

Abstract
Propionibacterium acnes induction of IL-1 cytokines through the NLRP3 inflammasome was recently highlighted as a dominant etiological factor for acne vulgaris. Therefore, therapeutics targeting both the stimulus and the cascade would be ideal. Nitric oxide (NO), a potent biological messenger, has documented broad-spectrum antimicrobial and immunomodulatory properties. To harness these characteristics to target acne, we utilized an established nanotechnology capable of generating/releasing nitric oxide over time (NO-np). P. acnes was found to be highly sensitive to all concentrations of NO-np tested, though human keratinocyte, monocyte, and embryonic zebra fish assays revealed no cytotoxicity. NO-np significantly suppressed IL-1β, TNF-α, IL-8 and IL-6 from human monocytes and IL-8 and IL-6 from human keratinocytes respectively. Importantly, silencing of NLRP3 expression by small interfering RNA did not limit NO-np inhibition of IL-1 β secretion from monocytes, and neither TNF-α, nor IL-6 secretion nor inhibition by NO-np was found to be dependent on this pathway. The observed mechanism by which NO-np impacts IL-1β secretion was through inhibition of caspase-1 and IL-1β gene expression. Together, these data suggest that NO-np can effectively prevent P. acnes induced inflammation by both clearing the organism and inhibiting microbial stimulation of the innate immune response.
Introduction
Acne's multifactorial etiology, resulting from a mix of hormone-induced elevations in sebum production, abnormal follicular epithelial desquamation and proliferation, hypercolonization of Propionibacterium acnes and host inflammatory reactions, make treatment often times challenging (Castro and Ferreira, 2008; Zouboulis et al., 2005). This challenge has fueled the development of innovative delivery platforms that improve delivery and efficacy of established and investigative agents. Various particulate delivery platforms have been evaluated for use in acne, including liposomes and nanoparticles. These carriers have the advantage of gradual release (Manconi et al., 2002), minimizing irritancy (Manconi et al., 2002), follicular targeting/enhanced follicular penetration (Jung et al., 2006), and cutaneous retention of the active ingredient (Manconi et al., 2006). More importantly, the unique properties conferred by the vehicles existing at the nanoscale are allowing for the investigation of new therapeutics which in their bulk form can not be delivered topically due to stability and solubility issues, including lauric acid (Nakatsuji et al., 2009) and chitosan (Friedman et al., 2013).
Along this vein, nanoparticle platforms can be a vehicle through which unstable therapeutically relevant agents are harnessed. Nitric oxide (NO) represents one such molecule - a diatomic, lipophilic gaseous molecule with functions described ranging from vascular modulation to cell cycle regulation, pro- and anti-inflammatory properties, and microbicidal/static activity (Friedman and Friedman, 2009). The simplicity of NO mixed with its biological potency and complexity makes it an extremely promising pharmacological agent, but utilization has been limited due to the lack of effective and safe delivery systems. We previously reported on a hybrid hydrogel-sugar based nanoparticulate system (np) capable of generating physiologic concentrations of NO over time. Upon exposure to an aqueous environment, the hydrogen bonding network that comprises the nanoparticle matrix loosens, allowing release of the NO. (Friedman et al., 2008b; Han et al., 2011). This platform is unique in that nanoparticle matrix facilitates the generation of nitric oxide from encapsulated sodium nitrite through a dinitrogen trioxide intermediate, as the rich disulfide bonding network allows for long range proton and electron transfer/movement. As opposed to organic nitrates, the most commonly used NO-donor in clinical practice, there is no potential for decreased efficacy with prolonged and continuous use, what is known as ‘nitrite tolerance,’ which results from the prerequisite host reduction of the administered nitrate to active nitric oxide (Bennett et al., 1994; Gori and Parker, 2002).
We have previously shown that NO-np significantly increase the healing of MRSA infected wounds in a murine excisional injury model (Martinez et al., 2009). NO-np also accelerated wound healing in non-obese, diabetic, severe combined immunodeficiency mice, with significantly decreased neutrophil and increased macrophage infiltration noted in the NO-np treated wound beds as compared to another topical NO-donating platform, diethylenetriamine (Blecher et al., 2012). Given the antimicrobial and immunological properties of NO, we investigated whether NO-np has antimicrobial activity against Propionibacterium acnes, and modifies the inflammatory cascade associated with the evolution of acne lesions.
Results
Characterization of nanoparticles, detection of NO release, and tissue penetration
Nitric oxide nanoparticles were formed using a previously described protocol (Friedman et al., 2008). In brief, the highly anhydrous interior of the nanoparticles facilitates the formation of N2O3 from nitrite and protons. N2O3 is both the source of NO and is a potent S-nitrosating agent that likely contributes to the biological efficacy of NO-np.
Using scanning electron microscopy (SEM), NO-nanoparticle aggregates were found to average approximately 127 nm in diameter (Figure 1a). We have previously shown by transmission EM that individual particles measure ∼10 nm in diameter (Friedman et al., 2008). Using Dynamic light scattering (DLS), NO-np revealed an average hydrodynamic diameter of 216.9 nm based on 40 acquisition attempts (Figure 1b). The standard deviation was 14.2 nm (3.3%), proving that NO-np are homogenous in size. Since NO-np swell with moisture, the average diameter is likely an overestimate as compared to the SEM. NO release from the NO-np has been previously reported using both amperometric detection and ozone chemiluminescence (Cabrales et al., 2010; Friedman et al., 2008).
Figure 1. Nanoparticles Characterization.
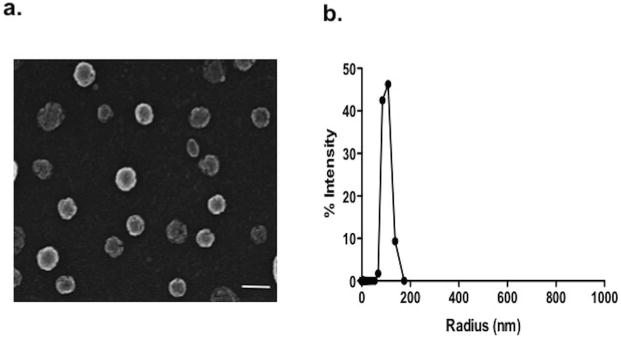
(a) Scanning electron microscopy revealed distinct spherical nanoparticles with irregular surface structure indicative of the porous matrix lattice (scale bar = 100 nm). (b) Using Dynamic light scattering (DLS), NO-np revealed an average hydrodynamic radius of 108.47 nm based on 40 acquisition attempts. The standard deviation was 14.2 nm (3.3%), proving that NO-np are homogenous in size.
Antimicrobial activity of NO-np
NO generated from the NO-np has demonstrated antimicrobial activity against various Gram positive and negative pathogens, including Staphylococcus aureus (Martinez et al., 2009) and Pseudomonas aeruginosa (Friedman et al., 2011), and therefore we sought to evaluate the impact on P. acnes.
P. acnes strains were incubated with various concentrations of NO-np and np (0.625, 1.25, 2.5, or 5 mg/mL) for 24 h (Figure 2a). For all isolates tested, all concentrations of NO-np significantly inhibited bacterial growth compared to controls in a dose-dependent manner for up to 24 h. Concentrations over 2.5 mg/ml completely inhibited growth over 24 hours while 1.25 and 2.5 mg/ml significantly inhibited growth after 24 hours of co-incubation respectively (p<0.0001) as compared to all controls as well as 0.625 mg/ml NO-np. Statistical significance was not met when comparing the two treatment groups (p=0.07). To demonstrate the bactericidal activity against P. acnes, bacteria was incubated with varying doses of NO-np, as well as np control alone for 4 hours. Subsequently, the bacteria were plated and the viability was determined by colony-forming unit (CFU) assay. The NO-np were effective at significantly killing P. acnes, resulting in only 8.9% and 4.6% survival for the two concentrations illustrated (Fig. 2b). Interestingly, the 2.5 and 5 mg/ml np control had a modest, albeit significant effect, compared to untreated cells, likely owing to the physical presence of the nanoparticles that probably imparted a steric effect, interfering with cell-cell interactions, noted in previous investigations (Friedman et al., 2011).
Figure 2. Antimicrobial effects of Nitric Oxide releasing nanoparticles.
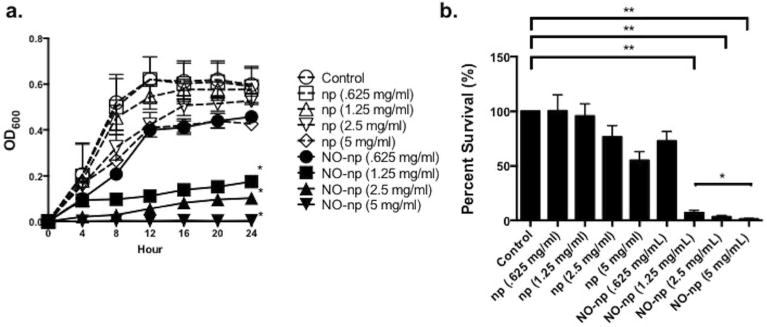
(a) Susceptibility of P. acnes isolates to varying concentrations of empty nanoparticles (np) and NO-np (0.625, 1.25, 2.5 and 5 mg/ml), was investigated by real-time Bioscreen analysis and (b) colony form unit assay determination (mean CFU per ml). Experiments were repeated in triplicate and performed at least thrice on separate days. Asterisks denote p value significance calculated by unpaired two-tailed t test analysis. (*, p < 0.05; **, p < 0.01).
NO-np inhibit P. acnes stimulated inflammatory cytokines
Given the importance of the inflammatory response in the pathophysiology of acne, we evaluated NO's ability to inhibit inflammatory cytokine production from a P. acnes challenged human keratinocyte cell line, HaCaT, and peripheral blood mononuclear cells (PBMCs). Previous studies showed that P. acnes induces the inflammatory cytokines IL-1, IL-8 and IL-12 in human monocytes and IL-6 in human keratinocytes (Friedman et al., 2013; Qin et al., 2014). At the same concentrations tested in the antimicrobial assays, NO-np significantly reduced P. acnes stimulated human monocyte (PBMC) expression of IL-1 β (52.0% reduction), TNF-α (91.3% reduction) at the highest dose tested (Fig 3a-b). Control nanoparticle (np) had no inhibitory effect on IL-1β secretion at all concentrations tested (S2). NO-np significantly inhibited P. acnes stimulated PMBC and HaCaT IL-6 and IL-8 secretion in a dose dependent manner as compared to controls (Fig 3c-f). For IL-6 and IL-8 secretion respectively, a statistically significant difference was noted when comparing control to 2.5 mg/ml (33.4% and 54.84% reduction) and 5mg/ml (54.8% and 88.47% reduction) for the PBMC group and to all treatment groups with the HaCaT cells (15.1%, 51.5%, and 65.0% and 48.31%, 90.76%, and 98.93% respectively).
Figure 3. NO-np significantly inhibits P. acnes induced pro-inflammatory cytokines in PMBC and keratinocytes.
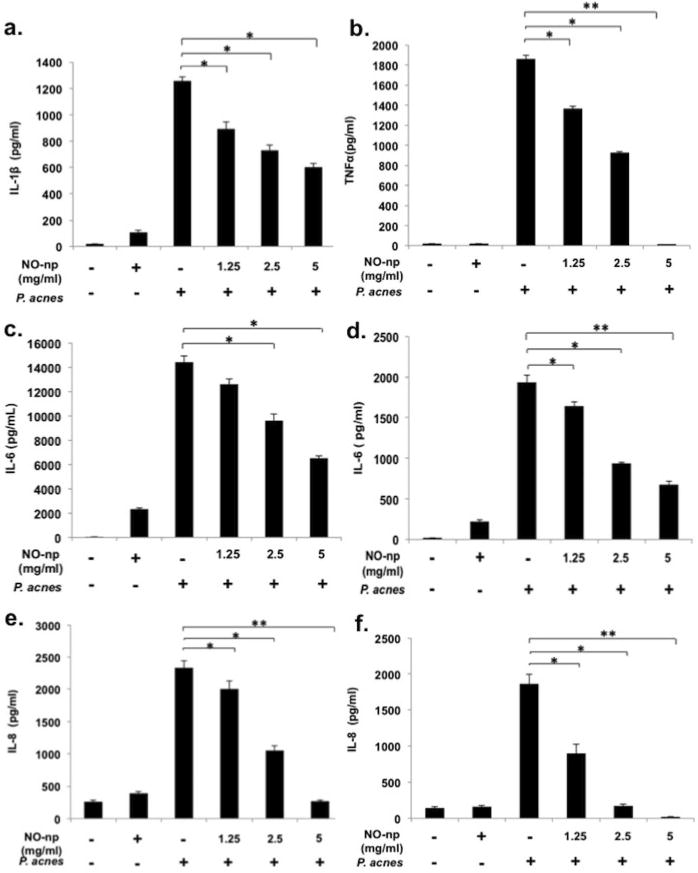
P. acnes stimulated PBMCs expression of IL-1β (a), TNF-α (b), IL-6 (c), and IL-8 (e), and HaCaT cell expression of IL-6 (d) and IL-8 (f) with and without various concentrations of NO-np. NO-np at 2.5 mg/ml was used as a control. IL-1β and TNF-α were undetectable in the HaCaT supernatant via ELISA (data not shown)). The data is presented as the mean of triplicate wells ± SD and is representative of three individual experiments. (*, p < 0.05; **, p < 0.01).
With the recent attention paid to IL-1β as the initial subclinical stimulus for follicular hyperkeratinization, its place in and importance of the NLRP3 inflammasome with respect to P. acnes induction of the host defense (Qin et al., 2014), we sought to determine if the aforementioned anti-inflammatory effects resulted from NO-np's impact on this multi-protein complex. To evaluate whether NO-np reduction of IL-1β, IL-6, and TNF-α secretion was mediated specifically through NO activity on NLRP1 and/or NLRP3, small interfering RNA (siRNA) were utilized silence specific gene expression as previously described (Qin et al., 2014). Transfection of monocytes with specific siRNA targeting NLRP3 resulted in a significant reduction of IL-1 β expression (93.8%, p<0.01) (Fig 4a), as compared to stimulated controls, respectively. A nonspecific control siRNA had no effect in all cases. While siNLRP3 transfection of PBMCs resulted in a 93.8% reduction in IL-1β secretion, NO-np treatment provided further, statistically significant reduction (>99%) as compared to stimulated siNLRP3 control. In congruence with past results, siNLRP1 transfection of PBMCs resulted in minimal reduction in P. acnes induced IL-1 β secretion, while a 55.27% reduction in IL-1 β expression following NO-np treatment compared to untreated was noted. Interestingly, knockout of NLRP3 did not influence P. acnes stimulated secretion of IL-6, IL-8 nor TNF-α from PMBCs, and NO-np treatment yielded similar reductions in cytokine secretion as noted above (S1a-c). However, knockdown of NLRP1 had a significant effect in NO-np inhibition of IL-6 secretion from PBMCs as compared to NLRP3 knockdown.
Figure 4. NO-np inhibits PBMC IL-1β secretion by directly down-regulating IL-1β gene and Caspase-1 gene expression via the inflammasome pathway.
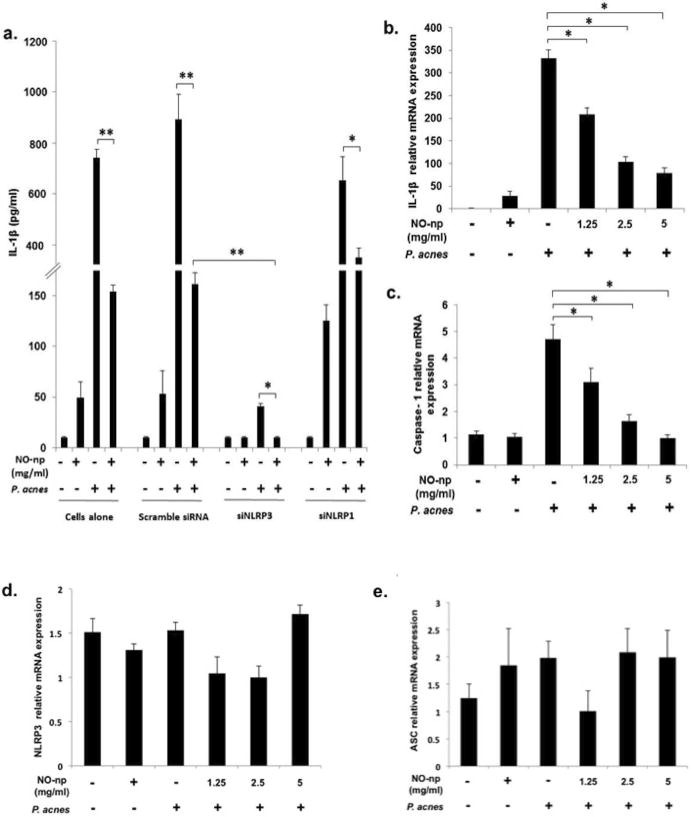
Cells were infected by P. acnes at MOI 0.5 with and without NO-np and with scramble siRNA, siNLRP3 or siNLRP1, and IL-1β secretion in culture supernatants was tabulated via ELISA (NO-np 2.5 mg/ml) (a). The expressions of Caspase-1, IL-1β, NLRP3, and ASC were measured by real-time PCR (b-e). Data are shown as the mean of triplicate wells ± SD and is representative of three individual experiments. (*, p < 0.05; **, p < 0.01).
NO-np inhibit P. acnes induced Caspase-1 and IL-1 β, but not NLRP3, gene expression
Given the additional reduction of IL-1 β secretion from siNLRP3 transfected PBMC beyond the effects of knockdown alone, we sought to evaluate NO-np activity on gene expression of key inflammasome components. IL-1β and caspase-1 mRNA levels were significantly reduced in PBMCs exposed to P. acnes and treated with NO-np as compared to controls (87.1% and 80.0% reduction, respectively; Fig 4a-b), with an associated decrease in caspase-1 production and activity (Fig 5a-b). NO-np treatment had minimal effect on both NLRP3 and ASC mRNA expression as compared to controls (Fig 4d-e). NO-np alone did cause a slight but significant increase in IL-1β mRNA expression.
Figure 5. NO-np down-regulates P. acnes induced Caspase-1 protein expression and activity in PBMC cells.
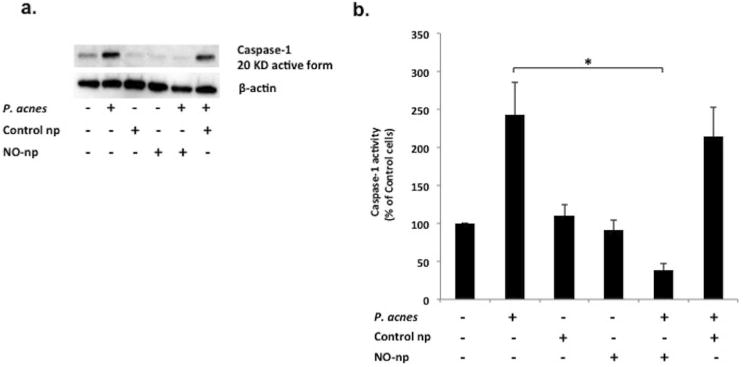
Peripheral blood mononuclear cells (PBMC) were isolated from whole blood of normal healthy donors. Transfected cells were plated into 6–well cell culture plates, cultured at 37°C overnight, and co-cultured with or without 2.5 mg/mL control-np and 2.5 mg/ml NO-nanoparticle for an hour. Cells were then infected by P. acnes at MOI 0.5 for 24 hours. Cells were lysed, and caspase-1 protein expression was measured with western blot (a) and activity assay (b). a) Western blot using anti-caspase-1: Cells alone (Lane 1); P. acnes (Lane 2); Control np (Lane 3); NO-np (Lane 4); P. acnes in the presence of NO-np (lane 5); P. acnes in the presence of Control np (lane 6). b) Caspase-1 activity assay: cells were normalized as percentage of control cells. NO-np significantly decreases PBMC caspase-1 protein activity in the presence of P. acnes compared to control-np. (*, p < 0.05)
NO-np are non-toxic
All concentrations of NO-np tested had minimal toxicity to keratinocytes and PBMCs using MTT assay (p=0.31 and 0.13, respectively, as compared to control) (Fig 6a-b), which was further substantiated using embryonic zebra fish assays (Fig 6c). The embryonic zebrafish assay was used to assess the toxicological impact of NO-np in an in vivo vertebrate model system. Embryos exposed to NO-nps did not exhibit differences in rates of malformations or mortality at any concentration tested compared to controls. Any differences in morphology, mortality, development, larval morphology, or behavioral endpoints between treatment and control were attributed to laboratory background. Therefore, our data suggest that NO-nps can inhibit P. acnes-induced cytokine production in human keratinocytes and PBMCs, and this is not a consequence of the release of cytokines at cell death.
Figure 6. Nitric Oxide releasing Nanoparticles are non-toxic.
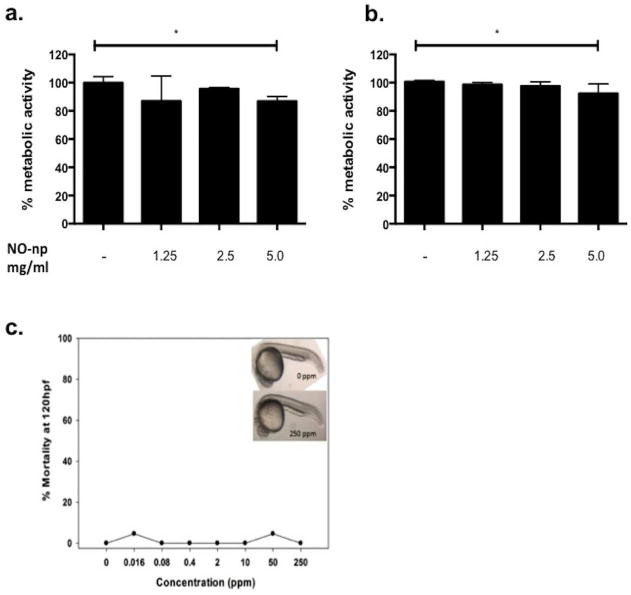
NO-np at various concentrations (1.25, 2.5 and 5 mg/ml, respectively) were incubated with HaCaT cells (a) or PBMCs (b). Cells were collected and evaluated for viability using MTT assay. (c) Representative images of zebrafish embryos at 120 hpf: control (top) and exposed to NO-np (bottom). No significant differences were observed in larval morphology or behavioral endpoints (p > 0.05 for each endpoint evaluated, Fisher's Exact test). Error bars denote SEM.
Discussion
In this work, we demonstrated that the NO-nps effectively decreased P. acnes viability and inhibited IL-1β, TNF-α, IL-8 and IL-6 secretion from P. acnes stimulated PMBCs and IL-8 and IL-6 from keratinocytes, while neither having in vitro nor in vivo cytotoxicity. While NO-np was found to impact components of the NLRP3 inflammasome pathway, its anti-inflammatory effects were not specific to NLRP3, and NLRP3 knockout did not impair NO-np's anti-inflammatory properties.
Nearly every member of the skin cell population expresses nitric oxide synthase (NOS) and is thereby able to produce NO to accomplish key physiologic processes. NO's role in skin host defense is concentration-dependent, and is imparted in a bimodal fashion. At low concentrations it stimulates the immune system by enhancing immune cell activity, cytokine production, expression of adhesion factors, and extracellular matrix constituent synthesis. High-output of endogenous NO is achieved through the stimulation of pathogen recognition receptors that promote iNOS transcription and subsequent L-arginine dependent NO release (Griffith and Stuehr, 1995). This calcium-independent enzymatic pathway along with nonenzymatic reduction of naturally occurring nitrosothiols (RSNO) and nitrate (NO3-) are the two key methods of endogenous NO synthesis pertinent to microbial eradication (Weller, 2009). The coupling of NO and O2- produces potentially toxic reactive nitrogen species (RNS), such as the peroxynitrite anion (ONOO-), and induces increased O2- production (Fang, 2011). RNS's can interfere with normal protein interactions as protein thiols become nitrosated (Han et al., 2011). Collectively, these molecules adversely affect microbial viability by traversing cellular membranes, interrupting DNA replication, instigating lipid peroxidation, and inactivating metalloproteinases essential for microbial metabolism (Wink and Mitchell, 1998).
P. acnes, which resides in pilosebaceous follicles in both acne and non-acne subjects, elicits an inflammatory responses essential for the pathogenesis and clinical phenotype of acne vulgaris (Bellew et al., 2011). P. acnes was found to be highly sensitive to NO derived from the NO-np. Perhaps the most significant aspect to be considered of NO's microbicidal activity is the lack of demonstrated resistance to date. This is thought to be a direct result of the aforementioned multiple mechanisms of NO's action - to overcome would require the simultaneous development of distinct resistance mutations. Some minor compensatory mechanisms against the actions of NO have been elucidated (Richardson et al., 2008); however, no protection against the high concentrations of NO released by donating materials is evident. Isolates of S. aureus, Staphylococcus epidermidis, Escherichia coli O157:H7, and P. aeruginosa that survived exposure to high NO concentrations showed no increase in minimum inhibitory concentration levels afterward (Privett et al., 2012). Given the ongoing concern regarding antibiotic resistance in P. acnes, NO-np represents a much needed multi-mechanistic antimicrobial approach (Eady, 1998; Ross et al., 2003).
While P. acnes is implicated in the pathophysiology of Acne Vulgaris, it serves more as a pro-inflammatory stimulus rather then a pathogenic organism, and therefore muting this response is central to clinical treatment. We found that NO-np inhibited IL-8 and IL-6 production from P. acnes stimulated PBMCs and keratinocytes in a dose dependent manner. IL-8 is a CXC chemokine that serves as a potent chemoattractant for neutrophils, a predominant cell type in acne-related lesions. IL-8 gene expression has been shown to be significantly elevated in biopsy specimens of acne lesions (Trivedi et al, 2006). IL-6 is pleiotropic cytokine playing an important role in acute and chronic inflammation. IL-6 has also been shown to contribute to persistent neutrophil trafficking in wounds, resulting in delayed wound healing in the setting of sustained stress (Kim et al, 2014). Interestingly, we found that NLRP1 may have a role in NO-np down-regulation of P. acnes induced IL-6 expression, as compared to NLRP3, warranting further investigation.
NO-np had a profound effect the secretion of TNF-α from PBMCs. TNF-α is central to stimulating the acute phase reaction of the immune response. Implicated in multiple dermatologic diseases including acne, several studies have highlighted that single nucleotide polymorphisms (SNPs) of the TNF-α gene (such as the -308 G/A polymorphism) are associated with an increased risk to develop chronic inflammatory disease (Yang et al., 2014). NO-np significantly inhibited PBMC induction of TNF-α triggered by P. acnes, almost completely at the highest concentration tested (5mg/ml).
NO is known to modulate several innate immune pathways, enhancing the host's own immune response; for example, NO is involved in lymphocyte activation and proliferation and numerous cytokine pathways, including TNF-α, TGF-β, p56, IL-6, and NF-κB (Fang, 1997). However, while an important recruiter of host defense against invading pathogens, NO's immunoregulatory function is clearly multi-faceted - NO can function as an anti-inflammatory agent when appropriate, as demonstrated in this study. For example, investigators have shown that NO inhibits key members of the inflammatory cascade such as NF-κB and STAT-1 in keratinocytes (Giustizieri et al., 2002), and down-regulates neutrophil aggregation, secretion and diapedesis (Dal Secco et al., 2003). We too demonstrated that NO-np inhibits secretion of various pro-inflammatory cytokines from different cell types stimulated with P. acnes.
In light of the recent data highlighting the role of caspase dependent IL-1 β generation through the NLRP3 inflammasome pathway in the pathophysiology of acne vulgaris, (Qin et al., 2014), we showed that NO-np inhibits P. acnes stimulated PBMC IL-1β and caspase-1 gene expression, likely explaining the additional reduction in IL-1β secretion from P. acnes stimulated siNLRP3 transfected monocytes treated with NO-np. The NLRP3 inflammasome is a multi-protein complex that triggers the maturation of the pro-inflammatory cytokines IL-1β and IL-18 (Hernandez-Cuellar et al., 2012). This complex can be activated by pathogen-associated molecular patterns (PAMPs) or endogenous danger-associated molecular patterns (DAMPs) generated in the setting of cellular injury or tissue damage (Dostert et al., 2008; Qin et al., 2014; Schroder and Tschopp, 2010). Interestingly, one of NO's roles may be as a negative regulator of inflammasome activity. It was previously shown that inflammasome activation was inversely correlated to iNOS expression, and that NO inhibited IL-1β production and caspase-1 activation (Mao et al., 2013). The mechanism by which NO is believed to inhibit the various elements of the inflammasome is via S-nitrosylation several proteins involved in cellular regulation, ultimately impacting their activity and function (Stamler et al., 2001). We demonstrated that NO-np did not reduce gene expression of NLRP3, however this finding does not provide insight into NLRP3 activity or degree of nitrosylation. We have previously shown that the NO-np are effective trans-nitrosating agents (Friedman et al., 2011), likely owing to the nanoparticles' forming a dinitrogen trioxide intermediate, capable of S-nitrosation under aerobic and anaerobic conditions, which therefore offers a mechanism for the observed activity.
Regardless of NLRP3 function, we demonstrated NLRP3 independent anti-inflammatory effects in a NLRP3 knockdown model, likely owing but not limited to NO-np's direct effect on caspase-1 and IL-1 β gene expression. This finding has far reaching implications in many disease states given the broad role of both in all inflammasome pathways. Interestingly, knockdown of NLRP3 did not impact NO-np suppression of P. acnes stimulated monocyte derived IL-6, IL-8, and TNF-α, once again supporting NO's multi-mechanistic anti-inflammatory activity and highlighting NO's target diversity.
Not surprisingly, we found that NO-np alone mildly induces PBMC IL-1β expression in the absence of pathogen stimulus, highlighting the duality of NO's functions in immune regulation. We and other investigators have shown that at low concentrations, NO stimulates PMBC generation of IL-6, but as NO levels increased using NO-donor molecules, both IL-6 expression and NF-κB activity decreased. A greater degree of nitrosative stress increases the likelihood of protein S-nitrosylation, in this case, of key NF-κB related proteins including IκB kinase b and p50, which was shown to directly inhibit pro-inflammatory activity (Marshall et al., 2004). This further supports any NO releasing technology that facilitates trans-nitrosylation. Therefore, the current theory is that in the early stage of infection, lower, initial concentrations of NO may accelerate the immune response by stimulating the synthesis of proinflammatory cytokines, while at a later stage, when NO concentrations have peaked and cells are activated, it rather counteracts the undesired development of inflammation.
Together, the presented and past data demonstrating that NO-np have multi-mechanistic antimicrobial and anti-inflammatory properties, suggest that NO-np can potentially target multiple pathophysiologic elements of Acne Vulgaris. Importantly, in the face of rising of antimicrobial resistance in the setting of prolonged antibiotic use, NO's bactericidal and static properties limit the risk of the emergence of resistant species. Therefore, our findings suggest that the NO-np offers a new approach to harness the therapeutic potential of nitric oxide, and that this platform should be translated to the bedside.
Materials and Methods
Production of NO-np
The production of the NO-np has been previously reported (Friedman et al., 2008). Briefly, the NO-np are prepared using the following sequence of steps: 1) Hydrolyzing Tetramethylorthosilicate (TMOS): Stock of 5 ml of TMOS, 600 μl of deioinized water, and 560 μl of 2 mM hydrochloric acid are added to a small vial. The contents of the vial are sonicated for approximately 20-30 minutes yielding a clear solution that is then placed on ice. 2) Mixing the sol-gel components: 1.49 g of sodium nitrite are dissolved in 4 ml of PBS buffer at pH 7.5 followed by sequential addition and mixing of 0.5 ml of PEG-200, 500μl of chitosan (1mg/ml). The resulting mixture is then vortexed thoroughly. Then, 2 ml of previously hydrolyzed TMOS is added to the solution followed by vigorous vortexing until complete gelation. 3) Lyophilizing the sol-gel: The resulting gelled material is then lyophilized for 24-48 hrs that removes all volatile components. 4) Ball-Milling the lyophilized sol-gel: Following lyophilization the dry material is ball milled at 150 rpm for 8 hours resulting in a very fine white powder.
Nanoparticle Imaging
Dynamic light scattering (DLS)
Nanoparticle skin penetration
Retired breeder male Sprague-Dawley rats (650 g, Charles River Breeding Laboratories (Wilmington, MA, USA)) were used in these studies. All animal protocols were approved by the Animal Use Committee at the Albert Einstein College of Medicine. Rats (n=3) were first anesthetized via intraperitoneal injection of sodium pentobarbital (35 mg/kg; Abbott Laboratory, Chicago, IL, USA). Four aliquots of 50 ul of Alexafluor-594nm labeled NO-np (5mg/ml) suspended in coconut oil were applied to the 4 abdominal quadrants of the abdomen. Coconut oil alone was used as a control (n=1). After ten minutes, quadrants were cleaned with alcohol wipes and 4-mm punch biopsies were taken from each application site. Specimens were subjected to frozen sectioning and vertical sections (4 mm thick) were fixed to glass slides. Images were captured with a confocal microscope (PerkinElmer) with a 10×, 1.4 NA objective and a digital camera (Orca ER; Hamamatsu).
CFU assay and Bioscreen Analysis
P. acnes strain ATCC 6919 was obtained from American Type Culture Collections and isolates grown in the miniMACS anaerobic workstation (Don Whitley Scientific, West Yorkshire, UK). Briefly, Brucella agar plates supplemented with blood, hemin, and vitamin K (Remel Fisher Scientific, Lenexa, KS) were streaked with P. acnes overnight and single colonies isolated. The cultures were grown in an anaerobic chamber for 4–5 days. A spectrophotometer OD600 was used to determine the bacterial log phase.
Various concentrations of control and NO-nps (0.625, 1.25, 2.5, 5 mg/ml) were incubated with 3.75 ×105 bacteria in a final volume of 30 ml at 37°C for 4 hours. After incubation, 10-fold dilutions were prepared and incubated for 4 days at 37°C under anaerobic conditions. Individual colonies were counted and the number of CFU was tabulated.
For growth kinetics, reinforced Clostridium medium was inoculated with a fresh colony grown on the Brucella Agar plates and suspended in 1 mL of medium. A suspension of 100 μL of bacteria was transferred to a 200-well plate with 100 μL of reinforced Clostridium medium per well containing NO-np or np (0.625, 1.25, 2.5, 5, mg/mL). Bacteria and nanoparticles were incubated for 24 h at 37°C. Controls included wells containing bacteria with reinforced Clostridium medium alone. Growth was assessed at OD 600 nm every 30 min using a microplate reader (Bioscreen C, Growth Curves USA, Piscataway, NJ).
P. acnes culture and cell stimulation
PBMCs were stimulated by P. acnes strain 6919 at MOI 0.5, while HaCaT keratinocytes were infected by P. acnes at MOI 20 (Qin et al., 2014). We used live P. acnes as opposed to sonicated-killed preparations of P. acnes due to the pathophysiological relevance of our model and consistency on the observed responses.
Isolation of Peripheral blood mononuclear cells, HaCaT cell culture and cytokine ELISA
PBMCs were isolated from whole blood of normal healthy donors as approved by the Institutional Review Board at UCLA using Ficoll-Paque gradients (GE Healthcare, Piscataway, NJ) and plated onto 24-well tissue culture plates (5 × 106 per well) in RPMI 1640 media containing 10% FBS (HyClone, South Logan, UT). Cells were co-cultured one hour with 0 mg/ml, 1.25 mg/ml, 2.5 mg/ml and 5 mg/ml of NO-nanoparticles respectively. Then cells were infected by P. acnes at MOI 0.5 for 24 hours at 37°C.
HaCaT cells, an immortalized human keratinocyte cell line, plated into 6-well tissue culture plates (5 × 105 per well) and cultured at 37°C with 5% CO2 in Dulbecco's Modified Eagle Medium (DMEM) (Gibco11995-060, Invitrogen Corp., USA) supplemented with 10% heat inactivated Fetal Bovine Serum (HyClone, South Logan, UT) and 1% Penicillin/Streptomycin (Gibco 15140-122, Invitrogen Corp., USA) for 18 hours. HaCat cells retain their ability to differentiate, to express Toll-like receptors involved in pathogen recognition and to produce a number of cytokines in response to bacterial stimulation in a similar manner to Primary human keratinocytes (Grange PA et al, 2009). HaCaT were treated one hour with the different doses of NO-nanoparticles as above. Then cells were infected by P. acnes at MOI20 for 24 hours at 37°C.
IL-6, IL-8, IL-1β and TNFα levels in culture supernatants were measured by ELISA following the manufacturer's recommendations (R&D ELISA Development System, Minneapolis, MN). Samples were assayed in triplicates. Results are expressed as mean±SD of at least three independent experiments, with PBMCs obtained from three independent donors.
RNA isolation, cDNA synthesis, and real-time PCR
(See Supplemental)
siRNA transfection
siRNA transfection into PBMCs was accomplished by using the Amaxa Nucleofector system and the Nucleofector Kit according to the manufacturer's recommendations with program U-001 for high viability. siRNA constructs (scramble, siNLRP3 and siNLRP1) were used at 1 μg per transfection. Transfection efficiency of expression constructs was assessed using the pMAX-GFP construct, and it yielded an average of 70% transfection rate (Lonza, Allendale, NJ). Transfected cells were plated into 6-well cell culture plates and cultured at 37 °C for overnight. Transfected cells were co-cultured with NO-nanoparticle 2.5 mg/ml for an hour, and then infected by P. acnes at MOI 0.5.
Western Blot analysis
Detection of caspase-1 was performed by western blot analysis. Isolated peripheral blood mononuclear cells (PBMC) were treated with 2.5 mg/mL NO-np or control np for an hour. Cells were then infected by P. acnes with MOI 0.5. After 24 hours, cells were harvested and lysed in 100 μL of lysis buffer (M-PER Mammalian Protein Extraction Reagents, Thermo Scientific) with protease inhibitors. Protein levels were normalized with the Bradford assay. Protein profiles were separated by electrophoresis in BIO-RAD mini PROTEAN TGX polyacrylamide precast gels and transferred onto 0.45 μm Millipore Immobilon P-transfer membranes, and caspase-1 was detected by caspase-1 monoclonal antibody (D7F10, Cell Signaling). The bands from western blot were visualized by using VersaDOC MP5000 (BIO-RAD).
Activity Assay
Isolated peripheral blood mononuclear cells (PBMC) were treated with 2.5 mg/mL NO-np or control np for an hour. Cells were then infected by P. acnes with MOI 0.5. After 24 hours, cells were harvested and lysed with 100 μL of cold lysis buffer. Caspase-1 protein activity was then measured using Caspase-1 Colorimetric Assay (R&D systems) per manufacturer's instructions and the activity was determined via percentage of the control (cells alone). After 16 hours of incubation at 37 °C, activity was read using BioTek Synergy 2 plate reader using a wavelength of 405 nm.
MTT toxicity assay
The MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrasodium bro-mide tetrazolium; Sigma Aldrich) was performed as previously described (Friedman et al., 2013) (see supplemental for complete methods).
Statistical analysis
GraphPad Prism 5.0 (GraphPad Software, La Jolla, CA) was used for statistical analyses. p val- ues were determined by analysis of variance and adjusted by use of the Bonferroni correction. p values of <0.05 were considered significant
In vivo toxicity assay
Zebrafish embryos (Danio rerio, wild type, 5D-Tropical strain) were obtained from Sinnhuber Aquatic Research Laboratory, Oregon State University, and exposures and evaluations were conducted according to Truong et al (Truong et al., 2011). Exposures were conducted over 5 days of development. All organ systems begin functioning during this time period and all of the molecular signaling pathways are active and necessary for normal development to occur. At 120 hpf, behavioral endpoints (motility, tactile response) and larval morphology (body axis, eye, snout, jaw, otic vesicle, notochord, heart, brain, somite, fin, yolk sac, trunk, circulation, pigment, swim bladder) were evaluated in vivo and scored in a binary fashion (present or absent).. Untreated control and exposed groups were compared using Fisher's exact test for each endpoint, and p-value < 0.05 for significance.
Supplementary Material
Supplementary Figure S1: NLRP1 may have a role in NO-np down-regulation of P. acnes induced IL-6 expression in PBMCs, Data shows that there are different levels of P. acnes induced IL-6 down regulation as affected by NLRP1 and NLRP3 (a). Knockout NLRP1 displays a more significant down-regulation of IL-6 than knockout NLRP3. In contrast, neither NLRP1 nor NLRP3 affect P. acnes-induced levels of TNF-α and IL-8 in PBMCs (b). Data are shown as the mean of triplicate wells ± SD and is representative of three individual experiments.
Supplementary Figure S2: Control nanoparticles have no impact on P. acnes stimulated PBMC secretion of IL-1β. PBMC cells isolated from normal donors were co-cultured for one hour with control nanopartciles (np) concentrations of 0 mg/ml, 1.25 mg/ml, and 5 mg/ml, respectively. Then, cells were infected by P. acnes at MOI 0.5 for 24 hours at 37 °C. The supernatants were harvested, and expression of IL-1β. Np has no affect in decreasing levels of cytokine secretion such as IL-1β secretion. The data is presented as the mean of triplicate wells ± SD and is representative of three individual experiments.
Acknowledgments
SLH acknowledges NIH grants ES017552-01A2, ES016896-01, P30 ES03850, T32 ES0007060, and AFRL FA8650-05-1-5041.
JDN acknowledges National Institutes of Health/National Institute of Allergy and Infectious Diseases grant 1RC2A1087612-01. AJF acknowledges Dermatology Foundation Career Development Award. JK acknowledges support from NIAMS R01 AR053542.
Abbreviations
- NO
Nitric Oxide
- np
nanoparticle
- SEM
scanning electrom microscopy
- MOI
multiplicity of infection
- NLR
nucleotide oligomerization domain-like receptor
- PBMC
human monocyte
- PEG
polyethylene glycol
- siRNA
small interfering RNA
Footnotes
Conflict of interest: AJF and JMF are co-inventors of the NO-np, a technology that has been licensed to Nano Biomed Inc for commercialization
References
- Bellew S, Thiboutot D, Del Rosso JQ. Pathogenesis of Acne Vulgaris: What's New, What's Interesting and What May Be Clinically Relevant. J Drugs Dermatol. 2011;10:582–5. [PubMed] [Google Scholar]
- Bennett BM, McDonald BJ, Nigam R, et al. Biotransformation of organic nitrates and vascular smooth muscle cell function. Trends Pharmacol Sci. 1994;15:245–9. doi: 10.1016/0165-6147(94)90319-0. [DOI] [PubMed] [Google Scholar]
- Blecher K, Martinez LR, Tuckman-Vernon C, et al. Nitric oxide-releasing nanoparticles accelerate wound healing in NOD-SCID mice. Nanomedicine. 2012 doi: 10.1016/j.nano.2012.02.014. [DOI] [PubMed] [Google Scholar]
- Cabrales P, Han G, Roche C, et al. Sustained release nitric oxide from long-lived circulating nanoparticles. Free radical biology & medicine. 2010;49:530–8. doi: 10.1016/j.freeradbiomed.2010.04.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castro G, Ferreira L. Novel vesicular and particulate drug delivery systems for topical treatment of acne. Expert Opin Drug Deliv. 2008;5:665–79. doi: 10.1517/17425247.5.6.665. [DOI] [PubMed] [Google Scholar]
- Dal Secco D, Paron JA, de Oliveira SH, et al. Neutrophil migration in inflammation: nitric oxide inhibits rolling, adhesion and induces apoptosis. Nitric Oxide. 2003;9:153–64. doi: 10.1016/j.niox.2003.11.001. [DOI] [PubMed] [Google Scholar]
- Dostert C, Petrilli V, Van Bruggen R, et al. Innate immune activation through Nalp3 inflammasome sensing of asbestos and silica. Science. 2008;320:674–7. doi: 10.1126/science.1156995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eady EA. Bacterial resistance in acne. Dermatology. 1998;196:59–66. doi: 10.1159/000017869. [DOI] [PubMed] [Google Scholar]
- Fang FC. Mechanisms of Nitric Oxide –reated Antimicrobial Activities. Journal of Clinical Investigation. 1997;99:2818–825. doi: 10.1172/JCI119473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman A, Blecher K, Sanchez D, et al. Susceptibility of Gram-positive and -negative bacteria to novel nitric oxide-releasing nanoparticle technology. Virulence. 2011;2:217–21. doi: 10.4161/viru.2.3.16161. [DOI] [PubMed] [Google Scholar]
- Friedman A, Friedman J. New biomaterials for the sustained release of nitric oxide: past, present and future. Expert Opinion on Drug Delivery. 2009;6:1113–22. doi: 10.1517/17425240903196743. [DOI] [PubMed] [Google Scholar]
- Friedman A, Han G, Navati M, et al. Sustained release nitric oxide nanoparticles: characterization of a novel delivery platform based on nitrite containing hydrogel/glass composites. Nitric Oxide. 2008;19:12–20. doi: 10.1016/j.niox.2008.04.003. [DOI] [PubMed] [Google Scholar]
- Friedman A, Phan J, Schairer D, et al. Antimicrobial and anti-inflammatory activity of chitosan-alginate nanoparticles: a targeted therapy for cutaneous pathogens. J Invest Dermatol. 2013;133:1231–9. doi: 10.1038/jid.2012.399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman AJ, Blecher K, Schairer D, et al. Improved antimicrobial efficacy with nitric oxide releasing nanoparticle generated S-nitrosoglutathione. Nitric Oxide. 2011 doi: 10.1016/j.niox.2011.09.001. [DOI] [PubMed] [Google Scholar]
- Giustizieri ML, Albanesi C, Scarponi C, et al. Nitric oxide donors suppress chemokine production by keratinocytes in vitro and in vivo. Am J Pathol. 2002;161:1409–18. doi: 10.1016/S0002-9440(10)64416-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gori T, Parker JD. The puzzle of nitrate tolerance: pieces smaller than we thought? Circulation. 2002b;106:2404–8. doi: 10.1161/01.cir.0000036742.52907.91. [DOI] [PubMed] [Google Scholar]
- Grange PA, Raingeaud J, Calvez V, et al. Journal of Dermatological Science. 2009;56:106–112. doi: 10.1016/j.jdermsci.2009.08.001. [DOI] [PubMed] [Google Scholar]
- Griffith OW, Stuehr DJ. Nitric oxide synthases: properties and catalytic mechanism. Annu Rev Physiol. 1995;57:707–36. doi: 10.1146/annurev.ph.57.030195.003423. [DOI] [PubMed] [Google Scholar]
- Han G, Friedman AJ, Friedman JM. Nitric oxide releasing nanoparticle synthesis and characterization. Methods Mol Biol. 2011;704:187–95. doi: 10.1007/978-1-61737-964-2_14. [DOI] [PubMed] [Google Scholar]
- Han G, Zippin JH, Friedman A. From Bench to Bedside: The Therapeutic Potential of Nitric Oxide in Dermatology. Journal of Drugs in Dermatology. 2009;8:586–96. [PubMed] [Google Scholar]
- Hernandez-Cuellar E, Tsuchiya K, Hara H, et al. Cutting edge: nitric oxide inhibits the NLRP3 inflammasome. J Immunol. 2012;189:5113–7. doi: 10.4049/jimmunol.1202479. [DOI] [PubMed] [Google Scholar]
- Jung S, Otberg N, Thiede G, et al. Innovative liposomes as a transfollicular drug delivery system: penetration into porcine hair follicles. J Invest Dermatol. 2006;126:1728–32. doi: 10.1038/sj.jid.5700323. [DOI] [PubMed] [Google Scholar]
- Manconi M, Sinico C, Valenti D, et al. Niosomes as carriers for tretinoin. III. A study into the in vitro cutaneous delivery of vesicle-incorporated tretinoin. Int J Pharm. 2006;311:11–9. doi: 10.1016/j.ijpharm.2005.11.045. [DOI] [PubMed] [Google Scholar]
- Manconi M, Sinico C, Valenti D, et al. Niosomes as carriers for tretinoin. I. Preparation and properties. Int J Pharm. 2002;234:237–48. doi: 10.1016/s0378-5173(01)00971-1. [DOI] [PubMed] [Google Scholar]
- Mao K, Chen S, Chen M, et al. Nitric oxide suppresses NLRP3 inflammasome activation and protects against LPS-induced septic shock. Cell research. 2013;23:201–12. doi: 10.1038/cr.2013.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marshall HE, Hess DT, Stamler JS. S-nitrosylation: physiological regulation of NF-kappaB. Proc Natl Acad Sci U S A. 2004;101:8841–2. doi: 10.1073/pnas.0403034101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez LR, Han G, Chacko M, et al. Antimicrobial and healing efficacy of sustained release nitric oxide nanoparticles against Staphylococcus aureus skin infection. J Invest Dermatol. 2009;129:2463–9. doi: 10.1038/jid.2009.95. [DOI] [PubMed] [Google Scholar]
- Nakatsuji T, Kao MC, Fang JY, et al. Antimicrobial property of lauric acid against Propionibacterium acnes: its therapeutic potential for inflammatory acne vulgaris. J Invest Dermatol. 2009;129:2480–8. doi: 10.1038/jid.2009.93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Privett BJ, Broadnax AD, Bauman SJ, et al. Examination of Bacterial Resistance to Exogenous Nitric Oxide. Nitric Oxide. 2012 doi: 10.1016/j.niox.2012.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin M, Pirouz A, Kim MH, et al. Propionibacterium acnes Induces IL-1beta secretion via the NLRP3 inflammasome in human monocytes. J Invest Dermatol. 2014;134:381–8. doi: 10.1038/jid.2013.309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross JI, Snelling AM, Carnegie E, et al. Antibiotic-resistant acne: lessons from Europe. The British journal of dermatology. 2003;148:467–78. doi: 10.1046/j.1365-2133.2003.05067.x. [DOI] [PubMed] [Google Scholar]
- Schroder K, Tschopp J. The inflammasomes. Cell. 2010;140:821–32. doi: 10.1016/j.cell.2010.01.040. [DOI] [PubMed] [Google Scholar]
- Stamler JS, Lamas S, Fang FC. Nitrosylation. the prototypic redox-based signaling mechanism. Cell. 2001;106:675–83. doi: 10.1016/s0092-8674(01)00495-0. [DOI] [PubMed] [Google Scholar]
- Trivedi NR, Gilliland KL, Zhao W, et al. Gene array expression profiling in acne lesions reveals marked upregulation of genes involved in inflammation and matrix remodeling. J Invest Dermatol; 2006;126:1071–9. doi: 10.1038/sj.jid.5700213. [DOI] [PubMed] [Google Scholar]
- Truong L, Harper SL, Tanguay RL. Evaluation of Embryotoxicity Using the Zebrafish Model. In: Gautier JC, editor. Drug Safety Evaluation: Methods and Protocols. Vol. 691. New York, NY: Humana Press; 2011. pp. 271–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weller RB. Nitric oxide-containing nanoparticles as an antimicrobial agent and enhancer of wound healing. J Invest Dermatol. 2009;129:2335–7. doi: 10.1038/jid.2009.149. [DOI] [PubMed] [Google Scholar]
- Wink DA, Mitchell JB. Chemical biology of nitric oxide: Insights into regulatory, cytotoxic, and cytoprotective mechanisms of nitric oxide. Free Radic Biol Med. 1998;25:434–56. doi: 10.1016/s0891-5849(98)00092-6. [DOI] [PubMed] [Google Scholar]
- Yang JK, Wu WJ, Qi J, et al. TNF-308 G/A polymorphism and risk of acne vulgaris: a meta-analysis. PloS one. 2014;9:e87806. doi: 10.1371/journal.pone.0087806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zouboulis C, Eady A, Philpott M, et al. What is the pathogenesis of acne? Exp Dermatol. 2005;14:143–52. doi: 10.1111/j.0906-6705.2005.0285a.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figure S1: NLRP1 may have a role in NO-np down-regulation of P. acnes induced IL-6 expression in PBMCs, Data shows that there are different levels of P. acnes induced IL-6 down regulation as affected by NLRP1 and NLRP3 (a). Knockout NLRP1 displays a more significant down-regulation of IL-6 than knockout NLRP3. In contrast, neither NLRP1 nor NLRP3 affect P. acnes-induced levels of TNF-α and IL-8 in PBMCs (b). Data are shown as the mean of triplicate wells ± SD and is representative of three individual experiments.
Supplementary Figure S2: Control nanoparticles have no impact on P. acnes stimulated PBMC secretion of IL-1β. PBMC cells isolated from normal donors were co-cultured for one hour with control nanopartciles (np) concentrations of 0 mg/ml, 1.25 mg/ml, and 5 mg/ml, respectively. Then, cells were infected by P. acnes at MOI 0.5 for 24 hours at 37 °C. The supernatants were harvested, and expression of IL-1β. Np has no affect in decreasing levels of cytokine secretion such as IL-1β secretion. The data is presented as the mean of triplicate wells ± SD and is representative of three individual experiments.