenin/Isl1 determines cardiac progenitor cell fate. Nature cell biology. 2009;11:951–957. doi: 10.1038/ncb1906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Shenje LT, et al. Precardiac deletion of Numb and Numblike reveals renewal of cardiac progenitors. Elife. 2014;3:e02164. doi: 10.7554/eLife.02164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Grevellec A, Tucker AS. The pharyngeal pouches and clefts: Development, evolution, structure and derivatives. Semin Cell Dev Biol. 2010;21:325–332. doi: 10.1016/j.semcdb.2010.01.022. [DOI] [PubMed] [Google Scholar]
- 10.Kwon C, Cordes KR, Srivastava D. Wnt/beta-catenin signaling acts at multiple developmental stages to promote mammalian cardiogenesis. Cell Cycle. 2008;7:3815–3818. doi: 10.4161/cc.7.24.7189. doi:10.4161/cc.7.24.7189 [pii] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cho GS, Fernandez L, Kwon C. Regenerative medicine for the heart: perspectives on stem-cell therapy. Antioxid Redox Signal. 2014;21:2018–2031. doi: 10.1089/ars.2014.6063. [DOI] [PMC free article] [PubMed] [Google Scholar]