

Abstract
Several different stoichiometries have been proposed for the Na+/monocarboxylate cotransporter SMCT1, including variable Na+/substrate stoichiometry. In this work, we have definitively established an invariant 2:1 cotransport stoichiometry for SMCT1. By using two independent means of assay, we first showed that SMCT1 exhibits a 2:1 stoichiometry for Na+/lactate cotransport. Radiolabel uptake experiments proved that, unlike lactate, propionic acid diffuses passively through oocyte membranes and, consequently, propionate is a poor candidate for stoichiometric determination by these methods. Although we previously determined SMCT1 stoichiometry by measuring reversal potentials, this technique produced erroneous values, because SMCT1 simultaneously mediates both an inwardly rectifying cotransport current and an outwardly rectifying anionic leak current; the leak current predominates in the range where reversal potentials are observed. We therefore employed a method that compared the effect of halving the external Na+ concentration to the effect of halving the external substrate concentration on zero-current potentials. Both lactate and propionate were cotransported through SMCT1 using 2:1 stoichiometries. The leak current passing through the protein has a 1 osmolyte/charge stoichiometry. Identification of cotransporter stoichiometry is not always a trivial task and it can lead to a much better understanding of the transport activity mediated by the protein in question.
INTRODUCTION
Cotransport proteins achieve the efficient movement of a substrate across a membrane (even against its own electrochemical gradient) by coupling this movement to the transport of an ion, enabling the electrochemical gradient of the ion to act as the driving force behind the cotransport. One of the basic parameters associated with this form of transport is its stoichiometry, i.e., the number of ions associated with the movement of each substrate molecule through the protein. This relationship can generally be described by a simple 2:1 or 1:1 ratio, such as those demonstrated for the Na+/glucose cotransporters SGLT1 (2 Na+/glucose) and SGLT2 (1 Na+/glucose) (1,2). More complex stoichiometric ratios have also been measured for some Na+ cotransporters, often involving Cl− ions (2,3) and some transport proteins have been shown to display variable stoichiometry (4–8).
A number of different techniques have been used to establish cotransport stoichiometries for individual proteins, and they are most often applied when the protein has been heterologously expressed in Xenopus laevis oocytes by injection of mRNA transcribed from the cloned transporter cDNA. This approach provides huge, convenient cells expressing high levels of one defined cotransporter. The most common procedure is to measure radiolabeled substrate uptake into voltage-clamped oocytes, permitting the inferral of stoichiometry by comparing the substrate-induced currents with the associated amount of radiolabel uptake (9). In 1995, we applied a thermodynamic method based on the effect of external substrate concentration on the reversal potential of SGLT1 (10). More recently, we presented a novel method based on concomitant current and volume measurements to establish the stoichiometry of inositol cotransporters (11).
Despite this body of work on transport stoichiometries and the existence of various techniques for measuring these values, the stoichiometry of the Na+/monocarboxylate cotransporter SMCT1 remains unclear several years after the protein function was first identified. Although we determined a 3:1 Na+/propionate stoichiometry by measuring transporter reversal potentials (12), another group identified a 2:1 Na+/lactate stoichiometry and a 4:1 Na+/propionate stoichiometry by combining current and uptake measurements, and concluded that the cotransporter stoichiometry is variable (13,14). Although variable transporter stoichiometries have been demonstrated for some transporters, they are not commonly encountered. This fact and the broad range of stoichiometric ratios published for this transporter suggests that its true stoichiometry remains to be firmly established. Further complicating the measurement of SMCT1 stoichiometry are the strong leak currents seen through this protein (12) (M. J. Coady, B. Wallendorff, and J.-Y. Lapointe, unpublished) and the passive permeation through membranes of some monocarboxylates.
In this study, we have conclusively established that SMCT1 has an invariant 2:1 cotransport stoichiometry for Na+ and monocarboxylates. In addition, we have examined how the method of establishing stoichiometries through reversal potentials can be confounded by cotransport and leak currents exhibiting opposite forms of rectification. Finally, we have established that the leak current through SMCT1 is most likely a purely anionic flux by determining a ratio of 1 net osmolyte transported per charge transported via this current.
MATERIALS AND METHODS
Materials
1-14C-propionic acid was obtained from MP Biomedicals (Aurora, OH); L-14C-U-lactic acid was obtained from Perkin-Elmer (Boston, MA). All other chemicals were purchased from Sigma-Aldrich (St. Louis, MO).
Preparation of mRNA and oocytes
mRNA encoding the SMCT1 cotransporter was transcribed from the vector pT7TS containing the full-length human coding region (12). Transcription of SMCT1 and aquaporin-1 were performed on cleaved templates using the T7 mMessage mMachine kit from Ambion (Austin, TX). Xenopus laevis oocytes (animals obtained from the University of Alberta, Edmonton, Alberta, Canada) were surgically removed under anesthesia (in accordance with animal handling regulations of the Université de Montréal) and individually separated. The follicular layers were removed by collagenase digestion and oocytes were maintained at 18°C in Barth's solution (90 mM NaCl, 3 mM KCl, 0.82 mM MgSO4, 0.41 mM CaCl2, 0.33 mM Ca(NO3)2, 5 mM HEPES, pH 7.6) supplemented with 100 units/ml penicillin and 0.1 mg/ml streptomycin. SMCT1 mRNA (46 nl, 0.25 μg/μl) was injected into the oocytes 1 day after surgical isolation; the oocytes were assayed for transporter activity at 5–8 days after injection. ROMK2 and AQP1 mRNA were utilized as previously described (11).
Steady-state current measurements in oocytes
Oocyte currents were measured using a standard two-microelectrode voltage-clamp technique (12). Briefly, a commercial amplifier (oocyte clamp model OC-725, Warner Instruments, Hamden, CT) and data acquisition system (Clampex 8.1 software, Axon Instruments, Union City, CA) conveyed voltage pulses to the oocyte and simultaneously recorded membrane current and voltage signals. The oocyte was superfused (∼1.5 ml/min) with a saline solution (ND96) containing 96 mM NaCl, 2 mM KCl, 1 mM MgCl2, 1.8 mM CaCl2, 10 mM HEPES-Tris, pH 7.6. After microelectrode impalement, the oocyte was left undisturbed for 1–10 min to allow the membrane potential to stabilize. Oocytes with membrane potentials less negative than −35 mV were discarded. The membrane potential was then clamped at −50 mV. When the current-voltage relationship was examined, a voltage range from +75 mV to −175 mV was covered in 25-mV steps; the oocyte membrane potential was held at the new levels for 250 ms intervals and traces were analyzed by averaging the signal in a window of 50 ms positioned after the decay of capacitive transient currents. Measurements were often taken in the absence and in the presence of a particular substrate (or inhibitor), and the substrate-specific current was determined by subtraction of one current from the other. When the concentration of NaCl was diminished, it was isotonically replaced with N-methyl-D-glucamine chloride. All of the steady-state current experiments were performed at room temperature (∼24°C). By standard convention, inward and outward currents are defined with respect to the movement of positive charges; thus, a movement of anions outward from the cell is defined as an inward current.
Uptake, current measurements
Stoichiometry measurements were performed as previously described (11) for 15 min with the exceptions that 1), three different substrate (lactate) concentrations (20, 100, and 200 μM) were employed; 2), uptakes were ended by addition of 1 mM ibuprofen (a potent inhibitor of SMCT1) and oocytes were then washed with ice-cold ND96 containing 1 mM ibuprofen; 3), electrophysiological data was recorded and processed using Clampex 8.1 software (Axon Instruments). The use of different substrate concentrations broadened the range of the data obtained, increasing the accuracy of measurement of the slope between the data points therein.
Volume, current measurements
High-accuracy volume measurements of Xenopus oocytes were performed as described (11) with minor modifications. Cotransport stoichiometry was assessed using a holding potential of −50 mV and exposure to 1 mM lactate for 10 min. For leak-current measurements, the holding potential was −25 mV, and 96 mM external Cl− was replaced by 96 mM external 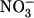 for 200 s.
for 200 s.
Other uptake experiments
The uptake of lactate or propionate into SMCT1-expressing oocytes without concomitant current measurements was performed at room temperature for 15 min using six oocytes in 500 μl ND96 solution containing 25 μM substrate. Na+ replacement was performed using N-methyl-D-glucamine (NMDG+). Ibuprofen, when present, was at 1 mM. The reaction was ended by rapidly washing the oocytes five times with 4 ml of ice-cold ND96 solution. Oocytes were dissolved overnight in 0.5 ml 10% SDS before addition of Betablend scintillation cocktail (ICN, Costa Mesa, CA) for liquid scintillation counting.
Reversal potential method
The measurement of reversal potentials in SMCT1-expressing oocytes was performed as previously described (12), except that lactic acid was added to the superfusing solution rather than propionic acid.
Zero-current clamp method
We followed a previously described procedure (16), with minor alterations. Oocytes expressing SMCT1 were voltage-clamped to −50 mV, exposed to ND96-MES medium (where 96 mM of Cl is replaced by 96 mM 2-(N-morpholino)ethanesulfonic acid (MES)) containing 5 mM lactic acid or 5 mM propionic acid, and left under these conditions for ∼10 min to allow preloading of the oocytes with substrate. The superfusing solution was then switched to ND96-MES and the voltage clamp was replaced by a zero-current clamp. Membrane potential was continuously monitored thereafter. The superfusing solution was altered to expose the oocyte to different concentrations of Na+ (replacement with NMDG+) or substrate (no replacement) to compare the changes in zero-current potential when either the external Na+ or monocarboxylate concentration was diminished by 50%.
To recap the theory underlying this procedure, the resting membrane potential Vm can be described as a summation of the reversal potential of each electrogenic transport pathway scaled by their partial conductance.
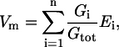 |
(1) |
where Gtot is the sum of all individual conductances (Gi). If a change in external ionic or substrate concentration affects one of the transport pathways, then
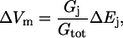 |
(2) |
assuming that these changes have not affected membrane conductances. If Ej represents Na+/lactate cotransport, it is defined as
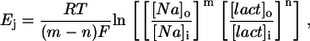 |
(3) |
where m and n are the numbers of Na+ ions and lactate molecules, respectively, per transport cycle.
Upon a change in external Na concentration from [Na]o to 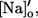 Ej will change by
Ej will change by
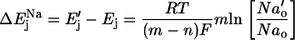 |
(4) |
Upon changing substrate concentration by an identical proportion from [lact]o to [lact]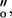 Ej will change by
Ej will change by
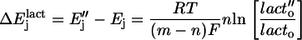 |
(5) |
Each change in the cotransport mechanism will be reflected in the overall Vm and, if the intracellular concentrations for Na+ and lactate remain constant, the ratio between the two changes in Vm will represent the ratio of m/n. A significant advantage to this method is that it does not require the use of an inhibitor, which can be problematic if the inhibitor simultaneously inhibits several distinct ionic pathways through the protein. This method simply requires that changing Na+ or lactic acid concentrations affects only SMCT1 cotransport, leaving the anionic leak through this protein and all other endogenous, electrogenic transport mechanisms untouched.
RESULTS
Our renewed efforts to characterize the stoichiometry of SMCT1 centered on lactate transport for several reasons, primarily due to the low level of passive diffusion of lactic acid across cell membranes. We began with the most common approach to measuring stoichiometry of cotransport, combining current measurements across the plasma membrane with uptake measurements of radiolabeled lactic acid. A separate measurement of the slope linking these data points was made during each of 5 weeks of experiments, each incorporating data from 8 to 11 oocytes of a particular “batch” of oocytes. The mean of these five slope measurements (Fig. 1) indicated that one charge was transported per lactate (1.04 ± 0.07 eq/mole), consistent with a 2:1 Na+/lactate stoichiometry (since lactate is an anion), similar to the stoichiometry observed for most members of the SLC5A family of Na+ cotransporter proteins. Similar results have been published elsewhere for the stoichiometry of lactate cotransport through SMCT1 (13). The oocytes exhibiting the lowest levels of SMCT1 current (all with 20 μM lactic acid superfusion) displayed unusually low levels of isotope accumulation. We do not yet know the cause of this.
FIGURE 1.

Estimation of SMCT1 cotransport stoichiometry for lactate using uptake data and current measurements. Oocytes were exposed to radiolabeled lactic acid (at a variety of concentrations) while the cell membrane potentials were clamped at −50 mV; the substrate-dependent currents were measured, and the total amount of current was deduced by integrating the area bound by the increased current due to substrate superfusion. At the end of the experiment, the oocyte was superfused with a solution lacking lactic acid but containing 1 mM ibuprofen; the voltage clamp was simultaneously ended. (A) A typical experiment is shown. The gray bar indicates the period during which lactic acid was present in the superfusing solution. (B) Comparison of uptake/current measurements. The data points for uptake and current measurement were used for linear regression; the best fit is shown by the dashed line.
As the absorption of monocarboxylates by colon involves co- and countertransport of other ionic species (and as we had initially thought that Cl− might be involved in SMCT1 activity), we also used an alternative means of measuring stoichiometry to better ascertain whether other ions might be involved in SMCT1 cotransport. One technique we have established (11) utilizes simultaneous measurement of current and volume in Xenopus oocytes that are expressing both the cotransporter under study and aquaporin. Rapid changes in volume, which maintain internal isotonicity, can be measured and are used to determine the number of osmolytes that have entered the cell during the time that substrate is present (and during which time transmembrane currents are measured). Analysis of the data requires measurement of the charge compensation provided by the intracellular, current-injecting electrode; when this is taken into account, a reliable estimate of the number of osmolytes passing through the cotransporter can be obtained. An example of this approach is shown in Fig. 2, and the average of 11 experiments produced a value of 3.1 ± 0.1 (mean ± SE) osmolytes per charge, in accordance with the 2:1 Na+/lactate stoichiometry established above by combining current and uptake measurements. As two different forms of measurement had yielded the same stoichiometry of cotransport, we considered this issue resolved for lactate cotransport.
FIGURE 2.

Estimation of SMCT1 Na+/lactate cotransport stoichiometry using volume measurements and current measurements. SMCT1 stoichiometry was determined by measuring the changes in oocyte volume and the substrate-induced current when the cells were exposed to 1 mM lactic acid. A typical experiment is shown; the upper trace indicates the transmembrane current (clamped at −50 mV), whereas the lower trace indicates the oocyte volume. Note dramatic changes in each tracing when 1 mM lactic acid was added to the superfusing solution. Coexpression of aquaporin was necessary to permit rapid volume change. Three lines obtained from modeling the volume changes are shown alongside the volume tracing; the numbers after each of these lines represent the numbers of osmolytes per unit charge that should yield that particular theoretical change in volume.
We did not attempt to establish the stoichiometry of Na+/propionate cotransport by combining uptake and current measurement because, in our hands, the passive diffusion of propionic acid is of considerable magnitude, preventing accurate measurement of flux through the SMCT1 transporter. To demonstrate this, we performed uptake experiments into SMCT1-expressing (and control) oocytes in the presence of either radiolabeled lactic acid or radiolabeled propionic acid (Fig. 3). Although there is negligible transport of lactate into control oocytes regardless of the presence of Na+ or ibuprofen, the expression of SMCT1 permits a large uptake of lactate that is Na+-dependent and ibuprofen-sensitive. In comparison to the uptake of lactate, there is a large, apparently passive uptake of propionic acid into oocytes, which is independent of external Na+ and ibuprofen and of similar magnitude to the parallel cotransport of propionate through SMCT1. We concluded that the large, passive diffusion of propionic acid through oocyte membranes prevents accurate assessment of stoichiometry when radiolabeled uptake is employed, since the passive diffusion would alter not only the amount of radiolabel that enters the cell during the current measurement but also the amount actually retained within the cell during washing after the external substrate is removed. Similarly, the passive diffusion of propionic acid seemed likely to confound stoichiometric measurements using oocyte volume since this method depends on restricting the influx of both Na+ and substrate to that which passes through the transporter over an extended period of time.
FIGURE 3.

Uptakes of (control) propionate and lactate. Oocytes were injected with water or with mRNA encoding SMCT1 (SMCT1) and uptake of radiolabeled lactic acid or propionic acid into the oocytes was assessed at 5–7 days after injection. Uptake experiments were performed for 15 min either: 1), in the absence of Na+ (replaced by NMDG+), 2), in the presence of 96 mM Na+, or 3), in the presence of 96 mM Na+ and 1 mM ibuprofen. Data shown represent one experiment with six oocytes per value shown, and are representative of three separate experiments. Both ordinate axes are labeled with respect to the transported anion, although the radiolabel uptake value actually reflects both anion transport and free acid diffusion.
Consequently, we utilized an alternative method that had recently been used to identify the stoichiometry of transporters expressed in Xenopus oocytes. This approach (16) employs a zero-current clamp of the oocyte membrane, and membrane potential is measured while external substrate and cation concentrations are varied by identical proportions. Initial experiments performed by superfusing a solution containing lactic acid indicated an unstable zero-current membrane potential as, presumably, the internal lactate levels increased rapidly during the experiment. For this reason, we subsequently began each experiment with a 10-min loading period during which the SMCT1-expressing oocytes were exposed to medium containing 96 mM Na+ and 5 mM lactic acid or propionic acid. Under these conditions, the current generated by SMCT1 rapidly fell as the internal lactate and Na+ concentrations rose. After this preloading step, superfusion with various concentrations of Na+ and lactate were unable to cause such rapid changes in the intracellular substrate concentrations, and steady currents were achieved. As Fig. 4 A illustrates, the effect of reducing the lactate concentration by a factor of 2 is about half the effect of reducing the Na+ concentration by a factor of 2. When [Na+] is further reduced from 48 to 24 mM, the effect on Vm is larger, but in the presence of 48 mM Na+, the effect of reducing [lactate] is also larger. On average, for a series of 11 experiments, the zero-current membrane potential approach yielded a calculated Na+/lactate stoichiometric ratio of 2.2 ± 0.2 (mean ± SE), in close agreement with the stoichiometric measurements described above, which were obtained using other methods. No difference was measured in the stoichiometric ratio whether the change in external [Na+] was from 96 to 48 mM, or from 48 to 24 mM. Application of this current-clamp procedure to propionate transport yielded a Na+/monocarboxylate stoichiometric ratio of 2.3 ± 0.3 (mean ± SE, n =7) (Fig. 4 B). As this is the only reliable means we have found for measuring stoichiometry of propionate uptake, we must conclude that SMCT1 exhibits a fixed 2:1 stoichiometry for both lactate and propionate. It should be noted that preliminary experiments indicated that decreasing the superfusing solution [Na+] by half caused a small change in membrane potential consistent with a low, partial conductance that would be equivalent to only 5% of the conductance recorded for the same oocyte in the presence of lactic acid, indicating that non-SMCT1 Na+ conductance was of negligible importance (data not shown). Similarly, exposure of control oocytes to 10 mM propionic acid did not significantly affect the current measured anywhere between −175 mV and +75 mV. We extended this technique to examine the stoichiometries for cotransport of pyruvate and nicotinate. These substrates are avidly transported through SMCT1 (12) and their cognate acids are more and less hydrophilic, respectively, than lactic and propionic acids (based on octanol/water partition coefficients). Na+/pyruvate cotransport displayed a stoichiometry of 1.91 ± 0.09, whereas Na+/nicotinate cotransport exhibited a stoichiometry of 2.0 ± 0.2, confirming an invariant 2:1 stoichiometry of cotransport through SMCT1.
FIGURE 4.

Current-clamp measurements of lactate and propionate cotransport stoichiometries. By preloading oocytes with Na+ and substrate via prior cotransport (not shown), rapid changes in the internal concentrations of these agents during subsequent current-clamp measurements were minimized. The lines at the bottom of the figure indicate where external superfusing solution contained Na+ at 96, 48, and 24 mM, and where the external monocarboxylic acid was present at 0.5 or 1 mM. The cotransport stoichiometry is estimated from the ratio between the changes in membrane voltage when either Na+ or substrate concentration was halved.
We had previously published a 3:1 stoichiometry for Na+:propionate through SMCT1 obtained by measuring reversal potentials in the presence of different concentrations of Na+ and propionate. The same result was obtained by measuring the reversal potentials of ibuprofen-sensitive currents in the presence of different external lactic acid concentrations (Fig. 5). The observed reversal potentials of ibuprofen-sensitive currents reflect the contributions of two different pathways: Na+/lactate cotransport and the anionic leak (12,15). These factors may explain why the reversal potentials observed are far more negative than would be expected. For instance, given an internal [Na+] of 8.5 mM (17), the observed reversal potential (+40 mV) for SMCT1 exposed to 1 mM external lactic acid in the presence of 96 mM Na+ would require nearly 50 mM internal lactate if it were determined solely by cotransport with a 2:1 Na+:substrate stoichiometry. The actual internal lactate concentration is presumably several orders of magnitude less than this (and thus outward cotransport is not a significant factor) and the leak current can clearly be seen to become the dominant feature of SMCT1 at positive potentials since the cotransport current is inwardly rectifying, the leak current is outwardly rectifying, and all of the ibuprofen-sensitive currents start to merge at these potentials. We sought to ameliorate the effects of the leak current by replacing most of the external Cl− by MES− (which is not transported through the leak current), but this did not measurably improve the stoichiometric predictions (data not shown). Since the leak currents are the overwhelming feature of SMCT1 activity over the range of membrane potentials where most reversal potentials would be measured, this method of determining stoichiometry is unsuited for this cotransporter or any other with similar rectifying currents.
FIGURE 5.

SMCT1 activity with different lactate concentrations. In an effort to determine why simple measurement of reversal potentials at different substrate concentrations did not accurately yield the stoichiometric ratio, oocytes expressing SMCT1 were superfused with different concentrations of lactic acid and the ibuprofen-sensitive currents were recorded at different membrane potentials. A typical experiment is shown. The ibuprofen-sensitive currents observed in the absence of external lactic acid represent the anionic leak current passing through SMCT1. Note that the ibuprofen-sensitive currents all merge into the leak current at positive potentials, where the reversal potentials would normally be observed.
To complete our understanding of the stoichiometries by which SMCT1 operates, we used volume measurement to determine the stoichiometry of the leak current passing through SMCT1. Although we had previously assigned this current to an influx of external anions, we had not ruled out the possibility that part of the leak current could be due to outward cotransport of endogenous, intracellular Na+ and monocarboxylates stimulated by superfusion with some specific anions such as I− or 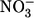 (15). The measured stoichiometry (Fig. 6) indicates 0.98 ± 0.21 osmolytes per negative charge passing through this current (mean ± SE, n = 16), in agreement with our original assessment of this current as an influx of anions. Thus, there is no observable efflux of Na+ and monocarboxylates through this protein in an oocyte that had not been recently exposed to monocarboxylates in the bathing medium.
(15). The measured stoichiometry (Fig. 6) indicates 0.98 ± 0.21 osmolytes per negative charge passing through this current (mean ± SE, n = 16), in agreement with our original assessment of this current as an influx of anions. Thus, there is no observable efflux of Na+ and monocarboxylates through this protein in an oocyte that had not been recently exposed to monocarboxylates in the bathing medium.
FIGURE 6.

Stoichiometry of the leak current. Current/volume measurements, similar to those of Fig. 2, were used to examine the stoichiometry of the SMCT1 leak current at −25 mV holding potential. The upper tracing indicates the current measurement during the experiment and the lower tracing indicates the oocyte volume measurement. Replacement of most of the Cl− in the superfusing solution by 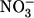 resulted in a large outward current (i.e., inward flux of anions) and concomitant swelling of the oocyte (a typical result is shown). Dotted lines indicate the mean change in volume under conditions before, during, and after exposure to
resulted in a large outward current (i.e., inward flux of anions) and concomitant swelling of the oocyte (a typical result is shown). Dotted lines indicate the mean change in volume under conditions before, during, and after exposure to 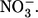
DISCUSSION
The possibility of variable stoichiometry through SMCT1, as well as the competing stoichiometric ratios described for cotransport through this protein, prompted us to definitively establish the cotransport stoichiometry for this protein. Although SMCT1 is best known for its cotransport of butyrate and other membrane-permeant short-chain fatty acids, it also cotransports lactate and exhibits a relatively large anionic leak current (15). In this article, we have used three distinct means of measuring Na+/lactate cotransport, and each method indicated a 2:1 Na+/lactate stoichiometry. Given the diverse nature of the methods by which these techniques measure stoichiometry, we feel that the legitimacy of the stoichiometric ratio established here is beyond question.
The application of the simplest technique (current/uptake measurements) for measuring stoichiometry of propionate transport through SMCT1 gave an incorrect result of 4 Na+ ions/propionate (13,14). These results were presumably skewed due to the highly hydrophobic nature of propionic acid, as well as its pKa value of 4.8, permitting a large degree of passive permeation into and out of the cells. Although it could be argued that the rapid, passive diffusion of radiolabeled propionic acid into cells observed in our hands could be attributable to a faulty batch of radiolabeled substrate, this seems highly unlikely, since we have obtained similar data regarding the permeability of unlabeled propionic acid and butyric acid through oocyte membranes (12), as have a number of other laboratories worldwide (18–23). Consequently, the published 4:1 Na+/propionate stoichiometry was presumably affected by diffusion of radiolabeled propionic acid out of the cell when washing the oocyte to remove adherent substrate at the end of the experiment. It should be noted that the rapid diffusion of short-chain fatty acids into Xenopus oocytes and many other cells may not hold true for the apical membranes of colonocytes in vivo, given observations made about the passive permeation of CO2 (24). The lack of passive diffusion of lactic acid is presumably due both to the more hydrophilic nature of the acid as well as to the lower pKa value (3.86).
The use of current-clamped oocytes to measure diffusion potentials enabled us to avoid the use of positive membrane potentials, where the leak current predominates, and preloading of the oocytes with substrate prevented rapid changes in the intracellular concentration. Most important, this technique was able to unequivocally measure the effects of changing Na+ and substrate concentrations on the cotransport current alone. Consequently, we feel that this method is likely to provide a true stoichiometric ratio for propionate and for other substrates that involve a high level of passive, nonelectrogenic diffusion and whose cotransport stoichiometry cannot therefore be measured using other techniques. For this technique to work, the effect of changing external Na+ and substrate concentrations needs to be almost exclusively through the cotransporter studied. In control (noninjected) oocytes and in SMCT1-expressing oocytes (before exposure to monocarboxylates), changing the external Na+ concentration produced a small depolarizing effect that became negligible when the conductance of the oocyte was quintupled by the cotransport current of SMCT1. Changing the level of lactic acid or propionic acid in the bathing medium did not change membrane potential, and consequently the substrate changes did not appreciably change any conductances other than those passing through SMCT1 cotransport.
As measurements of voltage-clamped reversal potentials indicated a 3:1 stoichiometry for both propionate and lactate cotransport, it is instructive to consider why this particular technique provided inaccurate stoichiometric measurements for SMCT1. One significant factor is the nature of the leak currents (currents passing through the protein in the absence of substrate and/or cotransported ions). First demonstrated through SGLT1 (25), leak currents could be due to outward cotransport through the protein or to uncoupled ionic flux through the cotransporter. The leak current through SGLT1 did not pose a significant problem for the measurement of stoichiometry because, as described by the kinetic model proposed for this cotransporter, the leak current is in competition with the cotransport current. It was estimated that the leak was equal to cotransport current when glucose reached 10 μM and rapidly became negligible when the glucose concentration was further increased (10). In the case of SMCT1, the leak current is an inward flux of inorganic anions (M. J. Coady, B. Wallendorff, and J.-Y. Lapointe, unpublished) and there is a low degree of competition observed between the leak and cotransport currents. More important, the leak current vastly predominates at positive membrane potentials, where the SMCT1 cotransport reversal potentials would lie, even if >90% of the external Cl− is replaced by an impermeant anion. Consequently, although this technique works well with some cotransport proteins, it failed to accurately measure SMCT1 stoichiometry.
In addition, we measured the stoichiometry of the leak current passing through SMCT1 to confirm that it was purely due to anionic passage. We measured a stoichiometric value of 1 osmolyte/charge, indicating a purely anionic influx without any evidence of outward cotransport of the oocyte's native intracellular Na+ and monocarboxylates.
In summary, the stoichiometry of cotransport through SMCT1 has been shown to be fixed at 2 Na+ ions/substrate molecule, along with a purely anionic leak current. Establishing such stoichiometric ratios is not always straightforward, and accurate identification of these ratios may require considerable insight into the activity of that particular cotransporter. Similarly, identification of the true stoichiometric ratio is an important step in understanding how any cotransporter functions.
Acknowledgments
This work was supported by the Canadian Institutes for Health Research (MOP-10580).
Editor: Tzyh-Chang Hwang.
References
- 1.Turner, R. J., and A. Moran. 1982. Further studies of proximal tubular brush border membrane D-glucose transport heterogeneity. J. Membr. Biol. 70:37–45. [DOI] [PubMed] [Google Scholar]
- 2.Wright, E. M., and E. Turk. 2004. The sodium/glucose cotransport family SLC5. Pflugers Arch. 447:510–518. [DOI] [PubMed] [Google Scholar]
- 3.Chen, N. H., M. E. Reith, and M. W. Quick. 2004. Synaptic uptake and beyond: the sodium- and chloride-dependent neurotransmitter transporter family SLC6. Pflugers Arch. 447:519–531. [DOI] [PubMed] [Google Scholar]
- 4.Coady, M. J., X. Z. Chen, and J. Y. Lapointe. 1996. rBAT is an amino acid exchanger with variable stoichiometry. J. Membr. Biol. 149:1–8. [DOI] [PubMed] [Google Scholar]
- 5.Alonso, G. L., D. A. Gonzalez, D. Takara, M. A. Ostuni, and G. A. Sanchez. 2001. Kinetic analysis of a model of the sarcoplasmic reticulum Ca-ATPase, with variable stoichiometry, which enhances the amount and the rate of Ca transport. J. Theor. Biol. 208:251–260. [DOI] [PubMed] [Google Scholar]
- 6.Sacher, A., A. Cohen, and N. Nelson. 2001. Properties of the mammalian and yeast metal-ion transporters DCT1 and Smf1p expressed in Xenopus laevis oocytes. J. Exp. Biol. 204:1053–1061. [DOI] [PubMed] [Google Scholar]
- 7.Gross, E., and I. Kurtz. 2002. Structural determinants and significance of regulation of electrogenic Na+-HCO(3)− cotransporter stoichiometry. Am. J. Physiol. Renal Physiol. 283:F876–F887. [DOI] [PubMed] [Google Scholar]
- 8.Iwamoto, H., R. D. Blakely, and L. J. De Felice. 2006. Na+, Cl−, and pH dependence of the human choline transporter (hCHT) in Xenopus oocytes: the proton inactivation hypothesis of hCHT in synaptic vesicles. J. Neurosci. 26:9851–9859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Eskandari, S., D. D. Loo, G. Dai, O. Levy, E. M. Wright, and N. Carrasco. 1997. Thyroid Na+/I− symporter. Mechanism, stoichiometry, and specificity. J. Biol. Chem. 272:27230–27238. [DOI] [PubMed] [Google Scholar]
- 10.Chen, X. Z., M. J. Coady, F. Jackson, A. Berteloot, and J. Y. Lapointe. 1995. Thermodynamic determination of the Na+: glucose coupling ratio for the human SGLT1 cotransporter. Biophys. J. 69:2405–2414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bourgeois, F., M. J. Coady, and J. Y. Lapointe. 2005. Determination of transport stoichiometry for two cation-coupled myo-inositol cotransporters: SMIT2 and HMIT. J. Physiol. 563:333–343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Coady, M. J., M. H. Chang, F. M. Charron, C. Plata, B. Wallendorff, J. F. Sah, S. D. Markowitz, M. F. Romero, and J. Y. Lapointe. 2004. The human tumour suppressor gene SLC5A8 expresses a Na+-monocarboxylate cotransporter. J. Physiol. 557:719–731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gopal, E., Y. J. Fei, M. Sugawara, S. Miyauchi, L. Zhuang, P. Martin, S. B. Smith, P. D. Prasad, and V. Ganapathy. 2004. Expression of slc5a8 in kidney and its role in Na+-coupled transport of lactate. J. Biol. Chem. 279:44522–44532. [DOI] [PubMed] [Google Scholar]
- 14.Miyauchi, S., E. Gopal, Y. J. Fei, and V. Ganapathy. 2004. Functional identification of SLC5A8, a tumor suppressor down-regulated in colon cancer, as a Na+-coupled transporter for short-chain fatty acids. J. Biol. Chem. 279:13293–13296. [DOI] [PubMed] [Google Scholar]
- 15.Reference deleted in proof.
- 16.Horita, S., H. Yamada, J. Inatomi, N. Moriyama, T. Sekine, T. Igarashi, Y. Endo, M. Dasouki, M. Ekim, L. Al-Gazali, M. Shimadzu, G. Seki, and T. Fujita. 2005. Functional analysis of NBC1 mutants associated with proximal renal tubular acidosis and ocular abnormalities. J. Am. Soc. Nephrol. 16:2270–2278. [DOI] [PubMed] [Google Scholar]
- 17.Charron, F. M., M. G. Blanchard, and J. Y. Lapointe. 2006. Intracellular hypertonicity is responsible for water flux associated with Na+/glucose cotransport. Biophys. J. 90:3546–3554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rodeau, J. L., S. Flament, E. Browaeys, and J. P. Vilain. 1998. Effect of procaine on membrane potential and intracellular pH in Xenopus laevis oocytes. Mol. Membr. Biol. 15:145–151. [DOI] [PubMed] [Google Scholar]
- 19.Leipziger, J., G. G. MacGregor, G. J. Cooper, J. Xu, S. C. Hebert, and G. Giebisch. 2000. PKA site mutations of ROMK2 channels shift the pH dependence to more alkaline values. Am. J. Physiol. Renal Physiol. 279:F919–F926. [DOI] [PubMed] [Google Scholar]
- 20.Stewart, A. K., M. N. Chernova, B. E. Shmukler, S. Wilhelm, and S. L. Alper. 2002. Regulation of AE2-mediated Cl− transport by intracellular or by extracellular pH requires highly conserved amino acid residues of the AE2 NH2-terminal cytoplasmic domain. J. Gen. Physiol. 120:707–722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Boldt, M., G. Burckhardt, and B. C. Burckhardt. 2003. NH4+ conductance in Xenopus laevis oocytes. III. Effect of NH(3). Pflugers Arch. 446:652–657. [DOI] [PubMed] [Google Scholar]
- 22.Cooper, G. J., and P. Fong. 2003. Relationship between intracellular pH and chloride in Xenopus oocytes expressing the chloride channel ClC-0. Am. J. Physiol. Cell Physiol. 284:C331–C338. [DOI] [PubMed] [Google Scholar]
- 23.Kim, K. H., N. Shcheynikov, Y. Wang, and S. Muallem. 2005. SLC26A7 is a Cl− channel regulated by intracellular pH. J. Biol. Chem. 280:6463–6470. [DOI] [PubMed] [Google Scholar]
- 24.Endeward, V., and G. Gros. 2005. Low carbon dioxide permeability of the apical epithelial membrane of guinea-pig colon. J. Physiol. 567:253–265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Umbach, J. A., M. J. Coady, and E. M. Wright. 1990. Intestinal Na+/glucose cotransporter expressed in Xenopus oocytes is electrogenic. Biophys. J. 57:1217–1224. [DOI] [PMC free article] [PubMed] [Google Scholar]