

Abstract
Basic and translational research on loiasis, a filarial nematode infection of medical importance, is impeded by a lack of suitable Loa loa infection models and techniques of obtaining and culturing life cycle stages. We describe the development of a new method for routine production of infective third-stage larvae (L3) of L. loa from the natural intermediate arthropod vector host, Chrysops silacea, following experimental infection with purified microfilariae. At 14-days post-infection of C. silacea, the fly survival rate was 43%. Survival was significantly higher in flies injected with 50 mf (55.2%) than those that received 100 mf (31.0%). However, yield per surviving fly and total yield of L3 was markedly higher in the group of flies inoculated with 100 mf (3474 vs 2462 L3 produced). The abdominal segment hosted the highest percentage recovery of L3 (47.7%) followed by head (34.5%) and thorax (17.9%). L. loa larval survival was higher than 90% after 30 days of in vitro culture. The in vitro moulting success rate to the L4 larval stage was 59.1%. After experimental infection of RAG2-/-IL-2γc-/—mice, the average L. loa juvenile adult worm recovery rate was 10.5% at 62 dpi. More than 87% of the worms were recovered from the muscles and subcutaneous tissues. Worms recovered measured an average 24.3 mm and 11.4 mm in length for females (n = 5) and males (n = 5), respectively. In conclusion, L. loa mf injected into C. silacea intrathoracically develop into infective larvae that remain viable and infective comparable to L3 obtained through natural feeding on the human host. This technique further advances the development of a full laboratory life cycle of L. loa where mf derived from experimentally-infected animals may be utilized to passage life cycle generations via intrathoracic injections of wild-caught vector hosts.
Author summary
The Neglected Tropical Disease, loiasis (tropical eye worm disease) affects 13–15 million individuals in Central Africa. Loiasis has recently been identified as a cause of premature mortality and is a barrier to ivermectin-based elimination of onchocerciasis or lymphatic filariasis where co-infections occur, due to the risk of serious adverse events. Loiasis lacks laboratory preclinical models for drug development, biomarker discovery and inflammatory pathology research. Here we detail the successful development of an experimental technique for the laboratory production of Loa loa infective larvae via injection of purified microfilariae into the thorax of wild-caught tabanid flies that are the natural transmission vector. The high yielding infective larvae produced in the laboratory were validated as biologically viable in culture and in a new mouse infection model whereby adult-stage parasites could be produced. The evidence reported herein is an important step to establishing a full laboratory life cycle of L. loa by passage between animal models and experimental injections of the wild-caught vector, Chrysops.
Introduction
Loiasis is a vector-borne, neglected tropical disease (NTD), caused by the tissue dwelling nematode parasite, Loa loa and transmitted by the tabinid flies Chrysops silacea and C. dimidiata [1, 2]. Geographically, the disease is limited by the distribution of its two biting tabanid vectors and is endemic in 11 Western and Central African countries particularly in forested areas [3]. It is estimated that some 14.4 million people live in high risk areas where the prevalence of loiasis is greater than 40%, with 15.2 million in intermediate risk areas where the estimated eye worm prevalence is between 20 and 40% [3]. The common clinical signs of loiasis are migration of the adult worm, Calabar swelling, pruritis and arthralgia [4]. Loiasis is a neglected but nevertheless epidemiologically relevant tropical disease which has, until latterly, attracted only limited attention in drug or vaccine research and development. In the past two decades, interest in this disease has increased due to reports of severe adverse events (SAEs) post-ivermectin during mass treatment of the priority NTD, onchocerciasis (river blindness) in areas of onchocerciasis / loiasis co-endemicity. This situation has hampered the mass drug administration (MDA) with ivermectin for the elimination of onchocerciasis in areas where L. loa prevalence exceeds 20% [5, 6]. Therefore, this has impeded the goals stated by the World Health Organization, to eliminate onchocerciasis as a public health problem by 2030. This includes the permanent protection of the remaining 120 million people at risk of this debilitating and disfiguring disease in 19 countries in Africa through the establishment of community-directed treatment with ivermectin (CDTi), in areas of co-endemicity. Presently, there is no satisfactory treatment for loiasis though two medications have so far been employed for clinical cases namely, diethylcarbamazine (DEC) and albendazole. Several cases of coma and even death have been reported in individuals with high microfilarial (mf) loads of L. loa when treated with DEC [7]. Whilst albendazole-based clinical trials have shown promise in lowering Loa microfilaraemias, more precise dose optimisations are required to define a minimally sufficient and safe dosing regimen. If achievable, albendazole short-courses or other, new, safe anti-loiasis drugs could be administered before MDA with ivermectin in areas of co-endemicity [8, 9]. Loiasis drug discovery and development necessitates the convenient and abundant availability of parasite material suitable for screening compounds both in vitro and in vivo. While the mf stages can be purified from the blood of infected humans or primates, the production of infective larvae requires infection of the Chrysops vectors with the mf. Several methods have been used to produce infective larvae of filariae from flies and include recovery of L3 from engorged flies on consenting microfilaraemic donors (human landing catch) [10–12], baited traps (wood fire) and multiple fly dissections from hyperendemic hot-spots of infection and artificial membrane feeding systems [13, 14]. However, applying the latter technique for infection of Chrysops for L3 production have been unsuccessful as these flies are refractory to feeding in captivity. Bianco et al. [15], Dohnal et al. [16] and Fukuda et al. [17] produced infective larvae of Onchocerca species by experimentally infecting Simulium blackfly intrathoracically with mf. Intrathoracic injections of flies with mf bypass feeding of flies on donors, and therefore address the ethical concerns of human landing catch. The use of animal sources of mf, such as splenectomised baboons [11] or immunodeficient mice [18] would potentially completely obviate the use of human donors and may achieve a complete laboratory life cycle local to a source of wild-caught vectors. Further, high yielding production of L3 following experimental infections of Chrysops would be a significant improvement in throughput for translational medicine applications compared to standard methods. In this study, we assessed if intrathoracic L. loa mf injected C. silacea can survive under laboratory conditions for a minimum of two weeks and whether L3 infective larvae recovered from lab maintained flies could maintain their viability and development both in vitro and in vivo taking advantage of recent advances in L. loa culture systems and mouse models developed by our laboratories [12, 18].
Methods
Ethics statement
The study design and protocols were approved by the REFOTDE Institutional Animal Ethics Committee (RIAEC), and ethical clearances (001/RIAEC/2015) were issued. Animal experimentation were in strict accordance with the international guidelines of rearing animals and use in medical research, the Animal Welfare Legislation and Policies and complied with the Animals (Scientific Procedures) Act 1986 (ASPA) and its associated codes of practice on animal housing and care [19]. Previous works that used the same procedures are found here [18].
Experimental animals
RAG2-/-IL-2γc-/- C57BL/6 mice used in the experiment were obtained from the Institute of Medical Microbiology, Immunology and Parasitology (IMMIP), University of Bonn, Germany. Mice were shipped in filter topped boxes to Research Foundation for Tropical Diseases and the Environment (REFOTDE), Buea, Cameroon. All mice were reared at the REFOTDE animal facility in a 12:12 light: dark cycle, maintained in individually ventilated caging (IVC) with HEPA filtered air system (Techniplast) with ad libitum provision of standard irradiated rodent chow and bottled mineral water. All mice used for the experiments were infected in the laboratory of the REFOTDE. All mouse procedures received ethical approval by the REFOTDE Institutional Ethics Animal Committee (RIEAC) and were undertaken in accordance with UK standards for use of animals in research.
Loa loa microfilariae (mf) were obtained from splenectomised infected baboons (Papio anubis) that were kept in captivity and infected with the L. loa human strain [11]. The acquisition, care and ethical concerns on the use of baboons as donors of mf have been previously documented [10, 11]. Ethical and administrative approvals for the use of baboons in this study were obtained from the Ministry of Scientific Research and Innovation of Cameroon (Research permit #028/ MINRESI/ B00/ C00/ C10/ C12) and the RIEAC. Procedures adhered to the NIH Guide for the Care and Use of Laboratory Animals.
Collection of wild C. silacea
Wild C. silacea (with an infection rate of less than 1%) were collected from Kindongi rainforest reserve [1] located in the South-West Region of Cameroon. Catches were made from 8 a. m. to 4 p.m. during the mild rainy and mild dry seasons especially during sunny hours. C. silacea is the only vector species found in this study site [1]. Flies were attracted by the smoke from wood fire, then caught using sweep nets (with multiple fly dissections to control for any naturally acquired infection) and transferred to individually labelled 50 mL Falcon tubes (Corning, USA) prepared to provide suitable conditions for their survival during their transportation to the laboratory in a cold box as described by Wanji et al. [11].
Isolation and purification of L. loa mf from blood
L. loa mf were obtained from experimentally infected baboons (Papio anubis) with the human strain of L. loa. The infection and follow up of these animals were previously described [11]. Approximately 8–10 mL venous blood was collected from microfilaraemic baboons around 9 am into 10 ml EDTA coated tubes (Corning, USA) and transported within 1 hour to the laboratory for processing.
L. loa mf were extracted using Percoll gradient centrifugation as described by Cesbron et al. [20] with modifications [21]. Briefly, an aliquot of 2.5 ml of baboon whole blood was pipetted and layered gently onto a Percoll (GE Healthcare, Pharmacia, Uppsala, Sweden) gradient (40,50 and 65%) in a 15 ml centrifuge tube (Corning, Kennebunk-ME, USA) and centrifuged (Humax 14k human, Wiesbaden, Germany) at 3000 rpm for 10 minutes at 25°C. The layer of Percoll gradient containing microfilariae was removed using a syringe and pumped gently through a 5 μm pore size cellulose filter into a 50 ml falcon tube. The syringe was carefully removed and 10 ml of RPMI-1640 (Sigma, USA) was aspirated into the syringe to wash the filter. The filter was transferred immediately to a Petri dish containing culture medium and incubated at 37°C for 5 minutes. The filter was then removed from the Petri dish and the suspension centrifuged at 1500 rpm for 10 minutes to concentrate the parasites which were then quantified using an inverted microscope. Concentrations of 50 mf and 100 mf load/ 15 μl of medium were prepared as previously described [22].
Injection of C. silacea with L. loa mf
Prior to injection, the flies were knocked down by placing the 50 ml Falcon tube containing the flies at -20°C for 5 minutes. Each fly was removed from the tube, orientated in a supine aspect, and held in place with sterile forceps on a cold light source platform of a dissecting microscope. The fly was then injected at the left side through the blind spot of the thoracic region with a 15μl of medium containing 50 (group 1) or 100 (group 2) mf using a 0.5 ml insulin syringe (Fig 1). After injection, flies were placed into individual rearing tubes and monitored for survival.
Fig 1. Injection of Loa loa mf into C. silacea.

Here, 0.5 ml insulin syringe containing Loa loa mf is introduced to a ventral positioned Chrysops fly held with a pair of forceps.
Laboratory maintenance of infected C. silacea
Flies were transferred to the insectarium and monitored daily for 14 days (intrinsic incubation period for mf development to L3 stage). During this period, flies were fed daily with a sterile 15% sucrose solution. The temperature of the insectarium was maintained between 21–23°C with 79% - 80% relative humidity [10]. Fly mortality was recorded daily to determine the survival rate.
Dissection of flies for L. loa L3 recovery
At 14 days post infection (p.i.), flies were placed in sterile distilled water containing 0.2% Tween 20 (Sigma-Aldrich, St Louis, USA) in a sterile Petri dish for 1 min, transferred to a cleaning solution (sterile distilled water) and placed into a Petri dish of diameter 5.5 cm containing dissecting medium (RPMI-1640 medium; Sigma-Aldrich, St Louis, USA) using sterile forceps. The head, thorax and abdomen were separated from each other, placed in Petri dishes and dissected with sterile needles under a dissecting microscope (Motic, Wetzlar, Germany). Emerging larvae were collected and transferred with a sterile micropipette into the first 3 wells of a depression or concentration plate filled with sterile RPMI 1640 medium. Infective larvae from head, thorax and abdomen were recovered and processed separately. The larvae were washed 3 times by transferring into different depression wells. Isolated parasites were quantified, and motility/survival rates assessed using an inverted microscope (Motic AE 21 series, Wetzlar, Germany).
In vitro culture
Infectious L3 larvae were transferred into centrifuge tubes containing dissection medium using a sterile pipette. The infective larvae were purified by a Percoll gradient technique [21]. Briefly, L3 were centrifuged (Centrifuge; HUMAX 14K HUMAN, Germany) at 1500 rpm for 10 mins and later concentrated in less than 1 ml RPMI. The pellet was slowly layered on the surface of a 15 ml tube containing stock iso-osmotic Percoll (GE Healthcare, Pharmacia, Uppsala, Sweden) and centrifuged at 800 rpm for 10 minutes. The process was repeated to reduce contaminants. At the end, the L3 were washed twice with RPMI-1640 by centrifugation at 1500 rpm for 10 minutes to remove residual Percoll. The L3 were then cultured in Dulbecco’s Modified Eagle Medium (DMEM; Gibco Life Technologies, Cergy-Pontoise, France) supplemented with 10% foetal bovine serum (FBS; Lonza Verviers, Belgium) on monkey kidney epithelial cells (LLC-MK2; ATCC, Virginia, USA) in a flat bottom 48 well plates (Corning, Kennebunk-ME, USA) containing lids for a maximum period of 30 days at 37° C, 5% CO2 in an incubator (CO2 series Sheldon Mfg. Inch, Cornelius, OR, USA) while monitoring for parasite motility, survival and moulting [12]. Parasite viability during culture was assessed daily by visual inspection under an inverted microscope for 30 days. Their motility was scored on a 4 point scale: 0; no movement or immotile, 1; intermittent shaking of head and tail, 2; sluggish (shaking of the whole worm on a spot), 3; vigorous movement (shaking of the whole worm and migration from one spot to another) as previously described [12, 22].
Experimental mouse infection
The L3 larvae were washed and quantified under a dissecting microscope and concentrated in approximately 100–200 μL of RPMI in a 0.5 ml insulin syringe. We have recently established mouse models of loiasis utilising Severe-combined / γc-/- compound immunodeficient mouse strains [18]. We experimentally infected groups of 5–6 mice subcutaneously in the lumbar area [10] with L3 (ranging from 50–140) derived from the head or abdomen of experimentally-injected C. silacea. Due to limited numbers of L3 recovered from the thorax of C. silacea, a single mouse was infected and an additional five mice were infected with a mixture of L3 derived from the thorax and abdomen. Infected animals were euthanized by a rising concentration of CO2 and cervical dislocation to confirm death before dissection 62 days p.i. for parasite recovery. The protocol used in the dissection of mice was described in previous studies [18, 24]. Initially, blood was collected by cardiac puncture and two slides of 50 μL each prepared (stained with Giemsa and viewed under a microscope). This was done to check for the absence of L. loa mf [11]. Animals were dissected, and the different tissues were gently teased out and incubated in RPMI medium to allow for free migration of larvae into the medium as previously described [10, 23]. Specifically, the skin (subcutaneous tissues), muscles, peritoneal and pleural cavities, heart and lung tissues were incubated in separate petri dishes. The larvae recovered were counted and the site of recovery noted. The larvae recovered were stored in 80% ethanol for measurement of their lengths and widths. For morphological observations, 5 female and 5 male worms were treated with Amann lactophenol and viewed under a light microscope as described by Ushirogawa et al. [24] and Bain et al. [25].
Statistical analysis
The survival rates of the flies at each time point was estimated as the percentage of the remaining number of flies alive for each batch. Mann-Whitney-U-Test and Chi square test were used to compare the survival rate (%) at day 14 between the two fly groups. For L3 recovery, test for normality was done using Anderson-Darling test and L3 recovery between groups was compared using independent-Samples median test. The L3 recovery from different fly anatomical locations were compared using Friedman test (Head vs Thorax vs Abdomen) and Wilcoxon Signed Rank test to compare pairwise (Head vs Thorax, Head vs Abdomen and Thorax vs Abdomen). Mean motility and mean survival were computed to assess the viability of the parasites in vitro.
Cuzick test for trend was used to compare the moulting rate of L. loa L3 from different parts of the C. silacea. The proportions of larvae recovered from each region of the mice and mean number (%) of larvae recovered (with respect to provenance of L3) were compared using the Chi-square and Kruskal-Wallis tests. All the tests were performed at 5% level of significance. Statistical analysis was undertaken using SPSS version 20, GraphPad Prism 8.0.2, and the PMCMRplus package in R version 3.6.1.
Results
C. silacea experimentally injected with L. loa mf survive and produced L3 larvae
Survival of flies injected with either 50 or 100 purified L. loa mf after 14 days are detailed in Table 1 and Additional flies 1 and 2. Out of 428 C. silacea injected with L. loa mf, 184 survived (42.0%) and were dissected for L3 recovery. At day 14 p.i., group 1 (50 mf inoculated) recorded a higher survival rate compared to group 2 (100 mf inoculated) (55.2% vs 31.0% survival, χ2 = 18.1, P<0.001)
Table 1. C. silacea survival rate and number of L3 of L. loa recovered 14 days following intrathoracic injection of mf.
| Fly group | mf load injected / fly | No. flies injected | No. of flies surviving, day 14 |
Fly survival rate, day 14 (%) |
Percentage of Chrysops with L3 (%) | Median L3 recovered / fly | Median L3 yield / fly (%) | Total L3 recovered |
|---|---|---|---|---|---|---|---|---|
| 1 | 50 | 212 | 117 | 55.19* | 100 | 19 | 42.08 | 2462 |
| 2 | 100 | 216 | 67 | 31.01 | 100 | 56* | 51.84 | 3473 |
*χ2 = 18.12 P<0.001 proportion survival of flies between groups, *Independent-Samples median test P<0.001 (median L3 recovered/fly). All surviving flies were dissected for L3 recovery
A total of 184 C. silacea were dissected and the recovered L3 were quantified. At least 2 infective L3 were recovered from each dissected fly. The median L3 recovered and parasite yields per injected Chrysops fly were calculated (Table 1). The median L3 recovered per surviving fly in group 2 was significantly greater than in group 1 (56 vs 19, P < 0.001, independent samples median test). Despite the significantly higher mortality rate in group 2, a higher total yield of L3 was produced from surviving flies in group 2 compared with group 1 (3473 vs 2462).
The highest recovery percentage of infective larvae was from the abdomen (47.7%) followed by the head (34.5%) and the thorax (17.9%). Variation in percentage recovery between the various fly segments was statistically significant (P<0.001, Friedman Test, Table 2). Subsequent post-hoc testing, comparing average recovery (%) of infective larvae between fly segments, determined that L3 recovery was significantly highest in the abdomen compared with either thorax or head (P<0.001 and P = 0.012, respectively, Wilcoxon Signed Rank Tests) but that yields in the head were also significantly higher than the thorax (P = 0.001, Wilcoxon Signed Rank Test). The distribution of L3 in the three anatomical locations was not different in flies receiving 50 or 100 injected mf.
Table 2. Average L3 recovered from different parts of C. silacea injected intrathoracically with 50 and 100 mf of L. loa at day 14 p.i.
| Group | Number of flies dissected | % L3 recovered from head (mean±SD) |
% L3 recovered from thorax (mean±SD) |
(%) L3 recovered from abdomen (mean±SD) |
|---|---|---|---|---|
| 1 | 117 | 36.1 (7.6±9.8) |
16.7 (3.5±3.4) |
47.2 (9.94±7.0) |
| 2 | 67 | 33.3 (17.2±21.0) |
18.7 (9.7±12.1) |
48.0 (24.87 ±21.43) |
| Total | 184 | 34.5^ (11.1±15.5) |
17.9 (5.8±8.3) |
47.7*#‡ (15.37 ±15.77) |
p-values for comparison of recovery rate of L. loa L3 from different sections of the C. silacea body:
*Head vs thorax vs abdomen: P <0.001; Friedman’s test
^Head vs thorax: P<0.001; Wilcoxon signed rank test
#Head vs abdomen: P = 0.012; Wilcoxon signed rank test
‡Thorax vs abdomen: P<0.001; Wilcoxon signed rank test
L. loa L3 generated from mf injections of C. silacea develop to fourth-stage larvae and survive within long-term in vitro cultures
We have recently developed a long-term mammalian cell co-culture system whereby L. loa L3 isolated from wild-caught C. silacea remain ≥90% viable for 18 days, with growth and 20–40% moulting success rate to the L4 stage [12]. We exploited these defined in vitro culture conditions to evaluate the in vitro fitness of experimentally-derived L. loa L3. Survival and average motility scores of developing L. loa larvae in culture are plotted in Fig 2. The survival of larvae remained >90% throughout the course of the experiment (30 days). At day 5, motility was greater than 80% and decreased to 36% (day 20) and stayed constant.
Fig 2. Survival and motility monitoring of L. loa L3 derived from mf injections of C. silacea, in mammalian in vitro culture conditions.
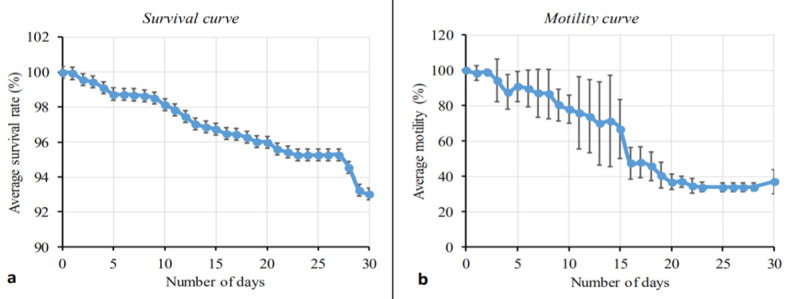
Percentage survival curve (A) and motility curve (B) of Loa loa larvae during 30 days of culture. Plotted is the average (mean) response. Bars indicate standard deviations (SD).
Of the 2462 L3 recovered from flies that were injected with 50 mf, 2453 were successfully placed into culture. Subsequently, 1450/2453 larvae underwent moulting (from L3 stage to L4 stage; Fig 3 & Table 3). Whilst there was a trend for higher moulting rate in larvae derived from the head (61.2%) followed by 60.1% and 56.4% for those derived from the thorax and abdominal segments, respectively (Table 3), this did not attain significance (P = 0.975, Cuzick trend test).
Fig 3. Representative image of an in vitro L. loa L4 larvae and cuticle cast.

Image taken by inverted tissue culture microscope. Arrows: red = fully moulted Loa L4; black = Cast cuticle from moulted Loa L3. Scale bar 100μM.
Table 3. in vitro culture moulting success of L. loa L3 derived from mf injection of C. silacea.
| Parameters | Head | Thorax | Abdomen | Total |
|---|---|---|---|---|
| n L3 plated | 1206 | 502 | 745 | 2453 |
| n L3 moulted | 738 | 297 | 415 | 1450 |
| % moulted | 61.2 | 59.2 | 55.7 | 59.1 |
| Statistics | Cuzick trend test: z = 0.0311, p = 0.975, | |||
L. loa L3 generated from mf injections of C. silacea can successfully infect immunodeficient mice and develop into adult filariae
All worms recovered from mice 62dpi were young adults (Fig 4). The average length and width of female worms recovered was 24.6 mm and 0.35 mm respectively and 11.4 mm and 0.27 mm for male worms (n = 5 worms measured per sex).
Fig 4. Loa loa adult morphology 62 days post-inoculation.

(A and B) Head of young adult female stage (anterior region); Mo–mouth opening; Eo—esophagus. (c): Vulva [Vu] of mature young adult female (lateral view). (D): Open vulva, intestine, uterus of female young adult worm with no mf present (indicating that the worm is not fully developed [lateral view]). (E and F): Posterior extremity of matured young adult female worm (E; lateral view [An–anus] and F; ventral view) with caudal papillae [Pa]. (G and H) Caudal region (lateral view) of Loa loa young adult male with caudal papillae (G) and spicule (H). [A, D, E, G viewed under x10 objective, B, C viewed under x40 objective F, H viewed under x100 objective].
Worms were recovered from the subcutaneous tissues, muscles, peritonea and pleural cavities (Table 4 and Additional file 3). Overall, more than 87% of worms were recovered from the muscles and subcutaneous tissues, the natural niche of adults in humans and primates [2, 11]. The proportions of worms recovered from the different anatomical regions of mice were therefore significantly different (χ2 = 33.8, P<0.001). The provenance of L3 from experimentally infected C. silacea, in terms of head, thorax and/or abdomen, did not significantly affect the recovery rate of adult L. loa worms from mice 62 dpi (Kruskal Wallis test P = 0.42), although there was a trend for higher recoveries when L3 derived from the head of the vector were used.
Table 4. Recovery of adult L. loa from RAG2-/-IL-2γc-/- mice infected with L3 derived from experimental injections of C. silacea.
| L3 Source |
Total L3 inoculated |
Mice N |
Site of parasite recovery (% total recovered) |
Total adult L. loa recovered (% L3 inoculate) |
Median adult L. loa recovererd / mouse % L3 inoculate (IR)* |
|||
|---|---|---|---|---|---|---|---|---|
| Sub- Cut. |
muscle | Peritonea | Pleura | |||||
| Head | 489 |
5 | 28 (33.3) | 44 (52.4) | 3 (3.6) |
9 (10.7) | 84 (17.2) |
20.8 (1.8–22.2) |
| Abdomen | 565 |
6 | 21 (45.7) | 19 (41.3) | 5 (10.9) |
1 (2.2) |
46 (8.1) |
7.5 (5.5–10.1) |
| Thorax | 123 |
1 | 2 (16.7) |
9 (75) |
1 (8.3) |
0 (0) |
12 (9.8) |
9.8 |
| Thorax + Abdomen | 472 |
6 | 15 (48.4) | 15 (48.4) | 1 (3.2) |
0 (0) |
31 (6.6) |
4.7 (2.2–10.9) |
| All tissues | 1649 | 18 | 66* (38.2) |
87 (50.3) |
10 (5.8) |
10 (5.8) |
173 (10.5) |
7.5 (3.3–12.9) |
Infective larvae of 50–140 were inoculated per mice depending on recovery from fly anatomical regions
*X2 = 33.8, P<0.001, proportion of adults parasitizing specific tissues
Discussion
In this study, we establish, for the first time, that L. loa mf purified from the blood of an experimental baboon host can be successfully injected into wild-caught, laboratory-maintained C. silacea to yield viable infectious stage L3 larvae. The yields, quality and infectivity of the infective larvae produced were confirmed via novel in vitro and in vivo systems recently developed by our laboratories to be comparable to L. loa L3 derived from C. silacea human landing catches [12] or wood-smoke baited traps [18].
We scrutinised two microfilaraemic doses because we predicted a trade-off between numbers of L3 produced and fly mortality due to high level L. loa infection and collateral damage to fly tissues as L. loa migrate and develop from the point of injection. Despite the higher mortality rate of flies injected with 100 mf, we determined that C. silacea surviving to day 14 yielded significantly higher numbers of L3 per batch compared with flies inoculated with 50 mf. The higher mortality in 100 versus 50 mf injections indicates that infectious dose is the major factor affecting survival of the flies. However, sham infections need to be performed in future to determine what impact the injection technique has on fly survival. Because of the reduced labour time required to dissect the fewer surviving, high-yielding flies, we conclude that 100 mf injections are most suitable for the production of L. loa L3.
We evaluated the biological fitness of L3 derived from experimental inoculations of C. silacea in three ways: 1. The ability to shed their cuticles (moult) to develop to the L4 stage 2. Their survival in culture over 30 days and 3. Their ability to infect and establish adult infections in susceptible immunodeficient mice after two months. In foetal calf serum supplemented DMEM medium and incubation at standard human tissue culture conditions, we have prior determined that between 20–40% of L3 derived from human-fed C. silacea complete the L3-L4 moult and survival, assessed by motility, continues in >90% of cultured larvae for 18 days [12]. In the experiments reported here for experimentally-derived L3, 59.1% completed the L3-L4 moult and >90% survival was recorded until the end of the experiment at day 30. In L. loa, the moulting of L3 to L4 stage occurs near the end of the first week of infection / incubation [25]. Bain and colleagues reported moults of L3 to L4 stage of L. loa at day 8 after inoculating infective larvae of L. loa recovered from Chrysops into mice, whilst our prior in vitro findings moulting started from day 9 [12]. Here we observed moulting of L3 to L4 stage was characterised by marked decrease in the activity of the larvae, that was observed as a drop in motility which stabilized at day 23 and a slight increase in motility observed towards day 30 when the majority of larvae had completed their moult.
Our in vitro observations suggest that in vitro ‘fitness’ of experimentally derived L3 is at least comparable, if not superior, to L3 derived from naturally-fed flies caught after landing on infected human volunteers and subsequently maintained in our laboratory insectary. Whilst several direct individual experimental comparisons would be required to determine the superior biological fitness of experimentally-reared L. loa L3, the data supports the use of experimentally-derived L3 in drug screening or other biological in vitro studies.
Utilising our recently established L. loa mouse model system [18] immature adult-stage worms were consistently recovered from RAG2-/-γc-/- mice at day 62 post-inoculation, with a 10.5% yield of the initial unit inoculum, predominantly from the muscle and subcutaneous tissues; the natural L. loa adult parasite niche. The immature adult L. loa stage was verified by identification of sex-specific organs and morphological characteristics, as well as the macroscopic size of the filariae recovered. These findings emulate our studies where experimental infections of immunodeficient mice were undertaken utilising L3 from blood fed Chrysops both in terms of the matching predominant anatomical locations of adult infections and the yields evident [10, 18]. Following infections of both baboons and immunodeficient mice, the development of patent infections, with mf production seeding the blood, are detectable at between 5 and 6 months [11,18]. Given comparable yields and presence of both male and female immature worms in mice infected with experimentally generated L3, we would predict that patent infections would be possible, and this is the focus of ongoing work. If patent infections can be established, the successful derivation of L3 via experimental injections of mf into C. silacea could be utilised to sequentially passage L. loa between mice and wild-caught flies to establish a laboratory experimental life-cycle of L. loa.
We determined that at day 14 post-injection, the abdominal fly segment recorded the highest proportion of L3, followed by L3 isolated from the head with a minority of L3 derived from the thorax. The abdominal fat bodies serve as the site of mf-L1-L2-L3 development in Chrysops [26]. Following mf development to L3 in the abdomen, L3 migrate to the head via the thorax and only the L3 in the head are positioned for transmission when infected flies take a subsequent blood meal [27]. Because it is speculated that L3 derived from the head might be more biologically adapted to successfully transition to mammalian host conditions, we scrutinised whether biological fitness of L3 to develop to L4 in vitro or immature adults in vivo was associated with the fly anatomical segments they were isolated from. We observed a trend for both increased moulting rate in vitro and increased immature adult L. loa yields in vivo when utilising ‘infective’ L3 derived from the head of C. silacea. However, the majority of L3 derived from the thorax and abdomen could also undergo moulting and L3 isolated from these fly segments could also develop to adult infections in mice. The differences in moulting and adult establishment were not statistically different between L3 isolated from the three fly segments. Our results support the use of pooled L3 derived from all segments of C. silacea in onward biological and translational medicine research such as in vitro and in vivo drug studies. This has the advantage of significantly increasing yields of L3 from experimental injections for onward applications.
Conclusions
This study is the first demonstrating the production of L. loa infective L3 larvae from C. silacea inoculated with L. loa mf through intrathoracic injections. The quality of infective L. loa larvae produced using this method were confirmed in in vitro and in vivo experimental systems. This technique creates a new platform for the generation of high yields of L. loa L3 to increase throughput for in vitro and in vivo drug screening experiments. We also conclude that via iterative experimental injections of C. silacea and passage in mice to provide patent infections, a complete L. loa laboratory life cycle is now achievable utilising wild-caught flies. These findings obviate the necessity for human volunteers and non-human primates to generate L. loa life-cycle stages and will accelerate translational medical research on this Neglected Tropical Disease by providing a more abundant source of parasitic material to researchers in need.
Acknowledgments
We thank the local communities for allowing access and assisting in the trapping of C. silacea.
Data Availability
All relevant data are within the manuscript.
Funding Statement
This work was supported by a Bill & Melinda Gates Foundation Grand Challenges Explorations grant (no. OPP1119043) to J.D.T., S.W., and M.J.T and a grant awarded to M.R., A.H., S.W from the German Research Council (DFG) (Grant DFG RI 3036/1-1) within the “African-German Cooperation Projects in Infectiology.” The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Wanji S, Tendongfor N, Esum M, Enyong P. Chrysops silacea biting densities and transmission potential in an endemic area of human loiasis in South-West Cameroon. Trop Med Int Health. 2001;7:371–377. [DOI] [PubMed] [Google Scholar]
- 2.Padgett JJ, Kathryn HJ. “Loiasis: African eye worm” Trans R Soc Trop Med Hyg. 2008;102:983–989. 10.1016/j.trstmh.2008.03.022 [DOI] [PubMed] [Google Scholar]
- 3.Zouré HGM, Wanji S, Mounkaïla N, Uche VA, Diggle PJ, Tekle AHT, Remme JHF. The Geographic Distribution of Loa loa in Africa: Results of Large-Scale Implementation of the Rapid Assessment Procedure for Loiasis (RAPLOA). Plos Neglected Tropical Diseases. 2011;5(6). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lukiana T, Mandina M, Situakibanza NH, Mbula MM, Lepira BF, Odio WT, Kamgo J, Boussinesq M. A possible case of spontaneous Loa loa encephalopathy associated with a glomerulopathy. Filarial Journal. 2006;5:6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Boussinesq M, Gardon J, Kamgno J, Pion SD, Gardon-Wendel N. Relationship between the prevalence and intensity of Loa loa infection in the Central province of Cameroon. Ann Trop Med Parasitol. 2001;95:495–507 10.1080/00034980120073184 [DOI] [PubMed] [Google Scholar]
- 6.Boussineq M, Kamgno J, Pion SD, Gardon J. What are the mechanisms associated with post–ivermectin serious adverse events? Trends in Parasitology. 2006;22:244–246. 10.1016/j.pt.2006.04.006 [DOI] [PubMed] [Google Scholar]
- 7.CDC–Center for Diseases Control and Prevention (2016) Parasites–Loiasis. http: //www.cdc.gov/parasites/loiasis/treatment.html. Accessed January 2016.
- 8.D'Ambrosio MV, Bakalar M, Bennuru S, Reber C, Skandarajah A, Nilsson L, Switz N, Kamgno J, Pion S, Boussinesq M, Nutman TB, Fletcher DA. Point-of-care quantification of blood-borne filarial parasites with a mobile phone microscope. Sci Transl Med. 2015;7(286):286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bennuru S, Pion SD, Kamgno J, Wanji S, Nutman TB. Repurposed automated handheld counter as a point-of-care tool to identify individuals 'at risk' of serious post-ivermectin encephalopathy. PLoS Negl Trop Dis. 2014;8(9): e3180 10.1371/journal.pntd.0003180 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Tendongfor N, Wanji S, Ngwa JC, Esum ME, Specht S, Enyong P, Matthaei KI, Hoerauf A. The human parasite Loa loa in cytokine and cytokine receptor gene knock out BALB/ c mice: survival, development and localization. Parasites & Vectors. 2012;5:43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wanji S, Eyong E-E, Tendongfor N, Ngwa C, Esuka E, Kengne-Ouafo A, Datchoua-Poutcheu F, Enyong P, Hopkins A, Mackenzie CD. Parasitological, Hematological and Biochemical Characteristics of a Model of Hyper-microfilariaemic Loiasis (Loa loa) in the Baboon (Papio anubis). PLoS Negl Trop Dis. 2015;9(11). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zofou D, Fombad FF, Gandjui NVT, Njouendou AJ, Kengne-Ouafo AJ, Ndongmo PWC, Datchoua-Poutcheu FR, Enyong PA, Bita DT, Taylor MJ, Turner JD, Wanji S. Evaluation of in vitro culture systems for the maintenance of microfilariae and infective larvae of Loa loa Parasites & Vectors. 2018;11:275 10.1186/s13071-018-2852-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Albuquerque CMR, Cavalcanti VMS, Melo MAAM, Verçosa P, Regis LN, Hurd H. Blood meal microfilariae density and the uptake and establishment of Wuchereria bancrofti infections in Culex quinquefasciastus and Aedes aegypti. Mem Inst Oswaldo Cruz. 1991;94 (5):591–596. [DOI] [PubMed] [Google Scholar]
- 14.Paily KP, Hoti SL, Balaraman K. Development of lymphatic filarial parasite Wuchereria bancrofti (Spirurida: Onchocercidae) in mosquito species (Diptera: Culicidae) fed artificially on microfilaremic blood. J Med Entomol. 2006;43(6):1222–6. 10.1603/0022-2585(2006)43[1222:dolfpw]2.0.co;2 [DOI] [PubMed] [Google Scholar]
- 15.Bianco AE, Ham PG, Townson S, Mustafa MB, Nelson GS. A semi-automated system of intrathoracic infection for the large scale production of Onchocerca Lienalis infective larvae. Trop Med Parasitol. 1989;40(1):57–64. [PubMed] [Google Scholar]
- 16.Dohnel J, Blinn J, Wahl G, Shhulz-Key H. Distribution of microfilariae of O. lienalis and O. gutturosa in the skin of cattle in Germany and their development in Simulium Ornatum and Culicoides nuberulosis following artificial infestation. Vet parasitol. 1990;36(3–4):325–32. 10.1016/0304-4017(90)90044-c [DOI] [PubMed] [Google Scholar]
- 17.Fukuda M, Takaoka H, Uni S, Bain O. Infective larvae of five Onchocerca species from experimentally infected Simulium species in an area of zoonotic onchocerciasis in Japan. Parasite. 2008;15:111–119. 10.1051/parasite/2008152111 [DOI] [PubMed] [Google Scholar]
- 18.Pionnier NP, Sjoberg H, Chunda VC, Fombad FF, Chounna PW, Njouendou AJ, Metuge HM, Ndzeshang BL, Gandjui NV, Akumtoh DN, Tayong DB, Taylor MJ, Wanji S, Turner JD. Mouse models of Loa loa. Nature Communications. 2019;10:1429 10.1038/s41467-019-09442-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hollands C. The Animals (Scientific Procedures) Act 1986. Lancet 1986;328(8497):32–33. [DOI] [PubMed] [Google Scholar]
- 20.Cesbron JY, Chandenier J, Taelman H, Henry D, Capron A. Density gradient separation of Loa loa and Dipetalonema perstans microfiariae from infected patients. Annales de la Societe Belge de Medecine Tropicale. 1989;66:77–78. [PubMed] [Google Scholar]
- 21.Wanji S, Amvongo-Adjia NN, Njouendou AJ, Kengne-Ouafo JA, Ndongmo WPC, Fombad FF, Koudou B, Enyong PA, Bockarie M. Further evidence of the cross-reactivity of the Binax NOW Filariasis ICT cards to non-Wuchereria bancrofti filariae: Experimental studies with Loa loa and Onchocerca ochengi. Parasites & Vectors. 2016;9:267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Njouendou AJ, Fombad FF, O’Neill M, Zofou D, Nutting C, Ndongmo PC, Kengne-Ouafo AJ, Geary TG, Mackenzie CD, Wanji S. Heterogeneity in the in vitro susceptibility of Loa loa microfilariae to drugs commonly used in parasitological infections. Parasites and Vectors. 2018;11:223 10.1186/s13071-018-2799-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wanji S, Tendongfor N, Vuong PN, Enyong P, Bain O. The migration and localization of Loa loa infective and fourth stage larvae in normal and immuno-suppressed rodents, consequences in human pathology. Ann Trop Med Parasit. 2002;96(8):823–830 10.1179/000349802125002220 [DOI] [PubMed] [Google Scholar]
- 24.Ushirogawa H, Orino T, Hatsushika R, Tabuchi A. Morphological studies of the adult female Loa loa (Nematoda: Filarioidea) and a review of the literature on imported human loiasis in Japan. Kawasaki med J. 2003;29(3.4):53–59. [Google Scholar]
- 25.Bain O, Wanji S, Enyong P, Petit G, Noireau F, Eberhard MI, Wahl G. New features on the moults and morphogenesis of the human filaria Loa loa by using rodents hosts consequences. Parasites. 1998;5:37–46. [DOI] [PubMed] [Google Scholar]
- 26.Orihel TC, Lowrie RC. Loa loa: development to the infective stage in an American deerfly, Chrysops atlanticus. Am J Trop Med Hyg. 1995;24(4):610–5. [DOI] [PubMed] [Google Scholar]
- 27.Lavoipierre MMG. The sites of development and the migration of Loa loa in Chrysops silacea, the escape of the infective forms from the head on the fly and the effect of the worm on its insect hosts. Trans. R. Soc. Med. Hyg. 1958;52:103–121. [DOI] [PubMed] [Google Scholar]