

Abstract
Breast cancer is a heterogeneous disease, comprising multiple entities associated with distinctive histological and biological features, clinical presentations and behaviours and responses to therapy. Microarray‐based technologies have unravelled the molecular underpinning of several characteristics of breast cancer, including metastatic propensity and histological grade, and have led to the identification of prognostic and predictive gene expression signatures. Furthermore, a molecular taxonomy of breast cancer based on transcriptomic analysis has been proposed. However, microarray studies have primarily focused on invasive ductal carcinomas of no special type. Owing to the relative rarity of special types of breast cancer, information about the biology and clinical behaviour of breast cancers conveyed by histological type has not been taken into account. Histological special types of breast cancer account for up to 25% of all invasive breast cancers. Recent studies have provided direct evidence of the existence of genotypic–phenotypic correlations. For instance, secretory carcinomas of the breast consistently harbour the t(12;15) translocation that leads to the formation of the ETV6–NTRK3 fusion gene, adenoid cystic carcinomas consistently display the t(6;9) MYB–NFIB translocation and lobular carcinomas consistently show inactivation of the CDH1 gene through multiple molecular mechanisms. Furthermore, histopathological and molecular analysis of tumours from conditional mouse models has provided direct evidence for the causative role of specific genes in the genesis of specific histological special types of breast cancer. Here we review the associations between the molecular taxonomy of breast cancer and histological special types, discuss the possible origins of the heterogeneity of breast cancer and propose an approach for the identification of novel therapeutic targets based on the study of histological special types of breast cancer.
Keywords: Breast cancer, Special types, Classification, Microarrays, Therapeutic targets
1. Introduction
Breast cancer is a heterogeneous disease, comprising numerous distinct entities that not only have different biological features but also clinical behaviours (Weigelt et al., 2010c; Weigelt and Reis‐Filho, 2009; Reis‐Filho and Lakhani, 2008; Simpson et al., 2005a; Vargo‐Gogola and Rosen, 2007). There are several lines of evidence to suggest that breast cancer is, in fact, a collection of different diseases with different risk factors, clinical presentations, pathological features, response to therapy and outcomes, which affect the same anatomical organ and originate in the same anatomical structure (i.e. the terminal duct‐lobular unit). Although histopathologists have been aware of the diversity of breast cancer and have endeavoured to devise approaches to classify the disease into meaningful groups (Weigelt and Reis‐Filho, 1979, 1982, 1973, 1987, 2001, 1968, 1999, 2009), this concept has only been brought to the forefront of breast cancer research after the publication of high‐throughput microarray‐based class discovery studies that unravelled the existence of multiple molecular subtypes (Perou et al., 2006, 2000, 2001, 2003).
Breast cancers can be classified into biologically and clinically meaningful subgroups according to histological grade (Elston and Ellis, 1991) and histological type (Ellis et al., 1992). Grade is an assessment of the degree of differentiation (i.e. tubule formation and nuclear pleomorphism) and proliferative activity (i.e. mitotic index) of a tumour, and mirrors its aggressiveness (Elston and Ellis, 1991). Histological grade has been incorporated in multiple, validated, prognostic algorithms to determine breast cancer therapy, such as the Nottingham Prognostic Index and Adjuvant! Online (Mook et al., 2009). Interestingly, grade has been shown to correlate with the genetic and transcriptomic features of breast cancers and microarray‐based genomic signatures for histological grade have been devised (Weigelt et al., 2010c; Sotiriou et al., 2006; Ivshina et al., 2006; Sotiriou and Pusztai, 2009).
Histological type, on the other hand, refers to the growth pattern of the tumours. The histological diversity of adenocarcinomas in the breast has long fascinated pathologists, who have identified specific morphological and cytological patterns that were consistently associated with distinctive clinical presentations and/or outcomes. These patterns are called ‘histological types’. The commonest type of breast carcinoma is the so‐called invasive ductal carcinomas not otherwise specified (IDC‐NOS) or of no special type (IDC‐NST) (Ellis et al., 2003) (Table 1), which is a diagnosis of exclusion and comprises adenocarcinomas that fail to exhibit sufficient characteristics to warrant their classification in one of the special types. Breast cancer special types account for up to 25% of all breast cancers and the latest edition of the World Health Organisation classification recognises the existence of at least 17 distinct histological special types (1, 2) (Ellis et al., 2003). It should be noted that grade and type provide complementary information (Rakha et al., 2008, 2008). Although grade identifies prognostic subgroups among special types of breast cancer, some entities that are by definition of high histological grade have a relatively good prognosis (e.g. medullary carcinomas) (Ellis et al., 2003).
Table 1.
Prevalence of histological types of breast cancer.
| Histological type | Prevalence (%) Rosen (2001) | Prevalence (%) WHO (Ellis et al., 2003) | Prevalence (%) Rakha et al. (2006, 2008c)a | Prevalence (%) Louwman et al. (2007)b |
|---|---|---|---|---|
| Invasive ductal carcinoma (NST) | 65–80 | 50–80 | 56.4 | 78 |
| Carcinoma with osteoclastic giant cells | 0.5–1.2 | |||
| Invasive lobular carcinoma | 5 | 5–15 | 8.2 | 11.1 |
| Classical | 7.4 | |||
| Alveolar | 0.1 | |||
| Solid | 0.3 | |||
| Tubulo‐lobular | 0.4 | |||
| Pure tubular carcinoma | <2 | <2 | 4.4 | 2.2 |
| Invasive cribriform carcinoma | <4 | 0.8–3.5 | 0.6 | 0.3 |
| Medullary carcinoma | <5–7 | 1–7 | 2.6 | 1.1 |
| Typical | 0.3 | |||
| Atypical | 2.3 | |||
| Mucinous carcinoma | <2 | 2 | 1.4 | 2.2 |
| Neuroendocrine carcinoma | 2–5 | 0.0 | ||
| Invasive papillary carcinoma | 1–2 | 1–2 | 0.4 | 0.7 |
| Invasive micropapillary carcinoma | <2.7 | <2 | ||
| Apocrine carcinoma | <1–4 | <4 | ||
| Metaplastic carcinoma | <5 | <1 | 0.2 | |
| Lipid‐rich carcinoma | <1 | <1–6 | ||
| Secretory carcinoma | Few cases | <0.15 | ||
| Oncocytic carcinoma | Few cases | |||
| Adenoid cystic carcinoma | Few cases | 0.1 | 0.1 | |
| Acinic‐cell carcinoma | Few cases | |||
| Glycogen‐rich clear cell carcinoma | <1–3 | 1–3 | ||
| Sebaceous carcinoma | Few cases | |||
| Mixed types | 25.3 | |||
| NST and invasive lobular carcinoma | 3.3 | 4.0 | ||
| NST and special type | 2.1 | |||
| Invasive lobular mixed | 3.1 | |||
| Tubular carcinoma mixed | 16.8 | |||
| Miscellaneous | 0.6 (Metaplastic and adenoid cystic carcinoma) | 0.0 (Signet ring cell carcinoma) | ||
2304 invasive breast cancers diagnosed in Nottingham, UK between 1987 and 1998 from two consecutive series (Rakha et al., 2006, 2008c; Emad Rakha, personal communication).
Adapted from Louwman et al. (2007), Netherlands Cancer Registry, considering only cases where correct morphological codes and histopathological review were available.
Figure 1.
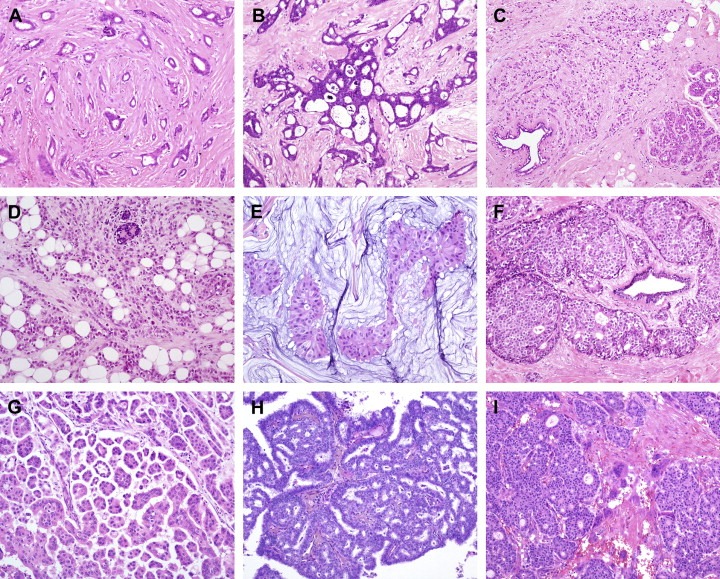
Histological special types of breast cancer preferentially oestrogen receptor positive. (A) Tubular carcinoma, (B) cribriform carcinoma, (C) classic invasive lobular carcinoma, (D) pleomorphic invasive lobular carcinoma, (E) mucinous carcinoma, (F) neuroendocrine carcinoma, (G) micropapillary carcinoma, (H) papillary carcinoma, (I) low grade invasive ductal carcinoma with osteoclast‐like giant cells.
Figure 2.
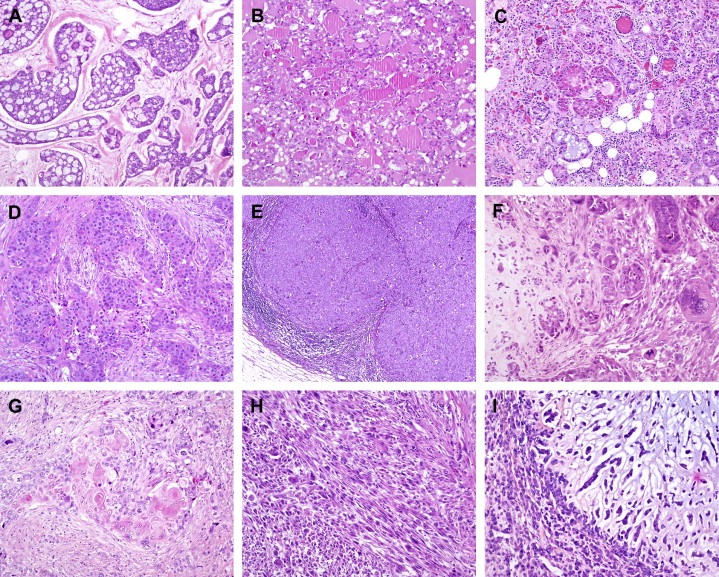
Histological special types of breast cancer preferentially oestrogen receptor negative. (A) Adenoid cystic carcinoma, (B) secretory carcinoma, (C) acinic‐cell carcinoma, (D) apocrine carcinoma, (E) medullary carcinoma, (F) metaplastic carcinoma with heterologous elements, (G) metaplastic carcinoma with squamous metaplasia, (H) metaplastic spindle cell carcinoma, (I) metaplastic matrix‐producing carcinoma.
Although special types of breast cancer have been shown to be associated with distinct biological features and carry important clinical implications (e.g. patients with tubular carcinomas have survival rates close to normal life expectancy, Rakha et al., 2010), the use of information on special types has been limited in tailoring the therapy for breast cancer patients, partly due to the lack of standardised criteria for their diagnosis and low inter‐observer reproducibility. In addition, owing to the relative low prevalence of special types of breast cancer, they have not been systematically investigated in class discovery (Perou et al., 2006, 2000, 2001, 2003) and class prediction (Sotiriou et al., 2006; Wang et al., 2005; van't Veer et al., 2002; van de Vijver et al., 2002; Wirapati et al., 2008) microarray‐based expression profiling studies.
Here we review the morphological and molecular subtypes of breast cancer, discuss the possible origins of the heterogeneity of breast cancers and, finally, propose that the study of special types of breast cancer may help identify novel therapeutic targets for specific subgroups of breast cancer patients.
2. Histological special types vs. molecular subtypes
The long‐standing believe that distinct histological types of breast cancer would arise from distinct microanatomical structures of the normal breast was challenged by Wellings and Jensen (1973) and Wellings et al. (1975) who demonstrated that the vast majority of invasive breast cancers, and their in situ precursors, originate from the terminal duct‐lobular unit regardless of the histological type. It should be emphasised that the terms ductal and lobular carcinoma by no means imply histogenesis or site of origin within the mammary ductal system. Instead, these are entities defined based on their discrete architectural patterns, cytological features and immunohistochemical profiles. For a concise description of the histopathology of special types of breast cancer the authors refer to a recent review by Yerushalmi et al. (2009).
In the past decade, high‐throughput microarray‐based gene expression profiling has been extensively applied to the study of breast cancer to unravel the molecular underpinning of biological features such as metastatic propensity (Wang et al., 2005; van't Veer et al., 2002; van de Vijver et al., 2002) or histological grade (Sotiriou et al., 2006), and to identify signatures associated with prognosis (Sotiriou et al., 2006; Wang et al., 2005; van't Veer et al., 2002; van de Vijver et al., 2002) and response to therapy (Potti et al., 2006). In addition, class discovery analysis of breast cancers subjected to microarray‐based expression profiling has led to a rediscovery at the molecular level of the long appreciated morphological and clinical heterogeneity in breast cancer.
In the seminal study by Perou et al. (2000), the authors performed cDNA microarray analysis of 38 invasive breast cancers (36 invasive ductal carcinomas and 2 lobular carcinomas), 1 ductal carcinoma in situ, 1 fibroadenoma and 3 normal breast samples, and a number of biological replicates from the same patient, and defined an ‘intrinsic’ gene list (i.e. genes that vary most between tumours from different patients compared to samples from the same tumour/patient). Hierarchical cluster analysis using this ‘intrinsic gene list’ revealed (i) the division of the cluster dendrogram into oestrogen receptor (ER)‐positive and ER‐negative breast cancers, a robust feature when analysing the transcriptome of breast cancers (Weigelt et al., 2010c; van't Veer et al., 2002; Correa Geyer and Reis‐Filho, 2009; Gruvberger et al., 2001), and (ii) the existence of four molecular subtypes of breast cancer: luminal, normal breast‐like, HER2 and basal‐like (Perou et al., 2000) (Table 2). The same group demonstrated in a subsequent study using an extended patient cohort that the ER‐positive luminal group could be separated into at least 2 subgroups, luminal A and luminal B, and that different molecular subtypes were associated with distinct clinical outcomes (Sorlie et al., 2001). These molecular subtypes of breast cancer were confirmed and expanded upon in subsequent microarray datasets (Sorlie et al., 2003; Hu et al., 2006) and to some extent by independent groups (Sotiriou et al., 2003).
Table 2.
Characteristics of molecular subtypes and assignment of histological special types of breast cancer.
| Molecular subtype | ER, PR, HER2 | Additional markers | Proliferation cluster microarrays | Histological special types |
|---|---|---|---|---|
| Basal‐like | ER− | CK5/6+ | High | Adenoid cystic |
| PR− | EGFR+ | Acinic cell | ||
| HER2− | Medullary | |||
| Metaplastic | ||||
| Pleomorphic lobular | ||||
| Secretory | ||||
| HER2/ER− | ER− | CK5/6+/− | High | Apocrine |
| PR− | EGFR+/− | Lobular | ||
| HER2+ | Micropapillary | |||
| Pleomorphic lobular | ||||
| Normal breast‐like | ER−/+ | CK5/6+ | Low | Medullary |
| PR unknown | EGFR+ | Metaplastic | ||
| HER2− | ||||
| Luminal | ER+ (−) | Low/high | Apocrine | |
| PR+/− | IDC osteoclastic | |||
| HER2− (+) | Lobular | |||
| Micropapillary | ||||
| Mucinous | ||||
| Neuroendocrine | ||||
| Pleomorphic lobular | ||||
| Tubular | ||||
| Molecular apocrine | ER− | AR+ | High | Apocrine |
| PR− | CK5/6+/− | Pleomorphic lobular | ||
| HER2+/− | EGFR+/− | |||
| Claudin‐low | ER− | CLDN low/− | High | Metaplastic |
| PR− | CDH1 low/− | Medullary (?) | ||
| HER2− | CK5/6+/− | |||
| EGFR+/− | ||||
| Interferon‐related | ER−/+ | STAT1 | High | Medullary (?) |
| PR unknown | ||||
| HER2− | ||||
AR: androgen receptor; CDH1: E‐cadherin; CLDN: claudin; CK: cytokeratin; EGFR: epidermal growth factor receptor; ER: oestrogen receptor; IDC: invasive ductal carcinoma; PR: progesterone receptor; STAT1: signal transducer and activator of transcription 1. −: negative; +: positive; +/−: occasionally positive; −/+: rarely positive.
The ER‐positive luminal tumours were originally described as those that show expression patterns reminiscent of ‘normal luminal epithelial cells’ of the mammary gland, including low molecular weight cytokeratins 8/18, ER and genes associated with an active ER pathway (Weigelt et al., 2009, 2000, 2001, 2003, 2010). Luminal A tumours have been shown to have high levels of expression of ER‐activated genes and low levels of proliferation related genes, to be usually of low histological grade and have a good outcome, whereas luminal B cancers are more often of higher histological grade, have higher proliferation rates and a significantly worse prognosis than luminal A tumours (Weigelt et al., 2009, 2009, 2000, 2001, 2003, 2010) (Table 2). It should be noted, however, that a recent large meta‐analysis of published available expression data suggested that luminal tumours form a continuum and that the separation of these tumours into two subgroups based on proliferation may be arbitrary (Wirapati et al., 2008).
In the ER‐negative branch of the cluster, three subtypes were identified: normal breast‐like, HER2 and basal‐like. Normal breast‐like cancers have been shown to consistently cluster together with fibroadenoma and normal breast samples and to be enriched for genes usually expressed in adipose tissue (Peppercorn et al., 2008). Tumours of this subtype are still poorly characterised and their clinical significance has yet to be determined (Correa Geyer and Reis‐Filho, 2009; Pusztai et al., 2006). Recently, the proponents of this molecular classification suggested that the normal breast‐like subtype may be a mere artefact of sample representation (i.e. disproportionately high content of normal tissue contamination) (Parker et al., 2009; Peppercorn et al., 2008). The HER2 and basal‐like molecular subtypes are associated with an aggressive clinical behaviour. HER2 tumours overexpress HER2 and genes associated with HER2 pathway and/or HER2 amplicon on 17q12 (e.g. GRB7) (Table 2). It should be noted that although the majority (>80%) of microarray‐defined HER2 cancers harbour HER2 gene amplification or HER2 3+ immunohistochemical expression (Rouzier et al., 2005), not all HER2‐amplified tumours fall into the HER2 cluster by expression arrays analysis (Parker et al., 2009; Rouzier et al., 2005). A significant number of HER2‐amplified, ER‐positive cancers actually fall into the luminal B subtype rather than the HER2‐microarray subtype (Parker et al., 2009; Rouzier et al., 2005). The basal‐like subtype, which has been extensively studied in the recent years, was named ‘basal‐like’ given that the neoplastic cells of these tumours consistently express genes usually found in normal basal/myoepithelial cells of the breast, including high molecular weight cytokeratins (5 and 17), P‐cadherin, caveolins 1 and 2 (Abd El‐Rehim et al., 2004, 2005, 2004, 2006, 2002, 2007, 2008, 2006), nestin (Parry et al., 2008), CD44 (Klingbeil et al., 2010) and epidermal growth factor receptor (EGFR) (Nielsen et al., 2004). In a minority of cases, they harbour EGFR gene amplification (Weigelt et al., 2010c) or aneusomy (Gilbert et al., 2008). However, basal‐like tumours may express genes characteristic of luminal epithelia, including cytokeratin 8/18, albeit at lower levels than luminal carcinomas, as well as KIT, which is expressed by luminal ER‐negative epithelia (Westbury et al., 2009). Basal‐like carcinomas are usually of high histological grade, have high mitotic indices, central necrotic zones, pushing borders, conspicuous lymphocytic infiltrate, typical/atypical medullary features and metaplastic areas (Fulford et al., 2006; Livasy et al., 2006; Reis‐Filho et al., 2006b; Turner et al., 2007). This subtype has been reported to be more prevalent in young women of African and Hispanic descent, to be associated with distinct risk factors and to show a high response rate to traditional chemotherapeutic regimens (Rakha et al., 2008a; Reis‐Filho and Tutt, 2008; Carey et al., 2006). The morphological and immunohistochemical features of basal‐like cancers are remarkably similar to those described for tumours arising in BRCA1 germ‐line mutation carriers (Turner and Reis‐Filho, 2003, 1998, 2002, 2005, 2006) and there is a growing body of evidence to suggest that BRCA1 pathway is dysfunctional in sporadic basal‐like cancers (Turner et al., 2007; Silver et al., 2010). Engineered mouse models targeting Brca1 and Trp53 genes in luminal or basal cells of the mouse mammary gland resulted in the development of tumours displaying morphological and immunohistochemical features that recapitulate those seen in human basal‐like breast carcinomas (Liu et al., 2007; McCarthy et al., 2007), which we will discuss below in greater detail.
In the past years, at least 3 additional ER‐negative molecular subtypes have been described: the ‘molecular apocrine’ group of tumours, which has been claimed by some to be similar to the HER2 subtype and appears to have activation of the androgen receptor signalling (Farmer et al., 2005b; Doane et al., 2006); the ‘interferon’ subtype, which is characterised by high expression of interferon regulated‐genes, including STAT1 (Hu et al., 2006); and the ‘claudin‐low’ subgroup, which comprises tumours that have transcriptomic features suggestive of a ‘cancer stem cell‐like’ phenotype (Herschkowitz et al., 2007; Hennessy et al., 2009). It should be noted that the clinical and biological significance of tumours pertaining to these newly described classes remains to be determined.
Despite the claim that expression profiling should be regarded as the ‘gold standard’ for breast cancer taxonomy (Peppercorn et al., 2008), the validity of this taxonomy (Gusterson, 2009), the stability of the molecular subgroups identified by microarray analysis (Pusztai et al., 2006) and their use in the management of breast cancer patients have been called into question (Weigelt et al., 2010, 2010). Perhaps it would be best to consider the current molecular taxonomy as a working model in development and not a final, definitive classification system, given that further molecular subtypes have been and may be identified. In addition, in each publication, different ‘intrinsic gene lists’ were employed and slightly different subtypes emerged (e.g. three categories of luminal cancers in some studies (Sorlie et al., 2001; Sotiriou et al., 2003)), an interferon‐rich group in another (Hu et al., 2006), and the claudin‐low subtype in two recent studies (Herschkowitz et al., 2007; Hennessy et al., 2009). Moreover, normal breast‐like tumours were part of the ER‐negative branch in some studies (Perou et al., 2007, 2000, 2001, 2003) and of the ER‐positive in others (Hu et al., 2006; Parker et al., 2009). The subjective nature of how the molecular subtypes were identified, the moderate stability of each cluster, and the lack of a definitive study demonstrating the reproducibility of each molecular subtype limit the application of this taxonomy in diagnostic practice (Pusztai et al., 2006). We performed a comparison of three different methods for classification of individual samples using three single sample predictors (Sorlie et al., 2003; Hu et al., 2006; Parker et al., 2009) that revealed a modest agreement between different methods and that only the patients with basal‐like cancers could be reproducibly identified (Weigelt et al., 2010a).
The current molecular taxonomy claims to provide accurate portraits of breast cancer that account for its diversity, however histological special types have not been systematically investigated, probably due to their low prevalence and limited availability of fresh/frozen samples. In fact, Perou et al. (2000) analysed only IDC‐NSTs and two invasive lobular carcinomas. Hence, it is rather arguable that these portraits could ‘accommodate all characters in the picture’.
The question of whether this taxonomy could be applied to histological special types of breast cancer has been recently addressed (Weigelt et al., 2006, 2008, 2005, 2003, 2007, 2007, 2008, 2009, 2004). Microarray‐based analysis of 11 special types of breast cancer has demonstrated that at the transcriptomic level each special type of breast cancer is composed of tumours that are more homogeneous than IDC‐NSTs and invasive lobular carcinomas (Weigelt et al., 2008). Hierarchical cluster analysis revealed that each histological special type studied pertained to only one molecular subtype with the exception of apocrine carcinomas. For example, tubular, mucinous and neuroendocrine carcinomas consistently displayed a luminal phenotype, whereas adenoid cystic, medullary and metaplastic carcinomas a basal‐like phenotype (Weigelt et al., 2008), in agreement with previous studies (Livasy et al., 2006; Reis‐Filho et al., 2006b; Bertucci et al., 2006; Jacquemier et al., 2005; Vincent‐Salomon et al., 2007; Azoulay et al., 2005) (Fig. 3). However, none of the tumours were of normal breast‐like subtype and 6% of special types of breast cancer displayed a molecular apocrine phenotype (Weigelt et al., 2008).
Figure 3.
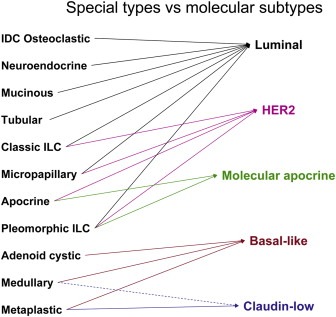
Histological special types and molecular subtypes. Tumours from each of the histological special types are more homogeneous than invasive ductal carcinomas (IDC) of no special type. IDC with osteoclastic‐like giant cells, mucinous and tubular carcinomas preferentially harbour a luminal phenotype, whilst adenoid cystic, medullary and metaplastic breast cancers preferentially display a basal‐like phenotype. Some metaplastic breast cancers and medullary carcinomas may display a claudin‐low phenotype. Classic lobular and micropapillary carcinomas are preferentially of luminal or HER2 phenotypes. Apocrine carcinomas are either of molecular apocrine or HER2 phenotype, whereas pleomorphic lobular carcinomas may be of luminal, HER2 or molecular apocrine phenotype. The dashed line represents a hypothetical association. Note that the HER2 group here is defined by the presence of HER2 gene amplification and not the ‘intrinsic gene’ defined HER2 group.
Interestingly, although special types of breast cancer could be classified into molecular subgroups, supervised analyses comparing special types of a given molecular subtype with IDC‐NSTs of the same molecular subtype have consistently revealed important genetic and transcriptomic differences (Weigelt et al., 2010c; Weigelt and Reis‐Filho, 2009). For instance, when metaplastic breast carcinomas were compared to basal‐like IDC‐NSTs, it was demonstrated that DNA repair pathways, including the BRCA1 DNA damage response pathway, PTEN, a gene whose loss of function is associated with resistance to chemotherapy, and TOP2A, the molecular target of anthracyclines, were significantly downregulated in metaplastic breast cancers (Weigelt et al., 2009a; Podo et al., 2010). These findings may explain the reported poor responses to chemotherapy of this histological subtype. In addition, metaplastic cancers were found to show significantly higher expression of genes related to myoepithelial differentiation and epithelial‐to‐mesenchymal transition compared to IDC‐NSTs of basal‐like molecular subtype (Lien et al., 2007; Weigelt et al., 2009a). Furthermore, mutations of the β‐catenin gene and genetic activation of the Wnt pathway were reported to be more prevalent in metaplastic breast cancers (Hayes et al., 2008); however these results were not confirmed in subsequent analyses (Hennessy et al., 2009; Geyer et al., unpublished data). Contrary to this body of evidence, Hennessy et al. have recently suggested that metaplastic breast carcinomas may not be of basal‐like phenotype (Hennessy et al., 2009). Out of 12 cases subjected to hierarchical clustering analysis, six samples formed a distinct subgroup, and the remaining cases clustered together with basal‐like (n = 2), normal breast‐like (n = 2) or claudin‐low groups (n = 2) (Hennessy et al., 2009). In addition, a high prevalence (53%) of mutations affecting the PI3K/AKT pathway was also reported in metaplastic breast carcinomas (Hennessy et al., 2009). Further studies with larger sample sizes and utilising objective methods for the classification of metaplastic breast cancers are required to reconcile these discrepancies. Based on the clinically relevant evidence available to date, it would be more appropriate to consider metaplastic breast carcinomas as a heterogeneous subgroup of triple negative breast cancers.
Medullary carcinomas and IDC‐NSTs of basal‐like phenotype have also been shown to have distinct molecular characteristics (Weigelt et al., 2008; Bertucci et al., 2006; Jacquemier et al., 2005; Vincent‐Salomon et al., 2007). Gene expression profiling analysis of medullary breast cancers revealed an upregulation of genes involved in TH1 immune response, including interleukins, transcription factors involved in TH1 differentiation, IFN regulatory factors and TH1 cytokines, as well as genes related to the apoptosis pathway, including members of the TNF receptor and ligand family, compared to basal‐like IDC‐NSTs (Weigelt et al., 2008; Bertucci et al., 2006). Conversely, genes involved in the architecture and remodelling of the cytoskeleton, and genes associated with cell invasiveness were downregulated in medullary carcinomas, which altogether may account for the reported favourable outcome of this ER‐negative histological special type (Weigelt et al., 2008; Bertucci et al., 2006).
Adenoid cystic carcinomas, although consistently displaying a basal‐like phenotype (Weigelt et al., 2008; Azoulay et al., 2005), have distinct morphological features (Ellis et al., 2003), clinical presentation and show downregulation of genes related to cell migration, proliferation and immune response (Weigelt et al., 2008), which in combination with their low histological grade may explain their excellent prognosis. In a way akin to adenoid cystic carcinomas of the breast, secretory carcinomas also have an indolent clinical behaviour and consistently display a triple negative and basal‐like phenotype (Lae et al., 2009; Lambros et al., 2009). Interestingly, these special histological types of breast cancer harbour recurrent balanced chromosomal translocations which lead to fusion transcripts, namely the t(12;15)(p13;q25) ETV6–NTRK3 fusion gene in secretory carcinomas (Tognon et al., 2002) and the t(6;9)(q22–23; p23–24) MYB–NFIB fusion gene in adenoid cystic carcinomas (Persson et al., 2009). Given that a recent massively parallel sequencing study identified multiple somatic rearrangements leading to the formation of fusion genes in breast cancers, it is plausible that distinct special types of breast cancer may be underpinned by somatic rearrangements leading to the formation of specific fusion genes (Stephens et al., 2009), similar to specific subtypes of renal tumours, aggressive midline carcinomas, soft tissue tumours, and lymphomas and leukaemias (Mitelman et al., 2007). Studies to address this hypothesis are eagerly awaited.
3. Cell of origin
There are several lines of evidence that cancer classifications may not reflect tumour histogenesis but tumour differentiation (Gusterson et al., 2005; Gusterson, 2009; Gould, 1986; Zhang et al., 2001; Moinfar, 2008). Nevertheless, the proponents of the molecular classification for breast cancer envisaged that the molecular subgroups identified by microarray analysis would indeed have histogenetic implications (i.e. that the phenotype of a breast cancer would mirror that of its cell of origin) (Sorlie et al., 2001). Sorlie et al. (2001) suggested that luminal and basal‐like cancers would originate from luminal and basal cells, respectively.
The identification of normal adult stem cells in the mouse mammary gland (reviewed in Stingl (2009)) and of tumourigenic cells within human breast tumours by means of cell surface markers and transplantation assays in immunodeficient mice has fuelled the concept that cancers are initiated and maintained by a population of tumour cells with stem cell‐like features (i.e. ‘cancer stem cells’) (Stingl, 2009; Al‐Hajj et al., 2003). This ‘cancer stem cell’ hypothesis further suggests that the type of genetic event and the cell of origin, either breast stem cells or their progenies, could be the underlying causes for the morphological and molecular heterogeneity found in breast cancers (Vargo‐Gogola and Rosen, 2007; Polyak, 2007; Behbod and Rosen, 2005; Dontu et al., 2004; Stingl and Caldas, 2007). It should be noted, however, that the intra‐tumour phenotypic heterogeneity observed in breast cancers may also stem from the existence of intra‐tumour genetic heterogeneity (Marusyk and Polyak, 2009; Torres et al., 2007; Geyer et al., 2010).
According to this model, it has been proposed that each of the five molecular breast cancer subtypes (i.e. basal‐like, HER2, normal breast‐like, luminal A, luminal B) might initiate in different types of stem/progenitor cells. These five cell types of origin would not only be of distinct cell lineages, as hypothesised by Sorlie et al. (2003), but also of different stages of mammary epithelial cell differentiation. For instance, a transforming event in an ER‐negative mammary stem cell would give rise to ER‐negative basal‐like or HER2 breast cancers, whereas a genetic aberration in a more differentiated progenitor would give rise to ER‐positive luminal breast cancers (Behbod and Rosen, 2005; Dontu et al., 2004). Alternatively, it has been suggested that luminal A tumours, which have a favourable prognosis, would originate from well‐differentiated cells of the ER‐positive lineage, whereas luminal B tumours, which show lower levels of ER expression and have a poorer prognosis than luminal A tumours, from more primitive ER‐positive progenitors (Stingl and Caldas, 2007). Likewise, the histological types of breast cancer have been suggested to be initiated in different types of stem or progenitor cells (Stingl and Caldas, 2007). However, there is limited direct experimental evidence to support these hypotheses. Furthermore, a recent study from Lim et al. (2009) provided strong evidence to suggest that tumours arising in BRCA1 mutation carriers and basal‐like breast cancers may, in fact, originate from luminal progenitor cells rather than ‘basal’ cells (Lim et al., 2009). Although some have argued that these findings would not impact on the use of the basal‐like nomenclature (Prat and Perou, 2009), others have claimed that basal‐like is a misnomer, given that these tumours neither display pure basal differentiation nor originate from basal progenitor cells (Weigelt et al., 2010c; Gusterson et al., 2005; Gusterson, 2009; Moinfar, 2008).
4. Genotypic–phenotypic correlations
The concept of clonal evolution explains the heterogeneity of breast cancer as a result of various distinct combinations of mutations acquired by a random cell over time, which provides a selective growth advantage under a specific set of selective pressures (Polyak, 2007; Marusyk and Polyak, 2009). It should be noted that the cancer stem cell and the Darwinian clonal evolution hypotheses are by no means mutually exclusive (Marusyk and Polyak, 2009; Geyer et al., 2010). The interplay of these combinations of genetic and also epigenetic aberrations and the cell of origin may provide the basis for the vast number of histological types of breast cancer (Polyak, 2007). In fact, there is substantial evidence to suggest that at least some breast cancer special types are underpinned by distinct arrays of genetic and epigenetic aberrations (Weigelt et al., 2008, 2008, 2002, 2007, 2008, 2010, 2009), which would ultimately determine not only their morphological features, but also their clinical behaviour.
There are several lines of evidence demonstrating that at least some histological special types of breast cancer are characterised by distinct genetic alterations (Hennessy et al., 2009, 2009, 2008, 2009, 2009, 2002, 2002). As mentioned above, adenoid cystic carcinomas, regardless of the anatomical site of origin, harbour the t(6;9)(q22–23;p23–24) involving the genes MYB and NFIB (Persson et al., 2009), whereas secretory carcinomas of the breast have been shown to harbour the t(12;15)(p13;q25) chromosomal translocation involving the genes ETV6 and NTRK3 (Tognon et al., 2002). This translocation leads to the formation of a fusion gene ETV6–NTRK3, which has been shown to have transformation activity in mouse mammary gland myoepithelial and epithelial cells (Tognon et al., 2002). Although the t(12;15)(p13;q25) can be also found in other human cancers (e.g. cellular mesoblastic nephroma, congenital fibrosarcoma and acute myeloid leukaemia), in the context of breast malignancies, this translocation seems to be specific to secretory carcinomas (Letessier et al., 2005; Makretsov et al., 2004; Reis‐Filho et al., 2008). Taken together, these two fusion genes (MYB–NFIB and ETV6–NTRK3) provide clear examples of phenotypic–genotypic correlations in breast cancer.
Genome‐wide genomic and transcriptomic studies have demonstrated that some histological special types of breast cancer are more than mere architectural patterns. In fact, apart from adenoid cystic and secretory carcinomas, also micropapillary carcinomas, metaplastic carcinomas, lobular carcinomas, mucinous carcinomas and neuroendocrine carcinomas have been shown to constitute entities distinct from IDC‐NSTs at the molecular level (Hennessy et al., 2008, 2009, 2008, 2009, 2008, 2009, 2010). For example, micropapillary carcinomas are characterised by the nests of cells with an inside‐out growth pattern. These tumours have been shown to have a higher proclivity to disseminate to axillary lymph nodes and a more aggressive clinical behaviour than IDC‐NSTs (Nassar, 2004). Invasive micropapillary carcinomas have been shown to preferentially be of luminal B molecular subtype (Weigelt et al., 2008; Marchio et al., 2008). However, when analysed by mircoarray‐based gene expression hierarchical clustering together with other special types of breast cancer, micropapillary cancers appear to form an almost exclusive cluster (Weigelt et al., 2008). Moreover, genomic analysis using comparative genomic hybridisation revealed that micropapillary tumours are characterised by a constellation of copy number aberrations that are distinct from those found in ER‐matched IDC‐NSTs of the same histological grade (Marchio et al., 2008; Thor et al., 2002). These observations suggest that micropapillary histology is not only a histological pattern, but actually a discrete morphological and molecular entity. Interestingly, when the morphologically distinct components of mixed micropapillary carcinomas (i.e. tumours with micropapillary and IDC‐NST areas) were subjected to genomic and immunohistochemical analysis, striking similarities were observed at the genetic and immunohistochemical level. When compared to IDC‐NSTs matched by grade and ER status, mixed micropapillary carcinomas displayed significant differences in the type and pattern of copy number changes and higher proliferative rates (Marchio et al., 2009). In fact, the genetic aberrations found in the ductal components of mixed micropapillary carcinomas were more similar to those of pure micropapillary carcinomas than to those of grade‐ and ER‐matched IDC‐NSTs (Marchio et al., 2009). These observations suggest that micropapillary differentiation in breast tumours, even in those featuring a mixed histology may identify a subgroup of more aggressive ER‐positive breast carcinomas, that mixed micropapillary carcinomas are closely related to pure micropapillary carcinomas and that the characteristic morphological features observed in micropapillary cancers may stem from a combination of genetic changes and epigenetic hits.
The hypothesis of genetic aberrations leading to the development of cancers of a specific morphological type is given further credence by conditional mouse models. The most common histological special type invasive lobular carcinoma (Table 1) is characterised by the growth of discohesive cells infiltrating the breast stroma as single cells or arranged in single cell files (Ellis et al., 2003; Weigelt et al., 2010b; Hanby and Hughes, 2008). The outcome of patients with lobular carcinomas is similar to that of patients with IDC‐NST, however, lobular cancers are often multifocal, have a distinct metastatic pattern (i.e. often metastasise to serosal surfaces, gastrointestinal tract and gynaecological organs) and less often respond to neoadjuvant chemotherapy (Rosen, 2001; Ellis et al., 2003). Several correlative studies have demonstrated that lobular carcinomas are characterised by the inactivation of the CDH1 gene, which encodes E‐cadherin, an adhesion molecule that mediates homophilic–homotypic adhesions (Vos et al., 1997). Our group performed a microarray analysis of invasive lobular carcinomas and grade‐ and molecular subtype‐matched IDC‐NSTs, which revealed that these tumours differ in the expression of not only E‐cadherin, but also genes related to cell adhesion, cell‐to‐cell signalling, and actin cytoskeleton signalling (Weigelt et al., 2010b). Several molecular mechanisms have been shown to lead to CDH1 inactivation in lobular carcinomas (Droufakou et al., 2001; Sarrio et al., 2003), including gene deletion, inactivating gene mutation, gene promoter methylation or transcriptional silencing. This has led several groups to hypothesise that lack of E‐cadherin expression would account for the so characteristic discohesiveness of lobular carcinoma cells and could determine the metastatic pattern of these lesions. Histological and molecular analyses of the K14cre;Cdh1 F/F/Trp53 F/F conditional mouse model revealed that these animals developed mammary tumours that were strikingly similar to human invasive lobular carcinomas (Derksen et al., 2006). Tumours from this conditional mouse model were often multifocal and characterised by monomorphic atypical neoplastic cells infiltrating the stroma as single cells and single cell files. Like human lobular carcinomas, K14cre;Cdh1 F/F/Trp53 F/F tumours also metastasised to gastrointestinal and urogenital tracts or diffusely disseminated throughout the peritoneal cavity (Ellis et al., 2003; Derksen et al., 2006). It should be noted that tumours from the K14cre;Cdh1 F/F/Trp53 F/F model also displayed important differences when compared to human lobular cancers, including lack of ER expression, presence of Trp53 inactivation, expression of ‘basal’ keratins, and lack of associated in situ lobular neoplasia.
Metaplastic breast cancer is a histological special type comprising tumours characterised by a complex admixture of adenocarcinomatous areas with metaplastic elements, which can be subdivided in homologous (i.e. squamous and spindle metaplasia) or heterologous (i.e. chondroid, osteous and rhabdomyoid metaplasia) (Huvos et al., 1973). Our group (Reis‐Filho et al., 2006, 2006, 2009) and others (Leibl et al., 2005; Leibl and Moinfar, 2005) have demonstrated that >90% of these cancers lack expression of ER and HER2, and display a basal‐like immunoprofile and transcriptome. Although a recent study has suggested that these tumours may be more heterogeneous at the transcriptomic level and enriched for cells with features of epithelial‐to‐mesenchymal transition and stem cell characteristics (Hennessy et al., 2009), metaplastic breast cancers in a way akin to IDC‐NSTs of basal‐like phenotype appear to have a dysfunctional BRCA1 pathway (Turner and Reis‐Filho, 2006; Turner et al., 2007; Silver et al., 2010; Kwei et al., 2010) and in >60% of these cancers, BRCA1 is downregulated due to gene promoter methylation (Turner et al., 2007). Based on the fact that the majority of basal‐like breast cancers show a dysfunctional BRCA1 pathway (Turner and Reis‐Filho, 2006; Turner et al., 2007; Weigelt et al., 2009a) and harbour TP53 gene mutations (Sorlie et al., 2001; Lien et al., 2004; Manie et al., 2009), we have engineered the conditional mouse Blg‐Cre;Brca1 F22–24/F22–24 ;Trp53 +/−, where the Brca1 gene is inactivated in β‐lactoglobulin‐expressing cells (i.e. luminal epithelial cells of the mouse mammary gland) and all cells of the animal have only one wild‐type allele of Trp53 (McCarthy et al., 2007). Consistent with the histological and immunophenotypic characteristics of human metaplastic breast cancers, pathological analysis of the tumours arising in the above mice revealed that 78% lacked expression of hormone receptors and HER2 and expressed basal markers (cytokeratins 14 and/or EGFR) and 88% showed homologous metaplastic elements (McCarthy et al., 2007) (Fig. 4). This mouse model (McCarthy et al., 2007) provides another line of evidence (Turner et al., 2007) for the link between basal‐like phenotype and BRCA1 pathway dysfunction and may prove useful for testing novel therapies for basal‐like cancers. Interestingly, another conditional mouse model K14cre;Brca1 F/F ;Trp53 F/F, where Brca1 and Trp53 were inactivated in basal cells of the mouse mammary gland (Liu et al., 2007), has been shown lead to the development of tumours whose morphological and phenotypic characteristics are remarkably similar to those observed in our study. Although the design of these animal models cannot provide definitive comments about the exact cell of origin of the tumours, these findings suggest that independent of the cell type in which Brca1 and Trp53 are inactivated, the concurrent inactivation of these genes may lead to the development of basal‐like breast cancers often with metaplastic elements (Fig. 4). This should not come as a surprise, given that BRCA1 has been shown to play a pivotal role in the regulation of ER expression and that RNA interference‐mediated silencing of BRCA1 in breast cancer cell lines leads to a marked reduction in expression of endogenous levels of ER (Hosey et al., 2007) and in increased expression of markers usually found in basal/myoepithelial cells (e.g. cytokeratins 5 and 17, and P‐cadherin) (Gorski et al., 2009; Mullan, 2006).
Figure 4.
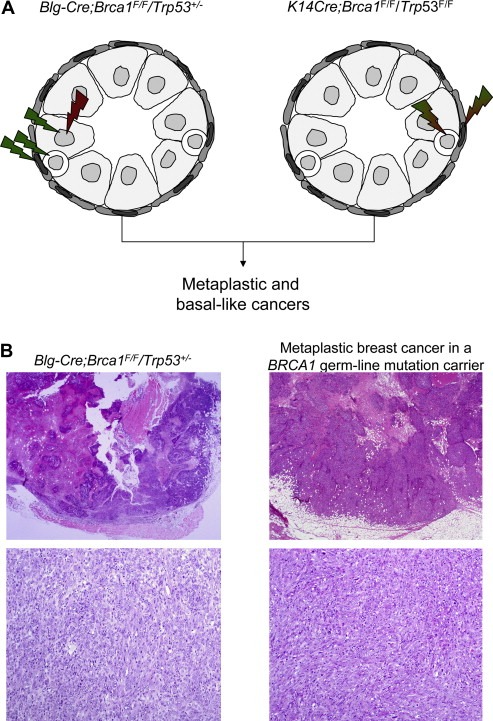
Histological special types may be driven by specific genomic hits regardless of the target cells. (A) In the Blg‐Cre;Brca1F/F/Trp53+/− mouse model, Brca1 was inactivated in luminal epithelial cells of the mouse mammary gland and all cells harboured only one functional copy of Trp53. In the K14cre;Brca1F/F;Trp53F/F mouse model, Brca1 and Trp53 were inactivated in the basal/myoepithelial cells of the mouse mammary gland. Histopathological and immunohistochemical analysis of tumours developing in mouse models where Brca1 and Trp53 were inactivated in different lineages of the mouse mammary gland revealed that tumours had morphological and immunohistochemical features that recapitulated those of human basal‐like breast cancers (Liu et al., 2007; McCarthy et al., 2007) (i.e. a convergent phenotype driven by the type of genetic hits). (B) Representative scanning and medium power magnification micrographs of a tumour developing in Blg‐Cre;Brca1F/F/Trp53+/− mouse model and in a patient with a pathogenic BRCA1 truncating germ‐line mutation.
Transcriptomic and genetic analysis of breast cancers has also helped understand the similarities in clinical presentation and outcome of some special types of breast cancer (Simpson et al., 2005a; Correa Geyer and Reis‐Filho, 2009). Low grade IDC‐NSTs, tubular, cribriform, tubulo‐lobular and classic lobular carcinomas are tumours that albeit having distinctive histological features display (i) similar clinical presentation and natural history, (ii) are associated with the same family of precursor and pre‐invasive lesions (i.e. flat epithelial atypia, atypical ductal hyperplasia, lobular neoplasia and low grade ductal carcinoma in situ), (iii) have similar immunophenotypes (i.e. consistent expression of hormone receptors and lack of HER2 expression), and (iv) are characterised by similar types and patterns of genetic aberrations (namely, diploid/near diploid karyotypes characterised by deletion of 16q coupled with gain of 1q and 16p, often resultant from a der(16)t(1; 16)/der(1; 16) unbalanced chromosomal translocation) (Simpson et al., 2007, 2008, 1998, 2005, 2005, 1999). These findings have led several groups to posit that low grade IDC‐NSTs, tubular and cribriform carcinomas would be more similar to classic lobular carcinomas than to high grade IDC‐NSTs and that these lesions and their respective precursors should be considered as members of a family of interrelated lesions (i.e. low grade breast neoplasia family) (Abdel‐Fatah et al., 2008). It should be noted, however, that although the main difference between lobular lesions and the other tumours has already been identified (i.e. the target gene of 16q deletions: whilst in lobular carcinomas it has been shown to be the CDH1 gene, in the remaining tumours the target gene is yet to be found (Simpson et al., 2005a)), the molecular differences between the other histological special types in this family of tumours are yet to be characterised.
Some of the morphological differences between special types of breast cancer cannot be explained by their patterns of genetic aberrations using current techniques. Tubular and cribriform carcinomas have strikingly similar genomic features by comparative genomic hybridisation; neuroendocrine and mucinous B carcinomas have undistinguishable transcriptomic profiles (Weigelt et al., 2008, 2009). Despite the paucity of evidence in support of a genetic basis to account for the full diversity of histological special types of breast cancer, it is possible that when these lesions are subjected to genome‐wide sequencing using massively parallel approaches a few pathognomonic mutations or specific fusion genes will account for their distinctive morphological features (Aparicio and Huntsman, 2010; Reis‐Filho, 2009). Alternatively, based on the evidence available to date, it is plausible that the diversity in the histological and phenotypic characteristics of special types of breast cancer may stem from a combination of distinct cells of origin and genetic/epigenetic aberrations (Weigelt and Reis‐Filho, 2009).
5. Special types of breast cancer as models for identification of novel therapeutic targets
The molecular heterogeneity of chromosomal aberrations present in unselected groups of breast tumours has constituted a formidable challenge for the identification of the actual ‘drivers’ of breast cancers (i.e. genes causally implicated in the survival and growth of cancer cells) and novel therapeutic targets.
There are several lines of evidence to suggest that tumours from each histological special type of breast cancer have more homogeneous transcriptomic patterns than IDC‐NSTs (Weigelt et al., 2008, 2009, 2009, 2010), suggesting that each histological special type may be driven by a less diverse constellation of genetic and possibly also epigenetic events (Weigelt and Reis‐Filho, 2009; Reis‐Filho and Lakhani, 2008; Weigelt et al., 2008; Aparicio and Huntsman, 2010). We have hypothesised that by focusing on the analysis of special histological types of breast cancer, the molecular complexity and heterogeneity of breast cancer can be significantly reduced and that this may facilitate the identification of biological drivers of breast cancers and novel therapeutic targets for patients with specific types of breast cancer.
Based on this rationale, we have studied a series of classic lobular carcinomas using high resolution comparative genomic hybridisation and in situ hybridisation and identified recurrent amplifications on 8p11.2–p12, encompassing the FGFR1 locus (Reis‐Filho et al., 2006c). Through a detailed genomic and transcriptomic analysis of breast cancer cell lines, we identified a model for lobular carcinomas harbouring 8p11.2–p12 amplification (i.e. MDA‐MB‐134) and demonstrated through a combination of RNA interference and chemical inhibition that FGFR1 silencing is selectively lethal in cell lines harbouring this amplification, suggesting that FGFR1 is a potential therapeutic target for a subgroup of lobular cancers (Reis‐Filho et al., 2006c). Analysis of large cohorts of breast cancer patients using either Southern blotting (Cuny et al., 2000), chromogenic in situ hybridisation (Courjal et al., 1997; Elbauomy Elsheikh et al., 2007) or microarray‐based comparative genomic hybridisation (Natrajan et al., 2009; Adelaide et al., 2007) has revealed that FGFR1 amplification is present in approximately 10% of all breast cancers, but is more frequently found in ER‐positive disease. In ER‐positive breast cancers, FGFR1 gene amplification is associated with lower levels of progesterone receptor expression and higher Ki‐67 labelling indices (Turner et al., 2010), and has been shown to be both a prognostic factor independent of size, grade and nodal stage in ER‐positive breast cancer patients treated with endocrine therapy (Elbauomy Elsheikh et al., 2007). Furthermore, a recent study (Turner et al., 2010) has demonstrated that breast cancer cell lines harbouring FGFR1 amplification display resistance to Tamoxifen in vitro, which is reversed by FGFR1 RNA interference silencing.
Understanding the underlying molecular features of special types of breast cancer may also provide new avenues for improvement of the type of adjuvant systemic therapy offered to breast cancer patients. As described above, metaplastic breast cancers have been shown to express low levels of Topoisomerase IIα, PTEN and BUB1 (Weigelt et al., 2009a) and to frequently harbour mutations that may lead to activation of the PI3K/AKT pathway (Hennessy et al., 2009); therefore, it is not surprising that these tumours do not respond to conventional chemotherapy regimens based on anthracycline and spindle cell poison therapies (Hennessy et al., 2006). On the other hand, transcriptomic analysis and conditional mouse models indicate that these cancers are likely to harbour a dysfunctional BRCA1 pathway. Seminal studies carried out by Alan Ashworth's lab have demonstrated that tumours with BRCA1, BRCA2 or PTEN loss of function lack a competent homologous recombination DNA repair and, therefore, show an exquisite sensitivity to cross‐linking agents (e.g. platinum salts) and inhibitors of the PARP enzyme (Farmer et al., 2005a; Ashworth, 2008; Mendes‐Pereira et al., 2009), suggesting that these approaches may be more efficacious in metaplastic cancers.
6. Conclusions
Special types of breast cancer account for up to one quarter of all invasive breast malignancies and their importance should not be disregarded. Studies focusing on specific subtypes of carcinomas from several sites have recently identified pathognomonic mutations (Shah et al., 2009a) and specific fusion genes (Tognon et al., 2002; Persson et al., 2009; Mitelman et al., 2007; French, 2008; Brenner and Chinnaiyan, 2009) that can be used not only for diagnostic purposes, but also therapeutically. Understanding the biological drivers of these entities may lead to a better understanding of the biology of breast cancer. Fortunately, the concept of special types of breast cancer has recently been incorporated by the International Cancer Genome Consortium and other large scale resequencing exercises. In fact, the first complete genome of a breast cancer was from a metastasis from a special type of breast cancer! (Shah et al., 2009b). It is likely that in the next 5 years, a complete catalogue of the type, pattern and complexity of genetic and transcriptomic aberrations of special types of breast cancer will become available. Until then, instead of arguing whether one should be a ‘grader’ or a ‘typer’ (Pereira et al., 1995), pathologists should strive for developing a standardised classification for these fascinating entities, which may lead to a better understanding of their biology and, most importantly, their clinical behaviour.
Acknowledgment
The authors are grateful to Dr. Kay Savage for the excellent histological sections. BW is funded by a Cancer Research UK post‐doctoral fellowship, FCG and JSR‐F are funded in part by Breakthrough Breast Cancer.
Weigelt Britta, Geyer Felipe C., Reis-Filho Jorge S., (2010), Histological types of breast cancer: How special are they?, Molecular Oncology, 4, doi: 10.1016/j.molonc.2010.04.004.
References
- Abd El-Rehim, D.M. , Pinder, S.E. , Paish, C.E. , Bell, J. , Blamey, R.W. , Robertson, J.F. , Nicholson, R.I. , Ellis, I.O. , 2004. Expression of luminal and basal cytokeratins in human breast carcinoma. J. Pathol. 203, 661–671. [DOI] [PubMed] [Google Scholar]
- Abdel-Fatah, T.M. , Powe, D.G. , Hodi, Z. , Lee, A.H. , Reis-Filho, J.S. , Ellis, I.O. , 2007. High frequency of coexistence of columnar cell lesions, lobular neoplasia, and low grade ductal carcinoma in situ with invasive tubular carcinoma and invasive lobular carcinoma. Am. J. Surg. Pathol. 31, 417–426. [DOI] [PubMed] [Google Scholar]
- Abdel-Fatah, T.M. , Powe, D.G. , Hodi, Z. , Reis-Filho, J.S. , Lee, A.H. , Ellis, I.O. , 2008. Morphologic and molecular evolutionary pathways of low nuclear grade invasive breast cancers and their putative precursor lesions: further evidence to support the concept of low nuclear grade breast neoplasia family. Am. J. Surg. Pathol. 32, 513–523. [DOI] [PubMed] [Google Scholar]
- Adelaide, J. , Finetti, P. , Bekhouche, I. , Repellini, L. , Geneix, J. , Sircoulomb, F. , Charafe-Jauffret, E. , Cervera, N. , Desplans, J. , Parzy, D. , Schoenmakers, E. , Viens, P. , Jacquemier, J. , Birnbaum, D. , Bertucci, F. , Chaffanet, M. , 2007. Integrated profiling of basal and luminal breast cancers. Cancer Res. 67, 11565–11575. [DOI] [PubMed] [Google Scholar]
- Al-Hajj, M. , Wicha, M.S. , Benito-Hernandez, A. , Morrison, S.J. , Clarke, M.F. , 2003. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. U.S.A. 100, 3983–3988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aparicio, S.A. , Huntsman, D.G. , 2010. Does massively parallel DNA resequencing signify the end of histopathology as we know it?. J. Pathol. 220, 307–315. [DOI] [PubMed] [Google Scholar]
- Ashworth, A. , 2008. A synthetic lethal therapeutic approach: poly(ADP) ribose polymerase inhibitors for the treatment of cancers deficient in DNA double-strand break repair. J. Clin. Oncol. 26, 3785–3790. [DOI] [PubMed] [Google Scholar]
- Azoulay, S. , Lae, M. , Freneaux, P. , Merle, S. , Al Ghuzlan, A. , Chnecker, C. , Rosty, C. , Klijanienko, J. , Sigal-Zafrani, B. , Salmon, R. , Fourquet, A. , Sastre-Garau, X. , Vincent-Salomon, A. , 2005. KIT is highly expressed in adenoid cystic carcinoma of the breast, a basal-like carcinoma associated with a favorable outcome. Mod. Pathol. 18, 1623–1631. [DOI] [PubMed] [Google Scholar]
- Azzopardi, J.G. , Ahmed, A. , Millis, R.R. , 1979. Problems in Breast Pathology W.B. Saunders Company Ltd; London: [PubMed] [Google Scholar]
- Azzopardi, J.G. , Chepick, O.F. , Hartmann, W.H. , Jafarey, N.A. , Llombart-Bosch, A. , Ozzello, L. , Rilke, F. , Sasano, N. , Sobin, L.H. , Sommers, S.C. , Stalsberg, H. , Sugar, J. , Williams, A.O. , 1982. The world health organization histological typing of breast tumors – second edition. The World Organization. Am. J. Clin. Pathol. 78, 806–816. [DOI] [PubMed] [Google Scholar]
- Behbod, F. , Rosen, J.M. , 2005. Will cancer stem cells provide new therapeutic targets?. Carcinogenesis. 26, 703–711. [DOI] [PubMed] [Google Scholar]
- Bertucci, F. , Finetti, P. , Cervera, N. , Charafe-Jauffret, E. , Mamessier, E. , Adelaide, J. , Debono, S. , Houvenaeghel, G. , Maraninchi, D. , Viens, P. , Charpin, C. , Jacquemier, J. , Birnbaum, D. , 2006. Gene expression profiling shows medullary breast cancer is a subgroup of basal breast cancers. Cancer Res. 66, 4636–4644. [DOI] [PubMed] [Google Scholar]
- Bertucci, F. , Orsetti, B. , Negre, V. , Finetti, P. , Rouge, C. , Ahomadegbe, J.C. , Bibeau, F. , Mathieu, M.C. , Treilleux, I. , Jacquemier, J. , Ursule, L. , Martinec, A. , Wang, Q. , Benard, J. , Puisieux, A. , Birnbaum, D. , Theillet, C. , 2008. Lobular and ductal carcinomas of the breast have distinct genomic and expression profiles. Oncogene. 27, 5359–5372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brenner, J.C. , Chinnaiyan, A.M. , 2009. Translocations in epithelial cancers. Biochim. Biophys. Acta. 1796, 201–215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carey, L.A. , Perou, C.M. , Livasy, C.A. , Dressler, L.G. , Cowan, D. , Conway, K. , Karaca, G. , Troester, M.A. , Tse, C.K. , Edmiston, S. , Deming, S.L. , Geradts, J. , Cheang, M.C. , Nielsen, T.O. , Moorman, P.G. , Earp, H.S. , Millikan, R.C. , 2006. Race, breast cancer subtypes, and survival in the Carolina Breast Cancer Study. JAMA. 295, 2492–2502. [DOI] [PubMed] [Google Scholar]
- Correa Geyer, F. , Reis-Filho, J.S. , 2009. Microarray-based gene expression profiling as a clinical tool for breast cancer management: are we there yet?. Int. J. Surg. Pathol. 17, 285–302. [DOI] [PubMed] [Google Scholar]
- Courjal, F. , Cuny, M. , Simony-Lafontaine, J. , Louason, G. , Speiser, P. , Zeillinger, R. , Rodriguez, C. , Theillet, C. , 1997. Mapping of DNA amplifications at 15 chromosomal localizations in 1875 breast tumors: definition of phenotypic groups. Cancer Res. 57, 4360–4367. [PubMed] [Google Scholar]
- Cuny, M. , Kramar, A. , Courjal, F. , Johannsdottir, V. , Iacopetta, B. , Fontaine, H. , Grenier, J. , Culine, S. , Theillet, C. , 2000. Relating genotype and phenotype in breast cancer: an analysis of the prognostic significance of amplification at eight different genes or loci and of p53 mutations. Cancer Res. 60, 1077–1083. [PubMed] [Google Scholar]
- Derksen, P.W. , Liu, X. , Saridin, F. , van der Gulden, H. , Zevenhoven, J. , Evers, B. , van Beijnum, J.R. , Griffioen, A.W. , Vink, J. , Krimpenfort, P. , Peterse, J.L. , Cardiff, R.D. , Berns, A. , Jonkers, J. , 2006. Somatic inactivation of E-cadherin and p53 in mice leads to metastatic lobular mammary carcinoma through induction of anoikis resistance and angiogenesis. Cancer Cell. 10, 437–449. [DOI] [PubMed] [Google Scholar]
- Doane, A.S. , Danso, M. , Lal, P. , Donaton, M. , Zhang, L. , Hudis, C. , Gerald, W.L. , 2006. An estrogen receptor-negative breast cancer subset characterized by a hormonally regulated transcriptional program and response to androgen. Oncogene. 25, 3994–4008. [DOI] [PubMed] [Google Scholar]
- Dontu, G. , El-Ashry, D. , Wicha, M.S. , 2004. Breast cancer, stem/progenitor cells and the estrogen receptor. Trends Endocrinol. Metab. 15, 193–197. [DOI] [PubMed] [Google Scholar]
- Droufakou, S. , Deshmane, V. , Roylance, R. , Hanby, A. , Tomlinson, I. , Hart, I.R. , 2001. Multiple ways of silencing E-cadherin gene expression in lobular carcinoma of the breast. Int. J. Cancer. 92, 404–408. [DOI] [PubMed] [Google Scholar]
- Elbauomy Elsheikh, S. , Green, A.R. , Lambros, M.B. , Turner, N.C. , Grainge, M.J. , Powe, D. , Ellis, I.O. , Reis-Filho, J.S. , 2007. FGFR1 amplification in breast carcinomas: a chromogenic in situ hybridisation analysis. Breast Cancer Res. 9, R23 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis, I.O. , Galea, M. , Broughton, N. , Locker, A. , Blamey, R.W. , Elston, C.W. , 1992. Pathological prognostic factors in breast cancer. II. Histological type. Relationship with survival in a large study with long-term follow-up. Histopathology. 20, 479–489. [DOI] [PubMed] [Google Scholar]
- Ellis, P. , Schnitt, S.J. , Sastre-Garau, X. , Bussolati, G. , Tavassoli, F.A. , Eusebi, V. , Peterse, J.L. , Mukai, K. , Tabar, L. , Jacquemier, J. , Cornelisse, C.J. , Sasco, A.J. , Kaaks, R. , Pisani, P. , Goldgar, D.E. , Devilee, P. , Cleton-Jansen, M.J. , Borresen-Dale, A.L. , van't Veer, L. , Sapino, A. , 2003. Invasive breast carcinoma. In Tavassoli F.A., Devilee P.(Eds.), WHO Classification of Tumours Pathology and Genetics of Tumours of the Breast and Female Genital Organs. Lyon Press; Lyon: [Google Scholar]
- Elston, C.W. , Ellis, I.O. , 1991. Pathological prognostic factors in breast cancer. I. The value of histological grade in breast cancer: experience from a large study with long-term follow-up. Histopathology. 19, 403–410. [DOI] [PubMed] [Google Scholar]
- Farmer, H. , McCabe, N. , Lord, C.J. , Tutt, A.N. , Johnson, D.A. , Richardson, T.B. , Santarosa, M. , Dillon, K.J. , Hickson, I. , Knights, C. , Martin, N.M. , Jackson, S.P. , Smith, G.C. , Ashworth, A. , 2005. Targeting the DNA repair defect in BRCA mutant cells as a therapeutic strategy. Nature. 434, 917–921. [DOI] [PubMed] [Google Scholar]
- Farmer, P. , Bonnefoi, H. , Becette, V. , Tubiana-Hulin, M. , Fumoleau, P. , Larsimont, D. , Macgrogan, G. , Bergh, J. , Cameron, D. , Goldstein, D. , Duss, S. , Nicoulaz, A.L. , Brisken, C. , Fiche, M. , Delorenzi, M. , Iggo, R. , 2005. Identification of molecular apocrine breast tumours by microarray analysis. Oncogene. 24, 4660–4671. [DOI] [PubMed] [Google Scholar]
- Flagiello, D. , Gerbault-Seureau, M. , Sastre-Garau, X. , Padoy, E. , Vielh, P. , Dutrillaux, B. , 1998. Highly recurrent der(1;16)(q10;p10) and other 16q arm alterations in lobular breast cancer. Genes Chromosomes Cancer. 23, 300–306. [PubMed] [Google Scholar]
- Foulkes, W.D. , Stefansson, I.M. , Chappuis, P.O. , Begin, L.R. , Goffin, J.R. , Wong, N. , Trudel, M. , Akslen, L.A. , 2003. Germline BRCA1 mutations and a basal epithelial phenotype in breast cancer. J. Natl. Cancer Inst. 95, 1482–1485. [DOI] [PubMed] [Google Scholar]
- French, C.A. , 2008. Molecular pathology of NUT midline carcinomas. J. Clin. Pathol. [DOI] [PubMed] [Google Scholar]
- Fulford, L.G. , Easton, D.F. , Reis-Filho, J.S. , Sofronis, A. , Gillett, C.E. , Lakhani, S.R. , Hanby, A. , 2006. Specific morphological features predictive for the basal phenotype in grade 3 invasive ductal carcinoma of breast. Histopathology. 49, 22–34. [DOI] [PubMed] [Google Scholar]
- Geyer, F.C. , Weigelt, B. , Natrajan, R. , Lambros, M.B.K. , de Biase, D. , Vatcheva, R. , Savage, K. , Mackay, A. , Ashworth, A. , Reis-Filho, J.S. , 2010. Molecular analysis reveals a genetic basis for the phenotypic diversity of metaplastic breast carcinomas. J. Pathol. 220, 561–573. [DOI] [PubMed] [Google Scholar]
- Gilbert, J.A. , Goetz, M.P. , Reynolds, C.A. , Ingle, J.N. , Giordano, K.F. , Suman, V.J. , Blair, H.E. , Jenkins, R.B. , Lingle, W.L. , Reinholz, M.M. , Adjei, A.A. , Ames, M.M. , 2008. Molecular analysis of metaplastic breast carcinoma: high EGFR copy number via aneusomy. Mol. Cancer Ther. 7, 944–951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorski, J.J., James, C.R., Quinn, J.E., Stewart, G.E., Staunton, K.C., Buckley, N.E., McDyer, F.A., Kennedy, R.D., Wilson, R.H., Mullan, P.B., Harkin, D.P., 2009. BRCA1 transcriptionally regulates genes associated with the basal-like phenotype in breast cancer. Breast Cancer Res. Treat., epub ahead of print. [DOI] [PubMed]
- Gould, V.E. , 1986. Histogenesis and differentiation: a re-evaluation of these concepts as criteria for the classification of tumors. Hum. Pathol. 17, 212–215. [DOI] [PubMed] [Google Scholar]
- Gruvberger, S. , Ringner, M. , Chen, Y. , Panavally, S. , Saal, L.H. , Borg, A. , Ferno, M. , Peterson, C. , Meltzer, P.S. , 2001. Estrogen receptor status in breast cancer is associated with remarkably distinct gene expression patterns. Cancer Res. 61, 5979–5984. [PubMed] [Google Scholar]
- Gusterson, B. , 2009. Do ‘basal-like’ breast cancers really exist?. Nat. Rev. Cancer. 9, 128–134. [DOI] [PubMed] [Google Scholar]
- Gusterson, B.A. , Ross, D.T. , Heath, V.J. , Stein, T. , 2005. Basal cytokeratins and their relationship to the cellular origin and functional classification of breast cancer. Breast Cancer Res. 7, 143–148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanby, A.M. , Hughes, T.A. , 2008. In situ and invasive lobular neoplasia of the breast. Histopathology. 52, 58–66. [DOI] [PubMed] [Google Scholar]
- Hayes, M.J. , Thomas, D. , Emmons, A. , Giordano, T.J. , Kleer, C.G. , 2008. Genetic changes of Wnt pathway genes are common events in metaplastic carcinomas of the breast. Clin. Cancer Res. 14, 4038–4044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hennessy, B.T. , Giordano, S. , Broglio, K. , Duan, Z. , Trent, J. , Buchholz, T.A. , Babiera, G. , Hortobagyi, G.N. , Valero, V. , 2006. Biphasic metaplastic sarcomatoid carcinoma of the breast. Ann. Oncol. 17, 605–613. [DOI] [PubMed] [Google Scholar]
- Hennessy, B.T. , Gonzalez-Angulo, A.M. , Stemke-Hale, K. , Gilcrease, M.Z. , Krishnamurthy, S. , Lee, J.S. , Fridlyand, J. , Sahin, A. , Agarwal, R. , Joy, C. , Liu, W. , Stivers, D. , Baggerly, K. , Carey, M. , Lluch, A. , Monteagudo, C. , He, X. , Weigman, V. , Fan, C. , Palazzo, J. , Hortobagyi, G.N. , Nolden, L.K. , Wang, N.J. , Valero, V. , Gray, J.W. , Perou, C.M. , Mills, G.B. , 2009. Characterization of a naturally occurring breast cancer subset enriched in epithelial-to-mesenchymal transition and stem cell characteristics. Cancer Res. 69, 4116–4124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herschkowitz, J.I. , Simin, K. , Weigman, V.J. , Mikaelian, I. , Usary, J. , Hu, Z. , Rasmussen, K.E. , Jones, L.P. , Assefnia, S. , Chandrasekharan, S. , Backlund, M.G. , Yin, Y. , Khramtsov, A.I. , Bastein, R. , Quackenbush, J. , Glazer, R.I. , Brown, P.H. , Green, J.E. , Kopelovich, L. , Furth, P.A. , Palazzo, J.P. , Olopade, O.I. , Bernard, P.S. , Churchill, G.A. , Van Dyke, T. , Perou, C.M. , 2007. Identification of conserved gene expression features between murine mammary carcinoma models and human breast tumors. Genome Biol. 8, R76 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosey, A.M. , Gorski, J.J. , Murray, M.M. , Quinn, J.E. , Chung, W.Y. , Stewart, G.E. , James, C.R. , Farragher, S.M. , Mulligan, J.M. , Scott, A.N. , Dervan, P.A. , Johnston, P.G. , Couch, F.J. , Daly, P.A. , Kay, E. , McCann, A. , Mullan, P.B. , Harkin, D.P. , 2007. Molecular basis for estrogen receptor alpha deficiency in BRCA1-linked breast cancer. J. Natl. Cancer Inst. 99, 1683–1694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu, Z. , Fan, C. , Oh, D.S. , Marron, J.S. , He, X. , Qaqish, B.F. , Livasy, C. , Carey, L.A. , Reynolds, E. , Dressler, L. , Nobel, A. , Parker, J. , Ewend, M.G. , Sawyer, L.R. , Wu, J. , Liu, Y. , Nanda, R. , Tretiakova, M. , Ruiz Orrico, A. , Dreher, D. , Palazzo, J.P. , Perreard, L. , Nelson, E. , Mone, M. , Hansen, H. , Mullins, M. , Quackenbush, J.F. , Ellis, M.J. , Olopade, O.I. , Bernard, P.S. , Perou, C.M. , 2006. The molecular portraits of breast tumors are conserved across microarray platforms. BMC Genomics. 7, 96 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huvos, A.G. , Lucas, J.C. , Foote, F.W. , 1973. Metaplastic breast carcinoma. Rare form of mammary cancer. N.Y. State J. Med. 73, 1078–1082. [PubMed] [Google Scholar]
- Ivshina, A.V. , George, J. , Senko, O. , Mow, B. , Putti, T.C. , Smeds, J. , Lindahl, T. , Pawitan, Y. , Hall, P. , Nordgren, H. , Wong, J.E. , Liu, E.T. , Bergh, J. , Kuznetsov, V.A. , Miller, L.D. , 2006. Genetic reclassification of histologic grade delineates new clinical subtypes of breast cancer. Cancer Res. 66, 10292–10301. [DOI] [PubMed] [Google Scholar]
- Jacquemier, J. , Padovani, L. , Rabayrol, L. , Lakhani, S.R. , Penault-Llorca, F. , Denoux, Y. , Fiche, M. , Figueiro, P. , Maisongrosse, V. , Ledoussal, V. , Martinez Penuela, J. , Udvarhely, N. , El Makdissi, G. , Ginestier, C. , Geneix, J. , Charafe-Jauffret, E. , Xerri, L. , Eisinger, F. , Birnbaum, D. , Sobol, H. , 2005. Typical medullary breast carcinomas have a basal/myoepithelial phenotype. J. Pathol. 207, 260–268. [DOI] [PubMed] [Google Scholar]
- Klingbeil, P. , Natrajan, R. , Everitt, G. , Vatcheva, R. , Marchio, C. , Palacios, J. , Buerger, H. , Reis-Filho, J.S. , Isacke, C.M. , 2010. CD44 is overexpressed in basal-like breast cancers but is not a driver of 11p13 amplification. Breast Cancer Res. Treat. 120, 95–109. [DOI] [PubMed] [Google Scholar]
- Korkola, J.E. , DeVries, S. , Fridlyand, J. , Hwang, E.S. , Estep, A.L. , Chen, Y.Y. , Chew, K.L. , Dairkee, S.H. , Jensen, R.M. , Waldman, F.M. , 2003. Differentiation of lobular versus ductal breast carcinomas by expression microarray analysis. Cancer Res. 63, 7167–7175. [PubMed] [Google Scholar]
- Kwei, K.A. , Kung, Y. , Salari, K. , Holcomb, I.N. , Pollack, J.R. , 2010. Genomic instability in breast cancer: Pathogenesis and clinical implications. Mol. Oncol. 4, (3) 255–266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lae, M. , Freneaux, P. , Sastre-Garau, X. , Chouchane, O. , Sigal-Zafrani, B. , Vincent-Salomon, A. , 2009. Secretory breast carcinomas with ETV6–NTRK3 fusion gene belong to the basal-like carcinoma spectrum. Mod. Pathol. 22, 291–298. [DOI] [PubMed] [Google Scholar]
- Lakhani, S.R. , Jacquemier, J. , Sloane, J.P. , Gusterson, B.A. , Anderson, T.J. , van de Vijver, M.J. , Farid, L.M. , Venter, D. , Antoniou, A. , Storfer-Isser, A. , Smyth, E. , Steel, C.M. , Haites, N. , Scott, R.J. , Goldgar, D. , Neuhausen, S. , Daly, P.A. , Ormiston, W. , McManus, R. , Scherneck, S. , Ponder, B.A. , Ford, D. , Peto, J. , Stoppa-Lyonnet, D. , Bignon, Y.J. , Struewing, J.P. , Spurr, N.K. , Bishop, D.T. , Klijn, J.G. , Devilee, P. , Cornelisse, C.J. , Lasset, C. , Lenoir, G. , Barkardottir, R.B. , Egilsson, V. , Hamann, U. , Chang-Claude, J. , Sobol, H. , Weber, B. , Stratton, M.R. , Easton, D.F. , 1998. Multifactorial analysis of differences between sporadic breast cancers and cancers involving BRCA1 and BRCA2 mutations. J. Natl. Cancer Inst. 90, 1138–1145. [DOI] [PubMed] [Google Scholar]
- Lakhani, S.R. , Van De Vijver, M.J. , Jacquemier, J. , Anderson, T.J. , Osin, P.P. , McGuffog, L. , Easton, D.F. , 2002. The pathology of familial breast cancer: predictive value of immunohistochemical markers estrogen receptor, progesterone receptor, HER-2, and p53 in patients with mutations in BRCA1 and BRCA2. J. Clin. Oncol. 20, 2310–2318. [DOI] [PubMed] [Google Scholar]
- Lakhani, S.R. , Reis-Filho, J.S. , Fulford, L. , Penault-Llorca, F. , van der Vijver, M. , Parry, S. , Bishop, T. , Benitez, J. , Rivas, C. , Bignon, Y.J. , Chang-Claude, J. , Hamann, U. , Cornelisse, C.J. , Devilee, P. , Beckmann, M.W. , Nestle-Kramling, C. , Daly, P.A. , Haites, N. , Varley, J. , Lalloo, F. , Evans, G. , Maugard, C. , Meijers-Heijboer, H. , Klijn, J.G. , Olah, E. , Gusterson, B.A. , Pilotti, S. , Radice, P. , Scherneck, S. , Sobol, H. , Jacquemier, J. , Wagner, T. , Peto, J. , Stratton, M.R. , McGuffog, L. , Easton, D.F. , 2005. Prediction of BRCA1 status in patients with breast cancer using estrogen receptor and basal phenotype. Clin. Cancer Res. 11, 5175–5180. [DOI] [PubMed] [Google Scholar]
- Lambros, M.B. , Tan, D.S. , Jones, R.L. , Vatcheva, R. , Savage, K. , Tamber, N. , Fenwick, K. , Mackay, A. , Ashworth, A. , Reis-Filho, J.S. , 2009. Genomic profile of a secretory breast cancer with an ETV6–NTRK3 duplication. J. Clin. Pathol. 62, 604–612. [DOI] [PubMed] [Google Scholar]
- Leibl, S. , Gogg-Kammerer, M. , Sommersacher, A. , Denk, H. , Moinfar, F. , 2005. Metaplastic breast carcinomas: are they of myoepithelial differentiation?: immunohistochemical profile of the sarcomatoid subtype using novel myoepithelial markers. Am. J. Surg. Pathol. 29, 347–353. [DOI] [PubMed] [Google Scholar]
- Leibl, S. , Moinfar, F. , 2005. Metaplastic breast carcinomas are negative for Her-2 but frequently express EGFR (Her-1): potential relevance to adjuvant treatment with EGFR tyrosine kinase inhibitors?. J. Clin. Pathol. 58, 700–704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Letessier, A. , Ginestier, C. , Charafe-Jauffret, E. , Cervera, N. , Adelaide, J. , Gelsi-Boyer, V. , Ahomadegbe, J.C. , Benard, J. , Jacquemier, J. , Birnbaum, D. , Chaffanet, M. , 2005. ETV6 gene rearrangements in invasive breast carcinoma. Genes Chromosomes Cancer. 44, 103–108. [DOI] [PubMed] [Google Scholar]
- Lien, H.C. , Lin, C.W. , Mao, T.L. , Kuo, S.H. , Hsiao, C.H. , Huang, C.S. , 2004. p53 overexpression and mutation in metaplastic carcinoma of the breast: genetic evidence for a monoclonal origin of both the carcinomatous and the heterogeneous sarcomatous components. J. Pathol. 204, 131–139. [DOI] [PubMed] [Google Scholar]
- Lien, H.C. , Hsiao, Y.H. , Lin, Y.S. , Yao, Y.T. , Juan, H.F. , Kuo, W.H. , Hung, M.C. , Chang, K.J. , Hsieh, F.J. , 2007. Molecular signatures of metaplastic carcinoma of the breast by large-scale transcriptional profiling: identification of genes potentially related to epithelial–mesenchymal transition. Oncogene. 26, 7859–7871. [DOI] [PubMed] [Google Scholar]
- Lim, E. , Vaillant, F. , Wu, D. , Forrest, N.C. , Pal, B. , Hart, A.H. , Asselin-Labat, M.L. , Gyorki, D.E. , Ward, T. , Partanen, A. , Feleppa, F. , Huschtscha, L.I. , Thorne, H.J. , Fox, S.B. , Yan, M. , French, J.D. , Brown, M.A. , Smyth, G.K. , Visvader, J.E. , Lindeman, G.J. , 2009. Aberrant luminal progenitors as the candidate target population for basal tumor development in BRCA1 mutation carriers. Nat. Med. 15, 907–913. [DOI] [PubMed] [Google Scholar]
- Liu, X. , Holstege, H. , van der Gulden, H. , Treur-Mulder, M. , Zevenhoven, J. , Velds, A. , Kerkhoven, R.M. , van Vliet, M.H. , Wessels, L.F. , Peterse, J.L. , Berns, A. , Jonkers, J. , 2007. Somatic loss of BRCA1 and p53 in mice induces mammary tumors with features of human BRCA1-mutated basal-like breast cancer. Proc. Natl. Acad. Sci. U.S.A. 104, 12111–12116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livasy, C.A. , Karaca, G. , Nanda, R. , Tretiakova, M.S. , Olopade, O.I. , Moore, D.T. , Perou, C.M. , 2006. Phenotypic evaluation of the basal-like subtype of invasive breast carcinoma. Mod. Pathol. 19, 264–271. [DOI] [PubMed] [Google Scholar]
- Louwman, M.W. , Vriezen, M. , van Beek, M.W. , Nolthenius-Puylaert, M.C. , van der Sangen, M.J. , Roumen, R.M. , Kiemeney, L.A. , Coebergh, J.W. , 2007. Uncommon breast tumors in perspective: incidence, treatment and survival in the Netherlands. Int. J. Cancer. 121, 127–135. [DOI] [PubMed] [Google Scholar]
- Makretsov, N. , He, M. , Hayes, M. , Chia, S. , Horsman, D.E. , Sorensen, P.H. , Huntsman, D.G. , 2004. A fluorescence in situ hybridization study of ETV6–NTRK3 fusion gene in secretory breast carcinoma. Genes Chromosomes Cancer. 40, 152–157. [DOI] [PubMed] [Google Scholar]
- Manie, E. , Vincent-Salomon, A. , Lehmann-Che, J. , Pierron, G. , Turpin, E. , Warcoin, M. , Gruel, N. , Lebigot, I. , Sastre-Garau, X. , Lidereau, R. , Remenieras, A. , Feunteun, J. , Delattre, O. , de The, H. , Stoppa-Lyonnet, D. , Stern, M.H. , 2009. High frequency of TP53 mutation in BRCA1 and sporadic basal-like carcinomas but not in BRCA1 luminal breast tumors. Cancer Res. 69, 663–671. [DOI] [PubMed] [Google Scholar]
- Marchio, C. , Iravani, M. , Natrajan, R. , Lambros, M.B. , Savage, K. , Tamber, N. , Fenwick, K. , Mackay, A. , Senetta, R. , Di Palma, S. , Schmitt, F.C. , Bussolati, G. , Ellis, L.O. , Ashworth, A. , Sapino, A. , Reis-Filho, J.S. , 2008. Genomic and immunophenotypical characterization of pure micropapillary carcinomas of the breast. J. Pathol. 215, 398–410. [DOI] [PubMed] [Google Scholar]
- Marchio, C. , Iravani, M. , Natrajan, R. , Lambros, M.B. , Geyer, F.C. , Savage, K. , Parry, S. , Tamber, N. , Fenwick, K. , Mackay, A. , Schmitt, F.C. , Bussolati, G. , Ellis, I. , Ashworth, A. , Sapino, A. , Reis-Filho, J.S. , 2009. Mixed micropapillary-ductal carcinomas of the breast: a genomic and immunohistochemical analysis of morphologically distinct components. J. Pathol. 218, 301–315. [DOI] [PubMed] [Google Scholar]
- Marusyk, A. , Polyak, K. , 2009. Tumor heterogeneity: causes and consequences. Biochim. Biophys. Acta. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCarthy, A. , Savage, K. , Gabriel, A. , Naceur, C. , Reis-Filho, J.S. , Ashworth, A. , 2007. A mouse model of basal-like breast carcinoma with metaplastic elements. J. Pathol. 211, 389–398. [DOI] [PubMed] [Google Scholar]
- Mendes-Pereira, A.M. , Martin, S.A. , Brough, R. , McCarthy, A. , Taylor, J.R. , Kim, J.S. , Waldman, T. , Lord, C.J. , Ashworth, A. , 2009. Synthetic lethal targeting of PTEN mutant cells with PARP inhibitors. EMBO Mol. Med. 1, 315–322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitelman, F. , Johansson, B. , Mertens, F. , 2007. The impact of translocations and gene fusions on cancer causation. Nat. Rev. Cancer. 7, 233–245. [DOI] [PubMed] [Google Scholar]
- Moinfar, F. , 2008. Is ‘basal-like’ carcinoma of the breast a distinct clinicopathological entity? a critical review with cautionary notes. Pathobiology. 75, 119–131. [DOI] [PubMed] [Google Scholar]
- Mook, S. , Schmidt, M.K. , Rutgers, E.J. , van de Velde, A.O. , Visser, O. , Rutgers, S.M. , Armstrong, N. , van't Veer, L.J. , Ravdin, P.M. , 2009. Calibration and discriminatory accuracy of prognosis calculation for breast cancer with the online Adjuvant! program: a hospital-based retrospective cohort study. Lancet Oncol. 10, 1070–1076. [DOI] [PubMed] [Google Scholar]
- Mullan, P.A. , 2006. Inherited Forms of Breast Cancer, including BRCA1. Abstracts from EMBO Molecular Medicine Conference Common Molecular Mechanisms of Mammary Gland Development and Breast Cancer Progression University College; Dublin: p. 30-1 [Google Scholar]
- Nassar, H. , 2004. Carcinomas with micropapillary morphology: clinical significance and current concepts. Adv. Anat. Pathol. 11, 297–303. [DOI] [PubMed] [Google Scholar]
- Natrajan, R. , Lambros, M.B. , Rodriguez-Pinilla, S.M. , Moreno-Bueno, G. , Tan, D.S. , Marchio, C. , Vatcheva, R. , Rayter, S. , Mahler-Araujo, B. , Fulford, L.G. , Hungermann, D. , Mackay, A. , Grigoriadis, A. , Fenwick, K. , Tamber, N. , Hardisson, D. , Tutt, A. , Palacios, J. , Lord, C.J. , Buerger, H. , Ashworth, A. , Reis-Filho, J.S. , 2009. Tiling path genomic profiling of grade 3 invasive ductal breast cancers. Clin. Cancer Res. 15, 2711–2722. [DOI] [PubMed] [Google Scholar]
- Nielsen, T.O. , Hsu, F.D. , Jensen, K. , Cheang, M. , Karaca, G. , Hu, Z. , Hernandez-Boussard, T. , Livasy, C. , Cowan, D. , Dressler, L. , Akslen, L.A. , Ragaz, J. , Gown, A.M. , Gilks, C.B. , van de Rijn, M. , Perou, C.M. , 2004. Immunohistochemical and clinical characterization of the basal-like subtype of invasive breast carcinoma. Clin. Cancer Res. 10, 5367–5374. [DOI] [PubMed] [Google Scholar]
- Page, D.L. , Anderson, T.J. , 1987. Diagnostic Histopathology of the Breast Churchill Livingstone; Edinburgh: [Google Scholar]
- Parker, J.S. , Mullins, M. , Cheang, M.C. , Leung, S. , Voduc, D. , Vickery, T. , Davies, S. , Fauron, C. , He, X. , Hu, Z. , Quackenbush, J.F. , Stijleman, I.J. , Palazzo, J. , Marron, J.S. , Nobel, A.B. , Mardis, E. , Nielsen, T.O. , Ellis, M.J. , Perou, C.M. , Bernard, P.S. , 2009. Supervised risk predictor of breast cancer based on intrinsic subtypes. J. Clin. Oncol. 27, 1160–1167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parry, S. , Savage, K. , Marchio, C. , Reis-Filho, J.S. , 2008. Nestin is expressed in basal-like and triple negative breast cancers. J. Clin. Pathol. 61, 1045–1050. [DOI] [PubMed] [Google Scholar]
- Peppercorn, J. , Perou, C.M. , Carey, L.A. , 2008. Molecular subtypes in breast cancer evaluation and management: divide and conquer. Cancer Invest. 26, 1–10. [DOI] [PubMed] [Google Scholar]
- Pereira, H. , Pinder, S.E. , Sibbering, D.M. , Galea, M.H. , Elston, C.W. , Blamey, R.W. , Robertson, J.F. , Ellis, I.O. , 1995. Pathological prognostic factors in breast cancer. IV: Should you be a typer or a grader? A comparative study of two histological prognostic features in operable breast carcinoma. Histopathology. 27, 219–226. [DOI] [PubMed] [Google Scholar]
- Perou, C.M. , Sorlie, T. , Eisen, M.B. , van de Rijn, M. , Jeffrey, S.S. , Rees, C.A. , Pollack, J.R. , Ross, D.T. , Johnsen, H. , Akslen, L.A. , Fluge, O. , Pergamenschikov, A. , Williams, C. , Zhu, S.X. , Lonning, P.E. , Borresen-Dale, A.L. , Brown, P.O. , Botstein, D. , 2000. Molecular portraits of human breast tumours. Nature. 406, 747–752. [DOI] [PubMed] [Google Scholar]
- Persson, M. , Andren, Y. , Mark, J. , Horlings, H.M. , Persson, F. , Stenman, G. , 2009. Recurrent fusion of MYB and NFIB transcription factor genes in carcinomas of the breast and head and neck. Proc. Natl. Acad. Sci. U.S.A. 106, 18740–18744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podo, F. , Buydens, L.M.C. , Degani, H. , Hilhorst, R. , Klipp, E. , Gribbestad, I.S. , van Huffel, S. , van Laarhoven, H.W.M. , Luts, J. , Monleon, D. , Postma, G.J. , Schneiderhan-Marra, N. , Santoro, F. , Wouters, H. , Russnes, H.G. , Sørlie, T. , Tagliabue, E. , Børresen-Dale, A.L. , 2010. Triple-negative breast cancer: present challenges and new perspectives. Mol. Oncol. 4, (3) 209–229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polyak, K. , 2007. Breast cancer: origins and evolution. J. Clin. Invest. 117, 3155–3163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Potti, A. , Dressman, H.K. , Bild, A. , Riedel, R.F. , Chan, G. , Sayer, R. , Cragun, J. , Cottrill, H. , Kelley, M.J. , Petersen, R. , Harpole, D. , Marks, J. , Berchuck, A. , Ginsburg, G.S. , Febbo, P. , Lancaster, J. , Nevins, J.R. , 2006. Genomic signatures to guide the use of chemotherapeutics. Nat. Med. 12, 1294–1300. [DOI] [PubMed] [Google Scholar]
- Prat, A. , Perou, C.M. , 2009. Mammary development meets cancer genomics. Nat. Med. 15, 842–844. [DOI] [PubMed] [Google Scholar]
- Pusztai, L. , Mazouni, C. , Anderson, K. , Wu, Y. , Symmans, W.F. , 2006. Molecular classification of breast cancer: limitations and potential. Oncologist. 11, 868–877. [DOI] [PubMed] [Google Scholar]
- Rakha, E.A. , Putti, T.C. , Abd El-Rehim, D.M. , Paish, C. , Green, A.R. , Powe, D.G. , Lee, A.H. , Robertson, J.F. , Ellis, I.O. , 2006. Morphological and immunophenotypic analysis of breast carcinomas with basal and myoepithelial differentiation. J. Pathol. 208, 495–506. [DOI] [PubMed] [Google Scholar]
- Rakha, E.A. , Reis-Filho, J.S. , Ellis, I.O. , 2008. Basal-like breast cancer: a critical review. J. Clin. Oncol. 26, 2568–2581. [DOI] [PubMed] [Google Scholar]
- Rakha, E.A. , El-Sayed, M.E. , Lee, A.H. , Elston, C.W. , Grainge, M.J. , Hodi, Z. , Blamey, R.W. , Ellis, I.O. , 2008. Prognostic significance of Nottingham histologic grade in invasive breast carcinoma. J. Clin. Oncol. 26, 3153–3158. [DOI] [PubMed] [Google Scholar]
- Rakha, E.A. , El-Sayed, M.E. , Menon, S. , Green, A.R. , Lee, A.H. , Ellis, I.O. , 2008. Histologic grading is an independent prognostic factor in invasive lobular carcinoma of the breast. Breast Cancer Res. Treat. 111, 121–127. [DOI] [PubMed] [Google Scholar]
- Rakha, E.A. , Lee, A.H. , Evans, A.J. , Menon, S. , Assad, N.Y. , Hodi, Z. , Macmillan, D. , Blamey, R.W. , Ellis, I.O. , 2010. Tubular carcinoma of the breast: further evidence to support its excellent prognosis. J. Clin. Oncol. 28, 99–104. [DOI] [PubMed] [Google Scholar]
- Reis-Filho, J.S. , 2009. Next-generation sequencing. Breast Cancer Res. 11, (Suppl. 3) S12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reis-Filho, J.S. , Pinheiro, C. , Lambros, M.B. , Milanezi, F. , Carvalho, S. , Savage, K. , Simpson, P.T. , Jones, C. , Swift, S. , Mackay, A. , Reis, R.M. , Hornick, J.L. , Pereira, E.M. , Baltazar, F. , Fletcher, C.D. , Ashworth, A. , Lakhani, S.R. , Schmitt, F.C. , 2006. EGFR amplification and lack of activating mutations in metaplastic breast carcinomas. J. Pathol. 209, 445–453. [DOI] [PubMed] [Google Scholar]
- Reis-Filho, J.S. , Milanezi, F. , Steele, D. , Savage, K. , Simpson, P.T. , Nesland, J.M. , Pereira, E.M. , Lakhani, S.R. , Schmitt, F.C. , 2006. Metaplastic breast carcinomas are basal-like tumours. Histopathology. 49, 10–21. [DOI] [PubMed] [Google Scholar]
- Reis-Filho, J.S. , Simpson, P.T. , Turner, N.C. , Lambros, M.B. , Jones, C. , Mackay, A. , Grigoriadis, A. , Sarrio, D. , Savage, K. , Dexter, T. , Iravani, M. , Fenwick, K. , Weber, B. , Hardisson, D. , Schmitt, F.C. , Palacios, J. , Lakhani, S.R. , Ashworth, A. , 2006. FGFR1 emerges as a potential therapeutic target for lobular breast carcinomas. Clin. Cancer Res. 12, 6652–6662. [DOI] [PubMed] [Google Scholar]
- Reis-Filho, J.S. , Natrajan, R. , Vatcheva, R. , Lambros, M.B. , Marchio, C. , Mahler-Araujo, B. , Paish, C. , Hodi, Z. , Eusebi, V. , Ellis, I.O. , 2008. Is acinic cell carcinoma a variant of secretory carcinoma? A FISH study using ETV6‘split apart’ probes. Histopathology. 52, 840–846. [DOI] [PubMed] [Google Scholar]
- Reis-Filho, J.S. , Lakhani, S.R. , 2008. Breast cancer special types: why bother?. J. Pathol. 216, 394–398. [DOI] [PubMed] [Google Scholar]
- Reis-Filho, J.S. , Tutt, A.N. , 2008. Triple negative tumours: a critical review. Histopathology. 52, 108–118. [DOI] [PubMed] [Google Scholar]
- van de Rijn, M. , Perou, C.M. , Tibshirani, R. , Haas, P. , Kallioniemi, O. , Kononen, J. , Torhorst, J. , Sauter, G. , Zuber, M. , Kochli, O.R. , Mross, F. , Dieterich, H. , Seitz, R. , Ross, D. , Botstein, D. , Brown, P. , 2002. Expression of cytokeratins 17 and 5 identifies a group of breast carcinomas with poor clinical outcome. Am. J. Pathol. 161, 1991–1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosen, P.P. , 2001. Rosen's Breast Pathology second ed. Lippincott Williams & Wilkins; Philadelphia: [Google Scholar]
- Rouzier, R. , Perou, C.M. , Symmans, W.F. , Ibrahim, N. , Cristofanilli, M. , Anderson, K. , Hess, K.R. , Stec, J. , Ayers, M. , Wagner, P. , Morandi, P. , Fan, C. , Rabiul, I. , Ross, J.S. , Hortobagyi, G.N. , Pusztai, L. , 2005. Breast cancer molecular subtypes respond differently to preoperative chemotherapy. Clin. Cancer Res. 11, 5678–5685. [DOI] [PubMed] [Google Scholar]
- Sarrio, D. , Moreno-Bueno, G. , Hardisson, D. , Sanchez-Estevez, C. , Guo, M. , Herman, J.G. , Gamallo, C. , Esteller, M. , Palacios, J. , 2003. Epigenetic and genetic alterations of APC and CDH1 genes in lobular breast cancer: relationships with abnormal E-cadherin and catenin expression and microsatellite instability. Int. J. Cancer. 106, 208–215. [DOI] [PubMed] [Google Scholar]
- Savage, K. , Lambros, M.B. , Robertson, D. , Jones, R.L. , Jones, C. , Mackay, A. , James, M. , Hornick, J.L. , Pereira, E.M. , Milanezi, F. , Fletcher, C.D. , Schmitt, F.C. , Ashworth, A. , Reis-Filho, J.S. , 2007. Caveolin 1 is overexpressed and amplified in a subset of basal-like and metaplastic breast carcinomas: a morphologic, ultrastructural, immunohistochemical, and in situ hybridization analysis. Clin. Cancer Res. 13, 90–101. [DOI] [PubMed] [Google Scholar]
- Savage, K. , Leung, S. , Todd, S.K. , Brown, L.A. , Jones, R.L. , Robertson, D. , James, M. , Parry, S. , Rodrigues Pinilla, S.M. , Huntsman, D. , Reis-Filho, J.S. , 2008. Distribution and significance of caveolin 2 expression in normal breast and invasive breast cancer: an immunofluorescence and immunohistochemical analysis. Breast Cancer Res. Treat. 110, 245–256. [DOI] [PubMed] [Google Scholar]
- Scarff, R.W. , Torloni, H. , i.c.w.t.p.i.t. countries1968. Histological Typing of Breast Tumours World Health Organisation; Geneva: [Google Scholar]
- Shah, S.P. , Kobel, M. , Senz, J. , Morin, R.D. , Clarke, B.A. , Wiegand, K.C. , Leung, G. , Zayed, A. , Mehl, E. , Kalloger, S.E. , Sun, M. , Giuliany, R. , Yorida, E. , Jones, S. , Varhol, R. , Swenerton, K.D. , Miller, D. , Clement, P.B. , Crane, C. , Madore, J. , Provencher, D. , Leung, P. , DeFazio, A. , Khattra, J. , Turashvili, G. , Zhao, Y. , Zeng, T. , Glover, J.N. , Vanderhyden, B. , Zhao, C. , Parkinson, C.A. , Jimenez-Linan, M. , Bowtell, D.D. , Mes-Masson, A.M. , Brenton, J.D. , Aparicio, S.A. , Boyd, N. , Hirst, M. , Gilks, C.B. , Marra, M. , Huntsman, D.G. , 2009. Mutation of FOXL2 in granulosa-cell tumors of the ovary. N. Engl. J. Med. 360, 2719–2729. [DOI] [PubMed] [Google Scholar]
- Shah, S.P. , Morin, R.D. , Khattra, J. , Prentice, L. , Pugh, T. , Burleigh, A. , Delaney, A. , Gelmon, K. , Guliany, R. , Senz, J. , Steidl, C. , Holt, R.A. , Jones, S. , Sun, M. , Leung, G. , Moore, R. , Severson, T. , Taylor, G.A. , Teschendorff, A.E. , Tse, K. , Turashvili, G. , Varhol, R. , Warren, R.L. , Watson, P. , Zhao, Y. , Caldas, C. , Huntsman, D. , Hirst, M. , Marra, M.A. , Aparicio, S. , 2009. Mutational evolution in a lobular breast tumour profiled at single nucleotide resolution. Nature. 461, 809–813. [DOI] [PubMed] [Google Scholar]
- Silver, D.P. , Richardson, A.L. , Eklund, A.C. , Wang, Z.C. , Szallasi, Z. , Li, Q. , Juul, N. , Leong, C.O. , Calogrias, D. , Buraimoh, A. , Fatima, A. , Gelman, R.S. , Ryan, P.D. , Tung, N.M. , De Nicolo, A. , Ganesan, S. , Miron, A. , Colin, C. , Sgroi, D.C. , Ellisen, L.W. , Winer, E.P. , Garber, J.E. , 2010. Efficacy of neoadjuvant Cisplatin in triple-negative breast cancer. J. Clin. Oncol. 28, 1145–1153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson, P.T. , Reis-Filho, J.S. , Gale, T. , Lakhani, S.R. , 2005. Molecular evolution of breast cancer. J. Pathol. 205, 248–254. [DOI] [PubMed] [Google Scholar]
- Simpson, P.T. , Gale, T. , Reis-Filho, J.S. , Jones, C. , Parry, S. , Sloane, J.P. , Hanby, A. , Pinder, S.E. , Lee, A.H. , Humphreys, S. , Ellis, I.O. , Lakhani, S.R. , 2005. Columnar cell lesions of the breast: the missing link in breast cancer progression? A morphological and molecular analysis. Am. J. Surg. Pathol. 29, 734–746. [DOI] [PubMed] [Google Scholar]
- Sorlie, T. , Perou, C.M. , Tibshirani, R. , Aas, T. , Geisler, S. , Johnsen, H. , Hastie, T. , Eisen, M.B. , van de Rijn, M. , Jeffrey, S.S. , Thorsen, T. , Quist, H. , Matese, J.C. , Brown, P.O. , Botstein, D. , Eystein Lonning, P. , Borresen-Dale, A.L. , 2001. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. U.S.A. 98, 10869–10874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sorlie, T. , Tibshirani, R. , Parker, J. , Hastie, T. , Marron, J.S. , Nobel, A. , Deng, S. , Johnsen, H. , Pesich, R. , Geisler, S. , Demeter, J. , Perou, C.M. , Lonning, P.E. , Brown, P.O. , Borresen-Dale, A.L. , Botstein, D. , 2003. Repeated observation of breast tumor subtypes in independent gene expression data sets. Proc. Natl. Acad. Sci. U.S.A. 100, 8418–8423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sotiriou, C. , Neo, S.Y. , McShane, L.M. , Korn, E.L. , Long, P.M. , Jazaeri, A. , Martiat, P. , Fox, S.B. , Harris, A.L. , Liu, E.T. , 2003. Breast cancer classification and prognosis based on gene expression profiles from a population-based study. Proc. Natl. Acad. Sci. U.S.A. 100, 10393–10398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sotiriou, C. , Wirapati, P. , Loi, S. , Harris, A. , Fox, S. , Smeds, J. , Nordgren, H. , Farmer, P. , Praz, V. , Haibe-Kains, B. , Desmedt, C. , Larsimont, D. , Cardoso, F. , Peterse, H. , Nuyten, D. , Buyse, M. , Van de Vijver, M.J. , Bergh, J. , Piccart, M. , Delorenzi, M. , 2006. Gene expression profiling in breast cancer: understanding the molecular basis of histologic grade to improve prognosis. J. Natl. Cancer Inst. 98, 262–272. [DOI] [PubMed] [Google Scholar]
- Sotiriou, C. , Pusztai, L. , 2009. Gene-expression signatures in breast cancer. N. Engl. J. Med. 360, 790–800. [DOI] [PubMed] [Google Scholar]
- Stephens, P.J. , McBride, D.J. , Lin, M.L. , Varela, I. , Pleasance, E.D. , Simpson, J.T. , Stebbings, L.A. , Leroy, C. , Edkins, S. , Mudie, L.J. , Greenman, C.D. , Jia, M. , Latimer, C. , Teague, J.W. , Lau, K.W. , Burton, J. , Quail, M.A. , Swerdlow, H. , Churcher, C. , Natrajan, R. , Sieuwerts, A.M. , Martens, J.W. , Silver, D.P. , Langerod, A. , Russnes, H.E. , Foekens, J.A. , Reis-Filho, J.S. , van't Veer, L. , Richardson, A.L. , Borresen-Dale, A.L. , Campbell, P.J. , Futreal, P.A. , Stratton, M.R. , 2009. Complex landscapes of somatic rearrangement in human breast cancer genomes. Nature. 462, 1005–1010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stingl, J. , 2009. Detection and analysis of mammary gland stem cells. J. Pathol. 217, 229–241. [DOI] [PubMed] [Google Scholar]
- Stingl, J. , Caldas, C. , 2007. Molecular heterogeneity of breast carcinomas and the cancer stem cell hypothesis. Nat. Rev. Cancer. 7, 791–799. [DOI] [PubMed] [Google Scholar]
- Tavassoli, F.A. , 1999. Pathology of the Breast second ed. Appleton & Lange; Stamford: [Google Scholar]
- Thor, A.D. , Eng, C. , Devries, S. , Paterakos, M. , Watkin, W.G. , Edgerton, S. , Moore, D.H. , Etzell, J. , Waldman, F.M. , 2002. Invasive micropapillary carcinoma of the breast is associated with chromosome 8 abnormalities detected by comparative genomic hybridization. Hum. Pathol. 33, 628–631. [DOI] [PubMed] [Google Scholar]
- Tognon, C. , Knezevich, S.R. , Huntsman, D. , Roskelley, C.D. , Melnyk, N. , Mathers, J.A. , Becker, L. , Carneiro, F. , MacPherson, N. , Horsman, D. , Poremba, C. , Sorensen, P.H. , 2002. Expression of the ETV6–NTRK3 gene fusion as a primary event in human secretory breast carcinoma. Cancer Cell. 2, 367–376. [DOI] [PubMed] [Google Scholar]
- Torres, L. , Ribeiro, F.R. , Pandis, N. , Andersen, J.A. , Heim, S. , Teixeira, M.R. , 2007. Intratumor genomic heterogeneity in breast cancer with clonal divergence between primary carcinomas and lymph node metastases. Breast Cancer Res. Treat. 102, 143–155. [DOI] [PubMed] [Google Scholar]
- Tsuda, H. , Takarabe, T. , Fukutomi, T. , Hirohashi, S. , 1999. der(16)t(1;16)/der(1;16) in breast cancer detected by fluorescence in situ hybridization is an indicator of better patient prognosis. Genes Chromosomes Cancer. 24, 72–77. [DOI] [PubMed] [Google Scholar]
- Turner, N.C. , Reis-Filho, J.S. , Russell, A.M. , Springall, R.J. , Ryder, K. , Steele, D. , Savage, K. , Gillett, C.E. , Schmitt, F.C. , Ashworth, A. , Tutt, A.N. , 2007. BRCA1 dysfunction in sporadic basal-like breast cancer. Oncogene. 26, 2126–2132. [DOI] [PubMed] [Google Scholar]
- Turner, N. , Pearson, A. , Sharpe, R. , Lambros, M. , Geyer, F. , Lopez-Garcia, M.A. , Natrajan, R. , Marchio, C. , Iorns, E. , Mackay, A. , Gillett, C. , Grigoriadis, A. , Tutt, A. , Reis-Filho, J.S. , Ashworth, A. , 2010. FGFR1 amplification drives endocrine therapy resistance and is a therapeutic target in breast cancer. Cancer Res. 70, 2085–2094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner, N.C. , Reis-Filho, J.S. , 2006. Basal-like breast cancer and the BRCA1 phenotype. Oncogene. 25, 5846–5853. [DOI] [PubMed] [Google Scholar]
- Vargo-Gogola, T. , Rosen, J.M. , 2007. Modelling breast cancer: one size does not fit all. Nat. Rev. Cancer. 7, 659–672. [DOI] [PubMed] [Google Scholar]
- van't Veer, L.J. , Dai, H. , van de Vijver, M.J. , He, Y.D. , Hart, A.A. , Mao, M. , Peterse, H.L. , van der Kooy, K. , Marton, M.J. , Witteveen, A.T. , Schreiber, G.J. , Kerkhoven, R.M. , Roberts, C. , Linsley, P.S. , Bernards, R. , Friend, S.H. , 2002. Gene expression profiling predicts clinical outcome of breast cancer. Nature. 415, 530–536. [DOI] [PubMed] [Google Scholar]
- van de Vijver, M.J. , He, Y.D. , van't Veer, L.J. , Dai, H. , Hart, A.A. , Voskuil, D.W. , Schreiber, G.J. , Peterse, J.L. , Roberts, C. , Marton, M.J. , Parrish, M. , Atsma, D. , Witteveen, A. , Glas, A. , Delahaye, L. , van der Velde, T. , Bartelink, H. , Rodenhuis, S. , Rutgers, E.T. , Friend, S.H. , Bernards, R. , 2002. A gene-expression signature as a predictor of survival in breast cancer. N. Engl. J. Med. 347, 1999–2009. [DOI] [PubMed] [Google Scholar]
- Vincent-Salomon, A. , Gruel, N. , Lucchesi, C. , MacGrogan, G. , Dendale, R. , Sigal-Zafrani, B. , Longy, M. , Raynal, V. , Pierron, G. , de Mascarel, I. , Taris, C. , Stoppa-Lyonnet, D. , Pierga, J.Y. , Salmon, R. , Sastre-Garau, X. , Fourquet, A. , Delattre, O. , de Cremoux, P. , Aurias, A. , 2007. Identification of typical medullary breast carcinoma as a genomic sub-group of basal-like carcinomas, a heterogeneous new molecular entity. Breast Cancer Res. 9, R24 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vos, C.B. , Cleton-Jansen, A.M. , Berx, G. , de Leeuw, W.J. , ter Haar, N.T. , van Roy, F. , Cornelisse, C.J. , Peterse, J.L. , van de Vijver, M.J. , 1997. E-cadherin inactivation in lobular carcinoma in situ of the breast: an early event in tumorigenesis. Br. J. Cancer. 76, 1131–1133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, Y. , Klijn, J.G. , Zhang, Y. , Sieuwerts, A.M. , Look, M.P. , Yang, F. , Talantov, D. , Timmermans, M. , Meijer-van Gelder, M.E. , Yu, J. , Jatkoe, T. , Berns, E.M. , Atkins, D. , Foekens, J.A. , 2005. Gene-expression profiles to predict distant metastasis of lymph-node-negative primary breast cancer. Lancet. 365, 671–679. [DOI] [PubMed] [Google Scholar]
- Weigelt, B. , Horlings, H.M. , Kreike, B. , Hayes, M.M. , Hauptmann, M. , Wessels, L.F. , de Jong, D. , Van de Vijver, M.J. , Van't Veer, L.J. , Peterse, J.L. , 2008. Refinement of breast cancer classification by molecular characterization of histological special types. J. Pathol. 216, 141–150. [DOI] [PubMed] [Google Scholar]
- Weigelt, B. , Kreike, B. , Reis-Filho, J.S. , 2009. Metaplastic breast carcinomas are basal-like breast cancers: a genomic profiling analysis. Breast Cancer Res. Treat. 117, 273–280. [DOI] [PubMed] [Google Scholar]
- Weigelt, B. , Geyer, F.C. , Horlings, H.M. , Kreike, B. , Halfwerk, H. , Reis-Filho, J.S. , 2009. Mucinous and neuroendocrine breast carcinomas are transcriptionally distinct from invasive ductal carcinomas of no special type. Mod. Pathol. 22, 1401–1414. [DOI] [PubMed] [Google Scholar]
- Weigelt, B. , Mackay, A. , A'Hern, R. , Natrajan, R. , Tan, D.S. , Dowsett, M. , Ashworth, A. , Reis-Filho, J.S. , 2010. Breast cancer molecular profiling: a retrospective analysis of molecular subtype assignment using single sample predictors. Lancet Oncol. 11, 339–349. [DOI] [PubMed] [Google Scholar]
- Weigelt, B. , Geyer, F.C. , Natrajan, R. , Lopez-Garcia, M.A. , Ahmad, A.S. , Savage, K. , Kreike, B. , Reis-Filho, J.S. , 2010. The molecular underpinning of lobular histological growth pattern: a genome-wide transcriptomic analysis of invasive lobular carcinomas and grade- and molecular subtype-matched invasive ductal carcinomas of no special type. J. Pathol. 220, 45–57. [DOI] [PubMed] [Google Scholar]
- Weigelt, B. , Baehner, F.L. , Reis-Filho, J.S. , 2010. The contribution of gene expression profiling to breast cancer classification, prognostication and prediction: a retrospective of the last decade. J. Pathol. 220, 263–280. [DOI] [PubMed] [Google Scholar]
- Weigelt, B. , Reis-Filho, J.S. , 2009. Histological and molecular types of breast cancer: is there a unifying taxonomy?. Nat. Rev. Clin. Oncol. 6, 718–730. [DOI] [PubMed] [Google Scholar]
- Wellings, S.R. , Jensen, H.M. , Marcum, R.G. , 1975. An atlas of subgross pathology of the human breast with special reference to possible precancerous lesions. J. Natl. Cancer Inst. 55, 231–273. [PubMed] [Google Scholar]
- Wellings, S.R. , Jensen, H.M. , 1973. On the origin and progression of ductal carcinoma in the human breast. J. Natl. Cancer Inst. 50, 1111–1118. [DOI] [PubMed] [Google Scholar]
- Westbury, C.B. , Reis-Filho, J.S. , Dexter, T. , Mahler-Araujo, B. , Fenwick, K. , Iravani, M. , Grigoriadis, A. , Parry, S. , Robertson, D. , Mackay, A. , Ashworth, A. , Yarnold, J.R. , Isacke, C.M. , 2009. Genome-wide transcriptomic profiling of microdissected human breast tissue reveals differential expression of KIT (c-Kit, CD117) and oestrogen receptor-alpha (ERalpha) in response to therapeutic radiation. J. Pathol. 219, 131–140. [DOI] [PubMed] [Google Scholar]
- Wirapati, P. , Sotiriou, C. , Kunkel, S. , Farmer, P. , Pradervand, S. , Haibe-Kains, B. , Desmedt, C. , Ignatiadis, M. , Sengstag, T. , Schutz, F. , Goldstein, D.R. , Piccart, M. , Delorenzi, M. , 2008. Meta-analysis of gene expression profiles in breast cancer: toward a unified understanding of breast cancer subtyping and prognosis signatures. Breast Cancer Res. 10, R65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yerushalmi, R. , Hayes, M.M. , Gelmon, K.A. , 2009. Breast carcinoma – rare types: review of the literature. Ann. Oncol. 20, 1763–1770. [DOI] [PubMed] [Google Scholar]
- Zhang, Z. , Yuan, X.M. , Li, L.H. , Xie, F.P. , 2001. Transdifferentiation of neoplastic cells. Med. Hypotheses. 57, 655–666. [DOI] [PubMed] [Google Scholar]
- Zhao, H. , Langerod, A. , Ji, Y. , Nowels, K.W. , Nesland, J.M. , Tibshirani, R. , Bukholm, I.K. , Karesen, R. , Botstein, D. , Borresen-Dale, A.L. , Jeffrey, S.S. , 2004. Different gene expression patterns in invasive lobular and ductal carcinomas of the breast. Mol. Biol. Cell. 15, 2523–2536. [DOI] [PMC free article] [PubMed] [Google Scholar]