

Abstract
Treatment of patients with glioblastoma multiforme (GBM) remains to be a challenge with a median survival of 14.6 months following diagnosis. Standard treatment options include surgery, radiation therapy, and systemic chemotherapy with temozolomide. Despite the fact that the brain constitutes an immunoprivileged site, recent observations after immunotherapies with lysate from autologous tumor cells pulsed on dendritic cells (DCs), peptides, protein, messenger RNA, and cytokines suggest an immunological and even clinical response from immunotherapies. Given this plethora of immunomodulatory therapies, this paper gives a structure overview of the state-of-the art in the field. Particular emphasis was also put on immunogenic antigens as potential targets for a more specific stimulation of the immune system against GBM.
1. Introduction
The primary brain tumor, glioblastoma multiforme (GBM), occurs in 3 to 4 adult patients per 100,000 inhabitants in Europe, thus being the most common and life threatening primary brain tumor [1]. GBM is invasive and infiltrates the surrounding brain tissue.
GBM is most common in adults older than 50 years and affects more men than women. Furthermore, around 9% of childhood brain tumors are GBMs. The median survival from the time of diagnosis without any treatment is 3 months. The major prognostic factors are age and Karnofsky performance status (KPS) at the time of diagnosis [2].
2. Standard Treatment of Glioblastoma (GBM)
2.1. Primary Treatment
Although GBM has a typical appearance in MRI, histological diagnosis is mandatory for proper diagnosis. A treatment option is gross tumor resection (GTR), which involves the contrast enhancing tumor in to the MRI without causing additional neurological deficit [3]. The extent of resection can be optimized using fluorescence-guided resection [4], which probably includes noncontrast enhancing tumor part and can be visualized by aminoacid positron emission tomography [5]. Placing carmustine-loaded wafers in the resection cavity has shown to prolong survival rates [6]. However, the clinical benefit seems to be limited to patients with GTR and good KPS [7]. If GTR cannot be achieved and the tumor mass does not cause a midline shift, stereotactic serial biopsy is a safe procedure to enable histological and genetic diagnosis [8–10].
Following surgery, a typical treatment consists of concomitant temozolomide and 60 Gy radiotherapy of the tumor region for six weeks followed by 6 adjuvant cycles of temozolomide [11, 12]. After applying this treatment, tumors with methylated promotor for O(6)-methyl-guanine DNA methyltransferase (MGMT) appear to have a favorable clinical course with a median overall survival of 21.7 months [13].
Although long-term adjuvant temozolomide is safe [14], there is no evidence that the continuation of temozolomide beyond six cycles gives any additional benefit.
2.2. Recurrent GBM
In recurrent GBM, surgery is an optional treatment. However, there is no evidence for the clinical benefit of second surgery. According to retrospective studies, a second resection should be restricted to patients with good KPS and feasible systemic salvage treatment [15, 16]. Alternatively, conformal reirradiation may be administrable [17, 18].
For systemic treatment of recurrent glioblastoma, bevacizumab is FDA approved but only with class 2 evidence [19–22]. In Europe, bevacizumab remains off-label treatment for glioblastoma patients. Rechallenge with TMZ is an alternative to bevacizumab. Thereby, protocols vary from dose-intensified treatment to metronomic schedules [23–26]. However, there are no positive randomized controlled trials defining a standard treatment in recurrent glioblastoma. However, salvage treatment should be applied as long as the patient's condition has not declined [27].
Due to the limited treatment options for GBM patients, there is a fervent need for novel therapies such as immunotherapies. However, the brain is known as one of the immunologically privileged sites and is able to tolerate the introduction of antigen without eliciting an inflammatory immune response [28]. Thus, immunotherapy for brain tumor seems to constitute a “mission impossible.” Fortunately, it is now known that the central nervous system at least maintains a reciprocal communication network with the immune system. Infectious or experimental autoimmune encephalomyelitis animal models allow us to understand better how the immune system operates in the brain [29]. Therefore, immunotherapy offers the opportunity to allow the patient's immune system a chance to eliminate the tumor. The strength of immunotherapy with DCs has been demonstrated on the Food and Drug Administration (FDA) approval of DCs as “Provenge” for prostate cancer [30]. As for GBM, it has been demonstrated that it efficiently treats relatively small tumors in experimental animal models.
This paper focuses on the cellular-based immunotherapy for brain cancers with emphasis on GBM. We will also highlight some of the possible directions that may be taken in the immediate future to improve this therapeutic option.
3. Immunotherapy
There are two important basic strategies for immunotherapy. Firstly, adoptive immunotherapy, which means the passive administration of sensitized immune cells to patients. Secondly, the strategy of “active immunotherapy” is based on the boosting of antitumor T-cell responses by antigen-presenting cells (APCs), especially by dendritic cells (DCs).
3.1. Adoptive Immunotherapy
In adoptive immunotherapy, in vitro activated immune cells are administered to tumor-bearing patients. Lymphokine-activated killer (LAK) cells, which were generally obtained by cultivating peripheral lymphocytes in the presence of T-cell growth factor interleukin-2 (IL-2) and other cytokines. These LAKs showed cytolytic properties but not specifically against tumor cells [31–42]. A novel therapeutic option was to collect lymphocytes from lymph nodes or peripheral blood mononuclear cells (PBMCs) after peripheral injection of irradiated autologous tumor cells and granulocyte-macrophage colony-stimulating factor (GM-CSF), stimulating them in vitro, and subsequently reinjecting them [43–47]. Allogeneic cytotoxic T lymphocytes (CTLs) stimulated by the patient's autologous lymphocytes have been tested for recurrent GBM as well [48]. They were generated by ex vivo antigenic stimulation of PBMCs. As Quattrocchi et al. have shown in a pilot study, CTLs can also be amplificated from tumor-infiltrating lymphocytes (TILs) in the presence of IL-2 [49].
Injection of CTLs or TILs appeared to allow higher objective responses compared to LAKs in some GBM patients. Due to the large variability observed between patients and the limited number of patients, the correlation between the number of effector cells, their cytotoxic activity, and the clinical outcome is still not clear. Kronik et al. have predicted that GBM would be eradicated by new dose-intensive strategies, for instance, 3 × 108 CTLs every 4 days for small tumor burden or 2 × 109 CTLs infused every 5 days for larger tumor burden [50]. Interestingly, in several clinical trials with LAK and CTL therapy, the eosinophilic infiltration at the tumor site and in cerebrospinal fluid (CSF) could be observed [38, 40, 48, 49]. The impact of chemotherapy or corticosteroids on the efficacy of the treatment might also be questioned. These drugs were completely avoided in some trials according to their immunosuppressive properties [31, 39, 41, 42]; however, other studies have shown no influence of steroids or chemotherapy on the stimulation and the cytotoxic activity of the effector cells [32, 34, 35].
Progress in the treatment of brain tumors using immunotherapy is slowly moving forward. Initial attempts used nonspecific approaches, like adjuvants and, LAK cells were only minimally effective. Nowadays, the general focus is directed towards specific cellular approaches including TILs and CTLs, and alloreactive CTL stimulated by mixed lymphocyte reactions. All of these approaches have yielded some clinical success. GBM cells seem to have a plethora of tumor-associated antigens. Active immunization with autologous DCs that have been loaded with tumorantigens also appear to generate long-term survivors. Identification of other strategies that can be combined with immunotherapy approaches might improve our success against GBM.
3.2. Active Immunotherapy
3.2.1. Active Immunotherapy in GBM Using Autologous Tumor Cells (ATCs)
Autologous tumor cells are removed from patients during surgery. Whole cells, parts of cells, or antigens can be used to create a vaccine to a specific tumor. To date, there are at least eight high qualified trials, which focused on the GBM treatment were reported [51–58]. One phase I clinical trial [58], two cases report [51, 54], and five pilot studies of antitumor vaccination [52, 53, 55–57] were included, and the vaccination was repeated in all of these studies. ATCs are generally inactivated by radiation, sometimes genetically modified [51, 54, 56], and could be infected with a virus [52, 55] to enhance the immune response. The strategy of using antisense oligonucleotides for insulin growth factor receptor 1 for ATCs prior to implantation was applied in one pilot study [53]. The cells were injected either subcutaneously or intradermally. In three studies, injections of ATCs were given concomitantly with IL-2 [51], IL-4 [54] or B7-2, and GM-CSF infusions [56]. Different amounts of cells were given for vaccination.
At least half of the patients in these studies showed an induction of immune responses both in peripheral blood and tumor site [53, 55]. Toxicity was addressed in all trials without any severe adverse events. Beside an immune response, a clinical response was demonstrated to be associated with survival benefit in five studies with three complete responses (CRs), four partial responses (PRs), two minor responses (MRs), and six stable diseases (SDs) in a total of 48 GBM patients [53–55, 57, 58].
3.2.2. Active Immunotherapy Using Dendritic Cells
Dendritic cells (DCs) are professional antigen-presenting cells (APCs), which play a key role in eliciting, maintaining, and regulating T-cell responses [59, 60]. DCs are present in almost all organs, even in immune-privileged sites such as the central nervous system, testis, and ovaries. DCs can be generated not only from lymphoid organs but also from the blood or lymph. DC vaccines are attractive and now widely used in GBM active immunotherapy based on their various antitumor effects [Figure 1].
Figure 1.
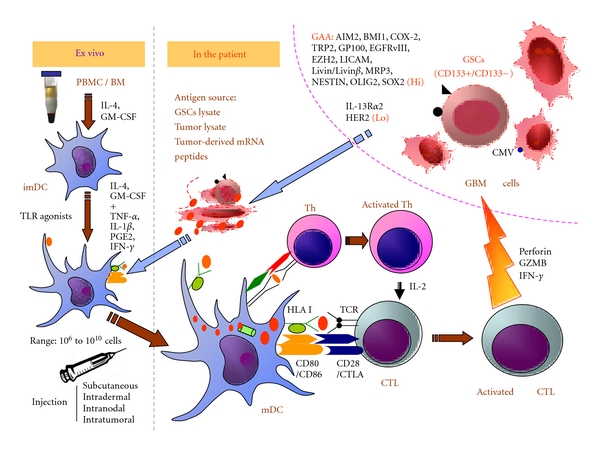
DC-based active immunotherapy for GBM. DCs display a unique capacity to induce and to maintain T-cell responses. Mature DCs are generated from PBMC in vitro in the presence of IL-4, GM-CSF, TNF-alpha, IL-1beta, PGE2, IFN-gamma, and other cytokines, in addition to TLR agonists. Subsequently, they are loaded with GBM or glioblastoma stem cell lysates, GBM-associated antigen-derived peptides, protein, or RNA. Due to their high surface expression of HLA-peptide-complexes and costimulatory molecules, DCs could efficiently activate and expand CD8+ CTLs and CD4+ Th cells. CD8+ CTLs are able to recognize and eliminate tumor cells, especially the GBM stem cells (CD133). CD4+ Th cells enhance the capacity of DCs to induce CTLs by the interaction between CD40 on DCs and CD40 ligand on activated CD4+ T cells. In addition, CD4+ T cells help in the maintenance and expansion of CTLs by secreting IL-2. CTLs: cytotoxic T cells; imDC: immature dendritic cells; GZMB: granzyme B; GSCs: glioblastoma stem cells, HLA: human leukocyte antigen; IL: interleukin; IFN: interferon; mDC: mature dendritic cells; PBMC: peripheral blood mononuclear cells; TCR: T-cell receptor; Th: T helper cell; TLR: toll-like receptor.
In Table 1, we summarized 15 clinical trials reported so far including 316 patients [61–75]: eight phase I trials [62, 64, 65, 69–72, 75], six phase I/II trials [61, 63, 66, 68, 73, 74], and one phase II trial [67]. Monocyte-derived DCs were used for most of the clinical trials. The preparation of DC is now well established, and a sufficient number of DC can be generated for injections into patients [76, 77]. Immature DCs were widely used in older trials [67, 69, 70, 74, 75]; however, some trials have used maturation factors like TNF-alpha [71], toll-like receptor (TLR) agonists: penicillin-killed streptococcus pyogenes (OK-432) [68] and imiquimod [62, 64], TLR ligand: poly ICLC [61, 62], IFN-gamma and TNF-alpha in combination with IL-4-secreting fibroblasts [78]. In several trials, DCs were matured using cocktails with IL-1beta, TNF-alpha, PGE2, or IFN-gamma [7, 63, 64, 74]. The number of DCs injected ranged from 1 × 106 to 1 × 1010. The frequency of the injections was highly variable. One phase I study focused on the dose of DCs, which did not result in any dose-limiting toxicity [69]. The sources of antigen were quite different: autologous tumor lysates, apoptotic glioma cells peptides eluded from ATCs, synthetic peptides, defined peptides, mRNA derived from ATCs, and irradiated single-cell suspension of ATCs. For vaccinations using ATCs, the ATCs were fused [71] or incubated with DC. Defined peptides were derived from EGFRvIII, the particular target as its frequent expression in GBM [65, 79]. Vaccines were injected intradermally, intranodally, or subcutaneously. Moreover, in one phase I/II trial, some patients even received intratumoral injections [74].
Table 1.
Synopsis of DC-based immunotherapy trials for GBM patients.
| Patients | Phase | Route | Antigen format | Immune response | Clinical response | References |
|---|---|---|---|---|---|---|
| 22 patients (13 recurrent GBM, 5 AA, 3 AO, 1 AOA) | Phase I/II | Intranodal + intramuscular injections of poly-ICLC | Synthetic peptides for GAAs | Induced positive immune responses against at least one of the GAAs in PBMCs in 58% of patients (after 4 vaccinations). Significant upregulation: interferon-alpha and CXCL10. | 4 recurrent GBM are progression free for at least 12 months; 1 CR (recurrent GBM). Median TTP: 4 months. | [61] |
| 23 patients (15 newly diagnosed GBM, 8 recurrent GBM) | Phase I | Intradermal + intramuscular injections of poly-ICLC | Autologous tumor lysate + imiquimod or poly-ICLC | No dose-limiting toxicity. Tumor samples with a mesenchymal gene expression signature had a higher number of CD3+ and CD8+ tumor-infiltrating lymphocytes | Newly diagnosed: median OS: 35.9 months, with a mean follow-up time of more than 4 years, and 1-, 2-, and 3-year survival rates of 93%, 77%, and 58%, respectively. Recurrent: median OS was 17.9 months from the time of initial glioblastoma diagnosis. OS was significantly longer for those who received DC vaccination at initial diagnosis compared with those who enrolled in this trial at the time of recurrence. | [62] |
| 8 patients (newly diagnosed GBM) | Phase I/II | Intradermal | Autologous tumor lysate | DTH (2/5) increased CD8+/CD25+ in PBL (6/7) ATR PBMC (5/8) IFN-gamma ELISPOT) | Median OS: 24 months | [63] |
| 45 children (23 recurrent GBM, 5 AA, 1 AOA, 16 other HGG) | Phase I | Intradermal | Autologous tumor lysate + imiquimod | No data available | Median PFS for relapsed GBM: 4.3 months; median OS for relapsed GBM: 12.2 months | [64] |
| 12 patients (newly diagnosed GBM) | Phase I | Intradermal | EGFRvIII antigen + KLH | DTH EGFRvIII (5/9); DTH KLH (9/9); ATR PBMC (10/12) (EGFRvIII-induced proliferation) | Median OS: 22.8 months | [65] |
| 56 patients (recurrent GBM) | Phase I/II | Intradermal | Autologous tumor lysate | DTH (9/21 at time of diagnosis, 9/17 after 2 vaccinations) | 3-month PFS; OS: 9.6 months; 24-month OS: 14.8%; total resection is a predictor for better PFS; younger age and total resection are predictors for better OS. | [66] |
| 34 patients (23 recurrent GBM, 11 newly diagnosed GBM) | Phase II | Subcutaneous | Autologous tumor lysate | Postvaccine antigen-directed IFNg response in PBMCs (17/34); DTH-test resulted in cutaneous GBM in 1 patient (DTH was subsequently discontinued) | Newly diagnosed: 8/17 (47%) vaccine responders versus. 3/15 (20%) nonresponders; Recurrent: TTS, 621 ± 81 and 402 ± 49 d; TTP, 28 ± 94 and 142 ± 22 d (8 responders and 13 nonresponders); TTP, 343 ± 116 and 136 ± 19 d (8 responders and 15 nonresponders) | [67] |
| 24 patients (18 recurrent GBM, 6 grade III glioma) | Phase I/II | Intradermal or intradermal + intratumoral (Ommaya reservoir) | Autologous tumor-lysate | DTH to tumor lysate (8/24); ATR PBMC (7/24) (IFN-gamma ELISPOT) | 1 PR, 3 MR, 6 SD (GBM); 4 SD (Grade III glioma); median OS: 16 months versus 13.3 months; longer survival if DC maturation or IC injection. One grade IV neurotoxicity event (stupor) observed. | [68] |
| 12 patients (7 newly diagnosed GBM, 5 recurrent GBM) | Phase I | Intradermal | Acid-eluted tumor associated peptides | CTL response (6/12); tumor infiltration CD8+ CD45RO+ cells (4/8) | Median TTP: 19.9 months—OS 18 to >58 months—median OS: 35.8 months. 1PR; Median OS: 23.4 versus 18.3 months. No dose-limiting toxicity observed | [69] |
| 14 patients (1 newly diagnosed GBM, 9 recurrent GBM, 4 AA) | Phase I | Subcutaneous | Autologous tumor lysate | Increased IFNγ RNA in PBMC (6/10) ATR T cells (4/9) (HER-2, gp100, MAGE-1 tetramers); CD8+, CD45RO+ cells infiltration (3/6) | Median survival: 33.3 versus 7.5 months (8/9 recurrent GBM). | [70] |
| 15 patients (6 recurrent GBM, 7 AA, 2 OAA) | Phase I | Intradermal | DC fusion with autologous glioma cells | DTH to tumor lysate (15/15); increased cytotoxic activity (2/15); increased intracellular IFN-gamma in CD8+ T cells (1/15) | 1 SD (GBM); 3 PR, 1 MR (AA); 1 PR, 1 SD (AOA) | [71] |
| 7 patients (2 recurrent GBM, 1 AA, 4 other HGG) | Phase I | Intradermal | Autologous tumor RNA | No anti-tumor responses (0/3) (IFN-gamma ELISA) | 1 PR (1XA); 4 SD (1AA, 3 other HGG) | [72] |
| 25 patients (newly diagnosed GBM: 13 plus chemotherapy, 12 without chemotherapy) | Phase I/II | Intradermal | Autologous tumor lysates or peptide elutions | Vaccine alone: ATR PBMC (4/11) Vaccine + chemotherapy: ATR PBMC (4/13) (lytic activity and IFN-gamma PCR) | Vaccine or chemotherapy alone: 24-month survival: 8% Vaccine + chemotherapy: 3 PR; 24-month survival: 42% | [73] |
| 10 patients (7 recurrent GBM after radiotherapy, 3 recurrent grade III glioma) | Phase I/II | Intradermal and/or intratumor (Ommaya) | Autologous tumor lysate | Increase in NK cells in PBMCs (5/10); DTH to tumor lysate (3/10); increased T-cell mediated antitumor activity (2/10) | 2 MR, 2SD (GBM), 2SD (Grade III glioma); OS > 50 months. | [74] |
| 9 patients (7 newly diagnosed GBM, 2 AA after radiotherapy) | Phase I | Subcutaneous | Tumor-specific MHC-I-associated peptides | Systemic CTL cytotoxicity against tumor (4/9) (lytic activity); tumor infiltration: CD4+, CD8+, CD45RO+ cells (2/4) | Prolonged median survival compared to control group: 15.2 versus 8.6 months (GBM) | [75] |
Abbreviations: AA: anaplastic astrocytoma; AO: anaplastic oligodendroglioma; AOA: anaplastic oligoastrocytoma; ATR: anti-tumor responses; CR: complete response; DTH: delayed-type hypersensitivity; GAA: glioblastoma associated antigen; GBM: glioblastoma multiform; HGG: high-grade glioma; KLH: keyhole limpet haemocyanin; MR: Minor response; OS: overall survival; PBMC: peripheral blood mononuclear cells; PFS: progression-free status; PR: partial response; PXA: pleomorphic xanthoastrocytoma; SD: stable disease; TMZ: Temozolomide; TTP: Time to tumor progression; TTS: Time to tumor survival; XA: xanthoastrocytoma.
From all of these clinical studies, only one patient had a large residual tumor and a perilesional edema suffered grade IV neurotoxicity (stupor) [68]. A peripheral immune response such as DTH (delayed-type hypersensitivity) lymphocyte infiltrations, particularly CD8+ cells, can be observed in more than half of patients. Activated CD8+ CTLs efficiently recognize and destroy tumor cells, which expose peptides derived from tumor-associated antigens (TAAs) in human leukocyte antigen (HLA) class I molecules [80]. CD4+ T cells recognizing peptides in the complex of HLA class II molecules also play an important role in antitumor immunity [81]. CD4+ T cells improve the capacity of DCs to induce CTLs by the interaction between CD40 on DCs and CD40 ligand on activated CD4+ T cells. In addition, CD4+ T cells help to maintain and expand CTLs by secreting cytokines such as IL-2. Beside their extraordinary capacity to elicit T-cell responses, DCs efficiently improve the immunomodulatory and cytotoxic potential of natural killer cells, which are also involved in the elimination of tumor [82, 83]. Furthermore, DCs can also directly mediate tumor-directed cytotoxicity [84].
Almost fifteen studies have reported on a survival benefit of patients receiving immunotherapies when compared with historical cohorts [61–70, 73–75]. Liau et al. [69] vaccinated four patients showing an increase of intratumoral infiltration by lymphocytes after vaccination at a time when the tumor was minimal. T-cell infiltration correlated with a decrease in intratumoral TGF-beta and was associated with a better survival. Patients without T-cell infiltration showed a reverse effect. Combined intravenous and intracranial administration of ATCs gave a superior response when compared to intravenous injection only [68]. Wheeler et al. reported on the large cohort of 34 GBM patients demonstrating that responders had an increase of IFN-gamma after vaccination when compared with the IFN-gamma level before vaccination using in vitro PBMC stimulation. Moreover, responders to vaccination showed a better response to chemotherapy which was delivered in a second phase [67]. Recent phase I and phase I/II studies with 10 newly diagnosed GBM and 13 recurrent GBM, vaccinated intranodally with autologous tumor lysate pulsed on DCs after radiation and chemotherapy or synthetic peptides for GAA epitopes showed a good immune response and a prolonged survival [61, 85].
As a consequence, active immunotherapy appears to have a beneficial effect in some patients, particularly in those with a limited tumor, without causing major toxicity. Both clinical trials using ATCs and DC demonstrate induced immune responses (DTH reaction, tumor infiltration, and/or anti-tumor responses of PBMC) and some clinical responses. The important take-home message for DC vaccination is that no dose-related toxicity has been demonstrated [69]. In addition, it seems better to use mature DC compared to immature DC. Due to the large variability of protocols tested, the source of ATCs, and the type and the cell number of DC injected, the type of adjuvants, no proven approach can be presented so far.
Various antigen sources can be used for DC active immunotherapy. Peptides are very popular; however, loading DCs with peptides requires a large culture of autologous tumor cells, which is a complex process. To break this limitation, some trials load DCs with tumor lysate instead of eluted peptides. Yu et al. [75] found T-cell-mediated cytotoxicity in 60% of the patients after immunization with tumor lysate-loaded DCs, a success rate higher than the 40% value seen with eluted peptides by the same team [70]. In another phase I/II trial, 24 patients with recurrent malignant gliomas were treated with intradermal or intratumoral (Ommaya reservoir) injections of DCs loaded with tumor lysate. Some patients also received intratumoral injections. One PR and three MR were observed [68]. A novel development in cancer vaccines consists of fusing tumor cells with DCs, a strategy that has been associated with clinical responses in patients with glioma [71].
3.2.3. Antigens for GBM Immunotherapy
(1) Glioblastoma-Associated Antigens (GAAs) —
Recently, many efforts have been made to identify tumor-associated proteins as targets of tumor-reactive T cells and to define peptide motifs within these proteins constituting T-cell epitopes. In this paper, we focus on glioblastoma-associated antigens (GAAs), which have already been used for DC-based vaccination trials enrolling GBM patients. GAAs such as EGFRvIII, EphA2, GP100, HER2, MAGE-1, IL-13Rα2, SOX11, and TRP2 [86–89], which were frequently overexpressed in GBMs, were able to initiate immune responses. Other antigens associated with GBM have been described including survivin, WT1, SOX2, AIM2, SART1, SART2, and SART3 [90]. T cells directed against IL-13Rα2 and EphA2 have been demonstrated in the PBMCs of a long surviving patient with anaplastic astrocytoma, showing that a spontaneous immune reaction can be observed in high-grade glioma [91]. Many glioblastoma-associated antigens were identified within either glioblastoma cell lines or GBM cells, such as ART and SART [92]. “Cancer-testis antigens” are differentially expressed in testis and tumors including MAGE-1, GAGE-1, and NY-ESO-1. These antigens were found in terminally differentiated melanocytes and also in GBMs [93–95]. TRP-1 and TRP-2 were not found in the testis but were detected in normal cells like melanocytes as well transformed tissues like melanomas and glioblastomas [89]. Since melanoma and glioblastoma cells share a common embryonic neuroectoderm precursor, it is not that surprising that these two cancer types share many common antigens. Here, we summarized the key GAAs in Table 2 [79, 89–123].
Table 2.
List of glioblastoma-associated antigens (GAAs).
| GAAs | Characteristic/potential function | References |
|---|---|---|
| *AIM2: absent in melanoma 2 | AIM-2 could be used as a tumor antigen target for monitoring vaccine trials or for developing antigen-specific active immunotherapy for glioma patients. | [90, 96–98] |
| *BMI1: BMI1 polycomb ring finger oncogene | Expressed in human GBM tumors and highly enriched in CD133+ GSC cells. | [99] |
| *COX-2: cyclooxygenase-2 | Overexpressed in many tumors including CD133+ GSC cells, COX-2 inhibitor celecoxib will become a nice weapon for GBM therapy. | [100] |
| *TRP-2: tyrosinase related protein 2 | Highly expressed in GSCs. | [89, 97, 98] |
| *GP100: glycoprotein 100 | Melanocyte lineage-specific antigen, expressed in GSCs as well. | [89, 98] |
| *EGFRvIII: epidermal growth factor receptor variant III | EGFRvIII is the most prevalent of several EGFR mutations found in human gliomas and is expressed in 20–25% of GBM. GSC-associated antigen. | [79, 89, 98] |
| *EZH2: enhancer of zeste homolog 2 | Upregulated in malignant gliomas and in GSC cells. | [97, 101] |
| *LICAM: human L1 cell adhesion molecule | Highly expressed in GSCs. Invasion-associated proteins. | [102] |
| *Livin and Livinβ | The expression of livin and livinβ in CD133+ U251 stem-like cells was much higher than that in cancer cells, Livinβ was more related with the high survival rate. It is a cancer-associated member of the inhibitor of apoptosis protein (IAP). | [103] |
| *MRP-3: multidrug-resistance protein 3, | GBMs overexpress MRP3 at both mRNA and protein levels. Multidrug-resistance protein 3, potential correlation with survival. Highly expressed in GSC cells as well. | [97, 98, 104] |
| *Nestin | Nestin plays important roles in cell growth, migration, invasion, and adhesion to extracellular matrices in glioma cells. Overexpressed in GSCs. | [105] |
| *OLIG2: oligodendrocyte transcription factor 2 | GSC marker, OLIG2 is highly expressed in all diffuse gliomas. Immunohistochemistry and microarray analyses demonstrated higher OLIG2 in anaplastic oligodendrogliomas versus glioblastomas, which are heterogeneous with respect to OLIG2 levels. | [106] |
| *SOX2: SRY-related HMG-box 2 | SOX2 expression and amplification in gliomas and GSC cell lines. | [98, 107] |
| ART1: antigen recognized by T cells 1 | Pediatric GBM express ART1, ART4, SART1, SART2, and SART3, they were identified within glioblastoma cell lines as well. | [92, 97] |
| ART4: antigen recognized by T cells 4 | ||
| SART1: squamous cell carcinoma antigen recognized by T cells 1 | ||
| SART2: squamous cell carcinoma antigen recognized by T cells 2 | ||
| SART3: squamous cell carcinoma antigen recognized by T cells 3 | ||
| B-cyclin | Overexpressed in GBM. | [97, 108] |
| β-catenin | β-catenin and Gli1 are prognostic markers in GBM. | [109] |
| Gli1: glioma-associated oncogene homolog 1 | Gli1 is correlated with glioma recurrence after chemotherapy, Gli1 plays a dominant role in chemoresistance of glioma cells. located in nuclear, might be fluctuating between the cytoplasm and the nucleus. | [109, 110] |
| Cav-1: caveolin-1 | Expressed in most HGG, correlated with proliferation and invasive potential of tumor. | [111] |
| Cathepsin B | Overexpression of cathepsin B during the progression of human gliomas. | [112] |
| CD74: cluster of Differentiation 74 | Contribute to TMZ resistance. Also known as HLA class II histocompatibility antigen gamma chain. | [113] |
| E-cadherin: epithelial calcium-dependent adhesion | Expression in gliomas correlated with an unfavorable clinic outcome. | [114] |
| EphA2/Eck: EPH receptor A2/epithelial cell kinase | Overexpressed in both pediatric and adult GBM. Used as a novel target for glioma vaccines. | [90, 97, 115] |
| Fra-1/Fosl 1: fos-related antigen 1 | Plays an important role in maintenance/progression of various cancers, including GBM. Highly expressed in pediatric GBM. | [97, 116] |
| GAGE-1: G antigen 1 | A potential target for specific immunotherapy and diagnostic markers in high-grade brain tumors. | [93] |
| Ganglioside/GD2 | Expressed in astrocytic tumors. | [117] |
| GnT-V, β1,6-N: acetylglucosaminyltransferase-V | Plays an important role in regulating invasivity of human glioma. | [97, 118] |
| Her2/neu: human epidermal growth factor receptor 2 | A tumor-associated antigen that is expressed by up to 80% of GBMs but not by normal postnatal neurons or glia. | [97, 98] |
| Ki67: nuclear proliferation-associatedantigen of antibody Ki67 | Prognostic marker for glioma, especially for the lower grades. | [79] |
| Ku70/80: human Ku heterodimer proteins subunits (molecular weight: 70 kDa/80 kDa) | A therapeutic potential target antigen. Highly expressed in GBM. | [119] |
| IL-13Rα2: interleukin-13 receptor subunit alpha-2 | Overexpressed in GBM but diminished in several GSC cell lines. | [89, 97, 98] |
| MAGE-A: melanoma-associated antigen 1 | MAGE-A1, MAGE-A3, and NY-ESO-1 can be upregulated in neuroblastoma cells to facilitate cytotoxic T-lymphocyte-mediated tumor cell killing. | |
| MAGE-A3: melanoma-associated antigen 3 | [94] | |
| NY-ESO-1: New York oesophageal squamous cell carcinoma 1 | ||
| MART-1: melanoma antigen recognized by T-cells | Melanoma antigen also associated with glioma. | [95] |
| PROX1: prospero homeobox protein 1 | Strongly express in GBM, frequently coexpress early neuronal proteins MAP2 and betaIII-tubulin but not the mature neuronal marker NeuN. | [120] |
| PSCA: prostate stem cell antigen | GPI-anchored cell surface protein, represented as a novel GAA. | [121] |
| SOX10: SRY-related HMG-box 10 | The SOX10 expression was restricted to gliomas and melanomas. All glioma types expressed SOX10, and tumors of low-grade glioma had a much broader distribution of SOX10 compared with high-grade gliomas. | [122] |
| SOX11: SRY-related HMG-box 11 | The transcription factor SOX11 highly specific overexpression of in human malignant gliomas. | [87, 97] |
| Survivin | Quantitatively determined survivin expression levels are of prognostic value in human gliomas. | [79, 97] |
| UPAR: urokinase-type plasminogen activator receptor | UPAR and Cathepsin B, known to be overexpressed in high-grade gliomas and strongly correlated with invasive cancer phenotypes. | [123] |
| WT-1: Wilms' tumor protein 1 | A transcription factor overexpressed in glioma. | [91] |
*: Glioblastoma stem cell (GSC) associated antigens as potential targets for immunotherapy.
(2) Viral Antigens —
Viral antigens act as good targets for anti-infectious immunity. Moreover, many viruses such as HTLV-1, hepatitis B and C virus, and EBV/JCV play a critical role in several human cancers as well. Cytomegalovirus (CMV, a common, typically harmless herpes virus) is frequently detected within chronically immunosuppressed patients. It is thought that up to 90% of the population might be chronically infected with CMV. However, our immune system keeps them under tight control. GBM patients are considered to be immunosuppressed through many mechanisms [124]. So the CMV can revive whenever the immune system is impaired. In 2002, Dr. Cobbs et al. [125] linked CMV with human GBM. They analyzed GBM samples from 22 patients and found that all of them harbored CMV. 80% people have this virus, which remains in the body for remaining lifespan. Whether CMV directly causes GBM is still a hot topic and is also controversial. The possibility that CMV attaches itself to GBM via the platelet-derived growth factor alpha allows some interesting therapies to be explored. One CMV antigen, pp65, induced a HLA-A2 restricted immune response in a GBM patient [126]. Freshly isolated GBM samples seem to highly express this CMVpp65 antigen, but cell lines lose this ability [127]. If a high number of GBM cells harbor CMV or express CMV antigen in vivo, this might open the door towards developing CMV peptides to vaccinate against the virus and the tumor at the same time [Figure 2]. Currently, after learning about Dr. Cobbs's work, Dr. Mitchell and his colleagues first confirmed the basic findings. They discovered CMV in the tumors of more than 90% of those patients with GBM, but not in healthy brain tissue nor in nonmalignant brain tumors [127]. Then used DC-based vaccines-targeting CMV antigens to treat 21 patients who had been diagnosed with GBM. Allogeneic CMV-specific CTLs have been used for treating glioma patients by the research group from Pennsylvania State University [128, 129]. Yao et al. [130] and Schmitt et al. [131] showed that the streptamer technology offered the advantage of selecting CMVpp65-specific CD8+ CTLs at the good manufacturing practice level in vitro. This strategy might then be used for adoptive immunotherapy for GBM patients in the future [Figure 2]. CMV-specific T cells might constitute a key of the useful immunological tool to attack GBM.
Figure 2.
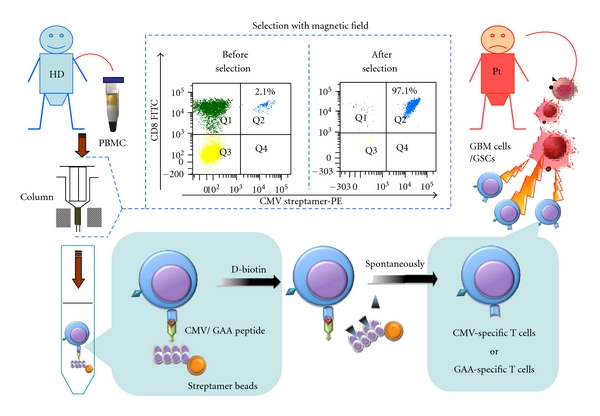
Adoptive immunotherapy for GBM patients with CMV or GAA peptides. CMV and GAAs are highly expressed in GBM, but neither in healthy brain tissue, nor in nonmalignant brain tumors. Therefore, GAAs constitute good targets for immunotherapy of GBM patients. The streptamer technology offers the advantage of selecting CMV- or GAA-specific CD8+ CTLs at the good manufacturing practice (GMP) level in vitro. PBMCs from healthy donors are collected and isolated by streptamer beads. Noninduced antigen-specific T cells are purified and accumulated through a magnetic field and released by D-biotin from the streptamer complex. Subsequently, these cells are administered to the GBM patient. CMV/GAA-specific cytotoxic T cells can recognize the target antigens which are presented on the surface of GBM cells or GSCs. Cytotoxicity is exerted directly through the Fas or perforin pathway and/or indirectly by the release of cytokines. CMV: cytomegalovirus; GAA: glioblastoma associated antigen; GBM: glioblastoma multiforme; GSCs: glioblastoma stem cells; HD: healthy donor; PBMC: peripheral blood mononuclear cells; Pt: patient.
3.3. Role of Regulatory T Cells and Th17 Cells in Immunotherapy
CD4+ regulatory T cells (Tregs) play a key role in maintaining immune homeostasis. They have been well characterized as a distinct subpopulation of T cells due to the identification of the forkhead box transcription factor 3 (Foxp3) as an essential transcription factor in Tregs [132]. The investigation of Treg in brain tumor has blossomed in the last five years. The CD4+, CD25+ (IL-2Rα +), and Foxp3+ Tregs were most frequently found in GBM but very rarely in low-grade astrocytomas and were not present in normal brain tissue. Treg infiltration differed significantly in the brain tumor according to lineage, pathology, and grade. Under the microenvironmental conditions in the GBM patients, Tregs work in several ways to inhibit the effect of T-cell response and act as immune suppressors [133–135]. This might cause the failure of elimination of GBM with glioblastoma infiltrating lymphocytes in clinical trials. Options to eliminate Treg function will likely improve clinical results in future trials. Daclizumab is an approved antibody against IL-2R, which can be used for Treg elimination. Tregs share a common early-stage pathway with another type of CD4+ IL-17A+ T-helper cell, called Th17 cells [136, 137]. Naive T cells upon exposure to antigen and TGF-beta can generate mouse Th17 cells, but not human Th17 cells. For generating Th17 cells, the presence of IL-6 is required. Both cytokines are produced by GBMs. In a melanoma-bearing mouse model, Th17 cells could be used to clear large-established tumor cells [138]. To date, the presence of Th17 cells was confirmed in both human and mouse glioma as well [139], but their beneficial or inhibitory actions have not been fully understood.
3.4. Myeloid-Derived Suppressor Cells (MDSCs)
GBM patients are immunosuppressed and have more circulating myeloid-derived suppressor cells (MDSCs) when compared to normal donors. Interestingly, MDSCs might be generated from glioma-conditioned monocytes in vitro [140]. As Raychaudhuri et al. reported in 2011, GBM patients have increased MDSCs counts (CD33+ HLA-DR−) in their peripheral blood. The accumulation of MDSCs in patients with GBM promotes T-cell immune suppression. Increased plasma levels of arginase and granulocyte colony-stimulating factor may relate to MDSC suppressor function and MDSC expansion, respectively. Removing MDSCs from the PBMCs with anti-CD33/CD15-coated beads significantly restored T-cell function [141].
3.5. GBM Stem Cells (GSCs)
Relapse of GBM is attributed to the persistence of hibernating tumor stem cells [142]. The existence of GBM stem cells is also correlated with multidrug and radiation resistance in GBM [143, 144]. In the past few years, one of the neural progenitor cells marker, CD133, was described as a reasonable marker for GSCs as well [145]. However, some GSCs were also reported to be CD133 negative [146]. So the actual concept of GSCs still needs to be defined.
A safe and effective immune response against rodent gliomas can be elicited by using GSC cell lines as a vaccine in rodent models [147]. Glioblastoma-associated antigens such as AIM2, BMI1, COX-2, TRP2, GP100, EGFRv III, EZH2, LICAM, Livin/Livin β, MRP3, NESTIN, OLIG2, and SOX2 are present on these human GSCs. In contrast, IL-13Rα2 and HER2 seem to be decreased in these GSCs [98]. Two studies suggest that GSCs can differentiate into glioma endothelial cells [148, 149]. GSCs might be considered as sources of antigens for DC vaccination against human GBM, with the aim of achieving GSC-targeting and better antitumor immunity [Figure 1].
4. Future Perspective
Recently, several studies have reported that the combination of chemotherapy and immunotherapy may be more effective than single-modality treatment alone [150, 151]. Kim et al. demonstrated that in a GL26 glioma model, a combination of low-dose TMZ chemotherapy and transactivation of transcription (TAT)-based DC immunotherapy may be a novel strategy for safe and effective treatment of malignant gliomas. TAT contains a protein transduction domain and could be used as an efficient carrier [152].
Our recent work also showed that TMZ might not be deleterious but rather beneficial to immunomodulatory therapy of GBM patients [153].
Future developments in DC vaccination for GBM might include transfection/nucleofection of DCs with RNA encoding for GAA, cytokines, or TLRs.
Acknowledgments
This paper was supported by the German José Carreras Leukemia Foundation (DJCLS, Project no. R09/14 and R10/03). The authors would like to thank Dr. Benjamin Pierce for making suggestions on the paper from the perspective of a native speaker.
References
- 1.Fleury A, Menegoz F, Grosclaude P, et al. Descriptive epidemiology of cerebral gliomas in France. Cancer. 1997;79(6):1195–1202. doi: 10.1002/(sici)1097-0142(19970315)79:6<1195::aid-cncr19>3.0.co;2-v. [DOI] [PubMed] [Google Scholar]
- 2.Mirimanoff RO, Gorlia T, Mason W, et al. Radiotherapy and temozolomide for newly diagnosed glioblastoma: recursive partitioning analysis of the EORTC 26981/22981-NCIC CE3 phase III randomized trial. Journal of Clinical Oncology. 2006;24(16):2563–2569. doi: 10.1200/JCO.2005.04.5963. [DOI] [PubMed] [Google Scholar]
- 3.Stummer W, Reulen HJ, Meinel T, et al. Extent of resection and survival in glioblastoma multiforme: identification of and adjustment for bias. Neurosurgery. 2008;62(3):564–576. doi: 10.1227/01.neu.0000317304.31579.17. [DOI] [PubMed] [Google Scholar]
- 4.Stummer W, Pichlmeier U, Meinel T, Wiestler OD, Zanella F, Reulen HJ. Fluorescence-guided surgery with 5-aminolevulinic acid for resection of malignant glioma: a randomised controlled multicentre phase III trial. The Lancet Oncology. 2006;7(5):392–401. doi: 10.1016/S1470-2045(06)70665-9. [DOI] [PubMed] [Google Scholar]
- 5.Stockhammer F, Misch M, Horn P, Koch A, Fonyuy N, Plotkin M. Association of F18-fluoro-ethyl-tyrosin uptake and 5-aminolevulinic acid-induced fluorescence in gliomas. Acta Neurochirurgica. 2009;151(11):1377–1383. doi: 10.1007/s00701-009-0462-7. [DOI] [PubMed] [Google Scholar]
- 6.Westphal M, Ram Z, Riddle V, Hilt D, Bortey E. Gliadel wafer in initial surgery for malignant glioma: long-term follow-up of a multicenter controlled trial. Acta Neurochirurgica. 2006;148(3):269–275. doi: 10.1007/s00701-005-0707-z. [DOI] [PubMed] [Google Scholar]
- 7.Menei P, Metellus P, Parot-Schinkel E, et al. Biodegradable carmustine wafers (Gliadel) alone or in combination with chemoradiotherapy: the French experience. Annals of Surgical Oncology. 2010;17(7):1740–1746. doi: 10.1245/s10434-010-1081-5. [DOI] [PubMed] [Google Scholar]
- 8.Kreth FW, Warnke PC, Scheremet R, Ostertag CB. Surgical resection and radiation therapy versus biopsy and radiation therapy in the treatment of glioblastoma multiforme. Journal of Neurosurgery. 1993;78(5):762–766. doi: 10.3171/jns.1993.78.5.0762. [DOI] [PubMed] [Google Scholar]
- 9.Kreth FW, Muacevic A, Medele R, Bise K, Meyer T, Reulen HJ. The risk of haemorrhage after image guided stereotactic biopsy of intra-axial brain tumours—a prospective study. Acta Neurochirurgica. 2001;143(6):539–546. doi: 10.1007/s007010170058. [DOI] [PubMed] [Google Scholar]
- 10.Grasbon-Frodl EM, Kreth FW, Ruiter M, et al. Intratumoral homogeneity of MGMT promoter hypermethylation as demonstrated in serial stereotactic specimens from anaplastic astrocytomas and glioblastomas. International Journal of Cancer. 2007;121(11):2458–2464. doi: 10.1002/ijc.23020. [DOI] [PubMed] [Google Scholar]
- 11.Stupp R, Mason WP, Van Den Bent MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. The New England Journal of Medicine. 2005;352(10):987–996. doi: 10.1056/NEJMoa043330. [DOI] [PubMed] [Google Scholar]
- 12.Stupp R, Hegi ME, Mason WP, et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. The Lancet Oncology. 2009;10(5):459–466. doi: 10.1016/S1470-2045(09)70025-7. [DOI] [PubMed] [Google Scholar]
- 13.Hegi ME, Diserens AC, Gorlia T, et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. The New England Journal of Medicine. 2005;352(10):997–1003. doi: 10.1056/NEJMoa043331. [DOI] [PubMed] [Google Scholar]
- 14.Hau P, Koch D, Hundsberger T, et al. Safety and feasibility of long-term temozolomide treatment in patients with high-grade glioma. Neurology. 2007;68(9):688–690. doi: 10.1212/01.wnl.0000255937.27012.ee. [DOI] [PubMed] [Google Scholar]
- 15.Barbagallo GMV, Jenkinson MD, Brodbelt AR. ’Recurrent’ glioblastoma multiforme, when should we reoperate? British Journal of Neurosurgery. 2008;22(3):452–455. doi: 10.1080/02688690802182256. [DOI] [PubMed] [Google Scholar]
- 16.Mandl ES, Dirven CMF, Buis DR, Postma TJ, Vandertop WP. Repeated surgery for glioblastoma multiforme: only in combination with other salvage therapy. Surgical Neurology. 2008;69(5):506–509. doi: 10.1016/j.surneu.2007.03.043. [DOI] [PubMed] [Google Scholar]
- 17.Combs SE, Bischof M, Welzel T, et al. Radiochemotherapy with temozolomide as re-irradiation using high precision fractionated stereotactic radiotherapy (FSRT) in patients with recurrent gliomas. Journal of Neuro-Oncology. 2008;89(2):205–210. doi: 10.1007/s11060-008-9607-4. [DOI] [PubMed] [Google Scholar]
- 18.Villavicencio AT, Burneikiene S, Romanelli P, et al. Survival following stereotactic radiosurgery for newly diagnosed and recurrent glioblastoma multiforme: a multicenter experience. Neurosurgical Review. 2009;32(4):417–424. doi: 10.1007/s10143-009-0212-6. [DOI] [PubMed] [Google Scholar]
- 19.Cloughesy T. FDA accelerated approval benefits glioblastoma. The Lancet Oncology. 2010;11(12):p. 1120. doi: 10.1016/S1470-2045(10)70269-2. [DOI] [PubMed] [Google Scholar]
- 20.Vredenburgh JJ, Desjardins A, Herndon JE, et al. Phase II trial of bevacizumab and irinotecan in recurrent malignant glioma. Clinical Cancer Research. 2007;13(4):1253–1259. doi: 10.1158/1078-0432.CCR-06-2309. [DOI] [PubMed] [Google Scholar]
- 21.Vredenburgh JJ, Desjardins A, Herndon JE, et al. Bevacizumab plus irinotecan in recurrent glioblastoma multiforme. Journal of Clinical Oncology. 2007;25(30):4722–4729. doi: 10.1200/JCO.2007.12.2440. [DOI] [PubMed] [Google Scholar]
- 22.Friedman HS, Prados MD, Wen PY, et al. Bevacizumab alone and in combination with irinotecan in recurrent glioblastoma. Journal of Clinical Oncology. 2009;27(28):4733–4740. doi: 10.1200/JCO.2008.19.8721. [DOI] [PubMed] [Google Scholar]
- 23.Brandes AA, Tosoni A, Cavallo G, et al. Temozolomide 3 weeks on and 1 week off as first-line therapy for recurrent glioblastoma: phase II study from gruppo italiano cooperativo di neuro-oncologia (GICNO) British Journal of Cancer. 2006;95(9):1155–1160. doi: 10.1038/sj.bjc.6603376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wick A, Felsberg J, Steinbach JP, et al. Efficacy and tolerability of temozolomide in an alternating weekly regimen in patients with recurrent glioma. Journal of Clinical Oncology. 2007;25(22):3357–3361. doi: 10.1200/JCO.2007.10.7722. [DOI] [PubMed] [Google Scholar]
- 25.Perry JR, Rizek P, Cashman R, Morrison M, Morrison T. Temozolomide rechallenge in recurrent malignant glioma by using a continuous temozolomide schedule: the "rescue" approach. Cancer. 2008;113(8):2152–2157. doi: 10.1002/cncr.23813. [DOI] [PubMed] [Google Scholar]
- 26.Stockhammer F, Misch M, Koch A, et al. Continuous low-dose temozolomide and celecoxib in recurrent glioblastoma. Journal of Neuro-Oncology. 2010;100(3):407–415. doi: 10.1007/s11060-010-0192-y. [DOI] [PubMed] [Google Scholar]
- 27.Hau P, Baumgart U, Pfeifer K, et al. Salvage therapy in patients with glioblastoma: is there any benefit? Cancer. 2003;98(12):2678–2686. doi: 10.1002/cncr.11845. [DOI] [PubMed] [Google Scholar]
- 28.Hong S, Van Kaer L. Immune privilege: keeping an eye on natural killer T cells. The Journal of Experimental Medicine. 1999;190(9):1197–1200. doi: 10.1084/jem.190.9.1197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kanaly CW, Ding D, Heimberger AB, Sampson JH. Clinical applications of a peptide-based vaccine for glioblastoma. Neurosurgery Clinics of North America. 2010;21(1):95–109. doi: 10.1016/j.nec.2009.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Thara E, Dorff TB, Pinski JK, Quinn DI. Vaccine therapy with sipuleucel-T (Provenge) for prostate cancer. Maturitas. 2011;69(4):296–303. doi: 10.1016/j.maturitas.2011.04.012. [DOI] [PubMed] [Google Scholar]
- 31.Jacobs SK, Wilson DJ, Kornblith PL, Grimm EA. Interleukin-2 or autologous lymphokine-activated killer cell treatment of malignant glioma: phase I trial. Cancer Research. 1986;46(4):2101–2104. [PubMed] [Google Scholar]
- 32.Merchant RE, Grant AJ, Merchant LH, Young HF. Adoptive immunotherapy for recurrent glioblastoma multiforme using lymphokine activated killer cells and recombinant interleukin-2. Cancer. 1988;62(4):665–671. doi: 10.1002/1097-0142(19880815)62:4<665::aid-cncr2820620403>3.0.co;2-o. [DOI] [PubMed] [Google Scholar]
- 33.Barba D, Saris SC, Holder C, Rosenberg SA, Oldfield EH. Intratumoral LAK cell and interleukin-2 therapy of human gliomas. Journal of Neurosurgery. 1989;70(2):175–182. doi: 10.3171/jns.1989.70.2.0175. [DOI] [PubMed] [Google Scholar]
- 34.Lillehei KO, Mitchell DH, Johnson SD, McCleary EL, Kruse CA. Long-term follow-up of patients with recurrent malignant gliomas treated with adjuvant adoptive immunotherapy. Neurosurgery. 1991;28(1):16–23. doi: 10.1097/00006123-199101000-00003. [DOI] [PubMed] [Google Scholar]
- 35.Jeffes EWB, III, Beamer YB, Jacques S, et al. Therapy of recurrent high grade gliomas with surgery, and autologous mitogen activated IL-2 stimulated killer (MAK) lymphocytes: I. Enhancement of MAK lytic activity and cytokine production by PHA and clinical use of PHA. Journal of Neuro-Oncology. 1993;15(2):141–155. doi: 10.1007/BF01053935. [DOI] [PubMed] [Google Scholar]
- 36.Blancher A, Roubinet F, Grancher AS, et al. Local immunotherapy of recurrent glioblastoma multiforme by intracerebral perfusion of interleukin-2 and LAK cells. European Cytokine Network. 1993;4(5):331–341. [PubMed] [Google Scholar]
- 37.Boiardi A, Silvani A, Adelchi Ruffini P, et al. Loco-regional immunotherapy with recombinant interleukin-2 and adherent lymphokine-activated killer cells (A-LAK) in recurrent glioblastoma patients. Cancer Immunology Immunotherapy. 1994;39(3):193–197. doi: 10.1007/BF01533386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hayes RL, Koslow M, Hiesiger EM, et al. Improved long term survival after intracavitary interleukin-2 and lymphokine-activated killer cells for adults with recurrent malignant glioma. Cancer. 1995;76(5):840–852. doi: 10.1002/1097-0142(19950901)76:5<840::aid-cncr2820760519>3.0.co;2-r. [DOI] [PubMed] [Google Scholar]
- 39.Sankhla SK, Nadkarni JS, Bhagwati SN. Adoptive immimotherapy using lymphokine-activated killer (LAK) cells and interleukin-2 for recurrentmalignant primary brain tumors. Journal of Neuro-Oncology. 1995;27(2):133–140. doi: 10.1007/BF00177476. [DOI] [PubMed] [Google Scholar]
- 40.Hayes RL, Arbit E, Odaimi M, et al. Adoptive cellular immunotherapy for the treatment of malignant gliomas. Critical Reviews in Oncology/Hematology. 2001;39(1-2):31–42. doi: 10.1016/s1040-8428(01)00122-6. [DOI] [PubMed] [Google Scholar]
- 41.Dillman RO, Duma CM, Schiltz PM, et al. Intracavitary placement of autologous lymphokine-activated killer (LAK) cells after resection of recurrent glioblastoma. Journal of Immunotherapy. 2004;27(5):398–404. doi: 10.1097/00002371-200409000-00009. [DOI] [PubMed] [Google Scholar]
- 42.Dillman RO, Duma CM, Ellis RA, et al. Intralesional lymphokine-activated killer cells as adjuvant therapy for primary glioblastoma. Journal of Immunotherapy. 2009;32(9):914–919. doi: 10.1097/CJI.0b013e3181b2910f. [DOI] [PubMed] [Google Scholar]
- 43.Holladay FP. Autologous tumor cell vaccination combined with adoptive cellular immunotherapy in patients with Grade III/IV astrocytoma. Journal of Neuro-Oncology. 1996;27(2):179–189. doi: 10.1007/BF00177482. [DOI] [PubMed] [Google Scholar]
- 44.Plautz GE, Barnett GH, Miller DW, et al. Systemic T cell adoptive immunotherapy of malignant gliomas. Journal of Neurosurgery. 1998;89(1):42–51. doi: 10.3171/jns.1998.89.1.0042. [DOI] [PubMed] [Google Scholar]
- 45.Plautz GE, Miller DW, Barnett GH, et al. T cell adoptive immunotherapy of newly diagnosed gliomas. Clinical Cancer Research. 2000;6(6):2209–2218. [PubMed] [Google Scholar]
- 46.Wood GW, Holladay FP, Turner T, Wang YY, Chiga M. A pilot study of autologous cancer cell vaccination and cellular immunotherapy using anti-CD3 stimulated lymphocytes in patients with recurrent grade III/IV astrocytoma. Journal of Neuro-Oncology. 2000;48(2):113–120. doi: 10.1023/a:1006456421177. [DOI] [PubMed] [Google Scholar]
- 47.Sloan AE, Dansey R, Zamorano L, et al. Adoptive immunotherapy in patients with recurrent malignant glioma: preliminary results of using autologous whole-tumor vaccine plus granulocyte-macrophage colony-stimulating factor and adoptive transfer of anti-CD3-activated lymphocytes. Neurosurgical Focus. 2000;9(6):p. e9. doi: 10.3171/foc.2000.9.6.10. [DOI] [PubMed] [Google Scholar]
- 48.Kruse CA, Cepeda L, Owens B, Johnson SD, Stears J, Lillehei KO. Treatment of recurrent glioma with intracavitary alloreactive cytotoxic T lymphocytes and interleukin-2. Cancer Immunology Immunotherapy. 1997;45(2):77–87. doi: 10.1007/s002620050405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Quattrocchi KB, Miller CH, Cush S, et al. Pilot study of local autologous tumor infiltrating lymphocytes for the treatment of recurrent malignant gliomas. Journal of Neuro-Oncology. 1999;45(2):141–157. doi: 10.1023/a:1006293606710. [DOI] [PubMed] [Google Scholar]
- 50.Kronik N, Kogan Y, Vainstein V, Agur Z. Improving alloreactive CTL immunotherapy for malignant gliomas using a simulation model of their interactive dynamics. Cancer Immunology, Immunotherapy. 2008;57(3):425–439. doi: 10.1007/s00262-007-0387-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Sobol RE, Fakhrai H, Shawler D, et al. Interleukin-2 gene therapy in a patient with glioblastoma. Gene Therapy. 1995;2(2):164–167. [PubMed] [Google Scholar]
- 52.Schneider T, Gerhards R, Kirches E, Firsching R. Preliminary results of active specific immunization with modified tumor cell vaccine in glioblastoma multiforme. Journal of Neuro-Oncology. 2001;53(1):39–46. doi: 10.1023/a:1011856406683. [DOI] [PubMed] [Google Scholar]
- 53.Andrews DW, Resnicoff M, Flanders AE, et al. Results of a pilot study involving the use of an antisense oligodeoxynucleotide directed against the insulin-like growth factor type I receptor in malignant astrocytomas. Journal of Clinical Oncology. 2001;19(8):2189–2200. doi: 10.1200/JCO.2001.19.8.2189. [DOI] [PubMed] [Google Scholar]
- 54.Okada H, Lieberman FS, Edington HD, et al. Autologous glioma cell vaccine admixed with interleukin-4 gene transfected fibroblasts in the treatment of recurrent glioblastoma: preliminary observations in a patient with a favorable response to therapy. Journal of Neuro-Oncology. 2003;64(1-2):13–20. doi: 10.1007/BF02700016. [DOI] [PubMed] [Google Scholar]
- 55.Steiner HH, Bonsanto MM, Beckhove P, et al. Antitumor vaccination of patients with glioblastoma multiforme: a pilot study to assess feasibility, safety, and clinical benefits. Journal of Clinical Oncology. 2004;22(21):4272–4281. doi: 10.1200/JCO.2004.09.038. [DOI] [PubMed] [Google Scholar]
- 56.Parney IF, Chang LJ, Farr-Jones MA, Hao C, Smylie M, Petruk KC. Technical hurdles in a pilot clinical trial of combined B7-2 and GM-CSF immunogene therapy for glioblastomas and melanomas. Journal of Neuro-Oncology. 2006;78(1):71–80. doi: 10.1007/s11060-005-9058-0. [DOI] [PubMed] [Google Scholar]
- 57.Ishikawa E, Tsuboi K, Yamamoto T, et al. Clinical trial of autologous formalin-fixed tumor vaccine for glioblastoma multiforme patients. Cancer Science. 2007;98(8):1226–1233. doi: 10.1111/j.1349-7006.2007.00518.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Clavreul A, Piard N, Tanguy JY, et al. Autologous tumor cell vaccination plus infusion of GM-CSF by a programmable pump in the treatment of recurrent malignant gliomas. Journal of Clinical Neuroscience. 2010;17(7):842–848. doi: 10.1016/j.jocn.2009.11.017. [DOI] [PubMed] [Google Scholar]
- 59.Banchereau J, Briere F, Caux C, et al. Immunobiology of dendritic cells. Annual Review of Immunology. 2000;18:767–811. doi: 10.1146/annurev.immunol.18.1.767. [DOI] [PubMed] [Google Scholar]
- 60.Steinman RM, Banchereau J. Taking dendritic cells into medicine. Nature. 2007;449(7161):419–426. doi: 10.1038/nature06175. [DOI] [PubMed] [Google Scholar]
- 61.Okada H, Kalinski P, Ueda R, et al. Induction of CD8+ T-cell responses against novel glioma-associated antigen peptides and clinical activity by vaccinations with α-type 1 polarized dendritic cells and polyinosinic-polycytidylic acid stabilized by lysine and carboxymethylcellulose in patients with recurrent malignant glioma. Journal of Clinical Oncology. 2011;29(3):330–336. doi: 10.1200/JCO.2010.30.7744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Prins RM, Soto H, Konkankit V, et al. Gene expression profile correlates with T-cell infiltration and relative survival in glioblastoma patients vaccinated with dendritic cell immunotherapy. Clinical Cancer Research. 2011;17(6):1603–1615. doi: 10.1158/1078-0432.CCR-10-2563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ardon H, Van Gool S, Lopes IS, et al. Integration of autologous dendritic cell-based immunotherapy in the primary treatment for patients with newly diagnosed glioblastoma multiforme: a pilot study. Journal of Neuro-Oncology. 2010;99(2):261–272. doi: 10.1007/s11060-010-0131-y. [DOI] [PubMed] [Google Scholar]
- 64.Ardon H, De Vleeschouwer S, Van Calenbergh F, et al. Adjuvant dendritic cell-based tumour vaccination for children with malignant brain tumours. Pediatric Blood and Cancer. 2010;54(4):519–525. doi: 10.1002/pbc.22319. [DOI] [PubMed] [Google Scholar]
- 65.Sampson JH, Archer GE, Mitchell DA, et al. An epidermal growth factor receptor variant III-targeted vaccine is safe and immunogenic in patients with glioblastoma multiforme. Molecular Cancer Therapeutics. 2009;8(10):2773–2779. doi: 10.1158/1535-7163.MCT-09-0124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.De Vleeschouwer S, Fieuws S, Rutkowski S, et al. Postoperative adjuvant dendritic cell—based immunotherapy in patients with relapsed glioblastoma multiforme. Clinical Cancer Research. 2008;14(10):3098–3104. doi: 10.1158/1078-0432.CCR-07-4875. [DOI] [PubMed] [Google Scholar]
- 67.Wheeler CJ, Black KL, Liu G, et al. Vaccination elicits correlated immune and clinical responses in glioblastoma multiforme patients. Cancer Research. 2008;68(14):5955–5964. doi: 10.1158/0008-5472.CAN-07-5973. [DOI] [PubMed] [Google Scholar]
- 68.Yamanaka R, Homma J, Yajima N, et al. Clinical evaluation of dendritic cell vaccination for patients with recurrent glioma: results of a clinical phase I/II trial. Clinical Cancer Research. 2005;11(11):4160–4167. doi: 10.1158/1078-0432.CCR-05-0120. [DOI] [PubMed] [Google Scholar]
- 69.Liau LM, Prins RM, Kiertscher SM, et al. Dendritic cell vaccination in glioblastoma patients induces systemic and intracranial T-cell responses modulated by the local central nervous system tumor microenvironment. Clinical Cancer Research. 2005;11(15):5515–5525. doi: 10.1158/1078-0432.CCR-05-0464. [DOI] [PubMed] [Google Scholar]
- 70.Yu JS, Liu G, Ying H, Yong WH, Black KL, Wheeler CJ. Vaccination with tumor lysate-pulsed dendritic cells elicits antigen-specific, cytotoxic T-cells in patients with malignant glioma. Cancer Research. 2004;64(14):4973–4979. doi: 10.1158/0008-5472.CAN-03-3505. [DOI] [PubMed] [Google Scholar]
- 71.Kikuchi T, Akasaki Y, Abe T, et al. Vaccination of glioma patients with fusions of dendritic and glioma cells and recombinant human interleukin 12. Journal of Immunotherapy. 2004;27(6):452–459. doi: 10.1097/00002371-200411000-00005. [DOI] [PubMed] [Google Scholar]
- 72.Caruso DA, Orme LM, Neale AM, et al. Results of a phase 1 study utilizing monocyte-derived dendritic cells pulsed with tumor RNA in children and young adults with brain cancer. Neuro-Oncology. 2004;6(3):236–246. doi: 10.1215/S1152851703000668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Wheeler CJ, Das A, Liu G, Yu JS, Black KL. Clinical responsiveness of glioblastoma multiforme to chemotherapy after vaccination. Clinical Cancer Research. 2004;10(16):5316–5326. doi: 10.1158/1078-0432.CCR-04-0497. [DOI] [PubMed] [Google Scholar]
- 74.Yamanaka R, Abe T, Yajima N, et al. Vaccination of recurrent glioma patients with tumour lysate-pulsed dendritic cells elicits immune responses: results of a clinical phase I/II trial. British Journal of Cancer. 2003;89(7):1172–1179. doi: 10.1038/sj.bjc.6601268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Yu JS, Wheeler CJ, Zeltzer PM, et al. Vaccination of malignant glioma patients with peptide-pulsed dendritic cells elicits systemic cytotoxicity and intracranial T-cell infiltration. Cancer Research. 2001;61(3):842–847. [PubMed] [Google Scholar]
- 76.Wheeler CJ, Black KL, Liu G, et al. Thymic CD8+ T cell production strongly influences tumor antigen recognition and age-dependent glioma mortality. The Journal of Immunology. 2003;171(9):4927–4933. doi: 10.4049/jimmunol.171.9.4927. [DOI] [PubMed] [Google Scholar]
- 77.Tuyaerts S, Aerts JL, Corthals J, et al. Current approaches in dendritic cell generation and future implications for cancer immunotherapy. Cancer Immunology, Immunotherapy. 2007;56(10):1513–1537. doi: 10.1007/s00262-007-0334-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Okada H, Lieberman FS, Walter KA, et al. Autologous glioma cell vaccine admixed with interleukin-4 gene transfected fibroblasts in the treatment of patients with malignant gliomas. Journal of Translational Medicine. 2007;5, article 67 doi: 10.1186/1479-5876-5-67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Kogiku M, Ohsawa I, Matsumoto K, et al. Prognosis of glioma patients by combined immunostaining for survivin, Ki-67 and epidermal growth factor receptor. Journal of Clinical Neuroscience. 2008;15(11):1198–1203. doi: 10.1016/j.jocn.2007.11.012. [DOI] [PubMed] [Google Scholar]
- 80.Rosenberg SA. Cancer vaccines based on the identification of genes encoding cancer regression antigens. Immunology Today. 1997;18(4):175–182. doi: 10.1016/s0167-5699(97)84664-6. [DOI] [PubMed] [Google Scholar]
- 81.Wang RF. The role of MHC class II-restricted tumor antigens and CD4+ T cells in antitumor immunity. Trends in Immunology. 2001;22(5):269–276. doi: 10.1016/s1471-4906(01)01896-8. [DOI] [PubMed] [Google Scholar]
- 82.Fernandez NC, Lozier A, Flament C, et al. Dendritic cells directly trigger NK cell functions: cross-talk relevant in innate anti-tumor immune responses in vivo. Nature Medicine. 1999;5(4):405–411. doi: 10.1038/7403. [DOI] [PubMed] [Google Scholar]
- 83.Wehner R, Löbel B, Bornhäuser M, et al. Reciprocal activating interaction between 6-sulfo LacNAc+ dendritic cells and NK cell. International Journal of Cancer. 2009;124(2):358–366. doi: 10.1002/ijc.23962. [DOI] [PubMed] [Google Scholar]
- 84.Schmitz M, Zhao S, Deuse Y, et al. Tumoricidal potential of native blood dendritic cells: direct tumor cell killing and activation of NK cell-mediated cytotoxicity. The Journal of Immunology. 2005;174(7):4127–4134. doi: 10.4049/jimmunol.174.7.4127. [DOI] [PubMed] [Google Scholar]
- 85.Fadul CE, Fisher JL, Hampton TH, et al. Immune response in patients with newly diagnosed glioblastoma multiforme treated with intranodal autologous tumor lysate-dendritic cell vaccination after radiation chemotherapy. Journal of Immunotherapy. 2011;34(4):382–389. doi: 10.1097/CJI.0b013e318215e300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Hatano M, Eguchi J, Tatsumi T, et al. EphA2 as a glioma-associated antigen: a novel target for glioma vaccines. Neoplasia. 2005;7(8):717–722. doi: 10.1593/neo.05277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Schmitz M, Wehner R, Stevanovic S, et al. Identification of a naturally processed T cell epitope derived from the glioma-associated protein SOX11. Cancer Letters. 2007;245(1-2):331–336. doi: 10.1016/j.canlet.2006.01.014. [DOI] [PubMed] [Google Scholar]
- 88.Liu G, Ying H, Zeng G, Wheeler CJ, Black KL, Yu JS. HER-2, gp100, and MAGE-1 are expressed in human glioblastoma and recognized by cytotoxic T cells. Cancer Research. 2004;64(14):4980–4986. doi: 10.1158/0008-5472.CAN-03-3504. [DOI] [PubMed] [Google Scholar]
- 89.Saikali S, Avril T, Collet B, et al. Expression of nine tumour antigens in a series of human glioblastoma multiforme: interest of EGFRvIII, IL-13Rα2, gp100 and TRP-2 for immunotherapy. Journal of Neuro-Oncology. 2007;81(2):139–148. doi: 10.1007/s11060-006-9220-3. [DOI] [PubMed] [Google Scholar]
- 90.Okada H, Kohanbash G, Zhu X, et al. Immunotherapeutic approaches for glioma. Critical Reviews in Immunology. 2009;29(1):1–42. doi: 10.1615/critrevimmunol.v29.i1.10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Ueda R, Low KL, Zhu X, et al. Spontaneous immune responses against glioma-associated antigens in a long term survivor with malignant glioma. Journal of Translational Medicine. 2007;5, article 68 doi: 10.1186/1479-5876-5-68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Zhang JG, Kruse CA, Driggers L, et al. Tumor antigen precursor protein profiles of adult and pediatric brain tumors identify potential targets for immunotherapy. Journal of Neuro-Oncology. 2008;88(1):65–76. doi: 10.1007/s11060-008-9534-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Scarcella DL, Chow CW, Gonzales MF, Economou C, Brasseur F, Ashley DM. Expression of MAGE and GAGE in high-grade brain tumors: a potential target for specific immunotherapy and diagnostic markers. Clinical Cancer Research. 1999;5(2):335–341. [PubMed] [Google Scholar]
- 94.Bao L, Dunham K, Lucas K. MAGE-A1, MAGE-A3, and NY-ESO-1 can be upregulated on neuroblastoma cells to facilitate cytotoxic T lymphocyte-mediated tumor cell killing. Cancer Immunology, Immunotherapy. 2011;60(9):1299–1307. doi: 10.1007/s00262-011-1037-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Jian GZ, Eguchi J, Kruse CA, et al. Antigenic profiling of glioma cells to generate allogeneic vaccines or dendritic cell-based therapeutics. Clinical Cancer Research. 2007;13(2):566–575. doi: 10.1158/1078-0432.CCR-06-1576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Liu G, Yu JS, Zeng G, et al. AIM-2: a novel tumor antigen is expressed and presented by human glioma cells. Journal of Immunotherapy. 2004;27(3):220–226. doi: 10.1097/00002371-200405000-00006. [DOI] [PubMed] [Google Scholar]
- 97.Driggers L, Zhang JG, Newcomb EW, Ge L, Hoa N, Jadus MR. Immunotherapy of pediatric brain tumor patients should include an immunoprevention strategy: a medical hypothesis paper. Journal of Neuro-Oncology. 2010;97(2):159–169. doi: 10.1007/s11060-009-0016-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Xu Q, Liu G, Yuan X, et al. Antigen-specific T-cell response from dendritic cell vaccination using cancer stem-like cell-associated antigens. Stem Cells. 2009;27(8):1734–1740. doi: 10.1002/stem.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Abdouh M, Facchino S, Chatoo W, Balasingam V, Ferreira J, Bernier G. BMI1 sustains human glioblastoma multiforme stem cell renewal. The Journal of Neuroscience. 2009;29(28):8884–8896. doi: 10.1523/JNEUROSCI.0968-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Shono T, Tofilon PJ, Bruner JM, Owolabi O, Lang FF. Cyclooxygenase-2 expression in human gliomas: prognostic significance and molecular correlations. Cancer Research. 2001;61(11):4375–4381. [PubMed] [Google Scholar]
- 101.Orzan F, Pellegatta S, Poliani PL, et al. Enhancer of Zeste 2 (EZH2) is up-regulated in malignant gliomas and in glioma stem-like cells. Neuropathology and Applied Neurobiology. 2011;37(4):381–394. doi: 10.1111/j.1365-2990.2010.01132.x. [DOI] [PubMed] [Google Scholar]
- 102.Cheng L, Wu Q, Guryanova OA, et al. Elevated invasive potential of glioblastoma stem cells. Biochemical and Biophysical Research Communications. 2011;406(4):643–648. doi: 10.1016/j.bbrc.2011.02.123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Jin F, Zhao L, Guo YJ, et al. Influence of Etoposide on anti-apoptotic and multidrug resistance-associated protein genes in CD133 positive U251 glioblastoma stem-like cells. Brain Research. 2010;1336:103–111. doi: 10.1016/j.brainres.2010.04.005. [DOI] [PubMed] [Google Scholar]
- 104.Kuan CT, Wakiya K, Herndon JE, et al. MRP3: a molecular target for human glioblastoma multiforme immunotherapy. BMC Cancer. 2010;10, article 468 doi: 10.1186/1471-2407-10-468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Ishiwata T, Teduka K, Yamamoto T, Kawahara K, Matsuda Y, Naito Z. Neuroepithelial stem cell marker nestin regulates the migration, invasion and growth of human gliomas. Oncology Reports. 2011;26(1):91–99. doi: 10.3892/or.2011.1267. [DOI] [PubMed] [Google Scholar]
- 106.Ligon KL, Alberta JA, Kho AT, et al. The oligodendroglial lineage marker OLIG2 is universally expressed in diffuse gliomas. Journal of Neuropathology and Experimental Neurology. 2004;63(5):499–509. doi: 10.1093/jnen/63.5.499. [DOI] [PubMed] [Google Scholar]
- 107.Annovazzi L, Mellai M, Caldera V, Valente G, Schiffer D. SOX2 expression and amplification in gliomas and glioma cell lines. Cancer Genomics and Proteomics. 2011;8(3):139–147. [PubMed] [Google Scholar]
- 108.Kakino S, Sasaki K, Kurose A, Ito H. Intracellular localization of cyclin B1 during the cell cycle in glioma cells. Cytometry. 1996;24(1):49–54. doi: 10.1002/(SICI)1097-0320(19960501)24:1<49::AID-CYTO6>3.0.CO;2-D. [DOI] [PubMed] [Google Scholar]
- 109.Rossi M, Magnoni L, Miracco C, et al. β-catenin and Gli1 are prognostic markers in glioblastoma. Cancer Biology and Therapy. 2011;11(8):753–761. doi: 10.4161/cbt.11.8.14894. [DOI] [PubMed] [Google Scholar]
- 110.Cui D, Xu Q, Wang K, Che X. Gli1 is a potential target for alleviating multidrug resistance of gliomas. Journal of the Neurological Sciences. 2010;288(1-2):156–166. doi: 10.1016/j.jns.2009.09.006. [DOI] [PubMed] [Google Scholar]
- 111.Senetta R, Miracco C, Lanzafame S, et al. Epidermal growth factor receptor and caveolin-1 coexpression identifies adult supratentorial ependymomas with rapid unfavorable outcomes. Neuro-Oncology. 2011;13(2):176–183. doi: 10.1093/neuonc/noq160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Sivaparvathi M, Sawaya R, Wang SW, et al. Overexpression and localization of cathepsin B during the progression of human gliomas. Clinical and Experimental Metastasis. 1995;13(1):49–56. doi: 10.1007/BF00144018. [DOI] [PubMed] [Google Scholar]
- 113.Kitange GJ, Carlson BL, Schroeder MA, et al. Expression of CD74 in high grade gliomas: a potential role in temozolomide resistance. Journal of Neuro-Oncology. 2010;100(2):177–186. doi: 10.1007/s11060-010-0186-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Lewis-Tuffin LJ, Rodriguez F, Giannini C, et al. Misregulated E-cadherin expression associated with an aggressive brain tumor phenotype. PLoS ONE. 2010;5(10) doi: 10.1371/journal.pone.0013665. Article ID e13665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Wykosky J, Gibo DM, Stanton C, Debinski W. EphA2 as a novel molecular marker and target in glioblastoma multiforme. Molecular Cancer Research. 2005;3(10):541–551. doi: 10.1158/1541-7786.MCR-05-0056. [DOI] [PubMed] [Google Scholar]
- 116.Wykosky J, Gibo DM, Stanton C, Debinski W. Interleukin-13 receptor α2, EphA2, and Fos-related antigen 1 as molecular denominators of high-grade astrocytomas and specific targets for combinatorial therapy. Clinical Cancer Research. 2008;14(1):199–208. doi: 10.1158/1078-0432.CCR-07-1990. [DOI] [PubMed] [Google Scholar]
- 117.Mennel HD, Lell B. Ganglioside (GD2) expression and intermediary filaments in astrocytic tumors. Clinical Neuropathology. 2005;24(1):13–18. [PubMed] [Google Scholar]
- 118.Yamamoto H, Swoger J, Greene S, et al. β1,6-N-acethylglucosamine-bearing N-glycans in human gliomas: implications for a role in regulating invasivity. Cancer Research. 2000;60(1):134–142. [PubMed] [Google Scholar]
- 119.Persson O, Salford LG, Fransson J, Widegren B, Borrebaeck CAK, Holmqvist B. Distribution, cellular localization, and therapeutic potential of the tumor-associated antigen Ku70/80 in glioblastoma multiforme. Journal of Neuro-Oncology. 2010;97(2):207–215. doi: 10.1007/s11060-009-0013-3. [DOI] [PubMed] [Google Scholar]
- 120.Elsir T, Eriksson A, Orrego A, Lindström MS, Nistér M. Expression of PROX1 is a common feature of high-grade malignant astrocytic gliomas. Journal of Neuropathology and Experimental Neurology. 2010;69(2):129–138. doi: 10.1097/NEN.0b013e3181ca4767. [DOI] [PubMed] [Google Scholar]
- 121.Geiger KD, Hendruschk S, Rieber EP, et al. The prostate stem cell antigen represents a novel glioma-associated antigen. Oncology Reports. 2011;26(1):13–21. doi: 10.3892/or.2011.1265. [DOI] [PubMed] [Google Scholar]
- 122.Ferletta M, Uhrbom L, Olofsson T, Pontén F, Westermark B. Sox10 has a broad expression pattern in gliomas and enhances platelet-derived growth factor-B—induced gliomagenesis. Molecular Cancer Research. 2007;5(9):891–897. doi: 10.1158/1541-7786.MCR-07-0113. [DOI] [PubMed] [Google Scholar]
- 123.Gondi CS, Lakka SS, Dinh DH, Olivero WC, Gujrati M, Rao JS. RNAi-mediated inhibition of cathepsin B and uPAR leads to decreased cell invasion, angiogenesis and tumor growth in gliomas. Oncogene. 2004;23(52):8486–8496. doi: 10.1038/sj.onc.1207879. [DOI] [PubMed] [Google Scholar]
- 124.Gomez GG, Kruse CA. Mechanisms of malignant glioma immune resistance and sources of immunosuppression. Gene Therapy and Molecular Biology. 2006;10(1):133–146. [PMC free article] [PubMed] [Google Scholar]
- 125.Cobbs CS, Harkins L, Samanta M, et al. Human cytomegalovirus infection and expression in human malignant glioma. Cancer Research. 2002;62(12):3347–3350. [PubMed] [Google Scholar]
- 126.Prins RM, Cloughesy TF, Liau LM. Cytomegalovirus immunity after vaccination with autologous glioblastoma lysate. New England Journal of Medicine. 2008;359(5):539–541. doi: 10.1056/NEJMc0804818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Mitchell DA, Xie W, Schmittling R, et al. Sensitive detection of human cytomegalovirus in tumors and peripheral blood of patients diagnosed with glioblastoma. Neuro-Oncology. 2008;10(1):10–18. doi: 10.1215/15228517-2007-035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Bao L, Dunham K, Stamer M, Mulieri KM, Lucas KG. Expansion of cytomegalovirus pp65 and IE-1 specific cytotoxic T lymphocytes for cytomegalovirus-specific immunotherapy following allogeneic stem cell transplantation. Biology of Blood and Marrow Transplantation. 2008;14(10):1156–1162. doi: 10.1016/j.bbmt.2008.07.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Bao L, Sun Q, Lucas KG. Rapid generation of CMV pp65-specific T cells for immunotherapy. Journal of Immunotherapy. 2007;30(5):557–561. doi: 10.1097/CJI.0b013e31803b945b. [DOI] [PubMed] [Google Scholar]
- 130.Yao J, Bechter C, Wiesneth M, et al. Multimer staining of cytomegalovirus phosphoprotein 65-specific T cells for diagnosis and therapeutic purposes: a comparative study. Clinical Infectious Diseases. 2008;46(10):e96–e105. doi: 10.1086/587749. [DOI] [PubMed] [Google Scholar]
- 131.Schmitt A, Tonn T, Busch DH, et al. Adoptive transfer and selective reconstitution of streptamer-selected cytomegalovirus-specific CD8+ T cells leads to virus clearance in patients after allogeneic peripheral blood stem cell transplantation. Transfusion. 2011;51(3):591–599. doi: 10.1111/j.1537-2995.2010.02940.x. [DOI] [PubMed] [Google Scholar]
- 132.Fontenot JD, Rasmussen JP, Gavin MA, Rudensky AY. A function for interleukin 2 in Foxp3-expressing regulatory T cells. Nature Immunology. 2005;6(11):1142–1151. doi: 10.1038/ni1263. [DOI] [PubMed] [Google Scholar]
- 133.El Andaloussi A, Lesniak MS. An increase in CD4+CD25+FOXP3+ regulatory T cells in tumor-infiltrating lymphocytes of human glioblastoma multiforme. Neuro-Oncology. 2006;8(3):234–243. doi: 10.1215/15228517-2006-006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Fecci PE, Mitchell DA, Whitesides JF, et al. Increased regulatory T-cell fraction amidst a diminished CD4 compartment explains cellular immune defects in patients with malignant glioma. Cancer Research. 2006;66(6):3294–3302. doi: 10.1158/0008-5472.CAN-05-3773. [DOI] [PubMed] [Google Scholar]
- 135.Heimberger AB, Abou-Ghazal M, Reina-Ortiz C, et al. Incidence and prognostic impact of FoxP3+ regulatory T cells in human gliomas. Clinical Cancer Research. 2008;14(16):5166–5172. doi: 10.1158/1078-0432.CCR-08-0320. [DOI] [PubMed] [Google Scholar]
- 136.Annunziato F, Cosmi L, Liotta F, Maggi E, Romagnani S. The phenotype of human Th17 cells and their precursors, the cytokines that mediate their differentiation and the role of Th17 cells in inflammation. International Immunology. 2008;20(11):1361–1368. doi: 10.1093/intimm/dxn106. [DOI] [PubMed] [Google Scholar]
- 137.Wan YY, Flave RA., I I How diverse—CD4 effector T cells and their functions. Journal of Molecular Cell Biology. 2009;1(1):20–36. doi: 10.1093/jmcb/mjp001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Muranski P, Boni A, Antony PA, et al. Tumor-specific Th17-polarized cells eradicate large established melanoma. Blood. 2008;112(2):362–373. doi: 10.1182/blood-2007-11-120998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Wainwright DA, Sengupta S, Han Y, Ulasov IV, Lesniak MS. The presence of IL-17A and T helper 17 cells in experimental mouse brain tumors and human gliom. PLoS ONE. 2010;5(10) doi: 10.1371/journal.pone.0015390. Article ID e15390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Rodrigues JC, Gonzalez GC, Zhang L, et al. Normal human monocytes exposed to glioma cells acquire myeloid-derived suppressor cell-like properties. Neuro-Oncology. 2010;12(4):351–365. doi: 10.1093/neuonc/nop023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Raychaudhuri B, Rayman P, Ireland J, et al. Myeloid-derived suppressor cell accumulation and function in patients with newly diagnosed glioblastoma. Neuro-Oncology. 2011;13(6):591–599. doi: 10.1093/neuonc/nor042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Ghebeh H, Bakr MM, Dermime S. Cancer stem cell immunotherapy: the right bullet for the right target. Hematology/Oncology and Stem Cell Therapy. 2008;1(1):1–2. doi: 10.1016/s1658-3876(08)50053-7. [DOI] [PubMed] [Google Scholar]
- 143.Bao S, Wu Q, McLendon RE, et al. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature. 2006;444(7120):756–760. doi: 10.1038/nature05236. [DOI] [PubMed] [Google Scholar]
- 144.Liu G, Yuan X, Zeng Z, et al. Analysis of gene expression and chemoresistance of CD133+ cancer stem cells in glioblastoma. Molecular Cancer. 2006;5, article 67 doi: 10.1186/1476-4598-5-67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Singh SK, Clarke ID, Terasaki M, et al. Identification of a cancer stem cell in human brain tumors. Cancer Research. 2003;63(18):5821–5828. [PubMed] [Google Scholar]
- 146.Prestegarden L, Svendsen A, Wang J, et al. Glioma cell populations grouped by different cell type markers drive brain tumor growth. Cancer Research. 2010;70(11):4274–4279. doi: 10.1158/0008-5472.CAN-09-3904. [DOI] [PubMed] [Google Scholar]
- 147.Pellegatta S, Poliani PL, Corno D, et al. Neurospheres enriched in cancer stem-like cells are highly effective in eliciting a dendritic cell-mediated immune response against malignant gliomas. Cancer Research. 2006;66(21):10247–10252. doi: 10.1158/0008-5472.CAN-06-2048. [DOI] [PubMed] [Google Scholar]
- 148.Wang R, Chadalavada K, Wilshire J, et al. Glioblastoma stem-like cells give rise to tumour endothelium. Nature. 2010;468(7325):829–835. doi: 10.1038/nature09624. [DOI] [PubMed] [Google Scholar]
- 149.Ricci-Vitiani L, Pallini R, Biffoni M, et al. Tumour vascularization via endothelial differentiation of glioblastoma stem-like cells. Nature. 2010;468(7325):824–830. doi: 10.1038/nature09557. [DOI] [PubMed] [Google Scholar]
- 150.Lake RA, Robinson BWS. Immunotherapy and chemotherapy—apractical partnership. Nature Reviews Cancer. 2005;5(5):397–405. doi: 10.1038/nrc1613. [DOI] [PubMed] [Google Scholar]
- 151.Bourquin C, Schreiber S, Beck S, Hartmann G, Endres S. Immunotherapy with dendritic cells and CpG oligonucleotides can be combined with chemotherapy without loss of efficacy in a mouse model of colon cancer. International Journal of Cancer. 2006;118(11):2790–2795. doi: 10.1002/ijc.21681. [DOI] [PubMed] [Google Scholar]
- 152.Kim CH, Woo SJ, Park JS, et al. Enhanced antitumour immunity by combined use of temozolomide and TAT-survivin pulsed dendritic cells in a murine glioma. Immunology. 2007;122(4):615–622. doi: 10.1111/j.1365-2567.2007.02680.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Xu X, Stockhammer F, Schmitt A, et al. Therapeutical doses of temozolomide do not impair the function of dendritic cells and CD8+ T cells. International Journal of Oncology. 40(3):764–772. doi: 10.3892/ijo.2011.1269. [DOI] [PubMed] [Google Scholar]