

Abstract
The yeast heme activator protein Hap1 binds to DNA and activates transcription of genes encoding functions required for respiration and for controlling oxidative damage, in response to heme. Hap1 contains a DNA-binding domain with a C6 zinc cluster motif, a coiled-coil dimerization element, typical of the members of the yeast Gal4 family, and an acidic activation domain. The regulation of Hap1 transcription-activating activity is controlled by two classes of Hap1 elements, repression modules (RPM1–3) and heme-responsive motifs (HRM1–7). Previous indirect evidence indicates that Hap1 may repress transcription directly. Here we show, by promoter analysis, by chromatin immunoprecipitation, and by electrophoretic mobility shift assay, that Hap1 binds directly to DNA and represses transcription of its own gene by at least 20-fold. We found that Hap1 repression of the HAP1 gene occurs independently of heme concentrations. While DNA binding is required for transcriptional repression by Hap1, deletion of Hap1 activation domain and heme-regulatory elements has varying effects on repression. Further, we found that repression by Hap1 requires the function of Hsp70 (Ssa), but not Hsp90. These results show that Hap1 binds to its own promoter and represses transcription in a heme-independent but Hsp70-dependent manner.
THE yeast heme activator protein (Hap)1 plays an important role in the activation of genes encoding functions required for respiration and for controlling oxidative damage, in response to oxygen or heme (Creusot et al. 1988; Pfeifer et al. 1989; Zhang and Hach 1999). The Hap1-activated genes include those encoding cytochrome c-iso-1 (CYC1), iso-2 (CYC7), cytochrome c1 (CYT1), catalase (CTT1), and flavohemoglobin (YHB1; Guarente and Mason 1983; Guarente et al. 1984; Pfeifer et al. 1987; Lowry and Zitomer 1988; Winkler et al. 1988; Lodi and Guiard 1991; Schneider and Guarente 1991; Zhao et al. 1996; Buisson and Labbe-Bois 1998). Hap1 also activates transcription of the ROX1 gene (Lowry and Zitomer 1988; Zitomer and Lowry 1992; Zitomer et al. 1997), encoding the aerobic repressor Rox1, which represses the expression of genes encoding functions specifically required for anaerobic growth, such as ANB1 (Lowry and Zitomer 1988; Ushinsky and Keng 1994).
The mechanism of transcriptional activation by Hap1 has been intensely investigated (Creusot et al. 1988; Pfeifer et al. 1989; Zhang and Hach 1999). Hap1 binds to DNA via a C6 zinc cluster motif, which is located near the N terminus and is found in members of the Gal4 transcriptional regulator family (Creusot et al. 1988; Pfeifer et al. 1989). Like Gal4, Hap1 binds as a dimer to DNA containing two CGG triplets, with each C6 zinc cluster motif recognizing one CGG (Zhang and Guarente 1994c, 1996). However, unlike Gal4, which binds to symmetrical DNA sites containing an inverted repeat of two CGGs, Hap1 binds to asymmetric sites containing a direct repeat of two CGGs (optimal sequence, CGGnnnTAnCGG; Zhang and Guarente 1994c, 1996). Hap1 transcription-activating activity is conferred by its acidic activation domain located at its C terminus (Pfeifer et al. 1989).
Hap1 transcription-activating activity is precisely controlled by intracellular heme concentration (Creusot et al. 1988; Pfeifer et al. 1989; Zhang and Hach 1999). Previous work showed that the regulation of Hap1 activity by heme involves a two-tier regulatory mechanism: inactivation in the absence of heme and activation by heme (Hach et al. 1999; Zhang and Hach 1999; Hon et al. 2000). Hap1 inactivation in the absence of heme requires the proper functioning of a class of Hap1 elements, repression modules (RPM1–3), and the molecular chaperone Hsp70 and cochaperones (Hach et al. 1999; Hon et al. 2001). Disrupting the function of RPMs or Hsp70 causes Hap1 to gain high levels of activity in the absence of heme (Hach et al. 1999; Hon et al. 2001). Hap1 activation by heme is mediated by another class of Hap1 elements, heme-responsive motifs (HRM1–6 and HRM7), and the molecular chaperone Hsp90 (Hach et al. 1999; Hon et al. 2000; Lee et al. 2002, 2003). Particularly, HRM7 plays a predominant role, while HRM1–6 play an auxiliary role, in heme activation (Hach et al. 1999; Lee et al. 2003).
Previous evidence suggests that Hap1 represses transcription of several genes, including ROX1 and ERG11 (Verdiere et al. 1991; Defranoux et al. 1994; Deckert et al. 1995; Pinkham et al. 1997), in the absence of heme. However, there is no direct evidence showing that Hap1 binds to the promoters of these genes and promotes repression. Furthermore, it is unclear how Hap1 may repress transcription. Does transcriptional repression by Hap1 also involve Hap1 elements HRMs and RPMs and molecular chaperones? Is DNA binding necessary for repression by Hap1? In this report, to answer these questions, we explore the possibility of the repression of the HAP1 gene by Hap1. We found that Hap1 indeed strongly represses the transcription of its own gene and that it does so by a heme-independent mechanism distinct from its mechanism of transcriptional activation.
MATERIALS AND METHODS
Yeast strains and reporter plasmids:
Yeast strains used were BWG 1-7aΔtrp1 (MATa, ura3-52, leu2-3, 112, his4-519, ade1-100, trp1::HisG; wild-type control for Δhap1), L51 (MATa, ura3-52, leu2-3, 112, his4-519, ade1-100, trp1::HisG, hap1::LEU2) (Δhap1; Zhang and Guarente 1994a), MHY100 (MATa, ura3-52, leu2-3, 112, his4-519, ade1-100, hem1-Δ100) (Δhem1; Haldi and Guarente 1989), MHY200 (MATa, ura3-52, leu2-3, 112, his4-519, ade1-100, hem1-Δ100, hap1::LEU2) (Δhap1Δhem1; Haldi and Guarente 1995), JN55 (MATa, ura3-52, leu2-3, 112, his3-11, trp1Δ1, lys2) (wild-type control for a2a3a4), JN516 (MATa, ura3-52, leu2-3, 112, his3-11, trp1Δ1, lys2, ssa2::LEU2, ssa3::TRP1, ssa4::LYS2) (a2a3a4; Becker et al. 1996), W303 (MATα, can1-100, ade2-1, his3-11,15, leu2-3,112, trp1-1, ura3-1) (wild-type control for iLEP), and iLEP (MATα, can1-100, ade2-1, his3-11,15, leu2-3,112, trp1-1, ura3-1, hsc82::LEU2, hsp82::LEU2, pLEP1HSP82; Chang and Lindquist 1994).
The full-length HAP1 promoter-lacZ reporter H1 (Table 1 and Figures 1, 2, and 5) and the reporter H2–H4 (Figure 2) were constructed by inserting the indicated DNA fragment containing the full-length or part of the HAP1 promoter into the pLG178 reporter plasmid (Guarente and Mason 1983; Guarente et al. 1984) digested with BamHI and XhoI. The resulting reporter plasmids do not contain the promoter of the CYC1 gene present in pLG178. The plasmids were confirmed by restriction digestion and sequencing. The reporter plasmid C1 was constructed by inserting the indicated DNA fragment containing part of the HAP1 promoter into the pLG178 reporter plasmid (Guarente and Mason 1983; Guarente et al. 1984) digested with SmaI and XhoI. The resulting reporter plasmid contains the basal promoter of the CYC1 gene present in pLG178. The pLG178 reporter plasmid contains only the basal promoter elements of the CYC1 gene (−1 to −178) and no upstream activation sequence (UAS; Guarente and Mason 1983; Guarente et al. 1984) and exhibits a low, basal level activity in all cells used here and elsewhere (Guarente and Mason 1983; Guarente et al. 1984; Lee et al. 2002). All expression plasmids for Hap1 and mutants have been described previously (Pfeifer et al. 1989; Zhang and Guarente 1994a; Hach et al. 1999; Lee et al. 2003).
TABLE 1.
The effects of Hap1 and heme on the promoter activity ofHAP1
| Cells | Wild type | Δhap1 | Δhem1: low heme |
Δhem1: high heme |
Δhem1 Δhap1: low heme |
Δhem1 Δhap1: high heme |
|---|---|---|---|---|---|---|
| HAP1-LacZ | 15 ± 2 | 265 ± 53 | 18 ± 2 | 19 ± 3 | 597 ± 62 | 475 ± 70 |
| UAS1/CYC1-LacZ | 314 ± 12 | 2 ± 0.3 | <1.0 | 439 ± 2 | <1.0 | <1.0 |
Yeast wild-type, Δhap1, Δhem1, and Δhem1 Δhap1 cells were transformed with a reporter fusion gene with the promoter sequence of the HAP1 gene (−461 to −1) inserted at the N terminus of the β-galactosidase-coding lacZ gene or with the UAS1-CYC1-lacZ reporter (for comparison). Δhem1 and Δhem1 Δhap1 cells were grown in 2 μm/ml δ-aminolevulinic acid for low heme levels (low heme) or 250 μ/ml for high heme levels (high heme).
Figure 1.—

(A) The repression of the HAP1 promoter reporter by Hap1 mutants. The domain structures of the Hap1 mutants with point mutation or deletion are shown. Wild-type Hap1 is shown to contain the C6 zinc cluster motif (Zn), the dimerization element (DE), three repression modules (RPM1–3), seven heme-responsive motifs (HRM1–6 and HRM7), and the activation domain (ACT). Mutant Hap1-18 contains a mutation of Ser-to-Arg at amino acid position 63. The deleted regions in mutants Δheme, H7-d1, H7-d2, and smHap1 are shown by angled lines. HB, HG, and HK contain residues 1–444, 1–746, and 1–1308, respectively. The expression plasmids for Hap1 mutants were transformed into Δhap1 cells bearing the full HAP1 promoter-lacZ reporter or the UAS1/CYC1-lacZ reporter, which contains the UAS1 sequence to which Hap1 binds, but not the UAS2 sequence to which the Hap2/3/4/5 complex binds (Guarente et al. 1984; Turcotte and Guarente 1992). Cells were grown in selective medium, and β-galactosidase activities were detected. The data shown are averages from at least three independent transformants. (B) Western blot showing the levels of wild-type and mutant Hap1 proteins. Extracts prepared from cells expressing wild-type Hap1 (lane 1), Hap1-18 (lane 2), Δheme (lane 3), H7-d1 (lane 4), H7-d2 (lane 5), smHap1 (lane 6), HB (lane 7), and HK (lane 8) were subjected to electrophoresis in a 7.5% SDS-polyacrylamide gel and transferred to a polyvinyl difluoride membrane. The levels of Hap1 proteins were detected by probing Western blots with antibodies against Hap1.
Figure 2.—
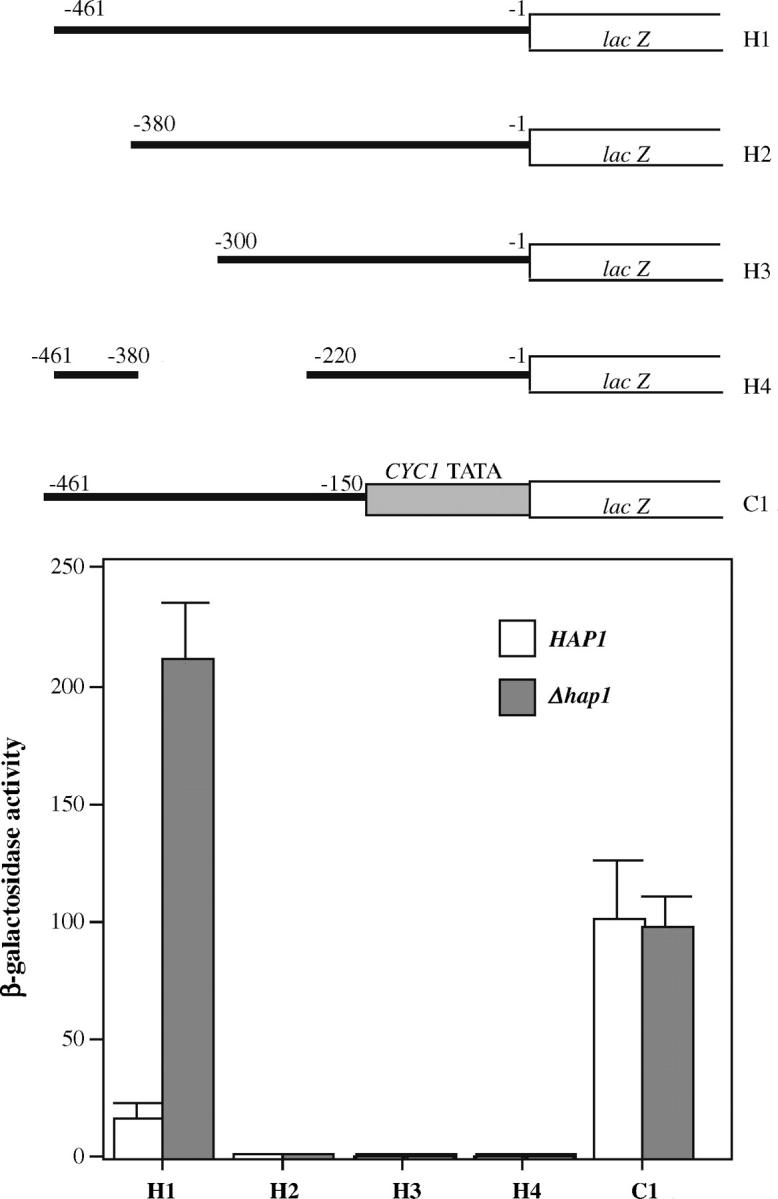
Deletion analysis of the HAP1 promoter. Shown in the top half are the sequence structures of the various constructed HAP1 promoter reporter genes. Yeast wild-type and Δhap1 cells were transformed with the indicated reporter plasmid. Cells were grown in selective medium, and β-galactosidase activities were detected. The plotted data are averages from at least three independent transformants.
β-galactosidase assays:
To measure β-galactosidase levels from the HAP1 promoter reporter gene at low and high heme concentrations, yeast cells were transformed with the indicated reporter plasmid. Cells were grown in synthetic complete media containing 2% glucose with a low level (2 μg/ml) or high level (250 μg/ml) of heme precursor δ-aminolevulinate to an optical density (OD) of ∼1.0. Cells were harvested for determination of β-galactosidase activity as described (Zhang et al. 1998; Hon et al. 2000). β-Galactosidase levels from the UAS1/CYC1-lacZ reporter (Guarente et al. 1984; Turcotte and Guarente 1992) and the HAP1 promoter reporter gene with deletions were similarly measured. The UAS1/CYC1-lacZ reporter contains the UAS1 sequence, to which Hap1 binds with high affinity and promotes transcription, and the basal CYC1 promoter sequence −1 to −178 (Guarente and Mason 1983; Guarente et al. 1984). SEMs were calculated from data from at least three independent transformants.
Preparation of yeast extracts and purified GST-Hap1 and electrophoretic mobility shift assays:
Extracts were prepared according to previously established protocols (Zhang and Guarente 1994b; Zhang et al. 1998). Briefly, yeast cells were grown to OD 0.8, harvested, and resuspended in 3 packed cell volumes of buffer (20 mm Tris, 10 mm MgCl2, 1 mm EDTA, 10% glycerol, 1 mm DTT, 0.3 m NaCl, 1 mm phenylmethylsulfonyl fluoride, 1 μg/ml pepstatin, and 1 μg/ml leupeptin). Cells bearing the GAL1-driven HAP1 expression vector (Turcotte and Guarente 1992) were induced by 2% galactose for 5–6 hr prior to collection, as described (Hach et al. 1999). Cells were then permeabilized by agitation with 4 packed cell volumes of glass beads, and extracts were collected. Protein concentrations were subsequently determined by Bradford assay.
The GST-Hap1 fusion protein (Figure 3) containing GST residues 1–222 and Hap1 residues 1–171 was expressed and purified from Escherichia coli (Zhang and Guarente 1994c). The construction of the expression plasmid was described previously (Zhang and Guarente 1994c), and its purification was carried out exactly as described (Zhang and Guarente 1994c). DNA-binding reactions were carried out in a 20-μl volume with 5% glycerol, 4 mm Tris, pH 8, 40 mm NaCl, 4 mm MgCl2, 10 mm DTT, 1.5 μg salmon sperm DNA, 10 μm ZnOAc2, and 300 μg/ml BSA as described (Zhang and Guarente 1994b; Zhang et al. 1998; Hon et al. 2000). Approximately 0.01 pmol of labeled DNA and 20 μg of protein extracts were used in each reaction. The Hap1 antibodies were included in reaction mixtures as described previously (Zhang et al. 1993). The reaction mixtures were incubated at 4° for 1 hr and then loaded onto 4% polyacrylamide gels in 1/3 TBE for gel electrophoresis at 4°. Radioactivity of the interested bands was visualized and quantified by using the PhosphoImage system (Molecular Dynamics, Sunnyvale, CA).
Figure 3.—

Hap1 binding to the HAP1 promoter region encompassing nucleotide sequence −50 to −300. Extracts prepared from Δhap1 cells with the HAP1 gene deleted (lane 1) or from cells expressing high levels of Hap1 (lanes 2–7), or the purified GST-Hap1 protein (100 nm, lanes 9 and 10) were incubated with radiolabeled DNA encompassing nucleotide sequence −50 to −300 of the HAP1 promoter, in the absence (lanes 1 and 2) or presence of preimmune serum (lane 3), Hap1 antibodies (lane 4), unlabeled DNA containing high-affinity Hap1-binding site 7/1 (lanes 5 and 10), unlabeled DNA encompassing nucleotide sequence −1 to −461 of the HAP1 promoter (lane 6), or unlabeled DNA encompassing nucleotide sequence −1 to −380 of the HAP1 promoter (lane 7). The free DNA probe is shown in lane 8. The reaction mixtures were analyzed on a 4% nondenaturing polyacrylamide gel. The positions of the Hap1-DNA complex (Hap1 and GST-Hap1) and the bands representing free probe (Free) are marked. The purified GST-Hap1 fusion protein contains the GST residues 1–222 (26 kD) and the Hap1 residues 1–171 (Zhang and Guarente 1994c) and is smaller than full-length Hap1 (residues 1–1483) expressed in yeast cells. Thus, the GST-Hap1-DNA complex migrated faster than the Hap1-DNA complex in the gel.
Chromatin immunoprecipitation assays:
Chromatin immunoprecipitation (ChIP) was carried out as described previously (Orlando 2000; Iyer et al. 2001). Briefly, yeast MHY100 cells bearing the high-level expression plasmid for Hap1 (under the control of the GAL1/10 promoter and marked by URA3) were grown to OD 0.6–0.8 under heme-deficient or heme-sufficient conditions. Cells were collected and fixed with formaldehyde and then resuspended in lysis buffer containing 50 mm Hepes KOH, pH 7.5, 140 mm NaCl, 1 mm EDTA, 1% Triton X-100, 0.1% Na-deoxycholate, 1 mm PMSF, 10 μg/ml aprotinin, 1 μg/ml leupeptin, 1 μg/ml pepstatin, 1 mm benzamidine, and 1× protease inhibitors (Roche Biochemicals, Indianapolis). Cells were permeabilized by agitation with equal volumes of glass beads. Permeabilized cells were sheared by using a sonicator. Cell extracts were collected and incubated with magnetic beads (Dynabeads M-280 sheep anti-rabbit IgG) bound with anti-Hap1 antibodies. Bound complexes were eluted by using elution buffer containing 50 mm Tris-HCl pH 8, 10 mm EDTA, and 1% SDS. Subsequently, crosslinked complexes were reversed by incubating with 1% SDS at 65° overnight. DNA was subsequently collected and subjected to PCR analysis by using the desired primer pairs for different genes.
Microarray analysis of DNA pulled down by chromatin immunoprecipitation:
Yeast MHY100 cells bearing the Hap1 expression plasmid under the control of the GAL1/10 promoter were grown under high- and low-heme conditions and were used for chromatin immunoprecipitation, as described above. To determine the genome-wide binding distribution of Hap1 by ChIP-chip, we amplified and labeled the ChIP samples with Cy5 as described before (Iyer et al. 2001; Hahn et al. 2004). Genomic DNA from the same strain was labeled in parallel with Cy3 and used as the reference sample. Fluorescently labeled ChIP samples were simultaneously hybridized to whole-genome yeast arrays containing nearly all ORFs and intergenic sequences as described previously (Iyer et al. 2001; Hahn et al. 2004). Microarrays were scanned with a GenePix 4000B scanner (Axon Instruments, Foster City, CA), and fluorescent intensities were quantified by using GenePix pro software (version 4). Data were normalized and filtered, and spots with obvious defects or with signals below an empirically determined threshold were excluded. Further data analyses were performed by using the Longhorn Array database (Killion et al. 2003). The Cy5-to-Cy3 fluorescent signal ratio of each locus represents the fold of enrichment. The ChIP enrichment score E for each locus is calculated as follows: E = median R(Cy5/Cy3)1, R(Cy5/Cy3)2, R(Cy5/Cy3)3 … R(Cy5/Cy3)n. R(Cy5/Cy3)i is the percentile rank of the Cy5-to-Cy3 ratio of each locus among all genomic loci in the ith independent ChIP experiment (Killion et al. 2003; Kim and Iyer 2004). Those with enrichment scores of >96% (median percentile ranking) are considered significant.
RESULTS
Hap1 represses the transcription of its own gene by >20-fold:
To determine whether Hap1 represses the transcription of its own gene, we constructed a reporter plasmid by inserting the promoter sequence (−461 to −1) of the HAP1 gene [see Saccharomyces Genome database (SGD)] into the pLG178 basal reporter plasmid at the N terminus of the β-galactosidase-coding lacZ gene (Guarente et al. 1984). In Δhap1 cells with the HAP1 gene deleted, the HAP1-lacZ reporter activity was >20-fold higher than that in the corresponding wild-type cells (Table 1). For controls, we showed that the activity of the Hap1-activated UAS1/CYC1-lacZ reporter was high in wild-type cells, but low in Δhap1 cells, as expected. This result shows that Hap1 greatly represses the transcription of the HAP1 gene. To determine whether the repression of the HAP1 gene by Hap1 depends on heme levels, we detected the HAP1-lacZ reporter activity in Δhem1 and Δhem1 Δhap1 cells grown under heme-deficient and heme-sufficient conditions (Table 1). We found that regardless of heme levels, the HAP1-lacZ reporter activity was low (repressed) in Δhem1 cells, but high (derepressed) in Δhem1 Δhap1 cells (Table 1). As expected, the activity of the Hap1-activated UAS1/CYC1-lacZ reporter was high in wild-type cells or in Δhem1 cells grown at high heme levels, but was very low in Δhap1 cells or in Δhem1 cells grown at low heme levels or in Δhem1 Δhap1 cells grown at both low and high heme levels (Table 1). These results show that the repression of the HAP1 gene is dependent on Hap1, but independent of heme levels.
Hap1 DNA binding is important for transcriptional repression by Hap1:
To better understand the mechanism of transcriptional repression by Hap1, we asked whether Hap1 domains critical for transcriptional activation and heme regulation are also important for transcriptional repression by Hap1. We measured and compared the repression activity of wild-type Hap1, the Hap1-18 mutant, and Hap1 proteins with various domains deleted (Figure 1A; Kim and Guarente 1989; Pfeifer et al. 1989; Hach et al. 1999). All these Hap1 proteins are shown to be stably expressed at high levels in yeast (Pfeifer et al. 1987; Hach et al. 1999; Figure 1B). Notably, mutant Hap1-18 was largely defective in the repression of the HAP1 promoter (Figure 1, Hap1-18). Hap1-18 contains a Ser-to-Arg mutation at amino acid position 63 in the zinc cluster motif (Kim and Guarente 1989; Turcotte and Guarente 1992); it binds only to the UAS/CYC7 site (CGCTATTATCGCTATTA), not the typical Hap1-binding sites (optimal: CGGnnnTAnCGG) (Zhang and Guarente 1994c). Therefore, this result from Hap1-18 shows that Hap1 DNA binding is necessary for transcriptional repression by Hap1.
The Hap1 heme domain has little effect on transcriptional repression by Hap1:
Hap1 domains critical for transcriptional activation and for heme regulation exerted varying effects on the repression activity of Hap1 (Figure 1). Notably, deletion of the heme domain, containing both the repression module RPM1 and heme-responsive motifs HRM1–6 (Figure 1, Δheme), virtually did not affect the repression activity of Hap1. This result, together with those in Table 1, demonstrates that the repression function of Hap1, unlike its activation function, is completely heme independent.
Interestingly, the deletion of the Hap1 activation domain caused a fivefold derepression of the HAP1 promoter (Figure 1A, HK). As the deleted region enlarges in HB and HG, Hap1 repression capacity further diminishes (Figure 1A). Due to lack of the activation domain, these deletion proteins were inactive at the UAS1/CYC1 site mediating transcriptional activation by Hap1 (see Figure 1, the UAS1/CYC1 column). The H7-d1 and H7-d2 deletion proteins, which cause Hap1 derepression in the absence of heme and exhibit higher transcriptional activity at UAS1/CYC1 (Hach et al. 1999; see also Figure 1, the UAS1/CYC1 column), were partially defective in Hap1 transcriptional repression. The smHap1 mutant protein (Figure 1), which is completely transcriptionally inactive and heme inactivable (Lee et al. 2003), completely lost its ability to repress the HAP1 gene. The data show that the Hap1 activation domain is required not only for transcriptional activation, but also for repression, although to a lesser extent. It appears that other than the DNA-binding domain, the activation domain and other internal domains of Hap1 all contribute to transcriptional repression, while the activation domain is both necessary and sufficient for transcriptional activation, as shown previously (Pfeifer et al. 1989).
Hap1 appears to bind to the HAP1 promoter and promotes repression directly:
To better understand how Hap1 represses transcription of the HAP1 gene, we tried to map the DNA site responsible for transcriptional repression. To this end, we constructed three 5′-end deletions (Figure 2, H1, H2, and H3), one internal deletion (H4), and one 3′-end deletion (C1) of the HAP1 promoter. The basal promoter elements for the 3′-end deletions are supplied by using the basal promoter of the CYC1 gene. We were unable to construct a reporter containing the complete HAP1 promoter and the basal CYC1 promoter even after repeated attempts, very likely because the repeated AT-rich regions in these promoters cause the construct to be unstable in E. coli.
Nonetheless, the deletion analysis provides important clues about transcriptional repression by Hap1 (Figure 2). The results show that a UAS is likely located around the nucleotide −380 region, because reporter constructs lacking sequences upstream or downstream of −380 were inactive in cells expressing Hap1 (Figure 2, HAP1) and in cells with the HAP1 gene deleted (Figure 2, Δhap1). It is not clear whether this region contains the DNA site mediating repression, because the absence of a UAS makes it impossible to identify the site by derepression in cells lacking Hap1. However, reporter construct C1 lacking sequence from nucleotide −150 to −1 exhibited high activity even in cells expressing wild-type Hap1, suggesting that the −150 to −1 region contains part of the or the whole site mediating repression by Hap1 (Figure 2). The site mediating Hap1 repression may overlap with sequence beyond −150 (the reporter C1 may contain part of the repression site and still be derepressed), but should not overlap the coding region beyond −1. These results show that the HAP1 promoter contains a DNA site responsible for transcriptional repression.
The HAP1 promoter does not have a Hap1-binding site containing an indirect repeat of two CGG/CGC triplets separated by a 6-bp spacer, which is found in promoters that are activated by Hap1 (Zhang and Guarente 1994c). However, the promoter contains at least 10 CGG/CGC half-sites (sequence from the SGD). These sites can make contacts with a Hap1 monomer or a subunit of a Hap1 dimer or oligomer in a repression complex with another protein partner. To ascertain whether Hap1 actually binds to the HAP1 promoter directly, we performed electrophoretic mobility shift assays. We generated radiolabled and unlabeled DNA fragments containing various regions of the HAP1 gene promoter, as described previously (Zhang and Guarente 1994c). All DNA fragments containing sequence between −300 and −50 showed at least one Hap1-bound complex when incubated with extracts prepared from cells expressing Hap1. Particularly, the DNA fragment containing −300 to −50 of the HAP1 promoter revealed a specific Hap1-binding activity by wild-type Hap1 in extracts or by purified GST-Hap1 (residues 1–171) fusion protein (Zhang and Guarente 1994c; see Figure 3).
As shown in Figure 3, a specific Hap1-DNA complex was formed when radiolabled −300 to −50 fragments were incubated with extracts prepared from cells expressing Hap1 (see Figure 3, lane 2). This band was largely disrupted by addition of Hap1 antibodies (Figure 3, compare lanes 4 and 2), but not the preimmune serum (Figure 3, lane 3; Zhang et al. 1993). Quantification showed that the intensity of the band was reduced by fivefold by antibodies but slightly increased by preimmune serum. This band was also titrated out fully by unlabeled DNA fragments containing the typical high-affinity Hap1-binding site 7/1 (Figure 3, lane 5) or by unlabeled DNA fragments containing the promoter sequence (Figure 3, lanes 6 and 7). The band intensity was reduced >20-fold by the competitor DNA. For a control, we showed that no specific protein binding was detected when extracts prepared from Δhap1 cells were used in the assay (see Figure 3, lane 1). Notably, a specific Hap1-DNA complex was also formed when the −300 to −50 fragment was incubated with a purified GST-Hap1 (residues 1–171) fusion protein (see Figure 3, lanes 9 and 10). However, this result showing binding by the purified Hap1 fragment (∼100 nm and at 4°) does not exclude the possibility that non-Hap1 proteins cobind with Hap1 at the HAP1 promoter in extracts or in vivo.
Further, by chromatin immunoprecipitation assays (Orlando 2000; Iyer et al. 2001) with purified anti-Hap1 antibodies (Figure 4; Zhang et al. 1998), coupled to DNA microarray hybridization (ChIP-chip), we showed that Hap1 is associated with the promoter of the HAP1 gene preferentially in vivo. Chromatin-immunoprecipitated DNAs were obtained from a strain where Hap1 is expressed from the GAL1/10 promoter. Chromatin-immunoprecipitated DNAs were first randomly amplified and then labeled with Cy5. Genomic DNAs from the same cells were amplified and labeled with Cy3 as a reference. The immunoprecipitated and reference DNAs were simultaneously hybridized to whole-genome arrays containing both ORFs and intergenic sequences. The microarrays were scanned, and data analyses were performed by using the Longhorn Array database (Killion et al. 2003). The fold of enrichment for each locus was obtained by calculating the ratio of Cy5 to Cy3 fluorescent intensities (red/green) at the corresponding locus in the ChIP experiments. To identify the most strongly and consistently enriched loci we determined the median percentile ranks (enrichment scores) of ChIP enrichment across multiple (nine in this case) independent replicate experiments as described before (Hahn et al. 2004; Kim and Iyer 2004). Within the top 2% of enriched loci across nine independent ChIP-chip experiments, we found that the two loci (Figure 4, iYLR255C and YLR256W) that contain part of the HAP1 gene were consistently immunoprecipitated. The fold of enrichment for the YLR256W, iYLR255C, and iYLRWdelta loci, shown in Figure 4, was 5.1±1.6, 3.5±0.9, and 4.8±2.2, respectively. The fold of enrichment for the CYC1 locus, which binds to Hap1 with high affinity and is strongly activated by Hap1, was 5.8±2.4. Several other known Hap1-regulated genes, including CYT1 (Schneider and Guarente 1991), MOT3 (Sertil et al. 2003), and COX15 (Choi et al. 1996), were also found to be repeatedly enriched in the ChIP-chip experiments. Also, PCR amplification of immunoprecipitated DNAs by Hap1 antibodies with primers flanking the HAP1 promoter region repeatedly generated the expected product while no specific product was detected if primers flanking the coding sequence of the SSA1 gene were used or if Hap1 was not expressed. Quantification showed that the level of PCR product from the HAP1 genes was at least 15-fold higher than the background level from PCR amplification of the SSA1 gene. These results strongly support the idea that Hap1 can directly bind to its own promoter in vivo. Together with data in Figures 1–3, these results demonstrate that Hap1 binds specifically to DNA in the HAP1 promoter region mediating transcriptional repression.
Figure 4.—

ChIP-chip analysis indicates that the promoter DNA of the HAP1 gene is preferentially associated with Hap1 in vivo. ChIP-chip analysis was performed as described in materials and methods. Shown is the genomic sequence map around the HAP1 locus. The boxes represent the sequences that are preferentially detected by coimmunoprecipitated DNAs with Hap1 antibodies. Three sequences, iYLR255C encompassing the promoter region of the HAP1 gene, YLR256W encompassing the coding sequence of the HAP1 gene, and iYLRWdelta14, which is distal to the HAP1 gene, are found to be preferentially enriched by coimmunoprecipitation.
Hsp70 (Ssa), not Hsp90, is important for transcriptional repression by Hap1:
To further compare the mechanisms by which Hap1 represses and activates transcription, we asked whether repression by Hap1 requires the functions of Hsp90 and Hsp70, which are involved in transcriptional activation by Hap1 and heme regulation of Hap1 activity (Zhang et al. 1998; Hon et al. 2001; Lee et al. 2002). We measured the HAP1 promoter activity in cells with the Hsp70 (Ssa)-coding genes SSA2, SSA3, and SSA4 deleted (Figure 5A, a2a3a4 cells; Becker et al. 1996) and in cells expressing a low level of Hsp90 from a truncated promoter (Figure 5B, iLEP cells; Chang and Lindquist 1994). The a2a3a4 cells express about one-third of Ssa proteins expressed in wild-type cells (Becker et al. 1996). In these cells, Hap1 gained low levels of transcription-activating activity in the absence of heme (Hon et al. 2001). The data in Figure 5A show that the HAP1 promoter-lacZ reporter activity was increased by about ninefold in a2a3a4 cells. As controls, we show that the activities of the Hap2/3/4/5-driven UAS2UP1-lacZ (Forsburg and Guarente 1989) and HIS4-lacZ (Hinnebusch 1984) reporters were largely unaffected in a2a3a4 cells, compared to that in wild-type cells (Figure 5A). As shown previously (Hon et al. 2001), the activity of the Hap1-activated UAS1/CYC1-lacZ reporter in a2a3a4 cells (heme sufficient) was largely unaffected. These results show that Ssa function is specifically required for the repression of the HAP1 gene promoter.
Figure 5.—

The effect of Ssa (A) and Hsp90 (B) on the repression of the HAP1 promoter by Hap1. Yeast cells with SSA2, SSA3, and SSA4 genes deleted (a2a3a4); cells expressing a low level of Hsp90 (iLEP); and their wild-type counterparts were transformed with the full-length HAP1 promoter-lacZ reporter plasmid. Cells were grown in selective medium, and β-galactosidase activities were detected. The plotted data are averages from at least three independent transformants. For controls, the activities of the Hap2/3/4/5-driven UAS2UP1-lacZ (Forsburg and Guarente 1989), HIS4-lacZ (Hinnebusch 1984), and UAS1/CYC1-lacZ (Guarente et al. 1984; Turcotte and Guarente 1992) reporters in a2a3a4 cells were also measured and shown. The activities of the Hap2/3/4/5-driven UAS2UP1-lacZ (Forsburg and Guarente 1989) and HIS4-lacZ (Hinnebusch 1984) in the iLEP strain have been shown previously (Zhang et al. 1998). Note that the strains used in A and B are derived from different laboratories and have different genotypes (see materials and methods). This difference likely accounts for the fourfold difference in the HAP1-lacZ reporter activity in the wild-type control strains in A (JN55) and B (W303).
The HAP1 reporter activity in iLEP cells expressing a low level of Hsp90 was reduced less than twofold. This reduction is likely attributable to the general effect of low levels of Hsp90 on cell growth and reporter activity (Zhang et al. 1998; Lee et al. 2002). Previous results show that the activities of various reporters, including the Hap2/3/4/5-driven UAS2UP1-lacZ and HIS4-lacZ reporters, are generally reduced two- to threefold in cells expressing low levels of Hsp90 (Zhang et al. 1998; Lee et al. 2002). For comparison, we showed that the activity of the Hap1-activated UAS1/CYC1-lacZ reporter was reduced by more than fivefold in iLEP cells, as was shown previously (Lee et al. 2002). Thus, the result in Figure 5B suggests that Hap1 repression function is likely not affected specifically by Hsp90.
DISCUSSION
In this report, we show that Hap1 binds to and represses directly the transcription of its own HAP1 gene, that the repression function of Hap1 is totally independent of heme and heme regulation, and that Ssa plays a role in transcriptional repression by Hap1. One may argue that Hap1 represses transcription by activating the expression of a repressor. If so, repression by Hap1 should occur only when Hap1 transcriptional activity is activated. Our data strongly argue against such an idea because Hap1 represses HAP1 in heme-deficient cells (Table 1). Hap1, when expressed from the chromosome or its own promoter, cannot activate transcription in heme-deficient cells (Zhang and Guarente 1994b). Thus, it is unlikely that the repression function of Hap1 derives from another repressor whose expression is activated by Hap1. Further, our in vitro DNA-binding analysis, ChIP-chip, and in vivo functional data show that Hap1 binds to the HAP1 promoter directly and promotes its repression.
How does Hap1 repress transcription? The mechanism by which Hap1 represses transcription is distinct from its mechanism of transcriptional activation, while they may share common characteristics. First, the repression function of Hap1 is heme independent whereas the activation function of Hap1 is heme dependent. This does not necessarily suggest that the repression function of Hap1 is irrelevant to transcriptional activation by Hap1. The repression function of Hap1 may greatly enhance its ability to precisely control transcription. For example, repression may be the default function in heme-deficient cells and may be part of heme regulation. By transforming from a repressor in the absence of heme to an activator in the presence of heme, Hap1 may exert a much greater degree of control on transcription than a simple activator, as in the case of ROX1 expression (Zhang and Hach 1999).
Second, the Hap1 activation domain is also required for its repressing function although its role is less dominant in repression than in activation. In addition to the DNA-binding domain, the Hap1 activation domain is both necessary and sufficient for transcriptional activation (Pfeifer et al. 1989). Repression by Hap1 appears to depend not only on the activation domain, but also on the internal domains (Figure 1). No single domain appears to play a dominant role in repression (Figure 1). Rather, all Hap1 domains appear to act together to exert a collective control over its repression function. Nevertheless, the finding that the Hap1 activation domain plays a role in both transcriptional activation and repression is intriguing. Recent studies indicate that certain coactivator complexes may also have a repressing function. For example, the Swi/Snf coactivator complex can directly repress transcription, although the requirement for specific components may differ in repression, compared to activation (Martens and Winston 2002, 2003). Thus, it is conceivable that the Hap1 activation domain is necessary to recruit its coactivator complex, while it can also help recruit its corepressor complex, because the coactivator and corepressor share a common component(s). Consistent with this idea, previous experiments (Ha et al. 2000) suggest that Swi2 plays a positive role in transcriptional activation by Hap1. Thus, the Hap1 activation domain may help recruit coactivator and corepressor complexes like the Swi/Snf complexes, thereby promoting both transcriptional activation and repression.
Third, the requirement for protein partners for repression and activation is different. While Hsp90 is critical for transcriptional activation by Hap1 (Zhang et al. 1998; Lee et al. 2002), it does not appear to be important for repression by Hap1. Ssa, however, appears to cooperate with Hap1 in transcriptional repression. This role of Ssa is consistent with its role in general protein folding and in the inactivation of Hap1 transcription-activating function in the absence of heme (Hon et al. 2001; Hartl and Hayer-Hartl 2002).
In addition to Ssa, Hap1 may require one more protein for repressing transcription. This protein likely cooperates with Hap1 in DNA binding and perhaps in repression. The HAP1 promoter does not contain a full Hap1-binding site for a Hap1 dimer (a direct repeat of two CGG/CGC triplets with a 6-bp spacer; Zhang and Guarente 1994c), but it contains many single CGG/CGCs that can make contacts with a Hap1 monomer or a subunit of a Hap1 dimer/oligomer. Either CGG or CGC may be involved in Hap1 binding, because Hap1 can bind to both triplets well (Pfeifer et al. 1987; Hon et al. 1999; King et al. 1999) and because Hap1-18 also conferred a slight (less than twofold) repression on the HAP1 promoter (Figure 1). Hap1 monomer-DNA contacts may be an essential part of a Hap1-DNA repression complex. Indeed, purified GST-Hap1 (residues 1–171; Figure 3), at a concentration of ∼100 nm and at 4°, can bind to the HAP1 promoter in vitro. However, this result does not exclude the possibility that in vivo, and even in extracts, Hap1 binds to the HAP1 promoter cooperatively with another corepressor. This corepressor may contact DNA and may bind to DNA cooperatively to enhance the DNA-binding affinity of the repressor complex. Hap1 may repress transcription by many mechanisms. For example, like glucocorticoid receptor (GR; Miner et al. 1991), Hap1 may bind to composite DNA sites with other factor(s) and thereby repress transcription. Alternatively, Hap1 may recruit a transcriptional repressor and thereby repress transcription. Or Hap1 may displace binding of another activator.
The repression function of Hap1 is likely to be important for many genes other than HAP1. Recent gene expression analyses show that >100 genes (Lombardia et al. 2000), including ANB1 and HEM13, exhibit increased transcript levels in cells with HAP1 deleted, compared to wild-type cells (also T. Hon and L. Zhang, unpublished results). Although the effect of Hap1 on both ANB1 and HEM13 appears to be indirect (Zitomer et al. 1997; Mennella et al. 2003), Hap1 may bind to and repress directly transcription of at least some of these genes. The mechanism of transcriptional repression by Hap1 may have general implications in understanding the regulation of numerous genes in yeast. The mechanisms by which Hap1 activates and represses transcription may provide insights into the mechanisms by which many other characterized and uncharacterized transcriptional regulators, such as Rap1 (Morse 2000), activate and repress transcription.
Acknowledgments
We are grateful to Dr. S. L. Lindquist and Dr. E. A. Craig for providing yeast strains. This work was supported by funds from the National Institutes of Health (GM62246 to L.Z.).
References
- Becker, J., W. Walter, W. Yan and E. A. Craig, 1996. Functional interaction of cytosolic hsp70 and a DnaJ-related protein, Ydj1p, in protein translocation in vivo. Mol. Cell. Biol. 16: 4378–4386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buisson, N., and R. Labbe-Bois, 1998. Flavohemoglobin expression and function in Saccharomyces cerevisiae. No relationship with respiration and complex response to oxidative stress. J. Biol. Chem. 273: 9527–9533. [DOI] [PubMed] [Google Scholar]
- Chang, H. C., and S. Lindquist, 1994. Conservation of Hsp90 macromolecular complexes in Saccharomyces cerevisiae. J. Biol. Chem. 269: 24983–24988. [PubMed] [Google Scholar]
- Choi, J. Y., J. Stukey, S. Y. Hwang and C. E. Martin, 1996. Regulatory elements that control transcription activation and unsaturated fatty acid-mediated repression of the Saccharomyces cerevisiae OLE1 gene. J. Biol. Chem. 271: 3581–3589. [DOI] [PubMed] [Google Scholar]
- Creusot, F., J. Verdiere, M. Gaisne and P. P. Slonimski, 1988. CYP1 (HAP1) regulator of oxygen-dependent gene expression in yeast. I. Overall organization of the protein sequence displays several novel structural domains. J. Mol. Biol. 204: 263–276. [DOI] [PubMed] [Google Scholar]
- Deckert, J., R. Perini, B. Balasubramanian and R. S. Zitomer, 1995. Multiple elements and auto-repression regulate Rox1, a repressor of hypoxic genes in Saccharomyces cerevisiae. Genetics 139: 1149–1158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Defranoux, N., M. Gaisne and J. Verdière, 1994. Functional analysis of the zinc cluster domain of the CYP1 (HAP1) complex regulator in heme-sufficient and heme-deficient yeast cells. Mol. Gen. Genet. 242: 699–707. [DOI] [PubMed] [Google Scholar]
- Forsburg, S. L., and L. Guarente, 1989. Identification and characterization of HAP4: a third component of the CCAAT-bound HAP2/HAP3 heteromer. Genes Dev. 3: 1166–1178. [DOI] [PubMed] [Google Scholar]
- Guarente, L., and T. Mason, 1983. Heme regulates transcription of the CYC1 gene of S. cerevisiae via an upstream activation site. Cell 32: 1279–1286. [DOI] [PubMed] [Google Scholar]
- Guarente, L., B. Lalonde, P. Gifford and E. Alani, 1984. Distinctly regulated tandem upstream activation sites mediate catabolite repression of the CYC1 gene of S. cerevisiae. Cell 36: 503–511. [DOI] [PubMed] [Google Scholar]
- Ha, N., K. Hellauer and B. Turcotte, 2000. Fusions with histone H3 result in highly specific alteration of gene expression. Nucleic Acids Res. 28: 1026–1035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hach, A., T. Hon and L. Zhang, 1999. A new class of repression modules is critical for heme regulation of the yeast transcriptional activator Hap1. Mol. Cell. Biol. 19: 4324–4333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hahn, J. S., Z. Hu, D. J. Thiele and V. R. Iyer, 2004. Genome-wide analysis of the biology of stress responses through heat shock transcription factor. Mol. Cell. Biol. 24: 5249–5256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haldi, M., and L. Guarente, 1989. N-terminal deletions of a mitochondrial signal sequence in yeast. Targeting information of delta-aminolevulinate synthase is encoded in non-overlapping regions. J. Biol. Chem. 264: 17107–17112. [PubMed] [Google Scholar]
- Haldi, M. L., and L. Guarente, 1995. Multiple domains mediate heme control of the yeast activator HAP1. Mol. Gen. Genet. 248: 229–235. [DOI] [PubMed] [Google Scholar]
- Hartl, F. U., and M. Hayer-Hartl, 2002. Molecular chaperones in the cytosol: from nascent chain to folded protein. Science 295: 1852–1858. [DOI] [PubMed] [Google Scholar]
- Hinnebusch, A. G., 1984. Evidence for translational regulation of the activator of general amino acid control in yeast. Proc. Natl. Acad. Sci. USA 81: 6442–6446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hon, T., A. Hach, D. Tamalis, Y. Zhu and L. Zhang, 1999. The yeast heme-responsive transcriptional activator Hap1 is a preexisting dimer in the absence of heme. J. Biol. Chem. 274: 22770–22774. [DOI] [PubMed] [Google Scholar]
- Hon, T., A. Hach, H. C. Lee, T. Chen and L. Zhang, 2000. Functional analysis of heme regulatory elements of the transcriptional activator Hap1. Biochem. Biophys. Res. Commun. 273: 584–591. [DOI] [PubMed] [Google Scholar]
- Hon, T., H. C. Lee, A. Hach, J. L. Johnson, E. A. Craig et al., 2001. The Hsp70-Ydj1 molecular chaperone represses the activity of the transcriptional activator Hap1 in the absence of heme. Mol. Cell. Biol. 21: 7923–7932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iyer, V. R., C. E. Horak, C. S. Scafe, D. Botstein, M. Snyder et al., 2001. Genomic binding sites of the yeast cell-cycle transcription factors SBF and MBF. Nature 409: 533–538. [DOI] [PubMed] [Google Scholar]
- Killion, P. J., G. Sherlock and V. R. Iyer, 2003. The Longhorn Array Database (LAD): an open-source, MIAME compliant implementation of the Stanford Microarray Database (SMD). BMC Bioinformatics 4: 32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, J., and V. R. Iyer, 2004. Global role of TATA box-binding protein recruitment to promoters in mediating gene expression profiles. Mol. Cell. Biol. 24: 8104–8112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, K. S., and L. Guarente, 1989. Mutations that alter transcriptional activation but not DNA binding in the zinc finger of yeast activator HAPI. Nature 342: 200–203. [DOI] [PubMed] [Google Scholar]
- King, D. A., L. Zhang, L. Guarente and R. Marmorstein, 1999. Structure of a HAP1/DNA complex reveals dramatically asymmetric DNA binding by a homodimeric protein. Nat. Struct. Biol. 6: 64–71. [DOI] [PubMed] [Google Scholar]
- Lee, H. C., T. Hon and L. Zhang, 2002. The Hsp90 molecular chaperone mediates heme activation of the yeast transcriptional activator Hap1. J. Biol. Chem. 277: 7430–7437. [DOI] [PubMed] [Google Scholar]
- Lee, H. C., T. Hon, C. Lan and L. Zhang, 2003. Structural environment dictates the biological significance of heme-responsive motifs and the role of Hsp90 in the activation of the heme activator protein Hap1. Mol. Cell. Biol. 23: 5857–5866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lodi, T., and B. Guiard, 1991. Complex transcriptional regulation of the Saccharomyces cerevisiae CYB2 gene encoding cytochrome b2: CYP1(HAP1) activator binds to the CYB2 upstream activation site UAS1–B2. Mol. Cell. Biol. 11: 3762–3772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lombardia, L. J., J. L. Cadahia-Rodriguez, M. A. Freire-Picos, M. I. Gonzalez-Siso, A. M. Rodriguez-Torres et al., 2000. Transcript analysis of 203 novel genes from Saccharomyces cerevisiae in hap1 and rox1 mutant backgrounds. Genome 43: 881–886. [DOI] [PubMed] [Google Scholar]
- Lowry, C. V., and R. S. Zitomer, 1988. ROX1 encodes a heme-induced repression factor regulating ANB1 and CYC7 of Saccharomyces cerevisiae. Mol. Cell. Biol. 8: 4651–4658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martens, J. A., and F. Winston, 2002. Evidence that Swi/Snf directly represses transcription in S. cerevisiae. Genes Dev. 16: 2231–2236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martens, J. A., and F. Winston, 2003. Recent advances in understanding chromatin remodeling by Swi/Snf complexes. Curr. Opin. Genet. Dev. 13: 136–142. [DOI] [PubMed] [Google Scholar]
- Mennella, T. A., L. G. Klinkenberg and R. S. Zitomer, 2003. Recruitment of Tup1-Ssn6 by yeast hypoxic genes and chromatin-independent exclusion of TATA binding protein. Eukaryot. Cell 2: 1288–1303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miner, J. N., M. I. Diamond and K. R. Yamamoto, 1991. Joints in the regulatory lattice: composite regulation by steroid receptor-AP1 complexes. Cell Growth Differ. 2: 525–530. [PubMed] [Google Scholar]
- Morse, R. H., 2000. RAP, RAP, open up! New wrinkles for RAP1 in yeast. Trends Genet. 16: 51–53. [DOI] [PubMed] [Google Scholar]
- Orlando, V., 2000. Mapping chromosomal proteins in vivo by formaldehyde-crosslinked-chromatin immunoprecipitation. Trends Biochem. Sci. 25: 99–104. [DOI] [PubMed] [Google Scholar]
- Pfeifer, K., T. Prezant and L. Guarente, 1987. Yeast HAP1 activator binds to two upstream activation sites of different sequence. Cell 49: 19–27. [DOI] [PubMed] [Google Scholar]
- Pfeifer, K., K. S. Kim, S. Kogan and L. Guarente, 1989. Functional dissection and sequence of yeast HAP1 activator. Cell 56: 291–301. [DOI] [PubMed] [Google Scholar]
- Pinkham, J. L., Z. Wang and J. Alsina, 1997. Heme regulates SOD2 transcription by activation and repression in Saccharomyces cerevisiae. Curr. Genet. 31: 281–291. [DOI] [PubMed] [Google Scholar]
- Schneider, J. C., and L. Guarente, 1991. Regulation of the yeast CYT1 gene encoding cytochrome c1 by HAP1 and HAP2/3/4. Mol. Cell. Biol. 11: 4934–4942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sertil, O., R. Kapoor, B. D. Cohen, N. Abramova and C. V. Lowry, 2003. Synergistic repression of anaerobic genes by Mot3 and Rox1 in Saccharomyces cerevisiae. Nucleic Acids Res. 31: 5831–5837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turcotte, B., and L. Guarente, 1992. HAP1 positive control mutants specific for one of two binding sites. Genes Dev. 6: 2001–2009. [DOI] [PubMed] [Google Scholar]
- Ushinsky, S. C., and T. Keng, 1994. A novel allele of HAP1 causes uninducible expression of HEM13 in Saccharomyces cerevisiae. Genetics 136: 819–831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verdiere, J., M. Gaisne and B. R. Labbe, 1991. CYP1 (HAP1) is a determinant effector of alternative expression of heme-dependent transcribed genes in yeast. Mol. Gen. Genet. 228: 300–306 (Erratum: Mol. Gen. Genet. 229 (2): 324). [DOI] [PubMed] [Google Scholar]
- Winkler, H., G. Adam, E. Mattes, M. Schanz, A. Hartig et al., 1988. Co-ordinate control of synthesis of mitochondrial and non-mitochondrial hemoproteins: a binding site for the HAP1 (CYP1) protein in the UAS region of the yeast catalase T gene (CTT1). EMBO J. 7: 1799–1804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, L., and L. Guarente, 1994. a Evidence that TUP1/SSN6 has a positive effect on the activity of the yeast activator HAP1. Genetics 136: 813–817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, L., and L. Guarente, 1994. b HAP1 is nuclear but is bound to a cellular factor in the absence of heme. J. Biol. Chem. 269: 14643–14647. [PubMed] [Google Scholar]
- Zhang, L., and L. Guarente, 1994. c The yeast activator HAP1—a GAL4 family member—binds DNA in a directly repeated orientation. Genes Dev. 8: 2110–2119. [DOI] [PubMed] [Google Scholar]
- Zhang, L., and L. Guarente, 1996. The C6 zinc cluster dictates asymmetric binding by HAP1. EMBO J. 15: 4676–4681. [PMC free article] [PubMed] [Google Scholar]
- Zhang, L., and A. Hach, 1999. Molecular mechanism of heme signaling in yeast: the transcriptional activator Hap1 serves as the key mediator. Cell. Mol. Life Sci. 56: 415–426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, L., M. O. Bermingham, B. Turcotte and L. Guarente, 1993. Antibody-promoted dimerization bypasses the regulation of DNA binding by the heme domain of the yeast transcriptional activator HAP1. Proc. Natl. Acad. Sci. USA 90: 2851–2855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, L., A. Hach and C. Wang, 1998. Molecular mechanism governing heme signaling in yeast: a higher-order complex mediates heme regulation of the transcriptional activator HAP1. Mol. Cell. Biol. 18: 3819–3828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao, X. J., D. Raitt, P. V. Burke, A. S. Clewell, K. E. Kwast et al., 1996. Function and expression of flavohemoglobin in Saccharomyces cerevisiae. Evidence for a role in the oxidative stress response. J. Biol. Chem. 271: 25131–25138. [DOI] [PubMed] [Google Scholar]
- Zitomer, R. S., and C. V. Lowry, 1992. Regulation of gene expression by oxygen in Saccharomyces cerevisiae. Microbiol. Rev. 56: 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zitomer, R. S., P. Carrico and J. Deckert, 1997. Regulation of hypoxic gene expression in yeast. Kidney Int. 51: 507–513. [DOI] [PubMed] [Google Scholar]