

Abstract
Aerobic exercise (e.g., wheel running (WR) extensively used in animal research) positively impacts many measures of neuroplastic potential in the brain, such as rates of adult neurogenesis, angiogenesis, and expression of neurotrophic factors in rodents. This intervention has also been shown to mitigate behavioral and neuroanatomical aspects of the negative impacts of teratogens (i.e., developmental exposure to alcohol) and age-related neurodegeneration in rodents. Environmental complexity (EC) has been shown to produce numerous neuroplastic benefits in cortical and subcortical structures and can be coupled with wheel running to increase the proliferation and survival of new cells in the adult hippocampus. The combination of these two interventions provides a robust "superintervention" (WR-EC) that can be implemented in a range of rodent models of neurological disorders. We will discuss the implementation of WR/EC and its constituent interventions for use as a more powerful therapeutic intervention in rats using the animal model of prenatal exposure to alcohol in humans. We will also discuss which elements of the procedures are absolutely necessary for the interventions and which ones may be altered depending on the experimenter's question or facilities.
Keywords: Neuroplasticity, rats, exercise, neurogenesis, alcohol, development, novelty
Introduction
Rearing in different environments has long been known to cause changes in various measures of neurological wellness. Many studies look at the beneficial effects of rearing in a complex environment (EC) starting with groundbreaking research by Diamond and Rosenzweig e.g., 1,2 and Greenough e.g., 3,4. EC has been demonstrated to have an undeniable positive effects on synaptic and cellular changes in the brain 5–7. EC can affect a multiplicity of brain regions including the hippocampus 8,9 and visual cortex 10,11, ventral striatum 12,13, as well as brain-wide neuroimmune function reviewed in 14. Particular interest has developed from the studies on hippocampus when it was demonstrated that EC can increase the survival rate of adult-born granule cells of the dentate gyrus through dendritic plasticity 9,13. This last point has gathered much interest due to the growing body of literature indicating that cardiovascular exercise promotes adult neurogenesis in both the healthy and damaged brain 15–18. Wheel running (WR) is an easy to implement form of voluntary cardiovascular activity that has been shown to be beneficial in rodent models of neurological disorders or aging 17,19,20. WR affects the expression of growth factors in both the central and peripheral nervous system 21–23.
Combining (subsequently) WR and EC into a "superintervention" (WR-EC) (i.e., 12 days of WR followed by 30 days in EC) provides a robust increase in hippocampal adult neurogenesis and increased survival of the newly proliferated cells 8, the effect that in the animal model of FASD is not achieved by individual components (see below). Since both components of WR-EC affect a diverse array of structures within the brain 13 (WR reviewed in 22, EC reviewed in 24), implementation of this intervention can easily be applied to rodent models of both developmental and later life onset models of neurological impairment (e.g., neonatal alcohol exposure, aging, early life stress).
Integration of WR-EC in the adolescent and early adult periods (i.e., postnatal days 30 - 72) can ameliorate some of the negative effects of a rat model of fetal alcohol spectrum disorders (FASDs) 8. A collection of studies have demonstrated that rodents exposed to alcohol from postnatal day (PD) 4 through 9 display significant deficits in neuroanatomical measures such as dendritic complexity 25, cerebellar development 26,27 and neuroimmune responsiveness 28 as well as manifestations of impaired learning and memory 29–31. Even a reduced amount of alcohol exposure within this time window (i.e., PD 7 through 9) can lead to deficits in learning and memory in adolescent and adult rats 32 while some structures no longer see significant neuroanatomical impairment 27. Many of these deficits - in addition to behavioral impairments in hippocampus-dependent tasks - have been mitigated following exposure to this WR-EC paradigm 8,33 or WR alone 25,31. Although WR alone has been a widely used intervention, the combination of WR-EC has not yet been utilized in the literature despite its ability to sustain the relatively shorter-term benefits of WR 8. This article will discuss the implementation of the WR-EC intervention during adolescence. Although this paradigm is used in the context of early postnatal alcohol exposure, it can be introduced to various rodent models to assess brain potential for neuroplasticity in the models of brain disorders.
Protocol
Ethics Statement: The following protocol was approved by the Institutional Animal Care and Use Committee (IACUC) of the University of Delaware.
1. Developmental Exposure (or Model of Binge-Like Ethanol Exposure)
-
On PD3, determine the sex of each animal and cross-foster any animals if necessary to keep litter size (8 animals) and sex distribution (4 males : 4 females) consistent within each litter.
NOTE: It is important to keep litter size and sex distribution as consistent as possible to avoid experimental confounds. Although this protocol uses 8 pups (4 males and 4 females) per litter, alternative litter sizes or sex distributions may be tailored to the needs of the experimental design.
Subcutaneously inject a small amount of black India ink into the paws to identify animals within each litter.
-
Pseudo-randomly assign litters as experimental (containing 50 % alcohol-exposed (AE) and 50 % sham-intubated control (SI) pups) or suckle control (SC) (animals that do not undergo any intubation, tail clipping, or separation protocols from PD 4 - 9 except for daily weighing and ear-punching).
To retain consistent group size, assign twice as many experimental litters as SC litters.
-
Weigh each animal then return it to its home cage. Animal weighing should occur daily during the intubation period (PD 4 – 9).
Remove the whole litter from the dam.
Place pups on heated pad.
Record the weight of each individual pup.
-
On PD4, after weighing each animal calculate the necessary alcohol amount for a total of 5.25 g/kg/day per each animal (based on pup weight from step 1.4)8.
Administer alcohol as 11.9 % ethanol-in-milk substitute (vol/vol).
Starting at 9 AM, remove one litter's pups from the mother at a time.
Administer the ethanol-in-milk to each AE pup34.
Sham-intubate each SI pup8.
Repeat steps 1.5. through 1.8. for each experimental litter.
Two hours following the first dose, repeat the dosing procedure (steps 1.5 through 1.8) for a second alcohol dose.
-
One and a half hours after the second alcohol dose (the point at which peak daily blood alcohol content is achieved), collect and centrifuge blood from the AE and SI pups via tail clipping for future blood alcohol content analysis 35.
Collect 60 μL of blood.
Place blood in a 1 mL microcentrifuge tube. Centrifuge blood at 1.5 x g for 25 min.
Carefully collect the supernatant serum from the centrifuge tube and save for future blood alcohol content analysis.
-
Repeat the dosing procedure (steps 1.5 through 1.8) using milk instead of ethanol-in-milk to prevent nutritional deficits from nursing inability in AE pups.
Perform a total of 2 supplemental milk doses 2 h apart on PD 4.
Repeat steps 1.4 through 1.12 on PD 5 - 9.
-
Following the final supplemental milk dose on PD9, ear punch all pups for identification in the EC cage.
Coordinate punched ear with some measure of litter number or identifier (e.g., odd numbered litters within a cohort would get their left ear punched while animals from even numbered litters would get their right ears punched). This will make it easier to identify animals in the EC cage should multiple animals from different litters have the same pawmark pattern.
2. Weaning
-
On PD 23, house all animals in cages of 2 – 3.
Ensure that all animals housed in the same cage are the same sex.
Include one SC, one SI, and one AE animal per cage when possible.
Minimize the number of cage mates that are from the same litter.
Make sure all animals are capable of accessing food and water.
3. Wheel Running
-
On PD30, allocate half of the cages with animals to WR. House these animals in cages with a free access to attached stainless steel running wheel.
Ensure that wheels have a counter to assess the total number of revolutions.
Weigh all animals on PD 30 and PD 36.
Check the number of revolutions of each wheel at 9 AM every day.
Leave animals in their respective housing condition for 12 days.
4. Environmental Complexity
-
Prepare the EC cage before 9 AM on the day that corresponds to PD 42 for experimental animals.
-
Get a 30 " x 18 " x 36 " galvanized steel cage.
NOTE: The cage should have multiple levels, be capable of supporting the weight of multiple rats, be filled with standard bedding, and have multiple locations to attach water bottles and food dispensers.
-
Place novel, colorful objects of variable sizes and shapes in the cage.
Place 6 large toys in the EC cage. Ensure that each toy is big enough for 3 or more rats to interact with concurrently.
Place 6 medium toys in the EC cage. Ensure that each toy is big enough for 3 – 4 rats to interact with concurrently.
Place a lot (at least 20) of smaller toys in the EC cage.
Use toys of varying colors, shapes, size, etc. Novelty is critical to this intervention (see discussion).
Place two dishes of food at opposite ends of the cage.
Place two bottles of water at opposite ends of the cage.
-
-
At 9 AM on PD 42, weigh all animals and relocate the WR animals to the EC cage. Each EC cage should contain 9 – 12 animals.
Make sure that no animals have both the same pawmark and ear-punch patterns.
Check all food and water daily.
Every two days, remove the toys from the EC cage and replace them (according to step 4.1.2.).
-
Every three days, clean the EC cage.
Remove the animals from the EC cage and put them in temporary holding cages of 2 - 3 animals.
Remove all of the bedding from the bottom of the cage.
Return the same toys to the cage unless this day coincides with the toy replacement schedule (according to step 4.4.).
Replace all of the food and water.
Replace the rats into the EC cage.
5. Collect Tissue
NOTE: Tissue collection (e.g., perfusion with paraformaldehyde) and storage (e.g., freezing, paraffin embedding) can be performed with a variety of methods. The following will explain the process of perfusion with 4 % paraformaldehyde in 0.1 M phosphate buffered saline (4 % paraformaldehyde in PBS) solution 8.
Caution: Paraformaldehyde is carcinogenic and may also cause skin irritation, allergic skin reaction, or eye damage. Use appropriate eye/skin protection.
Expose one rat at a time to isoflurane to lightly anesthetize the animal.
Intraperitoneally inject the rat with 2 mL/kg of Ketamine/xylazine mixture (1.5 mL xylazine mixed with 10 mL of ketamine). NOTE: Ketamine and xylazine are both at stock concentrations of 100 mg/mL before combining for injection mixture.
Once rat is no longer responsive, perfuse the animal with 0.1 M phosphate buffered saline (PBS; pH = 7.2) followed by 4 % paraformaldehyde in PBS (pH = 7.2).
Remove brain and store in 4 % paraformaldehyde in PBS at 4 °C for 48 h.
After 2 days, transfer to solution of 30 % sucrose added to 4 % paraformaldehyde in PBS at 4 °C.
Representative Results
In order to assess the effect of the super intervention, we must look at the effects of each of its constituent elements - WR and EC - on our measures of interest. Figures 1 through 3 (below) appeared in a previous publication utilizing this paradigm 8. Figure 4 appeared in a doctoral dissertation 36. These data illustrate the impact of WR-EC on hippocampal adult neurogenesis in the dentate gyrus. All graphs illustrate group means, with error bars indicating a single standard error from mean. Figure 1 demonstrates increases in cell proliferation following the WR portion of our intervention, indicating that the WR component is robustly capable of increasing cellular proliferation in the DG of the hippocampus in normally developing, early-life stressed, and alcohol-exposed animals. Figure 2 demonstrates the ability of EC to increase survival of adult generated cells in the DG in animals that were exposed to either stress or alcohol neonatally. Figure 3 demonstrates the increase in cells that differentiate into a neuronal phenotype, indicating that WR-EC can increase proliferation and survival of adult-born dentate gyrus granule cells in animals that undergo neonatal exposure to alcohol or intubation stress, implicating it as a therapeutic to rescue deficits in hippocampal adult neurogenesis. Finally, Figure 4 confirms the WR-EC effect on dendritic plasticity: the length of doublecortin-positive dendrites of dentate gyrus' granule cells in AE rats is no longer different from control. Blood alcohol content (BAC) on PD 4 was 321.19 ± 14.03 mg/dl (mean ± SEM), comparable to other studies using this exposure paradigm 28,37. Previous studies have demonstrated that animals across these treatment groups do not differ in distances run during WR 15.
Figure 1. WR Robustly Increases Cell Proliferation in the Hippocampal DG.

Photomicrographs illustrate differences in cell proliferation in the DG on PD42 (the cessation of WR) as labeled with Bromo-deoxyuridine (BrdU) in AE animals following WR (A) and social housing (B). WR robustly increases cell proliferations irrespective of neonatal treatment (C). A two-way ANOVA revealed a main effect of housing condition (WR vs. SH) (F1,40 = 19.703, p < 0.001), while no significant main effect of postnatal treatment (SC vs. SI vs. AE) or interaction between the two factors were observed. Post hoc comparisons were performed as Tukey's tests. All values represent mean ± standard error of the mean (SEM). *p < 0.05, #p < 0.01. This figure has been reproduced from Hamilton et al., 20128.
Figure 4. WR-EC Rescues Deficits in Dendritic Complexity of Hippocampal DG Granule Cells.

Sholl analyses of dendritic intersections illustrate WR-EC's ameliorative effects on dendritic complexity in the dentate gyrus of adult rats following neonatal alcohol exposure. In social housing conditions, AE animals have a decreased number of DG granule cell dendrite intersections relative to control animals (A). Housing in WREC increases the number of intersections in AE animals relative to socially housed controls (B). AE animals reared in our WREC paradigm display similar numbers of intersections relative to control animals housed in WREC (C). Repeated measure ANOVAs were performed on the data in each graph. Panel A demonstrates a main effect of postnatal treatment (F1,11 = 6.265, p = 0.029). Panel B demonstrates a trend toward a main effect between housing conditions (F1,6 = 4.181,p = 0.087). Panel C demonstrates no significant difference between SC and AE animals within the WREC housing condition. All post hoc comparisons were performed as Tukey's tests. All values represent mean ± SEM. ^ p < 0.01, *p < 0.05. This figure has been reproduced from Hamilton, 201236.
Figure 2. WR Followed by EC Rescues Deficits in Cell Survival Following Neonatal Alcohol Exposure or Sham Stress.
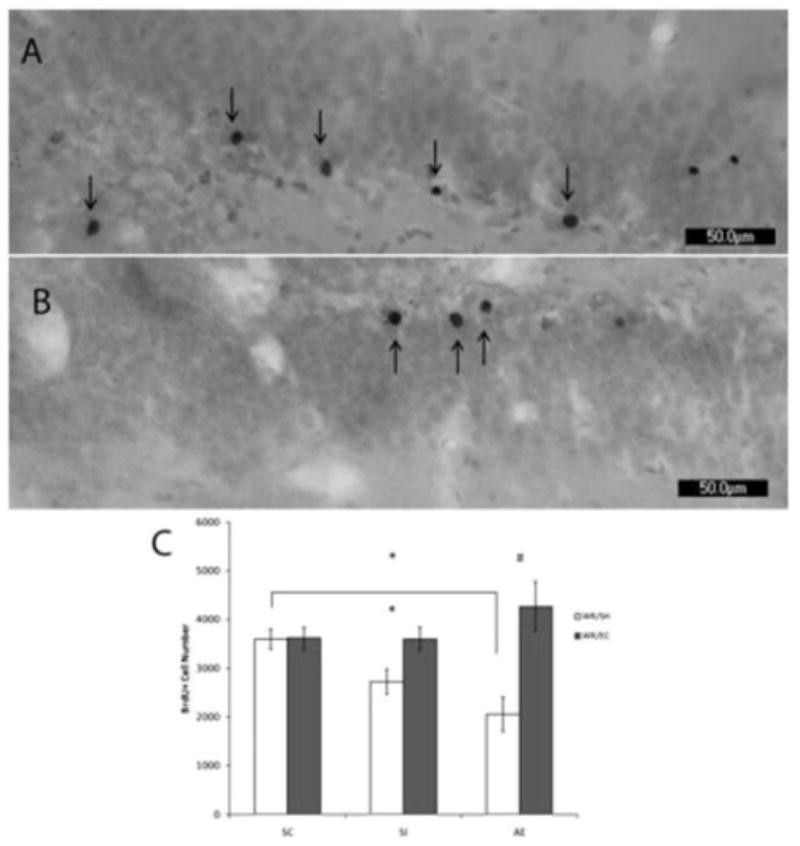
Photomicrographs illustrate differences in cells labeled with BrdU in AE animals from WR-EC (A) and social housed conditions (B) injected with BrdU on PD41. Socially housed animals displayed a decrease following alcohol exposure relative to suckle controls. Animals undergoing the WR-EC superintervention display increased survival rates of cells proliferating after PD41 in both SI and AE groups (C). A two-way ANOVA revealed a main effect of housing condition (WR vs. SH) (F1,29 = 11.402, p < 0.01) and a significant interaction between postnatal treatment and housing condition (F1,29 = 3.870, p < 0.05), while no significant main effect of postnatal treatment (SC vs. SI vs. AE) was observed. A one-way ANOVA within SH animals revealed a main effect of postnatal treatment (F1,19 = 3.727, p < 0.05) whereas a one-way ANOVA within WREC animals revealed no significant differences between postnatal treatments. Post hoc comparisons were performed as Tukey's tests. All values represent mean ± SEM. *p < 0.05, #p < 0.01. This figure has been reproduced from Hamilton et al., 20128.
Figure 3. WR-EC Rescues Deficits in Neurogenesis Following Neonatal Alcohol Exposure or Sham Stress.
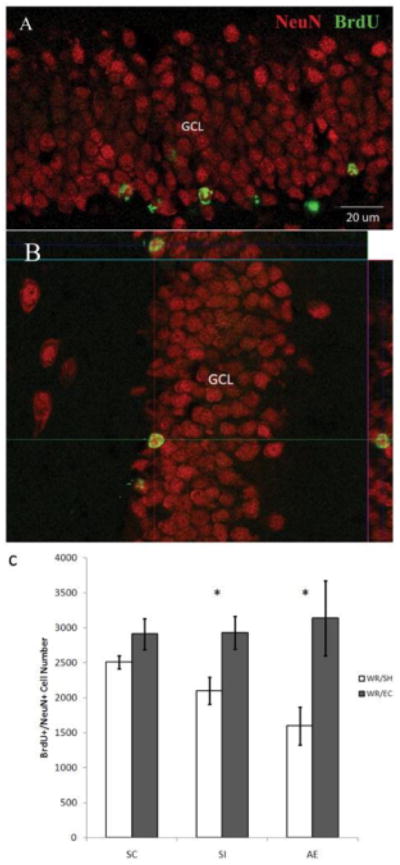
Co-localization of BrdU (green) expression and NeuN (red) in hippocampal granule cells. Fluorescent confocal images were acquired following immunohistochemical procedures. BrdU was injected on PD41 tissue was collected on PD72. Both BrdU and NeuN were observed in the DG (A, B). Although SC animals did not show a significant increase in number of proliferating neurons, both AE and SI animals showed an increase in neurogenesis (as indicated by double labeling with BrdU and NeuN) following the WR-EC paradigm compared to socially housed animals (C). A two-way ANOVA revealed a main effect of housing condition (WR vs. SH) (F1,28 = 20.48, p < 0.001), while no significant main effect of postnatal treatment (SC vs. SI vs. AE) or interaction between the two factors were observed. Post hoc comparisons were performed as Tukey's tests. All values represent mean ± SEM. *p < 0.05, #p < 0.01. This figure has been reproduced from Hamilton et al., 20128.
Discussion
In the above protocol, we demonstrated an expedient intervention to rescue neuroanatomical deficits following neonatal alcohol exposure. This intervention can be used as a therapeutic in other animal models due to the robustness of each of the components of the intervention. Voluntary cardiovascular activity in the form of WR has been shown to benefit several behavioral outcomes 38,39 and induce functional plastic alterations in brain regions such as the hippocampus (reviewed in 40). This is in part due to expression of growth factors and other neuroprotective mechanisms in the brain parenchyma in both rodents and humans 21,41. Supplementing these effects, EC can induce beneficial cellular 6,11,42,43, structural 2 and pharmacological 12,44 change in rodents.
In order for WR to be maximally effective in this particular model of human syndrome, it is critical for animals to have voluntary access to a functional running wheel; daily wheel access should last for an extended amount of time45 at least 10–12 h per day and preferably 24 h (some adverse effects of withdrawal from the running wheel were reported). This WR paradigm lasts for 12 days to allow for the combination of WR and EC to fit into adolescence and early adulthood. The duration, age at exposure, and modality of exercise (among other factors) can affect the efficacy of exercise as a therapeutic intervention46, and such critical factors should be considered when planning to implement this protocol or any other WREC paradigm. A key component of this EC paradigm is the novelty of the multiple objects in the environment and social interaction (reviewed in 14,47). Therefore, it is critical for the items in this paradigm to be replaced every 48 h. Based on the need for multiple items, the interaction with the items and their exploration, and social interaction, we find that our number of unique items, frequency of item replacement, and number of cage mates is sufficient to induce therapeutic outcomes on the neuroanatomical measures that we assess. We found that continuous exposure for 30 days is more appropriate to overcome deficits induced by neonatal alcohol exposure than limited exposure interaction to a novel environment.
The goal of this protocol is to introduce a WREC paradigm that addresses both the cardiovascular exercise and environmental novelty components of plastic intervention. For this reason, we will address the modification that can be made to the paradigm but would caution use of modifications that may alter the ways that animals interact within the paradigm as well as the experimental conclusions that can be drawn. One possible alteration would be the introduction of running wheels to the EC environment. In doing so, it would be difficult to determine the relative contributions of each component. It would additionally be difficult to assure that all animals participate in both the WR components and EC components of the paradigm as housing of 8 – 10 animals together is required for EC. However, since long-term access to exercise is critical in the efficacy of this intervention 45, further research may address the optimal ratio of WR access to EC access (although the methods in this protocol have shown robust neuroanatomical and behavioral implications 8,33). Modifications to individual items used within the EC environment are acceptable, but it is critical for the items to be interesting, complex, novel, stimulating and frequently refreshed 14.
This paradigm does contain several innate limitations in our hands, which should be considered when planning to implement this "super intervention". One limitation to the WR component of the paradigm is the inability to assess the distance run by individual animals. One of the obvious and straightforward solutions would be an individual housing of the animals during WR component. However, it needs to be stressed that individual housing is widely accepted as detrimental to animals and can even directly counteract the beneficial effects of wheel running 48. An additional alternative (although time-consuming and imperfect) would be to video record the running wheel at all times that the animals have access. This would require a unique identifier for each animal in a cage (e.g., painting unique colors or patterns on the fur of each animal)49. This technique would still be subject to confounds of multiple animals utilizing the wheel concurrently. A similar difficulty carries to EC where it becomes difficult to food restrict individual animals (without limiting the time period of food consumption). To reduce the impact of this, we would recommend housing in EC for a full 30 days followed by an immediate food restriction paradigm. Extended amounts of time out of EC could inhibit induced plasticity that occurs during this paradigm.
As mentioned previously, the importance of this article is to allow for consistent characterization of the EC paradigm and its implementation following cardiovascular exercise in the form of WR. Previous EC paradigms have exposed animals to EC housing without exposure to WR 12,50, WR inside of the EC cage for a shorter amount of time 51 or with less animals 52, or the animals were exposed to an EC environment for a longer amount of time with less frequent changing of cage items 13. It is likely that the beneficial effects of EC require the induced plasticity from WR in a temporally relevant time window to show long-term benefit. In this way, we believe that coupling WR and EC for 12 and 30 days respectively allows for a maximally beneficial and concise intervention.
At this point, the use of this model has been limited to the adolescent and early adult time periods. Further examination of the robustness of this intervention at different stages, and the ontogeny of neuroplastic benefit should be examined further in the future. Additionally, the use of different developmental deficits is greatly encouraged, as this will assist in developing effective therapeutic interventions for individuals afflicted by such disorders. Previous literature has demonstrated independent effects of WR or EC on adult neurogenesis, learning and memory, or anxiety-like behaviors in a genetic mouse model of anxiety 53. The robustness of these two interventions and the synergistic effect of EC to sustain the short-term effects of increased WR-induced benefits (i.e., hippocampal cell proliferation and neurogenesis) makes it well poised for integration into a diverse range of research questions.
Acknowledgments
We would like to dedicate this work to the memory of late Dr. William T. Greenough, a great mentor, a colleague and a friend. This work was supported by NIH/NIAAA grant number AA009838 and NIH/NIGMS COBRE: The Delaware Center for Neuroscience research grant 1P20GM103653 to AYK. We are grateful to the former and current members of Klintsova lab.
Footnotes
Video Link
The video component of this article can be found at http://www.jove.com/video/54947/
Disclosures
The authors have nothing to disclose.
References
- 1.Diamond MC, et al. Increases in cortical depth and glia numbers in rats subjected to enriched environment. J Comp Neurol. 1966;128(1):117–126. doi: 10.1002/cne.901280110. [DOI] [PubMed] [Google Scholar]
- 2.Rosenzweig MR, Bennett EL, Hebert M, Morimoto H. Social grouping cannot account for cerebral effects of enriched environments. Brain Res. 1978;153(3):563–576. doi: 10.1016/0006-8993(78)90340-2. [DOI] [PubMed] [Google Scholar]
- 3.Greenough WT. Experiential modification of the developing brain. Am Sci. 1975;63(1):37–46. [PubMed] [Google Scholar]
- 4.Volkmar FR, Greenough WT. Rearing complexity affects branching of dendrites in the visual cortex of the rat. Science. 1972;176(4042):1445–1447. doi: 10.1126/science.176.4042.1445. [DOI] [PubMed] [Google Scholar]
- 5.Greenough WT, Volkmar FR, Juraska JM. Effects of rearing complexity on dendritic branching in frontolateral and temporal cortex of the rat. Exp Neurol. 1973;41(2):371–378. doi: 10.1016/0014-4886(73)90278-1. [DOI] [PubMed] [Google Scholar]
- 6.Sampedro-Piquero P, Zancada-Menendez C, Begega A. Housing condition-related changes involved in reversal learning and its c-Fos associated activity in the prefrontal cortex. Neuroscience. 2015;307:14–25. doi: 10.1016/j.neuroscience.2015.08.038. [DOI] [PubMed] [Google Scholar]
- 7.Sirevaag AM, Greenough WT. Differential rearing effects on rat visual cortex synapses. III. Neuronal and glial nuclei, boutons, dendrites, and capillaries. Brain Res. 1987;424(2):320–332. doi: 10.1016/0006-8993(87)91477-6. [DOI] [PubMed] [Google Scholar]
- 8.Hamilton GF, Boschen KE, Goodlett CR, Greenough WT, Klintsova AY. Housing in environmental complexity following wheel running augments survival of newly generated hippocampal neurons in a rat model of binge alcohol exposure during the third trimester equivalent. Alcohol Clin Exp Res. 2012;36(7):1196–1204. doi: 10.1111/j.1530-0277.2011.01726.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kempermann G, Kuhn HG, Gage FH. More hippocampal neurons in adult mice living in an enriched environment. Nature. 1997;386(6624):493–495. doi: 10.1038/386493a0. [DOI] [PubMed] [Google Scholar]
- 10.Juraska JM, Greenough WT, Elliott C, Mack KJ, Berkowitz R. Plasticity in adult rat visual cortex: an examination of several cell populations after differential rearing. Behav Neural Biol. 1980;29(2):157–167. doi: 10.1016/s0163-1047(80)90482-3. [DOI] [PubMed] [Google Scholar]
- 11.Turner AM, Greenough WT. Differential rearing effects on rat visual cortex synapses. I. Synaptic and neuronal density and synapses per neuron. Brain Res. 1985;329(1–2):195–203. doi: 10.1016/0006-8993(85)90525-6. [DOI] [PubMed] [Google Scholar]
- 12.Brenes JC, Rodriguez O, Fornaguera J. Differential effect of environment enrichment and social isolation on depressive-like behavior, spontaneous activity and serotonin and norepinephrine concentration in prefrontal cortex and ventral striatum. Pharmacol Biochem Behav. 2008;89(1):85–93. doi: 10.1016/j.pbb.2007.11.004. [DOI] [PubMed] [Google Scholar]
- 13.Kolb B, Gorny G, Soderpalm AH, Robinson TE. Environmental complexity has different effects on the structure of neurons in the prefrontal cortex versus the parietal cortex or nucleus accumbens. Synapse. 2003;48(3):149–153. doi: 10.1002/syn.10196. [DOI] [PubMed] [Google Scholar]
- 14.Singhal G, Jaehne EJ, Corrigan F, Baune BT. Cellular and molecular mechanisms of immunomodulation in the brain through environmental enrichment. Front Cell Neurosci. 2014;8:97. doi: 10.3389/fncel.2014.00097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Helfer JL, Goodlett CR, Greenough WT, Klintsova AY. The effects of exercise on adolescent hippocampal neurogenesis in a rat model of binge alcohol exposure during the brain growth spurt. Brain Res. 2009;1294:1–11. doi: 10.1016/j.brainres.2009.07.090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Praag H, et al. Functional neurogenesis in the adult hippocampus. Nature. 2002;415(6875):1030–1034. doi: 10.1038/4151030a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Praag H, Shubert T, Zhao C, Gage FH. Exercise enhances learning and hippocampal neurogenesis in aged mice. J Neurosci. 2005;25(38):8680–8685. doi: 10.1523/JNEUROSCI.1731-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vivar C, Peterson BD, van Praag H. Running rewires the neuronal network of adult-born dentate granule cells. Neuroimage. 2015;1(131):29–41. doi: 10.1016/j.neuroimage.2015.11.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mustroph ML, et al. Increased adult hippocampal neurogenesis is not necessary for wheel running to abolish conditioned place preference for cocaine in mice. Eur J Neurosci. 2015;41(2):216–226. doi: 10.1111/ejn.12782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Patten AR, et al. Long-term exercise is needed to enhance synaptic plasticity in the hippocampus. Learn Mem. 2013;20(11):642–647. doi: 10.1101/lm.030635.113. [DOI] [PubMed] [Google Scholar]
- 21.Carro E, Nunez A, Busiguina S, Torres-Aleman I. Circulating insulin-like growth factor I mediates effects of exercise on the brain. J Neurosci. 2000;20(8):2926–2933. doi: 10.1523/JNEUROSCI.20-08-02926.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cotman CW, Berchtold NC. Exercise: a behavioral intervention to enhance brain health and plasticity. Trends Neurosci. 2002;25(6):295–301. doi: 10.1016/s0166-2236(02)02143-4. [DOI] [PubMed] [Google Scholar]
- 23.Praag H, Fleshner M, Schwartz MW, Mattson MP. Exercise, energy intake, glucose homeostasis, and the brain. J Neurosci. 2014;34(46):15139–15149. doi: 10.1523/JNEUROSCI.2814-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Praag H, Kempermann G, Gage FH. Neural consequences of environmental enrichment. Nat Rev Neurosci. 2000;1(3):191–198. doi: 10.1038/35044558. [DOI] [PubMed] [Google Scholar]
- 25.Hamilton GF, Criss KJ, Klintsova AY. Voluntary exercise partially reverses neonatal alcohol-induced deficits in mPFC layer II/III dendritic morphology of male adolescent rats. Synapse. 2015;69(8):405–415. doi: 10.1002/syn.21827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Goodlett CR, Thomas JD, West JR. Long-term deficits in cerebellar growth and rotarod performance of rats following "binge-like" alcohol exposure during the neonatal brain growth spurt. Neurotoxicol Teratol. 1991;13(1):69–74. doi: 10.1016/0892-0362(91)90029-v. [DOI] [PubMed] [Google Scholar]
- 27.Goodlett CR, Lundahl KR. Temporal determinants of neonatal alcohol-induced cerebellar damage and motor performance deficits. Pharmacol Biochem Behav. 1996;55(4):531–540. doi: 10.1016/s0091-3057(96)00248-1. [DOI] [PubMed] [Google Scholar]
- 28.Boschen K, Ruggiero M, Klintsova A. Neonatal binge alcohol exposure increases microglial activation in the developing rat hippocampus. Neuroscience. 2016;324:355–366. doi: 10.1016/j.neuroscience.2016.03.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Goodlett CR, Peterson SD. Sex differences in vulnerability to developmental spatial learning deficits induced by limited binge alcohol exposure in neonatal rats. Neurobiol Learn Mem. 1995;64(3):265–275. doi: 10.1006/nlme.1995.0009. [DOI] [PubMed] [Google Scholar]
- 30.Murawski NJ, Klintsova AY, Stanton ME. Neonatal alcohol exposure and the hippocampus in developing male rats: effects on behaviorally induced CA1 c-Fos expression, CA1 pyramidal cell number, and contextual fear conditioning. Neuroscience. 2012;206:89–99. doi: 10.1016/j.neuroscience.2012.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Thomas JD, Sather TM, Whinery LA. Voluntary exercise influences behavioral development in rats exposed to alcohol during the neonatal brain growth spurt. Behav Neurosci. 2008;122(6):1264–1273. doi: 10.1037/a0013271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hamilton GF, et al. Neonatal alcohol exposure disrupts hippocampal neurogenesis and contextual fear conditioning in adult rats. Brain Res. 2011;1412:88–101. doi: 10.1016/j.brainres.2011.07.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hamilton GF, et al. Exercise and environment as an intervention for neonatal alcohol effects on hippocampal adult neurogenesis and learning. Neuroscience. 2014;265:274–290. doi: 10.1016/j.neuroscience.2014.01.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kelly SJ, Lawrence CR. In: Alcohol: Methods and Protocols. Nagy LE, editor. 2008. [Google Scholar]
- 35.Helfer JL, et al. Binge-like postnatal alcohol exposure triggers cortical gliogenesis in adolescent rats. J Comp Neurol. 2009;514(3):259–271. doi: 10.1002/cne.22018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hamilton GF. Doctor of Philosophy thesis. University of Delaware; 2012. Behavioral Interventions to Alleviate the Impact of Neonatal Alcohol Exposure on Cell Morphology in the Rodent Hippocampus and Medial Prefrontal Cortex. [Google Scholar]
- 37.Klintsova AY, et al. Persistent impairment of hippocampal neurogenesis in young adult rats following early postnatal alcohol exposure. Alcohol Clin Exp Res. 2007;31(12):2073–2082. doi: 10.1111/j.1530-0277.2007.00528.x. [DOI] [PubMed] [Google Scholar]
- 38.Brockett AT, LaMarca EA, Gould E. Physical exercise enhances cognitive flexibility as well as astrocytic and synaptic markers in the medial prefrontal cortex. PLoS One. 2015;10(5):e0124859. doi: 10.1371/journal.pone.0124859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Creer DJ, Romberg C, Saksida LM, van Praag H, Bussey TJ. Running enhances spatial pattern separation in mice. Proc Natl Acad Sci U S A. 2010;107(5):2367–2372. doi: 10.1073/pnas.0911725107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Patten AR, et al. The benefits of exercise on structural and functional plasticity in the rodent hippocampus of different disease models. Brain Plast. 2015;1(1):97–127. doi: 10.3233/BPL-150016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Van der Borght K, et al. Physical exercise leads to rapid adaptations in hippocampal vasculature: temporal dynamics and relationship to cell proliferation and neurogenesis. Hippocampus. 2009;19(10):928–936. doi: 10.1002/hipo.20545. [DOI] [PubMed] [Google Scholar]
- 42.Johansson BB, Belichenko PV. Neuronal plasticity and dendritic spines: effect of environmental enrichment on intact and postischemic rat brain. J Cereb Blood Flow Metab. 2002;22(1):89–96. doi: 10.1097/00004647-200201000-00011. [DOI] [PubMed] [Google Scholar]
- 43.Sirevaag AM, Greenough WT. Differential rearing effects on rat visual cortex synapses. II. Synaptic morphometry. Brain Res. 1985;351(2):215–226. doi: 10.1016/0165-3806(85)90193-2. [DOI] [PubMed] [Google Scholar]
- 44.Pham TM, Winblad B, Granholm AC, Mohammed AH. Environmental influences on brain neurotrophins in rats. Pharmacol Biochem Behav. 2002;73(1):167–175. doi: 10.1016/s0091-3057(02)00783-9. [DOI] [PubMed] [Google Scholar]
- 45.Patten AR, et al. Long-term exercise is needed to enhance synaptic plasticity in the hippocampus. Learn Mem. 2013;20(11):642–647. doi: 10.1101/lm.030635.113. [DOI] [PubMed] [Google Scholar]
- 46.Patten AR, et al. The Benefits of Exercise on Structural and Functional Plasticity in the Rodent Hippocampus of Different Disease Models. Brain Plasticity. 2015;1(1):97–127. doi: 10.3233/BPL-150016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Abou-Ismail UA. Are the effects of enrichment due to the presence of multiple items or a particular item in the cages of laboratory rat? Appl Ani Behav Sci. 2011;134(1–2):72–82. [Google Scholar]
- 48.Stranahan AM, Khalil D, Gould E. Social isolation delays the positive effects of running on adult neurogenesis. Nat Neurosci. 2006;9(4):526–533. doi: 10.1038/nn1668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Boschen KE, Hamilton GF, Delorme JE, Klintsova AY. Activity and social behavior in a complex environment in rats neonatally exposed to alcohol. Alcohol. 2014;48(6):533–541. doi: 10.1016/j.alcohol.2014.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Artola A, et al. Long lasting modulation of the induction of LTD and LTP in rat hippocampal CA1 by behavioural stress and environmental enrichment. Eur J Neurosci. 2006;23(1):261–272. doi: 10.1111/j.1460-9568.2005.04552.x. [DOI] [PubMed] [Google Scholar]
- 51.Bergami M, et al. A critical period for experience-dependent remodeling of adult-born neuron connectivity. Neuron. 2015;85(4):710–717. doi: 10.1016/j.neuron.2015.01.001. [DOI] [PubMed] [Google Scholar]
- 52.Fréchette M, Rennie K, Pappas BA. Developmental forebrain cholinergic lesion and environmental enrichment: behaviour, CA1 cytoarchitecture and neurogenesis. Brain Res. 2009;1252:172–182. doi: 10.1016/j.brainres.2008.11.082. [DOI] [PubMed] [Google Scholar]
- 53.Rogers J, et al. Dissociating the therapeutic effects of environmental enrichment and exercise in a mouse model of anxiety with cognitive impairment. Transl Psychiatry. 2016;6:e794. doi: 10.1038/tp.2016.52. [DOI] [PMC free article] [PubMed] [Google Scholar]