

Abstract
Background/Objectives
High-fat diet (HFD)-induced obesity has significant negative effects on lymphatic function, but it remains unclear whether this is a direct effect of HFD or secondary to adipose tissue deposition.
Methods
We compared the effects of HFD on obesity-prone and obesity-resistant mice and analyzed lymphatic function in vivo and in vitro.
Results
Only obesity-prone mice had impaired lymphatic function, increased perilymphatic inflammation, and accumulation of lipid droplets surrounding their lymphatic endothelial cells (LECs). LECs isolated from obesity-prone mice, in contrast to obesity-resistant animals, had decreased expression of VEGFR-3 and Prox1. Exposure of LECs to a long-chain free fatty acid increased cellular apoptosis and decreased VEGFR-3 expression, while inhibition of intracellular inhibitors of VEGFR-3 signaling pathways increased cellular viability.
Conclusions
Collectively, our studies suggest that HFD-induced obesity decreases lymphatic function by increasing perilymphatic inflammation and altering LEC gene expression. Reversal of diminished VEGFR-3 signaling may rescue this phenotype and improve lymphatic function.
Keywords: Obesity, high-fat diet, lymphatic function, free fatty acid, lymphatic endothelial cell, inflammation
Introduction
The incidence of obesity is rapidly rising in Western countries; in the United States alone, it is estimated that 1 in 3 adults and 1 in 6 children are obese.(1) This is important because obesity is a significant risk factor for a variety of systemic disorders and adds nearly 200 billion dollars to our annual healthcare budget.(2) Therefore, understanding the mechanisms that regulate obesity-related pathology is an important goal.
Chronic adipose tissue inflammation is thought to contribute to a large number of pathologic responses in obesity, including metabolic syndrome and malignancy. These responses are modulated by T cells and macrophages, leading to increased local/systemic expression of inflammatory cytokines and release of free fatty acids (FFA) with direct toxic effects on surrounding tissues.(3–6) In addition, adipose tissue functions as an endocrine organ that produces substances that regulate function and obesity-related pathology.(5, 7)
Recent studies have shown that obesity has significant negative effects on the lymphatic system. For example, our group and others have shown that HFD-induced obesity markedly decreases interstitial fluid transport, immune cell trafficking, and lymphatic collecting vessel pumping.(8–10) These negative consequences were correlated with increasing body mass suggesting that adipose deposition plays a role in this response. In contrast, others have reported that high fructose diet-induced metabolic syndrome(11) or hypercholesterolemia,(12) decrease collecting vessel pumping and result in lymphatic system abnormalities independent of obesity. Thus, although it is clear that obesity can cause lymphatic dysfunction, the independent effects of dietary changes and weight gain on the lymphatic system remain unknown.
In this study, we sought to determine the independent effects of prolonged HFD intake with or without concomitant obesity on lymphatic function. By comparing the effects of HFD in obesity-prone and obesity-resistant mice, we show that dietary changes alone are insufficient to induce lymphatic dysfunction. In addition, we show that lymphatic dysfunction in obesity-prone mice correlates with perilymphatic accumulation of inflammatory cells and lipids, and alterations in LEC gene expression of lymphatic markers and inflammatory receptors. Finally, using correlative in vitro studies, we show that exposure of LECs to stearic acid, a long-chain fatty acid known to be increased in obese tissues,(13) results in decreased VEGFR-3 expression, apoptosis, and growth inhibition. Furthermore, intracellular modulation of VEGFR-3 signaling by inhibiting dephosphorylation of phosphatidylinositol (3,4,5)-trisphosphate-tyrosine-kinase (PIP3), addition of recombinant VEGF-C (direct activation of VEGFR-3), or recombinant insulin (indirect activation of PIP3 pathway) rescues LECs and markedly decreases apoptosis. Taken together, our findings suggest that obesity-induced inflammation, and not HFD, is responsible for the pathologic changes observed in the lymphovascular system and that these changes are due at least in part to direct injury to the LECs via byproducts of inflamed adipose tissues, including long-chain free fatty acids.
Materials and Methods
An extended version of Materials and Methods is available on the International Journal of Obesity website.
Animals, Diets, and Metabolic Analysis
Male C57BL/6J, BALB/cJ, and lean (Ln) allele of myostatin (MSTNln) mice were purchased from Jackson Laboratories (Bar Harbor, Maine) and maintained on a HFD (Purina Test Diet 60% kcal from fat; W.F Fisher & Son, Inc., NJ) for 10–12 weeks. Age-matched controls on the same genetic backgrounds were maintained on a normal chow diet (NCD, 13% kcal from fat; Purina PicoLab Rodent Diet 20) for the same period of time.
Analysis of Lymphatic Function
Lymphoscintigraphy was used to analyze lymph node uptake following peripheral injection of a technetium labeled colloid (99mTc) as previously reported.(14) We used a modification of previously reported methods to analyze dendritic cell (DCs) trafficking(15) and ferritin uptake.(16)
Histology
Tissues were analyzed with immunofluorescent staining as previously reported.(14)
LEC Isolation and PCR
Dermal LECs were isolated from the abdominal skin by digestion in 0.4% Collagenase IV digestion buffer (MP Biomedicals, Solon, OH) and flow cytometry to sort for LECs (CD45−, CD31+, podoplanin+).(17) RNA was extracted using TRIZOL (Invitrogen, Life Technologies, Carlsbad, CA) and converted to cDNA with TaqMan reverse transcriptase kit (Roche, Branchburg, NJ). Relative gene expression between groups was analyzed by normalizing gene expression with GAPDH.(18)
ELISA
Protein isolated from skin/subcutaneous tissues and serum was analyzed with ELISA to quantify CCL21, IL-1β, TNF-α (Sigma-Aldrich), VEGF-C (United States Biological) or FFA (Abcam) following the manufacturer’s protocol.
Cell Culture, Immunocytochemistry, and Western Blot
Human dermal LECs and adipose derived mesenchymal stem cells (ASCs) were obtained from PromoCell (Heidelberg, Germany) and stained in chamber slides for Prox1, Ki67 (Abcam, Cambridge, MA), p-AKT (Cell Signaling Technology, Danvers, MA), and VEGFR-3 (EMD Millipore, Billerica, MA). Cell signal intensity was quantified using MetaMorph (Molecular Devices, Sunnyvale, CA). Western blot analysis for VEGFR-3 (Abcam), p-AKT, and p-eNOS (Cell-signaling Technology) was performed with total cellular protein harvested from LECs and quantified with NIH Image J.(19)
Cellular Apoptosis and Viability assays
Apoptosis in response to stearic acid (SA; 0.1, 1, 10, 100μM) with/without 0.3nM Phosphatase and tensin homolog inhibitor (PTENi; SF1670, Sigma-Aldrich), (20) was quantified using caspase-3 assay (R and D systems, Minneapolis, MN) and Annexin V-FITC Apoptosis Detection Kit (eBioscience, San Diego, CA). Viability was measured using a WST-8 cytotoxicity cell assay kit (Sigma-Aldrich).
Statistical Analysis
Statistical analysis was performed using GraphPad Prism 6 software (La Jolla, CA) and differences between 2 groups were analyzed using a Student’s T test. Data are expressed as mean ± standard deviation with p<0.05 considered significant.
Results
High-fat diet (HFD) decreases lymphatic vascular density in obesity-prone mice
In order to study the effects of obesity and HFD on lymphatic function, obesity-prone C57BL/6J mice were compared to obesity-resistant BALB/cJ(21) and MSTNln mice.(22) As expected, feeding male C57BL/6J mice a HFD for 10 weeks resulted in obesity characterized by marked weight gain (164% increase), 9-fold increase in subcutaneous adipose deposition, mild glucose intolerance, and increased fasting serum insulin levels (Fig. 1A, C and Supplemental Fig. 1A, B). In contrast, BALB/cJ mice fed a HFD gained only a small though significant amount of weight (8%), with modest increases (2.86-fold) in subcutaneous adipose tissue thickness, mildly increased fasting serum insulin levels, and no signs of glucose intolerance. HFD-fed MSTNln mice remained lean throughout the study and displayed no abnormalities in glucose metabolism. All HFD-fed mice, regardless of strain, had modestly increased serum levels of total and HDL cholesterol and increased levels of FFA in serum; HFD-fed C57BL/6J and BALB/cJ mice also displayed increased serum LDL (Supplemental Fig. 1C, D). HFD-induced obesity in C57BL/6J mice correlated with a decrease in lymphatic (LYVE-1+) vessel density in the dermis and subcutaneous tissues as compared with their lean NCD-fed controls (Fig. 1B). In contrast, obesity-resistant HFD-fed BALB/cJ and MSTNln mice demonstrated no significant differences.
Fig. 1. HFD results in decreased lymphatic vascular density in obesity-prone mice.
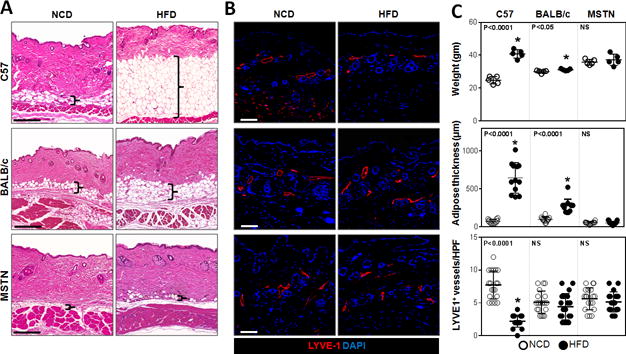
A. Representative H&E stained sections of back skin from NCD- or HFD-fed C57BL/6J, BALB/cJ, and MSTNln mice (Scale bar=200μm; bracket surrounds subcutaneous adipose tissues).
B. Representative immunofluorescent localization of LYVE-1+ vessels in upper limb tissues of NCD- or HFD-fed mice in all groups (Scale bar=100μm).
C. Upper panel: Body weights of mice on NCD (open circles) and HFD (filled circles) in all groups (n=5/group). C57BL/6J HFD vs. NCD (*p<0.0001). BALB/cJ HFD vs. NCD (*p<0.05), MSTNln mice had no significant difference. Middle panel: Quantification of subcutaneous soft tissue thickness in NCD- and HFD-fed mice in all groups (n=5–10/group). C57BL/6J HFD vs. NCD (*p<0.0001). MSTNln and BALB/cJ mice show no significant difference between NCD and HFD groups. Lower panel: Quantification of upper limb LYVE-1+ lymphatic vessel density per high powered field (HPF) and quadrant in NCD- and HFD-fed mice in all groups (n=5animals*4HPF/group). C57BL/6J HFD vs. NCD (*p<0.0001). MSTNln and BALB/cJ mice show no significant difference between NCD and HFD groups.
Obesity impairs lymphatic transport
The rate of uptake and peak transport of macromolecules by the lymphatic system can be quantified by injecting 99mTc labeled sulfur colloid in the distal hindlimb and analyzing its uptake by the inguinal lymph node.(14) Consistent with our previous studies, analysis of lymphatic transport in HFD-fed C57BL/6J revealed significant decreases in both total and rate of lymph node 99mTc uptake (Fig. 2A–C). In contrast, HFD-fed obesity-resistant mice had no significant decrease in peak or rate of 99mTc uptake as compared with their respective lean controls.
Fig. 2. Obesity impairs lymphatic transport of macromolecules to draining lymph nodes.
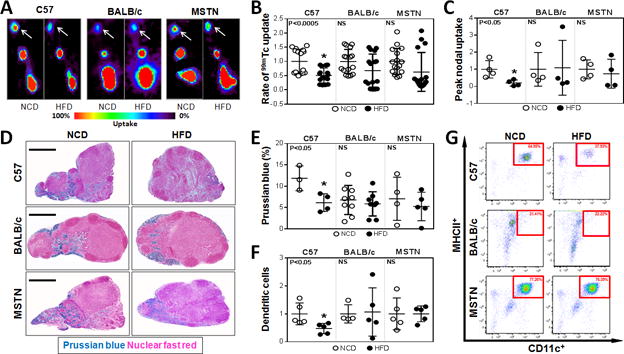
A. Representative hindlimb 99mTc heat maps for NCD- and HFD-fed C57BL/6J, BALB/cJ, and MSTNln mice. White arrows indicate the uptake in the inguinal lymph nodes (n=4/group).
B. Quantification of the rate of 99mTc uptake in inguinal lymph nodes following hindlimb injection. Data is presented as fold change relative to their NCD controls in each group (n=16/group). C57BL/6J HFD vs. NCD (*p<0.0005). MSTNln and BALB/cJ had no significant difference between NCD and HFD groups.
C. Quantification of peak nodal uptake of 99mTc in inguinal lymph nodes following hindlimb injection. Data are presented as fold change relative to their NCD controls in each group (n=4/group). C57BL/6J HFD vs. NCD (*p<0.05). MSTNln and BALB/cJ had no significant difference between NCD and HFD groups.
D. Representative Prussian blue stained histological cross sections of axillary lymph nodes harvested from NCD- or HFD-fed mice in all groups. (Scale bar=500μm).
E. Quantification of Prussian blue staining as a percentage of the total lymph node area in NCD- and HFD-fed mice (n=3–9/group). C57BL/6J HFD vs. NCD (*p<0.05). MSTNln and BALB/cJ had no significant difference between NCD and HFD groups.
F. Quantification of migrating CD45.1+ dendritic cells (MHCII+/CD11C+) in inguinal and popliteal lymph nodes of NCD- (open circles) and HFD- (filled circles) fed mice in all groups (n=5/group). Data are presented as fold change from respective NCD-fed controls. C57BL/6J HFD vs. NCD (*p<0.05). MSTNln and BALB/cJ had no significant difference between NCD and HFD groups.
G. Representative dot plot graphs of inguinal/popliteal lymph node flow cytometry analyzing migrating CD45.1+ dendritic cells in all groups. Red square surrounds dendritic cell population.
Peripherally injected ferritin is transported by, and remains bound to functional lymphatic vessels enabling histochemical localization using Prussian blue staining.(23) Consistent with our lymphoscintigraphic findings, only obese C57BL/6J animals had significantly decreased lymph node ferritin staining (2-fold) as compared to their NCD-fed controls, whereas HFD-fed BALB/cJ and MSTNln mice did not (Fig. 2D, E).
Functional lymphatic vessels also regulate migration of antigen presenting cells to regional lymph nodes where they interact with other immune cells.(24) Therefore, we analyzed migration of isolated DCs from the distal hindlimb (injection site) to the inguinal lymph node of HFD- and NCD-fed animals over a 24-hour period. Flow cytometry of draining lymph nodes harvested from HFD-fed obese C57BL/6J mice demonstrated a more than 50% reduction in the number of trafficked DCs as compared to their NCD-fed controls (Fig. 2F, G). In contrast, HFD-fed BALB/cJ and MSTNln mice demonstrated no changes when compared to their NCD-fed controls. Taken together, these findings show that obesity, rather than simply prolonged exposure to HFD, is necessary for impaired lymphatic function in mice.
Obesity results in perilymphatic inflammation
We next analyzed tissues harvested from multiple anatomical locations and co-localized lymphatic vessels (LYVE-1+) with T cells (CD3+) or macrophages (CD11b+) since previous studies have shown that these inflammatory cell types play a key role in the regulation of pathological changes in obesity.(3–6) This approach enabled us to quantify perilymphatic inflammation, which we defined as the number of inflammatory cells located within 50μm of a lymphatic vessel.
Analysis of ear tissues harvested from HFD-fed obese C57BL/6J mice demonstrated significant increases in the number of perilymphatic CD3+ and CD11b+ cells (3.6- and 3-fold increase from controls, respectively; Fig. 3A–B, Supplemental Fig. 2). Similar findings were noted in the trachea and back skin, suggesting that perilymphatic inflammation in obese C57BL/6J mice is a systemic phenomenon (Supplemental Fig. 3, 4). In contrast, HFD-fed BALB/cJ or MSTNln mice failed to show significant changes as compared with their NCD-fed controls, indicating that HFD alone is insufficient for perilymphatic inflammation.
Fig. 3. Obesity results in local tissue and perilymphatic inflammation.
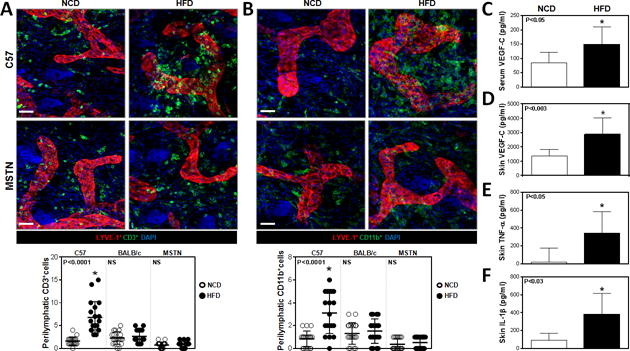
A. Representative immunofluorescence co-localization (upper panel) of CD3+ cells (green) and LYVE-1+ vessels (red) in ear skin whole mounts harvested from NCD- and HFD-fed C57BL/6J and MSTNln mice. Blue stain is DAPI (Scale bar=30μm). Quantification of perilymphatic CD3+ cells in skin ear cross-sections of NCD- (open circles) and HFD- (filled circles) fed mice in all groups (lower panel). C57BL/6J HFD vs. NCD (*p<0.0001, n=4 quadrants *4–5animals/group). MSTNln and BALB/cJ mice showed no significant difference between NCD and HFD groups.
B. Representative immunofluorescence co-localization (upper panel) of CD11b+ cells (green) and LYVE-1+ vessels (red) in ear skin whole mounts harvested from NCD- and HFD-fed C57BL/6J and MSTNln mice. Blue stain is DAPI (Scale bar=30μm). Quantification of perilymphatic CD11b+ cells in skin ear cross-sections of mice fed NCD (open circles) or HFD (filled circles) in all groups (lower panel). C57BL/6J HFD vs. NCD (*p<0.0001, n=4 quadrants * 5animals/group). MSTNln and BALB/cJ mice showed no significant difference between NCD and HFD groups.
C. Quantification of VEGF-C protein levels in serum harvested from lean and obese C57BL/6J mice (n=10/group, p<0.05).
D. Quantification of VEGF-C protein levels in back skin tissues harvested from lean and obese C57BL/6J mice (n=6–10/group, p<0.003).
E. Quantification of TNF-α protein levels in back skin tissues harvested from lean and obese C57BL/6J mice (n=4–6/group, p<0.05).
F. Quantification of IL-1β protein levels in back skin tissues harvested from lean and obese C57BL/6J mice (n=5/group, p<0.03).
Previous studies have shown that VEGF-C expression is increased in obese individuals and that this growth factor plays a key role in macrophage infiltration and differentiation.(25, 26) Consistent with these studies and our finding of increased macrophage infiltration in obese mice, we noted a marked increase in VEGF-C expression in both serum (1.75-fold) and skin (2-fold) of HFD-fed obese C57BL/6J mice, as compared to their NCD-fed controls (Fig. 3 C, D). In addition, analysis of skin tissues from obese HFD-fed C57BL/6J mice also demonstrated increased expression of inflammatory cytokines including TNF-α (20-fold) and IL-1β (8.32-fold) as compared to their NCD controls (Fig. 3 E, F).
Obesity downregulates expression of lymphatic markers in LECs
We next analyzed gene expression in LECs isolated from the subcutaneous tissues of obesity-prone and obesity-resistant mice fed either a HFD or NCD. PCR analysis of sorted LECs isolated from obese HFD-fed C57BL/6J mice demonstrated a nearly 2-fold decrease in the expression of Prox1 (a transcription factor necessary for lymphatic differentiation), VEGFR-3 (a signaling receptor necessary for LEC proliferation and function), and CCL21 (a chemotactic cytokine that regulates migration of DCs to lymphatic channels), as compared with their NCD-fed controls (Fig. 4A). Immunofluorescent co-localization of lymphatic vessels (LYVE-1+) with CCL21 confirmed our PCR findings demonstrating a marked reduction in CCL21 expression in lymphatic vessels of HFD-fed obese C57BL/6J mice as compared to their NCD-fed controls (Fig. 4B). Furthermore, LECs isolated from obese HFD-fed C57BL/6J mice demonstrated a more than 2-fold increase in the expression of the inflammatory cell receptor ICAM-1 and the pro-apoptotic gene Bax. In contrast, we noted no significant gene expression changes in cell sorted LECs isolated from HFD-fed BALB/cJ or MSTNln mice as compared with their respective NCD-fed controls.
Fig. 4. Obesity downregulates expression of lymphatic markers in LECs.
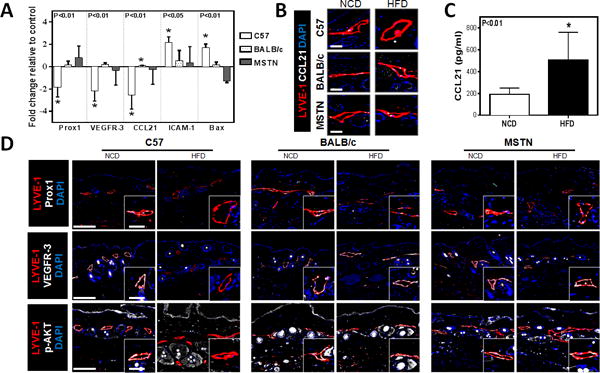
A. Quantification of mRNA expression in LECs sorted from NCD- and HFD-fed mice skin tissues for all groups, demonstrating relative expression of Prox1, VEGFR-3, CCL21, ICAM-1 and Bax (n=4–6/group). Data are presented as fold change relative to the NCD-fed controls in all groups. Note the downregulation of Prox1, VEGFR-3, and CCL21 (*p<0.01 for all) and the upregulation of ICAM-1 (*p<0.05) and Bax (*p<0.01) in obese C57BL/6J when compared to their lean controls.
B. Representative immunofluorescent localization of LYVE-1+(red) and CCL21+(white) in NCD- and HFD-fed mice in all groups (DAPI is blue; Scale bar=25μm).
C. Quantification of CCL21 protein levels in hindlimb tissues harvested from lean and obese C57BL/6J mice (n=6–8/group, p<0.01).
D. Representative low and high power (dashed box shown in lower right inset) immunofluorescent localization of LYVE-1+ cells (red) and Prox1 (top, white), VEGFR-3 (middle, white) and p–AKT (bottom, white). (DAPI nuclear stain is blue; Scale bars=50μm and 25μm.)
Previous studies have shown that gradients of CCL21 expression are necessary for migration of antigen presenting cells (e.g. dendritic cells) to lymphatic channels.(27, 28) In light of our observation that obese C57BL/6J mice had decreased DC trafficking and that their LECs had downregulated CCL21 expression, we next sought to determine if tissue gradients of CCL21 expression are altered in obesity by analyzing protein levels in the skin and subcutaneous tissues. Interestingly, we found that cutaneous tissues harvested from HFD-fed C57BL/6J mice had a more than 2-fold increase in CCL21 expression as compared to NCD-fed controls suggesting that clearance of inflammatory cells by the lymphatic system in obese animals is decreased, at least in part, due to a loss of gradients of CCL21 (i.e. low expression in LEC and high expression in tissues) (Fig. 4C).
To confirm our PCR findings, tissues were immunostained for LYVE-1 with VEGFR-3, Prox1, and p-AKT (a downstream mediator of VEGFR-3 signaling). Consistent with our PCR findings, we noted a marked decrease in VEGFR-3 and Prox1 expression by lymphatic vessels in HFD-fed obese C57BL/6J mice but not in HFD-fed BALB/cJ or MSTNln mice (Fig. 4D, Supplemental Fig. 5). In addition, only the lymphatic vessels in obese C57BL/6J mice had a marked decrease in expression of p-AKT as compared to their lean controls.
Obesity results in perilymphatic lipid accumulation and in vitro exposure of LECs to FFA decreases cellular viability
We subsequently sought to identify potential mechanisms that may regulate LEC injury and gene expression changes in obesity. Previous studies have shown obesity results in macrophage accumulation, which subsequently engulf necrotic adipocytes resulting in the formation of “crown-like structures” (CLS).(29, 30) Adipocyte breakdown by CLS releases toxic by-products such as long-chain FFA, leading to increased expression of inflammatory mediators and worsening subcutaneous tissue inflammation.(31, 32) Indeed, immunohistochemical analysis of skin harvested from obese HFD-fed C57BL/6J mice demonstrated the presence of CLS in the subcutaneous fat (Fig. 5A). Additionally, co-localization of lymphatic vessels (LYVE-1+) and lipids (BODIPY+), demonstrated the presence of numerous large lipid droplets in close proximity to LECs of obese HFD-fed C57BL/6J mice (Fig. 5B). In contrast, we found little lipid accumulation in NCD-fed C57BL/6J controls.
Fig. 5. Obesity results in accumulation of large lipid droplets around lymphatic vessels.
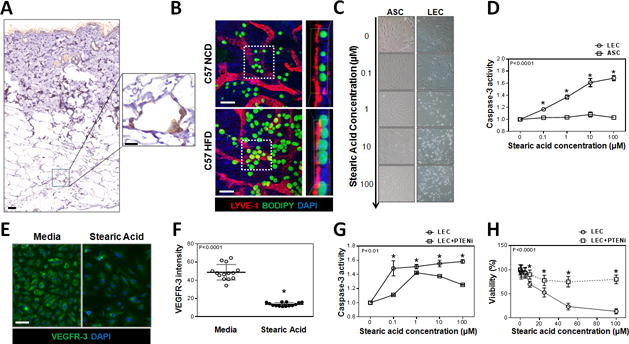
A. Representative low (left) and high power (right) immunohistological localization of CD45+ cells (brown) surrounding adipocytes (white) in back skin of obese C57BL/6J mice (Scale bars=100μm and 25μm, respectively).
B. Representative immunofluorescent localization (left) and orthoslice (right) from daughter region (white dotted square) of LYVE+ cells (red), BODIPY+ lipids (green) and nuclear DAPI stain (blue), in whole mount ear tissues of NCD- or HFD-fed C57BL/6J mice (Scale bar=20μm).
C. Representative bright field images showing LECs (top) and adipose derived mesenchymal stem cells (ASCs; bottom) cultured in media containing increasing concentrations of SA for 12hrs.
D. Quantification of caspase-3 activity in LECs (circles) and ASCs (squares) after treatment with SA for 12hrs (n=3–5/group, p<0.0001).
E. Representative immunofluorescent localization of VEGFR-3 expression (green) in LECs cultured with 10μM of SA for 12hrs (DAPI nuclear stain is shown in blue, Scale bar=25μm).
F. Quantification of VEGFR-3 expression intensity in LECs cultured in media (open circle) or media with 10μM SA (closed circle) for 12hrs (n=14/group, p<0.0001).
G. Quantification of caspase-3 activity in LECs cultured with media containing SA (circle) or SA with 0.3nM PTENi (square) for 12hrs (n=3/group, p<0.01).
H. Quantification of LEC viability treated with media containing SA or SA with 0.3nM PTENi for 12hrs (n=12/group, p<0.0001).
We next performed corroborative in vitro studies on isolated LECs in order to better understand the effects of lipid by-products and FFA at the cellular level. SA is known to be increased in obesity where it has been shown to have toxic effects on blood endothelial cells.(6, 33) We therefore compared the effects of SA on LECs with those on ASCs, since these cells are also present in adipose tissues. Interestingly, we found that SA, even in micromolar concentrations, was highly toxic to LECs resulting in a dose-dependent increase in cellular apoptosis as reflected by increasing activity of caspase-3 and annexin V (Fig. 5C, D, and Supplemental Fig. 6A). In contrast, ASCs were highly resistant to SA-induced apoptosis or injury. In addition, similar to our PCR findings, we noted that exposing LECs to even low doses of SA resulted in a significant decrease in VEGFR-3 expression (3-fold) as compared with LECs cultured in control media (Fig. 5E, F). This is important since a major function of VEGFR-3 signaling in LECs is prevention of apoptosis through activation of AKT and subsequent downregulation of apoptotic pathways.(34)
VEGFR-3, similar to the insulin receptor, is a tyrosine kinase receptor (TKR) and activates intracellular signaling via a variety of mechanisms. A major intracellular signaling pathway of TKRs is regulated by phosphorylation of Phosphatidylinositol 4,5-bisphosphate (PIP2) to PIP3, which then activates downstream signaling pathways, including AKT phosphorylation. PTEN is an intracellular protein that dephosphorylates PIP3 to PIP2, thereby functioning as an intracellular inhibitor of VEGFR-3 signaling. To determine if VEGFR-3 downregulation and subsequent decreased PIP3 activation in response to SA exposure plays a causal role in increased apoptosis, we cultured LECs with SA, either with or without media supplementation with PTENi. Analysis of caspase-3 activity and cellular viability in these cells demonstrated a statistically significant protective effect of PTEN inhibition in LECs as compared to cells cultured without PTENi (Fig. 5G, H). Similarly, we found that culturing isolated LECs with SA and either recombinant VEGF-C (direct activation of VEGFR3 downstream pathways) or recombinant insulin (indirect activation of PIP3 pathway via tyrosine kinase activation) restored the viability of LECs cultured with SA (Supplemental Figure 6B).
Our VEGFR-3 immunocytochemistry findings were confirmed with Western blot analysis of protein isolated from LECs cultured in media containing SA, with or without the addition of PTENi, VEGF-C, or insulin (Fig. 6A, B). Consistent with our immunocytochemistry findings, we noted that exposure of LECs to SA resulted in a 2.9-fold decrease in expression of VEGFR-3, 3.7-fold decrease in expression of p-AKT, and a 2.6-fold decrease in the expression of phosphorylated eNOS (p-eNOS, another downstream mediator of VEGFR-3). Addition of PTENi, recombinant VEGF-C, or insulin to LECs cultured in media containing SA normalized VEGFR-3, p-AKT, and p-eNOS expression to levels noted in LECs cultured in control (i.e. no SA) media. Addition of PTENi, recombinant VEGF-C, or insulin alone to LECs also significantly increased expression of these proteins independent of SA, suggesting that intracellular PTEN function acts as a feedback mechanism regulating VEGFR-3 signaling.
Fig. 6. Exposure of LECs to SA in vitro downregulates expression of VEGFR-3 and its downstream mediators.

A, B. Representative Western blot images (A) for VEGFR-3, p-AKT, p-eNOS and quantification (B) normalized to GAPDH expression of total cellular protein harvested from LECs treated with 1μM SA, 10μM SA+ 0.3nM PTENi, PTENi (0.3nM), SA+ 100ng/mL VEGF-C, VEGF-C (100ng/mL), SA+ 100nM insulin or insulin (100nM) (n=4–5/group, p<0.05 for all antibodies).
C. Left panels: Representative immunofluorescent co-localization of VEGFR-3 (green) and Prox1, Ki67, and p-AKT (all shown in pink) in LECs cultured in control media, media containing 10μM SA, media containing 10μM SA with PTENi (0.3nM), 10μM SA with VEGF-C (100ng/mL) or 10μM SA with insulin (100nM) for 12hrs. (Scale bar=25μm). Right panels: Quantification of expression of Prox1, Ki67, and p-AKT in groups shown in the left panels (n=10–15/group, p<0.0005).
Consistent with our cell-sorted PCR on LECs obtained from obese C57BL/6J mice, we noted a marked reduction in the expression of Prox1 by LECs after exposure to SA (Fig. 6C). This was normalized to control levels with the addition of PTENi, VEGF-C or insulin. We noted similar findings with Ki67 (a marker of cellular proliferation) and p-AKT expression. Taken together, our in vitro findings suggest that long-chain FFA are harmful to LECs by decreasing the expression of master lymphatic regulators and that these effects can be mitigated by modulating intracellular pathways downstream of VEGFR-3 through PIP3 pathway and subsequently AKT activation.
Discussion
Previous clinical and laboratory studies have shown that obesity is a negative regulator of lymphatic function.(3–10) However, it remains unclear if the observed effects on lymphatic function were secondary to weight gain, prolonged exposure to HFD, or toxic metabolic by-products of adipose tissue. In order to address this question, we compared lymphatic function in obesity-prone and obesity-resistant mice after prolonged exposure to HFD. Our results are consistent with previous reports demonstrating that C57BL/6J mice become morbidly obese on HFD feeds,(8, 9) while BALB/cJ and MSTNln mice do not(21, 22) even though serum FFA levels were significantly increased in all HFD-fed mice.(13, 35) This approach is clinically relevant since the incidence and severity of obesity in patients is also highly variable and modulated by genetic factors. Our findings imply that obesity, but not HFD, is necessary for the development of lymphatic dysfunction since obesity-resistant mice maintained essentially normal lymphatic function despite prolonged exposure to HFD feeds. Our study also suggests that obesity-induced tissue changes or alterations in glucose and/or FA metabolism (or a combination of all) are necessary for lymphatic dysfunction and that prolonged exposure to HFD alone does not cause direct injury to the lymphatic system.
Alternate hypotheses could explain the differences we noted in lymphatic function of obesity-prone and obesity-resistant mice after prolonged HFD feeds. For example, it is possible that strain-related differences may contribute to the disparities we observed in lymphatic function. Although our study does not definitely disprove this possibility, we believe that genetic differences in baseline lymphatic function are less important than the effects of obesity since MSTNln mice are derived from a B6 background, and because both obesity-resistant mice strains (MSTNln and BALB/cJ) were protected from lymphatic injury despite dissimilar genetic backgrounds. Additionally, although previous studies reported lymphatic dysfunction in ApoE deficient mice with very high serum cholesterol levels (1000–2000mg/mL),(36) it is unlikely that changes in cholesterol levels are responsible for the lymphatic dysfunction we observed in our obese mice since the total cholesterol levels in our study were an order of a magnitude lower (<200mg/mL) than those reported by Angeli et al. In addition, total and LDL cholesterol levels were elevated in all HFD-fed mice (i.e. both obesity-prone and obesity-resistant mice). In fact, BALB/cJ mice had the highest serum levels of LDL cholesterol. Nonetheless, despite increases in total and LDL cholesterol in all HFD-fed animals, only the obese C57BL/6J mice displayed decreased lymphatic function indicating that modestly increased levels of cholesterol do not have deleterious effects on the lymphatic system.
A key histological difference between obesity-prone and obesity-resistant mice in our study was perilymphatic inflammation. This finding extends the results of previous studies demonstrating chronic, low-grade perivascular inflammatory responses in obesity.(37, 38) This is important because perivascular inflammation plays a key role in the pathological outcomes of obesity on the blood vascular system by diverse mechanisms including altered expression of adipokines, impaired nitric oxide signaling, and increased production of reactive oxygen species.(7, 39, 40) These effects are compounded by infiltration of macrophages that phagocytose necrotic adipocytes, leading to the release of long-chain FFA(41) that, in turn, have deleterious effects on endothelial cells.(6, 31, 42) Indeed, in our studies we observed the presence of numerous CLS, and close association of large lipid droplets with lymphatic vessels, in the subcutaneous tissues of obese mice. Thus, chronic perilymphatic inflammatory responses in obese animals may act via a variety of mechanisms to inhibit lymphatic function in this setting.
We analyzed gene expression changes in cell sorted LECs from NCD and HFD mice, as well as isolated LECs cultured with SA, a long-chain fatty acid known to be increased in obesity.(41) This analysis demonstrated that obese animals (but not obesity-resistant mice fed a HFD) and LECs cultured with SA had markedly decreased expression of Prox1 and VEGFR-3. Consistent with this effect, we noted that LECs harvested from obese C57BL/6J mice had a marked increase in the expression of the pro-apoptotic gene Bax and decreased expression of p-AKT, the downstream mediator of VEGFR-3 activation. These findings are important as previous studies have shown that the transcription factor Prox1 is a master lymphatic regulator necessary for the maintenance of the lymphatic phenotype and VEGFR-3 expression.(43) Similarly, VEGFR-3 signaling plays a crucial role in a variety of physiologic functions in LECs, including proliferation, migration, differentiation, protection from apoptosis, and expression of nitric oxide.(34, 44) To further explore the role of VEGFR-3 and TKR signaling in LEC response to injury, we cultured LECs with SA with or without an inhibitor of PTEN, VEGF-C, or insulin. This analysis demonstrated that direct activation of VEGFR-3 signaling pathways with VEGF-C or indirect activation of tyrosine kinase signaling with a PTEN inhibitor or insulin confers significant protective effects to LECs cultured with SA. Thus, downregulation of Prox1 and VEGFR-3 in obesity may serve as a key cellular mechanism regulating lymphatic dysfunction in this setting and represents a potential target for therapeutic interventions. Our findings also suggest that obesity or exposure to high levels of long-chain fatty acids decrease insulin sensitivity in lymphatic endothelial cells and that subsequent intracellular downregulation of tyrosine kinase activity (e.g. AKT phosphorylation, eNOS, etc.) decreases LEC viability and proliferation.
A large number of studies have shown that inflammation increases lymphangiogenesis.(45, 46) Thus, it may seem contradictory that we found decreased lymphatic vessel density in obese animals in which we also noted marked subcutaneous infiltration of T cells and macrophages. However, numerous other studies have shown that chronic inflammation in some physiologic settings can markedly inhibit lymphangiogenesis(47, 48) and that the initial lymphangiogenic response to acute inflammation subsides over time.(17, 47, 48) Furthermore, these studies have shown that an important mechanism regulating this effect is decreased VEGFR-3 expression. For example, Huggenberger et al. showed that chronic inflammation can decrease VEGFR-3 protein and gene expression during oxazolone challenge, despite concomitantly increased expression of pro-inflammatory cytokines TNF-α, IL-1β, and IFN-γ.(49) Others have reported that micro-RNAs regulate inflammation-induced decreased VEGFR-3 and Prox1 expression.(50, 51) Finally, T cell-derived cytokines including IFN-γ, IL-4 and IL-13 have been shown to directly inhibit lymphangiogenesis.(46, 52)
Consistent with previous studies in obese mice and patients,(26) we noted a marked increase in VEGF-C expression in tissues and serum harvested from obese HFD-fed C57BL/6J mice. This effect plays a key role on glucose homeostasis in obesity by regulating macrophage infiltration and differentiation.(25) In addition, the combination of increased VEGF-C in the context of decreased LEC VEGFR-3 expression in obese mice suggest that homeostatic mechanisms contribute to changes in VEGF-C expression. This scenario may be similar to insulin resistance in obesity (i.e. increased insulin/decreased insulin receptor function), suggesting that similar mechanisms regulate lymphatic function, a condition we have termed VEGF-C resistance. Our hypothesis is supported by the fact that receptor downregulation/dysfunction is a common pathologic outcome in obesity, regulating sensitivity to a number of molecules including insulin and leptin.(53, 54) In addition, VEGFR-3, similar to the insulin receptor, is a TKR,(34) and its expression in obesity may therefore be down-regulated by similar mechanisms, including inflammatory cytokines(55) and free fatty acids.(56) Finally, similar to our findings in LECs in vitro, previous studies have shown that deletion of PTEN is protective against insulin resistance by increasing intracellular PI3K/AKT signaling.(57)
An important function of the lymphatic system is the transport of antigen presenting cells to regional lymph nodes. In the current study, we found that obesity markedly decreases DC migration to regional lymph nodes and that this response may be related to decreased clearance of interstitial fluid as well as changes in gradients of CCL21 expression.(27, 58) These findings are in-line with previous studies demonstrating decreased CCL21 gradients and increased LEC ICAM-1 expression in response to contact hypersensitivity.(17) Our findings are also consistent with the known roles of interstitial flow and VEGFR-3 expression in regulating CCL21 expression by LECs.(59) Thus, changes in CCL21 expression gradients and increased expression of ICAM by LECs may effectively trap antigen presenting cells in peripheral tissues and prevent trafficking of activating or suppressive DCs to regional lymph nodes. These data suggest a new cellular mechanism that regulates impaired T cell recall responses and may contribute to increased incidence of autoimmunity in obesity.(60)
In summary, we have shown that obesity, rather than prolonged exposure to HFD is necessary for induction of lymphatic dysfunction in mice and that this response correlates with perilymphatic inflammation and down-regulation of LEC VEGFR-3 and Prox1 expression. In addition, we have shown that LECs are highly sensitive to the toxic effects of long-chain fatty acids that are known to be increased in obesity and that direct or indirect intracellular activation of PIP3 confers significant protection to LECs.
Supplementary Material
Acknowledgments
The authors thank Mesruh Turkekul, Navid Paknejad, Sho Fujisawa, and Yevgeniy Romin of the Molecular Cytology Core at Memorial Sloan Kettering Cancer Center for assistance with both histology and tissue imaging (Core Grant (P30 CA008748)).
Source of Funding:
NIH R01 HL111130-01 awarded to BJM
NIH T32 CA009501-27 grant to DAC
NIH T32 CA009685-21A1 grant to GGN
NIH/NCI Cancer Center Support Grant P30 CA008748
Plastic Surgery Foundation Pilot Research Grant awarded to JCG
Plastic Surgery Foundation Pilot Research Grant awarded to GGN
Footnotes
Competing Financial Interests:
The authors declare no competing financial interests.
Supplementary information is available on the International Journal of Obesity’s website.
References
- 1.Ogden CL, Carroll MD, Kit BK, Flegal KM. Prevalence of childhood and adult obesity in the United States, 2011–2012. Jama. 2014;311(8):806–14. doi: 10.1001/jama.2014.732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cawley J, Meyerhoefer C. The medical care costs of obesity: an instrumental variables approach. Journal of health economics. 2012;31(1):219–30. doi: 10.1016/j.jhealeco.2011.10.003. [DOI] [PubMed] [Google Scholar]
- 3.Ross R. Atherosclerosis–an inflammatory disease. The New England journal of medicine. 1999;340(2):115–26. doi: 10.1056/NEJM199901143400207. [DOI] [PubMed] [Google Scholar]
- 4.Lumeng CN, Bodzin JL, Saltiel AR. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. The Journal of clinical investigation. 2007;117(1):175–84. doi: 10.1172/JCI29881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hajer GR, van Haeften TW, Visseren FL. Adipose tissue dysfunction in obesity, diabetes, and vascular diseases. European heart journal. 2008;29(24):2959–71. doi: 10.1093/eurheartj/ehn387. [DOI] [PubMed] [Google Scholar]
- 6.Harvey KA, Walker CL, Pavlina TM, Xu Z, Zaloga GP, Siddiqui RA. Long-chain saturated fatty acids induce pro-inflammatory responses and impact endothelial cell growth. Clinical nutrition. 2010;29(4):492–500. doi: 10.1016/j.clnu.2009.10.008. [DOI] [PubMed] [Google Scholar]
- 7.Cao H. Adipocytokines in obesity and metabolic disease. The Journal of endocrinology. 2014;220(2):T47–59. doi: 10.1530/JOE-13-0339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Weitman ES, Aschen SZ, Farias-Eisner G, Albano N, Cuzzone DA, Ghanta S, et al. Obesity impairs lymphatic fluid transport and dendritic cell migration to lymph nodes. PloS one. 2013;8(8):e70703. doi: 10.1371/journal.pone.0070703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Savetsky IL, Torrisi JS, Cuzzone DA, Ghanta S, Albano NJ, Gardenier JC, et al. Obesity increases inflammation and impairs lymphatic function in a mouse model of lymphedema. American journal of physiology Heart and circulatory physiology. 2014;307(2):H165–72. doi: 10.1152/ajpheart.00244.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Savetsky IL, Albano NJ, Cuzzone DA, Gardenier JC, Torrisi JS, Garcia Nores GD, et al. Lymphatic Function Regulates Contact Hypersensitivity Dermatitis in Obesity. The Journal of investigative dermatology. 2015 doi: 10.1038/jid.2015.283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zawieja SD, Wang W, Wu X, Nepiyushchikh ZV, Zawieja DC, Muthuchamy M. Impairments in the intrinsic contractility of mesenteric collecting lymphatics in a rat model of metabolic syndrome. American journal of physiology Heart and circulatory physiology. 2012;302(3):H643–53. doi: 10.1152/ajpheart.00606.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lim RS, Kratzer A, Barry NP, Miyazaki-Anzai S, Miyazaki M, Mantulin WW, et al. Multimodal CARS microscopy determination of the impact of diet on macrophage infiltration and lipid accumulation on plaque formation in ApoE-deficient mice. Journal of lipid research. 2010;51(7):1729–37. doi: 10.1194/jlr.M003616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Li M, Fu W, Li XA. Differential fatty acid profile in adipose and non-adipose tissues in obese mice. International journal of clinical and experimental medicine. 2010;3(4):303–7. [PMC free article] [PubMed] [Google Scholar]
- 14.Avraham T, Daluvoy S, Zampell J, Yan A, Haviv YS, Rockson SG, et al. Blockade of transforming growth factor-beta1 accelerates lymphatic regeneration during wound repair. The American journal of pathology. 2010;177(6):3202–14. doi: 10.2353/ajpath.2010.100594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Jolly K, Bradley F, Sharp S, Smith H, Mant D. Follow-up care in general practice of patients with myocardial infarction or angina pectoris: initial results of the SHIP trial. Southampton Heart Integrated Care Project. Family practice. 1998;15(6):548–55. doi: 10.1093/fampra/15.6.548. [DOI] [PubMed] [Google Scholar]
- 16.Aschen SZ, Farias-Eisner G, Cuzzone DA, Albano NJ, Ghanta S, Weitman ES, et al. Lymph node transplantation results in spontaneous lymphatic reconnection and restoration of lymphatic flow. Plastic and reconstructive surgery. 2014;133(2):301–10. doi: 10.1097/01.prs.0000436840.69752.7e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Vigl B, Aebischer D, Nitschke M, Iolyeva M, Rothlin T, Antsiferova O, et al. Tissue inflammation modulates gene expression of lymphatic endothelial cells and dendritic cell migration in a stimulus-dependent manner. Blood. 2011;118(1):205–15. doi: 10.1182/blood-2010-12-326447. [DOI] [PubMed] [Google Scholar]
- 18.Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative CT method. Nat Protocols. 2008;3(6):1101–8. doi: 10.1038/nprot.2008.73. [DOI] [PubMed] [Google Scholar]
- 19.Zampell JC, Yan A, Elhadad S, Avraham T, Weitman E, Mehrara BJ. CD4(+) cells regulate fibrosis and lymphangiogenesis in response to lymphatic fluid stasis. PloS one. 2012;7(11):e49940. doi: 10.1371/journal.pone.0049940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Li Y, Prasad A, Jia Y, Roy SG, Loison F, Mondal S, et al. Pretreatment with phosphatase and tensin homolog deleted on chromosome 10 (PTEN) inhibitor SF1670 augments the efficacy of granulocyte transfusion in a clinically relevant mouse model. Blood. 2011;117(24):6702–13. doi: 10.1182/blood-2010-09-309864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Montgomery MK, Hallahan NL, Brown SH, Liu M, Mitchell TW, Cooney GJ, et al. Mouse strain-dependent variation in obesity and glucose homeostasis in response to high-fat feeding. Diabetologia. 2013;56(5):1129–39. doi: 10.1007/s00125-013-2846-8. [DOI] [PubMed] [Google Scholar]
- 22.McPherron AC, Lee SJ. Suppression of body fat accumulation in myostatin-deficient mice. The Journal of clinical investigation. 2002;109(5):595–601. doi: 10.1172/JCI13562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Isaka N, Padera TP, Hagendoorn J, Fukumura D, Jain RK. Peritumor lymphatics induced by vascular endothelial growth factor-C exhibit abnormal function. Cancer research. 2004;64(13):4400–4. doi: 10.1158/0008-5472.CAN-04-0752. [DOI] [PubMed] [Google Scholar]
- 24.Randolph GJ, Angeli V, Swartz MA. Dendritic-cell trafficking to lymph nodes through lymphatic vessels. Nature reviews Immunology. 2005;5(8):617–28. doi: 10.1038/nri1670. [DOI] [PubMed] [Google Scholar]
- 25.Karaman S, Hollmen M, Robciuc MR, Alitalo A, Nurmi H, Morf B, et al. Blockade of VEGF-C and VEGF-D modulates adipose tissue inflammation and improves metabolic parameters under high-fat diet. Molecular metabolism. 2015;4(2):93–105. doi: 10.1016/j.molmet.2014.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gomez-Ambrosi J, Catalan V, Rodriguez A, Ramirez B, Silva C, Gil MJ, et al. Involvement of serum vascular endothelial growth factor family members in the development of obesity in mice and humans. The Journal of nutritional biochemistry. 2010;21(8):774–80. doi: 10.1016/j.jnutbio.2009.05.004. [DOI] [PubMed] [Google Scholar]
- 27.MartIn-Fontecha A, Sebastiani S, Hopken UE, Uguccioni M, Lipp M, Lanzavecchia A, et al. Regulation of dendritic cell migration to the draining lymph node: impact on T lymphocyte traffic and priming. The Journal of experimental medicine. 2003;198(4):615–21. doi: 10.1084/jem.20030448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Saeki H, Moore AM, Brown MJ, Hwang ST. Cutting edge: secondary lymphoid-tissue chemokine (SLC) and CC chemokine receptor 7 (CCR7) participate in the emigration pathway of mature dendritic cells from the skin to regional lymph nodes. Journal of immunology. 1999;162(5):2472–5. [PubMed] [Google Scholar]
- 29.Murano I, Barbatelli G, Parisani V, Latini C, Muzzonigro G, Castellucci M, et al. Dead adipocytes, detected as crown-like structures, are prevalent in visceral fat depots of genetically obese mice. Journal of lipid research. 2008;49(7):1562–8. doi: 10.1194/jlr.M800019-JLR200. [DOI] [PubMed] [Google Scholar]
- 30.Cinti S, Mitchell G, Barbatelli G, Murano I, Ceresi E, Faloia E, et al. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. Journal of lipid research. 2005;46(11):2347–55. doi: 10.1194/jlr.M500294-JLR200. [DOI] [PubMed] [Google Scholar]
- 31.Suganami T, Nishida J, Ogawa Y. A paracrine loop between adipocytes and macrophages aggravates inflammatory changes: role of free fatty acids and tumor necrosis factor alpha. Arteriosclerosis, thrombosis, and vascular biology. 2005;25(10):2062–8. doi: 10.1161/01.ATV.0000183883.72263.13. [DOI] [PubMed] [Google Scholar]
- 32.Suganami T, Tanimoto-Koyama K, Nishida J, Itoh M, Yuan X, Mizuarai S, et al. Role of the Toll-like receptor 4/NF-kappaB pathway in saturated fatty acid-induced inflammatory changes in the interaction between adipocytes and macrophages. Arteriosclerosis, thrombosis, and vascular biology. 2007;27(1):84–91. doi: 10.1161/01.ATV.0000251608.09329.9a. [DOI] [PubMed] [Google Scholar]
- 33.Opie LH, Walfish PG. Plasma free fatty acid concentrations in obesity. The New England journal of medicine. 1963;268:757–60. doi: 10.1056/NEJM196304042681404. [DOI] [PubMed] [Google Scholar]
- 34.Coso S, Bovay E, Petrova TV. Pressing the right buttons: signaling in lymphangiogenesis. Blood. 2014;123(17):2614–24. doi: 10.1182/blood-2013-12-297317. [DOI] [PubMed] [Google Scholar]
- 35.Machado MV, Michelotti GA, Xie G, Almeida Pereira T, Boursier J, Bohnic B, et al. Mouse models of diet-induced nonalcoholic steatohepatitis reproduce the heterogeneity of the human disease. PloS one. 2015;10(5):e0127991. doi: 10.1371/journal.pone.0127991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lim HY, Rutkowski JM, Helft J, Reddy ST, Swartz MA, Randolph GJ, et al. Hypercholesterolemic mice exhibit lymphatic vessel dysfunction and degeneration. The American journal of pathology. 2009;175(3):1328–37. doi: 10.2353/ajpath.2009.080963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nieman DC, Henson DA, Nehlsen-Cannarella SL, Ekkens M, Utter AC, Butterworth DE, et al. Influence of obesity on immune function. Journal of the American Dietetic Association. 1999;99(3):294–9. doi: 10.1016/S0002-8223(99)00077-2. [DOI] [PubMed] [Google Scholar]
- 38.Womack J, Tien PC, Feldman J, Shin JH, Fennie K, Anastos K, et al. Obesity and immune cell counts in women. Metabolism: clinical and experimental. 2007;56(7):998–1004. doi: 10.1016/j.metabol.2007.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Greenstein AS, Khavandi K, Withers SB, Sonoyama K, Clancy O, Jeziorska M, et al. Local inflammation and hypoxia abolish the protective anticontractile properties of perivascular fat in obese patients. Circulation. 2009;119(12):1661–70. doi: 10.1161/CIRCULATIONAHA.108.821181. [DOI] [PubMed] [Google Scholar]
- 40.Ketonen J, Shi J, Martonen E, Mervaala E. Periadventitial adipose tissue promotes endothelial dysfunction via oxidative stress in diet-induced obese C57Bl/6 mice. Circulation journal: official journal of the Japanese Circulation Society. 2010;74(7):1479–87. doi: 10.1253/circj.cj-09-0661. [DOI] [PubMed] [Google Scholar]
- 41.Neels JG, Olefsky JM. Inflamed fat: what starts the fire? The Journal of clinical investigation. 2006;116(1):33–5. doi: 10.1172/JCI27280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kosteli A, Sugaru E, Haemmerle G, Martin JF, Lei J, Zechner R, et al. Weight loss and lipolysis promote a dynamic immune response in murine adipose tissue. The Journal of clinical investigation. 2010;120(10):3466–79. doi: 10.1172/JCI42845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wigle JT, Harvey N, Detmar M, Lagutina I, Grosveld G, Gunn MD, et al. An essential role for Prox1 in the induction of the lymphatic endothelial cell phenotype. The EMBO journal. 2002;21(7):1505–13. doi: 10.1093/emboj/21.7.1505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Dumont DJ, Jussila L, Taipale J, Lymboussaki A, Mustonen T, Pajusola K, et al. Cardiovascular failure in mouse embryos deficient in VEGF receptor-3. Science. 1998;282(5390):946–9. doi: 10.1126/science.282.5390.946. [DOI] [PubMed] [Google Scholar]
- 45.Kataru RP, Jung K, Jang C, Yang H, Schwendener RA, Baik JE, et al. Critical role of CD11b+ macrophages and VEGF in inflammatory lymphangiogenesis, antigen clearance, and inflammation resolution. Blood. 2009;113(22):5650–9. doi: 10.1182/blood-2008-09-176776. [DOI] [PubMed] [Google Scholar]
- 46.Savetsky IL, Ghanta S, Gardenier JC, Torrisi JS, Garcia Nores GD, Hespe GE, et al. Th2 cytokines inhibit lymphangiogenesis. PloS one. 2015;10(6):e0126908. doi: 10.1371/journal.pone.0126908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Baluk P, Tammela T, Ator E, Lyubynska N, Achen MG, Hicklin DJ, et al. Pathogenesis of persistent lymphatic vessel hyperplasia in chronic airway inflammation. The Journal of clinical investigation. 2005;115(2):247–57. doi: 10.1172/JCI22037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Johnson LA, Prevo R, Clasper S, Jackson DG. Inflammation-induced uptake and degradation of the lymphatic endothelial hyaluronan receptor LYVE-1. The Journal of biological chemistry. 2007;282(46):33671–80. doi: 10.1074/jbc.M702889200. [DOI] [PubMed] [Google Scholar]
- 49.Huggenberger R, Siddiqui SS, Brander D, Ullmann S, Zimmermann K, Antsiferova M, et al. An important role of lymphatic vessel activation in limiting acute inflammation. Blood. 2011;117(17):4667–78. doi: 10.1182/blood-2010-10-316356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Jones D, Li Y, He Y, Xu Z, Chen H, Min W. Mirtron microRNA-1236 inhibits VEGFR-3 signaling during inflammatory lymphangiogenesis. Arteriosclerosis, thrombosis, and vascular biology. 2012;32(3):633–42. doi: 10.1161/ATVBAHA.111.243576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kazenwadel J, Michael MZ, Harvey NL. Prox1 expression is negatively regulated by miR-181 in endothelial cells. Blood. 2010;116(13):2395–401. doi: 10.1182/blood-2009-12-256297. [DOI] [PubMed] [Google Scholar]
- 52.Kataru RP, Kim H, Jang C, Choi DK, Koh BI, Kim M, et al. T lymphocytes negatively regulate lymph node lymphatic vessel formation. Immunity. 2011;34(1):96–107. doi: 10.1016/j.immuni.2010.12.016. [DOI] [PubMed] [Google Scholar]
- 53.Zhou Y, Rui L. Leptin signaling and leptin resistance. Frontiers of medicine. 2013;7(2):207–22. doi: 10.1007/s11684-013-0263-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lee BC, Lee J. Cellular and molecular players in adipose tissue inflammation in the development of obesity-induced insulin resistance. Biochimica et biophysica acta. 2014;1842(3):446–62. doi: 10.1016/j.bbadis.2013.05.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Su D, Coudriet GM, Hyun Kim D, Lu Y, Perdomo G, Qu S, et al. FoxO1 links insulin resistance to proinflammatory cytokine IL-1beta production in macrophages. Diabetes. 2009;58(11):2624–33. doi: 10.2337/db09-0232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hotamisligil GS, Arner P, Caro JF, Atkinson RL, Spiegelman BM. Increased Adipose-Tissue Expression of Tumor-Necrosis-Factor-Alpha in Human Obesity and Insulin-Resistance. Journal of Clinical Investigation. 1995;95(5):2409–15. doi: 10.1172/JCI117936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Wijesekara N, Konrad D, Eweida M, Jefferies C, Liadis N, Giacca A, et al. Muscle-specific Pten deletion protects against insulin resistance and diabetes. Molecular and cellular biology. 2005;25(3):1135–45. doi: 10.1128/MCB.25.3.1135-1145.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ohl L, Mohaupt M, Czeloth N, Hintzen G, Kiafard Z, Zwirner J, et al. CCR7 governs skin dendritic cell migration under inflammatory and steady-state conditions. Immunity. 2004;21(2):279–88. doi: 10.1016/j.immuni.2004.06.014. [DOI] [PubMed] [Google Scholar]
- 59.Iwami D, Brinkman CC, Bromberg JS. Vascular endothelial growth factor c/vascular endothelial growth factor receptor 3 signaling regulates chemokine gradients and lymphocyte migration from tissues to lymphatics. Transplantation. 2015;99(4):668–77. doi: 10.1097/TP.0000000000000561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Aritomi K, Kuwabara T, Tanaka Y, Nakano H, Yasuda T, Ishikawa F, et al. Altered antibody production and helper T cell function in mice lacking chemokines CCL19 and CCL21-Ser. Microbiology and immunology. 2010;54(11):691–701. doi: 10.1111/j.1348-0421.2010.00266.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.